Back to Journals » International Journal of Nanomedicine » Volume 21
Plant-Derived Exosome-Like Nanoparticles in Neurodegenerative Diseases: From Dual Bioactive-Delivery Roles to Translational Challenges
Authors Sun Y ![]() , Xu Z, Cui L, Guo J, Zhang X
, Xu Z, Cui L, Guo J, Zhang X ![]() , Xiao Y
, Xiao Y ![]()
Received 10 March 2026
Accepted for publication 2 June 2026
Published 9 June 2026 Volume 2026:21 608292
DOI https://doi.org/10.2147/IJN.S608292
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Prof. Dr. Anderson Oliveira Lobo
Yue Sun,1– 3 Zongqiang Xu,2,3 Lianhui Cui,2– 4 Jianfei Guo,2,3 Xiaohui Zhang,2,3 Yilei Xiao2,3
1School of Clinical Medicine, Shandong Second Medical University, Weifang, Shandong, People’s Republic of China; 2Department of Neurosurgery, Liaocheng People’s Hospital Affiliated to Shandong First Medical University, Liaocheng, Shandong, People’s Republic of China; 3Liaocheng Key Laboratory of Brain Science and Neurological Disorders Innovation Technology, Liaocheng, Shandong, People’s Republic of China; 4Postgraduate Department, Shandong First Medical University, Jinan, Shandong, People’s Republic of China
Correspondence: Xiaohui Zhang, Email [email protected] Yilei Xiao, Email [email protected]
Abstract: Neurodegenerative diseases, particularly Alzheimer’s disease (AD) and related disorders, remain difficult to treat because of their multifactorial pathogenesis, limited disease-modifying therapies, and insufficient central nervous system exposure of many therapeutic agents. Plant-derived exosome-like nanoparticles (PELNs) are emerging as biogenic nanovesicles that combine intrinsic bioactivity with natural nanocarrier properties. Enriched with lipids, proteins, small RNAs, and phytochemicals, PELNs may exert neuroprotective effects while offering opportunities for gastrointestinal stability, systemic transport, and potential central nervous system delivery. This review critically summarizes the dual bioactive-delivery roles of PELNs in AD and related neurodegenerative disorders. We discuss their potential mechanisms in modulating neuroinflammation, glial cell-mediated immune responses, redox imbalance, mitochondrial dysfunction, pathological protein aggregation, neural repair, and gut-brain axis regulation. We further examine how administration routes, biodistribution patterns, cellular uptake, and blood-brain barrier (BBB) models influence the interpretation of evidence for central nervous system (CNS) targeting. In addition, recent advances in isolation, purification, characterization, cargo loading, and surface engineering strategies are reviewed in the context of improving stability, targeting capacity, and translational feasibility. Despite their promise, the clinical development of PELNs remains constrained by source-dependent heterogeneity, non-standardized isolation methods, insufficiently defined critical quality attributes, inconsistent dosing metrics, limited pharmacokinetic and biodistribution data, and unresolved long-term biosafety concerns. Establishing rigorous Chemistry, Manufacturing, and Controls (CMC) frameworks, reproducible quality-control assays, and evidence-based translational pathways will be essential for advancing PELNs from experimental bioactive vesicles to clinically relevant neurotherapeutic platforms. The infographic highlights the bioactivity and nanocarrier role of PELNs (plant-derived extracellular vesicle-like nanoparticles). Their bioactivity includes antioxidant, anti-inflammatory, immunomodulatory effects and gut-brain axis regulation. As nanocarriers, they protect cargo, facilitate cellular uptake, enable cargo loading, surface engineering and interact with barriers. These functions help reduce neuroinflammation, offer antioxidant and mitochondrial protection, support neuronal survival, modulate Aβ/Tau pathology, regulate gut-brain axis and microbiota and enhance synaptic repair and plasticity. Clinical application requires scalable sources, standardized isolation, quality control, pharmacokinetics, safety, targeted engineering, manufacturing and regulatory compliance. Challenges include source variability, non-standardized isolation, undefined attributes, limited pharmacokinetics data, long-term safety concerns and regulatory issues.Infographic on PELNs’ bioactivity, nanocarrier function and clinical application process.
Keywords: plant-derived exosome-like nanoparticles, PELNs, Alzheimer’s disease, AD, neurodegenerative diseases, blood-brain barrier, BBB, bioactive nanocarriers
Introduction
Neurodegenerative diseases represent a spectrum of chronic, progressive disorders characterized by the selective and gradual loss of neuronal structure and function, leading to profound cognitive and motor impairments.1–3 As global life expectancy increases, the burden of these conditions has escalated into a major public health crisis, imposing substantial medical, social, and economic pressures worldwide.1,2 Among these disorders, Alzheimer’s disease (AD) is the most prevalent form of dementia and serves as a leading model for understanding proteinopathy-driven neurodegeneration, while Parkinson’s disease (PD), Huntington’s disease (HD), amyotrophic lateral sclerosis (ALS), and neural injury-related disorders share overlapping pathological cascades.3,4
Despite extensive research into the complex biology of neurodegeneration, current clinical management remains insufficient for fundamentally halting disease progression. Traditional symptomatic therapies, such as cholinesterase inhibitors, can provide limited cognitive or behavioral benefits but do not reverse the underlying neurodegenerative process.4 Although recent therapeutic advances have expanded the disease-modifying landscape of AD, the clinical efficacy of available interventions remains constrained by disease-stage specificity, heterogeneous patient responses, safety monitoring requirements, and limited ability to simultaneously target the multifactorial pathological networks that drive neurodegeneration.5 Consequently, there remains an urgent unmet need for innovative, precision-oriented therapeutic strategies capable of modulating multiple disease mechanisms and improving central nervous system (CNS) exposure.
The pathological features of AD primarily include the extracellular accumulation of β-amyloid (Aβ) plaques and the intracellular formation of neurofibrillary tangles composed of hyperphosphorylated tau protein. The misfolding, aggregation, and abnormal deposition of these proteins are widely recognized as central events associated with disease onset and progression.6,7 Beyond these classical hallmarks, increasing evidence suggests that disrupted intercellular communication, neuroinflammation, oxidative stress, mitochondrial dysfunction, impaired mitophagy, gut-brain axis dysregulation, and defective proteostasis form interconnected pathological networks across AD and related neurodegenerative disorders.8–11 In particular, systems-level studies have emphasized that gut microbial metabolism and hepatic biotransformation may reshape the bioavailability, systemic exposure, and potential CNS relevance of plant-derived bioactive compounds, supporting the importance of gut-brain axis and systemic metabolic regulation in neurotherapeutic development.12
However, the therapeutic efficacy of many systemic treatments is severely compromised by the formidable blood-brain barrier (BBB), which remains a major obstacle for CNS drug delivery, particularly for biologics, nucleic acids, and many nanomedicine formulations.13 As classically estimated, the BBB excludes almost all large-molecule neurotherapeutics and the majority of small-molecule drug candidates, making insufficient brain exposure a central bottleneck in neurotherapeutic development.13 Therefore, delivery systems capable of improving CNS accessibility while maintaining biocompatibility and functional cargo integrity are urgently needed.
In recent years, nanomedicine-based strategies have gained increasing attention for CNS disorders, especially those designed to improve the stability, bioavailability, tissue delivery, and multi-target activity of natural products and biologically derived therapeutic cargoes. More broadly, the integration of natural-product pharmacology with nanocarrier design has provided new opportunities to overcome the poor solubility, limited bioavailability, and insufficient tissue distribution of many plant-derived bioactives.14 Exosomes and other extracellular vesicles (EVs) have been identified as key mediators of intercellular communication, transporting proteins, nucleic acids, lipids, and metabolites to recipient cells and thereby regulating diverse physiological and pathological processes.15–17 Owing to their nanoscale size, membrane-enclosed structure, and biological origin, EVs have attracted considerable interest as potential diagnostic and therapeutic tools for neurological diseases.15–19 However, their CNS delivery efficiency and biological effects vary substantially depending on vesicle origin, administration route, disease state, and detection method. In addition, mammalian cell-derived EVs face translational hurdles such as limited yield, high production cost, donor- or cell-source variability, and potential safety concerns.20
Against this background, plant-derived exosome-like nanoparticles (PELNs) have emerged as a compelling class of biogenic nanovesicles at the interface of natural-product pharmacology and nanomedicine. Compared with synthetic nanoparticles or mammalian cell-derived EVs, PELNs may offer several practical advantages, including sustainable sourcing, relatively scalable production, favorable biosafety profiles, and intrinsic bioactive cargoes.20–23 Importantly, PELNs should not be interpreted as pharmacological equivalents of whole plants or crude plant extracts. Rather, they represent nanoscale, membrane-enclosed bioactive fractions isolated from plant tissues, with molecular cargoes including lipids, proteins, nucleic acids/small RNAs, and small-molecule metabolites or phytochemicals. These vesicle-associated features may confer biological activity and delivery potential that differ from those of unfractionated plant materials or conventional plant extracts. This conceptual distinction is essential for interpreting their biological activity, standardizing their preparation, and designing clinically translatable products.21–26
Accumulating studies suggest that selected PELNs may exert neuroprotective effects by modulating neuroinflammatory responses, oxidative stress, mitochondrial dysfunction, proteinopathy-related pathways, and gut-brain axis regulation.11,22,27–30 More broadly, plant-derived phytocompounds have also been reported to regulate stress-response and proteostasis-related pathways implicated in neurodegenerative diseases, including AD.31 However, evidence supporting their BBB transport and CNS targeting remains heterogeneous. Administration route, vesicle labeling strategy, disease-associated BBB disruption, cellular uptake pathway, and biodistribution analysis can substantially influence the interpretation of BBB-crossing results. Therefore, the therapeutic potential of PELNs should be evaluated not only through efficacy outcomes, but also through rigorous assessment of vesicle identity, physicochemical properties, administration-route-dependent biodistribution, and evidence strength for intact-vesicle transport.
Despite growing interest in PELNs, the field still lacks a comprehensive framework that integrates their biological identity, physicochemical properties, delivery behavior, therapeutic mechanisms, engineering strategies, and translational requirements. Previous studies and reviews have provided valuable insights into the separation, characterization, and biomedical applications of PELNs.11,21–23,27 However, critical questions remain regarding source-dependent heterogeneity, standardized isolation and purification, quality-control benchmarks, critical quality attributes, batch-to-batch reproducibility, dosing normalization, pharmacokinetics, long-term safety, and regulatory classification.22,23 From a manufacturing perspective, scalable purification strategies, including tangential flow filtration (TFF), size-exclusion chromatography, and combined workflows, have been increasingly discussed as approaches to improve yield, recovery, purity, and process reproducibility; however, their suitability for clinical-grade PELN production remains insufficiently validated.22,23
Therefore, this review aims to provide a mechanism- and translation-oriented framework for evaluating PELNs in AD and related neurodegenerative disorders. We first clarify the biological identity, molecular composition, and source-dependent heterogeneity of PELNs, followed by a critical discussion of isolation, purification, characterization, and quality control. We then examine administration-route-dependent biodistribution, cellular uptake, BBB transport evidence, and disease-shared therapeutic mechanisms involving neuroinflammation, redox imbalance, mitochondrial dysfunction, proteinopathy, neural repair, and gut-brain axis regulation. Finally, we discuss engineering strategies, pharmacokinetic uncertainties, long-term safety, Chemistry, Manufacturing, and Controls (CMC) requirements, regulatory classification, and a tiered clinical translation pathway. Through this framework, we seek to define both the therapeutic promise and the translational boundaries of PELNs as bioactive nanocarriers for neurodegenerative diseases.
Overview of PELNs as Bioactive Nanocarriers
Definition, Nomenclature, and Biological Identity of PELNs
PELNs are nanoscale vesicle-like particles isolated from plant-derived materials, including plant tissues, fruits, vegetables, medicinal plants, and plant cell cultures. Many PELNs preparations are commonly reported within the exosome-like nanoscale range of approximately 30–150 nm, although broader size distributions extending beyond this range may occur depending on plant source, tissue type, physiological status, sample processing, and isolation method.25,32–34
Although PELNs share several exosome-like features with mammalian EVs, including nanoscale morphology, membrane-enclosed architecture, and the ability to carry bioactive molecular cargoes, they should not be considered direct equivalents of mammalian exosomes. Mammalian exosomes are generally associated with endosomal biogenesis and relatively established EV nomenclature, reporting frameworks, and characterization guidelines, whereas PELNs are more often operationally defined by isolation procedure, morphology, particle size distribution, density, surface charge, and molecular composition.25,32,33,35,36
In this review, the term “exosome-like” is used to describe the exosome-resembling nanoscale morphology, membrane-enclosed structure, and cargo-carrying capacity of PELNs, rather than to imply complete equivalence to mammalian exosomes in biogenesis or marker identity. This distinction is important because different plant sources and preparation methods may produce vesicle populations with distinct size distributions, cargo profiles, biological activities, and translational properties.25,32,33
Accordingly, PELNs are discussed as bioactive nanocarriers at the interface of plant-derived natural products and nanomedicine. Their biological identity is defined not only by their exosome-like nanoscale morphology, but also by membrane organization, molecular cargoes, source-dependent heterogeneity, and vesicle-associated delivery potential.
Structure, Molecular Composition, and Bioactive Cargoes of PELNs
Structurally, PELNs are generally characterized by a lipid bilayer or membrane-like envelope that protects internal bioactive cargoes. These cargoes include lipids, proteins, nucleic acids, and small-molecule metabolites.25,34,37 The structural and compositional characteristics of PELNs are illustrated in Figure 1. This membrane-enclosed architecture provides a physical basis for their colloidal stability, cargo protection, and potential interactions with recipient cells.
 |
Figure 1 Structural and compositional characteristics of plant-derived exosome-like nanoparticles (PELNs). (A) Representative transmission electron microscopy (TEM) image illustrating the vesicle-like morphology and nanoscale size distribution of PELNs (adapted from Ref.38). (B) Schematic comparison of lipid-composition tendencies between PELNs and mammalian extracellular vesicles (EVs). The upward arrows (↑) indicate relative enrichment tendencies or frequently reported abundance in the indicated vesicle category, rather than universal diagnostic markers or quantitative increases in every preparation. PELNs may display a plant-associated lipid profile, including phosphatidic acid (PA), digalactosyldiacylglycerol/monogalactosyldiacylglycerol (DGDG/MGDG), phosphatidylethanolamine (PE), plant sterols, and source-dependent ether-linked lipids, whereas mammalian EVs are often associated with cholesterol, sphingomyelin, ceramide, and phosphatidylserine. (C) Schematic illustration of the diverse molecular cargoes carried by PELNs, including lipids, proteins, nucleic acids, and plant-derived secondary metabolites or phytochemicals. (D) Overview of key biological features of PELNs as bioactive nanocarriers, including cargo protection, cellular interaction, favorable biocompatibility, relatively low immunogenicity, and intrinsic bioactivity. These structural, compositional, and functional features should be interpreted as source- and preparation-dependent characteristics because PELNs are heterogeneous and may vary according to plant species, tissue origin, physiological state, processing conditions, and isolation method. Panels B–D were created by the authors based on Refs.20,38–41 |
Lipids are among the most important structural and functional components of PELNs because they contribute to membrane integrity, environmental stability, cellular interaction, and cargo protection. Lipidomic analyses have shown that PELNs may contain phospholipids, glycolipids, triglycerides, phosphatidic acid (PA), phosphatidylethanolamine, phosphatidylcholine, phosphatidylglycerol, phosphatidylinositol, plant sterols, galactolipids, and, in certain preparations, ether-linked lipid species.34,35,37,38,42,43 However, the relative abundance of these lipid classes varies substantially across plant species, tissue types, physiological states, and preparation methods.
In some plant-derived vesicles, enrichment of PA, galactolipids, plant sterols, or ether-linked lipids has been associated with enhanced membrane stability, resistance to gastrointestinal conditions, cargo protection, and interactions with recipient cell membranes.34,35,37,38,43,44 Nevertheless, these properties should not be generalized to all PELNs, because lipid composition is highly source-dependent and can be affected by plant species, cultivation environment, tissue origin, developmental stage, extraction procedure, and purification workflow.25,33,34,37
Compared with many mammalian EVs, which are often enriched in cholesterol, sphingomyelin, ceramide, and phosphatidylserine, PELNs may display a more plant-associated lipid profile, including PA, galactolipids, plant sterols, and other plant-specific lipid species.20,37–39,42 However, this distinction should be interpreted as a general compositional tendency rather than a universal diagnostic criterion, because both mammalian EVs and PELNs are heterogeneous populations whose lipid profiles depend strongly on cell type, biological source, and isolation method.
Protein cargoes represent another important component of PELNs. Proteomic studies have identified proteins associated with membrane trafficking, vesicle transport, stress responses, defense signaling, cell-wall remodeling, metabolism, and transmembrane transport.23,37,45–47 In particular, plant vesicle preparations have been reported to contain proteins such as tetraspanins, aquaporins, adenosine triphosphatases (ATPases), ATP-binding cassette (ABC) transporters, heat shock proteins, pathogenesis-related proteins, and cell-wall-modifying enzymes.37,45,47,48
Unlike mammalian EVs, which are frequently characterized using EV-associated proteins such as the tetraspanins CD9, CD63, and CD81, tumor susceptibility gene 101 (TSG101), and ALG-2-interacting protein X (ALIX) within established EV reporting frameworks, PELNs currently lack universally accepted surface markers.36 Candidate plant EV-associated proteins, including tetraspanin 8 (TET8) and penetration 1 (PEN1), have been reported in specific plant vesicle populations, but their applicability across different plant species, tissue origins, and PELN preparations remains limited.23,25,45–47 These protein components may contribute to vesicle stability, stress adaptation, membrane trafficking, cell-wall passage, and interactions with recipient cells. However, receptor-specific uptake mechanisms and universal PELN marker systems remain insufficiently defined.23,25,46,47 This uncertainty highlights the need for standardized proteomic profiling and source-specific marker validation.
Although direct evidence for protein corona formation on PELNs remains limited, studies on synthetic nanoparticles and mammalian extracellular vesicles indicate that nanoscale vesicles rapidly acquire biomolecular or protein coronas upon exposure to biological fluids. This corona could redefine their biological identity, cellular uptake, immune recognition, biodistribution, and therapeutic performance.25,49,50
PELNs have also been reported to contain diverse RNA species, including miRNA-like small RNAs, small interfering RNAs, and other regulatory RNAs. For example, miRNA-like small RNAs have been identified in exosome-like nanoparticles isolated from multiple edible fruits and vegetables, supporting the presence of plant-derived regulatory RNA cargoes in selected PELN preparations.40,44,47,51–54 These RNA cargoes have attracted interest because of their potential roles in cross-kingdom communication, gene-expression modulation, host–microbe interactions, and plant defense responses.40,47,51–53 Nevertheless, the abundance, stability, delivery efficiency, and functional relevance of plant small RNAs in mammalian recipient cells remain context-dependent and should be interpreted with methodological caution. Therefore, plant small RNAs should be discussed as potentially functional cargoes rather than universally validated mediators of mammalian gene regulation.
In addition to lipids, proteins, and RNAs, PELNs may carry plant-derived secondary metabolites, including flavonoids, phenolic compounds, terpenoids, stilbene derivatives, and other phytochemicals.37,55–57 These small molecules may contribute to anti-inflammatory, antioxidant, immunomodulatory, or antitumor activities reported for selected PELNs preparations.55–58 However, the metabolite profile of PELNs is not necessarily identical to that of crude plant extracts; vesicle isolation may enrich, exclude, or alter the relative abundance of specific bioactive compounds.26,56 Thus, PELNs should be viewed as nanoscale, membrane-enclosed bioactive fractions rather than simple miniature versions of whole plants or unfractionated plant extracts.
Together, lipids, proteins, small RNAs, and secondary metabolites form an integrated cargo system that provides a mechanistic basis for considering PELNs as bioactive nanocarriers. At the same time, their complex and source-dependent composition creates challenges for identity definition, potency evaluation, quality control, dosing normalization, and batch-to-batch reproducibility. These compositional features therefore provide the rationale for subsequent discussions on standardized isolation, characterization, and critical quality attributes.
Comparison of PELNs with Crude Plant Extracts, Mammalian EVs, and Synthetic Nanocarriers
A key conceptual issue in the PELNs field is that PELNs should not be interpreted as pharmacological equivalents of whole plants or crude plant extracts. Rather, they represent nanoscale, membrane-enclosed bioactive fractions isolated from plant materials, with molecular cargoes including lipids, proteins, nucleic acids, and small-molecule metabolites or phytochemicals.25,26 These vesicle-associated features may confer biological activity, stability, and delivery properties that differ from those of unfractionated plant materials or conventional plant extracts.26
This distinction is particularly important in the broader context of natural-product nanomedicine. The integration of natural-product pharmacology with nanocarrier design has been proposed as a strategy to overcome the poor solubility, instability, low oral bioavailability, and limited tissue distribution of many plant-derived bioactive compounds.16 PELNs fit within this framework because they combine plant-derived bioactivity with vesicle-associated delivery properties, thereby serving not merely as extracts, but as biologically derived nanocarriers.
Compared with mammalian EVs, PELNs may offer several practical advantages, including abundant plant sources, relatively low production cost, dietary or medicinal plant compatibility, favorable biocompatibility, and potential scalability.32,33,47,59 However, mammalian EVs currently benefit from more mature nomenclature, marker systems, and characterization guidelines, whereas PELNs still lack universally accepted identity markers, potency assays, and critical quality attributes.25,32,33 Therefore, the translational advantage of PELNs should be framed as potential rather than established.
Although many PELN preparations have been reported to exhibit favorable biocompatibility and low immunogenicity, their safety profiles should be evaluated according to plant source, dose, administration route, and preparation method. Differences in plant allergens, residual contaminants, protein corona formation, and purification quality may influence biological responses after administration.25,26
Compared with synthetic nanoparticles, PELNs possess a naturally derived membrane structure and endogenous bioactive cargoes. These features may support cargo protection, biological compatibility, and interactions with mammalian cells or gut microbiota.35,43,60,61 In addition, engineered plant-derived lipid nanovectors, such as ginger-derived nanolipids loaded with doxorubicin, have been explored as drug-delivery platforms, supporting the broader concept that plant-derived nanoscale lipid systems can be adapted for therapeutic cargo delivery.41 However, synthetic nanoparticles often provide more controllable composition, reproducible manufacturing, and tunable physicochemical properties. Thus, PELNs should be viewed as bioactive, source-dependent nanocarriers with unique biological potential but also greater compositional complexity and standardization challenges.
A systematic comparison of crude plant extracts, PELNs, mammalian EVs, and synthetic nanoparticles is summarized in Table 1. This comparison emphasizes that PELNs occupy an intermediate position between natural products and engineered nanomedicines: they are biologically derived and cargo-rich, but their clinical development requires nanomedicine-level characterization, quality control, and manufacturing standards. Because mammalian EVs remain the most commonly used biological comparator for PELNs, a more focused comparison of delivery-relevant molecular features between PELNs and mammalian EVs is provided in Table 2.
