Back to Journals » Drug Design, Development and Therapy » Volume 20
PJ34 Prevents Trauma-Induced Heterotopic Ossification without Adverse Bone Healing: An in vivo and in vitro Investigation
Authors Zhang W, Hou J, Feng H, Zhu J, Liang Y, Mu W, Xu P, Qian Y ![]() , Jiang S
, Jiang S
Received 24 September 2025
Accepted for publication 16 February 2026
Published 23 March 2026 Volume 2026:20 561571
DOI https://doi.org/10.2147/DDDT.S561571
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Tuo Deng
Weiying Zhang,1,* Jing Hou,2,* Hao Feng,3,* Jingshu Zhu,4,* Yuqing Liang,5 Weidong Mu,6 Peng Xu,6 Yun Qian,7 Shichao Jiang6
1Health Management Center, Shandong Provincial Hospital Affiliated to Shandong First Medical University, Jinan, Shandong, People’s Republic of China; 2Department of Rehabilitation Medicine, Tongliao People’s Hospital, Tongliao, Inner Mongolia, China; 3State Key Laboratory for Modification of Chemical Fibers and Polymer Materials, Shanghai Engineering Research Center of Nano-Biomaterials and Regenerative Medicine, College of Biological Science and Medical Engineering, Donghua University, Shanghai, People’s Republic of China; 4Alberta Institute, Wenzhou Medical University, Wenzhou, Zhejiang, People’s Republic of China; 5Department of Sports Medicine, Qingdao Traditional Chinese Medicine Hospital, Qingdao Hiser Hospital Affiliated of Qingdao University, Qingdao, Shandong, People’s Republic of China; 6Department of Orthopedics, Shandong Provincial Hospital Affiliated to Shandong First Medical University, Jinan, Shandong, People’s Republic of China; 7National Center for Orthopaedics, Shanghai Sixth People’s Hospital Affiliated to Shanghai Jiao Tong University School of Medicine, Shanghai, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Shichao Jiang, Email [email protected] Yun Qian, Email [email protected]
Introduction: Trauma-induced heterotopic ossification (THO) is a complication characterized by ectopic lamellar bone formation in soft tissues after trauma. Current treatments are limited by low efficiency, recurrence, and potential impairment of bone healing, highlighting the need for new strategies. Mesenchymal stem cell (MSC) recruitment and osteogenic differentiation are critical in THO pathogenesis. Poly [ADP-ribose] polymerase 1 (PARP1) is involved in regulating osteogenic differentiation, and PJ34, a PARP1 inhibitor, may have potential in THO prevention.
Methods: This study investigated PJ34’s effects on THO and bone healing through in vitro and in vivo experiments. In vitro experiments, the effects of different concentrations of PJ34 on the proliferation, recruitment, migration, alkaline phosphatase (ALP) activity, mineralization and the expression of osteogenesis-related genes of bone marrow stromal cells (BMSCs) were assessed. In vivo experiments involved constructing mouse THO models and tibial fracture models to evaluate the impact of PJ34 on ectopic bone formation and fracture healing.
Results: In vitro, PJ34 dose-dependently inhibited BMSC proliferation, recruitment, migration, alkaline phosphatase (ALP) activity, and mineralization. In vivo, PJ34 significantly reduced heterotopic bone formation in a mouse THO model without impairing fracture healing. PJ34 also downregulated the expression of osteogenic genes (Runx2, BMP-2, ALP, OPN, BGLAP) in BMSCs.
Conclusion: These findings preliminarily indicate that PJ34 prevents THO by inhibiting proliferation, migration, and osteogenic differentiation of BMSC, without adverse effects on normal bone healing. This supports PJ34’s potential as a novel therapeutic agent for THO prevention, offering a safer alternative to current treatments, though further mechanistic validation is needed.
Keywords: trauma-induced heterotopic ossification, PARP1, PJ34, bone healing, BMSC
Introduction
Heterotopic ossification (HO) is an ectopic formation of bone in soft tissue or joints, which impedes physical function and activities, leading to chronic pain, joint stiffness, pressure ulcers, venous thrombosis, or other health complications. There are two forms of HO: genetic and acquired. Acquired HO is a kind of common clinical complication after traumatic orthopedics, deep burns, and central nervous system injury. THO primarily occurs after skeletal muscle trauma or fracture, while neurogenic heterotopic ossification often follows central nervous system injuries (such as spinal cord injuries). The two exhibit significant differences in terms of etiological factors, lesion sites, and pathological processes. It has a high rate of recurrence and complications during treatment, especially invasive procedures, which is one difficult problem to be solved clinically.1,2
At present, there are various treatments for THO, containing surgery and conservative treatments, such as non-steroidal anti-inflammatory drugs (NSAIDs), radiotherapy and physical therapy.3 While THO typically occurs in soft tissue injuries with or without fractures, for HO associated with fractures, treating the HO can easily affect fracture healing. Therefore, the non-surgical management has some disadvantages, which contain delayed union, nonunion, cardiovascular risk, gastrointestinal side effects,4,5 and periarticular fibrosis. Among of them, delayed union and nonunion are the most common complications after conservative treatment of THO accompanied by fractures.6,7 Until now, despite the risks of recurrence and re-fracture, surgical resection remains the definitive management. Therefore, finding a new therapeutic method with fewer side effects is crucial.
Currently, the pathological mechanisms of THO are not clear. However, fibrodysplasia ossificans progressiva (FOP), which is driven by mutations in the ACVR1 gene, went through inflammation-mediated mesenchymal stem cell osteogenic differentiation abnormalities as the core pathological mechanism. Previous lineage tracing studies have confirmed that the ectopic bones in FOP originate from mesenchymal stem cells, and the abnormal activation of related signaling pathways (such as the TGF-β/Smad pathway) plays a pivotal role.8 Most studies suggest that post-traumatic local tissue injury can trigger a persistent inflammatory response. The inflammatory microenvironment, by releasing cytokines such as IL-6 and TNF-α, recruits BMSCs and vascular cells to the injury site. Simultaneously, it activates osteogenic signaling pathways, ultimately promoting ectopic bone formation. Therefore, inflammation is a driving factor in the occurrence and development of THO.9,10 Among the progress, MSCs’ migration and osteogenic differentiation play an essential role in THO.11 Consequently, blocking migration and osteogenic differentiation of MSCs could be a viable treatment for HO.
Some animal experiments have found that signaling pathways such as TGF-β/Smad, NF-κB and Wnt/β-catenin, as well as inflammatory cytokine networks, play a crucial role in THO pathogenesis.12,13 Poly ADP ribose polymerase 1 (PARP1), as a transcriptional co-regulator, can interact with the above pathway like TGF-β/Smad and NF-κB in some diseases.14,15 Recently, PARP inhibitors have garnered considerable interest from researchers and clinicians, since its approval by the Food and Drug Administration (FDA) for use against cancer. In addition to cancer, PARP1 is also associated with other diseases, including inflammation, ischemia, and vascular calcification.16–19 PJ34, as a classic PARP1 inhibitor, was shown to attenuate diabetic atherosclerotic calcification in vivo by downregulating PARP1/STAT1-mediated vascular smooth muscle cell phenotype switching and macrophage polarization.18 Furthermore, related studies have identified an inhibitory effect of PJ34 on MSCs’ differentiation,19 speculating the possibility of PARP1 as a potential target for THO prevention. This study aimed to explore the effect of PJ34 on THO through in vitro and in vivo experiments and to evaluate its impact on normal bone healing, providing a potential therapeutic option for THO prevention.
