Back to Journals » Infection and Drug Resistance » Volume 11
Plasmodium knowlesi malaria: current research perspectives
Authors Amir A, Cheong FW ![]() , de Silva JR
, de Silva JR ![]() , Liew JWK
, Liew JWK ![]() , Lau YL
, Lau YL
Received 26 February 2018
Accepted for publication 22 May 2018
Published 10 August 2018 Volume 2018:11 Pages 1145—1155
DOI https://doi.org/10.2147/IDR.S148664
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Professor Suresh Antony
Amirah Amir, Fei Wen Cheong, Jeremy Ryan de Silva, Jonathan Wee Kent Liew, Yee Ling Lau
Department of Parasitology, Faculty of Medicine, University of Malaya, Kuala Lumpur, Malaysia
Abstract: Originally known to cause simian malaria, Plasmodium knowlesi is now known as the fifth human malaria species. Since the publishing of a report that largely focused on human knowlesi cases in Sarawak in 2004, many more human cases have been reported in nearly all of the countries in Southeast Asia and in travelers returning from these countries. The zoonotic nature of this infection hinders malaria elimination efforts. In order to grasp the current perspective of knowlesi malaria, this literature review explores the different aspects of the disease including risk factors, diagnosis, treatment, and molecular and functional studies. Current studies do not provide sufficient data for an effective control program. Therefore, future direction for knowlesi research is highlighted here with a final aim of controlling, if not eliminating, the parasite.
Keywords: zoonosis, epidemiology, diagnosis, treatment, molecular, functional studies
Introduction
Plasmodium knowlesi, originally known to cause simian malaria, is now recognized as the fifth human malarial parasite.1 Human knowlesi infections have been reported in nearly all the countries in Southeast Asia and in travelers returning from these countries.2–4 The cumulative cases of knowlesi malaria in the Southeast Asia region from 2004 to 2015 is 3413 cases with 91.47% of these found in Malaysian Borneo.5
The parasite’s natural reservoir hosts are the long-tailed macaque (Macaca fascicularis),6 pig-tailed macaque (Macaca nemestrina),7 and the banded-leaf monkey (Presbytis melalophos).8 Thus far, several mosquito species belonging to the Leucosphyrus group have been incriminated as vectors for knowlesi malaria, namely, Anopheles hackeri,9 Anopheles latens,10 Anopheles cracens,11 Anopheles balabacensis,12 Anopheles dirus,13 and Anopheles introlatus.14 It is not surprising that the geographical distribution of P. knowlesi is confined to Southeast Asia since it follows the limits of natural distribution of both its natural hosts and vectors.15 To date, the pattern of knowlesi infection in humans does not indicate that they are transmitted via human–mosquito–human.16–18
Several factors have led to an increased reporting of P. knowlesi cases. These include better diagnostic capacity, decreasing human malaria cases which in turn reduces relative immunity, increased awareness to P. knowlesi, and close proximity of humans with natural reservoir hosts or infected vectors due to changes in human land use.5,18 Many studies have been carried out on P. knowlesi ever since it was first described in 1932 by Knowles and Das Gupta.6 This has led to important findings in malarialogy, which include the discovery of antigenic variation in malaria19 and demonstration of an absolute requirement for the Duffy receptor for the invasion of red cells by the parasite.20 Furthermore, since the publishing of report of Singh et al21 that largely focused on naturally acquired P. knowlesi infection in humans, the disease and the parasite have been further characterized, both clinically and molecularly. In order to grasp the current perspective of knowlesi malaria, this literature review explores the different aspects of the disease including risk factors, diagnosis, treatment, molecular and functional studies, mainly focusing on publications from the past five years.
Ecological and individual-level factors associated with infection
Risk factors associated with P. knowlesi infection have been an area of interest over the past few years as researchers have begun to investigate the links between environmental, occupational, sociodemographic, and domestic factors that may contribute to an increased risk of infection. The understanding and identification of these risk factors would be invaluable in designing appropriate and effective public health interventions for knowlesi malaria.
With the advancement of technology, including the use of unmanned aircraft systems or drones, the task of mapping spatial and geographic data to identify environmental factors has been greatly improved.22 Fornace et al studied the association between P. knowlesi incidence and various environmental variables via satellite-based remote sensing data in Kudat and Kota Marudu, Sabah.23 The study found that factors such as more than 65% forest cover in a 2 km radius, higher historical forest loss, and lower elevations were significantly associated with higher incidence of P. knowlesi infection. The authors postulated that the association may be a result of shifts in human settlement as well as changes in the macaque and mosquito habitat due to deforestation and agricultural activities.
Sociodemographic and individual-level factors may also carry increased risk for knowlesi malaria infection. A two-year case–control study was carried out by Grigg et al from 2012 to 2015 in Sabah, Malaysia.24 From a demographic standpoint, participants with ages over 15 years were found to have increased risk of P. knowlesi, whereas other Plasmodium infections were found to be higher in individuals who were younger than 15 years of age. There was also a strong gender bias toward male participants indicating that males over the age of 15 years carried a higher risk of P. knowlesi infection. This suggests that health intervention programs focusing on maternal and child health may be inadequate for P. knowlesi malaria intervention. Other studies have also indicated higher parasitemia and increased P. knowlesi disease severity in association with age.25–27 Higher risk in association with age may relate to occupational risk, as older individuals seek job opportunities in the area. Farming and palm oil plantation work were also identified to be associated with higher risk of infection.
Domestic and peri-domestic factors, such as having open eaves and gaps in the walls, having long grass around the house, sleeping outside of the house, and recent clearing of vegetation, were also associated with a high risk of P. knowlesi infection. A reduction of risk was observed in households with indoor residual spraying being practiced, although bed nets only proved to have a marginal effect on risk.
It was noted that recent presence of monkeys was a strong predictor for risk indicating a high likelihood that monkey to human transmission is still the main transmission pathway rather than human to human transmission. Separately, it was also noted that G6PD deficiency conferred some form of protection against P. knowlesi, with it being associated with decreased risk of P. knowlesi infection, similar to what has been observed in Plasmodium vivax malaria.28 The presence of young sparse forest and rice paddy around the house similarly were significantly associated with reduced P. knowlesi risk.24
A study by Herdiana et al in 2016 also looked at malaria risk factor assessment using both active and passive surveillance in Indonesia.29 Initial screening revealed a total of 19 P. knowlesi cases which was an unexpected discovery in the study. A comparison of P. knowlesi cases to non-cases indicated risk factor findings similar to the study by Grigg et al, with increased risk for P. knowlesi infection: male gender, participants of adult age, and forest exposure or forest-related work, as well as spending time overnight in the forest. When comparing P. knowlesi cases with Plasmodium falciparum and P. vivax infections, it was noted that P. knowlesi cases were more likely linked to forest exposure and peri-domestic factors.
A study by De Silva et al compared the distribution of the different Duffy genotypes among P. knowlesi-infected patients and healthy donors to determine if there was an association between Duffy genotypes and susceptibility to P. knowlesi infection.30 The authors argued that due to the overwhelming homogeneity of the Duffy distribution in the region, assessment of susceptibility was not feasible. However, further studies into the Duffy distribution between P. knowlesi patients in Peninsular and Borneo Malaysia may allow for P. knowlesi susceptibility studies, especially if the Duffy distribution between both regions is markedly different.
