Back to Journals » OncoTargets and Therapy » Volume 11
Pleiotropic functions of miR107 in cancer networks
Authors Luo Z ![]() , Zheng Y, Zhang W
, Zheng Y, Zhang W
Received 8 September 2017
Accepted for publication 27 December 2017
Published 18 July 2018 Volume 2018:11 Pages 4113—4124
DOI https://doi.org/10.2147/OTT.S151236
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Faris Farassati
Zhiying Luo,1,2 Yi Zheng,3 Wei Zhang1,2
1Department of Clinical Pharmacology, Xiangya Hospital, Central South University, Changsha, Hunan, China; 2Institute of Clinical Pharmacology, Hunan Key Laboratory of Pharmacogenetics, Central South University, Changsha, Hunan, China; 3Department of Pharmacy, Hunan Province Maternal and Child Health, Changsha, Hunan, China
Abstract: MicroRNAs are short regulatory RNAs that posttranscriptionally modulate gene expression and thus play crucial roles in controlling cancer-onset, growth, and progression processes. miR107, a highly conserved microRNA that maps to intron 5 of the PANK1 gene, contributes to the regulation of normal and tumor biological processes. Studies have reported that miR107 has oncogenic or tumor-suppressor functions in different human tumors. The pleiotropic functions of miR107 in various cancers are achieved via its targeting different genes that are involved in tumor proliferation, invasiveness, metastasis, angiogenesis, and chemotherapy-response pathways. The carcinogenicity or cancer-suppressor effects of miR107 occur in a tissue- and cell-specific manner, and the expression level of miR107 can be affected by various factors, including epigenetic and genetic factors, treatment exposure, and daily diet. A comprehensive analysis of the current literature suggests that miR107 functions as a central element in the regulation of cancer networks and can be used as a potential diagnostic and prognostic biomarker and drug target for therapeutic intervention.
Keywords: microRNA, miR107, cancer, suppressor, proto-oncogene
Background
MicroRNAs (miRNAs) are a family of small (20–25 base pair nucleotides long) noncoding RNAs, most of which play important regulatory roles in regulating normal development and physiology.1,2 miRNAs cause these biological effects through sequence-specific binding of a seed sequence to the 3′-end of the untranslated regions (UTRs) of a target mRNA, which causes either mRNA degradation or inhibition of translation and further leads to posttranscriptional downregulation of target-protein expression.3 Thousands of miRNAs have been identified in humans, and it is thought that expression of two-thirds of all genes are regulated by miRNAs.4 Deregulated expression levels of many miRNAs are associated with carcinogenesis, and have been proposed to be novel prognostic and predictive biomarkers.5–7
Tumor cells are distinguished from normal cells by several peculiar biological traits, such as sustaining proliferative signaling, evading growth suppressors, resisting cell death, enabling replicative immortality, angiogenesis, and activating invasion and metastasis.8 The alternation of miRNA expression has a crucial role in cancer, as miRNAs can act either as oncogenes or tumor suppressors in the early disease stages, and they can also influence the progression of invasion and metastasis, as well as predict the clinical outcomes of antitumor therapies.9,10
This review focuses on the pleiotropic functions of miR107 in cancers. miR107 is a highly conserved miRNA that maps to intron 5 of the PANK1 gene. miR107 belongs to the miR15/107 group, whose members regulate genes involved in cell division, metabolism, stress response, and angiogenesis in vertebrate species. All miRNAs in this group share the sequence AGCAGC, starting at either the first or the second nucleotide from the 5′-end of the mature miRNA sequence.11 miR107 is ubiquitously expressed in a range of tissues, with relatively high abundance in the brain. Altered expression of miR107 has been reported in metabolism diseases, such as adipogenesis and diabetes, as well as in neurological diseases and cancers.12
The biological functions of miR107 vary significantly in physiological and pathological processes of various cancers, probably as a result of being targeted to different pathways or genes.13,14 Both clinical and non- or preclinical studies have shown that miR107 is deregulated in several human tumors and its expression level significantly associated with disease staging, metastasis, and treatment outcomes.15,16 We present a systematic review of the complex roles of miR107 in tumor pathogenesis, progression, and prognosis, and of the factors that regulate miR107-expression levels.
Carcinogenic effects of miR107 in various types of cancer
The association between miR107-expression level and carcinogenic effects has been widely researched in various human cancers. However, the functions of miR107 in carcinogenesis are controversial and highly cell-type-dependent, as it acts as either a tumor suppressor or an oncogene by presenting different expression levels in various human tumors. The opposing results found between and within studies are not an uncommon phenomenon in the field of miR107 biology, and these inconsistencies may be due to variations in its mechanism of action.17 To help decipher miR107’s function in specific cancer, Table 1 lists detail of miR107’s function by tumor type.
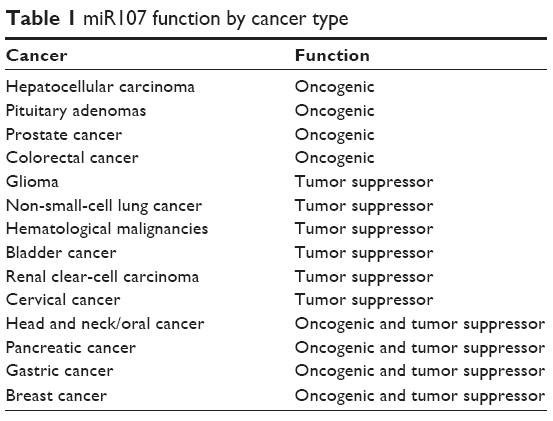 | Table 1 miR107 function by cancer type |
miR107 functions as an oncogenic miRNA
Hepatocellular carcinoma
Hepatocellular carcinoma (HCC) is the most common type of live cancer and the third leading cause of cancer-related death in the world. Previous studies have shown that miRNAs play a fundamental role in HCC, thereby opening a novel avenue for investigating the molecular mechanisms of HCC pathogenesis. The function of miR107 in HCC continues to be researched, as it has been found to have increased expression in HCC tumors compared to paired non-HCC tissue in cancer patients.18,19 Serum miR107 levels are higher in HCC patients compared with healthy controls, and a logit prediction model has shown that the serum levels of miR107, miR92a-3p, miR3126-5p, and α-fetoprotein can serve as sensitive, specific, and noninvasive biomarkers for the diagnosis of HCC, especially in patients at early stages or with low α-fetoprotein levels.20 Moreover, high miR107 expression has been significantly correlated with poor outcome (tumor differentiation and tumor vascular invasion) in HCC by repressing the expression of HMGCS2.21
Pituitary adenomas
Pituitary adenomas are common benign neoplasms that often grow invasively, but very rarely progress to true carcinomas. miR107 expression is significantly upregulated in both sporadic growth hormone-secreting pituitary adenomas and in nonfunctioning pituitary adenomas when compared with normal pituitaries.22 miR107 acts as an oncogenic miRNA in pituitary adenomas, negatively regulating the expression of the pituitary tumor-suppressor gene AIP.