Materials and Methods
Materials
Cell culture reagents, including fetal bovine serum (FBS), minimum essential medium α (α-MEM), penicillin/streptomycin and trypsin were purchased from Gibco (Grand Island, NY, USA). Alizarin red S solution (1%, pH 4.2), ascorbic acid, dexamethasone and β-glycerophosphate disodium were purchased from Sigma (St. Louis, MO, USA). PJ34 were purchased from MedChemExpress (MCE, USA). Alkaline phosphatase assay kit, 4’, 6’-diamidino-2-phenylindole hydrochloride (DAPI), and BCA protein assay kit were purchased from Beyotime (Shanghai, China). Phalloidin, cell counting kit-8 (CCK-8), live/dead staining kit and the primary antibodies of GAPDH, Col III, Col I, PARP1, OPN and Runx2 were acquired from Proteintech (Wuhan, China).
Isolation and Culture of Rat Bone Marrow MSCs
The animal care committee of Shandong Provincial Hospital affiliated to Shandong First Medical University approved the animal experimental procedures. Two 3-week-old male Sprague Dawley (SD) rats were sacrificed and then sterilized by 75% alcohol for 10 minutes. The femur and tibia were peeled under aseptic condition, the surrounding tissues and muscles were removed, then, the ends were trimmed and the bone marrow cavity was repeatedly rinsed with α-MEM (with 1% penicillin/streptomycin) medium. The cell suspension was transferred to a centrifuge tube and spun for 5 minutes at 100×g. After the supernatant was discarded, the cells were resuspended in α-MEM (containing 1% penicillin/streptomycin and 15% FBS) media before being inoculated into a culture dish and cultured at 37°C in 5% CO2. The cells were trypsinized, centrifuged, and passaged once they had reached 80% confluence.
Cytotoxicity/Cell Viability Assay
Acute cytotoxicity and cell viability following PJ34 treatment was assessed using CCK-8. BMSCs were first inoculated onto 96-well plates. After being cultured for 24 h, the cells were treated with different concentrations of PJ34 (0.0, 1.0, 2.5, 5.0, 10.0, 20.0 μM) for various durations (0.5, 1, 4, 7 days). After the addition of 10 μL CCK-8 solution, the cells were incubated in incubator for 1 h, the optical density (OD) value was measured by spectrophotometry (Multiskan GO 1510, ThermoFisher Scientific, USA) at 450 nm. The visualization of cell viability is displayed with live-dead cell staining. The cells were cleaned with PBS three times when the incubation days reached 1, 4, and 7 days. The medium in the well plate was then removed. After that, 200 µL of living-dead cell dye were arranged in each well. The dye was rinsed twice with PBS after being incubated for 45 min in the dark. A fluorescent microscope (Axio Observer A1, Carl Zeiss, Germany) took pictures of samples to observe the viability of the cells. To determine how PJ34 affects cell shape, the cytoskeleton and nuclei were dyed. BMSCs were inoculated on 24-well plates and treated with various dosages of PJ34. When the incubation time reached 1, 4, and 7 days, the cells were fixed with 4% paraformaldehyde (PFA) for 15 min, permeabilized with 0.5% Triton X-100, and then stained with phalloidin and DAPI to highlight the cytoskeleton and nucleus, respectively. The images of the samples were obtained by fluorescence microscope.
Transwell Migration Assay
To verify if PJ34 affects cell recruitment, an in vitro Transwell migration assay was performed using a polycarbonate membrane (Corning, USA) with pore size of 8 μm. Firstly, 1 mL of serum-free medium containing the corresponding concentrations of PJ34 (0.0, 1.0, 2.5, 5.0 μM) was added to the bottom plate. The inserts were then filled with 200 μL of BMSC suspension (1×105 cells/mL) on the cell plate and cultured for 24 h in the incubator. The inserts were washed with PBS three times before the cells were fixed with 100% methanol for 20 min on ice. Then, 500 μL of the crystal violet solution was added to each insert and well, followed by incubation for 15 min. The upper part of the membrane was gently cleaned with a cotton-tipped swab after the crystal violet had been removed. An optical microscope (Eclipse TS100, Nikon, Japan) was used to observe cells. Experiments were carried out five times, the number of cells entering and exiting the chamber was measured for statistical analysis in order to measure cell migration.
Scratch Wound Healing Assay
To assess PJ34’s effect on cell migration in vitro, an experiment of scratch wound healing was performed. A confluent monolayer of human umbilical vein endothelial cells (HUVECs) (2×104 cells/cm2) was formed after being cultured for up to 24 h on 24-well plates. The monolayer was then gently scratched with the same 200 μL pipette tip, and they were then gently rinsed twice with phosphate-buffered saline (PBS). After that, different concentrations of PJ34 (0.0, 1.0, 2.5, 5.0 μM) were added to low-serum culture medium (2.0 mL) for each group. To quantify the size of the original scratch region (B0) and unhealed scratch area (B1), images of the migrating cells at 0 and 24 h of culture were evaluated by Image J software. An equation (1) below was used to calculate the wound closure:
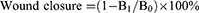
Alkaline Phosphatase (ALP) Activity and Alizarin Red S (ARS) Staining
BMSCs were seeded with a density of 4.5×103 cells/cm2 in 12-well plates and cultivated in α-MEM (with 1% antibiotics and 10% FBS) for 24 h until cells achieved 80% confluence. For osteogenesis, each well added 1 mL of osteogenic medium which contains dexamethasone (10−7M), β-glycerophosphate (10 mM), and vitamin C (50 µg/mL), PJ34 was diluted to concentrations of 0.0, 1.0, 2.5, 5.0 μM and added to the inducing media, which was replaced every 3 days. PBS washed each well three times and 4% PFA fixed cells for 15 min after the stimulation of osteogenic differentiation for 7 days. After washing, ALP activity was measured semi-quantitatively using p-nitrophenyl phosphate as a substrate after cells were stained by a 5-bromo-4-chloro-3-indolyl phosphate/nitro-blue tetrazolium (BCIP/NBT) ALP color development kit following the manufacturer’s instructions. PBS washed the cells thrice after 14 days of osteogenic induction, 4% PFA fixed the cells for 15 min, and then ARS stained for 5 min. At 37 °C, 100 μL of 10% acetylpyridine chloride were added to each well. After 30 min of complete dissolution, the samples’ absorbance at 562 nm was measured.