Diagnosis
Microscopy is the gold standard for malaria diagnosis, but it has its limitations as ring forms of P. knowlesi resemble P. falciparum and trophozoites and schizonts resemble those of P. malariae and, hence, cannot be reliably differentiated.2,31,32 Most P. knowlesi infections have been identified as infections of more benign P. malariae, which has been associated with failure to diagnose severe malaria and consequent delayed parenteral artesunate, with fatal outcomes.33 In P. knowlesi endemic areas, microscopic diagnosis of P. malariae should be reported and treated as P. knowlesi to reduce P. knowlesi case-fatality rates.33 Limitation of miscroscopic diagnosis has been reported in Sabah, where 21% and 38% of blood films of P. malariae and P. knowlesi were diagnosed as P. falciparum and P. vivax, respectively, by PCR.34 Low parasitemia is fairly common in knowlesi malaria and can cause fever.35 Knowlesi malaria with low parasitemia may not be detected by microscopy. Better methods for the diagnosis of knowlesi malaria such as molecular methods are needed.
To date, there are no immunochromatographic rapid diagnostic tests (RDTs) specifically designed for P. knowlesi detection. Currently available malaria RDTs have been mainly used for detection of P. falciparum and P. vivax infections. OptiMAL-IT, one of the first RDT that could detect P. knowlesi, could mistakenly identify it as P. falciparum, as the monoclonal antibody used to detect P. falciparum histidine rich protein II (HRP-2) cross-reacts with P. knowlesi. Among all the other RDTs, OptiMAL-IT has the highest sensitivity for detecting P. knowlesi, although the percentage is still low (32%–72%).36–38 Other RDTs include BinaxNow that measures antibody response to nonspecific pan malarial aldolase, and Paramax-3 and Entebe Malaria Cassette specific for P. vivax lactate dehydrogenase, which can detect P. knowlesi as P. vivax, is mostly still unable to distinguish P. knowlesi from P. vivax.37,38 Sensitivity of RDTs to knowlesi malaria is poor, particularly at low parasite densities,36.37,39 and they are not currently suitable for clinical use. However, RDTs are so far the only commercially available point-of-care diagnostic tool due to it being user-friendly, rapid, and cost-effective. Other limitations of RDTs include HRP2 deletion, inadequate quality control for RDT, and limited heat stability to be used on the field.40
While microscopy remains the gold standard for malaria diagnosis and RDTs as supplementary test, diagnostic method for clinical purposes must consider the potential inaccuracies of these two; thus, molecular methods such as PCR are needed to further confirm and differentiate the human Plasmodium species. Confirmation of species by PCR will also allow appropriate administration of primaquine to patients with P. vivax or P. ovale infection.36
Molecular methods, such as PCR and real-time PCR, are more accurate in detecting P. knowlesi and are valuable for species identification in cases of mixed malaria infection.41 Nested and real-time PCR based on 18S rRNA gene can detect P. knowlesi in as low as 1 parasite/µL of blood.42,43 However, due to lack of facilities and the lengthy procedure, PCR is not widely used in all endemic areas. Loop-mediated isothermal amplification (LAMP) is another promising molecular diagnostic technique that is applicable for bedside use as it is sensitive, specific, rapid (60–90 min), and easy to use. LAMP does not require costly machine and expertise.44 The Eiken Loopamp™ MALARIA Pan Detection kit targeted at Plasmodium genus can detect all malaria species. It has been shown to be highly sensitive to P. falciparum, P. vivax,45–48 and P. knowlesi.49 However, this method is unable to identify species of infecting Plasmodium and quantification of positive samples is also impossible. Species-specific LAMP assays for all five human malaria species have been developed and were highly sensitive and specific.50 However, due to its high sensitivity nature of LAMP, cross contamination can occur easily.50
Point-of-care molecular tools for rapid malaria diagnosis have also been reported. A lab-on-chip PCR diagnostic platform for malaria, the Accutas system, can detect five species of human malaria with high sensitivity (97.4%) and specificity (93.8%), and it can be performed directly with unprocessed blood (time saving).51 Truelab Uno, based on TaqMan chemistry, has 100% sensitivity and specificity compared to the nested PCR.52 Other point-of-care molecular tools for malaria include Illumigene Malaria LAMP workflow, nanomal, and nucleic acid lateral flow immunoassay DIAGMAL.53 However, these molecular methods were not tested specifically for P. knowlesi.
Although molecular methods are the most efficacious in diagnosing P. knowlesi infection, most of these tests, especially nested PCR, can produce a false-positive P. knowlesi result in P. vivax infections.54 On top of that, it requires expensive equipment and clean facility. An ideal molecular diagnostic test for point-of-care diagnosis of all five human malaria species that is cost-effective and suitable for the resource-limited setting is yet to be developed.
Treatment and drug resistance
Due to its short asexual cycle of 24 h, infection with P. knowlesi can rapidly progress into severe malaria that can be fatal.55 The treatment guidelines developed by the World Health Organization (WHO) for all human malaria including knowlesi malaria is based on four core principles: early diagnosis and prompt effective treatment, rational use of antimalarial agents, combination therapy, and appropriate weight-based dosing.56 The proposed choice of treatment depends mainly on whether the patient presents with uncomplicated or severe infection. In 2015 guidelines for the treatment of malaria, WHO recommends that adults and children with uncomplicated knowlesi malaria be treated with artemisinin-based combination therapy (ACT) (except for pregnant women in their first trimester) or chloroquine. However, the latter is not recommended in areas with chloroquine-resistant Plasmodium species. Whereas in severe malaria, WHO suggests the administration of parenteral artesunate for at least 24 h followed by ACT in both adults and children.56
Since the history of P. knowlesi infection in humans is relatively short compared to other human malaria, only few studies have been done to assess the sensitivity of P. knowlesi toward different antimalarials. A prospective observational study in adults demonstrated that chloroquine together with primaquine was successful in treating uncomplicated knowlesi malaria.57 In a randomized control trial, Grigg et al found both artesunate–mefloquine combination therapy and chloroquine monotherapy to be very effective in treating adults and children with uncomplicated P. knowlesi infection. However, the clearance of parasite and fever was notably faster in those receiving artesunate–mefloquine combination therapy.58 A more recent randomized controlled trial comparing the efficacy of arthemeter–lumefantrine and chloroquine concluded that the former was effective in treating uncomplicated knowlesi malaria with good tolerability and rapid therapeutic response, thus supporting its use as a first-line ACT treatment policy for malaria in Malaysia.27 In severe knowlesi malaria, intravenous artesunate has been shown to be effective with reduction in fatality rate.25,59,60 Additionally, deaths from knowlesi malaria have been linked to the delay in administering intravenous artesunate.61,62
An ex vivo drug sensitivity assay using clinical P. knowlesi isolates revealed that they were sensitive to artemisinins and chloroquine but were less sensitive toward mefloquine.63 Despite no evidence of mefloquine resistance from these studies and its monotherapy being reported to successfully treat uncomplicated knowlesi malaria,64,65 using mefloquine as monotherapy for knowlesi malaria is discouraged.55 This is likely due to concerns arising from treatment failures reported in rhesus monkeys66,67 and in a single human knowlesi case.68
In studying the drug resistance mutations, Tiyagi et al found that the orthologs for known P. falciparum drug resistance genes, namely, chloroquine resistance transporter (Pkcrt) and dihydrofolate reductase (Pkdhfr) of their P. knowlesi isolates were all wild type.69 Grigg et al did a similar study and found moderately diversed Pkdhfr sequence among their P. knowlesi isolates. Nevertheless, there was no evidence of selective drug pressure in humans.70 In addition to crt and dhfr, other orthologs of P. falciparum drug resistance genes, including multidrug resistance-1 (mdr1), dihydropteroate synthase (dhps), and kelch K13, were also looked at, with no signs of positive selection.71 Since only human hosts would be expected to have antimalarial drug exposure and as long as the transmission of P. knowlesi remains zoonotic, the absence of drug selection pressure will make it unlikely to develop antimalarial drug resistance.
