Back to Journals » Journal of Inflammation Research » Volume 16
PINK1/Parkin-Mediated Mitochondrial Autophagy Participates in H2O2-Induced Abnormal Proliferation of Fibroblast-Like Synoviocytes in Rheumatoid Arthritis
Authors Wang G, Chen X ![]() , Shao Y, Xu B
, Shao Y, Xu B ![]()
Received 29 November 2022
Accepted for publication 22 February 2023
Published 23 March 2023 Volume 2023:16 Pages 1271—1282
DOI https://doi.org/10.2147/JIR.S398690
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Ning Quan
Gaoyuan Wang,1 Xiaoyu Chen,2 Yubao Shao,2 Bin Xu1
1Department of Orthopaedics, the First Affiliated Hospital of Anhui Medical University, Hefei, Anhui, People’s Republic of China; 2Department of Histology and Embryology, Anhui Medical University, Hefei, Anhui, People’s Republic of China
Correspondence: Bin Xu, Email [email protected]
Introduction: To explore the role of PINK1/Parkin-mediated mitochondrial autophagy in H2O2-induced abnormal proliferation of rheumatoid arthritis fibroblast-like synoviocytes (RA-FLS).
Methods: Firstly, we isolated fibroblast like synoviocytes (RA-FLS) from RA patients. H2O2-induced oxidative stress, and NAC (a ROS inhibitor) or FCCP (a mitochondrial autophagy activator) treatment inhibited ROS level or activate mitochondrial autophagy in RA-FLS. MitoSOX Red, JC-1 kit, DCFH-DA kit and CCK8 kit were used to evaluate mitochondrial redox status, mitochondrial membrane potential, intracellular ROS level and cell activity, respectively. Western blot was used to detect the protein expression. The rat model of Freund’s complete adjuvant arthritis (AA) was established and treated with NAC and FCCP, respectively. The pathological changes of synovium and the percentage of apoptotic cells in synovium were detected by H&E and TUNEL staining, respectively.
Results: We have successfully isolated synovial cells from RA patients. Using 5μM H2O2 to stimulate RA-FLS could induce mitochondrial abnormalities of RA-FLS and inhibit RA-FLS autophagy. FCCP could reverse the effect of H2O2 on RA-FLS cell proliferation and apoptosis. NAC could reverse the effect of H2O2 on PINK1/Parkin. Overexpression of PINK1 or Parkin reversed the effect of H2O2 on RA-FLS mitochondrial autophagy, proliferation and apoptosis. The in vivo experiment results showed that both NAC and FCCP could prevent the pathogenesis of RA, reduce RA-FLS cell viability and increase RA-FLS cell apoptosis.
Conclusion: The PINK1/Parkin-mediated mitochondrial autophagy participates in H2O2-induced abnormal proliferation of RA-FLS, and targeting of PINK1/Parkin-mediated mitochondrial autophagy may be the key mechanism in the treatment of RA.
Keywords: mitochondrial autophagy, rheumatoid arthritis, PINK1, Parkin
Introduction
Rheumatoid arthritis (RA) is a chronic, systemic and destructive autoimmune disease.1 The synovial surface of normal joints consists of only 1–2 layers of cells, of which more than 70% are type B fibroblast-like synoviocytes (FLS). In the synovium of RA patients, FLS increases to 5~6 layers, or even more layers,2 and FLS proliferation is significantly accelerated.3 Although many kinds of cells are involved in RA progression, the abnormal proliferation of FLS is an important pathological process of RA. FLS is the effector cell of RA.4 However, the cause and mechanism of FLS hyperproliferation have not been clarified yet.