Quantitative Real-Time Polymerase Chain Reaction (PCR)
Cells were collected for extraction of RNA, followed by quantitative reverse transcription polymerase chain reaction. SteadyPure Universal RNA Extraction Kit (Accurate Biotechnology, Hunan, China) was used to extract total RNA. The Evo M-MLV RT Premix (Accurate Biotechnology, Hunan, China) for qPCR was used to make the cDNA. The SYBR Green Premix Pro Taq HS qPCR Kit (Accurate Biotechnology, Hunan, China) was used for the qRT-PCR following the manufacturer’s instructions. All of primer sequences ultimately that we employed can be found in Table 1, and the QuantStudio 3 was utilized for the PCR amplification.
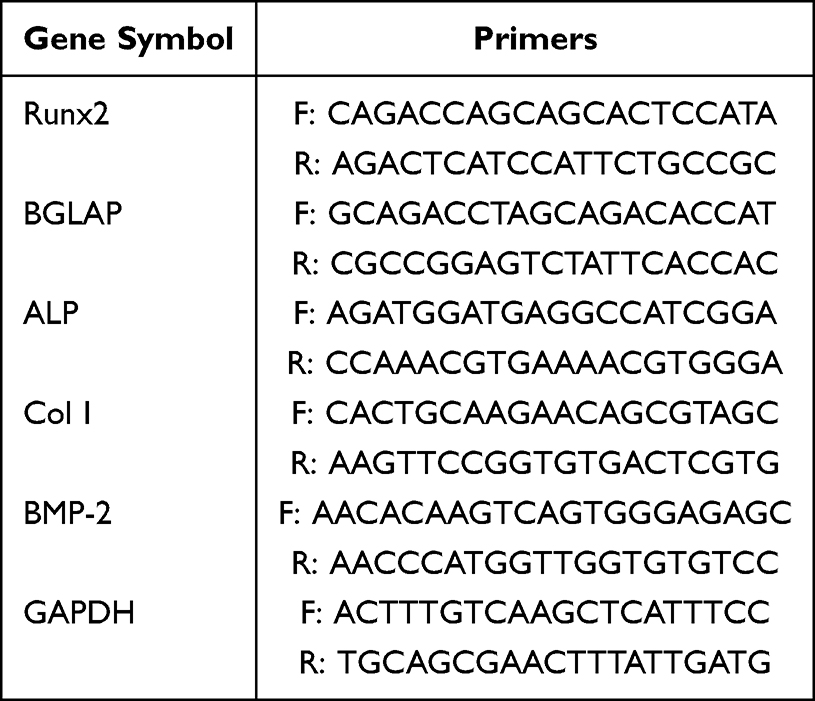 |
Table 1 Primer Sequences Used for qRT-PCR |
Western Blot
BMSCs were seeded in 6-well plates and cultivated in α-MEM (with 1% antibiotics and 10% FBS) to 70% confluency before being treated with various doses of PJ34 as previously mentioned. After 3 days, BMSCs were lysed with RIPA buffer containing protease and phosphatase inhibitors, and each sample was centrifuged at 1.2×104×g, 4°C for 20 min, and a bicinchoninic acid assay (BCA) protein quantification kit was used to determine the protein content. Thirty micrograms protein was added into each well of 10% SDS-PAGE gel (Epizyme, Shanghai, China) and electrophoretically separated. The bands were separated and transferred to a polyvinylidene fluoride (PVDF) membrane (Millipore, Massachusetts, USA). After incubating the membrane with primary antibodies of Col I, Col III, OPN, Runx2, PARP1, and β-actin overnight at 4°C, it was blocked with blocking buffer for 1 h. The membrane was incubated for 2 h with a secondary antibody (1:5000) on rotary shaker after being washed thrice with PBST. Amersham Imager 600 (GE, Boston, MA, USA) was utilized to observe the particular bands. Each sample was repeated five times, and Image J software was used to quantitatively assess each Western blot band.
Animal Experimental Model
Animal experiments were conducted in accordance with animal care guidelines of the institutional animal care and use committee (IACUC) of Shandong Provincial Hospital Affiliated to Shandong First Medical University. All protocols were approved by shandong provincial hospital IACUC (reference number: 2019–008). All methods are reported in accordance with ARRIVE (Animal Research: Reporting of In Vivo Experiments) guidelines. Pengyue experimental animal company (Jinan, China) provided 8-week-old male C57 mice for this experiment. Transection of the Achilles tendon combined with burn injury was used to simulate musculoskeletal trauma in a mouse model. The mice were anesthetized by intraperitoneal injection of ketamine (80 mg/kg) at first, then received a 30% partial-thickness burn on the shaved dorsum using a metal block heated to 70°C for 20s continuously. Secondly, a small incision was performed at the distal achilles tendon of the gastrocnemius muscle to find the achilles tendon and midpoint tenotomy was made, then the wound was sutured with 5–0 vicryl stitch. After modeling, the mice were separated into two groups randomly: PJ34 treatment group (n = 6) and control group (n = 6). Previous studies have demonstrated that PJ34 (10 mg/kg) administration in animal model has an effective therapeutic effect without obvious toxic and side effects.18,20,21 So, during eight weeks, in the PJ34 treatment group, we dissolved PJ34 (10 mg/kg) in 100 µL normal saline, and then gavaged the mice every other day. The mice in the control group received 100 µL normal saline every other day. After 8 weeks, these mice were sacrificed for micro-CT analysis and pathological observation.
In order to determine whether PJ34 affects normal fracture healing, a mouse tibial fracture model was subsequently developed. The mice were anesthetized by intraperitoneal injection of ketamine, placed supinely on the operating table and disinfected on their knees. Through an incision close to the patella, the tibial tubercle was exposed, and a 0.8 mm intramedullary needle was inserted. After the muscle has been stripped away, the tibia was cut in the middle part with tiny scissors, and the surgical wound was cleansed with saline before being stitched with 5–0 vicryl suture. Following the operation, the mice were divided into two groups randomly: the control group (n = 20) and the PJ34 treatment group (n = 20). The mice in the PJ34 treatment group received 100 µL normal saline containing PJ34 (10mg/kg) by oral administration every other day. The control group received the same equivalent normal saline. For micro-CT analysis and HE stain, the mice were sacrificed at 1, 2, 3, and 4 weeks, respectively.
Micro-CT and Histological Analysis
At the scheduled time point, mice were sacrificed by CO2 inhalation. The ankle together with the achilles tendons and tibia were fixed in 10% formalin for scanning by high-resolution micro-CT (viva CT80, SCANCO Medical AG, Switzerland). The 3D images were reconstructed and microstructural characteristics were employed to analyze the bone volume (BV) of HO and bone callus. Then, these samples were decalcified in 10% EDTA solution for 1 month, following by dehydration, paraffin embedding, and then slices were prepared for HE staining and observed by a bright field scanning microscope (TissueGnostics, Austria).