Although current state of P. knowlesi treatment shows no resistance toward antimalarials, research on new therapeutic candidates should not be halted as multiple antimalarial-resistant Plasmodium sp. strains are emerging in Southeast Asian countries particularly in Cambodia, Myanmar, and Thailand.72 These therapeutic candidates could aid in resolving antimalarial-resistant problems in other Plasmodium species or could be used as synergists and drug combinations to increase the efficiency of currently available antimalarials.
Molecular epidemiology and diversity
Genetic polymorphism studies not only guide malaria vaccine design73 but also help us understand the population history and structure of a parasite and thus its adaptive potential. Hitherto, proteins of P. knowlesi involved in invasion of erythrocytes are generally under purifying (negative) selection (Table 1), in most cases different from those of P. vivax and P. falciparum.73–83 Negative selection may imply that the genes are under functional constrains. At the same time, the mutations are deleterious to the parasite and the P. knowlesi population is screening for best-adapted variants.75 Long-term population expansion of P. knowlesi in Malaysia has been suggested to be the cause of this selection.73,79,83 There are, however, some differences in natural selection of these proteins in different geographical regions and different parts of the protein.71,78,84 These studies have also demonstrated that most of these genes are genetically diverse and to some extent more polymorphic than their counterparts in P. falciparum and P. vivax. Whole genome sequencing of P. knowlesi clinical isolates from Sarawak revealed them to have much higher nucleotide diversity than P. falciparum and P. vivax.71 Furthermore, the gamma protein region II (γRII)79 and circumsporozoite protein (CSP)78 of different isolates from varying regions and isolation time exhibit different lengths.
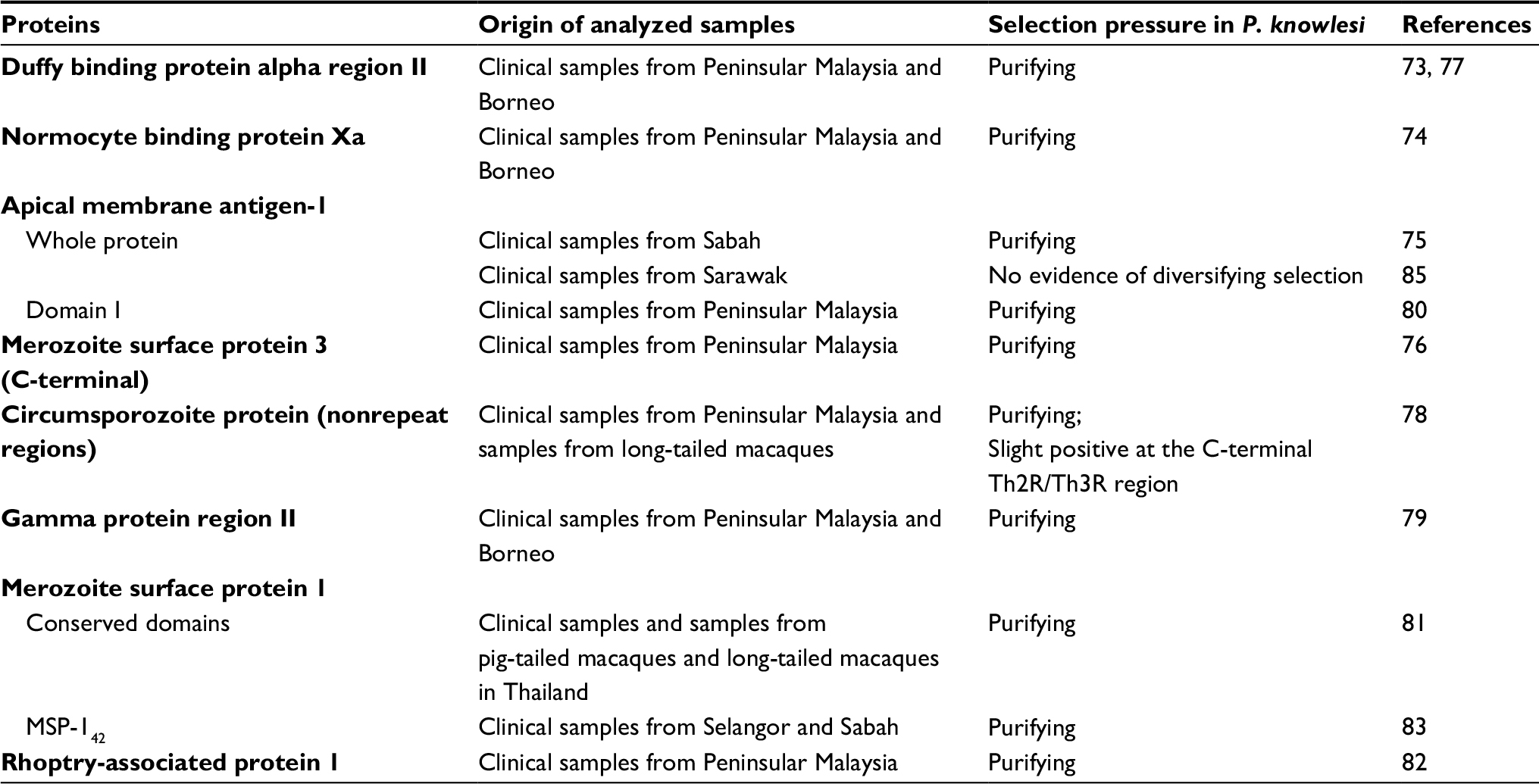 | Table 1 Selection pressure on proteins of P. knowlesi involved in erythrocyte invasion |
Assuming that these sequenced proteins from various sources (human and macaque) are representative of the gene pool in macaques, the high genetic diversity and deviation of selection pressure on these proteins from those of P. vivax and P. falciparum demonstrate intrinsic differences in the immunological targets used by the respective host species (macaques and humans).85 The P. knowlesi proteins’ immunogenic roles have placed them under selection pressure to generate high polymorphism for immune evasion.79 These polymorphisms are probably results of recombination and reproduction cycle of P. knowlesi in multiple hosts.75 Indeed, the Anopheles vector may exert strong evolutionary pressure on vector-related genes of the parasite, which is an evidence of adaptation of the parasite to the vector distribution or vectorial capacity.86
These genetic diversity studies also uncovered the existence of multiple P. knowlesi lineages. It began with the observation of consistent dimorphisms (two distinct groups) in many of the studied proteins originating from humans and macaques. This distinction is not only associated with two separate geographical regions, that is, Peninsular Malaysia/Thailand and Borneo,73,76,77,79,82,84,87–90 but also correlated with macaque host types, that is, with the long-tailed and pig-tailed macaque.91 Further genome sequencing revealed three distinct clusters. The two major sympatric clusters are associated with the long-tailed or pig-tailed macaque, and one cluster consisted of isolates from Peninsular Malaysia.71,86,91 This was also evident from the study on NBPXa protein, with the third cluster found only in Peninsular Malaysia. This led Ahmed et al to postulate that the negative selection on this gene could be the driving force in the evolution and separation of the protein into three clusters.74
The data show that genetic divergence of P. knowlesi resulted not only from long-term geographic isolation (between Peninsular and Bornean isolates) but also through extended, isolated transmission cycles within different macaque hosts but with evidence of recombination when coming in contact with each other.86 These phenomena, clearly driven by evolution, are pertinent to understanding knowlesi infection in humans and macaques. The genomic diversity seen in P. knowlesi is likely caused by geography, as well as the myriad of hosts, vector distribution, and ecological changes.