Prostate cancer
Prostate cancer is the most commonly diagnosed male malignancy and the second leading cause of male cancer-related death. A study investigating changes in circulating miRNA levels associated with prostate cancer found that miR107 had higher concentrations in the urine of men with prostate cancer compared with controls.23 There is also evidence that the growth factor granulin is dysregulated via miR107 in prostate cancer, which may provide a potential common therapeutic target.24
Colorectal cancer
Colorectal cancer (CRC) is the fourth most common cancer in the world, with a high mortality rate. Researchers first found that miR107 is a prometastatic miRNA in CRC, in which the expression level of miR107 was positively correlated with the metastatic potential of different CRC cell lines.13 Elevated levels of miR107 have been observed in stage III compared with stage II tumors, and higher expression of miR107 is associated with lymph node metastasis and distant metastasis.13 miR107 exerts its prometastatic function by negatively regulating two metastasis suppressors: DAPK and KLF4. The expression level of miR107 has also been associated with both progression-free survival and overall survival (OS), and overexpression of miR107 is significantly associated with improved objective response to chemotherapy in CRC patients.25 A recent study showed that miR107 expression was aberrantly increased in human CRC tumor tissue and cell lines when compared with colonic control tissues and colon epithelial cells.26
miR107 as a tumor suppressor
Glioma
Glioma is the most common primary malignant tumor in the central nervous system, and is associated with poor prognosis and rapid mortality. Perhaps the strongest evidence of miR107 acting as a tumor suppressor has been found in glioma studies. miR107 is ubiquitously expressed in a range of tissue, with relatively high abundance in the brain. The expression level of miR107 has been shown to be significantly downregulated in human glioma tissue and cell lines when compared with normal brain tissue.15 Low levels of miR107 expression are also significantly associated with advanced pathological features and poor prognosis of human gliomas, such as larger tumor size, lower Karnofsky performance score, and shorter OS and progression-free survival.16
The antitumorigenic roles of miR107 in glioma include the inhibition of cell apoptosis, migration, and invasion. Upregulation of miR107 inhibits the proliferation of gliomas by repressing the expression of SALL4 and activating the FADD–caspase 8–caspase 3/7 cell-apoptosis pathway.15 miR107 exerts its anti-invasion effects through the NOTCH2-signaling pathway.14,27 Overexpression of miR107 inhibits the proliferation of gliomas by arresting the cell cycle at the G0–G1 phase, and also downregulates the expression of CDK6 and NOTCH.28
Non-small-cell lung cancer
Lung cancer (LC) is one of the most common and deadliest cancers in the world, and non-small-cell LC (NSCLC) accounts for 80%–85% of all LC. Two studies have shown that miR107 expression is reduced in lung tumors and LC cell lines compared to normal lungs.29,30 Furthermore, low miR107 expression has been found to be significantly correlated with higher clinicopathological staging stage, regional lymph-node involvement, and tumor differentiation, as well as poor prognosis.29 Recently, it was shown that miR107 suppresses LC-cell proliferation, inhibits metastasis, impedes the cell cycle, and promotes apoptosis by directly targeting EGFR.31
Hematological malignancies
There is limited information on miR107 in hematological cancers. miR107 was initially reported to be upregulated in acute promyelocytic leukemia (APL) cells treated with all-trans retinoic acid (ATRA) compared with untreated cells.32 Further study showed that miR107 was downregulated in APL blasts compared with normal promyelocytes differentiated in vitro from CD34+ progenitors, and its expression level was upregulated after cells had been treated with ATRA.33 Chronic lymphocytic leukemia (CLL) is the most common leukemic disorder, and is characterized by an accumulation of mature B cells in the blood, bone marrow, and secondary lymphoid organs. A recent study assessed global miRNA expression between purified B cells from treatment-naïve CLL patients and healthy controls, and found that miR107 was downregulated in CLL patients.34 Evidence has shown that miR107 acts in CLL by targeting a calcium-channel protein (Cacna2d1) to promote erythroid differentiation.35
Bladder cancer
Bladder cancer is the most prevalent malignant tumor of the urinary system, ranks ninth in cancer incidence around the world, and is the 13th most common cause of death by cancer. Expression of miR107 is decreased in bladder carcinomas in situ when compared with normal bladders from UPK II SV40 mice.36 CircRNAs, a type of noncoding RNA, have been shown to exert major functions in gene regulation. A recent study showed that mechanisms underlying circRNAs in cancer-related pathway might involve miRNA sponge effects via interactions with miRNAs.37 Further results on circRNA in bladder carcinoma found that circTCF25 was significantly more highly expressed in carcinoma tissue compared with control tissue.38 An additional study on the circRNA–miRNA–mRNA axis in bladder cancer showed that overexpression of circTCF25 downregulated miR107, increased CDK6 expression, and promoted proliferation and migration in vitro and in vivo.38
Renal clear-cell carcinoma
Renal cell carcinoma (RCC), which accounts for approximately 85% of all kidney cancers, is the most common type of kidney cancer. Clear-cell RCC occupies the major proportion of RCC cases.39 miR107 plays a tumor-suppressor role in clear-cell RCC by inhibiting cell proliferation and invasiveness.39
Cervical cancer
Cervical cancer is the second most prevalent type of cancer among women worldwide and a complex disease involving numerous oncogenes or the abnormal expression of tumor suppressors. miR107 contributes to the development of cervical cancer by directly targeting CCR5, which acts as a tumor oncogene in cervical cancer.40
Controversy
Head and neck/oral cancer
Head and neck/oral cancer (HNOC) is the sixth most common cancer worldwide, accounting for 4% of cancers in men and 2% of cancers in women.41 miR107 was first found to be highly expressed in nine HNOC cell lines.41,42 However, subsequent studies showed that miR107 is downregulated in oral squamous cell carcinoma (SCC) cell lines and tongue SCC.43–45 Molecular research has found that downregulation of miR107, which targets the PRKCE gene, is a pathogenetic event in HNSCC, and miR107 may thus be a potential anticancer therapeutic for this patient population. Another recent study unraveled the tumor-suppressor role of miR107 in esophageal carcinogenesis, by targeting CDC42.46 HNOC can divide into a severe subtype, and the roles of miR-107 were poorly studied in some kinds of HNOC.47 Based on these in vitro studies, we concluded that further clinical and in vivo studies should be conducted to identify roles of miR107 in HNOC.
Pancreatic cancer
Pancreatic cancer (PCa) is a devastating disease with a poor 5-year survival rate, and local recurrence and systematic metastasis are the major reasons for treatment failure. A study analyzing global miRNA expression in 12 nontumor pancreas and 44 pancreas primary tumors, including 12 insulinomas, 28 nonfunctioning endocrine tumors, and four acinar carcinomas, showed that miR107 is overexpressed in pancreatic endocrine tumors versus normal pancreatic tissue.48 Data from this study suggest that alteration in miR107 expression is related to endocrine and acinar neoplastic transformation, but its expression cannot be used as a biomarker of PCa, due to the very small sample.
In contrast to this result, there is evidence suggesting that miR107 is a tumor-suppressive factor in PCa. BxPC3LN, a lymphatic PCa cell line that exhibits highly metastatic stem-cell-like properties, expresses lower levels of miR107 than the parental BxPC3 cell line.49 Another recent study found that plasma miR107 levels were significantly downregulated in PCa patients compared with healthy volunteers.50 Moreover, low plasma miR107 levels were significantly associated with advanced T and N stage, liver metastasis, and shown to be an independent factor predicting poor prognosis in PCa patients.50 miR107 plasma levels were increased in PCa patients who had undergone curative pancreatectomy compared with preoperative expression levels.50
Gastric cancer
Gastric cancer (GC) is the fourth most common malignancy and the second leading cause of cancer-related death across the world.51 Gastric adenocarcinoma accounts for over 90% of GC cases. miR107 is commonly upregulated in GC and can promote GC-cell migration, invasion, and metastasis, both in vitro and in vivo.52 The first study on miR107 in GC investigated miR107 expression levels in tissues from 50 cases of GC and matched normal tissue by quantitative reverse-transcription polymerase chain reaction.53 The authors discovered that miR107 is frequently upregulated in GC and its overexpression significantly associated with GC metastasis.53 A subsequent study conducted with a larger sample showed that miR107 expression was significantly higher in tumor tissue, and showed significant associations with tumor invasion, lymph-node metastasis, and stage.54 In addition, OS and disease-free survival of patients with high miR107 expression were significantly worse than those of patients with low miR107 expression.54 The overexpression of miR107 in GC has also been evidenced in other studies.55–58 Furthermore, circulating miR107-expression levels are also higher in diffuse-type GC in a mouse model.59
In contrast to previous findings, one study reported that miR107 expression was significantly decreased in GC (determined by comparing miRNA-expression profiles between 80 gastric tumor tissues and matched adjacent nonneoplastic tissues), and the reexpression of miR107 in GC cells significantly decreased proliferation by targeting CDK6.60 In contrast to the mouse model, plasma miR107 levels are not altered in the patients with diffuse-type GC.60
Because of the complex results, it is critical to expand sample sizes and affirm the function of miR107 in the pathogenesis of GC. Also, several questions remain. Dose the expression level of miR107 alter with cancer stage? Why does miR107 expression correlate with response to GC-treatment outcome? Finally, how can miR107 have opposing effects in the same cancer in two different studies? As these questions are answered, researchers will gain a better understanding of the precise function of miR107 in GC.