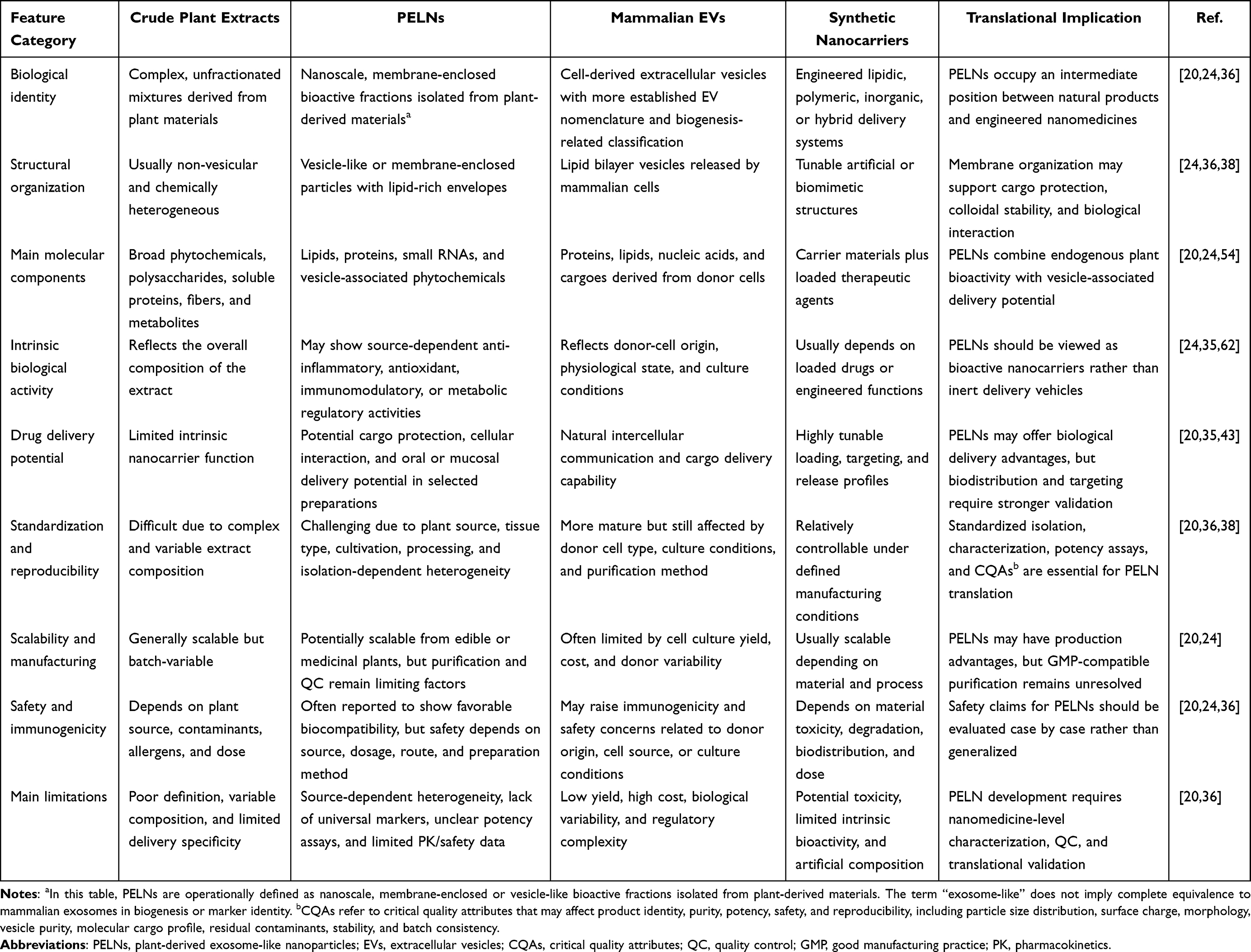 |
Table 1 Conceptual Comparison of Crude Plant Extracts, PELNs, Mammalian EVs, and Synthetic Nanocarriers |
 |
Table 2 Focused Comparison of Delivery-Relevant Molecular Features Between PELNs and Mammalian EVs |
Source-Dependent Heterogeneity of PELNs: Effects of Plant Source, Cultivation, and Processing Conditions
Source-dependent heterogeneity is a defining feature of PELNs. Variations in plant species, tissue type, developmental stage, cultivation environment, harvest conditions, storage, extraction buffer, centrifugation parameters, filtration membrane, and downstream purification strategy can influence vesicle yield, size distribution, surface charge, lipid composition, RNA cargoes, protein profiles, metabolite enrichment, biological activity, and safety profile.25,32,33,37,66
For example, different plant sources may generate PELNs with distinct lipidomic and metabolomic signatures. Some edible or medicinal plant-derived vesicles are enriched in phosphatidic acid, plant sterols, ether-linked lipids, flavonoids, phenolic compounds, or other secondary metabolites, whereas others may show different protein or RNA cargo profiles.34,37,42,55,57 These differences may partly explain why PELNs from different plants display diverse biological activities, including immunomodulatory, anti-inflammatory, antioxidant, intestinal regulatory, or drug-delivery-related effects.35,37,55,57,60,61
Several edible PELNs have been reported to interact with intestinal epithelial cells, immune cells, or gut-associated biological pathways, suggesting a potential role in gut-mediated systemic regulation.35,60,61 For example, grape exosome-like nanoparticles were reported to induce intestinal stem cells and protect mice from dextran sulfate sodium (DSS)-induced colitis, supporting the concept that selected edible PELNs may exert intrinsic biological activity through intestinal mechanisms. However, these effects should not be assumed to be universal across all PELNs.62 However, these effects should not be assumed to be universal across all PELNs. Whether gut-related activity is mediated by intact vesicles, released cargoes, plant metabolites, microbiota-dependent transformation, or host immune responses remains to be clarified.
For plant-derived bioactives with low oral bioavailability, systemic exposure may be shaped not only by the parent compound but also by gut microbial metabolism, hepatic biotransformation, and host metabolic networks.13 This perspective is relevant to PELNs because their biological effects may arise from both vesicle-mediated cargo delivery and indirect regulation of intestinal, immune, and metabolic pathways.
This heterogeneity is both an opportunity and a challenge. On one hand, it allows different plant sources to provide distinct therapeutic cargoes and biological functions. On the other hand, it complicates identity definition, quality control, dosing normalization, potency assay development, and batch-to-batch reproducibility. These issues are particularly important for clinical translation, where source material, cultivation conditions, processing workflows, and storage parameters must be controlled to ensure product consistency.
Therefore, understanding the biological identity and source-dependent heterogeneity of PELNs provides the conceptual basis for subsequent discussions of standardized isolation, scalable purification, physicochemical characterization, quality control, and critical quality attributes.
Isolation, Purification, Characterization, and Quality Control of PELNs
The source-dependent heterogeneity of PELNs makes standardized isolation, purification, characterization, and quality control essential for mechanistic interpretation and translational development. Unlike chemically defined synthetic nanoparticles, PELNs are isolated from complex plant matrices that contain soluble proteins, polysaccharides, phenolic compounds, pigments, cell-wall fragments, organelle-derived membranes, and other plant-derived colloidal materials. Therefore, the apparent physicochemical properties and biological activities of PELN preparations can be strongly influenced by plant source, tissue type, extraction buffer, homogenization procedure, centrifugation conditions, filtration steps, purification strategy, and storage conditions.21–25,33,47,59,66,67
Current PELN studies have used a range of isolation and purification workflows, including ultracentrifugation (UC), density-gradient ultracentrifugation (DGUC), size-exclusion chromatography (SEC), immunoaffinity capture, tangential flow filtration (TFF), ion-exchange or other chromatography-based purification methods, and emerging microfluidic technologies.21–23,25,66,68–73 However, no single method can simultaneously maximize yield, purity, vesicle integrity, cargo preservation, scalability, and manufacturing compatibility. A comprehensive comparison of yield, recovery, purity, contaminant risk, vesicle integrity, cargo preservation, scalability, and translational relevance across major PELN isolation and purification strategies is provided in Table 3. For translational applications, PELN production should move beyond simple particle enrichment toward standardized workflows that define product identity, purity, potency, safety, and batch-to-batch reproducibility.21–23,25,47,59,66,73
 |
Table 3 Comparison of Major Isolation and Purification Strategies for PELNs |
Conventional Isolation Methods
Ultracentrifugation remains one of the most widely used laboratory-scale methods for PELN isolation. UC relies on centrifugal force to sediment particles according to size and density, and is commonly performed after sequential low-speed centrifugation to remove fibers, cell debris, cell-wall fragments, organelles, and larger vesicular structures.68,69 This approach is technically accessible and suitable for initial enrichment. However, UC is often limited by insufficient purity, protein aggregation, and co-isolation of soluble proteins, polysaccharides, ribonucleoprotein complexes, phenolic aggregates, pigment-containing materials, and other non-vesicular particles.25,66,70,71,73 Prolonged exposure to high centrifugal forces may also promote vesicle aggregation, deformation, or loss of fragile subpopulations. Therefore, UC-derived pellets should not be assumed to represent highly purified PELNs without additional purity assessment and orthogonal characterization.25,66,68,70,71,73
DGUC can improve sample purity by separating vesicle-like particles according to buoyant density. This method is useful for reducing contamination from soluble macromolecules, protein aggregates, pigment complexes, and non-vesicular plant-derived materials.25,66,71,73 Nevertheless, DGUC is labor-intensive, time-consuming, difficult to scale, and may reduce vesicle recovery. Moreover, particles with overlapping buoyant densities may include both vesicular and non-vesicular components. Thus, DGUC should be considered a useful refinement step rather than definitive proof of PELN identity.25,66,71,73
SEC preserves vesicle integrity more effectively than harsh sedimentation-based methods and can separate vesicle-like particles from soluble proteins and small molecular contaminants.66,70,72–76 However, SEC throughput is limited by column capacity, sample dilution, processing time, and flow-rate constraints. Therefore, SEC is often most useful as a polishing step after initial enrichment by UC, TFF, or other concentration methods.66,70,72–76
Immunoaffinity capture can achieve high specificity in mammalian EV research, but its application to PELNs remains constrained by cost, low yield, and the lack of universally validated PELN markers.25,66,73,77 Classical mammalian EV markers such as CD63, CD81, CD9, TSG101, and ALIX are not generally applicable to PELNs. Although candidate plant EV-associated proteins such as TET8 and PEN1 have been reported in specific plant vesicle populations, their broad applicability across plant species, tissue origins, and isolation workflows remains unclear.34,40,45 Therefore, affinity-based isolation currently appears more suitable for studying defined plant EV subpopulations than for general PELN production.
Scalable Purification Strategies
To address the bottleneck of scalability, TFF has emerged as a promising strategy for large-volume processing of plant extracts.74–76,78,79 Unlike dead-end filtration, TFF uses cross-flow filtration to reduce membrane clogging and allows continuous concentration, diafiltration, and buffer exchange.74–76,78,79 A TFF-centered workflow, illustrated in Figure 2, may improve large-volume processing and help preserve vesicle integrity; however, its performance depends on membrane cutoff, pore size, transmembrane pressure, flow rate, feed viscosity, processing time, and downstream polishing steps.74–76,78,79,83,84 Therefore, TFF should be optimized and reported in detail rather than treated as a universally superior method.
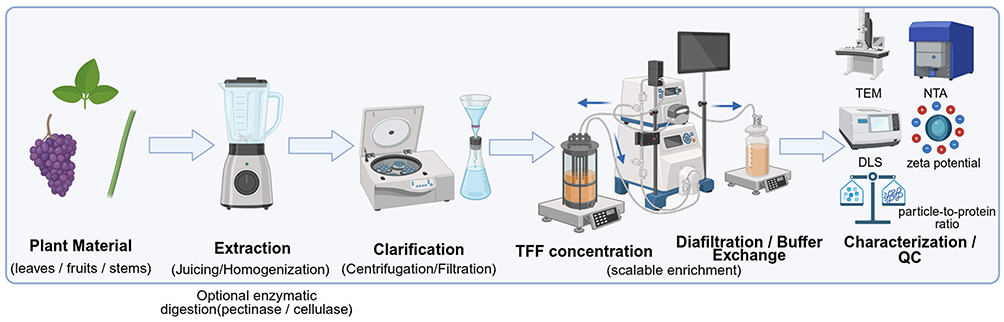 |
Figure 2 Representative tangential flow filtration (TFF)-centered workflow for the isolation and purification of plant-derived exosome-like nanoparticles (PELNs). Left-to-right schematic of a representative TFF-centered PELN preparation workflow: plant material → extraction by juicing or homogenization, with optional enzymatic digestion using pectinase or cellulase → clarification by centrifugation and/or filtration → TFF-based concentration → diafiltration/buffer exchange → characterization and quality control (QC). The rightward arrows indicate the sequential order of processing steps in this representative workflow and do not imply that this is the only applicable or universally required isolation strategy for all PELN preparations. The final preparation is assessed using characterization and QC parameters, including transmission electron microscopy (TEM), nanoparticle tracking analysis (NTA), dynamic light scattering (DLS), particle-to-protein ratio, and zeta potential. Created with BioRender.com. |
Previous studies have reported the use of TFF or TFF-related workflows for extracting PELNs from sources such as lotus and Catharanthus roseus, supporting its potential utility for scalable production.38,74–76,78,85 Nevertheless, plant-source-dependent differences in viscosity, fiber content, soluble macromolecules, and particle heterogeneity may affect filtration performance and final product quality.38,78,85
Current trends favor hybrid purification strategies rather than reliance on a single isolation method. For example, TFF can be used for rapid volume reduction and buffer exchange, followed by SEC, DGUC, or chromatography-based polishing to improve purity.21,72,74–76,79,80,86 Such combined workflows may better balance recovery, structural integrity, contaminant removal, scalability, and reproducibility. However, the optimal workflow should be selected according to the intended application, required purity, route of administration, and downstream quality-control criteria.
Emerging microfluidic technologies based on viscoelastic flow, acoustic separation, electrophoretic manipulation, or affinity-assisted capture are also being explored for extracellular vesicle isolation and analysis.73,81,82,87 These automated platforms may offer precise manipulation of nanoparticles with low sample consumption and minimal shear stress. However, most microfluidic systems remain at the proof-of-concept or analytical-scale stage for plant-derived vesicles. Their compatibility with high-throughput PELN manufacturing, plant-derived complex matrices, and good manufacturing practice (GMP)-compatible production remains to be validated.73,81,82,87
Physicochemical and Molecular Characterization
Reliable characterization is essential because particle enrichment alone does not confirm vesicle identity or product quality. A multiparameter characterization strategy informed by the Minimal Information for Studies of Extracellular Vesicles (MISEV) recommendations should be adopted for PELNs, while recognizing that PELNs lack universally accepted marker systems and may require source-specific validation.25,36,66,68,73,77,88 Characterization should include orthogonal measurements of particle size, concentration, morphology, surface charge, density, molecular cargoes, and potential contaminants.25,36,66,68,73
Particle size and concentration should be assessed using complementary methods such as nanoparticle tracking analysis (NTA), dynamic light scattering (DLS), tunable resistive pulse sensing (TRPS), or related single-particle technologies.25,36,66,68,73 NTA provides particle size distribution and concentration but can be influenced by dilution, refractive index, background particles, camera settings, and operator-dependent thresholds. DLS is useful for assessing hydrodynamic size and polydispersity but is highly sensitive to larger aggregates. TRPS can provide single-particle size and concentration information, although its application depends on appropriate pore selection and sample compatibility. Therefore, size analysis should be interpreted together with imaging, purity measurements, and molecular profiling rather than used as a stand-alone indicator of PELN identity.25,36,66,68,73
Morphological characterization by transmission electron microscopy (TEM), scanning electron microscopy (SEM), cryo-electron microscopy, or atomic force microscopy can provide direct visual evidence of vesicle-like structures.25,36,66,68,73,89 However, conventional electron microscopy may introduce dehydration, staining, or fixation artifacts, and cup-shaped morphology should not be considered definitive evidence of exosome identity. For PELNs, imaging should ideally be combined with particle counting, density assessment, surface charge measurement, and molecular characterization.25,36,66,68,73
Surface charge, commonly measured as zeta potential, provides information on colloidal stability and potential interactions with biological membranes.25,66,68,89 However, zeta potential can vary according to buffer composition, ionic strength, pH, purification method, and residual plant-derived macromolecules. Therefore, zeta potential should be reported together with sample preparation details and measurement conditions.25,66,68,73
Molecular characterization should include lipidomic, proteomic, small RNA, and metabolomic profiling whenever possible.37,38,40,42,45,51,55,63,86,90 Lipidomic analysis can identify plant-associated lipid species such as phosphatidic acid, phosphatidylethanolamine, galactolipids, plant sterols, and source-dependent ether-linked lipids.37,38,42,91 Proteomic analysis can reveal membrane-associated proteins, transporters, stress-response proteins, and candidate plant EV-associated proteins.34,40,45,63,86,90 Small RNA sequencing can identify miRNA-like small RNAs and other regulatory RNA species, while metabolomic analysis can determine whether specific phytochemicals are enriched, depleted, or altered during vesicle isolation.33,40,51,55 These analyses are important because PELN bioactivity may arise from an integrated cargo system rather than from a single molecular component.37,38,40,42,51,55,63
Contamination assessment is equally important. Plant extracts contain abundant soluble proteins, polysaccharides, phenolic compounds, pigments, ribonucleoprotein complexes, cell-wall-derived materials, and organelle fragments that may co-purify with PELNs and influence functional assays.25,66,68–73 Future studies should therefore report not only particle number and morphology, but also particle-to-protein ratio, particle-to-lipid ratio, residual soluble protein content, RNA distribution, density range, and negative controls for non-vesicular contaminants.25,66,68–73 Without these controls, it remains difficult to distinguish vesicle-mediated effects from biological activities caused by co-isolated soluble plant compounds or extraction artifacts.25,66,70,71,73
Advanced analytical and biosensing technologies may further support PELN quality assessment in the future. For example, nanozyme-based sensing platforms and nanoparticle-based exosome biosensing systems have recently been developed for small-molecule detection or tumor-derived exosome detection, and these strategies may inspire future analytical assays for PELN quality control.73,92,93 However, these studies should be cited as methodological references rather than direct evidence for PELN therapeutic activity.
Critical Quality Attributes and Batch-to-Batch Reproducibility
The translational development of PELNs requires the identification of critical quality attributes (CQAs) linked to product identity, purity, potency, safety, and reproducibility.21–23,25,33,36,47,59,66,73,77,88 Potential CQAs include plant source authentication, particle size distribution, particle concentration, morphology, surface charge, vesicle purity, lipid profile, protein cargo, RNA cargo, metabolite composition, sterility, residual contaminants, endotoxin-like activity, colloidal stability, storage stability, and batch consistency.21–23,25,33,47,59,66,73,87 These attributes should be selected according to the intended route of administration, therapeutic indication, and proposed mechanism of action.
Batch-to-batch reproducibility is a major challenge because PELN composition can be affected by plant species, cultivar, tissue type, developmental stage, cultivation environment, harvest season, storage conditions, and processing workflow.22,23,25,33,38,47,59,66,89 For medicinal plants, additional variables such as geographic origin, pesticide residues, microbial burden, heavy metals, and phytochemical variability may further influence the final PELN product. Therefore, source-material control should be considered an upstream component of PELN quality control rather than a separate agricultural issue.22,23,25,33,47,59,66,89
Dosing normalization remains unresolved in the PELN field. Current studies report PELN doses using different metrics, including particle number, protein mass, lipid amount, RNA content, plant tissue equivalent, or extract volume equivalent.22,23,25,59,66 These inconsistencies make it difficult to compare biological effects across studies. For greater reproducibility, future studies should report multiple dose metrics simultaneously, including particle number, total protein, total lipid, RNA amount, and source-material equivalent. When specific active cargoes are proposed, their abundance should also be quantified and related to biological potency.22,23,25,59,66
Potency assays are particularly important but remain underdeveloped. Depending on the intended application, potency evaluation may include assays for anti-inflammatory activity, antioxidant effects, epithelial uptake, macrophage or microglial polarization, cargo delivery, gut barrier regulation, or neuroprotective effects.35,38,46,51,56,60,78,80,94–99 However, no universal potency assay is currently available for PELNs. Therefore, potency assays should be mechanism-informed and product-specific. For example, PELNs intended for gut-mediated neuroprotection may require assays related to intestinal epithelial uptake, gut barrier integrity, microbiota interaction, or immune modulation, whereas engineered CNS delivery systems may require assays evaluating cargo retention, cellular uptake, and transport across relevant barrier models.28,35,46,56,60,94–99
Physicochemical and storage stability should also be incorporated into quality-control workflows. Previous studies suggest that selected PELN preparations may retain vesicle morphology or cargo stability under simulated gastrointestinal conditions, variable pH environments, or storage conditions, partly depending on lipid composition and vesicle–cargo interactions.57,61,87,96,100,101 However, these findings should be interpreted as source- and preparation-specific observations rather than universal properties of all PELNs. Freezing, thawing, lyophilization, buffer composition, cryoprotectants, pH, ionic strength, and storage duration may alter particle size distribution, aggregation state, cargo integrity, and biological activity.57,61,87,100,101 Therefore, basic reporting of storage conditions and freeze-thaw history should be included in all PELN studies, and stability-indicating assays should be developed for translational products.
Together, standardized isolation, scalable purification, orthogonal characterization, CQA definition, potency testing, dosing normalization, and reproducibility assessment form the methodological foundation for PELN translation.21–23,25,33,47,59,66,73 These requirements are particularly important for neurodegenerative disease applications, where chronic administration, low tolerance for impurities, and the need for reliable biodistribution and safety data impose higher standards on product quality. After PELN preparations are rigorously defined through reproducible quality-control frameworks, the next key issue is how administration route, biodistribution, and BBB transport influence their CNS delivery potential.100,102
Administration Routes, Biodistribution, Cellular Uptake and BBB Transport
After isolation, purification, and quality control have been defined, the biological interpretation of PELNs depends strongly on the route of administration. Oral, intranasal, and intravenous delivery expose PELNs to distinct anatomical barriers, biological fluids, immune compartments, clearance mechanisms, and cellular targets. Therefore, the targeting ability of PELNs should not be discussed as an intrinsic and route-independent property. Instead, biodistribution, cellular uptake, BBB transport, and CNS-related effects should be evaluated in a route-specific and preparation-specific manner.11,13,22,100,102–107
This distinction is particularly important for neurodegenerative disease applications. Evidence of cellular uptake in vitro does not necessarily demonstrate BBB transport in vivo, and brain-associated fluorescence or tissue signal does not always confirm intact vesicle entry into brain parenchyma or functional cargo delivery. Accordingly, administration-route studies should be interpreted together with particle stability, dose normalization, biodistribution imaging, tissue perfusion controls, cellular localization, cargo release, and functional readouts.102,105–108
Oral Administration
Oral administration is one of the most attractive routes for edible and medicinal plant-derived PELNs because it is non-invasive, patient-friendly, and compatible with repeated dosing. The oral route is also biologically relevant for PELNs derived from dietary plants, as these vesicle-like particles are first exposed to saliva, gastric acid, digestive enzymes, bile salts, mucus, intestinal epithelial cells, immune cells, and gut microbiota. Unlike intravenous or intranasal administration, oral delivery does not necessarily require direct BBB crossing to produce CNS-related effects; instead, PELNs may influence neurodegeneration through intestinal absorption, immune modulation, microbial regulation, metabolite production, and gut-brain axis signaling.11,28,35,60,61,64,102,109,110
Several studies suggest that selected plant-derived vesicle preparations can retain nanoscale morphology or protect vesicle-associated cargoes under simulated gastrointestinal conditions. For example, sesame leaf-derived nanovesicles were reported to improve the stability and bioavailability of luteolin compared with free luteolin, while Cyperus-derived exosome-like particles loaded with selenium nanoparticles showed stability under variable pH and storage conditions.57,101 These observations support the possibility that certain PELN preparations may protect labile bioactive compounds during gastrointestinal transit. However, such stability should be interpreted as source- and preparation-dependent rather than a universal property of all PELNs.57,61,87,101
Following oral administration, PELNs may interact with intestinal epithelial cells, macrophages, dendritic cells, and gut microbiota. These interactions may affect epithelial barrier function, inflammatory tone, microbial composition, and microbial metabolites such as short-chain fatty acids, all of which are relevant to gut-brain axis regulation. Citrus, ginger, grape, Allium tuberosum, pueraria, and other edible plant-derived vesicle-like particles have been associated with intestinal immune regulation, microbiota remodeling, or anti-inflammatory effects in selected models.35,46,60,61,64,94,109–111 Nevertheless, it remains difficult to determine whether oral PELN effects are mediated by intact vesicle absorption, released cargoes, vesicle-associated phytochemicals, microbiota-dependent transformation, or host immune responses. Therefore, future oral delivery studies should combine simulated digestion, intestinal uptake assays, microbiota analysis, systemic exposure assessment, and CNS-related functional outcomes.