When studying the adaptation of P. knowlesi in vitro to different host erythrocytes, deletion of 13 genes in clones adapted to cynomolgus blood and deletion of two and duplication of four genes in clones adapted to human blood were observed. In vitro adaptation from macaque erythrocyte to human erythrocyte shows that there is a gradual increase in human erythrocyte invasion efficiency. Hence, P. knowlesi shows immense ability to adapt via improved ability to invade human erythrocytes, thereby increasing virulence and multiplication rates.92
Currently, we know very little of the differences in characteristics and pathogenesis of the three subpopulations of P. knowlesi.91 There are some early evidences that they may exhibit differential pathogenesis.87,88,93 Moreover, the dimorphisms seen in MDR2 and MRP1 (transporter genes related to antimalarial drug resistance)89 should not be overlooked as their phenotypic characteristics and how they react to current antimalarials remains obscure to us. Isolation and investigation of these three subpopulations will be needed to shed more light on these. More importantly, we do not know if human to human transmission happens naturally. The genetic diversity in MSP1 and apical membrane antigen 1 (AMA1) observed among isolates of humans than those from monkeys in Thailand and non-human isolates of Malaysia suggests that it is possible.81,85
Functional studies in P. knowlesi
In 2013, P. knowlesi was reported to be successfully adapted to continuous culture in human erythrocytes by Moon et al94 and Lim et al.95 This breakthrough serves as an important model for P. knowlesi studies especially in validation of vaccine and drug targets. Several P. knowlesi vaccine/therapeutic candidates with their recent findings are discussed in this review.
An important parasite adhesin, the reticulocyte binding-like (RBL) family, is found to be involved in host cell erythrocyte receptor binding to facilitate merozoite invasion. NBPXa is one of the members in P. knowlesi RBL family and is expressed within the microneme organelles.96 With the human erythrocytes-adapted P. knowlesi strain, Moon et al further identified NBPXa as a key mediator for knowlesi infection in human, as this protein is crucial for invasion of human erythrocytes but not cynomolgus erythrocytes.92 By disrupting the NBPXa gene, in vitro multiplicative growth of parasites in human erythrocytes is prevented through impaired merozoite invasion, which support the potential of NBPXa in vaccine development.
Erythrocyte binding-like (EBL) family is another group that mediates interaction with host cell erythrocyte receptors.97 One of the important members in P. knowlesi EBL family, Duffy binding protein α (PkDBPα), interacts with the Duffy antigen receptor for chemokines to invade human erythrocytes.98,99 Genetic polymorphisms in PkDBPα have been postulated to lead to improved parasite binding ability to erythrocytes, thus enhancing the disease severity. This postulation was supported by Lim et al, who in their recent finding showed that two genetically distinct PkDBPα haplotypes from different geographical area demonstrated a different binding activity level to human erythrocytes by using erythrocyte rosetting assay.93
Tryptophan-rich antigens (TRAgs) are involved in rosetting formation and merozoite invasion, thereby contributing to disease severity.100–102 P. knowlesi TRAgs are mostly expressed in its blood stage.103 By using erythrocyte binding assay and parasite growth inhibition assay, Tyagi et al identified three TRAgs in P. knowlesi that were able to bind to human erythrocytes.104 Two of them competed with P. vivax TRAgs for receptors on human erythrocytes. Besides, all of these PkTRAgs were found to inhibit the P. falciparum growth in vitro, further demonstrating the biological significance of this TRAgs-receptor interaction in heterologous parasites by sharing host receptors.
AMA1 and its interaction with rhoptry neck protein 2 (RON2) are essential for merozoite invasion. P. knowlesi AMA1 (PkAMA1) has been tested in vaccine trials, and the results revealed that immunized macaques were protected against infection with controlled parasitemia.105 In 2015, the crystal structure of ectoplasmic region of PkAMA1 and its invasion-inhibitory monoclonal antibody R31C2 were developed.106 R31C2 binds to the hydrophobic groove and interferes with the exposition of complete binding site on AMA1. This monoclonal antibody is found to be cross-strain reactive as it targets on a non-polymorphic epitope. Besides, no polymorphism near RON2-binding site of AMA1 was detected. The lack of polymorphism suggests that P. knowlesi has not developed a mechanism to evade the host’s humoral response.
Proteins that demonstrated different mechanisms in parasite metabolic pathways from those found in human host are also of particular interest. Garg et al have carried out a study on P. knowlesi enzyme phosphoethanolamine methyltransferases (PkPMTs), which regulates the synthesis of phosphatidylcholine. Structural, biochemical properties, and inhibition profile of PkPMTs were evaluated using X-ray crystallography, enzyme kinetics, and mutant gene expression studies. PkPMTs enzyme activity could be inhibited by amodiaquine, chloroquine, and NCI compound NSC158011, whereas the yeasts carrying mutant PkPMT genes are unable to carry out phosphatidylcholine biosynthesis from phosphoethanolamine.107 Disruption of phosphatidylcholine synthesis in Plasmodium would lead to failure of intact Plasmodium membrane formation and thus inhibit parasite proliferation. These findings suggest that PkPMTs are suitable targets for chemotherapy, and the data from this study could further contribute to a better design of more selective antimalarial drugs against P. knowlesi.
In infected erythrocytes, P. falciparum skeleton-binding protein 1 (PfSBP1) is required to transport the erythrocyte surface ligand, erythrocyte membrane protein 1, to the surface of erythrocyte for mediating the surface ligands exposition.108 Its ortholog in P. knowlesi, PkSBP1, has been recently localized to the “Sinton and Mulligan” stipplings in the cytoplasm of infected erythrocytes.109 By using immunofluorescence assay and immunoelectron microscopy, transgenic P. knowlesi expressing the tagged recombinant PkSBP1 demonstrated an analogous trafficking pattern as in P. falciparum, supporting the hypothesis that Plasmodium has evolutionarily conserved protein export pathways. These pathway-related proteins could be used as malaria intervention targets as the disruption of the protein export structures would then interfere with the exposition of virulent ligands at the surface of infected erythrocytes.
Hitherto, P. knowlesi vaccine target antigens have not been evaluated in human trial, as the use of non-human primate models involves serious ethical concerns. Despite higher physiological similarity between human and primate compared to other animal models, vaccine studies of P. knowlesi in primate models are unable to completely represent the efficacy and safety of the targets in human due to the differences in host immunity regulations and pathogenic responses toward malaria. Taking into account that majority of P. falciparum single antigen was unable to raise specific antibodies up to protective levels in human vaccine trials,110–112 the importance of multi-antigens combination and incorporation of immunostimulants should be considered in P. knowlesi vaccine development.
Future directions and conclusion
There remains a clear risk of continuous P. knowlesi infections, especially when land-use change and human behavior may have driven behavioral changes in the reservoir and vector, enabling closer contact and higher chances of spillover to the human population.113 The current literature on risk factors associated with P. knowlesi has identified significant key risks that will prove helpful for the development of effective intervention programs and has highlighted the important environmental factors that intermingle with individual risk factors to P. knowlesi transmission. Further research into the way these risk factors associate with each other and further spatial mapping and geographic monitoring of P. knowlesi hotspots will prove to be an essential part of P. knowlesi research as a whole going forward.
WHO (2018) recommends to perform parasite-specific diagnosis in malaria-suspected patients before treatment to prevent unnecessary drug wastage and to halt antimalarial drug resistance.114 Therefore, efforts should be invested to develop a point-of-care tool that will be able to detect and differentiate all human Plasmodium species to be used in limited-resources environment.