Breast cancer
Breast cancer is a common highly heterogeneous malignancy and one of the main gynecological cancers worldwide. In breast cancer, hormone-receptor status is one of the most important predictive factors. In one study, miR107 expression was decreased in 30 breast cancer specimens compared with adjacent normal breast tissue,61 and miR107 can inhibit the proliferation and migration of MDA-MB231 cells in a dose- and time-dependent manner. There is also evidence that miR107 is markedly downregulated in both breast cancer cell lines and breast tumors.62
These results are different from previous studies, in which miR107 was shown to be upregulated in breast tumors and cells and elevated miR107 levels associated with more advanced tumor status, increased lymph-node metastasis, and increased metastasis in distant organs.63–65 The complex behavior of miR107 presented in breast cancer are possibly the result of high tumor heterogeneity and hormone responsiveness. Since the expression of miR107 can be downregulated by estrogen treatment, endogenous estrogen levels may significantly influence miR107 levels.66 As breast cancer can be classified into different subgroups based on the state of ER, PR, and HER2, differences in the carcinogenicity of miR107 may also be caused by the statuses of ER, PR, and HER2 in different breast cancer patient groups.
miR107 functions and downstream targets in cancer
Understanding the functions of miRNAs will provide broad prospects for understanding and overcoming tumors. Altered miR107-expression levels in several cancers have led numerous researchers to investigate its tumor-specific functions and identify its targets in various steps of tumor formation and development. The pleiotropic roles and direct targets of miR107 are further summarized in Figure 1 and discussed in detail herein.
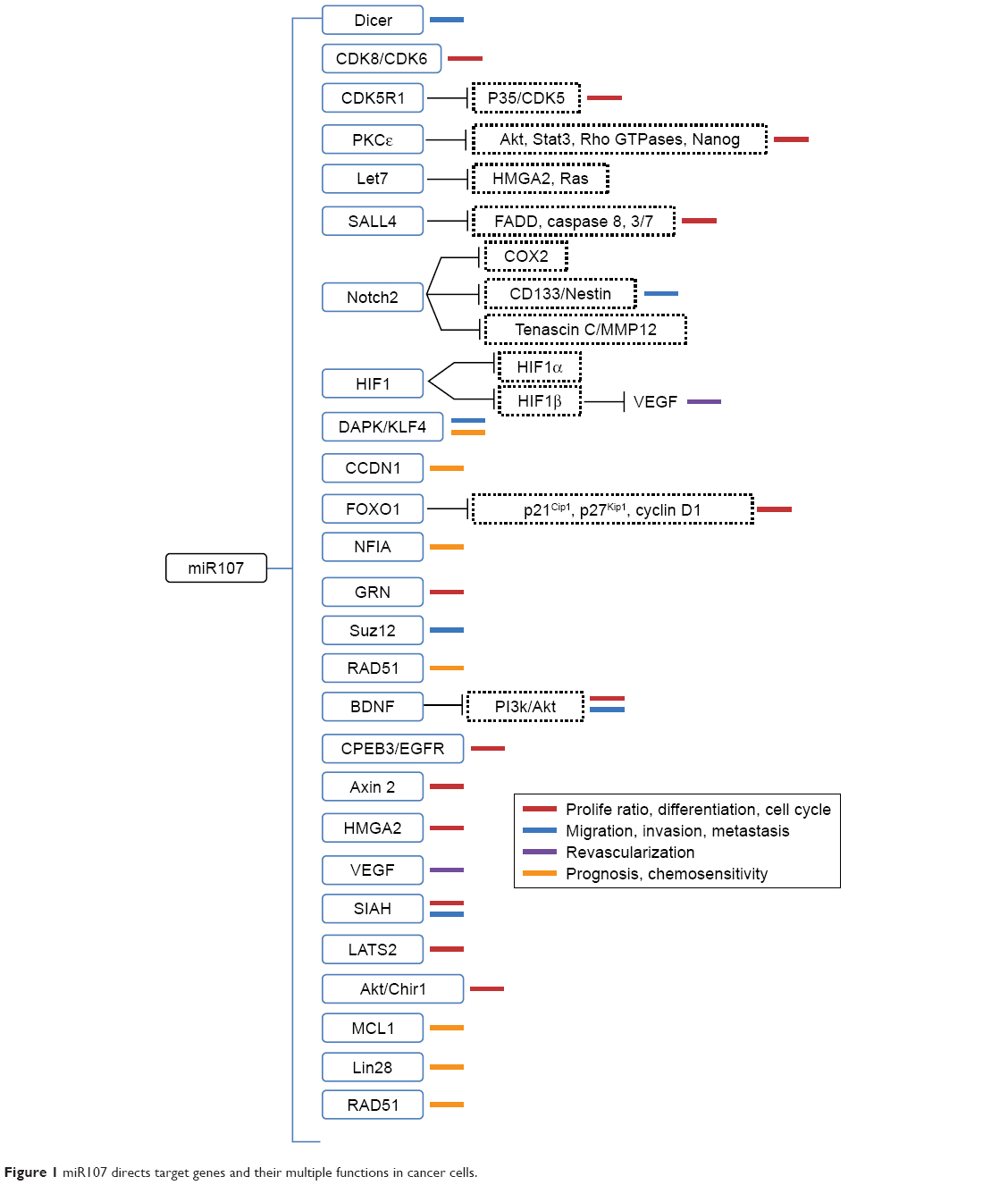 | Figure 1 miR107 directs target genes and their multiple functions in cancer cells. |
Proliferation, cell cycle, and apoptosis
Altered levels of miR107 have been demonstrated to be associated with proliferation, differentiation, and cell-cycle progression in cancer cells. However, the existing data are controversial. miR107 is effective in suppressing the tumorigenicity of HNSCC in vitro and in vivo by inhibiting proliferation, blocking DNA replication and colony formation, and inhibiting cell invasion. It acts by downregulating the expression of PRKCE, which plays critical roles in the signal-transduction pathways involved in proliferation, differentiation, apoptosis, and migration.67 Evidence has also shown that miR107 can suppress cell proliferation in two LC cell lines and induces G1 cell-cycle arrest by downregulating CCNE1 and CDK6.30 Upregulation of miR107 suppresses glioma cell growth through direct targeting of SALL4, CDK6, and NOTCH, leading to the activation of the FADD–caspase 8–caspase-3/7 apoptosis-signaling pathway.15,28 The antitumor effects of miR107 in inhibiting proliferation and inducing cell-cycle arrest in HNOC, LC, and glioma need further verification.
Data from Song et al suggested that miR107 promotes the proliferation of GC cells by posttranscriptionally targeting CDK8, a key protein in the regulation of cell cycle and cell growth, and thus promoting the development and progression of GC.56 In addition, it also promotes cell proliferation by targeting the transcription factor FOXO1, repressing expression of the cell-cycle inhibitors p21Cip1 and p27Kip1 and increasing the expression of cell-cycle regulator cyclin D1.57 Furthermore, miR107 regulates the expression of GRN, a mitogen and growth factor, in multiple human cancers, implying that a therapeutic strategy of restoring the expression of the miR-15/107 gene group may decrease the malignant potential of tumors by attenuating GRN expression.24,68 Li et al found that miR107 expression was decreased in breast cancer, while overexpression of miR107 suppressed MDA-MB231 breast cell proliferation by targeting CDK8, leading cells to arrest at the G0–G1 phase.61
Revascularization
Tumor vascularization, partly driven by hypoxia, plays a critical role in the progression of solid tumors. HIF1 is significant in hypoxic signaling in tumors. miR107 can decrease hypoxia signaling and inhibit the differentiation of endothelial progenitor cells by targeting HIF1B.69 Overexpression of miR107 in HCT116 colon cancer cells suppresses angiogenesis, tumor growth, and tumor VEGF expression in mice. These in vitro results support the hypothesis that miR107 can act as a tumor suppressor by inhibiting tumor vascularization in tumor-cell lines and tissue.