Another unresolved issue is dose interpretation after oral administration. Current studies often normalize oral PELN doses by plant mass, extract volume, particle number, protein amount, or RNA content, making cross-study comparison difficult. Because oral bioavailability may be affected by digestion, mucus penetration, microbiota interaction, and epithelial uptake, future studies should report multiple dose metrics and distinguish local intestinal effects from systemic and CNS-related outcomes.22,23,25,59,66,102,104
Intranasal Administration
Intranasal administration provides a non-invasive route that may partially bypass the BBB through olfactory and trigeminal nerve-associated pathways. This route is particularly attractive for CNS delivery because it can reduce first-pass metabolism and may enable more direct access to the brain or cerebrospinal fluid-associated compartments. However, intranasal delivery is also constrained by limited dosing volume, mucociliary clearance, nasal enzymatic degradation, interspecies anatomical differences, and formulation-dependent residence time.103,104,106,107,112
A representative example is grapefruit-derived nanovectors delivering therapeutic miR-17 through the intranasal route, which inhibited brain tumor progression in a preclinical model.74 This study supports the feasibility of plant-derived lipid nanovectors for nose-to-brain delivery. However, it should not be interpreted as definitive evidence that all native PELNs intrinsically cross the BBB or enter brain parenchyma after intranasal administration. The formulation, cargo loading, particle composition, dosing schedule, and disease model may all influence delivery efficiency and therapeutic outcome.103,104,112
For intranasal PELNs, future studies should distinguish between nasal epithelial uptake, olfactory bulb accumulation, cerebrospinal fluid exposure, vascular absorption, and true neuronal or glial delivery. Evidence should ideally include time-resolved biodistribution imaging, tissue clearing or high-resolution microscopy, cell-type-specific colocalization, quantitative cargo detection, and functional rescue in disease-relevant models. Without such validation, intranasal brain-associated signals may reflect mucosal retention, vascular redistribution, or imaging artifacts rather than efficient CNS cargo delivery.103–108,112
Intravenous Administration
Intravenous administration bypasses the gastrointestinal tract and provides direct systemic exposure, but it also exposes PELNs to plasma proteins, complement components, immune cells, and rapid clearance by the mononuclear phagocyte system. After intravenous injection, nanoscale vesicles are commonly distributed to clearance organs such as the liver, spleen, lung, and kidney, depending on particle size, surface charge, lipid composition, protein corona formation, dose, and disease state.77,102,104,105,108,113 Therefore, systemic administration may be useful for evaluating whole-body biodistribution and immune interaction, but it does not automatically confer efficient CNS delivery.
Catharanthus roseus-derived nanovesicles have been reported to display distinctive lipid composition, including ether-linked lipid species, and to interact with immune-related tissues and cells in preclinical studies.38 Other plant-derived nanovesicle systems have also shown route- and context-dependent biodistribution or immune-cell-associated interactions. For example, ginseng-derived nanoparticles were reported to modulate macrophage polarization in a melanoma model, while grapefruit-derived nanovectors were shown to exploit an activated leukocyte trafficking pathway to deliver therapeutic agents to inflammatory tumor sites.29,44 These findings suggest that lipid composition, surface molecules, immune-cell interactions, and disease-associated vascular environments may influence stability, immune recognition, and organ distribution in selected PELN or plant-derived nanovector preparations. However, apparent tissue-targeting tendencies should be interpreted cautiously because tissue accumulation may also reflect vascular exposure, macrophage uptake, clearance organ retention, inflammation-associated permeability, or nonspecific nanoparticle biodistribution.38,44,104,105,108
Compared with oral and intranasal routes, intravenous administration allows more controlled dosing and more direct pharmacokinetic analysis. It is therefore useful for evaluating circulation time, tissue distribution, clearance pathways, immune recognition, and off-target accumulation. However, systemic injection may increase the risk of complement activation, immune cell uptake, organ retention, and dose-dependent toxicity, especially during repeated administration. For neurodegenerative disease applications, intravenous PELNs should be evaluated with whole-body biodistribution imaging, blood clearance kinetics, organ histology, inflammatory markers, brain perfusion controls, and cell-type-specific localization in the CNS.102,104–108
Cellular Uptake and Intracellular Trafficking
Cellular uptake is a key step linking biodistribution to biological activity, but uptake mechanisms of PELNs remain incompletely defined. Available evidence suggests that PELN internalization may vary according to plant source, vesicle size, lipid composition, surface proteins, glycan structures, protein or biomolecular corona, recipient cell type, and route of administration. Potential uptake mechanisms include clathrin-mediated endocytosis, caveolin-mediated endocytosis, lipid raft-associated uptake, macropinocytosis, phagocytosis, and membrane fusion-like processes.35,60,61,77,98,102,104,113,114 However, direct pathway validation remains limited for many PELN preparations.
The recipient cell type is particularly important in CNS-related studies. Intestinal epithelial cells and macrophages may dominate oral-route interactions, whereas brain endothelial cells, pericytes, neurons, astrocytes, and microglia are more relevant to BBB transport and CNS parenchymal delivery. Microglia-like cells and macrophage-related models have been used to evaluate anti-inflammatory responses to selected plant-derived vesicle-like particles, including Allium tuberosum-derived nanoparticles and ginger-derived exosome-like nanoparticles.115,116 In addition, plant-derived exosome-like nanoplatforms have been explored for biomacromolecule delivery and mitochondrial protection in Parkinson’s disease-related models.117 These findings support the biological relevance of cell-type-specific uptake studies, but they do not yet establish a universal uptake pathway for all PELNs.
After internalization, PELNs or their cargoes may traffic through early endosomes, late endosomes, lysosomes, recycling compartments, or cytosolic release pathways. For therapeutic delivery, intracellular trafficking is as important as uptake because vesicles that remain trapped in endolysosomal compartments may have limited access to cytosolic or nuclear targets. Future studies should therefore combine pharmacological inhibitors, genetic knockdown of uptake mediators, live-cell imaging, endosomal and lysosomal colocalization, cargo-release assays, and quantitative uptake analysis. Such approaches would help distinguish surface adhesion from true internalization and clarify whether PELN cargoes can escape degradation and reach functional intracellular compartments.77,102,104,113,114
BBB Transport Mechanisms and Evidence Hierarchy
The BBB remains one of the major obstacles for CNS drug delivery because it restricts the entry of most therapeutic agents into the brain while maintaining neural homeostasis.13,103,105 For PELNs, several potential BBB-related mechanisms have been proposed, including receptor-mediated transcytosis, adsorptive-mediated transcytosis, lipid raft-associated uptake, endothelial endocytosis, and indirect gut-brain axis modulation.102–107 However, current evidence is still heterogeneous, and these mechanisms should be described as proposed or context-dependent rather than established universal pathways.
For systemic administration, PELNs may interact with brain microvascular endothelial cells through surface lipids, glycans, proteins, or adsorbed biomolecular coronas. These interactions could theoretically support adsorptive uptake or receptor-associated transport, but direct evidence for specific transporter engagement remains limited. Claims involving receptors such as transferrin receptor or glucose transporter 1 (GLUT1) should therefore be supported by blocking experiments, receptor knockdown, competitive inhibition, or receptor-specific imaging.103–107 Similarly, lipid raft-associated uptake or membrane fusion-like processes should be distinguished from nonspecific endothelial adhesion or endocytic uptake.
A practical evidence hierarchy is needed to evaluate BBB transport claims. The lowest level of evidence includes in vitro uptake by endothelial or neural cells, which demonstrates cellular interaction but not barrier crossing. A higher level includes Transwell or microfluidic BBB models with validated tight junction integrity, transendothelial electrical resistance, and paracellular permeability controls. Stronger evidence requires in vivo biodistribution imaging with vascular perfusion, quantitative brain-to-blood ratios, cellular colocalization in brain parenchyma, and exclusion of signal from blood vessels or meninges. The strongest evidence combines brain delivery of intact vesicles or defined cargoes with pharmacodynamic effects in disease-relevant CNS models.105–108
BBB-related results should also account for disease state and model artifacts. Neuroinflammation, tumors, ischemia, and neurodegenerative pathology may disrupt BBB integrity and increase nanoparticle entry independently of active transport. Therefore, apparent brain accumulation may reflect barrier leakage, inflammatory vascular permeability, peripheral immune cell trafficking, or residual blood signal rather than receptor-mediated transcytosis. Studies should include healthy and diseased controls, vascular perfusion, endothelial and parenchymal markers, and, when possible, live imaging or quantitative mass balance analysis.13,105,107,108,118
Selected studies provide proof-of-concept that plant-derived nanovesicles or plant-derived lipid nanovectors can be adapted for CNS-related delivery. Grapefruit-derived nanovectors delivered miR-17 via the intranasal route in a brain tumor model, while plant-derived exosome-like platforms have also been explored for biomacromolecule delivery and mitochondrial protection in Parkinson’s disease-related models.112,117 These examples support the broader feasibility of plant-derived nanoscale delivery systems for CNS applications. Nevertheless, they should be interpreted as source-specific and formulation-specific evidence, not as proof that all PELNs can intrinsically cross the BBB.
Overall, administration route, biodistribution, cellular uptake, intracellular trafficking, and BBB model selection jointly determine how CNS-targeting evidence should be interpreted. Oral PELNs may act largely through intestinal and gut-brain axis mechanisms; intranasal delivery may bypass parts of the BBB through nose-to-brain pathways; and intravenous delivery enables systemic exposure but faces strong clearance and BBB restrictions. Therefore, future PELN studies should integrate route-specific pharmacokinetics, quantitative biodistribution, validated BBB models, and functional cargo delivery assays before making claims of CNS targeting. This route- and evidence-based framework provides the basis for discussing the molecular therapeutic mechanisms of PELNs in neurodegenerative diseases.
Disease-Shared Therapeutic Mechanisms of PELNs in Neurodegeneration
Neurodegenerative diseases differ in their initiating triggers, affected neuronal populations, and clinical manifestations, but they share several convergent pathological processes, including chronic neuroinflammation, redox imbalance, mitochondrial dysfunction, impaired proteostasis, synaptic failure, impaired neural repair, and systemic immune-metabolic dysregulation.8–11,27,100,102 However, the extent to which PELNs can engage these mechanisms depends strongly on their plant source, cargo composition, engineering strategy, and administration route. As shown in Figure 3, orally, intranasally, and intravenously administered PELNs may follow distinct biological fates. Specifically, orally administered PELNs may act primarily through intestinal epithelial cells, gut microbiota, mucosal immune cells, and gut–brain axis-related immune, vagal, and metabolic signaling, as illustrated in Figure 3A. Intranasal delivery may partially bypass the BBB through olfactory and trigeminal nerve-associated pathways, thereby providing a potential non-invasive nose-to-brain transport route, as shown in Figure 3B. Intravenously administered PELNs enter systemic circulation but may undergo protein corona formation, liver and spleen accumulation, and mononuclear phagocyte system-mediated clearance, which can reduce effective brain exposure, as summarized in Figure 3C. Proposed BBB-associated transport mechanisms include receptor-associated transcytosis, lipid-dependent membrane interaction, and endocytosis, followed by potential uptake by CNS-resident cells, as depicted in Figure 3D. Therefore, the biological effects of PELNs in neurodegenerative models should be interpreted in a route-dependent and evidence-dependent manner. Therefore, the biological effects of PELNs in neurodegenerative models should be interpreted in a route-dependent and evidence-dependent manner.
 |
Figure 3 Route-dependent delivery, systemic biodistribution, and putative blood–brain barrier (BBB) transport of plant-derived exosome-like nanoparticles (PELNs). The biological fate of PELNs is strongly influenced by the administration route. (A) Orally administered PELNs may act primarily through the gastrointestinal tract by interacting with intestinal epithelial cells, gut microbiota, and mucosal immune cells, thereby modulating the gut–brain axis through immune, vagal, and metabolic signaling. (B) Intranasal delivery may enable partial blood–brain barrier (BBB) bypass via olfactory and trigeminal nerve-associated pathways, offering a non-invasive strategy for nose-to-brain transport. (C) Intravenously administered PELNs enter systemic circulation but are subject to protein Corona formation, liver and spleen accumulation, and mononuclear phagocyte system (MPS)-mediated clearance, which may reduce effective brain exposure. (D) Proposed BBB transport mechanisms include receptor-associated transcytosis, lipid-dependent membrane interaction, and endocytosis, followed by uptake by microglia, astrocytes, and neurons. Because direct evidence for intact PELN penetration across the BBB remains limited, future studies should combine quantitative pharmacokinetics, biodistribution analysis, co-localization of vesicle membrane and cargo, and pathway-inhibition experiments to distinguish true vesicle transport from free dye or degraded cargo signals. Created with BioRender.com. |
AD remains the most extensively investigated model for PELN-based neurotherapy, whereas PD, HD, ALS, ischemic stroke, and neural injury models currently provide varying levels of mechanistic or preclinical support. The present section therefore discusses disease-shared therapeutic mechanisms, while the following section evaluates disease-specific evidence and therapeutic applications. Figure 4 provides an AD-focused schematic depicting representative PELNs after putative BBB-associated CNS access and their downstream modulation of AD-related neuroinflammation, Aβ and Tau pathology, neural repair, and antioxidant signaling. In this framework, Figure 4A specifically highlights the proposed BBB-associated CNS access routes of PELNs and emphasizes that direct in vivo BBB penetration, transport of intact vesicles across the BBB, and quantitative brain biodistribution remain insufficiently validated for most plant sources. Downstream AD-relevant mechanisms are further summarized in Figure 4B–E as representative, source-associated modules involving microglial inflammatory regulation, Aβ/Tau-associated proteinopathy, neural repair and synaptic plasticity, and antioxidant signaling.
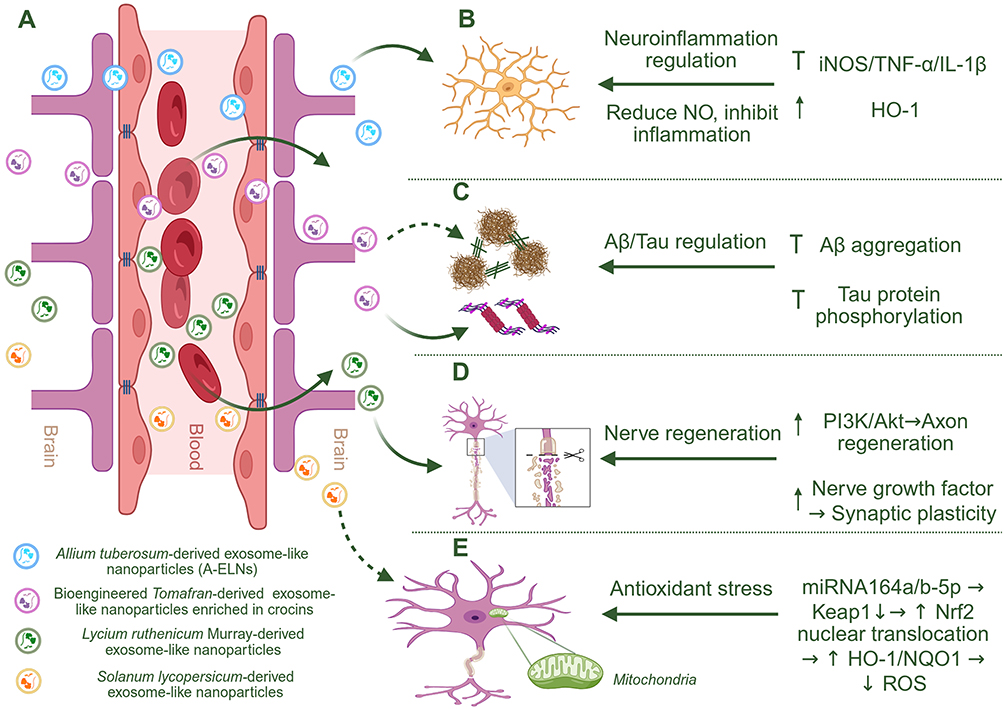 |
Figure 4 Proposed Alzheimer’s disease (AD)-relevant neuroprotective mechanisms of selected plant-derived exosome-like nanoparticles (PELNs). Schematic overview of three representative PELNs derived from Allium tuberosum, Solanum lycopersicum, and Lycium ruthenicum Murray, highlighting their potential blood–brain barrier (BBB)-associated transport routes and their reported regulation of AD-related pathological processes in the central nervous system (CNS). (A) PELNs may access the CNS through BBB-associated transport routes; however, direct in vivo BBB penetration, intact vesicle transport, and quantitative brain biodistribution remain to be further validated for most plant sources. (B) Allium tuberosum-derived ELNs suppress microglia-mediated neuroinflammatory responses by downregulating inducible nitric oxide synthase (iNOS), tumor necrosis factor-α (TNF-α), and interleukin-1β (IL-1β), while upregulating heme oxygenase-1 (HO-1), thereby reducing nitric oxide (NO) production and neuroinflammation. (C) Bioengineered Tomafran-derived ELNs enriched in crocins are proposed to attenuate AD-related Aβ/Tau pathology, particularly Tau-related toxicity, although direct evidence for reducing Aβ aggregation remains limited and requires further validation. (D) Lycium ruthenicum Murray-derived ELNs may promote axonal repair and synaptic plasticity through activation of phosphoinositide 3-kinase/protein kinase B (PI3K/Akt) signaling and upregulation of nerve growth factor. (E) Solanum lycopersicum-derived ELN-associated miRNA164a/b-5p suppresses Kelch-like ECH-associated protein 1 (Keap1) and activates nuclear factor erythroid 2–related factor 2 (Nrf2) signaling, leading to increased Nrf2 nuclear translocation, upregulation of downstream antioxidant enzymes, including HO-1 and NAD(P)H:quinone oxidoreductase 1 (NQO1), and reduced reactive oxygen species (ROS), thereby mitigating oxidative stress; however, this evidence currently derives mainly from vascular injury models rather than direct AD models. In the schematic, dashed arrows indicate proposed or indirect associations, including mechanisms supported by non-AD models or evidence requiring further validation; ↑ indicates upregulation or activation, ↓ indicates downregulation or reduction, and blunt-ended lines indicate inhibition or suppression. Created with BioRender.com. |
Neuroinflammation and Glial Immune Regulation
Neuroinflammation is a central pathological feature of AD, PD, ischemic injury, traumatic neural injury, and other neurodegenerative conditions. It is largely mediated by activated microglia, reactive astrocytes, infiltrating immune cells, and sustained production of inflammatory mediators, including nitric oxide (NO), tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β), interleukin-6, and inducible nitric oxide synthase (iNOS). 10, 20, 67 Persistent activation of nuclear factor-κB (NF-κB), inflammasome-related pathways, and glial immune responses can amplify neuronal injury, synaptic dysfunction, oxidative stress, and BBB disruption.10,27,100,119
Several PELN preparations have been reported to attenuate inflammatory responses in immune or neural-related models. Allium tuberosum-derived exosome-like nanoparticles (A-ELNs) provide a representative example. A-ELNs reduced iNOS expression and inflammatory cytokine production in lipopolysaccharide (LPS)-stimulated BV-2 microglia and MG-6 cells, while increasing heme oxygenase-1 (HO-1), a cytoprotective enzyme associated with antioxidant and anti-inflammatory responses.115 These findings support the concept that PELNs may function not merely as passive carriers but also as biologically active immunomodulatory nanoparticles. Moreover, dexamethasone-loaded A-ELNs further enhanced the suppression of NO production and inflammatory responses, indicating that endogenous PELN bioactivity and exogenous anti-inflammatory drug loading may act synergistically.115
Beyond general cytokine suppression, PELNs may also regulate upstream inflammatory hubs. Ginger-derived exosome-like nanoparticles have been shown to inhibit NLR family pyrin domain containing 3 (NLRP3) inflammasome activation, a critical event involved in IL-1β maturation and inflammatory pyroptosis.116 Because NLRP3 activation contributes to microglial dysfunction in AD and PD, this pathway provides a mechanistic bridge between PELN bioactivity and neurodegenerative inflammation.116,119 In addition, ginseng-derived nanoparticles have been reported to modulate macrophage polarization in peripheral immune contexts, suggesting that some PELNs may influence neuroinflammation indirectly through systemic immune remodeling.29
The relevance of inflammatory and antioxidant pathway crosstalk is also supported by comparative pharmacological neuroprotection studies. For example, dl-3-n-butylphthalide and edaravone dexborneol improved neurological function and suppressed neuroinflammation and oxidative stress in cerebral ischemic stroke rats through NF-κB/iNOS and kelch-like ech-associated protein 1 (Keap1)/ nuclear factor erythroid 2-related factor 2 (Nrf2) -related pathways.120 Similarly, edaravone dexborneol protected neurological function in cerebral ischemia-reperfusion injury by targeting Nrf2/ antioxidant response element (ARE) and NF-κB/ absent in melanoma 2 (AIM2) pathways.121 These studies should not be presented as direct PELN evidence, but they strengthen the biological rationale for targeting inflammatory-redox axes that PELNs may also modulate.
However, the anti-inflammatory effects of PELNs should be interpreted cautiously. Different PELN preparations contain distinct lipids, proteins, miRNAs, and phytochemicals, and their effects may depend on plant source, purification method, dose, administration route, and recipient cell type. Moreover, reductions in inflammatory markers do not necessarily prove direct CNS delivery or glial-cell-specific targeting. Therefore, future studies should distinguish whether PELNs act on microglia, astrocytes, endothelial cells, peripheral macrophages, or gut-associated immune cells, and should include appropriate controls such as vesicle-free plant extracts, disrupted vesicles, and cargo-depleted preparations.
Redox Homeostasis and Mitochondrial Protection
Oxidative stress and mitochondrial dysfunction are closely linked to neuronal vulnerability in neurodegenerative diseases. Excessive reactive oxygen species (ROS) production, impaired antioxidant defenses, mitochondrial membrane potential loss, defective mitophagy, and altered energy metabolism can promote neuronal apoptosis, synaptic failure, protein aggregation, and glial activation.8,9 Therefore, redox homeostasis and mitochondrial protection represent disease-shared therapeutic axes for PELNs.
PELNs may regulate oxidative stress through endogenous antioxidant cargos, vesicle-associated lipids, plant-derived small molecules, and miRNAs. Citrus reticulata-derived extracellular vesicles have been reported to possess antioxidant activity and drug delivery potential, supporting the concept that fruit-derived vesicles can carry redox-active components.96 Solanum lycopersicum-derived exosome-like nanovesicles were shown to alleviate vascular injury through the Keap1/Nrf2 pathway by delivering miR-164a, thereby promoting Nrf2 activation and downstream antioxidant gene expression.97 Although this model is not a neurodegenerative disease model, it provides mechanistic support for the ability of plant vesicle cargos to regulate redox-responsive transcriptional programs.