Statistical Analysis
Quantitative data were expressed as mean ± standard deviation (SD) and were collected from at least five different samples. Prior to statistical comparisons, data distribution was assessed using the Shapiro–Wilk test to verify normality. For normally distributed data, two-tailed Student’s t-test was used for comparisons between two groups, while one-way analysis of variance (ANOVA) followed by Tukey’s post-hoc test was applied for multiple group comparisons. Non-normally distributed data (if any) would be analyzed using non-parametric tests (eg, Kruskal–Wallis test). A P-value < 0.05 was considered statistically significant. All statistical analyses were performed using GraphPad Prism 9.0 software (GraphPad Software, Inc., La Jolla, CA, USA).
Results
PJ34 Reduced Proliferation of BMSCs
According to the results of acute cytotoxicity test, over 10 μM PJ34 has acute toxicity on cell growth, so we employed concentrations of PJ34 (0, 1.0, 2.5, and 5.0 μM) in the subsequent studies (Figure 1a). The CCK-8 assay showed that PJ34 reduced proliferation and viability of BMSCs in a dose- and time-dependent manner (Figure 1b). The cytoplasm-damaged cells are brightly stained with red by the live/dead reagent, whereas the healthy cells are brightly stained with green (Figure 1c). The outcomes demonstrated that the cells could increase along with the time at the same concentration. However, at the same time point, the number of dead cells increased with the increase of drug concentration. After the treatment of different concentration of PJ34, the average area of BMSCs was 2146.04 ± 333.91 μm2, 2560.12 ± 202.29 μm2, 2537.73 ± 291.47 μm2, 2293.47 ± 298.07 μm2 at day 1, 2283.67 ± 232.75 μm2, 2640.52 ± 366.42 μm2, 2459.02 ± 276.46 μm2, 2231.01 ± 188.9 μm2 at day 4, and 2420.92 ± 309.93 μm2, 2575.26 ± 174.43 μm2, 2411.12 ± 291.73 μm2, 2315.33 ± 304.51 μm2 at day 7 for PJ34 of 0 μΜ, 1.0 μΜ, 2.5 μΜ, and 5.0 μΜ, respectively, there was no significant change in cell shape, which was formed by the cytoskeleton (Figure 1d, S1). These results indicated that PJ34 may inhibit cell proliferation but did not change cell morphology.
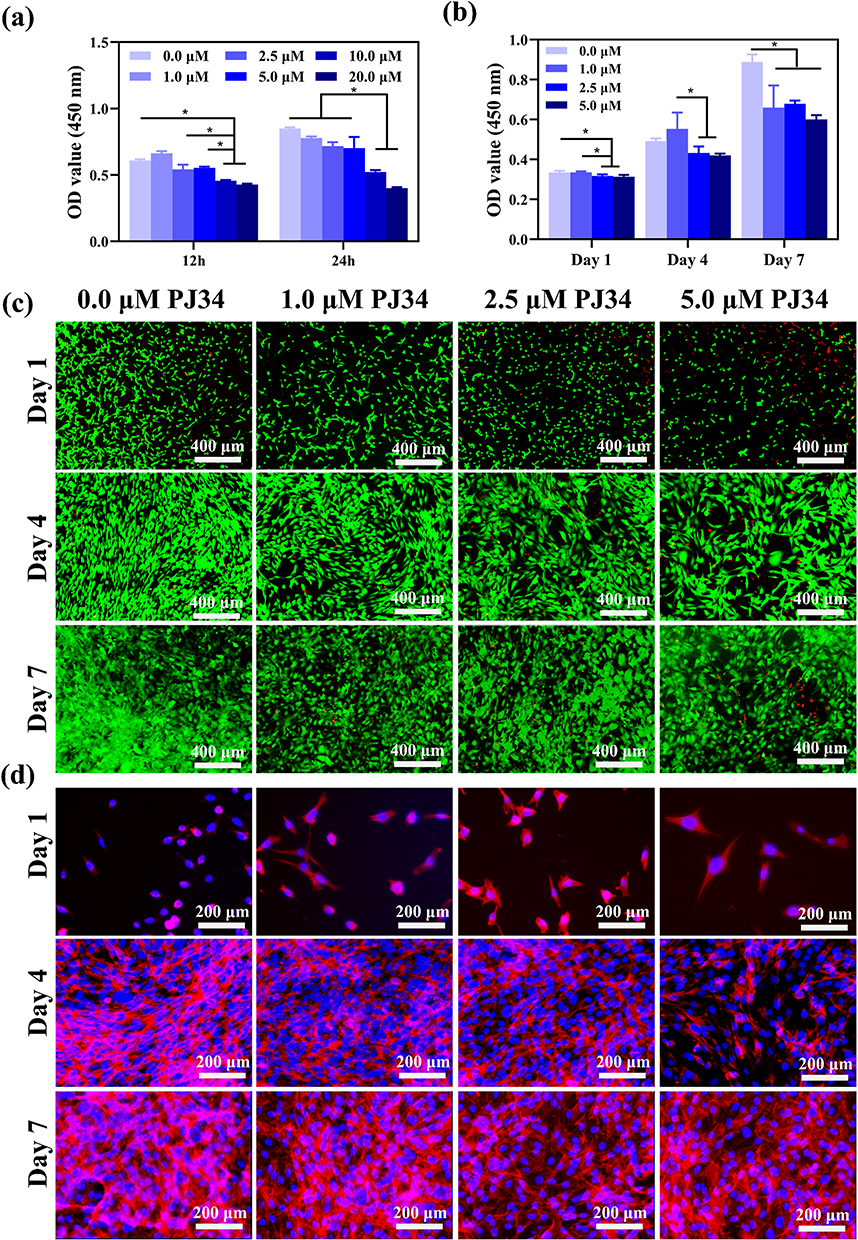 |
Figure 1 Cytotoxicity and cell viability with PJ34 in vitro. The effect of PJ34 on BMSCs cytotoxicity at 12 and 24 h (a). The effect of PJ34 on BMSCs proliferation at 1, 4, 7 days (b). Live (green) /dead (red) staining of BMSCs at 1, 4, 7 days (c). Cell morphology of BMSCs at 1, 4, 7 days (d). The cytoplasm of BMSCs was stained with phalloidin (d, red), and cell nucleus were stained with DAPI (d, blue) to further observe the cell morphology. n=5 per group. *p < 0.05. |
PJ34 Inhibited Migration of BMSCs and HUVECs
Cell recruitment is an important process in the formation of HO. Cell migration of HUVECs was observed by the wound scratch test, and it was slower in the PJ34 group than in the control group (Figure 2a). An in vitro transwell experiment was also used to confirm the capability of BMSC migration. The BMSCs that crossed the membrane exhibited a considerable, dose-dependent reduction after PJ34 treatment (Figure 2b). After 24h of treatment, the percentage of wound closure was 58.2 ± 11.8%, 40.2 ± 3.8%, 33.4 ± 3.1% for PJ34 of 1.0 μΜ, 2.5 μΜ, 5.0 μΜ, respectively, which were significantly less than the control group (83.5 ± 5.9%), compared to the wound size at day 0 (Figure 2c). The number of BMSCs passing through the chamber was 62 ± 4, 33 ± 4, 14 ± 4, 10 ± 2 for PJ34 of 0 μΜ, 1.0 μΜ, 2.5 μΜ, 5.0 μΜ, respectively (Figure 2d). Therefore, our findings revealed that PJ34 considerably inhibited cell migration.