Reactive oxygen species (ROS) is one of the important factors in the pathophysiological process of RA.5 Compared with normal synovial fluid, the ROS level in RA patients’ synovial fluid is obvious increased.6 Our previous research also confirmed that low concentrations of H2O2 promoted the abnormal proliferation of RA-FLS.7 More than 90% of ROS in the body comes from the mitochondrial electron respiratory chain. High levels of ROS can cause cell degeneration, aging, malignant change and death, while the lower level of ROS is an important regulatory basis for cell cycle and cell proliferation.8 Long-term exposure to high levels of ROS can lead to mitochondrial dysfunction, oxidative damage of mitochondrial membrane lipids, and increase the excessive production of ROS in cells, thus changing mitochondrial membrane potential.9
ROS-mediated mitochondrial oxidative damage, which is eliminated by autophagy.9 In mammals, PINKl/Parkin pathway is a classical way to regulate mitochondrial autophagy.10 Under normal conditions, with the guidance of mitochondrial raking sequence, PINK1 is transferred from cytoplasm to outer membrane of mitochondria, transported into the matrix, hydrolyzed by related proteases, then released to the cytoplasm again and finally degraded by the proteasome. Once the mitochondria are damaged, the mitochondrial membrane potential is depolarized, PINK1 will not enter the matrix but rapidly accumulate in the outer membrane of the damaged mitochondria, and then recruit cytoplasmic Parkin to migrate to the mitochondria, finally causing the damaged mitochondria to degrade through autophagy-lysosome pathway.11
The relationship between ROS and mitochondrial dysfunction or autophagy has attracted more and more attention.12 The PINK1/Parkin pathway mediated mitophagy could effectively eliminate damaged mitochondria, avoiding excessive ROS production.13 Therefore, we speculated that the accumulation of ROS caused mitochondrial damage; meanwhile, ROS inhibited PINKl/Parkin pathway. So the mitochondrial autophagy is blocked and damaged mitochondria cannot be cleared in time, which leads to FLS abnormal proliferation and promotes the occurrence and development of RA.
Materials and Methods
Clinical Sample Collection and Ethics
Synovial biopsies were obtained from RA patients who underwent knee replacement surgery in the First Affiliated Hospital of Anhui Medical University. All samples and clinical information in this study were collected after obtaining the written informed consent of all patients. This study complies with the Declaration of Helsinki and was approved by the Clinical Research Ethics Committee of First Affiliated Hospital of Anhui Medical University (PJ20230231).
Isolation and Identification of Synovial Cells
Fresh synovial tissues were washed and minced into small pieces, which were digested with type I collagenase and trypsin. Then, the obtained cell was suspended in DMEM medium for routine culture. RA-FLS were stained with the vimentin and CD68 antibody for immunohistochemistry staining and observed under a microscope. The cell experiment in this project uses the 3–7th generation RA-FLS.
Cell Infection and Treatment
Lentivirus-mediated overexpression of PINK1, Parkin and the blank control were constructed by GeneChem. For infection, the RA-FLS was transfected with PINK1, Parkin overexpression lentivirus or control lentivirus. Twenty-four hours later, cells were observed by fluorescence microscope to prove the efficiency of infection. Transfected RA-FLS were seeded into the 96-well plate for overnight, exposed to H2O2 (5 μM) and treated with trifluoromethoxy carbonylcyanide phenylhydrazone (FCCP, 2 µM, a mitochondrial autophagy activator) to active FCCP or NAC (10 μmol/L, a ROS inhibitor). Cells were harvested for the subsequent experiments.
RT-qPCR
The Trizol reagent (Invitrogen) was used to extract total RNA. The isolated RNA was reverse transcribed. RT-qPCR was conducted with SYBR Green qPCR Master Mix (Takara). β-actin was used for normalization, and relative expression levels of Pink1 and Parkin were calculated by the 2−ΔΔCT method.
Primer Sequence (5’-3’), PINK1, F: GCCTCATCGAGGAAAAACAGG, R: GTCTCGTGTCCAACGGGTC; Parkin, F: GTGTTTGTCAGGTTCAACTCCA, R: GAAAATCACACGCAACTGGTC; β-actin, F: CATGTACGTTGCTATCCAGGC; R: CTCCTTAATGTCACGCACGAT.
Western Blot
The cells were lysed using RIPA lysis buffer to extract total protein. After protein quantification, load about 30g protein into SDS-PAGE gels, incubated with primary and secondary antibodies in turn. The blots were developed using an enhanced chemiluminescence kit and the band intensity was quantified with ImageJ software. β-actin and COX-IV were used as control.