In the AD-focused schematic shown in Figure 4E, PELN-associated miRNA164a/b-5p is presented as a representative mechanism that activates Nrf2 signaling and promotes downstream antioxidant enzymes, including HO-1 and NAD(P)H:quinone oxidoreductase 1 (NQO1). More broadly, Nrf2-mediated redox regulation may represent a disease-shared neuroprotective axis, although its validation remains source- and model-dependent.96,97,122,123
In neuronal injury models, Lycium ruthenicum Murray-derived exosome-like nanovesicles inhibited Aβ-induced apoptosis and oxidative stress in HT22 and PC12 cells, partly through modulation of mitogen-activated protein kinase (MAPK) and phosphoinositide 3-kinase/protein kinase B (PI3K/Akt) signaling pathways.122,123 These pathways are closely linked to neuronal survival, mitochondrial integrity, and stress resistance. Activation of Nrf2/HO-1 signaling may further contribute to restoration of antioxidant defenses and reduction of Aβ-associated oxidative injury.122,123
Mitochondrial quality control is another critical point of intervention. In PD-related models, engineered plant-derived exosome-like nanoplatforms have been used to deliver brain biomacromolecules and ameliorate mitochondrial dysfunction.117 This strategy suggests that PELNs can be adapted not only for small-molecule delivery but also for functional biomacromolecule delivery targeting mitochondrial repair. Potential mechanisms include regulation of apoptosis-associated proteins, preservation of mitochondrial membrane potential, and activation of PTEN-induced kinase 1 (PINK1)/Parkin-dependent mitophagy.117 Since mitophagy failure contributes to both AD and PD, mitochondrial repair may be one of the most disease-shared mechanisms of PELN-based neuroprotection.8,9,117
Fruit-derived PELNs may also provide indirect evidence that vesicle-associated plant bioactives can regulate redox and barrier-related injury pathways. For example, pomegranate-derived exosome-like nanovesicles enriched in ellagic acid were reported to alleviate gut leakage and liver injury in a metabolic disease model.99 However, because this evidence derives from gut–liver and systemic metabolic injury rather than AD or CNS neurodegeneration models, pomegranate-derived vesicles should currently be interpreted only as indirect support for antioxidant and barrier-protective activity, not as direct evidence of anti-AD efficacy.
Nevertheless, antioxidant and mitochondrial effects should not automatically be attributed to intact PELNs. Plant extracts contain abundant polyphenols, flavonoids, pigments, and other redox-active compounds. Therefore, future studies should compare intact PELNs with vesicle-depleted extracts, disrupted vesicles, purified phytochemicals, and cargo-depleted vesicles. Such controls are necessary to determine whether observed redox effects arise from the vesicle structure, the enclosed cargo, co-isolated phytochemicals, or a combination of these components.
Proteinopathy Modulation and Proteostasis
Pathological protein aggregation is a hallmark of several neurodegenerative diseases. AD is characterized by extracellular Aβ deposition and intracellular hyperphosphorylated Tau accumulation; PD is associated with α-synuclein aggregation; HD involves mutant huntingtin accumulation; and ALS may involve TAR DNA-binding protein 43 (TDP-43) pathology, superoxide dismutase 1 (SOD1) misfolding or aggregation, or proteostatic disruption associated with chromosome 9 open reading frame 72 (C9orf72) repeat expansion.6,7,124,125 Therefore, proteinopathy modulation and proteostasis restoration should be discussed as disease-shared mechanisms rather than as AD-specific events.
In an AD-like okadaic acid-induced cellular model, bioengineered Tomafran-derived exosome-like nanoparticles enriched in crocins showed neuroprotective potential against Tau phosphorylation-associated toxicity.126 Tomafran-derived exosomes protected SH-SY5Y cells from okadaic acid-induced injury, whereas wild-type tomato-derived exosomes did not significantly protect SH-SY5Y cells from okadaic acid-induced injury, suggesting that crocin enrichment may contribute to the observed neuroprotective effect.126 Because okadaic acid induces Tau hyperphosphorylation through phosphatase inhibition, these findings are best interpreted as evidence for a Tau-related AD-like proteinopathy module rather than direct evidence for Aβ aggregation or clearance.126 In the AD-focused context, this Tau-related effect corresponds to the proteinopathy module illustrated in Figure 4C. However, direct validation of Tomafran-derived ELNs in Aβ aggregation, Aβ clearance, animal AD models, and other proteinopathies, including α-synuclein, mutant huntingtin, TDP-43, or SOD1 pathology, remains insufficient.
Plant lipid species may also participate in amyloid regulation. Plant-derived sphingolipids, particularly konjac-derived glucosylceramides or ceramides, have been reported to promote the release of host neuronal extracellular vesicles, including exosomes, and alleviate Aβ pathology in an AD mouse model, suggesting that plant lipid species may indirectly modulate amyloid clearance through EV-dependent pathways.127,128 Although these findings do not demonstrate that native PELNs themselves are the sole therapeutic agent, they support the broader rationale that plant lipid cargos or plant-derived lipid metabolites may influence EV-associated amyloid metabolism and proteostasis.
Exosome-based systems can also deliver plant-derived anti-proteinopathy compounds. An exosome-mediated co-delivery system carrying curcumin and methylene blue reduced abnormal Tau phosphorylation through Akt/glycogen synthase kinase-3β (GSK-3β) signaling and improved cognitive outcomes in an AD mouse model.129 Curcumin has also been reported to attenuate Tau hyperphosphorylation and neurotoxicity through modulation of stress and kinase pathways.130,131 Similarly, epigallocatechin gallate (EGCG), a major polyphenol from Camellia sinensis, can redirect Aβ aggregation into less toxic conformations, remodel mature Aβ and α-synuclein fibrils, and interfere with Tau aggregation or phosphorylation.132–135 These examples support the broader concept that vesicle-based delivery may enhance the neurotherapeutic value of plant-derived anti-proteinopathy compounds, although not all of these systems are native PELNs.
Stress-response pathways may further connect plant bioactives with proteostasis. Phytocompounds that modulate heat shock factor 1 (HSF1) and related chaperone systems may influence protein folding, aggregate handling, and neuronal stress resistance.31 However, evidence for direct PELN-mediated regulation of chaperone networks remains limited. Therefore, future studies should distinguish direct PELN evidence from supportive plant-bioactive or exosome-delivery evidence.
Neural Repair and Synaptic Plasticity
Neurodegeneration is not only a process of neuronal loss but also a failure of repair. Synaptic dysfunction, impaired axonal regeneration, reduced neurotrophic support, and maladaptive glial scarring all contribute to irreversible functional decline. PELNs may support neural repair through several complementary mechanisms, including promotion of neuronal differentiation, enhancement of axonal outgrowth, regulation of neurotrophic signaling, suppression of apoptosis, and remodeling of the inflammatory microenvironment.84,136,137
PELNs have been reported to transfer microRNAs that stimulate neural differentiation of stem cells in vitro and in vivo, supporting their potential role as natural regulators of neurodevelopmental and regenerative pathways.136 This finding is particularly relevant for neurodegenerative and injury-related disorders because miRNA-mediated regulation can influence cell fate, neurite extension, synaptic maturation, and survival signaling.136
Lycium barbarum L.-derived exosome-like nanoparticles loaded with isoliquiritigenin and incorporated into 3D-printed biomimetic scaffolds promoted spinal cord injury repair, suggesting that PELNs can be integrated with tissue-engineering platforms to provide both biological and structural support.96 In this hybrid system, plant-derived vesicles may regulate the post-injury inflammatory microenvironment, reduce excessive apoptosis or autophagy, and support axonal regeneration.95,138–140 This example is especially valuable because it moves PELNs beyond simple drug delivery and positions them as components of bioactive regenerative systems.
PELNs may also interact with classical pro-survival and pro-regenerative pathways, including PI3K/Akt, MAPK/extracellular signal-regulated kinase (ERK), and mammalian target of rapamycin (mTOR) signaling.138–140 These pathways regulate neuronal survival, axonal extension, synaptic plasticity, and cellular stress responses. In addition, neurotrophic factors such as nerve growth factor, brain-derived neurotrophic factor, and glial cell-derived neurotrophic factor may be indirectly modulated by PELN treatment or by plant bioactives delivered through vesicle systems.29,136,141,142 In the AD-oriented schematic, Figure 4D depicts axonal repair and synaptic plasticity as representative downstream outcomes associated with increased PI3K/Akt signaling and nerve growth factor-related repair pathways. These pathways may also be relevant to broader neural injury and regeneration models, but disease-specific validation remains necessary.95,136,138–140
Hydroxysafflor yellow A has been shown to promote neurogenesis and axon regeneration after traumatic brain injury, as supported by metabolomics-based analysis.143 Although this finding is not direct evidence for PELN-mediated neural repair, it supports the broader rationale for investigating whether PELN-delivered or PELN-associated plant bioactives can modulate regenerative responses in neural repair models. Similarly, 3D-bioprinted models of adult neural stem cell microenvironments in AD may provide future platforms for evaluating PELN effects on neural stem cells, glial interactions, and disease-relevant regenerative niches.144
Recent biohybrid platforms further illustrate the regenerative potential of vesicle-based strategies. Conductive microneedles loaded with polyphenol-engineered exosomes reshaped diabetic neurovascular niches and promoted chronic wound healing, partly by improving neurovascular interactions and tissue repair signaling.145 Although this model is not a CNS neurodegeneration model, it provides conceptual support for combining natural vesicle bioactivity with biomaterial-guided delivery.84,137,145
Gut-Brain Axis and Systemic Regulation
The gut–brain axis is increasingly recognized as a disease-modifying pathway in neurodegeneration. Intestinal barrier dysfunction, microbiota imbalance, microbial metabolites, systemic inflammation, vagal signaling, and immune-cell trafficking can influence neuroinflammation and neuronal vulnerability.11,28,46,64 For orally administered PELNs, the gastrointestinal tract is likely not only an absorption site but also a primary therapeutic interface.
Edible plant-derived exosome-like nanoparticles can mediate interspecies communication between plant and mammalian gut host cells, providing early evidence that plant vesicle-like structures can interact with intestinal physiology.61 Ginger-derived exosome-like nanoparticles have shown therapeutic activity in inflammatory bowel disease models,35 while grapefruit-derived exosome-like nanoparticles and other plant-derived exsome-like systems have been explored for immune and inflammatory targeting.35,43,44 These studies support the concept that orally delivered PELNs can regulate intestinal epithelial cells, macrophages, and inflammatory pathways before any potential CNS exposure occurs.
Microbiota modulation may be particularly important for neurological disorders. Plant-derived exosomal miRNAs can shape the gut microbiota,64 and citrus exosome-like nanoparticles have been shown to reverse high-fat diet-induced microbiota dysbiosis and alter short-chain fatty acid production.46 In a PD-related context, oral administration of antimicrobial nucleic acid-loaded exosome-like nanovesicles modulated intestinal flora and improved PD-related phenotypes, providing one of the most direct examples linking PELNs, gut microbiota, and neurodegenerative outcomes.28
Allium tuberosum-derived exosome-like nanoparticles also exhibit anti-inflammatory properties in colitis and can modify the gut microbiome, supporting a systemic immune-metabolic role for PELNs.111 In addition, microRNA-enriched plant-derived exosomes have been reported to alleviate colitis by modulating systemic immunity, metabolic homeostasis, and gut microbiota.110 Although these findings are not direct evidence of neuroprotection, they strengthen the biological plausibility that oral PELNs may regulate the gut–immune–metabolic axis relevant to neurodegeneration.
Microbial and hepatic biotransformation may provide an important conceptual framework for understanding the oral bioactivity of plant-derived nanotherapeutics. A recent study showed that low-bioavailability herbal products can exert neurological effects through gut microbial and hepatic biotransformation, generating metabolites with improved absorption and disease-relevant bioactivity.12 This concept is relevant to PELNs because their biological effects after oral administration may arise not only from vesicle-mediated cargo delivery, but also from released phytochemicals, microbiota-derived metabolites, and immune-metabolic regulation.12
Importantly, gut–brain axis regulation should not be interpreted as equivalent to direct CNS delivery. As summarized in Figure 3A, orally administered PELNs may act primarily through intestinal epithelial cells, gut microbiota, mucosal immune cells, metabolic mediators, and vagal or systemic immune pathways, rather than requiring intact vesicle penetration into the brain.11,12,61,64,110 Therefore, future studies should combine microbiome analysis, metabolomics, intestinal permeability assays, systemic cytokine profiling, quantitative biodistribution, and CNS-specific endpoints to distinguish gut-mediated systemic regulation from direct brain delivery.11,12,105
Disease-Specific Evidence and Therapeutic Applications
The mechanisms described above provide a mechanism-based framework for understanding PELN-mediated neuroprotection across neurological disease contexts. However, the level of experimental support varies markedly among disease models. AD is currently one of the most extensively investigated models for PELN-related neurotherapy, particularly in relation to oxidative stress regulation, neuroinflammation, Aβ/Tau-associated pathology, and neural repair. PD-related evidence is also emerging, especially in the context of gut–brain axis regulation, mitochondrial protection, and engineered cargo delivery. In contrast, HD and ALS remain largely conceptual application areas, with limited direct PELN-specific evidence. Ischemic stroke, spinal cord injury, and peripheral nerve injury provide complementary neurological injury models for evaluating PELN-mediated neuroprotection and regenerative repair. A structured overview of representative disease contexts, plant sources, mechanistic pathways, experimental models, and therapeutic implications is provided in Table 4.
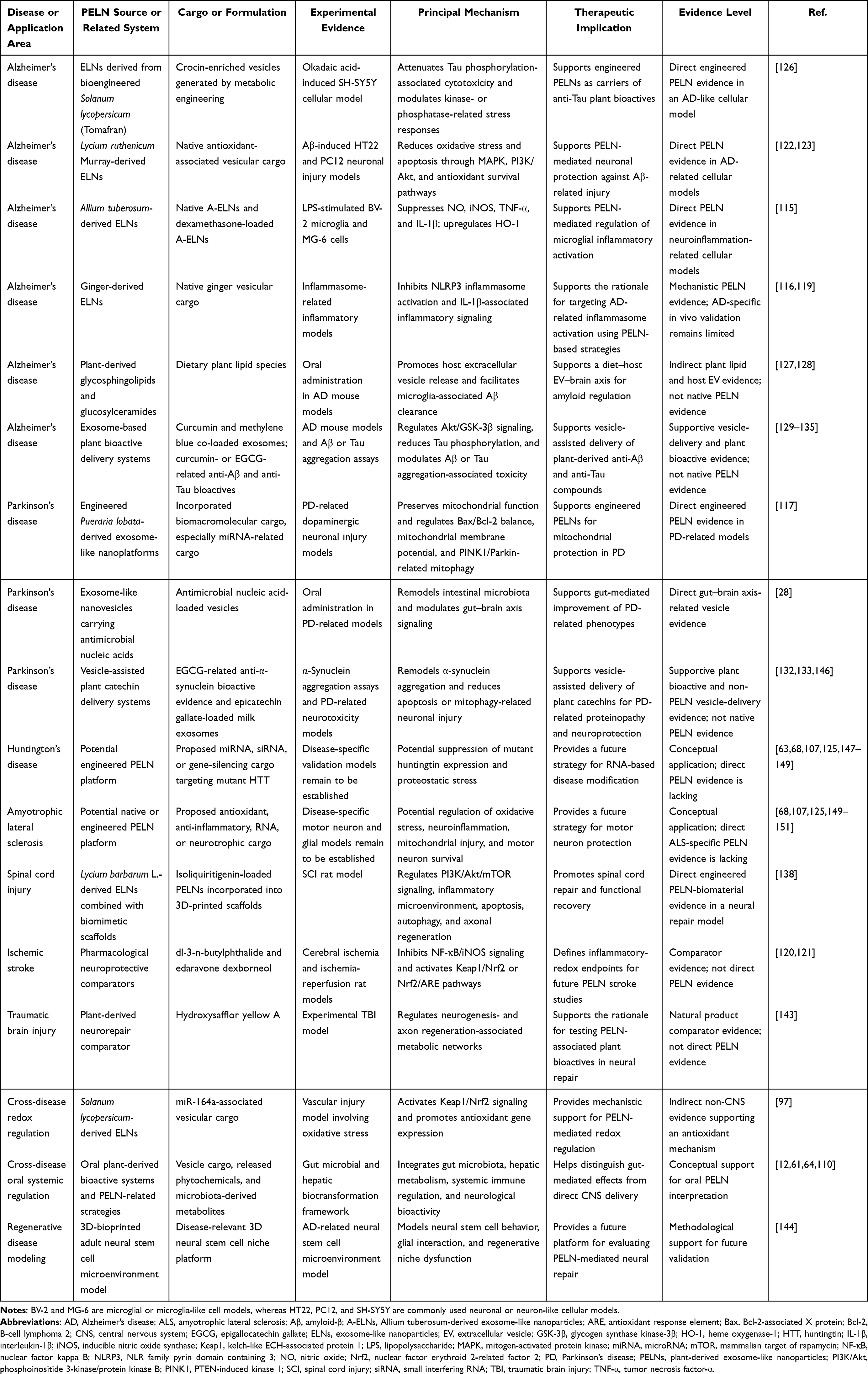 |
Table 4 Therapeutic Mechanisms and Applications of PELNs in Neurodegenerative Disease-Related Models: an Evidence-Stratified Overview |
Alzheimer’s Disease as the Leading Model for PELNs-Based Neurotherapy
AD is currently the most extensively discussed neurodegenerative indication for PELNs because its pathology integrates multiple PELN-relevant mechanisms, including Aβ deposition, Tau hyperphosphorylation, chronic neuroinflammation, oxidative stress, mitochondrial dysfunction, synaptic failure, and gut–brain axis dysregulation.3,5–9 These overlapping mechanisms make AD a useful leading model for evaluating the dual role of PELNs as intrinsic bioactive agents and natural delivery platforms.
In Aβ-related cellular models, ELNs derived from Lycium ruthenicum Murray inhibited Aβ-induced apoptosis and oxidative stress in neuronal cells, suggesting that these PELNs may protect neurons by activating survival pathways and reducing ROS burden.122,123 These findings are consistent with the redox and mitochondrial protection mechanisms discussed above. However, most of these data are derived from cellular models, and additional in vivo studies are needed to confirm whether intact PELNs reach AD-relevant brain regions at therapeutically meaningful concentrations.
Bioengineered Tomafran-derived exosome-like nanoparticles enriched in crocins provide another AD-relevant example. In an okadaic acid-induced SH-SY5Y cellular model associated with Tau hyperphosphorylation, Tomafran-derived exosomes protected cells from okadaic acid-induced injury.126 These findings support a Tau phosphorylation-associated AD-like toxicity module, but they do not provide direct evidence that Tomafran-derived ELNs attenuate Aβ aggregation, Aβ clearance, or Aβ-related toxicity. The value of this model lies in its combination of plant metabolic engineering and vesicle-mediated delivery; however, future work should compare intact vesicles, free crocins, disrupted vesicles, and cargo-depleted vesicles to define the specific contribution of the PELN structure.126
Neuroinflammation is another major AD-relevant target. In microglia-like inflammatory models, A-ELNs suppress iNOS, TNF-α, IL-1β, and NO production, while upregulating HO-1.115 Ginger-derived exosome-like nanoparticles have also been reported to inhibit NLRP3 inflammasome activation, providing additional support for the anti-inflammatory potential of PELNs.116,119 These findings are consistent with the pathological importance of microglial activation and inflammasome signaling in AD and suggest that PELNs may attenuate neuroinflammatory amplification. The A-ELN-associated microglial anti-inflammatory module is illustrated in Figure 4B, whereas Figure 3 summarizes the route-dependent delivery uncertainty, including whether orally, intranasally, or intravenously administered PELNs can reach brain-resident glial cells as intact vesicles.
Plant sphingolipids and vesicle-related lipid biology may also contribute to AD therapy. Plant sphingolipids have been reported to promote extracellular vesicle release and alleviate Aβ pathologies in an AD mouse model.127,128 This suggests that plant lipid components may influence amyloid clearance and extracellular vesicle-associated proteostasis, although more work is needed to determine whether similar mechanisms operate in isolated PELNs from different plant sources.
Exosome-based delivery of plant bioactives further expands AD therapeutic strategies. Curcumin and methylene blue co-loaded exosomes reduced Tau phosphorylation through Akt/GSK-3β signaling and improved cognitive function in an AD mouse model.129 Curcumin alone has also been shown to modulate neurotoxicity and Tau-related signaling.130,131 Epigallocatechin gallate (EGCG) can remodel Aβ and α-synuclein fibrils, reduce amyloid toxicity, and interfere with Tau aggregation.132–135 These examples support the broader concept that vesicle-based delivery may improve the CNS therapeutic utility of plant-derived anti-proteinopathy compounds. However, not all of these systems are native PELNs, so they should be discussed as supportive nanomedicine evidence rather than direct PELN evidence.
Overall, AD provides the strongest mechanistic rationale for PELN-based neurotherapy, but the field still requires better evidence standards, including quantitative brain biodistribution, intact-vesicle tracking, dose-response studies, comparison with parent plant extracts, and behavioral validation in disease-relevant animal models.
Parkinson’s Disease
PD is characterized by dopaminergic neuron degeneration, α-synuclein aggregation, mitochondrial dysfunction, oxidative stress, neuroinflammation, and gut microbiota alterations.124,146 These features make PD an attractive indication for PELNs, particularly because PELNs may regulate mitochondrial quality control, deliver nucleic acid cargos, and modulate the gut–brain axis.27,28,117
Mitochondrial dysfunction is one of the most important therapeutic targets in PD. Engineered Pueraria lobata-derived exosome-like nanoplatforms have been reported to deliver incorporated biomacromolecular cargos, particularly miRNAs, to dopaminergic neuronal cells and ameliorate mitochondrial dysfunction in PD-related models.117 This strategy suggests that PELNs can be designed not only to carry small molecules but also to deliver functional RNA cargos that regulate mitochondrial repair, apoptosis, and mitophagy. Potential mechanisms include regulation of the Bcl-2-associated X protein/B-cell lymphoma 2 (Bax/Bcl-2) balance, PTEN-induced kinase 1/Parkin RBR E3 ubiquitin-protein ligase (PINK1/Parkin) signaling, mitochondrial membrane potential, and mitochondrial respiratory chain integrity.117
Gut microbiota modulation provides another disease-specific opportunity. Oral antimicrobial nucleic acid-loaded exosome-like nanovesicles improved PD-related outcomes by targeting intestinal flora, supporting the concept that PELNs may affect PD through gut–brain axis regulation rather than only through direct BBB penetration.28 This is particularly relevant because gastrointestinal dysfunction and microbiota changes often precede or accompany PD progression. As shown in Figure 3A, oral PELNs may primarily influence gut epithelial, microbial, immune, and metabolic pathways, whereas intranasal or intravenous administration may be more relevant for direct CNS delivery attempts.
Proteinopathy remains a more challenging target in PD. Although plant polyphenols such as EGCG can remodel α-synuclein fibrils and reduce aggregate-associated toxicity, direct evidence that native PELNs clear α-synuclein aggregates in vivo remains limited.132,133 In a related vesicle-assisted phytochemical delivery context, bovine milk-derived exosomes loaded with epicatechin gallate (ECG), another plant-derived catechin, have shown neuroprotective effects against PD-related toxicity, mainly through anti-apoptotic and anti-mitophagy mechanisms.146 Because this platform uses mammalian milk-derived exosomes rather than native PELNs, it should be considered supportive evidence for vesicle-assisted phytochemical delivery rather than direct proof that PELNs can clear α-synuclein aggregates or reverse PD proteinopathy in vivo.146
Therefore, PD represents an emerging but not yet mature application area for PELNs. The most convincing current directions are mitochondrial protection, gut microbiota regulation, and engineered cargo delivery. Future PD studies should quantify dopaminergic neuron survival, α-synuclein burden, striatal dopamine function, motor and non-motor behavior, vesicle biodistribution, and long-term safety after repeated administration.