 |
Figure 2 Biological function of PJ34 in vitro. Migration of HUVECs in a scratch wound healing assay in vitro at 0 and 24 h after treated with PJ34 (a). Transwell migration assay of BMSCs (b). Percentage of wound closure determined by the microscopic photographs (c). Quantitative analysis of the number of BMSC cells passing through the chamber (per field) (d). n=5 per group. *p < 0.05. |
PJ34 Inhibited Osteogenic Differentiation of BMSCs in vitro
For the healing of bone, osteogenic differentiation is essential. The stage of osteogenic differentiation contains early stage and late stage. The early osteogenic effects of PJ34 were assessed by measuring ALP activity and observing calcium deposition both quantitatively and qualitatively. Figure 3 depicts the ALP activity of BMSCs cultured in osteogenic medium at different concentrations of PJ34. It’s worth noting that ALP staining was less in PJ34 treatment group on days 7 and 14 when compared to the control group (Figure 3a). ARS staining which indicates the late stage was carried out at 14 and 21 days to determine the mineral deposition of BMSCs following treatment with PJ34 (Figure 3b). The ALP activity in the PJ34 group was substantially lower than that in the control group, according to the quantitative results (Figure 3c). Due to the osteo-inductive media incubation, optical and microscopic images revealed that mineralized nodules were generated in each group; however, mineralization was dramatically reduced in the PJ34-treated group (Figure 3d). Therefore, it can be concluded that PJ34 may significantly inhibit all stages of osteogenic differentiation in BMSCs.
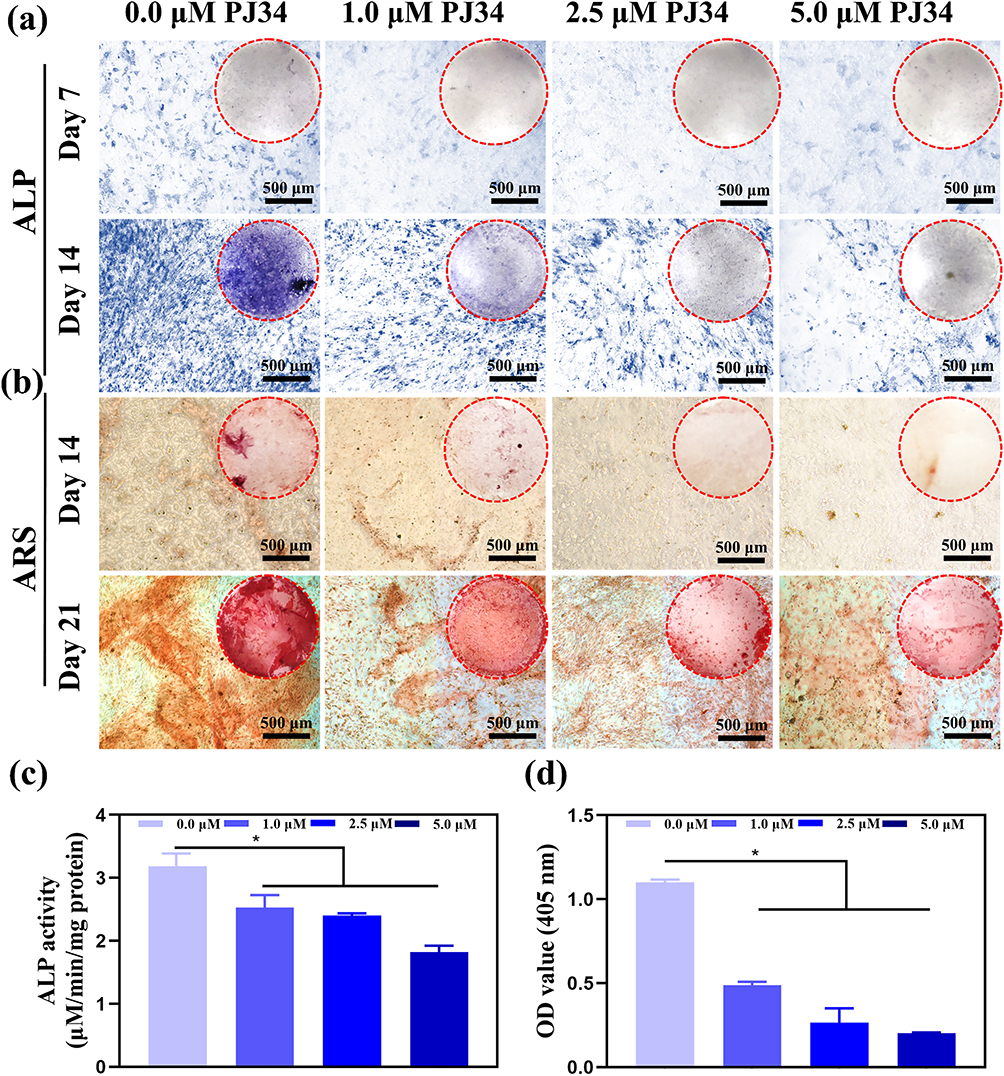 |
Figure 3 Osteogenesis-inducing capacity of PJ34 in vitro. The ALP activity of BMSCs for 7 and 14 days (a). ARS staining of BMSCs for 14 and 21 days (b). Quantitative analysis of the ALP activity of BMSCs (c). Quantitative analysis of the OD value for ARS staining of BMSCs (d). Dotted circles represent the gross images of each well in 12-well plate. n=5 per group. *p < 0.05. |
The expression of osteogenic genes indicates that PJ34 has an effect on osteogenic differentiation. In this work, qRT-PCR analysis revealed that PJ34 reduced the expression of osteogenic genes like Col I, Runx2, BMP-2, ALP, and BGLAP in BMSCs, especially when the concentration of PJ34 is up to 5.0 μΜ (Figure 4a–e). In addition, when the inhibitory action of PJ34 on protein of PARP1 was confirmed (Figure 4f and g), the expression of osteogenic proteins such as Col III, Col I, OPN, and Runx2 in BMSCs was also decreased with a dose-dependent manner of PJ34 (Figure 4h–l). All these results indicated that PJ34 affected the osteogenic ability of BMSCs, which implied a new approach to treating HO.