In-depth population genetic studies for both human and primate isolates are needed to shed light on possible human-to-human transmission of P. knowlesi and more importantly for us to understand the disease epidemiology and to guide knowlesi infection control. Knowlesi malaria cases have been showing an upward trend in Malaysian Borneo for the past decade and is also forecasted to increase.34,115,116 Experimental transmission of P. knowlesi from monkey to man, from man to man, and from man back to monkey has been shown by Chin et al.117 Besides, gametocytes can be found in infected patients,118,119 and most vectors of P. knowlesi (e.g. An. dirus, An. balabacensis, and An. cracens) are also vectors of human malarias.120 Although there is no direct evidence of natural human-to-human transmission of P. knowlesi, the likelihood of it occurring cannot be ignored until proven otherwise. Experimental studies on the vectors that transmit this parasite and knowing the distribution of vectors harboring this parasite will also lend credence to solving this issue.
Rapid development in genome editing tools such as clustered regularly interspaced short palindromic repeats highly increase the efficiency to engineer the genome.121,122 Transfections for labeling, knockout/knockdown, and gene editing can be carried out in in vitro P. knowlesi parasites to understand its pathogenesis, drug susceptibility, and gene functions.94 These in vitro culture-transfection systems coupled with suitable animal models could overcome the bottleneck that is hampering the translational malaria research.
The WHO Global Malaria Program has appropriate interventions focused on the malaria caused by P. falciparum and P. vivax, which cause major global morbidity and mortality. However, efforts for malaria control and elimination in certain Southeast Asia countries, such as Malaysia, are greatly impeded by the uncontrollable passage of P. knowlesi in macaque populations, which could lead to human malaria outbreak via zoonotic transmission. In conclusion, different strategies and interventions are needed to prevent P. knowlesi transmission from macaques to human. Effective vaccines are in need to control, if not eliminate the parasite.
Acknowledgments
AA was supported by BKP grant, University of Malaya (BK013-2016). FWC was supported by BKP grant, University of Malaya (BK005-2017). JRDS was supported by the Fogarty International Center and the Office of AIDS Research at the National Institutes of Health under the Global Health Equity Scholars Consortium at Yale University (D43TW010540).
Disclosure
The authors report no conflicts of interest in this work.
References
White NJ. Plasmodium knowlesi: the fifth human malaria parasite. Clin Infect Dis. 2008;46(2):172–173. | ||
Singh B, Daneshvar C. Human infections and detection of Plasmodium knowlesi. Clin Microbiol Rev. 2013;26(2):165–184. | ||
Ministry of Health. Annual Report Ministry of Health 2012. 2012 ed. Malaysia: Ministry of Health; 2012:69–70, 95–96. | ||
Yusof R, Lau YL, Mahmud R, et al. High proportion of knowlesi malaria in recent malaria cases in Malaysia. Malar J. 2014;13(1):168. | ||
Abeysinghe R. Outcomes from the evidence review group on Plasmodium knowlesi. In: Presented at the Malaria Policy Advisory Committee Meeting; 22–24 March; 2017; Geneva. | ||
Knowles R, Das Gupta BM. A study of monkey-malaria, and its experimental transmission to man (a preliminary report). Indian Med Gaz. 1932;67(6):301–320. | ||
Eyles D, Laing A, Dobrovolny C. The malaria parasites of the pig-tailed macaque, Macaca nemestrina (Linnaeus), in Malaya. Indian J Malariol. 1962;16:285–298. | ||
Eyles DE, Laing AB, Warren M, Sandosham AA, Wharton R. Malaria parasites of the Malayan leaf monkeys of the genus Presbytis. Medl J Malaya. 1962;17:85–86. | ||
Wharton R, Eyles DE. Anopheles hackeri, a vector of Plasmodium knowlesi in Malaya. Science. 1961;134(3474):279–2280. | ||
Vythilingam I, Tan CH, Asmad M, Chan ST, Lee KS, Singh B. Natural transmission of Plasmodium knowlesi to humans by Anopheles latens in Sarawak, Malaysia. Trans R Soc Trop Med Hyg. 2006;100(11):1087–1088. | ||
Jiram AI, Vythilingam I, NoorAzian YM, Yusof YM, Azahari AH, Fong MY. Entomologic investigation of Plasmodium knowlesi vectors in Kuala Lipis, Pahang, Malaysia. Malar J. 2012;11:213. | ||
Wong ML, Chua TH, Leong CS, et al. Seasonal and spatial dynamics of the primary vector of Plasmodium knowlesi within a major transmission focus in Sabah, Malaysia. PLoS Negl Trop Dis. 2015;9(10):e0004135. | ||
Marchand RP, Culleton R, Maeno Y, Quang NT, Nakazawa S. Co-infections of Plasmodium knowlesi, P. falciparum, and P. vivax among humans and Anopheles dirus mosquitoes, Southern Vietnam. Emerg Infect Dis. 2011;17(7):1232–1239. | ||
Vythilingam I, Lim YA, Venugopalan B, et al. Plasmodium knowlesi malaria an emerging public health problem in Hulu Selangor, Selangor, Malaysia (2009–2013): epidemiologic and entomologic analysis. Parasit Vectors. 2014;7:436. | ||
Singh B, Daneshvar C. Human infections and detection of Plasmodium knowlesi. Clin Microbiol Rev. 2013;26(2):165–184. | ||
Kantele A, Jokiranta TS. Review of cases with the emerging fifth human malaria parasite, Plasmodium knowlesi. Clin Infect Dis. 2011;52(11):1356–1362. | ||
Lee KS, Divis PC, Zakaria SK, et al. Plasmodium knowlesi: reservoir hosts and tracking the emergence in humans and macaques. PLoS Pathog. 2011;7(4):e1002015. | ||
Imai N, White MT, Ghani AC, Drakeley CJ. Transmission and control of Plasmodium knowlesi: a mathematical modelling study. PLoS Negl Trop Dis. 2014;8(7):e2978. | ||
Brown KN, Brown IN. Immunity to malaria: antigenic variation in chronic infections of Plasmodium knowlesi. Nature. 1965;208(5017):1286–1288. | ||
Miller LH, Aikawa M, Dvorak JA. Malaria (Plasmodium knowlesi) merozoites: immunity and the surface coat. J Immunol. 1975;114(4):1237–1242. | ||
Singh B, Sung LK, Matusop A, et al. A large focus of naturally acquired Plasmodium knowlesi infections in human beings. Lancet. 2004;363(9414):1017–1024. | ||
Fornace KM, Drakeley CJ, William T, Espino F, Cox J. Mapping infectious disease landscapes: unmanned aerial vehicles and epidemiology. Trends Parasitol. 2014;30(11):514–519. | ||
Fornace KM, Abidin TR, Alexander N, et al. Association between landscape factors and spatial patterns of Plasmodium knowlesi infections in Sabah, Malaysia. Emerg Infect Dis. 2016;22(2):201–208. | ||
Grigg MJ, Cox J, William T, et al. Individual-level factors associated with the risk of acquiring human Plasmodium knowlesi malaria in Malaysia: a case-control study. Lancet Planet Health. 2017;1(3):e97–e104. | ||