Migration, invasion, and metastasis
Migration is one of the main reasons for failure of tumor therapy and neoplasm recurrence. High miR107 expression can increase the tumorigenic and metastatic potential of breast cancer by inhibiting LET7 and DICER,64,65 and the expression level of miR107 is significantly elevated in triple-negative metastatic breast cancer patients compared with disease-free patients.63 However, there is also evidence that overexpression of miR107 suppresses breast cancer-cell migration by targeting CDK8.61 miR107 has been shown to function as a prometastatic factor in CRC by negatively regulating the expression of two metastasis suppressors: DAPK and KLF4.13
miR107 overexpression is not only an important factor in the pathological process of GC but also related to increased tumor invasion, metastasis, prognosis, and tumor stage.53,54 In addition, circulating miR107 was significantly increased in a mouse model of early- and late-stage diffuse-type GC.59 miR107 functions as a positive metastatic regulator for GC by targeting DICER.53,54 In contrast, Feng et al found that miR107 was decreased in gastric tumor tissues and could inhibit the invasion of GC cells by targeting CDK6.60
High miR107 expression probably serves a metastasis-inhibiting role in PCa.49,50 Additional studies have shown that the inhibitory effects on the migration and invasion exerted by miR107 in glioma and cervical cancer are due to its involvement in the modulation of NOTCH2–TNC–MMP12 and NOTCH2–COX2 signaling through direct targeting of NOTCH214,27 and through activation of the ATR–CHK1 pathway,70 respectively.
Cancer therapy resistance mediators
Vast evidence has shown that the expression level of miR107 is associated with chemotherapeutic sensitivity in different cancers, as shown in Table 2. For example, low miR107 expression significantly correlated with poor prognosis for NSCLC patients, and mimics transfected with miR107 significantly increased cisplatin resistance by targeting the CDK8 protein in NSCLC cells.29,73 miR107 expression can mediate the sensitivity of PARP inhibitors by targeting genes or factors in the DNA-damage-response pathway, such as RAD51.74
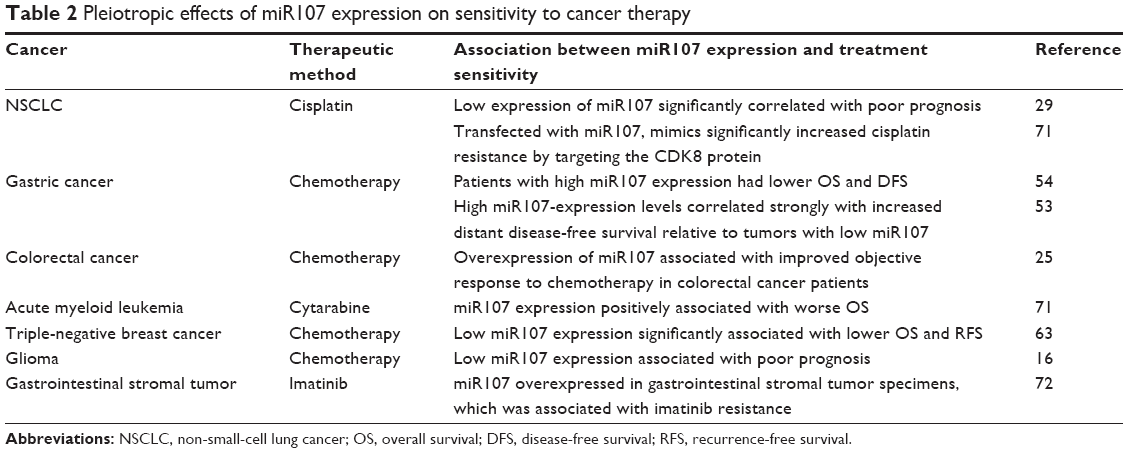 | Table 2 Pleiotropic effects of miR107 expression on sensitivity to cancer therapy |
In the previous section, we summarized that miR107 has oncogenic and tumor-suppressive effects by suppressing both tumor-suppressive mRNAs and oncogenic mRNAs in different cancer types. Due to the opposite roles of miR107 in the pathogeneses of difference cancer types, it also has various influences on treatment outcomes. Most studies on miRNA-expression levels in cancer have viewed positive correlations between miRNA-expression levels and increased survival as evidence that the miRNA is tumor-suppressive. However, this kind of interpretation can be misleading sometimes. It is critical first to validate the association between miR107-expression levels and clinical treatment outcomes if further studies aim to explore the roles of miR107 in the treatment outcome of cancers. Authors should also further illuminate molecular mechanisms via in vitro and animal studies. We are still far from using miR107 as a potential biomarker for personalized treatment strategies and a potential therapeutic target.
Factors influencing miR107 expression in cancer cell lines
The roles of miRNAs in human disease and treatment response have been widely studied. However, which factors influence the expression and stability of miRNAs have barely been considered. In this section, we summarize various factors that can influence miR107 expression, including genetic and epigenetic factors, treatment exposure, and multiple dietary factors.
Epigenetic and genetic factors
Epigenetic factors
Epigenetic mechanisms, including promoter methylation and histone modification, are critical in the regulation of gene expression. Previous studies have underscored the role of epigenetic regulation of noncoding miRNAs in human cancer, typically through the methylation of CpG islands within miRNA primary transcript-promoter regions.75–77 Recently, Lee et al reported that miR107 was a candidate miRNA that may undergo transcriptional silencing through the methylation of a conserved CpG island in its promoter sequence in pancreatic carcinomas.78 The primary transcript of miR107 (located at chromosome 10q23.31) is coexpressed from an intronic segment of PANK1, a coding gene on chromosome 10. We used two commonly used DNA-methylation prediction programs (MethPrimer and CpG Island Searcher) to predict CpG-island distribution 1,000 base pairs upstream from the 5′UTR, and confirmed that a CpG island exists immediately upstream (−442 to −338) of the PANK1-transcription starting site. Interestingly, a recent study showed that the promoter CpG islands of mir107 were not methylated in NSCLC,79 suggesting that epigenetic silencing of miR107 by DNA methylation is cell-specific and any epigenetic alteration of miR107 in other tumors should not be ignored in further studies.