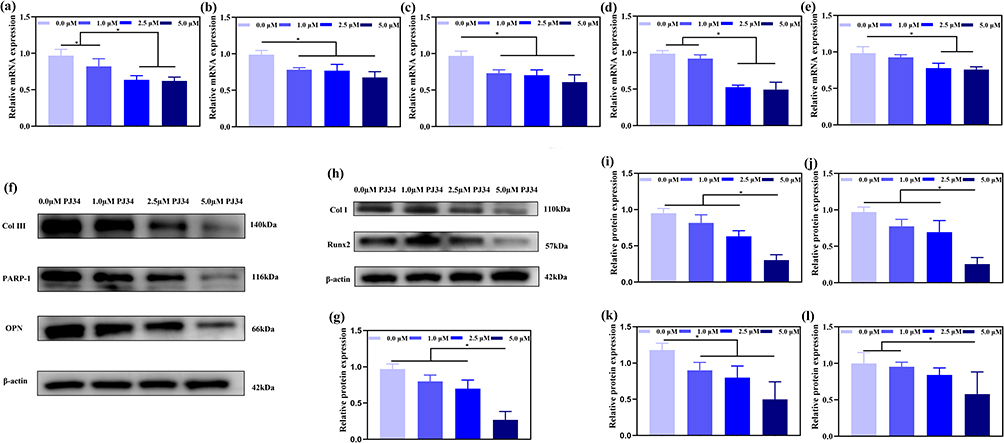 |
Figure 4 Evaluation of osteogenic potential of PJ34 under different concentrations using qRT-PCR and Western blot analysis in vitro. Effect of the osteogenic medium with PJ34 on the osteogenic gene expression in BMSCs at 3 days, including Col I (a), Runx2 (b), BMP2 (c), ALP (d), BGLAP (e). Verification of PARP-1 inhibition by PJ34 (f and g). Effect of the osteogenic medium with PJ34 on the osteogenic protein expression in BMSCs at 3 days (f–h), including Col III (i), Col I (j), OPN (k), Runx2 (l). n=5 per group. *p < 0.05. |
PJ34 Inhibited HO Formation without Impairing Bone Healing in vivo
To determine whether PJ34 inhibits HO in vivo, we tested a typical mouse model that underwent tenotomy and back burn. In comparison to control group, PJ34-treated animals had less ectopic ossification tissues at the surgical site close to the tendon rupture by micro-CT evaluation (Figure 5a and b). The quantitative analysis revealed the BV value of HO in the control group (1.19 ± 0.72 mm3) was significantly higher than that in the PJ34-treated group (0.56 ± 0.16 mm3) at 8 weeks (n = 6 per group) (Figure 5c). Further evidence that PJ34 inhibits heterotopic bone formation comes from histological examination by HE staining (Figure 5d), which revealed that HO masses in control mice contained significant amounts of mature bone, but it was significantly less or even did not form in the PJ34-treated mice.
 |
Figure 5 Evaluation inhibition of HO by PJ34 in vivo. Micro-CT of HO in control groups (a) and PJ34 groups (b) at 8 weeks. Quantitative analysis of BV at 8 weeks (c). Representative photographs of HE-stained section of control and PJ34 groups at 8 weeks (d). Red dotted squares indicating distal area of Achilles tendon transection, yellow dotted squares indicating proximal area of Achilles tendon transection. Red arrows indicating HO. n=6 per group. *p < 0.05. |
However, in our clinical cases, we discovered that fracture and HO frequently happened together. Therefore, we used a tibial fracture model to investigate whether PJ34 might affect fracture healing (Figure 6a). The extent of fracture healing was observed under micro-CT scanning each week and the results indicated that the trend of bone union between control group and PJ34-treated group was similar at each time point (Figure 6b and c), and there was no significant difference in the quantitative result of BV between the control group and PJ34 treated group in the whole process (Figure 6d–g). Moreover, the results of HE staining showed the average area of new bone tissues, which contain bone trabeculae and lamellar bone formed at the fracture site in control group and PJ34 treated group, was 1.63 ± 0.21 mm2, 1.63 ± 0.32 mm2 at first week, 1.77 ± 0.34 mm2, 1.54 ± 0.23 mm2 at second week, 1.99 ± 0.34 mm2, 2.13 ± 0.31 mm2 at third week, 2.08 ± 0.24 mm2, 2.08 ± 0.19 mm2 at fourth week, respectively. There is no significant difference between each group (Figure 7a and b, Figure S2).
 |
Figure 6 Effect of PJ34 on fracture healing. Photographs showing tibia fracture process in a mouse model (a). Micro-CT and X-ray showing healing process of tibia fracture in control group (b) and PJ34 group (c) at 1, 2, 3, and 4 weeks. BV analysis determined by micro-CT at 1, 2, 3, and 4 weeks (d–g). n=5 per time. |
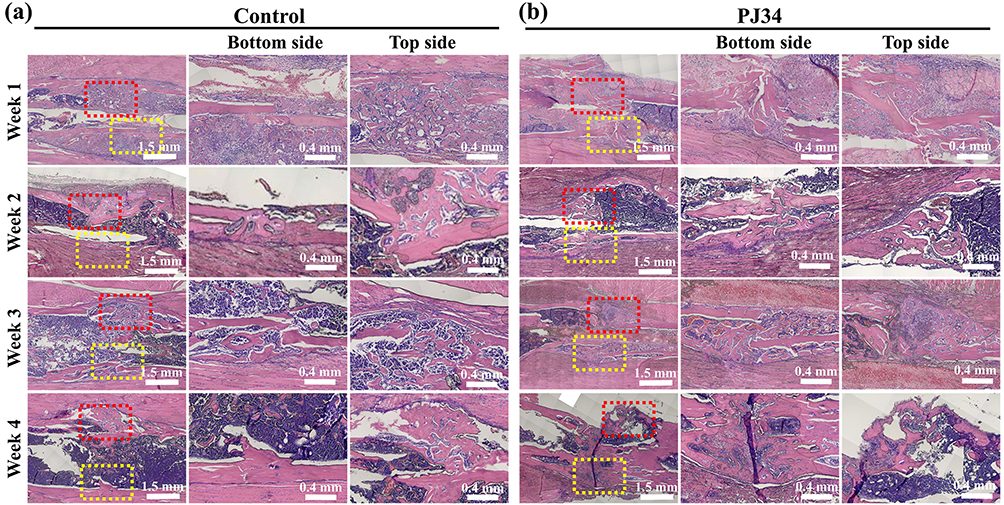 |
Figure 7 Effect of PJ34 on fracture healing. Histological observation of tibia fracture healing process by HE staining in control group (a) and PJ34 group (b) at 1, 2, 3, and 4 weeks. Red dotted squares indicating top side of tibial fracture healing area, yellow dotted squares indicating bottom side of tibial fracture healing area. n=5 per time. |
Discussion
Our in vitro results clearly show that PJ34 dose-dependently inhibits key processes in THO pathogenesis, including BMSC proliferation, migration, and osteogenic differentiation at both the early and late stages of osteogenesis. These findings align with previous reports19 indicating that suppressing BMSC activity can reduce ectopic ossification. Specifically, PJ34 reduced ALP activity, mineralization, and the expression of osteogenic markers, such as Col I, Runx2, BMP-2, ALP and BGLAP, confirming its ability to hinder osteogenic differentiation of BMSCs.