Barber BE, William T, Grigg MJ, et al. A prospective comparative study of knowlesi, falciparum, and vivax malaria in Sabah, Malaysia: high proportion with severe disease from Plasmodium knowlesi and Plasmodium vivax but no mortality with early referral and artesunate therapy. Clin Infect Dis. 2013;56(3):383–397. | ||
Barber BE, Grigg MJ, William T, et al. Effects of aging on parasite biomass, inflammation, endothelial activation, microvascular dysfunction and disease severity in Plasmodium knowlesi and Plasmodium falciparum malaria. J Infect Dis. 2017;215(12):1908–1917. | ||
Grigg MJ, William T, Barber BE, et al. Artemether-lumefantrine versus chloroquine for the treatment of uncomplicated Plasmodium knowlesi malaria: an open-label randomized controlled trial CAN KNOW. Clin Infect Dis. 2018;66(2):229–236. | ||
Leslie T, Briceno M, Mayan I, et al. The impact of phenotypic and genotypic G6PD deficiency on risk of Plasmodium vivax infection: a case-control study amongst Afghan refugees in Pakistan. PLoS Med. 2010;7(5):e1000283. | ||
Herdiana H, Cotter C, Coutrier FN, et al. Malaria risk factor assessment using active and passive surveillance data from Aceh Besar, Indonesia, a low endemic, malaria elimination setting with Plasmodium knowlesi, Plasmodium vivax, and Plasmodium falciparum. Malar J. 2016;15:468. | ||
De Silva JR, Lau YL, Fong MY. Genotyping of the Duffy blood group among Plasmodium knowlesi-infected patients in Malaysia. PLoS One. 2014;9(9):e108951. | ||
Barber BE, William T, Grigg MJ, Yeo TW, Anstey NM. Limitations of microscopy to differentiate Plasmodium species in a region co-endemic for Plasmodium falciparum, Plasmodium vivax and Plasmodium knowlesi. Malar J. 2013;12:8. | ||
Lee WC, Chin PW, Lau YL, et al. Hyperparasitaemic human Plasmodium knowlesi infection with atypical morphology in peninsular Malaysia. Malar J. 2013;12:88. | ||
World Health Organization. Expert Consultation on Plasmodium knowlesi Malaria to Guide Malaria Elimination Strategies, Kota Kinabalu, Malaysia, 1–2 March 2017: Meeting Report. Manila: WHO Regional Office for the Western Pacific; 2017. | ||
William T, Jelip J, Menon J, et al. Changing epidemiology of malaria in Sabah, Malaysia: increasing incidence of Plasmodium knowlesi. Malar J. 2014;13:390. | ||
Grigg MJ, William T, Barber BE, et al. Age-related clinical spectrum of Plasmodium knowlesi malaria and predictors of severity. Clin Infect Dis. 2018;6(12):19-21. | ||
Barber BE, William T, Grigg MJ, Piera K, Yeo TW, Anstey NM. Evaluation of the sensitivity of a pLDH-based and an aldolase-based rapid diagnostic test for diagnosis of uncomplicated and severe malaria caused by PCR-confirmed Plasmodium knowlesi, Plasmodium falciparum, and Plasmodium vivax. J Clin Microbiol. 2013;51:1118–1123. | ||
Foster D, Cox-Singh J, Mohamad DS, Krishna S, Chin PP, Singh B. Evaluation of three rapid diagnostic tests for the detection of human infections with Plasmodium knowlesi. Malar J. 2014;13:60. | ||
Kawai S, Hirai M, Haruki K, Tanabe K, Chigusa Y. Cross-reactivity in rapid diagnostic tests between human malaria and zoonotic simian malaria parasite Plasmodium knowlesi infections. Parasitol Int. 2009;58(3):300–302. | ||
Grigg MJ, William T, Barber BE, et al. Combining parasite lactate dehydrogenase-based and histidine-rich protein 2-based rapid tests to improve specificity for diagnosis of malaria due to Plasmodium knowlesi and other Plasmodium species in Sabah, Malaysia. J Clin Microbiol. 2014;52(6):2053–2060. | ||
McMorrow ML, Aidoo M, Kachur SP. Malaria rapid diagnostic tests in elimination settings – can they find the last parasite? Clin Microbiol Infect. 2011;17(11):1624–1631. | ||
Lau YL, Lai MY, Anthony CN, et al. Comparison of three molecular methods for the detection and speciation of five human Plasmodium species. Am J Trop Med Hyg. 2015;92:28–33. | ||
Snounou G, Viriyakosol S, Zhu XP, et al. High sensitivity of detection of human malaria parasites by the use of nested polymerase chain reaction. Mol Biochem Parasitol. 1993;61(2):315–320. | ||
Reller ME, Chen WH, Dalton J, Lichay MA, Dumler JS. Multiplex 5′ nuclease quantitative real-time PCR for clinical diagnosis of malaria and species-level identification and epidemiologic evaluation of malaria-causing parasites, including Plasmodium knowlesi. J Clin Microbiol. 2013;51(9):2931–2938. | ||
Britton S, Cheng Q, Grigg MJ, William T, Anstey NM, McCarthy JS. A sensitive, colorimetric, high-throughput loop-mediated isothermal amplification assay for the detection of Plasmodium knowlesi. Am J Trop Med Hyg. 2016;95(1):120–122. | ||
Polley S, Gonzalez I, Mohamed D, et al. Clinical evaluation of a loop-mediated amplification kit for diagnosis of imported malaria. J Infect Dis. 2013;208:637–644. | ||
Vallejo A, Martínez N, González IJ, Arévalo-Herrera M, Herrera S. Evaluation of the loop mediated isothermal DNA amplification (LAMP) kit for malaria diagnosis in Plasmodium vivax endemic settings of Colombia. PLoS Negl Trop Dis. 2015;9:e3453. | ||
Aydin-Schmidt B, Weiping X, Gonzalez I, et al. Loop mediated isothermal amplification (LAMP) accurately detects malaria DNA from filter paper blood samples of low density parasitaemias. PLoS One. 2014;9:e103905. | ||
Ponce C, Kaczorowski F, Perpoint T, et al. Diagnostic accuracy of loop-mediated isothermal amplification (LAMP) for screening patients with imported malaria in a non-endemic setting. Parasite. 2017;24:53. | ||
Piera KA, Aziz A, William T, et al. Detection of Plasmodium knowlesi, Plasmodium falciparum and Plasmodium vivax using loop-mediated isothermal amplification (LAMP) in a co-endemic area in Malaysia. Malar J. 2017;16:29. | ||
Lau YL, Lai MY, Fong MY, Jelip J, Mahmud R. Short report: loop-mediated isothermal amplification assay for identification of five human Plasmodium species in Malaysia. Am J Trop Med Hyg. 2016;94(2):336–339. | ||
Taylor BJ, Howell A, Martin KA, et al. A lab-on-chip for malaria diagnosis and surveillance. Malar J. 2014;13:179. | ||
Nair CB, Manjula J, Subramani PA, et al. Differential diagnosis of malaria on Truelab Uno®, a portable, real-time, microPCR device for point-of-care applications. PLoS One. 2016;11(1): e0146961. | ||
UNITAID. Malaria Diagnostics Technology and Market Landscape. 3rd ed. Geneva: UNITAID; 2016. | ||
Sulistyaningsih E, Fitri LE, Löscher T, Berens-Riha N. Diagnostic difficulties with Plasmodium knowlesi infection in humans. Emerg Infect Dis. 2010;16(6):1033. | ||
Cox-Singh J, Davis TM, Lee KS, et al. Plasmodium knowlesi malaria in humans is widely distributed and potentially life threatening. Clin Infect Dis. 2008;46(2):165–171. | ||