circRNAs are a class of single-stranded closed RNA molecules and produced from precursor mRNA back-splicing of thousands of genes in eukaryotes.80 circRNAs function as an miRNA- or RNA-binding protein sponge and regulate splicing or transcription. circRNAs are involved in cancer pathogenesis.81 Recent research demonstrated that circTCF25 has a miRNA sponge effect that sequesters miR107 and finally increases CDK6 expression and promotes proliferation and migration in bladder carcinoma.38
lncRNAs are transcripts with no protein-coding function that are longer than 200 nucleotides. lncRNAs are emerging as new factors in the cancer paradigm, demonstrating potential roles in both oncogenic and tumor-suppressive pathways.82 lncRNAs can regulate the expression of miRNA by various mechanisms. miR107-expression levels are regulated by lncRNAs. As an lncRNA, HULC is correlated with the development of HCC. HULC sequesters miR107 and promotes tumor angiogenesis in liver cancer through miR107–E2F1–SPHK1 signaling in liver cancer.83 NEAT1 plays an oncogenic role in human laryngeal SCC and regulates CDK6 expression of laryngeal SCC cells, mediated by miR107.84 c-Myc is an inducible gene that is regulated by specific growth signals in a cell-cycle-dependent manner. c-Myc upregulates the expression and strengthens the activity of lncRNA H19, and H19 promotes cell-cycle progression of NSCLC cells by downregulating miR107 and upregulating CDK6 expression.85
Genomic and transcription factors
The tumor suppressor p53 is a transcription factor that responds to stress and maintains cellular control by deregulating stress responses and thereby maintaining cell and tissue integrity. Mutations in P53 or other disruptions in the p53 pathway are associated with tumor growth and angiogenesis.86,87 P53 can activate the transcription of a group of miRNAs, which in turn suppresses the transcription of genes that regulate apoptosis, DNA repair, and cell-cycle progression.88,89
Computer analysis reveals a potential p53-binding site (1,811 base pairs upstream) of the PANK1/miR107-transcription starting site, and several studies have demonstrated that miR107 belongs to the group of miRNAs regulated by p53.89 Chen et al demonstrated that p53 inhibits the proliferation of glioma cells by targeting miR107, and wild-type p53 protein binding to the promoter region of miR107, leading to increased miR107 expression relative to that of glioma cells expressing mutated P53.28 Furthermore, Toll-like receptors modulate the expression of multiple miRNAs. TLR4 can downregulate miR107 expression through the NFKB pathway, and it can further influence the innate immune system by increasing macrophage adhesion via CDK6.90
Treatment exposure
Surgery
Circulating miRNAs have been identified as potentially convenient biomarkers for many diseases, as they can be easily measured without invasive biopsies. miRNAs in circulation are protected from RNase degradation by binding to protein complexes. It has been hypothesized that circulating miRNAs enter the circulation by passive leakage from apoptotic or necrotic cells or by active secretion of microvesicle-free miRNA or miRNA containing microvesicles,91 and many circulating miRNAs have been used to determine the diagnosis and prognosis of human cancer.92,93 Changes in circulating miRNA levels between pre- and postsurgery samples may be used as biomarkers for cancer diagnosis.67–69 However, it has not yet been explained why miRNA levels change between before and after surgery.
Kodahl et al investigated changes in the circulating miRNA profiles of 24 postmenopausal women with ER-positive early-stage breast cancer before surgery and 3 weeks after tumor resection, and found that the postoperative patients had significant higher miR107 levels compared with preoperative samples.91 On the other hand, Cookson et al showed the opposite result, in which circulating miR107 levels were decreased after surgery in a group of 10 breast cancer patients assessed before and after resection.92–94 To explain this difference, we propose several possible hypotheses. First, if surgery can influence miRNA expression, the time at which samples are taken after surgery should be considered carefully. Second, as miR107 levels in breast cancer are associated with tumor status, increased lymph-node metastasis, and increased metastasis in distant organs, the histopathological stage of the cancer before surgery should be taken into account. Third, since estrogen levels can influence miR107 expression, pre- and post-surgery ER status should also be considered carefully.
Drug exposure
miRNAs are not only associated with disease pathogenesis but also with treatment response to many drugs. Interestingly, disease and drug exposure can inversely influence miRNA expression.95–99 As shown by studies on miR107 and drug treatment, miR107-expression level is not only associated with treatment response but can also be regulated by treatment with various drugs.
Evidence has shown that miR107 is upregulated during ATRA and arsenic trioxide treatment in APL cells.32,100,101 Zhou et al showed that the effects of taxol in attenuating migration and invasion are due to the upregulation of miR107 expression in cervical cancer cells, in which miR107 plays an important role in regulating the expression of MCL1.102 In addition to chemotherapy, radiotherapy can also significantly influence miR107 expression. miR107 is overexpressed in mouse renal cortical tissue after administration of 177Lu-octreotate, which is a radiopharmaceutical used for treatment of neuroendocrine cancers.103 However, a recent study found that long-term and excessive use of 900 MHz radiofrequency radiation downregulated rno-miR107 expression in Wistar albino adult male rat brains.104
There is also evidence that chronic morphine treatment can significantly upregulate miR107 and miR103 levels, leading to downregulation of downstream target genes in vivo and in vitro.105 Soluble β-amyloid peptides have specific repression effects on miR107 expression on neuronal.106 Since miR107 plays a critical role in many cancers, we propose the hypothesis that the antineoplastic effects and treatment resistance of some antitumor drugs are partly due to the exceptional expression of miR107 under treatment exposure.
Lipopolysaccharide (LPS) is recognized as the most potent microbial mediator presaging the threat of invasion of Gram-negative bacteria. Blood miR107 expression levels are upregulated in mice exposed to LPS.107 Another study showed that LPS downregulated miR107 expression level in an MyD88- and p65-dependent manner, as well as PANK1α and PPARα, and then upregulated the expression of CDK6.90
Dietary factors
In addition to drug factors, dietary factors (such as fats, carbohydrates, proteins, vitamins, minerals, and trace elements) may also influence cellular processes by regulating endogenous miRNA expression.108 Davidson et al showed that exposure to fish oil, in which the main effective constituent is a chemoprotective n3-polyunsaturated fatty acid, can selectively upregulate the expression of miR107 in rat colons.109 Studies have further demonstrated that dietary lipids modulate the expression of miR107110 and plant-derived polyphenols can regulate the expression of miR103/107 and prevent diet-induced fatty liver disease in hyperlipidemic mice.111 There is also evidence that miR107 expression can be altered by chronic ethanol feeding.112
Conclusion
We can conclude that miR107 has pleiotropic functions in cancer by controlling the expression of genes involved in several cancer cell-signaling networks. miR107 is involved in tumor proliferation, cell-cycle progression, apoptosis, invasiveness, metastasis, angiogenesis, and chemotherapy response.
As miR107 levels are potential biomarkers for the diagnosis and prognosis of multiple cancers,13,16,29 the diagnostic/prognostic value of circulating miR107 should be explored further in various tumors. Almost all clinical studies on the association between miR107 deregulation and cancer onset compared the expression levels between tumor tissue and corresponding healthy tissue. The results of a study by Boominathan supported the view that circulating miR107 can be used as a novel diagnostic biomarker for breast cancer, as patients with newly diagnosed ER-positive breast cancer had higher circulating miR107 levels than healthy controls.89 Whether circulating miR107 can be used as a novel cancer diagnostic and prognostic biomarker requires further investigation in various cancers.
miR107 can significantly influence the treatment outcomes of various cancers. Conversely, exposure to therapy can also affect the expression of miR107 through unknown mechanisms, a similar phenomenon as observed for other miRNAs.90,91 Based on the complex association between miR107 expression and drug exposure, the relationship between miR107-expression levels, cancer susceptibility, and therapy sensitivity must be considered in future studies. Finally, and most importantly, as miRNA-based in vivo therapy attempts have already been successful,113,114 the potential use of miR107 as a therapeutic target merits careful investigation.
Acknowledgments
This research was supported by grants from the National Key Research and Development Program (2016YFC0905000 and 2016YFC0905001), National High Technology Research and Development Program of China 863 Project (2012AA02A518), National Natural Scientific Foundation of China (81522048, 81573511, 81273595, and 81403017), NSFC (81373477), Innovation-Driven Project of Central South University (2016CX024), and Central South University Innovation Foundation for Postgraduates (2015zzts117).
Disclosure
The authors report no conflicts of interest in this work.