The observed effects of PJ34 are consistent with its known role as a PARP1 inhibitor, and previous studies have linked PARP1 to osteogenic differentiation regulation.18 The majority of recent research makes the assumption that the TGF-β/Smad pathway mediates the recruitment of BMSCs and osteogenic differentiation during bone formation, and that inhibiting the TGF-β/Smad pathway can lessen the osteogenic effect. Meanwhile, it was found that PARP-1 interacted with Smad, PARP1 could regulate the ADP-ribosylation of Smad3 and the flow of Smad signals,22 while inhibiting PARP1 can lower TGF-β expression and disrupt the TGF-β/Smad transduction pathway.23 Thereby, we infer that the mechanism of PJ34 inhibiting ectopic osteogenesis may be that, after PJ34 inhibition of PARP-1, the TGF-β/Smad signaling pathway in BMSCs was blocked, which in turn inhibited osteogenic differentiation. Currently, IL-11 inhibition is considered as a potential THO therapy.24,25 This raises the question of whether PARP1 influences IL-11 signaling, forming a functional crosstalk that regulates THO progression. Though direct evidence for this interaction is lacking, PARP1 is a known regulator of NF-κB signaling,26 and NF-κB can be regulated by IL-11.27 By inhibiting PARP1, PJ34 may downregulate NF-κB activation, thereby reducing IL-11 expression in trauma-induced inflammatory microenvironments. Clarifying whether PARP1 inhibition suppresses IL-11 expression or signaling will not only deepen our understanding of PJ34’s mechanism of action but also open avenues for combinatorial therapies targeting both pathways to enhance THO prevention. However, the specific molecular pathways through which PJ34 exerts these effects require further investigation. It is worth noting that our findings do not rely on defining this mechanism; rather, they demonstrate a clear phenotypic effect of PJ34 on THO prevention and bone healing safety.
In in vivo experiments, PJ34 significantly reduced heterotopic bone volume in a mouse THO model, as evidenced by micro-CT and histological analysis. Critically, unlike conventional treatments,6,28,29 PJ34 did not compromise fracture healing, as shown by comparable bone callus formation, bone volume, and histological features between PJ34-treated and control groups throughout the healing process. This is a key advantage, as preserving normal bone repair is essential for clinical applicability. Compared to existing therapies, PJ34’s ability to target THO without disrupting normal bone healing addresses a critical unmet need. This seemingly contradictory mechanism may be explained by the following ways. 1) BMSCs involved in THO are recruited to traumatic soft tissue microenvironments characterized by persistent inflammation, high cytokine levels (eg, IL-6, TNF-α), and pathological remodeling. In contrast, fracture healing occurs in a physiological bone repair niche with spatiotemporally regulated cytokine gradients (eg, BMP-2, PDGF) and mechanical cues. PJ34 may preferentially target BMSCs in pathological inflammatory microenvironments, where PARP1 is abnormally activated, while having minimal impact on BMSCs in the physiological fracture healing niche. This selectivity is supported by previous findings that PJ34 induces cycle arrest in pathological cells (eg, cancer cells) without affecting normal cell proliferation.30 2) In vivo fracture healing involves multiple cell types (osteoblasts, chondrocytes, periosteal cells) and redundant regulatory pathways (eg, Wnt/β-catenin, IGF-1) that may compensate for PARP1 inhibition. Periosteal cells, a key contributor to fracture callus formation, may be less sensitive to PJ34 than BMSCs, ensuring unimpaired bone repair. 3) PJ34’s bioavailability, tissue distribution, and metabolism may result in local concentrations at fracture sites that are insufficient to inhibit physiological osteogenesis, while reaching effective levels in soft tissue trauma sites to suppress THO. It also implies that ectopic ossification is a pathological process of hyperproliferation, whereas fracture healing is a normal physiological process. Healthy osteoblasts play a predominant role in the whole process, so that PJ34 may not negatively affect the physiological processes of healthy cells. PJ34’s favorable profile in our models suggests it could be a safer alternative, though further studies in larger animal models and clinical trials are needed to confirm this.
The preliminary results of this study, in sum, demonstrated that the PARP1 inhibitor PJ34 prevented BMSCs from osteogenic differentiation and the formation of HO, without impairing fracture healing, which fully demonstrates the potential of PJ34 on the prevention of THO in clinical practice. However, there are certain limitations on this study, including the lack of mechanistic validation and the use of a single mouse model, which may restrict generalizability. Future research should focus on elucidating the molecular targets of PJ34 in BMSCs and evaluating its efficacy in diverse trauma models.
Conclusions
In summary, our study provides evidence that PJ34 prevents THO by inhibiting BMSC proliferation, migration, and osteogenic differentiation, without adverse effects on normal bone healing. This supports PJ34’s potential as a novel therapeutic agent for THO prevention, offering a safer alternative to current treatments.
We explicitly acknowledge the limitations of this study: 1) The tibial fracture model and the THO model were validated separately, thus failing to replicate the clinical scenario where fractures and THO co-occur; 2) Quantitative assessments of callus composition and biomechanical strength were not performed, which would further strengthen the conclusion that fracture healing is unimpaired; 3) The expression pattern of PARP1 during THO formation and its potential signaling pathway, like TGF-β/Smad or IL-11, remain uninvestigated; 4) The study relies on a single mouse model, which may limit the generalizability of the findings to other trauma settings or species.
To advance PJ34 toward pre-clinical testing, first, mechanistic validation will be conducted to elucidate the precise molecular pathways underlying PJ34’s effects, focusing on its interaction with the TGF-β/Smad signaling pathway and potential crosstalk with IL-11 signaling. Second, the efficacy of PJ34 will be evaluated in clinically relevant large animal models of THO (such as rabbits and sheep), while simulating the combination of fractures and severe soft tissue trauma, which is a common scenario in clinical practice but not fully covered by current mouse models. Thirdly, we will optimize the drug administration regimen and route to improve bioavailability at the trauma site while minimizing systemic exposure. Fourthly, long-term safety also needs further evaluation to monitor potential off-target effects, including changes in normal bone metabolism and organ function. Overall, these efforts will provide crucial data support for the development of PJ34 as a targeted therapeutic agent for THO, thus meeting the urgent clinical need for safe and effective prevention of THO.
Funding
This work was supported by Taishan Scholars Program of Shandong Province (tsqn201812141).