Measurement of the Mitochondrial Membrane Potential
The mitochondrial membrane potential was assessed by the MitoProbe TM JC-1 assay kit. At 37°C, we first incubate RA-FLS with JC-1 for about 0.5h. The JC-1 monomer/aggregate was observed by fluorescence microscope and photos were taken. Then, the ImageJ imaging system software was used to evaluate the mitochondrial membrane potential by calculating the green/red fluorescence ratio.
MtROS Assessment
The levels of mtROS were detected by MitoSOX Red Mitochondrial Superoxide Indicator. After being treated with H2O2, the RA-FLS were cultured with MitoSOX Red Mitochondrial Superoxide Indicator, and washed with fresh growth medium. A confocal laser scanning microscope was used to collect images. The red fluorescence signal was quantitative analysis by the built-in evaluation software.
Intracellular ROS Level
The intracellular ROS level was detected using 2’,7’-dichlorofluorescein diacetate (DCFH-DA). After treatment, RA-FLS was washed with PBS, incubated with 5 µM DCFH-DA for 30 min at 37°C in darkness, and measured fluorescence using an inverted fluorescence microscope. Then, the Image J software was used to analyze the fluorescence intensity.
Mitochondrial Isolation
The cell mitochondrial extraction kit was used for the mitochondrial isolation of RA-FLS cells. The mitochondria and cytoplasm were separated by differential centrifugation and stored in a storage solution containing phenylmethylsulfonyl fluoride for subsequent immunoblot analysis.
CCK-8 Assay for Cell Viability
After treatment, CCK-8 solution was added to cell culture well, and incubated for 4 h. The microplate reader was used to measure the optical density (OD) at 450 nm.
Adjuvant Arthritis (AA) Rat Model and Arthritis Index Score
Male SD rats weighed 180±20 g were freely available for standard food and tap water and acclimatized in temperature and humidity-controlled rooms for one week. The SD rats were randomly divided into two groups: control (n = 6) and AA model group (n = 18). The control rats were injected with 150 μL physiological saline. The AA model group rats left were injected with 150 μL Freund’s complete adjuvant for 20 days at hind toe. Then, AA rats were randomly assigned to three groups treated with DMSO (n = 6), NAC (200 mg/kg/d/, n = 6) and FCCP (50 nM/kg/d/, n = 6) by continuous intra-articular injection for 12 d. The degree of arthritis index was determined based on the previous reference,7 whereby severity was scored on a scale of 0–4, where 0 = absent, 1 = minimal, 2 = mild, 3 = moderate, and 4 = severe. After the experiment, all rats were sacrificed, and synovial tissue was collected and stored at −80°C or fixed in 4% paraformaldehyde for later analysis.
All animal experiments were approved by the Animal Care Committee of Anhui Medical University, and were performed in accordance with the animal ethics guidelines of Agricultural Animals for Research and Teaching at Anhui Medical University.
H&E Staining
The synovial tissue fixed with 4% paraformaldehyde was dehydrated, permeabilized, embedded, and cut into slices for H&E staining. Then, we observed and evaluated the pathological conditions of stained sections under the microscope.
TUNEL Assay
The cell apoptosis of the synovial tissues was evaluated using the TUNEL assay. Firstly, we mounted the synovial slice on the glass slide. The slices were incubated with DNase free proteinase K and 3% H2O2. Then, we performed section dyeing according to TUNEL kit instructions. Lastly, the sections were cleared, mounted, covered, and examined using a microscope.
Statistical Analysis
Data were presented as means ± standard deviation (SD) and analyzed using Prism 7. Student’s t-test was performed to analyze differences between two groups. One-way analysis of variance followed by Tukey’s post-tests was performed to analyze differences among multiple groups. P < 0.05 was considered significant.
Results
Oxidative Stress Induced Mitochondrial Abnormalities of RA-FLS
Firstly, we isolated synovial cells from the synovial tissue of RA patients and named RA-FLS. After cell passage and culture, the adherent cells were uniformly distributed in a long spindle shape. The RA-FLS cell grew in a “vortex” shape, and other miscellaneous cells gradually decreased and disappeared (Figure 1A). Subsequently, the RA-FLS was identified by Vimentin and CD68 protein immunohistochemistry. Most of the cells showed Vimentin positive, and the cells did not express CD68 molecule (Figure 1B), which suggested that we have successfully isolated synovial cells.