References
Shukla GC, Singh J, Barik S. MicroRNAs: processing, maturation, target recognition and regulatory functions. Mol Cell Pharmacol. 2011;3(3):83–92. | ||
Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell. 2009;136(2):215–233. | ||
Chen K, Rajewsky N. The evolution of gene regulation by transcription factors and microRNAs. Nat Rev Genet. 2007;8(2):93–103. | ||
Friedman RC, Farh KK, Burge CB, Bartel DP. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009;19(1):92–105. | ||
Blenkiron C, Goldstein LD, Thorne NP, et al. MicroRNA expression profiling of human breast cancer identifies new markers of tumor subtype. Genome Biol. 2007;8(10):R214. | ||
Rodriguez-Montes JA, Sanchez PM. Role of micro-RNA in colorectal cancer screening. Cir Esp. 2014;92(10):654–658. | ||
Guo Z, Zhao C, Wang Z. MicroRNAs as ideal biomarkers for the diagnosis of lung cancer. Tumour Biol. 2014;35(10):10395–10407. | ||
Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144(5):646–674. | ||
Huang J, Lyu H, Wang J, Liu B. MicroRNA regulation and therapeutic targeting of survivin in cancer. Am J Cancer Res. 2015;5(1):20–31. | ||
Penna E, Orso F, Taverna D. miR-214 as a key hub that controls cancer networks: small player, multiple functions. J Invest Dermatol. 2015;135(4):960–969. | ||
Finnerty JR, Wang WX, Hébert SS, Wilfred BR, Mao G, Nelson PT. The miR-15/107 group of microRNA genes: evolutionary biology, cellular functions, and roles in human diseases. J Mol Biol. 2010;402(3):491–509. | ||
Foley NH, O’Neill LA. miR-107: a Toll-like receptor-regulated miRNA dysregulated in obesity and type II diabetes. J Leukoc Biol. 2012;92(3):521–527. | ||
Chen HY, Lin YM, Chung HC, et al. miR-103/107 promote metastasis of colorectal cancer by targeting the metastasis suppressors DAPK and KLF4. Cancer Res. 2012;72(14):3631–3641. | ||
Chen L, Chen XR, Zhang R, et al. MicroRNA-107 inhibits glioma cell migration and invasion by modulating Notch2 expression. J Neurooncol. 2013;112(1):59–66. | ||
He J, Zhang W, Zhou Q, et al. Low-expression of microRNA-107 inhibits cell apoptosis in glioma by upregulation of SALL4. Int J Biochem Cell Biol. 2013;45(9):1962–1973. | ||
Ji Y, Wei Y, Wang J, Ao Q, Gong K, Zuo H. Decreased expression of microRNA-107 predicts poorer prognosis in glioma. Tumour Biol. 2015;36(6):4461–4466. | ||
Svoronos AA, Engelman DM, Slack FJ. OncomiR or tumor suppressor? The duplicity of microRNAs in cancer. Cancer Res. 2016;76(13):3666–3670. | ||
Wang Y, Chen F, Zhao M, et al. miR-107 suppresses proliferation of hepatoma cells through targeting HMGA2 mRNA 3′UTR. Biochem Biophys Res Commun. 2016;480(3):455–460. | ||
Zhang JJ, Wang CY, Hua L, Yao KH, Chen JT, Hu JH. miR-107 promotes hepatocellular carcinoma cell proliferation by targeting Axin2. Int J Clin Exp Pathol. 2015;8(5):5168–5174. | ||
Zhang Y, Li T, Qiu Y, et al. Serum microRNA panel for early diagnosis of the onset of hepatocellular carcinoma. Medicine (Baltimore). 2017;96(2):e5642. | ||
Su SG, Yang M, Zhang MF, et al. miR-107-mediated decrease of HMGCS2 indicates poor outcomes and promotes cell migration in hepatocellular carcinoma. Int J Biochem Cell Biol. 2017;91(Pt A):53–59. | ||
Trivellin G, Butz H, Delhove J, et al. MicroRNA miR-107 is overexpressed in pituitary adenomas and inhibits the expression of aryl hydrocarbon receptor-interacting protein in vitro. Am J Physiol Endocrinol Metab. 2012;303(6):E708–E719. | ||
Bryant RJ, Pawlowski T, Catto JW, et al. Changes in circulating microRNA levels associated with prostate cancer. Br J Cancer. 2012;106(4):768–774. | ||
Wang WX, Kyprianou N, Wang X, Nelson PT. Dysregulation of the mitogen granulin in human cancer through the miR-15/107 microRNA gene group. Cancer Res. 2010;70(22):9137–9142. | ||
Molina-Pinelo S, Carnero A, Rivera F, et al. miR-107 and miR-99a-3p predict chemotherapy response in patients with advanced colorectal cancer. BMC Cancer. 2014;14:656. | ||
Liu F, Liu S, Ai F, et al. miR-107 promotes proliferation and inhibits apoptosis of colon cancer cells by targeting prostate apoptosis response-4 (Par4). Oncol Res. 2017;25(6):967–974. | ||
Chen L, Chen XR, Chen FF, et al. MicroRNA-107 inhibits U87 glioma stem cells growth and invasion. Cell Mol Neurobiol. 2013;33(5):651–657. | ||
Chen L, Zhang R, Li P, et al. P53-induced microRNA-107 inhibits proliferation of glioma cells and down-regulates the expression of CDK6 and Notch-2. Neurosci Lett. 2013;534:327–332. | ||
Zhong KZ, Chen WW, Hu XY, Jiang AL, Zhao J. Clinicopathological and prognostic significance of microRNA-107 in human non small cell lung cancer. Int J Clin Exp Pathol. 2014;7(7):4545–4551. | ||
Takahashi Y, Forrest AR, Maeno E, Hashimoto T, Daub CO, Yasuda J. miR-107 and miR-185 can induce cell cycle arrest in human non small cell lung cancer cell lines. PLoS One. 2009;4(8):e6677. | ||
Wang P, Liu X, Shao Y, et al. MicroRNA-107-5p suppresses non-small cell lung cancer by directly targeting oncogene epidermal growth factor receptor. Oncotarget. 2017;8(34):57012–57023. | ||
Garzon R, Pichiorri F, Palumbo T, et al. MicroRNA gene expression during retinoic acid-induced differentiation of human acute promyelocytic leukemia. Oncogene. 2007;26(28):4148–4157. | ||
Careccia S, Mainardi S, Pelosi A, et al. A restricted signature of miRNAs distinguishes APL blasts from normal promyelocytes. Oncogene. 2009;28(45):4034–4040. | ||
Pallasch CP, Patz M, Park YJ, et al. miRNA deregulation by epigenetic silencing disrupts suppression of the oncogene PLAG1 in chronic lymphocytic leukemia. Blood. 2009;114(15):3255–3264. | ||
Ruan J, Liu X, Xiong X, et al. miR107 promotes the erythroid differentiation of leukemia cells via the downregulation of Cacna2d1. Mol Med Rep. 2015;11(2):1334–1339. | ||
de la Peña FA, Kanasaki K, Kanasaki M, Tangirala N, Maeda G, Kalluri R. Loss of p53 and acquisition of angiogenic microRNA profile are insufficient to facilitate progression of bladder urothelial carcinoma in situ to invasive carcinoma. J Biol Chem. 2011;286(23):20778–20787. | ||
Chen LL, Yang L. Regulation of circRNA biogenesis. RNA Biol. 2015;12(4):381–388. | ||
Zhong Z, Lv M, Chen J. Screening differential circular RNA expression profiles reveals the regulatory role of circTCF25-miR-103a-3p/miR-107-CDK6 pathway in bladder carcinoma. Sci Rep. 2016;6:30919. | ||
Song N, Ma X, Li H, et al. MicroRNA-107 functions as a candidate tumor suppressor gene in renal clear cell carcinoma involving multiple genes. Urol Oncol. 2015;33(5):201–205. | ||