World Health Organization.Guidelines for the Treatment of Malaria. 3rd ed. Geneva: World Health Organization; 2015. | ||
Daneshvar C, Davis TM, Cox-Singh J, et al. Clinical and parasitological response to oral chloroquine and primaquine in uncomplicated human Plasmodium knowlesi infections. Malar J. 2010;9(1):238. | ||
Grigg MJ, William T, Menon J, et al. Artesunate–mefloquine versus chloroquine for treatment of uncomplicated Plasmodium knowlesi malaria in Malaysia (ACT KNOW): an open-label, randomised controlled trial. Lancet Infect Dis. 2016;16(2):180–188. | ||
William T, Menon J, Rajahram G, et al. Severe Plasmodium knowlesi malaria in a tertiary care hospital, Sabah, Malaysia. Emerg Infect Dis. 2011;17(7):1248. | ||
Barber BE, Grigg MJ, William T, Yeo TW, Anstey NM. The treatment of Plasmodium knowlesi malaria. Trends Parasitol. 2017;33(3):242–253. | ||
Rajahram GS, Barber BE, William T, Menon J, Anstey NM, Yeo TW. Deaths due to Plasmodium knowlesi malaria in Sabah, Malaysia: association with reporting as Plasmodium malariae and delayed parenteral artesunate. Malar J. 2012;11(1):284. | ||
Rajahram GS, Barber BE, William T, et al. Falling Plasmodium knowlesi malaria death rate among adults despite rising incidence, Sabah, Malaysia, 2010–2014. Emerg Infect Dis. 2016;22(1):41. | ||
Fatih FA, Staines HM, Siner A, et al. Susceptibility of human Plasmodium knowlesi infections to anti-malarials. Malar J. 2013;12(1):425. | ||
Bronner U, Divis PC, Färnert A, Singh B. Swedish traveller with Plasmodium knowlesi malaria after visiting Malaysian Borneo. Malar J. 2009;8(1):15. | ||
Tanizaki R, Ujiie M, Kato Y, et al. First case of Plasmodium knowlesi infection in a Japanese traveller returning from Malaysia. Malar J. 2013;12(1):128. | ||
Singh PP, Dutta GP. Antimalarial activity of mefloquine and chloroquine against blood induced Plasmodium knowlesi infection in rhesus monkeys. Indian J Med Res. 1981;73:23. | ||
Tripathi R, Awasthi A, Dutta GP. Mefloquine resistance reversal action of ketoconazole – a cytochrome P 450 inhibitor, against mefloquine-resistant malaria. Parasitology. 2005;130(5):475–479. | ||
Lau YL, Tan LH, Chin LC, Fong MY, Noraishah MA, Rohela M. Plasmodium knowlesi reinfection in human. Emerg Infect Dis. 2011;17(7):1314. | ||
Tyagi RK, Das MK, Singh SS, Sharma YD. Discordance in drug resistance-associated mutation patterns in marker genes of Plasmodium falciparum and Plasmodium knowlesi during coinfections. J Antimicrob Chemother. 2013;68(5):1081–1088. | ||
Grigg MJ, Barber BE, Marfurt J, et al. Dihydrofolate-reductase mutations in Plasmodium knowlesi appear unrelated to selective drug pressure from putative human-to-human transmission in Sabah, Malaysia. PLoS One. 2016;11(3):e0149519. | ||
Assefa S, Lim C, Preston MD, et al. Population genomic structure and adaptation in the zoonotic malaria parasite Plasmodium knowlesi. Proc Natl Acad Sci USA. 2015;112(42):13027–13032. | ||
World Health Organization. Status Report on Artemisinin and ACT Resistance. Geneva: World Health Organization; 2017. | ||
Fong MY, Lau YL, Chang PY, Anthony CN. Genetic diversity, haplotypes and allele groups of Duffy binding protein (PkDBPαII) of Plasmodium knowlesi clinical isolates from Peninsular Malaysia. Parasit Vectors. 2014;7:161–161. | ||
Ahmed MA, Fong MY, Lau YL, Yusof R. Clustering and genetic differentiation of the normocyte binding protein (nbpxa) of Plasmodium knowlesi clinical isolates from Peninsular Malaysia and Malaysia Borneo. Malar J. 2016;15:241. | ||
Chua CY, Lee PC, Lau TY. Analysis of polymorphisms and selective pressures on ama1 gene in Plasmodium knowlesi isolates from Sabah, Malaysia. J Genet. 2017;96(4):653–663. | ||
De Silva JR, Lau YL, Fong MY. Genetic clustering and polymorphism of the merozoite surface protein-3 of Plasmodium knowlesi clinical isolates from Peninsular Malaysia. Parasit Vectors. 2017;10(1):2. | ||
Fong MY, Rashdi SA, Yusof R, Lau YL. Distinct genetic difference between the Duffy binding protein (PkDBPαII) of Plasmodium knowlesi clinical isolates from North Borneo and Peninsular Malaysia. Malar J. 2015;14(1):91. | ||
Fong MY, Ahmed MA, Wong SS, Lau YL, Sitam S. Genetic diversity and natural selection of the Plasmodium knowlesi circumsporozoite protein nonrepeat regions. PLoS One. 2015;10(9):e0137734. | ||
Fong MY, Rashdi SA, Yusof R, Lau YL. Genetic diversity, natural selection and haplotype grouping of Plasmodium knowlesi gamma protein region II (PkgammaRII): comparison with the Duffy binding protein (PkDBPalphaRII). PLoS One. 2016;11(5):e0155627. | ||
Fong MY, Wong SS, Silva JR, Lau YL. Genetic polymorphism in domain I of the apical membrane antigen-1 among Plasmodium knowlesi clinical isolates from Peninsular Malaysia. Acta Trop. 2015;152:145–150. | ||
Putaporntip C, Thongaree S, Jongwutiwes S. Differential sequence diversity at merozoite surface protein-1 locus of Plasmodium knowlesi from humans and macaques in Thailand. Infect Genet Evol. 2013;18(Suppl C):213–219. | ||
Rawa MSA, Fong MY, Lau YL. Genetic diversity and natural selection in the rhoptry-associated protein 1 (RAP-1) of recent Plasmodium knowlesi clinical isolates from Malaysia. Malar J. 2016;15(1):62. | ||
Yap NJ, Goh XT, Koehler AV, et al. Genetic diversity in the C-terminus of merozoite surface protein 1 among Plasmodium knowlesi isolates from Selangor and Sabah Borneo, Malaysia. Infect Genet Evol. 2017;54:39–46. | ||
Putaporntip C, Kuamsab N, Jongwutiwes S. Sequence diversity and positive selection at the Duffy-binding protein genes of Plasmodium knowlesi and P. cynomolgi: analysis of the complete coding sequences of Thai isolates. Infect Genet Evol. 2016;44(Suppl C):367–375. | ||
Faber BW, Abdul Kadir K, Rodriguez-Garcia R, et al. Low levels of polymorphisms and no evidence for diversifying selection on the Plasmodium knowlesi apical membrane antigen 1 gene. PLoS One. 2015;10(4):e0124400. | ||
Diez Benavente E, Florez de Sessions P, Moon RW, et al. Analysis of nuclear and organellar genomes of Plasmodium knowlesi in humans reveals ancient population structure and recent recombination among host-specific subpopulations. PLoS Genet. 2017;13(9):e1007008. | ||
Ahmed AM, Pinheiro MM, Divis PC, et al. Disease progression in Plasmodium knowlesi malaria is linked to variation in invasion gene family members. PLoS Negl Trop Dis. 2014;8(8):e3086. | ||
Ahmed MA, Cox-Singh J. Plasmodium knowlesi – an emerging pathogen. Isbt Sci Ser. 2015;10(Suppl 1):134–140. | ||
Pinheiro MM, Ahmed MA, Millar SB, et al. Plasmodium knowlesi genome sequences from clinical isolates reveal extensive genomic dimorphism. PLoS One. 2015;10(4):e0121303. | ||