Huntington’s Disease and Amyotrophic Lateral Sclerosis
Compared with AD and PD, direct evidence for PELNs in HD and ALS remains limited. Nevertheless, these disorders are conceptually relevant to PELN-based neuroprotection because they share several neurodegenerative hallmarks, including pathological protein aggregation, proteostatic disruption, mitochondrial or energy-metabolism impairment, oxidative stress, neuroinflammation, and progressive neuronal loss.125,152,153 HD is caused by a cytosine–adenine–guanine (CAG) trinucleotide repeat expansion in the huntingtin (HTT) gene, resulting in the production of mutant huntingtin with an expanded polyglutamine tract, followed by mutant huntingtin accumulation, aggregation, and neuronal dysfunction.147 ALS involves motor neuron degeneration associated with TDP-43 pathology, SOD1 mutation or aggregation, C9orf72-related pathology, glutamate excitotoxicity, neuroinflammation, oxidative stress, and mitochondrial injury.125,150,152
The potential value of PELNs in HD lies mainly in nucleic acid delivery. Because HD is caused by a defined genetic mutation, PELNs capable of protecting and delivering miRNAs, siRNAs, or other gene-silencing cargos could theoretically be engineered to reduce mutant huntingtin expression. Plant extracellular vesicles have been shown to transport small RNAs across kingdoms, and plant-derived nanovectors have been engineered to deliver therapeutic miRNAs in brain tumor models.40,112 More recent reviews further highlight the potential bioactivity of PELN-derived microRNAs and the feasibility of PELNs as nucleic acid delivery platforms.59,63,125 However, direct proof that PELNs can deliver anti-huntingtin RNA to striatal neurons in vivo is still lacking.
For ALS, PELNs may be relevant through anti-inflammatory, antioxidant, and neurotrophic mechanisms. Their ability to regulate Nrf2-related antioxidant responses, suppress inflammatory signaling, and potentially support neural repair could be useful for protecting vulnerable motor neurons.27,100,154 However, ALS has complex cellular pathology involving motor neurons, astrocytes, microglia, neuromuscular junctions, and peripheral immune components. Therefore, any PELN-based ALS strategy would require disease-specific delivery validation and cell-type-specific mechanistic studies.
At present, HD and ALS should be presented as future-oriented applications rather than established therapeutic indications. The strongest near-term research direction is to use PELNs as editable nanocarriers for RNA therapeutics, antioxidant cargos, or neurotrophic modulators, while systematically testing delivery to striatal neurons, spinal motor neurons, astrocytes, and microglia.
Ischemic Stroke, Nerve Injury, and Neural Repair Models
Ischemic stroke, traumatic brain injury, spinal cord injury, and peripheral nerve injury differ from chronic neurodegenerative diseases, but they share several pathological mechanisms with AD and PD, including oxidative stress, mitochondrial dysfunction, neuroinflammation, BBB disruption, neuronal apoptosis, impaired neurogenesis, and defective axonal regeneration.8,139,140 These models are therefore useful for evaluating the neuroprotective and regenerative potential of PELNs.
Direct PELN evidence in ischemic stroke remains limited. However, stroke models provide an important comparative framework for evaluating inflammatory-redox mechanisms that overlap with reported PELN-associated pathways, including Nrf2 activation, NF-κB inhibition, oxidative stress suppression, glial immune regulation, and neurovascular protection. A comparative pharmacological study of dl-3-n-butylphthalide and edaravone dexborneol in cerebral ischemic stroke rats showed that both agents improved neurological function and cognitive behavior while suppressing inflammation and oxidative stress through the NF-κB/iNOS and Keap1/Nrf2 pathways.120 Although this study does not provide direct evidence for PELNs, it identifies pathway-level endpoints that could be used to evaluate whether native or engineered PELNs exert comparable neuroprotective effects in ischemic injury models. Similarly, edaravone dexborneol protected neurological function in cerebral ischemia-reperfusion injury rats by targeting Nrf2/ARE and NF-κB/AIM2 pathways, further highlighting inflammatory-redox crosstalk as a shared mechanism in neural injury.121 Given that PELNs have been explored as natural nanocarriers for phytochemicals and therapeutic agents, future studies could investigate whether PELN-based formulations can deliver stroke-relevant neuroprotective compounds or modulate these pathways through endogenous vesicular cargos; however, direct PELN loading or delivery evidence for dl-3-n-butylphthalide or edaravone dexborneol remains to be established.
Plant-derived bioactives and exosome-based delivery systems also provide supportive evidence for ischemic and traumatic neural repair. Curcumin-loaded embryonic stem cell exosomes restored the neurovascular unit after ischemia-reperfusion injury, suggesting that exosome-mediated delivery of plant bioactives can improve ischemic neural repair.142 Lycium barbarum polysaccharide has been reported to protect against ischemia-related neuronal injury and to regulate autophagy-related pathways through PI3K/Akt/mTOR signaling.139,140,155 Although these studies do not all directly investigate native PELNs, they support the neuroprotective potential of plant-derived bioactive systems.
Among neural repair models, Lycium barbarum L.-derived exosome-like nanoparticles loaded with isoliquiritigenin and incorporated into 3D-printed biomimetic scaffolds provide a strong example of PELN-based regenerative therapy.138 This hybrid platform promoted spinal cord injury repair by combining the biological activity of plant-derived vesicles with the structural support of a biomimetic scaffold.138 The effects were associated with inflammatory modulation, regulation of PI3K/Akt/mTOR-related signaling, reduction of neuronal apoptosis or dysregulated autophagy, and enhancement of axonal regeneration.138–140
Hydroxysafflor yellow A has been shown to promote neurogenesis and axon regeneration after traumatic brain injury, providing supportive evidence that plant-derived bioactives can regulate neural repair processes.143 However, this evidence should be interpreted as a natural product-based neural repair comparator rather than direct evidence for PELN-mediated regeneration. It nevertheless supports the rationale for future studies testing whether PELN-delivered or PELN-associated plant bioactives can enhance neuroregeneration in neural injury models. In addition, 3D-bioprinted models of adult neural stem cell microenvironments in AD may serve as future platforms for evaluating PELN effects on neural stem cells, glial interactions, and disease-relevant regenerative niches.144
PELNs are also increasingly being explored as sustainable nanosystems for tissue engineering and regenerative medicine.84,137,145,156 Their low immunogenicity, natural lipid bilayer structure, and bioactive cargo profiles may allow them to regulate inflammatory microenvironments, promote angiogenesis, and support interactions among neural, vascular, and immune cells. However, most tissue-repair evidence comes from wound, diabetic ulcer, or scaffold-based models, and its direct relevance to CNS repair must be validated carefully.
Future studies should distinguish neuroprotection from true neural repair. Reduced cell death or inflammatory suppression does not necessarily indicate axonal regeneration, synaptic reconnection, or functional circuit recovery. Therefore, injury-related PELN studies should include axonal tracing, electrophysiology, synaptic markers, behavioral recovery, and long-term safety endpoints.
Evidence Gaps and Disease-Specific Limitations
Despite growing interest in PELNs for neurodegenerative and neural repair applications, the evidence base remains uneven. AD-related studies currently provide one of the most developed mechanistic contexts for PELN-based neuroprotection, particularly in relation to Aβ/Tau-associated pathology, oxidative stress reduction, and neuroinflammatory regulation.97,115,122,123,126 PD evidence is emerging, especially in relation to gut–brain axis modulation, mitochondrial protection, and engineered cargo delivery.28,117 In contrast, HD, ALS, and ischemic stroke remain less directly supported by PELN-specific data, and current discussions in these areas are largely extrapolated from shared neurodegenerative mechanisms, non-PELNs vesicle-delivery systems, or pharmacological neuroprotection comparators.27,100,102,125,152
A major limitation is the lack of standardized disease-specific evidence. Many studies use different plant sources, isolation methods, dosing metrics, administration routes, cell models, and outcome measures.21,22,25,33,34,66 As a result, it is difficult to compare efficacy across studies or determine whether observed benefits arise from intact PELNs, co-isolated plant compounds, soluble phytochemicals, vesicle-associated small RNAs, or engineered cargos.
Another important limitation concerns BBB and CNS delivery evidence. As shown in Figure 3, oral, intranasal, and intravenous administration routes differ substantially in biological barriers, biodistribution, clearance, and brain exposure. Oral PELNs may act primarily through the gut–brain axis; intranasal PELNs may provide partial nose-to-brain access through olfactory and trigeminal pathways; and intravenous PELNs may undergo protein corona formation, liver and spleen accumulation, and mononuclear phagocyte system-mediated clearance.11,28,102,105 Therefore, fluorescent brain signals should not be interpreted as definitive evidence of intact vesicle penetration unless supported by co-localization of vesicle membrane and cargo, quantitative pharmacokinetics, organ biodistribution, and pathway-inhibition experiments.
Future studies should also include appropriate controls, such as vesicle-free plant extracts, disrupted vesicles, purified phytochemicals, cargo-depleted vesicles, and synthetic or mammalian extracellular vesicle comparators. These controls are essential for distinguishing intrinsic PELN bioactivity from general plant extract effects, free cargo effects, or nonspecific nanoparticle effects.25,32,33,47,59
Disease-specific validation should be strengthened. AD studies should include chronic transgenic models, cognitive outcomes, quantitative Aβ/Tau pathology, and glial activation endpoints.3,6,7,115,122,123,126 PD studies should include α-synucleinopathy models, dopaminergic neuron survival, striatal dopamine function, mitochondrial endpoints, and motor/non-motor behavioral outcomes.8,9,28,117,124,146 HD and ALS studies require direct evaluation in genetically relevant models before therapeutic claims can be made.125,147,150,152 Stroke and nerve injury studies should include neurovascular, electrophysiological, regenerative, and functional recovery endpoints rather than relying solely on anti-inflammatory or antioxidant markers.120,121,138,142–144
Finally, neurological applications that require repeated or long-term administration require rigorous evaluation of pharmacokinetics, biodistribution, clearance, immune responses, off-target accumulation, microbiota perturbation, allergenicity, and long-term safety.20,32,47,61,64,87,105,110 These challenges do not diminish the therapeutic promise of PELNs, but they highlight the need to move beyond descriptive bioactivity studies toward mechanism-informed, disease-specific, and translationally standardized research. Future studies should clearly distinguish direct evidence from native PELNs, engineered PELN-based delivery systems, supportive evidence from plant-derived bioactives, and mechanistic extrapolations from non-PELN models.27,100,102,148,149 Such evidence stratification will be essential for defining realistic therapeutic claims and guiding the rational development of PELNs for neurodegenerative and neural repair applications.
Engineering Strategies for CNS-Targeted PELNs
As summarized in Figure 5, the clinical translation of PELNs for neurodegenerative therapy requires a rational engineering framework that integrates cargo loading, surface functionalization, stability preservation, quality control, and safety assessment. Native PELNs possess several properties that make them attractive as biological nanocarriers, including lipid-bilayer structures, nanoscale dimensions, plant-derived bioactive cargos, relatively low immunogenicity, and potential compatibility with oral or mucosal administration.20,22,27,32,34,148,149,157,158 Their vesicular membranes may protect small molecules, nucleic acids, proteins, and other bioactive cargos from premature degradation. However, these intrinsic features do not automatically ensure efficient brain delivery, disease-specific targeting, or reproducible therapeutic performance. For neurodegenerative applications, engineering strategies should therefore be designed to improve cargo loading, protect cargo integrity, enhance CNS exposure when appropriate, and minimize off-target effects without disrupting the vesicular structure or intrinsic bioactivity of PELNs.
 |
Figure 5 Clinical potential and translational barriers of plant-derived exosome-like nanoparticles (PELNs) for neurodegenerative therapy. (A) PELNs are derived from natural and scalable plant sources, including edible plants, medicinal plants, and plant-based production systems, with abundant plant materials, cost-effective sourcing, and scale-up potential. (B) PELNs have often been reported to show favorable biocompatibility and relatively low immunogenicity; however, their safety profile may vary with plant source, dose, administration route, and preparation quality. (C) The lipid membrane of PELNs may protect vesicle-associated bioactive cargos from enzymatic degradation and help preserve stability and bioactivity. The rightward arrow (→) indicates a directional process toward cargo degradation or inactivation, whereas the cross mark (X) indicates prevention or reduction of this process by membrane protection. (D) PELNs may support versatile cargo capacity, including small molecules, nucleic acids, proteins or peptides, and engineered payloads. (E) PELN delivery and biological interaction are route-dependent. Oral administration may mainly involve the gut–brain axis, intranasal delivery may involve potential nose-to-brain pathways, and intravenous delivery may result in systemic exposure; blood–brain barrier (BBB) interaction should be interpreted as evidence-dependent. (F) Representative engineering and cargo-loading strategies include surface ligand functionalization, PEGylation, electroporation, and ultrasound-assisted loading, which may improve barrier interaction or target-site exposure when supported by appropriate pharmacokinetic and biodistribution evidence. (G) In Alzheimer’s disease (AD)-related studies, plant glycosphingolipids (GSLs) and glucosylceramide (GlcCer)-derived metabolites have been associated with GSL–Aβ binding, microglia-mediated clearance, increased neuronal exosome release, reduced Aβ burden in AD mouse models, and emerging early clinical safety and efficacy signals. In panel G, the rightward arrow (→) indicates a proposed or reported directional association between sequential biological processes, the upward arrow (↑) indicates an increase or promotion, and the downward arrow (↓) indicates a reduction. These arrows summarize reported or conceptual directional effects and should not be interpreted as universal quantitative changes across all PELN-related preparations or disease models. (H) Key translational barriers and potential solutions include scalable gentle purification, such as TFF-based workflows, standardized operating procedures (SOPs) and quality control (QC), batch-to-batch consistency, long-term safety and toxicology evaluation, and biomarker-driven multicenter trials. Created with BioRender.com. |
Importantly, engineered PELNs should not be considered a single homogeneous therapeutic category. Depending on the source plant, isolation method, loading technique, surface modification strategy, and administration route, engineered PELNs may behave as oral gut-active vesicles, systemic nanocarriers, intranasal nose-to-brain delivery systems, or biomaterial-integrated regenerative platforms.35,43,82,107,112,117,138,148 Therefore, CNS-targeted PELN engineering should be evaluated in a route-specific and indication-specific manner rather than assuming universal BBB-crossing capacity.
Cargo Loading Strategies: Incubation, Electroporation, Sonication, Extrusion, and Microfluidics
Cargo loading is a central step in transforming native PELNs into programmable therapeutic nanoplatforms. Current loading strategies can be broadly divided into passive loading and active loading approaches. Passive incubation is technically simple and is particularly suitable for hydrophobic small molecules that can partition into or associate with vesicle membranes. This strategy has been used in plant vesicle-based delivery studies involving doxorubicin-loaded celery-derived nanovesicles, avocado-derived vesicles, sesame leaf-derived vesicles for luteolin delivery, and turmeric-derived exosome-like nanoparticles.57,89,159,160 Passive loading has the advantage of relatively mild processing conditions, but loading efficiency depends strongly on cargo hydrophobicity, vesicle lipid composition, incubation conditions, and post-loading purification.
Active loading methods, including electroporation, sonication, extrusion, freeze–thaw cycling, and membrane permeabilization, may improve encapsulation of hydrophilic drugs, nucleic acids, proteins, or other biomacromolecules.43,56,67,113,148 Electroporation can transiently disrupt membrane integrity to facilitate nucleic acid loading, but it may also induce vesicle aggregation, cargo precipitation, or RNA degradation if conditions are not carefully optimized. Sonication and extrusion can enhance cargo entry by mechanically remodeling vesicle membranes, but excessive energy input may alter particle size distribution, surface charge, protein conformation, lipid organization, and biological activity.67,148 These concerns are especially important for PELNs because vesicle-associated lipids, proteins, small RNAs, and phytochemicals may all contribute to biological effects.26,37,63,149
Microfluidic technologies provide a promising next-generation platform for controlled vesicle engineering. Compared with bulk loading methods, microfluidic systems may improve mixing precision, reduce sample loss, enable continuous processing, and support scalable, low-damage manufacturing.82 For CNS-related applications, microfluidics may be particularly valuable for loading fragile nucleic acids, neuroprotective proteins, or plant-derived bioactive compounds while maintaining vesicle size uniformity and cargo retention.82,107,148 Although most microfluidic loading data still come from mammalian extracellular vesicles or synthetic nanocarriers, such platforms may provide a useful technical basis for future PELN engineering. In addition, a biocompatible tea polyphenol nanoplatform was recently reported to enable efficient cytosolic delivery of protein therapeutics, supporting the broader feasibility of plant-derived or plant-inspired nanomaterials for biomacromolecule delivery.161 This evidence should be interpreted as supportive nanomedicine evidence rather than direct PELN evidence.
Several disease-relevant examples support the feasibility of engineered plant vesicle platforms. Grapefruit-derived nanovectors have been used to deliver therapeutic miR-17 through the intranasal route in a brain tumor model, demonstrating the feasibility of non-invasive vesicle-assisted CNS delivery.112 Engineered Pueraria lobata-derived exosome-like nanoplatforms have been reported to deliver biomacromolecular cargos to dopaminergic neuronal cells and ameliorate mitochondrial dysfunction in Parkinson’s disease-related models.117 In regenerative medicine, Lycium barbarum L.-derived exosome-like nanoparticles loaded with isoliquiritigenin and incorporated into 3D-printed biomimetic scaffolds promoted spinal cord injury repair.138 These studies collectively suggest that PELNs can be engineered for diverse cargo types and neurological contexts, but they also highlight the need to match the loading method with the cargo, disease target, and administration route.
Surface Functionalization for BBB Targeting and Cell-Specific Delivery
Surface functionalization is a major strategy for improving the tissue selectivity and cellular specificity of engineered PELNs. For CNS applications, potential modification approaches include ligand conjugation, peptide decoration, receptor-targeting moieties, membrane hybridization, lipid remodeling, polyethylene glycol modification, and polymer-assisted stabilization.18,103,106,107,118,148 These strategies are intended to enhance BBB interaction, prolong circulation, improve mucosal retention, promote receptor-mediated uptake, or redirect vesicles toward disease-relevant cells such as neurons, microglia, astrocytes, brain endothelial cells, or infiltrating immune cells.
However, BBB targeting should be interpreted cautiously. The BBB is a highly selective neurovascular interface that restricts the entry of most macromolecules and many nanoparticle systems.13 Evidence from mammalian extracellular vesicle studies suggests that vesicle transport across the BBB can be influenced by vesicle source, surface proteins, inflammation, disease state, and detection method.105 For PELNs, direct evidence of intact-vesicle transport across the BBB remains heterogeneous. Fluorescent signals in brain tissue may reflect intact vesicles, free dye, degraded vesicle fragments, released cargo, uptake by vascular or immune cells, or disease-associated BBB disruption. Therefore, surface-engineered PELNs intended for BBB targeting should be evaluated using quantitative biodistribution, dual-label tracking of vesicle membrane and cargo, endothelial transcytosis assays, organ-level pharmacokinetics, and cell-type-specific co-localization.
Intranasal delivery represents a particularly relevant route for CNS-oriented PELNs because it may partially bypass the BBB through olfactory and trigeminal pathways.112 Nevertheless, intranasal delivery does not guarantee broad brain distribution or neuronal targeting. Particle size, mucosal retention, enzymatic degradation, nasal clearance, formulation viscosity, and disease state can all influence delivery efficiency. For orally administered PELNs, surface features may be more important for intestinal uptake, gut epithelial interaction, macrophage uptake, microbiota modulation, and systemic immune-metabolic regulation than for direct BBB transport.35,61,64,94,110 Therefore, the design of surface-functionalized PELNs should be aligned with the intended biological interface, including the intestinal barrier, nasal mucosa, vascular endothelium, immune cells, or CNS parenchymal cells.
Cell-specific delivery also remains an important challenge. Microglia-targeted PELNs could be useful for neuroinflammatory disorders, whereas neuron-targeted vesicles may be more relevant for proteinopathy, mitochondrial dysfunction, or synaptic failure.18,115–117 Surface engineering may theoretically enhance such selectivity, but rigorous validation requires comparison across neurons, astrocytes, microglia, endothelial cells, macrophages, and peripheral immune cells. Quantitative uptake assays, receptor-blocking experiments, live-cell imaging, and single-cell omics could help define whether engineered PELNs truly achieve cell-specific delivery or merely alter nonspecific nanoparticle uptake.
Stability, Cargo Retention, and Functional Integrity After Engineering
Engineering can improve the therapeutic versatility of PELNs, but it can also compromise vesicle integrity. Therefore, engineered PELNs should be assessed not only for loading efficiency but also for structural stability, cargo retention, biological activity, and reproducibility after modification. Important quality attributes include particle size distribution, polydispersity, zeta potential, morphology, membrane integrity, cargo encapsulation efficiency, loading capacity, residual free cargo, sterility, endotoxin level, and functional potency.22,23,25,36,59,87,148
Cargo retention is especially important for CNS applications because premature cargo leakage during storage, circulation, or mucosal transit may reduce brain exposure and increase off-target toxicity. Hydrophobic drugs may remain associated with vesicle membranes, whereas hydrophilic small molecules, nucleic acids, and proteins may be more vulnerable to leakage or degradation.35,56,87,148 For orally delivered PELNs, cargo stability should be tested under simulated gastric and intestinal conditions. For systemic or intranasal administration, stability should be assessed in serum, nasal fluid, cerebrospinal fluid-relevant conditions, and inflammatory microenvironments.
Storage stability is another key translational issue. Freeze–thaw cycles, lyophilization, cryoprotectants, buffer composition, temperature, and storage duration can affect particle aggregation, membrane rupture, cargo release, and potency.73–76,87,109 Although TFF, SEC, and combined purification workflows may improve recovery and purity, they may also alter vesicle subpopulations or remove loosely associated bioactive components.71–76,79,80,109 Therefore, post-engineering and post-storage comparisons should be performed against freshly isolated native PELNs, unloaded engineered PELNs, free cargo, disrupted vesicles, and vesicle-depleted plant extracts. These controls are necessary to determine whether therapeutic activity arises from intact vesicles, vesicle-associated cargo, co-isolated phytochemicals, or nonspecific nanoparticle effects.
Functional integrity should also be measured using disease-relevant bioassays. For neuroinflammatory applications, assays may include microglial iNOS, NO, TNF-α, IL-1β, NLRP3 inflammasome activation, and HO-1 expression.115,116,119 For redox and mitochondrial mechanisms, endpoints may include ROS production, mitochondrial membrane potential, Nrf2/HO-1 activation, Bax/Bcl-2 balance, and PINK1/Parkin-related mitophagy.97,117,122 For proteinopathy-related applications, Aβ, Tau, or α-synuclein aggregation and toxicity assays may be used.129–135 Such potency assays should become part of the functional characterization package for engineered PELNs and should be aligned with broader extracellular vesicle reporting standards.36
Engineered PELNs versus Mammalian EVs and Synthetic Nanocarriers
PELNs occupy an intermediate position between natural-product therapeutics and engineered nanomedicine. Compared with synthetic nanocarriers such as liposomes, polymeric nanoparticles, inorganic nanoparticles, and lipid nanoparticles, PELNs may offer better biological compatibility, intrinsic bioactivity, edible or sustainable sourcing, and natural cargo complexity.20,32,34,56,149,157,158 Compared with mammalian extracellular vesicles, PELNs may offer advantages in scalability, cost, food-based sourcing, and reduced risk of mammalian pathogen contamination.20–24 However, these advantages should not be overstated because PELNs also face major challenges in source heterogeneity, marker uncertainty, batch consistency, cargo variability, and regulatory classification.