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Pavey GJ, Polfer EM, Nappo KE, Tintle SM, Forsberg JA, Potter BK. What risk factors predict recurrence of heterotopic ossification after excision in combat-related amputations? Clin Orthopaed Related Res. 2015;473(9):2814–14. doi:10.1007/s11999-015-4266-1
2. Xu R, Hu J, Zhou X, Yang Y. Heterotopic ossification: mechanistic insights and clinical challenges. Bone. 2018;109:134–142. doi:10.1016/j.bone.2017.08.025
3. Shapira J, Yelton MJ, Chen JW, et al. Efficacy of NSAIDs versus radiotherapy for heterotopic ossification prophylaxis following total Hip arthroplasty in high-risk patients: a systematic review and meta-analysis. HIP Int. 2022;32(5):576–590. doi:10.1177/1120700021991115
4. Sagi HC, Jordan CJ, Barei DP, Serrano-Riera R, Steverson B. Indomethacin prophylaxis for heterotopic ossification after acetabular fracture surgery increases the risk for nonunion of the posterior wall. J Orthop Trauma. 2014;28(7):377–383. doi:10.1097/BOT.0000000000000049
5. Leese PT, Hubbard RC, Karim A, Isakson PC, Yu SS, Geis GS. Effects of celecoxib, a novel cyclooxygenase-2 inhibitor, on platelet function in healthy adults: a randomized, controlled trial. J Clin Pharmacol. 2000;40(2):124–132. doi:10.1177/00912700022008766
6. Hamid N, Ashraf N, Bosse MJ, et al. Radiation therapy for heterotopic ossification prophylaxis acutely after elbow trauma: a prospective randomized study. J Bone Joint Surg Am. 2010;92(11):2032–2038. doi:10.2106/JBJS.I.01435
7. Balboni TA, Gobezie R, Mamon HJ. Heterotopic ossification: pathophysiology, clinical features, and the role of radiotherapy for prophylaxis. Int J Radiat Oncol Biol Phys. 2006;65(5):1289–1299. doi:10.1016/j.ijrobp.2006.03.053
8. Kaliya-Perumal A-K, Carney TJ, Ingham PW. Fibrodysplasia ossificans progressiva: current concepts from bench to bedside. Dis Model Mech. 2020;13(9). doi:10.1242/dmm.046441
9. Dey D, Wheatley BM, Cholok D, et al. The traumatic bone: trauma-induced heterotopic ossification. Transl Res. 2017;186:95–111. doi:10.1016/j.trsl.2017.06.004
10. Li J, Sun Z, Luo G, et al. Quercetin attenuates trauma-induced heterotopic ossification by tuning immune cell infiltration and related inflammatory insult. Front Immunol. 2021;12:649285. doi:10.3389/fimmu.2021.649285
11. Zhang Q, Zhou D, Wang H, Tan J. Heterotopic ossification of tendon and ligament. J Cell Mol Med. 2020;24(10):5428–5437. doi:10.1111/jcmm.15240
12. Wang X, Li F, Xie L, et al. Inhibition of overactive TGF-β attenuates progression of heterotopic ossification in mice. Nature Communications. 2018;9(1):551. doi:10.1038/s41467-018-02988-5
13. Liu F, Zhao Y, Pei Y, Lian F, Lin H. Role of the NF-kB signalling pathway in heterotopic ossification: biological and therapeutic significance. Cell Commun Signal. 2024;22(1):159. doi:10.1186/s12964-024-01533-w
14. Lu Y, Fu W, Xing W, Wu H, Zhang C, Xu D. Transcriptional regulation mechanism of PARP1 and its application in disease treatment. Epigenetics Chromatin. 2024;17(1):26. doi:10.1186/s13072-024-00550-w
15. Wang S, Wu Y, Zong W, Wang Z-Q. “Yin-Yang” of PARP1 in genotoxic and inflammatory response. DNA Repair. 2025;152:103858. doi:10.1016/j.dnarep.2025.103858
16. Ba X, Garg NJ. Signaling mechanism of poly(ADP-ribose) polymerase-1 (PARP-1) in inflammatory diseases. Am J Pathol. 2011;178(3):946–955. doi:10.1016/j.ajpath.2010.12.004
17. Strosznajder RP, Czubowicz K, Jesko H, Strosznajder JB. Poly(ADP-ribose) metabolism in brain and its role in ischemia pathology. Mol Neurobiol. 2010;41(2–3):187–196. doi:10.1007/s12035-010-8124-6
18. Li P, Wang Y, Liu X, et al. Loss of PARP-1 attenuates diabetic arteriosclerotic calcification via Stat1/Runx2 axis. Cell Death Disease. 2020;11(1):22. doi:10.1038/s41419-019-2215-8
19. Kishi Y, Fujihara H, Kawaguchi K, et al. PARP inhibitor PJ34 suppresses osteogenic differentiationin mouse mesenchymal stem cells by modulating BMP-2 signaling pathway. Int J Mol Sci. 2015;16(10):24820–24838. doi:10.3390/ijms161024820
20. Huang S, Zhang B, Chen Y, et al. Poly(ADP-ribose) polymerase inhibitor PJ34 attenuated hepatic triglyceride accumulation in alcoholic fatty liver disease in mice. J Pharmacol Exp Ther. 2018;364(3):452–461. doi:10.1124/jpet.117.243105
21. Bonnin P, Vitalis T, Schwendimann L, et al. Poly(ADP-ribose) polymerase inhibitor PJ34 reduces brain damage after stroke in the neonatal mouse brain. Curr Issues Mol Biol. 2021;43(1):301–312. doi:10.3390/cimb43010025
22. Dahl M, Maturi V, Lonn P, et al. Fine-tuning of smad protein function by poly (ADP-ribose) polymerases and poly(ADP-ribose) glycohydrolase during transforming growth factor β signaling. PLoS One. 2014;9(8):e103651. doi:10.1371/journal.pone.0103651
23. Lucarini L, Durante M, Lanzi C, et al. HYDAMTIQ, a selective PARP −1 inhibitor, improves bleomycin-induced lung fibrosis by dampening the TGF -β/ SMAD signalling pathway. J Cell Mol Med. 2017;21(2):324–335. doi:10.1111/jcmm.12967
24. Kaliya-Perumal A-K, Celik C, Carney TJ, Harris MP, Ingham PW. Genetic regulation of injury-induced heterotopic ossification in adult zebrafish. Dis Model Mech. 2024;17(5). doi:10.1242/dmm.050724
25. Dong B, Zhu J, Chen X, et al. The emerging role of interleukin-(IL)-11/IL-11R in bone metabolism and homeostasis: from cytokine to osteokine. Aging Dis. 2023;14(6):2113–2126. doi:10.14336/AD.2023.0306
26. Jin L, Chen J, Fu J, et al. PARP1 exacerbates prostatitis by promoting M1 macrophages polarization through NF-κB pathway. Inflammation. 2025;48(5):3022–3035. doi:10.1007/s10753-025-02247-y
27. Ng B, Widjaja AA, Viswanathan S, et al. Similarities and differences between IL11 and IL11RA1 knockout mice for lung fibro-inflammation, fertility and craniosynostosis. Sci Rep. 2021;11(1):14088. doi:10.1038/s41598-021-93623-9
28. Su B, O’Connor JP. NSAID therapy effects on healing of bone, tendon, and the enthesis. J Appl Physiol. 2013;115(6):892–899. doi:10.1152/japplphysiol.00053.2013
29. Wheatley BM, Nappo KE, Christensen DL, Holman AM, Brooks DI, Potter BK. Effect of NSAIDs on bone healing rates: a meta-analysis. J Am Acad Orthop Surg. 2019;27(7):e330–e336. doi:10.5435/JAAOS-D-17-00727
30. Cohen-Armon M. The modified phenanthridine PJ34 unveils an exclusive cell-death mechanism in human cancer cells. Cancers. 2020;12(6):1628. doi:10.3390/cancers12061628
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.