Yusof R, Ahmed MA, Jelip J, et al. Phylogeographic evidence for 2 genetically distinct zoonotic Plasmodium knowlesi parasites, Malaysia. Emerg Infect Dis. 2016;22(8):1371. | ||
Divis PC, Lin LC, Rovie-Ryan JJ, et al. Three divergent subpopulations of the malaria parasite Plasmodium knowlesi. Emerg Infect Dis. 2017;23(4):616–624. | ||
Moon RW, Sharaf H, Hastings CH, et al. Normocyte-binding protein required for human erythrocyte invasion by the zoonotic malaria parasite Plasmodium knowlesi. Proc Natl Acad Sci USA. 2016;113(26):7231–7236. | ||
Lim KL, Amir A, Lau YL, Fong MY. The Duffy binding protein (PkDBPalphaII) of Plasmodium knowlesi from Peninsular Malaysia and Malaysian Borneo show different binding activity level to human erythrocytes. Malar J. 2017;16(1):331. | ||
Moon RW, Hall J, Rangkuti F, et al. Adaptation of the genetically tractable malaria pathogen Plasmodium knowlesi to continuous culture in human erythrocytes. Proc Natl Acad Sci USA. 2013;110(2):531–536. | ||
Lim C, Hansen E, DeSimone TM, et al. Expansion of host cellular niche can drive adaptation of a zoonotic malaria parasite to humans. Nat Commun. 2013;4:1638. | ||
Meyer EV, Semenya AA, Okenu DM, et al. The reticulocyte binding-like proteins of P. knowlesi locate to the micronemes of merozoites and define two new members of this invasion ligand family. Mol Biochem Parasitol. 2009;165(2):111–121. | ||
Adams JH, Sim BK, Dolan SA, Fang X, Kaslow DC, Miller LH. A family of erythrocyte binding proteins of malaria parasites. Proc Natl Acad Sci USA. 1992;89(15):7085–7089. | ||
Miller LH, Mason SJ, Dvorak JA, McGinniss MH, Rothman IK. Erythrocyte receptors for (Plasmodium knowlesi) malaria: Duffy blood group determinants. Science. 1975;189(4202):561–563. | ||
Singh AP, Puri SK, Chitnis CE. Antibodies raised against receptor-binding domain of Plasmodium knowlesi Duffy binding protein inhibit erythrocyte invasion. Mol Biochem Parasitol. 2002;121(1):21–31. | ||
Pain A, Bohme U, Berry AE, et al. The genome of the simian and human malaria parasite Plasmodium knowlesi. Nature. 2008;455(7214):799–803. | ||
Bozdech Z, Mok S, Hu G, et al. The transcriptome of Plasmodium vivax reveals divergence and diversity of transcriptional regulation in malaria parasites. Proc Natl Acad Sci USA. 2008;105(42):16290–16295. | ||
Alam MS, Choudhary V, Zeeshan M, Tyagi RK, Rathore S, Sharma YD. Interaction of Plasmodium vivax tryptophan-rich antigen PvTRAg38 with band 3 on human erythrocyte surface facilitates parasite growth. J Biol Chem. 2015;290(33):20257–20272. | ||
Lapp SA, Mok S, Zhu L, et al. Plasmodium knowlesi gene expression differs in ex vivo compared to in vitro blood-stage cultures. Malar J. 2015;14:110. | ||
Tyagi K, Gupta D, Saini E, et al. Recognition of human erythrocyte receptors by the tryptophan-rich antigens of monkey malaria parasite Plasmodium knowlesi. PLoS One. 2015;10(9):e0138691. | ||
Mahdi AHM, Remarque EJ, van Duivenvoorde LM, et al. Vaccination with Plasmodium knowlesi AMA1 formulated in the novel adjuvant co-vaccine HT protects against blood-stage challenge in rhesus macaques. PLoS One. 2011;6(5):e20547. | ||
Vulliez-Le Normand B, Faber BW, Saul FA, et al. Crystal structure of Plasmodium knowlesi apical membrane antigen 1 and its complex with an invasion-inhibitory monoclonal antibody. PLoS One. 2015;10(4):e0123567. | ||
Garg A, Lukk T, Kumar V, et al. Structure, function and inhibition of the phosphoethanolamine methyltransferases of the human malaria parasites Plasmodium vivax and Plasmodium knowlesi. Sci Rep. 2015;5:9064. | ||
Maier AG, Rug M, O’Neill MT, et al. Skeleton-binding protein 1 functions at the parasitophorous vacuole membrane to traffic PfEMP1 to the Plasmodium falciparum-infected erythrocyte surface. Blood. 2007;109(3):1289–1297. | ||
Lucky AB, Sakaguchi M, Katakai Y, et al. Plasmodium knowlesi skeleton-binding protein 1 localizes to the ‘Sinton and Mulligan’ stipplings in the cytoplasm of monkey and human erythrocytes. PLoS One. 2016;11(10):e0164272. | ||
Ogutu BR, Apollo OJ, McKinney D, et al. Blood stage malaria vaccine eliciting high antigen-specific antibody concentrations confers no protection to young children in Western Kenya. PLoS One. 2009;4(3):e4708. | ||
Ellis RD, Martin LB, Shaffer D, et al. Phase 1 trial of the Plasmodium falciparum blood stage vaccine MSP1(42)-C1/Alhydrogel with and without CPG 7909 in malaria naive adults. PLoS One. 2010;5(1):e8787. | ||
Otsyula N, Angov E, Bergmann-Leitner E, et al. Results from tandem phase 1 studies evaluating the safety, reactogenicity and immunogenicity of the vaccine candidate antigen Plasmodium falciparum FVO merozoite surface protein-1 (MSP1(42)) administered intramuscularly with adjuvant system AS01. Malar J. 2013;12(1):29. | ||
Brock PM, Fornace KM, Parmiter M, et al. Plasmodium knowlesi transmission: integrating quantitative approaches from epidemiology and ecology to understand malaria as a zoonosis. Parasitology. 2016;143(4):389–400. | ||
World Health Organization. Overview of Diagnostic Testing. Geneva: World Health Organization; 2018. | ||
William T, Rahman HA, Jelip J, et al. Increasing incidence of Plasmodium knowlesi malaria following control of P. falciparum and P. vivax malaria in Sabah, Malaysia. PLoS Negl Trop Dis. 2013;7(1):e2026. | ||
Ooi CH, Bujang MA, Tg Abu Bakar Sidik TMI, Ngui R, Lim YA-L. Over two decades of Plasmodium knowlesi infections in Sarawak: trend and forecast. Acta Trop. 2017;176:83–90. | ||
Chin W, Contacos PG, Collins WE, Jeter MH, Alpert E. Experimental mosquito-transmission of Plasmodium knowlesi to man and monkey. Am J Trop Med Hyg. 1968;17(3):355–358. | ||
Maeno Y, Culleton R, Quang NT, Kawai S, Marchand RP, Nakazawa S. Plasmodium knowlesi and human malaria parasites in Khan Phu, Vietnam: gametocyte production in humans and frequent co-infection of mosquitoes. Parasitology. 2017;144(4):527–535. | ||
Lee K-S, Cox-Singh J, Singh B. Morphological features and differential counts of Plasmodium knowlesi parasites in naturally acquired human infections. Malar J. 2009;8(1):73. | ||
Vythilingam I, Wong ML, Wan-Yussof WS. Current status of Plasmodium knowlesi vectors: a public health concern? Parasitology. 2018;145(1):32–40. | ||
Ran FA, Hsu PD, Wright J, Agarwala V, Scott DA, Zhang F. Genome engineering using the CRISPR-cas9 system. Nat Protoc. 2013;8(11):2281–2308. | ||
Ghorbal M, Gorman M, Macpherson CR, Martins RM, Scherf A, Lopez-Rubio JJ. Genome editing in the human malaria parasite Plasmodium falciparum using the CRISPR-cas9 system. Nat Biotechnol. 2014;32(8):819–821. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.