Mammalian extracellular vesicles have advantages in homologous cell signaling, well-developed surface marker systems, and potentially more predictable interactions with mammalian recipient cells.15–18,77,88,151 However, mammalian extracellular vesicles often face limitations related to donor variability, cell culture cost, low yield, tumorigenic or immunological concerns depending on source, and complex manufacturing requirements.20,83,88 By contrast, fully synthetic nanocarriers, including liposomes, polymeric nanoparticles, lipid nanoparticles, micelles, dendrimers, and inorganic nanoparticles, offer high controllability, reproducible composition, defined chemistry, and established manufacturing processes, but they generally lack the intrinsic bioactive cargo and natural membrane identity of biogenic vesicles and may induce material-dependent toxicity, complement activation, or rapid clearance.77,113,118
Engineered PELNs may therefore be most valuable when engineering strategies enhance cargo loading, stability, delivery efficiency, or tissue targeting without substantially compromising vesicle integrity and source-derived bioactive features. This dual bioactive-delivery concept aligns with broader developments in natural-product nanomedicine, in which nanocarriers are designed to improve the stability, solubility, bioavailability, and tissue distribution of plant-derived bioactives.14,22,23,56,67,149,161 However, the added value of engineered PELNs should be demonstrated against both unmodified PELNs and established delivery platforms, including mammalian extracellular vesicles, liposomes, lipid nanoparticles, and polymeric nanoparticles. At present, standardized head-to-head comparisons remain limited. Future studies should compare cargo-loading capacity, encapsulation efficiency, cargo retention and release kinetics, vesicle integrity after engineering, cellular uptake pathways, performance in validated BBB models, biodistribution, immune activation, toxicity, storage stability, scalability, manufacturing cost, and disease-specific potency under harmonized experimental conditions.
Potential Risks Introduced by Engineering Modifications
Engineering modifications may introduce risks that are absent or less pronounced in native PELNs. Physical loading methods may disrupt membrane integrity, change vesicle size distribution, expose internal plant components, induce aggregation, or alter biodistribution. Chemical conjugation may introduce residual reagents, alter surface charge, mask natural uptake ligands, or change protein corona formation.67,148 Surface ligands designed to improve targeting may also redirect vesicles toward unintended tissues or immune cell populations. Polyethylene glycol modification may improve circulation or formulation stability in some nanomedicine contexts, but it may also alter cellular uptake, biodistribution, and immune recognition. Therefore, polyethylene glycol-modified or ligand-functionalized PELNs should be evaluated as modified biological nanoproducts rather than simple extensions of native vesicles.
Another concern is cargo-related toxicity. Nucleic acid-loaded PELNs may trigger innate immune sensing, off-target gene regulation, or unintended microbiota modulation.63,125,148 Small-molecule-loaded PELNs may alter pharmacokinetics and tissue exposure compared with free drug, potentially increasing accumulation in the liver, spleen, kidney, or inflamed tissues.43,44,105,160 For CNS applications, repeated dosing raises additional concerns regarding chronic neuroimmune activation, glial reactivity, endothelial perturbation, and long-term changes in gut microbiota or systemic immune metabolism.11,22,60,64,110
Plant source-related risks should also be considered. PELNs may contain plant allergens, pesticide residues, environmental contaminants, microbial products, or source-specific bioactive molecules. Plant cultivation conditions, harvest time, tissue type, ripeness, processing, and extraction method can all affect vesicle composition.23,26,37,47,59 Therefore, engineered PELNs should undergo safety evaluation not only as nanocarriers but also as plant-derived biological products. Risk assessment should include acute toxicity, repeat-dose toxicity, immunogenicity, cytokine release, complement activation, hemocompatibility, genotoxicity when relevant, microbiota perturbation, reproductive toxicity when appropriate, and long-term organ biodistribution.
Translational Challenges: Pharmacokinetics, Safety, CMC, and Regulatory Pathways
Although PELNs have shown considerable promise in preclinical models, their clinical translation remains constrained by unresolved challenges related to pharmacokinetics, long-term safety, CMC, and regulatory pathways. These challenges are particularly important for neurodegenerative diseases because therapeutic intervention may require repeated dosing, chronic administration, long-term safety monitoring, and reliable CNS- or gut–brain axis-related endpoints.22,32,33,47,59,100,102 The translational pathway for PELNs should therefore move beyond descriptive bioactivity and establish quantitative standards for dosing, biodistribution, clearance, release testing, and mechanism-based potency.
Dosing Normalization and Pharmacokinetic Uncertainties
A major barrier in PELN research is the lack of standardized dosing metrics. Current studies frequently normalize PELN dose by total protein content, particle number, plant weight, extract volume, RNA amount, or loaded drug concentration.21–23,34,59 These metrics are not interchangeable. Protein content may be influenced by co-isolated soluble proteins; particle number may be affected by aggregation or non-vesicular particles; plant weight does not account for extraction efficiency; and loaded drug concentration does not define vesicle-associated versus free drug fractions. As a result, cross-study comparison remains difficult.
For translational development, dosing should ideally report multiple parameters, including particle number, particle-to-protein ratio, lipid content, RNA content, cargo amount, encapsulation efficiency, residual free cargo, and biological potency.22,23,25,36,86 Dose–response relationships should be established for both efficacy and toxicity. In CNS applications, this is especially important because a biologically active oral dose may act through the gut–brain axis, whereas an intranasal or intravenous dose may aim for more direct CNS exposure.11,61,64,105,112
Pharmacokinetic interpretation is also complicated by the structural complexity of PELNs. After administration, intact vesicles may remain stable, undergo partial degradation, release cargo, fuse with cell membranes, be taken up by phagocytes, or be transformed by digestive and metabolic processes.11,12,35,60,61 Therefore, the pharmacokinetics of PELNs should not be reduced to a single plasma half-life value. Future studies should distinguish intact-vesicle kinetics, cargo kinetics, membrane lipid kinetics, loaded drug kinetics, and downstream pharmacodynamic effects. Dual-label or multi-label tracking strategies, quantitative mass spectrometry, RNA tracking, lipidomics, and functional bioassays may help clarify these relationships.
Biodistribution, Clearance, and Off-Target Accumulation
PELNs may show different biodistribution patterns depending on administration route. Oral PELNs are likely to interact first with intestinal mucus, epithelial cells, macrophages, gut microbiota, and mucosal immune structures.35,61,64,94,110 Intranasal PELNs may be influenced by mucociliary clearance, olfactory and trigeminal pathways, nasal-associated lymphoid tissue, and local immune responses.112 Intravenous PELNs may undergo protein corona formation, opsonization, uptake by mononuclear phagocyte system cells, and accumulation in liver, spleen, lung, or kidney.43,44,105
Therefore, biodistribution studies should avoid relying solely on qualitative fluorescence imaging. Fluorescent dye artifacts, dye transfer, vesicle degradation, and signal persistence can lead to overinterpretation of tissue accumulation.105 Robust biodistribution analysis should combine quantitative organ imaging, ex vivo tissue measurement, co-localization of membrane and cargo signals, pharmacokinetic modeling, and cell-type-specific analysis. When brain signals are observed, studies should determine whether the signal is associated with endothelial cells, perivascular macrophages, microglia, neurons, astrocytes, or infiltrating immune cells.
Advanced imaging and tracer technologies may help improve the evaluation of brain-targeted nanomedicine, although their use in PELN research remains underdeveloped. For example, multivoid magnetic nanoparticles have been explored as high-performance magnetic particle imaging tracers for precise glioma detection, illustrating how sensitive imaging platforms may support localization and monitoring of CNS-directed nanosystems.162 This type of imaging evidence should be considered methodological support rather than direct evidence for PELN biodistribution. Future PELN studies may benefit from integrating fluorescence imaging, magnetic resonance imaging, magnetic particle imaging, positron emission tomography, mass spectrometry imaging, and single-particle analytical approaches.
Clearance pathways also require systematic investigation. Vesicles and vesicle-associated cargos may be cleared through hepatic metabolism, biliary excretion, renal filtration of released small molecules, splenic macrophage uptake, or gastrointestinal elimination.43,44,60,105 For oral PELNs, microbial and hepatic biotransformation may generate metabolites with different absorption, tissue distribution, and biological effects compared with the original vesicle cargo.12 This possibility is particularly relevant for plant-derived bioactives with low intrinsic bioavailability but significant systemic or neurological effects after metabolic transformation.
Immunogenicity, Chronic Toxicity, and Long-Term CNS Safety
PELNs are often described as biocompatible and low-immunogenic, but this should be treated as a hypothesis requiring context-specific validation rather than a universal property.20,22,32,33,47 Acute tolerability does not necessarily predict chronic safety, especially for neurodegenerative diseases that may require repeated administration over months or years. Potential safety concerns include immune activation, cytokine release, complement activation, allergic responses, microbiota perturbation, off-target organ accumulation, and unintended gene regulation by vesicle-associated small RNAs.22,60,63,64,110
Some plant-derived vesicles may have immunomodulatory or immunostimulatory effects depending on source and cargo.38,91,111,115 These effects may be therapeutically useful in some settings but undesirable in chronic CNS disorders if they promote systemic inflammation or neuroimmune activation. Conversely, anti-inflammatory PELNs may suppress protective immune functions or alter host–microbiota balance after long-term use. Therefore, both immunostimulatory and immunosuppressive risks should be evaluated.
Long-term CNS safety requires particular attention. Even if PELNs do not directly enter the brain in large quantities, they may influence CNS function through peripheral immune regulation, gut microbiota remodeling, vagal signaling, metabolic products, or endothelial effects.11,60,61,64,110 Chronic studies should therefore assess behavior, cognition, motor function, neuroinflammation, glial activation, synaptic markers, BBB integrity, microbiota composition, systemic cytokines, and histopathology in major organs. For engineered PELNs, additional safety testing should evaluate the loaded cargo, surface ligand, residual reagents, and altered biodistribution.
GMP-Compatible PELN Manufacturing: Scalable Purification, Storage Stability, and Release Control
Clinical translation of PELNs requires scalable, reproducible, and GMP-compatible manufacturing workflows rather than simple particle enrichment. As discussed in Isolation, Purification, Characterization, and Quality Control of PELNs, current PELN studies have used diverse isolation and purification strategies, including UC, DGUC, SEC, immunoaffinity capture, TFF, ion-exchange or other chromatography-based purification methods, and emerging microfluidic technologies. However, these approaches differ in yield, recovery, purity, vesicle integrity, cargo preservation, contaminant risk, scalability, and manufacturing compatibility. Therefore, translational process development should focus not only on particle recovery, but also on product identity, purity, potency, safety, and batch-to-batch reproducibility.23,28–30,60,84,85,101
TFF is particularly relevant for processing large volumes and concentrating vesicle-containing fluids, while SEC may help separate vesicles from soluble proteins and small molecules.74–76,79,109 Anion-exchange chromatography has also been explored for plant-derived nanovesicle purification and may provide a route toward more defined manufacturing processes.80 However, each purification method may enrich different vesicle subpopulations and affect cargo composition, potency, and yield. Therefore, process development should include comparability studies across purification methods.
Raw material control is another major issue. Plant species, cultivar, tissue type, growth conditions, harvest season, geographic origin, storage, pesticide exposure, microbial contamination, and extraction conditions may influence PELN composition.23,26,37,47,59 A GMP-compatible workflow should define plant sourcing, authentication, cultivation standards, harvest conditions, extraction buffer, filtration parameters, purification steps, storage conditions, and release criteria. For edible or nutraceutical-grade products, food safety standards may be relevant; for engineered CNS-targeted nanodrugs, pharmaceutical-grade standards will be required.
Storage stability must be addressed early in development. PELN formulations should be evaluated under refrigerated, frozen, lyophilized, and reconstituted conditions.73–76,87,109 Stability-indicating assays should include particle size, morphology, aggregation, membrane integrity, cargo retention, sterility, endotoxin, and potency. Cryoprotectants or lyoprotectants may improve stability but may also affect biological activity or regulatory classification. Therefore, storage conditions should be treated as part of the product design rather than as a late-stage formulation issue. In this context, MISEV2023 provides useful general guidance for extracellular vesicle reporting, characterization, and functional assessment, although PELNs will still require plant-specific analytical standards.36
CMC Requirements, Release Testing, and Regulatory Classification
CMC requirements will be central to the clinical translation of PELNs. However, regulatory classification remains ambiguous because PELNs may be interpreted as natural products, biological products, nanomedicines, drug delivery systems, dietary components, or combination products depending on their source, processing, cargo, claims, and route of administration.20,22,32,47,59,148,163 A native edible PELN formulation intended for gut–brain axis modulation may face a different regulatory pathway from an engineered, ligand-modified, drug-loaded PELN intended for CNS disease treatment.
A minimal release testing panel for PELNs should include identity, purity, potency, safety, and stability parameters. Identity testing may include particle size distribution, morphology, lipid profile, protein profile, RNA profile, and source-specific molecular fingerprints.23,25,36,37,86 Purity testing should evaluate soluble protein contamination, plant debris, aggregates, non-vesicular particles, residual extraction reagents, endotoxin, microbial contamination, pesticide residues, and heavy metals. Potency testing should be mechanism-specific, such as anti-inflammatory activity, antioxidant activity, cargo delivery efficiency, neuronal survival, microglial modulation, or disease-relevant pathway activation.97,115–117,122
Because universal PELN markers remain unavailable, marker specificity is a major limitation.25,37,45,90 Unlike mammalian extracellular vesicles, PELNs do not yet have a standardized marker panel equivalent to classical mammalian EV markers. Therefore, source-dependent fingerprinting may be more realistic than universal marker-based identification. Advanced analytical methods such as proteomics, lipidomics, metabolomics, small RNA sequencing, nanoparticle tracking analysis, electron microscopy, chromatography, mass spectrometry, and emerging biosensing technologies may collectively support product characterization.36,82,86
Regulatory development should also require comparability criteria after process changes. Changes in plant source, extraction scale, purification method, storage condition, loading strategy, or surface modification may alter vesicle composition and potency. Therefore, batch-to-batch consistency should be evaluated using predefined critical quality attributes, including particle concentration, size distribution, particle-to-protein ratio, lipid signature, cargo profile, sterility, stability, and functional potency.22,23,36,47,59 Establishing such criteria will be essential for moving PELNs from exploratory preclinical studies toward standardized clinical products.
Future Perspectives: A Tiered Clinical Translation Framework
The future development of PELNs for neurodegenerative diseases should be guided by a tiered clinical translation framework rather than a single therapeutic pathway. This is because PELNs may be positioned as dietary bioactive vesicles, standardized natural-product formulations, engineered nanodrug delivery systems, or regenerative biomaterial components, each of which entails distinct requirements for product characterization, evidence generation, manufacturing control, regulatory evaluation, and clinical indication selection.22,23 A tiered framework may therefore help align vesicle design, quality standards, preclinical evidence, regulatory expectations, and clinical development strategies.
Nutraceutical and Dietary PELN-Related Formulations
The first translational tier involves nutraceutical or dietary PELN-related formulations. This tier is most relevant for edible plant-derived vesicles, plant lipid fractions, and oral formulations intended to modulate gut barrier function, gut microbiota, systemic metabolism, or low-grade inflammation.61,64,94,110,127,128,164 Such products may be particularly suitable for early prevention, health maintenance, or adjunctive management rather than disease-modifying treatment claims.
Dietary plant glycosphingolipids and glucosylceramides provide an example of how plant-derived lipid species may influence host extracellular vesicle biology and Aβ-related pathology through a potential diet–host EV–brain axis.127,128 Similarly, plant-derived exosomal microRNAs and microRNA-enriched plant exosomes have been reported to reshape gut microbiota, regulate systemic immunity, and influence metabolic homeostasis.64,110 Citrus exosome-like nanoparticles that reverse high-fat diet-induced microbiota dysbiosis and alter short-chain fatty acid production further support the potential of oral PELN-related systems in gut-mediated regulation.46
However, nutraceutical development should not rely on vague claims of “brain targeting.” For oral PELNs, the most plausible primary interface may be the gut epithelium, microbiota, mucosal immune cells, and systemic metabolism rather than direct brain entry.11,12,60,61,64 Therefore, future dietary PELN studies should prioritize microbiome profiling, metabolomics, intestinal permeability assays, inflammatory biomarkers, systemic pharmacodynamic markers, and long-term tolerability. If cognitive or neurological claims are made, they should be supported by disease-relevant behavioral endpoints and biomarkers.
Standardized Natural PELNs Formulations
The second translational tier involves standardized natural PELN formulations. These products would retain the intrinsic bioactivity of native PELNs while being manufactured under defined sourcing, isolation, characterization, and storage conditions. Compared with nutraceutical formulations, standardized natural PELNs would require stronger evidence of composition, mechanism, dose–response relationship, and batch consistency.22,23,25,47,59
This tier may be appropriate for plant sources with reproducible anti-inflammatory, antioxidant, gut-regulatory, or neuroprotective effects. Examples include Allium tuberosum-derived ELNs with anti-inflammatory effects in microglia-like cells, ginger-derived ELNs that inhibit NLRP3 inflammasome activation, Lycium ruthenicum Murray-derived ELNs that protect neuronal cells against Aβ-induced oxidative injury, and Solanum lycopersicum-derived ELNs that regulate Keap1/Nrf2 signaling in vascular injury models.97,115,116,122,123 These examples support the feasibility of source-specific natural PELN formulations, but disease-specific validation remains necessary.
Standardized natural PELNs should be developed using defined critical quality attributes. These may include botanical authentication, vesicle yield, particle size distribution, morphology, particle-to-protein ratio, RNA profile, lipid profile, key bioactive components, storage stability, and mechanism-based potency.22,23,25,36,86 For neurodegenerative applications, potency assays should be selected based on the intended mechanism, such as microglial inflammation, Nrf2 activation, mitochondrial protection, Aβ toxicity reduction, or gut barrier regulation. Such standardization would help distinguish PELN-specific bioactivity from crude plant extract effects.
Engineered CNS-Targeted PELNs Nanodrugs
The third translational tier involves engineered CNS-targeted PELN nanodrugs. These products would be designed to deliver defined therapeutic cargos, such as small molecules, nucleic acids, proteins, peptides, antioxidants, anti-inflammatory agents, or anti-proteinopathy compounds.14,18,56,107,112,117,138,142,146,148,161 Compared with nutraceutical or natural PELN formulations, engineered PELN nanodrugs will require the highest level of quality control, pharmacokinetic characterization, safety evaluation, and regulatory scrutiny.
Potential indications include AD-related Aβ or Tau pathology, PD-related mitochondrial dysfunction and α-synuclein toxicity, RNA-based strategies for genetic neurodegenerative diseases, and regenerative repair after neural injury.107,117,125,129,138,146,148 For example, engineered Pueraria lobata-derived nanoplatforms provide a model for biomacromolecule delivery in PD-related mitochondrial dysfunction, while Lycium barbarum L.-derived ELNs incorporated into 3D-printed scaffolds illustrate the potential of engineered PELN-biomaterial systems for neural repair.117,138 In addition, the integration of nanomedicine with traditional medicinal bioactives provides a broader conceptual framework for improving the delivery, stability, and tissue distribution of natural products, although such evidence should be distinguished from direct PELN-specific data.138
For CNS-targeted PELN nanodrugs, the key development questions are whether the engineered vesicles reach the intended biological site, retain their cargo, engage the expected target pathway, and produce meaningful functional benefit. These products should be evaluated using quantitative pharmacokinetics, organ biodistribution, brain-region analysis, cell-type-specific uptake, behavioral outcomes, and long-term safety endpoints. In addition, engineered products should be compared with free drug, unloaded vesicles, disrupted vesicles, mammalian extracellular vesicles, and synthetic nanocarriers to define their added value.
Multidisciplinary Integration
The successful development of PELNs for neurodegenerative diseases will require integration across botany, food science, nanotechnology, neuroscience, pharmacology, bioengineering, analytical chemistry, and regulatory science. Plant science is needed to control source variability, cultivation conditions, harvest quality, and bioactive composition.37,55,80 Nanotechnology and pharmaceutical sciences are needed to optimize loading, targeting, stability, and scalable manufacturing.82,107,148 Neuroscience is required to define disease-relevant mechanisms, models, biomarkers, and functional outcomes.15,18,100,102
Advanced disease models may improve translational prediction. Three-dimensional neural stem cell microenvironment models, organoids, BBB-on-chip platforms, co-culture systems, and biomaterial-based neural repair models can provide more disease-relevant platforms for evaluating PELN effects on neurons, astrocytes, microglia, endothelial cells, and neural stem cells.84,88,136–138,144 These systems may help bridge the gap between simple cellular assays and complex in vivo disease models. Natural product-based neural repair comparators, such as hydroxysafflor yellow A in traumatic brain injury, may also guide the selection of regenerative endpoints for future PELN studies.143
Future studies should also integrate multi-omics and imaging technologies. Proteomics, lipidomics, metabolomics, small RNA sequencing, single-cell transcriptomics, spatial omics, and in vivo imaging can help define PELN composition, tissue distribution, cell-type-specific responses, and mechanism of action.36,82,86,108 Emerging imaging platforms, including magnetic particle imaging tracers for precise glioma detection, may provide useful technical inspiration for future tracking and localization of CNS-directed nanosystems, although their application to PELNs remains to be established.162 Such approaches are essential for distinguishing direct CNS delivery from gut-mediated systemic effects, identifying predictive biomarkers, and building mechanism-informed clinical trial designs.
Ultimately, PELN translation should proceed through stepwise evidence generation: first establishing vesicle identity and source-dependent composition; then validating mechanism-specific potency; then defining biodistribution, safety, and dose–response relationships; and finally conducting indication-specific clinical studies. This tiered strategy can prevent overstatement while preserving the therapeutic promise of PELNs as sustainable, bioactive, and engineerable nanoplatforms.