Che LF, Shao SF, Wang LX. Downregulation of CCR5 inhibits the proliferation and invasion of cervical cancer cells and is regulated by microRNA-107. Exp Ther Med. 2016;11(2):503–509. | ||
Liu X, Chen Z, Yu J, Xia J, Zhou X. MicroRNA profiling and head and neck cancer. Comp Funct Genomics. 2009:837514. | ||
Tran N, Mclean T, Zhang X, et al. MicroRNA expression profiles in head and neck cancer cell lines. Biochem Biophys Res Commun. 2007;358(1):12–17. | ||
Kozaki K, Imoto I, Mogi S, Omura K, Inazawa J. Exploration of tumor-suppressive microRNAs silenced by DNA hypermethylation in oral cancer. Cancer Res. 2008;68(7):2094–2105. | ||
Wong TS, Liu XB, Ho AC, Yuen AP, Ng RW, Wei WI. Identification of pyruvate kinase type M2 as potential oncoprotein in squamous cell carcinoma of tongue through microRNA profiling. Int J Cancer. 2008;123(2):251–257. | ||
Wong TS, Liu XB, Wong BY, Ng RW, Yuen AP, Wei WI. Mature miR-184 as potential oncogenic microRNA of squamous cell carcinoma of tongue. Clin Cancer Res. 2008;14(9):2588–2592. | ||
Sharma P, Saini N, Sharma R. miR-107 functions as a tumor suppressor in human esophageal squamous cell carcinoma and targets Cdc42. Oncol Rep. 2017;37(5):3116–3127. | ||
Chu Y, Zhu H, Lv L, Zhou Y, Huo J. miRNAs in oesophageal squamous cancer. Neth J Med. 2013;71(2):69–75. | ||
Roldo C, Missiaglia E, Hagan JP, et al. MicroRNA expression abnormalities in pancreatic endocrine and acinar tumors are associated with distinctive pathologic features and clinical behavior. J Clin Oncol. 2006;24(29):4677–4684. | ||
Luo G, Long J, Cui X, et al. Highly lymphatic metastatic pancreatic cancer cells possess stem cell-like properties. Int J Oncol. 2013;42(3):979–984. | ||
Imamura T, Komatsu S, Ichikawa D, et al. Depleted tumor suppressor miR-107 in plasma relates to tumor progression and is a novel therapeutic target in pancreatic cancer. Sci Rep. 2017;7(1):5708. | ||
Ma YY, Tao HQ. Microribonucleic acids and gastric cancer. Cancer Sci. 2012;103(4):620–625. | ||
Shrestha S, Hsu SD, Huang WY, et al. A systematic review of microRNA expression profiling studies in human gastric cancer. Cancer Med. 2014;3(4):878–888. | ||
Li X, Zhang Y, Shi Y, et al. MicroRNA-107, an oncogene microRNA that regulates tumour invasion and metastasis by targeting DICER1 in gastric cancer. J Cell Mol Med. 2011;15(9):1887–1895. | ||
Inoue T, Iinuma H, Ogawa E, Inaba T, Fukushima R. Clinicopathological and prognostic significance of microRNA-107 and its relationship to DICER1 mRNA expression in gastric cancer. Oncol Rep. 2012;27(6):1759–1764. | ||
Li X, Zhang Y, Zhang H, et al. miRNA-223 promotes gastric cancer invasion and metastasis by targeting tumor suppressor EPB41L3. Mol Cancer Res. 2011;9(7):824–833. | ||
Song YQ, Ma XH, Ma GL, et al. MicroRNA-107 promotes proliferation of gastric cancer cells by targeting cyclin dependent kinase 8. Diagn Pathol. 2014;9:164. | ||
Li F, Liu B, Gao Y, et al. Upregulation of microRNA-107 induces proliferation in human gastric cancer cells by targeting the transcription factor FOXO1. FEBS Lett. 2014;588(4):538–544. | ||
Zhang M, Wang X, Li W, Cui Y. miR-107 and miR-25 simultaneously target LATS2 and regulate proliferation and invasion of gastric adenocarcinoma (GAC) cells. Biochem Biophys Res Commun. 2015;460(3):806–812. | ||
Rotkrua P, Shimada S, Mogushi K, Akiyama Y, Tanaka H, Yuasa Y. Circulating microRNAs as biomarkers for early detection of diffuse-type gastric cancer using a mouse model. Br J Cancer. 2013;108(4):932–940. | ||
Feng L, Xie Y, Zhang H, Wu Y. miR-107 targets cyclin-dependent kinase 6 expression, induces cell cycle G1 arrest and inhibits invasion in gastric cancer cells. Med Oncol. 2012;29(2):856–863. | ||
Li XY, Luo QF, Wei CK, Li DF, Li J, Fang L. miRNA-107 inhibits proliferation and migration by targeting CDK8 in breast cancer. Int J Clin Exp Med. 2014;7(1):32–40. | ||
Gao B, Hao S, Tian W, et al. MicroRNA-107 is downregulated and having tumor suppressive effect in breast cancer by negatively regulating BDNF. J Gene Med. 2017;19(12):e2932. | ||
Shen S, Sun Q, Liang Z, et al. A prognostic model of triple-negative breast cancer based on miR-27b-3p and node status. PLoS One. 2014;9(6):e100664. | ||
Martello G, Rosato A, Ferrari F, et al. A microRNA targeting dicer for metastasis control. Cell. 2010;141(7):1195–1207. | ||
Chen PS, Su JL, Cha ST, et al. miR-107 promotes tumor progression by targeting the let-7 microRNA in mice and humans. J Clin Invest. 2011;121(9):3442–3455. | ||
Cicatiello L, Mutarelli M, Grober OM, et al. Estrogen receptor α controls a gene network in luminal-like breast cancer cells comprising multiple transcription factors and microRNAs. Am J Pathol. 2010;176(5):2113–2130. | ||
Datta J, Smith A, Lang JC, et al. MicroRNA-107 functions as a candidate tumor-suppressor gene in head and neck squamous cell carcinoma by downregulation of protein kinase Cε. Oncogene. 2012;31(36):4045–4053. | ||
Wang WX, Wilfred BR, Madathil SK, et al. miR-107 regulates granulin/progranulin with implications for traumatic brain injury and neurodegenerative disease. Am J Pathol. 2010;177(1):334–345. | ||
Yamakuchi M, Lotterman CD, Bao C, et al. P53-induced microRNA-107 inhibits HIF-1 and tumor angiogenesis. Proc Natl Acad Sci U S A. 2010;107(14):6334–6339. | ||
Zhou C, Li G, Zhou J, Han N, Liu Z, Yin J. miR-107 activates ATR/Chk1 pathway and suppress cervical cancer invasion by targeting MCL1. PLoS One. 2014;9(11):e111860. | ||
Bhise NS, Chauhan L, Shin M, et al. MicroRNA-mRNA pairs associated with outcome in AML: from in vitro cell-based studies to AML patients. Front Pharmacol. 2015;6:324. | ||
Akcakaya P, Caramuta S, Ahlen J, et al. MicroRNA expression signatures of gastrointestinal stromal tumours: associations with imatinib resistance and patient outcome. Br J Cancer. 2014;111(11):2091–2102. | ||
Zhang Z, Zhang L, Yin ZY, et al. miR-107 regulates cisplatin chemosensitivity of A549 non small cell lung cancer cell line by targeting cyclin dependent kinase 8. Int J Clin Exp Pathol. 2014;7(10):7236–7241. | ||
Huang JW, Wang Y, Dhillon KK, et al. Systematic screen identifies miRNAs that target RAD51 and RAD51D to enhance chemosensitivity. Mol Cancer Res. 2013;11(12):1564–1573. | ||
Han L, Witmer PD, Casey E, Valle D, Sukumar S. DNA methylation regulates microRNA expression. Cancer Biol Ther. 2007;6(8):1284–1288. | ||
Lujambio A, Ropero S, Ballestar E, et al. Genetic unmasking of an epigenetically silenced microRNA in human cancer cells. Cancer Res. 2007;67(4):1424–1429. | ||
Lehmann U, Hasemeier B, Christgen M, et al. Epigenetic inactivation of microRNA gene hsa-mir-9-1 in human breast cancer. J Pathol. 2008;214(1):17–24. | ||