Conclusion
PELNs represent an emerging class of bioactive nanocarriers with potential relevance for AD and related neurodegenerative disorders. In contrast to conventional plant extracts, PELNs are nanoscale, membrane-enclosed vesicle-like fractions that carry source-dependent lipids, proteins, nucleic acids, and phytochemicals, thereby combining intrinsic bioactivity with delivery-related properties.24,25,32–34,37 Accumulating evidence suggests that PELNs may modulate several pathological processes shared by neurodegenerative diseases, including neuroinflammation, oxidative stress, mitochondrial dysfunction, protein aggregation, impaired intercellular communication, and gut–brain axis dysregulation.11,22,27,100,102,104
Nevertheless, the therapeutic translation of PELNs remains at an early stage. Although preclinical studies have reported neuroprotective effects of selected plant-derived vesicles in models related to AD, Parkinson’s disease, neural injury, and CNS inflammation, current evidence remains heterogeneous across plant sources, isolation methods, administration routes, disease models, and outcome measures.100,102,104,115,117,122,123,126,128 In particular, BBB transport and CNS targeting should be interpreted cautiously, because vesicle labeling strategy, disease-associated BBB disruption, biodistribution analysis, and the distinction between intact-vesicle delivery and cargo-derived signal can strongly affect experimental conclusions.13,105–107
Future development of PELN-based neurotherapeutics will require a shift from descriptive efficacy studies toward mechanism-driven and translation-oriented investigation. Standardized nomenclature, rigorous vesicle characterization, reproducible isolation and purification, source-dependent quality control, dosing normalization, pharmacokinetic assessment, and long-term safety evaluation are essential for defining critical quality attributes and improving cross-study comparability.21,23,33,36,66,71–76 Scalable manufacturing approaches, including TFF, SEC, chromatography-based purification, and microfluidic technologies, may improve yield, purity, recovery, and process reproducibility, but their suitability for clinical-grade PELN production still requires systematic validation.79–82
Overall, PELNs provide a promising but still developing platform at the intersection of natural-product pharmacology, extracellular vesicle biology, and nanomedicine. Their greatest translational value may lie not in assuming universal BBB-crossing or disease-modifying efficacy, but in rationally matching plant source, vesicle composition, engineering strategy, route of administration, and disease mechanism.14,148,149,157,158 With rigorous standardization, mechanistic validation, and carefully designed preclinical-to-clinical translation pathways, PELNs may become valuable bioactive nanocarriers for multi-target intervention in AD and related neurodegenerative disorders.15,18,20,59,102,104
Abbreviations
3D, three-dimensional; ABC, ATP-binding cassette; AD, Alzheimer’s disease; Aβ, β-amyloid; A-ELNs, Allium tuberosum-derived exosome-like nanoparticles; AGEs, advanced glycation end products; AIM2, absent in melanoma 2; Akt, protein kinase B; ALIX, ALG-2-interacting protein X; ALS, amyotrophic lateral sclerosis; ARE, antioxidant response element; ATPase(s), adenosine triphosphatase(s); Bax, Bcl-2-associated X protein; BBB, blood–brain barrier; Bcl-2, B-cell lymphoma 2; BDNF, brain-derived neurotrophic factor; C9orf72, chromosome 9 open reading frame 72; CAG, cytosine–adenine–guanine; CD9, cluster of differentiation 9; CD63, cluster of differentiation 63; CD81, cluster of differentiation 81; CMC, Chemistry, Manufacturing, and Controls; CNS, central nervous system; CQA/CQAs, critical quality attribute(s); DGUC, density-gradient ultracentrifugation; DGDG, digalactosyldiacylglycerol; DLS, dynamic light scattering; DSS, dextran sulfate sodium; ECG, epicatechin gallate; EGCG, epigallocatechin gallate; ELNs, exosome-like nanoparticles; ERK, extracellular signal-regulated kinase; EVs, extracellular vesicles; GDNF, glial cell-derived neurotrophic factor; GI, gastrointestinal; GMP, good manufacturing practice; GSK-3β, glycogen synthase kinase-3β; GSLs, glycosphingolipids; HD, Huntington’s disease; HO-1, heme oxygenase-1; HSF1, heat shock factor 1; HSPs, heat shock proteins; HSP70, heat shock protein 70; HTT, huntingtin; IL-1β, interleukin-1β; iNOS, inducible nitric oxide synthase; ISL, isoliquiritigenin; Keap1, kelch-like ECH-associated protein 1; LPS, lipopolysaccharide; MAPK, mitogen-activated protein kinase; MGDG, monogalactosyldiacylglycerol; miRNA, microRNA; MISEV, Minimal Information for Studies of Extracellular Vesicles; MPS, mononuclear phagocyte system; mRNA, messenger RNA; MPTP, 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine; mTOR, mammalian target of rapamycin; MVBs, multivesicular bodies; NF-κB, nuclear factor-κB; NGF, nerve growth factor; NLRP3, NLR family pyrin domain containing 3; NO, nitric oxide; NQO1, NAD(P)H:quinone oxidoreductase 1; Nrf2, nuclear factor erythroid 2-related factor 2; NTA, nanoparticle tracking analysis; PA, phosphatidic acid; PD, Parkinson’s disease; PE, phosphatidylethanolamine; PELNs, plant-derived exosome-like nanoparticles; PEN1, penetration 1; PI3K, phosphoinositide 3-kinase; PI3K/Akt, phosphoinositide 3-kinase/protein kinase B; PINK1, PTEN-induced kinase 1; PK, pharmacokinetics; QC, quality control; RNA, ribonucleic acid; ROS, reactive oxygen species; SCI, spinal cord injury; SEC, size-exclusion chromatography; SEM, scanning electron microscopy; siRNA, small interfering RNA; SM, sphingomyelin; SOD1, superoxide dismutase 1; TBI, traumatic brain injury; TDP-43, TAR DNA-binding protein 43; TEM, transmission electron microscopy; TET8, tetraspanin 8; TFF, tangential flow filtration; TNF-α, tumor necrosis factor-α; TRPS, tunable resistive pulse sensing; TSG101, tumor susceptibility gene 101; UC, ultracentrifugation.
Data Sharing Statement
Data sharing is not applicable to this article as no new data were created or analyzed in this study.
Ethical Approval and Informed Consent
Not applicable as this is a review article and does not involve direct research on human or animal subjects.
Acknowledgments
We thank BioRender.com for their assistance in creating the figures.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the Taishan Scholar Project of Shandong Province of China (tsqn202103200); the Traditional Chinese Medicine Science and Technology Project of Shandong Province (Z20241405); and the Shandong Provincial Natural Science Foundation (ZR2025QC1878Z); and the General Program for Scientific and Technological Development of Affiliated Hospital of Shandong Second Medical University (2025FYQ066).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Feigin VL, Vos T, Nichols E, et al. The global burden of neurological disorders: translating evidence into policy. Lancet Neurol. 2020;19(3):255–47. doi:10.1016/S1474-4422(19)30411-9
2. Wang S, Jiang Y, Yang A, Meng F, Zhang J. The expanding burden of neurodegenerative diseases: an unmet medical and social need. Aging Dis. 2024;16(5):2937–2952. doi:10.14336/AD.2024.1071
3. Ramachandran AK, Das S, Joseph A, Shenoy GG, Alex AT, Mudgal J. Neurodegenerative pathways in Alzheimer’s disease: a review. Curr Neuropharmacol. 2021;19(5):679–692. doi:10.2174/1570159X18666200807130637
4. Annicchiarico R, Federici A, Pettenati C, Caltagirone C. Rivastigmine in Alzheimer’s disease: cognitive function and quality of life. Ther Clin Risk Manag. 2007;3(6):1113–1123.
5. Sun MK, Alkon DL. Treating Alzheimer’s disease: focusing on neurodegenerative consequences. J Alzheimers Dis. 2024;101(s1):S263–S274. doi:10.3233/JAD-240479
6. Crestini A, Santilli F, Martellucci S, et al. Prions and neurodegenerative diseases: a focus on Alzheimer’s disease. J Alzheimers Dis. 2022;85(2):503–518. doi:10.3233/JAD-215171
7. Li YY, Guo DD, Duan RN, Li Y. Interactions between beta-amyloid and pericytes in Alzheimer’s disease. Front Biosci. 2024;29(4):136. doi:10.31083/j.fbl2904136
8. Monzio Compagnoni G, Di Fonzo A, Corti S, Comi GP, Bresolin N, Masliah E. The role of mitochondria in neurodegenerative diseases: the lesson from Alzheimer’s disease and Parkinson’s disease. Mol Neurobiol. 2020;57(7):2959–2980. doi:10.1007/s12035-020-01926-1
9. Cai Q, Jeong YY. Mitophagy in Alzheimer’s disease and other age-related neurodegenerative diseases. Cells. 2020;9(1):150. doi:10.3390/cells9010150
10. Fołta J, Rzepka Z, Wrześniok D. The role of inflammation in neurodegenerative diseases: Parkinson’s disease, Alzheimer’s disease, and multiple sclerosis. Int J Mol Sci. 2025;26(11):5177. doi:10.3390/ijms26115177
11. Ding L, Bian Q, Mou X, Chang X. Plant-derived exosome-like nanovesicles for CNS drug delivery and gut-brain axis modulation: a narrative review. Int J Nanomed. 2025;20:16093–16123. doi:10.2147/IJN.S562305
12. Hu E, Li Z, Li T, et al. A novel microbial and hepatic biotransformation-integrated network pharmacology strategy explores the therapeutic mechanisms of bioactive herbal products in neurological diseases: the effects of Astragaloside IV on intracerebral hemorrhage as an example. Chin Med. 2023;18(1):40. doi:10.1186/s13020-023-00745-5
13. Pardridge WM. The blood-brain barrier: bottleneck in brain drug development. Neurotherapeutics. 2005;2(1):3–14. doi:10.1602/neurorx.2.1.3
14. Zeng M, Guo D, Fernández-Varo G, et al. The integration of nanomedicine with traditional chinese medicine: drug delivery of natural products and other opportunities. Mol Pharm. 2023;20(2):886–904. doi:10.1021/acs.molpharmaceut.2c00882
15. Chen J, Tian C, Xiong X, Yang Y, Zhang J. Extracellular vesicles: new horizons in neurodegeneration. EBioMedicine. 2025;113:105605. doi:10.1016/j.ebiom.2025.105605
16. Huo L, Du X, Li X, Liu S, Xu Y. The emerging role of neural cell-derived exosomes in intercellular communication in health and neurodegenerative diseases. Front Neurosci. 2021;15:738442. doi:10.3389/fnins.2021.738442
17. Beatriz M, Vilaça R, Lopes C. Exosomes: innocent bystanders or critical culprits in neurodegenerative diseases. Front Cell Dev Biol. 2021;9:635104. doi:10.3389/fcell.2021.635104
18. Xu M, Feng T, Liu B, et al. Engineered exosomes: desirable target-tracking characteristics for cerebrovascular and neurodegenerative disease therapies. Theranostics. 2021;11(18):8926–8944. doi:10.7150/thno.62330
19. Bu CT, Zhu XD, Zhang QY, Shao WY. [Recent advances on the role of exosomes in neurodegenerative diseases]. Se Pu. 2025;43(5):487–497. Chinese. doi:10.3724/SP.J.1123.2024.10035
20. Sergazy S, Adekenov S, Khabarov I, Adekenova K, Maikenova A, Aljofan M. Harnessing mammalian- and plant-derived exosomes for drug delivery: a comparative review. Int J Mol Sci. 2025;26(10):4857. doi:10.3390/ijms26104857
21. Liu Y, Xiao S, Wang D, Qin C, Wei H, Li D. A review on separation and application of plant-derived exosome-like nanoparticles. J Sep Sci. 2024;47(8):e2300669. doi:10.1002/jssc.202300669
22. Liu M, Wang H, Yang Y, et al. Therapeutic potential of plant-derived exosome-like nanoparticles for CNS diseases: advancements in preparation and characterization. Ann Med. 2025;57(1):2537345. doi:10.1080/07853890.2025.2537345
23. Nueraihemaiti N, Dilimulati D, Baishan A, et al. Advances in plant-derived extracellular vesicle extraction methods and pharmacological effects. Biology. 2025;14(4):377. doi:10.3390/biology14040377
24. Kim J, Li S, Zhang S, Wang J. Plant-derived exosome-like nanoparticles and their therapeutic activities. Asian J Pharm Sci. 2022;17(1):53–69. doi:10.1016/j.ajps.2021.05.006
25. Sha A, Luo Y, Xiao W, et al. Plant-derived exosome-like nanoparticles: a comprehensive overview of their composition, biogenesis, isolation, and biological applications. Int J Mol Sci. 2024;25(22):12092. doi:10.3390/ijms252212092
26. Han X, Zheng W, Sun Z, et al. Plant-derived exosomes: unveiling the similarities and disparities between conventional extract and innovative form. Phytomedicine. 2025;145:157087. doi:10.1016/j.phymed.2025.157087
27. Isik S, Alhelwani S, Sahsahi A, Balcilar H, Yeman-Kiyak B. Plant-derived exosome-like nanovesicles: mechanisms and molecular understanding in neurological disorders with potential therapeutic applications. Drug Deliv Transl Res. 2025. doi:10.1007/s13346-025-01955-0
28. Cui W, Guo Z, Chen X, et al. Targeting modulation of intestinal flora through oral route by an antimicrobial nucleic acid-loaded exosome-like nanovesicles to improve Parkinson’s disease. Sci Bull. 2024;69(24):3925–3935. doi:10.1016/j.scib.2024.10.027
29. Cao M, Yan H, Han X, et al. Ginseng-derived nanoparticles alter macrophage polarization to inhibit melanoma growth. J Immunother Cancer. 2019;7(1):326. doi:10.1186/s40425-019-0817-4
30. Bahri F, Mansoori M, Vafaei S, et al. A comprehensive review on ginger-derived exosome-like nanoparticles as feasible therapeutic nano-agents against diseases. 2024. doi:10.1039/D3MA00856H.
31. Nelson VK, Begum MY, Dera AA, Ali SP, Suryadevara PR. Phytocompounds as modulators of HSF1 in neurodegenerative disease: special emphasis on Alzheimer’s disease. Eur J Pharmacol. 2025;1005:178034. doi:10.1016/j.ejphar.2025.178034
32. Lian MQ, Chng WH, Liang J, et al. Plant-derived extracellular vesicles: recent advancements and current challenges on their use for biomedical applications. J Extracell Vesicles. 2022;11(12):e12283. doi:10.1002/jev2.12283
33. Bai C, Liu J, Zhang X, et al. Research status and challenges of plant-derived exosome-like nanoparticles. Biomed Pharmacother. 2024;174:116543. doi:10.1016/j.biopha.2024.116543
34. Mu N, Li J, Zeng L, et al. Plant-derived exosome-like nanovesicles: current progress and prospects. Int J Nanomed. 2023;18:4987–5009. doi:10.2147/IJN.S420748
35. Zhang M, Viennois E, Prasad M, et al. Edible ginger-derived nanoparticles: a novel therapeutic approach for the prevention and treatment of inflammatory bowel disease and colitis-associated cancer. Biomaterials. 2016;101:321–340. doi:10.1016/j.biomaterials.2016.06.018
36. Welsh JA, Goberdhan DCI, O’Driscoll L, et al. Minimal information for studies of extracellular vesicles (MISEV2023): from basic to advanced approaches. J Extracell Vesicle. 2024;13(2):e12404. doi:10.1002/jev2.12404
37. Woith E, Guerriero G, Hausman JF, et al. Plant extracellular vesicles and nanovesicles: focus on secondary metabolites, proteins and lipids with perspectives on their potential and sources. IJMS. 2021;22(7):3719. doi:10.3390/ijms22073719
38. Ou X, Wang H, Tie H, et al. Novel plant-derived exosome-like nanovesicles from Catharanthus roseus: preparation, characterization, and immunostimulatory effect via TNF-α/NF-κB/PU.1 axis. J Nanobiotechnology. 2023;21(1):160. doi:10.1186/s12951-023-01919-x
39. Skotland T, Sandvig K, Llorente A. Lipids in exosomes: current knowledge and the way forward. Prog lipid res. 2017;66:30–41. doi:10.1016/j.plipres.2017.03.001
40. Cai Q, Qiao L, Wang M, et al. Plants send small RNAs in extracellular vesicles to fungal pathogen to silence virulence genes. Science. 2018;360(6393):1126–1129. doi:10.1126/science.aar4142
41. Zhang M, Xiao B, Wang H, et al. Edible ginger-derived nano-lipids loaded with doxorubicin as a novel drug-delivery approach for colon cancer therapy. Mol Ther. 2016;24(10):1783–1796. doi:10.1038/mt.2016.159
42. Wang F, Li L, Deng J, et al. Lipidomic analysis of plant-derived extracellular vesicles for guidance of potential anti-cancer therapy. Bioact Mater. 2025;46:82–96. doi:10.1016/j.bioactmat.2024.12.001
43. Wang Q, Zhuang X, Mu J, et al. Delivery of therapeutic agents by nanoparticles made of grapefruit-derived lipids. Nat Commun. 2013;4(1):1867. doi:10.1038/ncomms2886
44. Wang Q, Ren Y, Mu J, et al. Grapefruit-derived nanovectors use an activated leukocyte trafficking pathway to deliver therapeutic agents to inflammatory tumor sites. Cancer Res. 2015;75(12):2520–2529. doi:10.1158/0008-5472.CAN-14-3095
45. Rutter BD, Innes RW. Extracellular vesicles isolated from the leaf apoplast carry stress-response proteins. Plant Physiol. 2017;173(1):728–741. doi:10.1104/pp.16.01253
46. Zhan M, Cai Y, Zhao C, et al. Citrus exosome-like nanoparticles reversed high-fat diet-induced microbiota dysbiosis and associated short-chain fatty acid production. J Agric Food Chem. 2025;73(37):23344–23356. doi:10.1021/acs.jafc.5c04523
47. Jo HY, Kang SJ, Kim G, Gwak S, Baek G, Rhee WJ. Plant-derived extracellular vesicles: current status and challenges for developing a new paradigm in therapeutics development. VIEW. 2025;6(1):20240115. doi:10.1002/VIW.20240115
48. Stanly C, Moubarak M, Fiume I, Turiák L, Pocsfalvi G. Membrane transporters in citrus clementina fruit juice-derived nanovesicles. IJMS. 2019;20(24):6205. doi:10.3390/ijms20246205
49. Tóth EÁ, Turiák L, Visnovitz T, et al. Formation of a protein Corona on the surface of extracellular vesicles in blood plasma. J Extracell Vesicle. 2021;10(11):e12140. doi:10.1002/jev2.12140
50. Dietz L, Oberländer J, Mateos-Maroto A, et al. Uptake of extracellular vesicles into immune cells is enhanced by the protein Corona. J Extracell Vesicle. 2023;12(12):e12399. doi:10.1002/jev2.12399
51. Che K, Wang C, Chen H. Advancing functional foods: a systematic analysis of plant-derived exosome-like nanoparticles and their health-promoting properties. Front Nutr. 2025;12:1544746. doi:10.3389/fnut.2025.1544746
52. Baldrich P, Rutter BD, Karimi HZ, Podicheti R, Meyers BC, Innes RW. Plant extracellular vesicles contain diverse small RNA species and are enriched in 10- to 17-Nucleotide “Tiny” RNAs. Plant Cell. 2019;31(2):315–324. doi:10.1105/tpc.18.00872
53. He B, Cai Q, Qiao L, et al. RNA-binding proteins contribute to small RNA loading in plant extracellular vesicles. Nat Plants. 2021;7(3):342–352. doi:10.1038/s41477-021-00863-8
54. Xiao J, Feng S, Wang X, et al. Identification of exosome-like nanoparticle-derived microRNAs from 11 edible fruits and vegetables. PeerJ. 2018;6:e5186. doi:10.7717/peerj.5186
55. Shkryl Y, Tsydeneshieva Z, Menchinskaya E, et al. Exosome-like nanoparticles, high in Trans-δ-Viniferin derivatives, produced from grape cell cultures: preparation, characterization, and anticancer properties. Biomedicines. 2024;12(9):2142. doi:10.3390/biomedicines12092142
56. Kathait P, Patel PK, Sahu AN. Harnessing exosomes and plant-derived exosomes as nanocarriers for the efficient delivery of plant bioactives. Nanomedicine. 2024;19(30):2679–2697. doi:10.1080/17435889.2024.2354159
57. Jiang D, Li Z, Liu H, Liu H, Xia X, Xiang X. Plant exosome-like nanovesicles derived from sesame leaves as carriers for luteolin delivery: molecular docking, stability and bioactivity. Food Chem. 2024;438:137963. doi:10.1016/j.foodchem.2023.137963
58. Kilasoniya A, Garaeva L, Shtam T, et al. Potential of Plant Exosome Vesicles from Grapefruit (Citrus × paradisi) and Tomato (Solanum lycopersicum) juices as functional ingredients and targeted drug delivery vehicles. Antioxidants. 2023;12(4):943. doi:10.3390/antiox12040943
59. Song Y, Feng N, Yu Q, et al. Exosomes in disease therapy: plant-derived exosome-like nanoparticles current status, challenges, and future prospects. Int J Nanomed. 2025;20:10613–10644. doi:10.2147/IJN.S540094
60. Liu X, Mei L, Wang J, et al. Cutting-edge insights into the mechanistic understanding of plant-derived exosome-like nanoparticles: implications for intestinal homeostasis. Food Res Int. 2025;208:116186. doi:10.1016/j.foodres.2025.116186
61. Mu J, Zhuang X, Wang Q, et al. Interspecies communication between plant and mouse gut host cells through edible plant derived exosome-like nanoparticles. Mol Nutr Food Res. 2014;58(7):1561–1573. doi:10.1002/mnfr.201300729
62. Ju S, Mu J, Dokland T, et al. Grape exosome-like nanoparticles induce intestinal stem cells and protect mice from DSS-induced colitis. Mol Ther. 2013;21(7):1345–1357. doi:10.1038/mt.2013.64
63. Tembo KM, Wang X, Bolideei M, et al. Exploring the bioactivity of MicroRNAs Originated from Plant-derived Exosome-like Nanoparticles (PELNs): current perspectives. J Nanobiotechnology. 2025;23(1):563. doi:10.1186/s12951-025-03602-9
64. Teng Y, Ren Y, Sayed M, et al. Plant-derived exosomal MicroRNAs shape the gut microbiota. Cell Host Microbe. 2018;24(5):637–652.e8. doi:10.1016/j.chom.2018.10.001
65. Luo Y, Liu K, Ling D, Gu T, Zhang L, Chen W. Isolation and characterization of exosomes derived from mouse spleen tissues. JoVE. 2024;(211):67234. doi:10.3791/67234
66. Mun JG, Song DH, Kee JY, Han Y. Recent advances in the isolation strategies of plant-derived exosomes and their therapeutic applications. Curr Issues Mol Biol. 2025;47(3):144. doi:10.3390/cimb47030144
67. Cao M, Diao N, Cai X, et al. Plant exosome nanovesicles (PENs): green delivery platforms. Mater Horiz. 2023;10(10):3879–3894. doi:10.1039/d3mh01030a
68. Wang W, Sun H, Duan H, et al. Isolation and usage of exosomes in central nervous system diseases. CNS Neurosci Ther. 2024;30(3):e14677. doi:10.1111/cns.14677
69. Fu S, Zhu L, Yang X, Jiao Y, Hao G, Liu Y. Extracellular vesicles separated from goat milk by differential centrifugation coupled with sodium citrate pretreatments. Food Chem. 2024;446:138807. doi:10.1016/j.foodchem.2024.138807
70. Huang K, Garimella S, Clay-Gilmour A, et al. Comparison of human urinary exosomes isolated via ultracentrifugation alone versus ultracentrifugation followed by SEC Column-Purification. J Pers Med. 2022;12(3):340. doi:10.3390/jpm12030340
71. Van Deun J, Mestdagh P, Sormunen R, et al. The impact of disparate isolation methods for extracellular vesicles on downstream RNA profiling. J Extracell Vesicle. 2014;3(1):24858. doi:10.3402/jev.v3.24858
72. Kaddour H, Tranquille M, Okeoma CM. The past, the present, and the future of the size exclusion chromatography in extracellular vesicles separation. Viruses. 2021;13(11):2272. doi:10.3390/v13112272
73. Mukerjee N, Bhattacharya A, Maitra S, et al. Exosome isolation and characterization for advanced diagnostic and therapeutic applications. Mater Today Bio. 2025;31:101613. doi:10.1016/j.mtbio.2025.101613
74. Marsh SR, Pridham KJ, Jourdan J, Gourdie RG. Novel protocols for scalable production of high quality purified small extracellular vesicles from bovine milk. Nanotheranostics. 2021;5(4):488–498. doi:10.7150/ntno.62213
75. Gurriaran-Rodriguez U, De Repentigny Y, Kothary R, Rudnicki MA. Isolation of small extracellular vesicles from regenerating muscle tissue using tangential flow filtration and size exclusion chromatography. Skelet Muscle. 2024;14(1):22. doi:10.1186/s13395-024-00355-1
76. Visan KS, Lobb RJ, Ham S, et al. Comparative analysis of tangential flow filtration and ultracentrifugation, both combined with subsequent size exclusion chromatography, for the isolation of small extracellular vesicles. J Extracell Vesicle. 2022;11(9):12266. doi:10.1002/jev2.12266
77. Zou Z, Li H, Xu G, Hu Y, Zhang W, Tian K. Current knowledge and future perspectives of exosomes as nanocarriers in diagnosis and treatment of diseases. Int J Nanomed. 2023;18:4751–4778. doi:10.2147/IJN.S417422
78. Lo KJ, Wang MH, Kuo CY, Pan MH. Optimizing isolation methods and exploring the therapeutic potential of lotus-derived extracellular vesicles in modulating inflammation and promoting wound healing. ACS Biomater Sci Eng. 2025;11(7):4424–4435. doi:10.1021/acsbiomaterials.5c00377
79. Busatto S, Vilanilam G, Ticer T, et al. Tangential flow filtration for highly efficient concentration of extracellular vesicles from large volumes of fluid. Cells. 2018;7(12):273. doi:10.3390/cells7120273
80. Zanotti C, Arena S, De Pascale S, et al. Anion exchange chromatography-based purification of plant-derived nanovesicles from Brassica oleracea L.: molecular profiling and bioactivity in human cells. Front Bioeng Biotechnol. 2025;13:1617478. doi:10.3389/fbioe.2025.1617478
81. Wysor SK, Marcus RK. In-line coupling of capillary-channeled polymer fiber columns with optical absorbance and multi-angle light scattering detection for the isolation and characterization of exosomes. Anal Bioanal Chem. 2024;416(14):3325–3333. doi:10.1007/s00216-024-05283-z
82. Zhu L, Du J, Cheng X, et al. Microfluidic innovations for enhanced extracellular vesicle isolation and analysis: a comprehensive review. Anal Chem. 2025;97(9):4695–4705. doi:10.1021/acs.analchem.4c05801
83. Ni F, Zhu Q, Li H, Liu F, Chen H. Efficient preparation of high-purity and intact mesenchymal stem cell-derived extracellular vesicles. Anal Bioanal Chem. 2024;416(8):1797–1808. doi:10.1007/s00216-024-05193-0
84. Feng H, Yue Y, Zhang Y, et al. Plant-derived exosome-like nanoparticles: emerging nanosystems for enhanced tissue engineering. Int J Nanomed. 2024;19:1189–1204. doi:10.2147/IJN.S448905
85. Cook K, Li H. Advancing extracellular vesicle production: improving physiological relevance and yield with 3D cell culture. Nanoscale. 2025;17(25):15110–15131. doi:10.1039/D5NR00707K
86. Pocsfalvi G, Stanly C, Vilasi A, et al. Mass spectrometry of extracellular vesicles. Mass Spectrom Rev. 2016;35(1):3–21. doi:10.1002/mas.21457
87. Rawat S, Arora S, Dhondale MR, Khadilkar M, Kumar S, Agrawal AK. Stability dynamics of plant-based extracellular vesicles drug delivery. JoX. 2025;15(2):55. doi:10.3390/jox15020055
88. Zhang J, Tian X, Li Y, et al. Stem cell-derived exosomes: a comprehensive review of biomedical applications, challenges, and future directions. Int J Nanomed. 2025;20:10857–10905. doi:10.2147/IJN.S527137
89. Wei Y, Cai X, Wu Q, et al. Extraction, isolation, and component analysis of turmeric-derived exosome-like nanoparticles. Bioengineering. 2023;10(10):1199. doi:10.3390/bioengineering10101199
90. Shen C, Li X, Qin J, Duan L. Characterization of miRNA profiling in konjac-derived exosome-like nanoparticles and elucidation of their multifaceted roles in human health. Front Plant Sci. 2024;15:1444683. doi:10.3389/fpls.2024.1444683
91. Taşlı PN. Usage of celery root exosome as an immune suppressant; Lipidomic characterization of Apium graveolens originated exosomes and its suppressive effect on PMA/ionomycin mediated CD4+ T lymphocyte activation. J Food Biochem. 2022;46(12):e14393. doi:10.1111/jfbc.14393
92. Li T, Xiao L, Ling H, Yang Y, Zhong S. Preparation of artificial substrate binding sites of nanozyme with “modular structure” strategy used for the construction of visual sensing analysis platform for levodopa. Microchem J. 2025;212:113292. doi:10.1016/j.microc.2025.113292
93. Wu MY, Tian YQ, Lu JY, Fu TT, Liang HC, Huang WT. Synthesis of sustainable chromium-based nanoparticles with fluorescence tunable ability for biosensing of tumor-derived exosomes and molecular information protection. SusMat. 2026;6(1):e70053. doi:10.1002/sus2.70053
94. Ning H, Huang X, Deng N, et al. Plant-derived exosome-like nanovesicles: a novel strategy for targeted oral therapy in ulcerative colitis. Int J Nanomed. 2025;20:10595–10611. doi:10.2147/IJN.S536056
95. Ramírez O, Pomareda F, Olivares B, et al. Aloe vera peel-derived nanovesicles display anti-inflammatory properties and prevent myofibroblast differentiation. Phytomedicine. 2024;122:155108. doi:10.1016/j.phymed.2023.155108
96. Li S, Ye Z, Zhao L, Yao Y, Zhou Z. Evaluation of antioxidant activity and drug delivery potential of cell-derived extracellular vesicles from Citrus reticulata Blanco cv. “Dahongpao. Antioxidants. 2023;12(9):1706. doi:10.3390/antiox12091706
97. Shen H, Zhang M, Liu D, et al. Solanum lycopersicum derived exosome-like nanovesicles alleviate restenosis after vascular injury through the Keap1/Nrf2 pathway. Food Funct. 2025;16(2):539–553. doi:10.1039/d4fo03993a
98. Trentini M, Zanolla I, Tiengo E, et al. Link between organic nanovescicles from vegetable kingdom and human cell physiology: intracellular calcium signalling. J Nanobiotechnology. 2024;22(1):68. doi:10.1186/s12951-024-02340-8
99. Kim JS, Song BJ, Cho YE. Pomegranate-derived exosome-like nanovesicles containing ellagic acid alleviate gut leakage and liver injury in MASLD. Food Sci Nutr. 2025;13(4):e70088. doi:10.1002/fsn3.70088
100. Yu Y, Xu Z, Xu L, Lu D, Tang Y, Mai H. Plant extracellular vesicles as emerging neuroprotective agents for central nervous system disorders. J Adv Res. 2025;S2090–1232(25)00202–4. doi:10.1016/j.jare.2025.03.042
101. Zhao D, Yang X, Kelimu A, et al. Preparation and characterization of cyperus-derived exosomes loaded with selenium nanoparticles for selenium delivery based on exosome protein quantitation. Foods. 2025;14(15):2724. doi:10.3390/foods14152724
102. Wang R, Zhang Y, Guo Y, et al. Plant-derived nanovesicles: promising therapeutics and drug delivery nanoplatforms for brain disorders. Fundam Res. 2025;5(2):830–850. doi:10.1016/j.fmre.2023.09.007
103. Liu S, Jin X, Ge Y, et al. Advances in brain-targeted delivery strategies and natural product-mediated enhancement of blood–brain barrier permeability. J Nanobiotechnol. 2025;23(1):382. doi:10.1186/s12951-025-03415-w
104. Zheng M, Hong X, Liao P, et al. Plant-derived exosome-like nanoparticles: a promising therapeutic for neurological disorders and drug delivery. Int J Nanomed. 2025;20:15769–15791. doi:10.2147/IJN.S572417
105. Banks WA, Sharma P, Bullock KM, Hansen KM, Ludwig N, Whiteside TL. Transport of extracellular vesicles across the blood-brain barrier: brain pharmacokinetics and effects of inflammation. IJMS. 2020;21(12):4407. doi:10.3390/ijms21124407
106. Sun M, Qin F, Bu Q, et al. Exosome-based therapeutics: a natural solution to overcoming the blood-brain barrier in neurodegenerative diseases. MedComm. 2025;6(9):e70386. doi:10.1002/mco2.70386
107. Huang Q, Wang S, Liu Z, Rao L, Cheng K, Mao X. Engineering exosomes for targeted neurodegenerative therapy: innovations in biogenesis, drug loading, and clinical translation. Theranostics. 2026;16(1):545–579. doi:10.7150/thno.117143
108. Jiang A, Nie W, Xie HY. In vivo imaging for the visualization of extracellular vesicle-based tumor therapy. ChemistryOpen. 2022;11(9):e202200124. doi:10.1002/open.202200124
109. Lu Y, Xu J, Tang R, et al. Edible Pueraria lobata-derived exosome-like nanovesicles ameliorate dextran sulfate sodium-induced colitis associated lung inflammation through modulating macrophage polarization. Biomed Pharmacother. 2024;170:116098. doi:10.1016/j.biopha.2023.116098
110. Shi R, Tan W, Jin H, et al. MicroRNA-enriched plant-derived exosomes alleviate colitis by modulating systemic immunity, metabolic homeostasis, and gut microbiota. Adv Sci. 2025:e05921. doi:10.1002/advs.202505921
111. Kang M, Kang M, Lee J, Yoo J, Lee S, Oh S. Allium tuberosum -derived nanovesicles with anti-inflammatory properties prevent DSS-induced colitis and modify the gut microbiome. Food Funct. 2024;15(14):7641–7657. doi:10.1039/D4FO01366B
112. Zhuang X, Teng Y, Samykutty A, et al. Grapefruit-derived nanovectors delivering therapeutic miR17 through an intranasal route inhibit brain tumor progression. Mol Ther. 2016;24(1):96–105. doi:10.1038/mt.2015.188
113. Baruah H, Sarma A, Basak D, Das M. Exosome: from biology to drug delivery. Drug Deliv Transl Res. 2024;14(6):1480–1516. doi:10.1007/s13346-024-01515-y
114. Liu W, Yin X, Zhang Z, et al. Plant-derived exosome-like nanoparticles: mechanisms of cross-kingdom regulation and perspectives as natural drug carriers for disease treatment. IJN. 2026;21:1–22. doi:10.2147/IJN.S581774
115. Ishida T, Kawada K, Jobu K, et al. Exosome-like nanoparticles derived from Allium tuberosum prevent neuroinflammation in microglia-like cells. J Pharm Pharmacol. 2023;75(10):1322–1331. doi:10.1093/jpp/rgad062
116. Chen X, Zhou Y, Yu J. Exosome-like nanoparticles from ginger rhizomes inhibited NLRP3 inflammasome activation. Mol Pharm. 2019;16(6):2690–2699. doi:10.1021/acs.molpharmaceut.9b00246
117. Xu Y, Yan G, Zhao J, et al. Plant-derived exosomes as cell homogeneous nanoplatforms for brain biomacromolecules delivery ameliorate mitochondrial dysfunction against Parkinson’s disease. Nano Today. 2024;58:102438. doi:10.1016/j.nantod.2024.102438
118. Agrawal M, Ajazuddin, Tripathi DK, et al. Recent advancements in liposomes targeting strategies to cross blood-brain barrier (BBB) for the treatment of Alzheimer’s disease. J Control Release. 2017;260:61–77. doi:10.1016/j.jconrel.2017.05.019
119. Lee J, Kim H, Kim J, et al. A novel treatment strategy by natural products in NLRP3 inflammasome-mediated neuroinflammation in Alzheimer’s and Parkinson’s disease. IJMS. 2021;22(3):1324. doi:10.3390/ijms22031324
120. Zhang H, Wang L, Zhu B, et al. A comparative study of the neuroprotective effects of dl-3-n-butylphthalide and edaravone dexborneol on cerebral ischemic stroke rats. Eur J Pharmacol. 2023;951:175801. doi:10.1016/j.ejphar.2023.175801
121. Zhang H, Zhu C, Zhou X, et al. Edaravone dexborneol protected neurological function by targeting NRF2/ARE and NF-κB/AIM2 pathways in cerebral ischemia/reperfusion injury. Front Pharmacol. 2025;16:1581320. doi:10.3389/fphar.2025.1581320
122. Zhang Y, Lu L, Li Y, Liu H, Zhou W, Zhang L. Response surface methodology optimization of exosome-like nanovesicles extraction from Lycium ruthenicum murray and their inhibitory effects on Aβ-induced apoptosis and oxidative stress in HT22 cells. Foods. 2024;13(20):3328. doi:10.3390/foods13203328
123. Zhang Y, Zhang X, Kai T, Zhang L, Li A. Lycium ruthenicum Murray derived exosome-like nanovesicles inhibit Aβ-induced apoptosis in PC12 cells via MAPK and PI3K/AKT signaling pathways. Int J Biol Macromol. 2024;277(Pt 2):134309. doi:10.1016/j.ijbiomac.2024.134309
124. Wegrzynowicz M, Bar-On D, Calo’ L, et al. Depopulation of dense α-synuclein aggregates is associated with rescue of dopamine neuron dysfunction and death in a new Parkinson’s disease model. Acta Neuropathol. 2019;138(4):575–595. doi:10.1007/s00401-019-02023-x
125. Sataer X, Qifeng Z, Yingying Z, et al. Exosomal microRNAs as diagnostic biomarkers and therapeutic applications in neurodegenerative diseases. Neurol Res. 2023;45(3):191–199. doi:10.1080/01616412.2022.2129768
126. Etxebeste-Mitxeltorena M, Niza E, Fajardo CM, et al. Neuroprotective properties of exosomes and chitosan nanoparticles of Tomafran, a bioengineered tomato enriched in crocins. Nat Prod Bioprospect. 2024;14(1):9. doi:10.1007/s13659-023-00425-9
127. Yuyama K, Igarashi Y. Linking glycosphingolipids to Alzheimer’s amyloid-ß: extracellular vesicles and functional plant materials. Glycoconjugate J. 2022;39(5):613–618. doi:10.1007/s10719-022-10066-8
128. Yuyama K, Takahashi K, Usuki S, et al. Plant sphingolipids promote extracellular vesicle release and alleviate amyloid-β pathologies in a mouse model of Alzheimer’s disease. Sci Rep. 2019;9(1):16827. doi:10.1038/s41598-019-53394-w
129. Yang D, Deng Z, Zhou H, Zhang Q, Zhang X, Gong J. Exosome-mediated dual drug delivery of curcumin and methylene blue for enhanced cognitive function and mechanistic elucidation in Alzheimer’s disease therapy. Front Cell Dev Biol. 2025;13:1562565. doi:10.3389/fcell.2025.1562565
130. Yan D, Wang N, Yao J, Wu X, Yuan J, Yan H. Curcumin attenuates the PERK-eIF2α signaling to relieve acrylamide-induced neurotoxicity in SH‑SY5Y neuroblastoma cells. Neurochem Res. 2022;47(4):1037–1048. doi:10.1007/s11064-021-03504-w
131. Yan D, Yao J, Liu Y, et al. Tau hyperphosphorylation and P-CREB reduction are involved in acrylamide-induced spatial memory impairment: suppression by curcumin. Brain Behav Immun. 2018;71:66–80. doi:10.1016/j.bbi.2018.04.014
132. Ehrnhoefer DE, Bieschke J, Boeddrich A, et al. EGCG redirects amyloidogenic polypeptides into unstructured, off-pathway oligomers. Nat Struct Mol Biol. 2008;15(6):558–566. doi:10.1038/nsmb.1437
133. Bieschke J, Russ J, Friedrich RP, et al. EGCG remodels mature alpha-synuclein and amyloid-beta fibrils and reduces cellular toxicity. Proc Natl Acad Sci U S A. 2010;107(17):7710–7715. doi:10.1073/pnas.0910723107
134. Sonawane SK, Chidambaram H, Boral D, et al. EGCG impedes human Tau aggregation and interacts with Tau. Sci Rep. 2020;10(1):12579. doi:10.1038/s41598-020-69429-6
135. Guéroux M, Fleau C, Slozeck M, Laguerre M, Pianet I. Epigallocatechin 3-Gallate as an inhibitor of tau phosphorylation and aggregation: a molecular and structural insight. J Prev Alzheimers Dis. 2017;4(4):218–225. doi:10.14283/jpad.2017.35
136. Xu XH, Yuan TJ, Dad HA, et al. Plant exosomes as novel nanoplatforms for MicroRNA transfer stimulate neural differentiation of stem cells in vitro and in vivo. Nano Lett. 2021;21(19):8151–8159. doi:10.1021/acs.nanolett.1c02530
137. Wang Q, Huang Z, Guo J, et al. Plant-derived vesicle-like nanoparticles: pioneering sustainable and effective approaches for tissue repair and regeneration. Biomolecules. 2025;15(8):1055. doi:10.3390/biom15081055
138. Wang Q, Liu K, Cao X, et al. Plant-derived exosomes extracted from Lycium barbarum L. loaded with isoliquiritigenin to promote spinal cord injury repair based on 3D printed bionic scaffold. Bioeng Transl Med. 2024;9(4):e10646. doi:10.1002/btm2.10646
139. Jiang HF, Guo YQ, Rehman FU, Jing L, Zhang JZ. Potential cerebrovascular protective functions of lycium barbarum polysaccharide in alleviating hyperglycemia-aggravated cerebral ischemia/reperfusion injury in hyperglycemic rats. Eur Rev Med Pharmacol Sci. 2022;26(20):7379–7394. doi:10.26355/eurrev_202210_30007
140. Yu Y, Wu X, Pu J, et al. Lycium barbarum polysaccharide protects against oxygen glucose deprivation/reoxygenation-induced apoptosis and autophagic cell death via the PI3K/Akt/mTOR signaling pathway in primary cultured hippocampal neurons. Biochem Biophys Res Commun. 2018;495(1):1187–1194. doi:10.1016/j.bbrc.2017.11.165
141. Balusamy SR, Perumalsamy H, Huq MDA, et al. A comprehensive and systemic review of ginseng-based nanomaterials: synthesis, targeted delivery, and biomedical applications. Med Res Rev. 2023;43(5):1374–1410. doi:10.1002/med.21953
142. Kalani A, Chaturvedi P, Kamat PK, et al. Curcumin-loaded embryonic stem cell exosomes restored neurovascular unit following ischemia-reperfusion injury. Int J Biochem Cell Biol. 2016;79:360–369. doi:10.1016/j.biocel.2016.09.002
143. Hu E, Li T, Li Z, et al. Metabolomics reveals the effects of hydroxysafflor yellow A on neurogenesis and axon regeneration after experimental traumatic brain injury. Pharm Biol. 2023;61(1):1054–1064. doi:10.1080/13880209.2023.2229379
144. Ferreira ND, Ferreira PS, Pacheco Soares C, Aparecida Porcionatto M, Rodrigues Salles G. 3D-bioprinted model of adult neural stem cell microenvironment in Alzheimer’s disease. IJB. 2024;10:3751. doi:10.36922/ijb.3751
145. Liu D, Gao J, Wu X, Hao X, Hu W, Han L. Conductive microneedles loaded with polyphenol-engineered exosomes reshape diabetic neurovascular niches for chronic wound healing. Adv Sci. 2025;e07974. doi:10.1002/advs.202507974
146. Luo S, Sun X, Huang M, Ma Q, Du L, Cui Y. Enhanced neuroprotective effects of epicatechin gallate encapsulated by bovine milk-derived exosomes against Parkinson’s disease through antiapoptosis and antimitophagy. J Agric Food Chem. 2021;69(17):5134–5143. doi:10.1021/acs.jafc.0c07658
147. Bates GP, Dorsey R, Gusella JF, et al. Huntington disease. Nat Rev Dis Primers. 2015;1(1):15005. doi:10.1038/nrdp.2015.5
148. Li Y, Wang Y, Zhao H, Pan Q, Chen G. Engineering strategies of plant-derived exosome-like nanovesicles: current knowledge and future perspectives. Int J Nanomed. 2024;19:12793–12815. doi:10.2147/IJN.S496664
149. Langellotto MD, Rassu G, Serri C, Demartis S, Giunchedi P, Gavini E. Plant-derived extracellular vesicles: a synergetic combination of a drug delivery system and a source of natural bioactive compounds. Drug Deliv Transl Res. 2025;15(3):831–845. doi:10.1007/s13346-024-01698-4
150. Hardiman O, Al-Chalabi A, Chio A, et al. Amyotrophic lateral sclerosis. Nat Rev Dis Primers. 2017;3(1):17071. doi:10.1038/nrdp.2017.71
151. Guy R, Offen D. Promising opportunities for treating neurodegenerative diseases with mesenchymal stem cell-derived exosomes. Biomolecules. 2020;10(9):1320. doi:10.3390/biom10091320
152. Varshney V, Gabble BC, Bishoyi AK, et al. Exploring exosome-based approaches for early diagnosis and treatment of neurodegenerative diseases. Mol Neurobiol. 2025;62(11):14683–14705. doi:10.1007/s12035-025-05026-w
153. Wilson DM, Cookson MR, Van Den Bosch L, Zetterberg H, Holtzman DM, Dewachter I. Hallmarks of neurodegenerative diseases. Cell. 2023;186(4):693–714. doi:10.1016/j.cell.2022.12.032
154. Kahroba H, Davatgaran-Taghipour Y. Exosomal Nrf2: from anti-oxidant and anti-inflammation response to wound healing and tissue regeneration in aged-related diseases. Biochimie. 2020;171–172:103–109. doi:10.1016/j.biochi.2020.02.011
155. Li R, Duan W, Feng T, et al. Lycium barbarum polysaccharide inhibits ischemia-induced autophagy by promoting the biogenesis of neural stem cells-derived extracellular vesicles to enhance the delivery of miR-133a-3p. Chin Med. 2023;18(1):117. doi:10.1186/s13020-023-00831-8
156. Weng J, Chen Y, Zeng Y, et al. A novel hydrogel loaded with plant exosomes and stem cell exosomes as a new strategy for treating diabetic wounds. Mater Today Bio. 2025;32:101810. doi:10.1016/j.mtbio.2025.101810
157. Wu P, Wu W, Zhang S, et al. Therapeutic potential and pharmacological significance of extracellular vesicles derived from traditional medicinal plants. Front Pharmacol. 2023;14:1272241. doi:10.3389/fphar.2023.1272241
158. Barzin M, Bagheri AM, Ohadi M, Abhaji AM, Salarpour S, Dehghannoudeh G. Application of plant-derived exosome-like nanoparticles in drug delivery. Pharm Dev Technol. 2023;28(5):383–402. doi:10.1080/10837450.2023.2202242
159. Lu X, Han Q, Chen J, et al. Celery (Apium graveolens L.) exosome-like nanovesicles as a new-generation chemotherapy drug delivery platform against tumor proliferation. J Agric Food Chem. 2023;71(22):8413–8424. doi:10.1021/acs.jafc.2c07760
160. Salem D, Abdel-Ghany S, Mohamed E, et al. Natural nanoparticles for drug delivery: proteomic insights and anticancer potential of doxorubicin-loaded avocado exosomes. Pharmaceuticals. 2025;18(6):844. doi:10.3390/ph18060844
161. Wang H, Tang C, Zhang H, et al. A biocompatible tea polyphenol nanoplatform for efficient cytosolic delivery of protein therapeutics. J Control Release. 2025;388:114347. doi:10.1016/j.jconrel.2025.114347
162. Liang Q, Zeng J, Jiang Z, et al. Multivoid magnetic nanoparticles as high-performance magnetic particle imaging tracers for precise glioma detection. Bioconjugate Chem. 2026;37(1):118–127. doi:10.1021/acs.bioconjchem.5c00527
163. Orefice NS, Di Raimo R, Mizzoni D, Logozzi M, Fais S. Purposing plant-derived exosomes-like nanovesicles for drug delivery: patents and literature review. Expert Opin Ther Pat. 2023;33(2):89–100. doi:10.1080/13543776.2023.2195093
164. Cai J, Pan J. Beta vulgaris-derived exosome-like nanovesicles alleviate chronic doxorubicin-induced cardiotoxicity by inhibiting ferroptosis. J Biochem Mol Toxicol. 2024;38(1):e23540. doi:10.1002/jbt.23540
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.