Lee KH, Lotterman C, Karikari C, et al. Epigenetic silencing of microRNA miR-107 regulates cyclin-dependent kinase 6 expression in pancreatic cancer. Pancreatology. 2009;9(3):293–301. | ||
Rykov SV, Khodyrev DS, Pronina IV, Kazubskaya TP, Loginov VI, Braga EA. [Novel miRNA genes methylated in lung tumors]. Genetika. 2013;49(7):896–901. Russian. | ||
Chen LL. The biogenesis and emerging roles of circular RNAs. Nat Rev Mol Cell Biol. 2016;17(4):205–211. | ||
Qu S, Liu Z, Yang X, et al. The emerging functions and roles of circular RNAs in cancer. Cancer Lett. 2017;414:301–309. | ||
Hu X, Sood AK, Dang CV, Zhang L. The role of long noncoding RNAs in cancer: the dark matter matters. Curr Opin Genet Dev. 2017;48:8–15. | ||
Lu Z, Xiao Z, Liu F, et al. Long non-coding RNA HULC promotes tumor angiogenesis in liver cancer by up-regulating sphingosine kinase 1 (SPHK1). Oncotarget. 2016;7(1):241–254. | ||
Wang P, Wu T, Zhou H, et al. Long noncoding RNA NEAT1 promotes laryngeal squamous cell cancer through regulating miR-107/CDK6 pathway. J Exp Clin Cancer Res. 2016;35:22. | ||
Cui J, Mo J, Luo M, et al. c-Myc-activated long non-coding RNA H19 downregulates miR-107 and promotes cell cycle progression of non-small cell lung cancer. Int J Clin Exp Pathol. 2015;8(10):12400–12409. | ||
Yeudall WA. P53 mutation in the genesis of metastasis. Subcell Biochem. 2014;85:105–117. | ||
Li XL, Jones MF, Subramanian M, Lal A. Mutant p53 exerts oncogenic effects through microRNAs and their target gene networks. FEBS Lett. 2014;588(16):2610–2615. | ||
Hermeking H. P53 enters the microRNA world. Cancer Cell. 2007;12(5):414–418. | ||
Boominathan L. The tumor suppressors p53, p63, and p73 are regulators of microRNA processing complex. PLoS One. 2010;5(5):e10615. | ||
Hennessy EJ, Sheedy FJ, Santamaria D, Barbacid M, O’Neill LA. Toll-like receptor-4 (TLR4) down-regulates microRNA-107, increasing macrophage adhesion via cyclin-dependent kinase 6. J Biol Chem. 2011;286(29):25531–25539. | ||
Kodahl AR, Zeuthen P, Binder H, Knoop AS, Ditzel HJ. Alterations in circulating miRNA levels following early-stage estrogen receptor-positive breast cancer resection in post-menopausal women. PLoS One. 2014;9(7):e101950. | ||
Heneghan HM, Miller N, Lowery AJ, Sweeney KJ, Newell J, Kerin MJ. Circulating microRNAs as novel minimally invasive biomarkers for breast cancer. Ann Surg. 2010;251(3):499–505. | ||
Cheng H, Zhang L, Cogdell DE, et al. Circulating plasma miR-141 is a novel biomarker for metastatic colon cancer and predicts poor prognosis. PLoS One. 2011;6(3):e17745. | ||
Cookson VJ, Bentley MA, Hogan BV, et al. Circulating microRNA profiles reflect the presence of breast tumours but not the profiles of microRNAs within the tumours. Cell Oncol (Dordr). 2012;35(4):301–308. | ||
Lewis A, Nijhuis A, Mehta S, et al. Intestinal fibrosis in Crohn’s disease: role of microRNAs as fibrogenic modulators, serum biomarkers, and therapeutic targets. Inflamm Bowel Dis. 2015;21(5):1141–1150. | ||
Taipaleenmaki H, Browne G, Akech J, et al. Targeting of Runx2 by miRNA-135 and miRNA-203 impairs progression of breast cancer and metastatic bone disease. Cancer Res. 2015;75(7):1433–1444. | ||
Szabo G, Satishchandran A. MicroRNAs in alcoholic liver disease. Semin Liver Dis. 2015;35(1):36–42. | ||
Zhang Y, Geng L, Talmon G, Wang J. MicroRNA-520 g confers drug resistance by regulating p21 expression in colorectal cancer. J Biol Chem. 2015;290(10):6215–6225. | ||
Macdonagh L, Gray SG, Finn SP, Cuffe S, O’Byrne KJ, Barr MP. The emerging role of microRNAs in resistance to lung cancer treatments. Cancer Treat Rev. 2015;41(2):160–169. | ||
Garzon R, Pichiorri F, Palumbo T, et al. MicroRNA gene expression during retinoic acid-induced differentiation of human acute promyelocytic leukemia. Oncogene. 2007;26(28):4148–4157. | ||
Wu Y, Li XF, Yang JH, Liao XY, Chen YZ. [MicroRNAs expression profile in acute promyelocytic leukemia cell differentiation induced by all-trans retinoic acid and arsenic trioxide]. Zhonghua Xue Ye Xue Za Zhi. 2012;33(7):546–551. Chinese. | ||
Zhou C, Li G, Zhou J, Han N, Liu Z, Yin J. miR-107 activates ATR/Chk1 pathway and suppress cervical cancer invasion by targeting MCL1. PLoS One. 2014;9(11):e111860. | ||
Schüler E, Parris TZ, Helou K, Forssell-Aronsson E. Distinct microRNA expression profiles in mouse renal cortical tissue after 177Lu-octreotate administration. PLoS One. 2014;9(11):e112645. | ||
Dasdag S, Akdag MZ, Erdal ME, et al. Long term and excessive use of 900 MHz radiofrequency radiation alter microRNA expression in brain. Int J Radiat Biol. 2015;91(4):306–311. | ||
Lu ZG, Xu J, Xu MM, Pasternak GW, Pan YX. Morphine regulates expression of μ-opioid receptor MOR-1A, an intron-retention carboxyl terminal splice variant of the μ-opioid receptor (OPRM1) gene via miR-103/miR-107. Mol Pharmacol. 2014;85(2):368–380. | ||
Li JJ, Dolios G, Wang R, Liao FF. Soluble beta-amyloid peptides, but not insoluble fibrils, have specific effect on neuronal microRNA expression. PLoS One. 2014;9(3):e90770. | ||
Hsieh CH, Rau CS, Jeng JC, et al. Whole blood-derived microRNA signatures in mice exposed to lipopolysaccharides. J Biomed Sci. 2012;19:69. | ||
Wagner AE, Piegholdt S, Ferraro M, Pallauf K, Rimbach G. Food derived microRNAs. Food Funct. 2015;6(3):714–718. | ||
Davidson LA, Wang N, Shah MS, Lupton JR, Ivanov I, Chapkin RS. n-3 Polyunsaturated fatty acids modulate carcinogen-directed non-coding microRNA signatures in rat colon. Carcinogenesis. 2009;30(12):2077–2084. | ||
Daimiel-Ruiz L, Klett-Mingo M, Konstantinidou V, et al. Dietary lipids modulate the expression of miR-107, an miRNA that regulates the circadian system. Mol Nutr Food Res. 2015;59(3):552–565. | ||
Joven J, Espinel E, Rull A, et al. Plant-derived polyphenols regulate expression of miRNA paralogs miR-103/107 and miR-122 and prevent diet-induced fatty liver disease in hyperlipidemic mice. Biochim Biophys Acta. 2012;1820(7):894–899. | ||
Dippold RP, Vadigepalli R, Gonye GE, Patra B, Hoek JB. Chronic ethanol feeding alters miRNA expression dynamics during liver regeneration. Alcohol Clin Exp Res. 2013;37 Suppl 1:E59–E69. | ||
Tominaga N, Yoshioka Y, Ochiya T. A novel platform for cancer therapy using extracellular vesicles. Adv Drug Deliv Rev. 2015;95:50–55. | ||
Li F, Mahato RI. miRNAs as targets for cancer treatment: therapeutics design and delivery. Adv Drug Deliv Rev. 2015;81:v–vi. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.