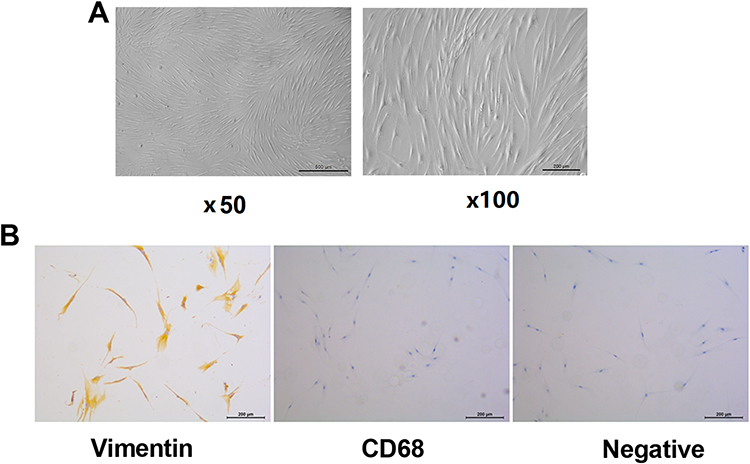 |
Figure 1 Isolation, culture and identification of fibroblast-like synoviocytes from rheumatoid arthritis (RA-FLS). (A) RA-FLS was isolated and cultured in vitro. (B) The selected RA-FLS was identified by Vimentin and CD68 immunohistochemistry. |
According to previous research,7 we used 5 μM H2O2 to stimulate RA-FLS cells for 12 h. The mitochondrial membrane potential (Δψm) results showed that there was a significantly decreased red fluorescence and increased green fluorescence in the H2O2 group (Figure 2A and B). Then, the mitochondrial and intracellular ROS levels were detected by MitoSOXTMRed and DCFH-DA fluorescence, respectively. As shown in Figure 2C-F, the H2O2 treatment induced the mitochondrial and intracellular ROS levels. Moreover, the down-regulated LC3II/I ratio and the Beclin 1 expression, and up-regulated P62 expression suggested that the H2O2 treatment inhibited autophagy in RA-FLS (Figure 2G). These data showed that oxidative stress induced mitochondrial abnormalities of RA-FLS.
 |
Figure 2 Oxidative stress induced mitochondrial abnormalities of RA-FLS. The RA-FLS cells were stimulated with 5µM H2O2 for 12 h, and the normal cultured cells as a control group. (A and B) The mitochondrial membrane potential was detected by the JC-1 assay kit. (C and D) The mitochondrial ROS was detected by MitoSOXTMRed fluorescence. (E and F) The intracellular ROS level was detected by DCFH-DA fluorescence. (G) Western blot was used to detect the expression of autophagy-related proteins (LC3II, Beclin 1 and P62). ***P < 0.001 vs Ctrl group. |
Oxidative Stress Mediated Mitochondrial Autophagy to Regulate RA-FLS Cells Proliferation and Apoptosis
To study the effect of mitochondrial autophagy on the RA-FLS cells proliferation and apoptosis under oxidative stress, we used the trifluoromethoxy carbonylcyanide phenylhydrazone (FCCP) to activate mitochondrial autophagy. H2O2 treatment significantly increased RA-FLS viability, while FCCP treatment reversed the effect of H2O2 treatment, reducing RA-FLS viability (Figure 3A). As shown in Figure 3B, the effect of H2O2 treatment on the LC3II/I ratio and the expression of Beclin 1 and P62 were reversed by FCCP treatment. Meanwhile, the effect of H2O2 treatment on the apoptosis-related proteins (Bax, Bcl-2 and cleaved-caspase 3) were reversed by FCCP treatment (Figure 3C). We used cell viability to assess cell proliferation. These results demonstrated that oxidative stress mediated mitochondrial autophagy to regulate the RA-FLS cells proliferation and apoptosis.
 |
Figure 3 Oxidative Stress mediated mitochondrial autophagy to regulate proliferation and apoptosis of RA-FLS cells. RA-FLS cells were treated with H2O2 and mitochondrial autophagy activator (FCCP). (A) The RA-FLS cell viability was detected by CCK-8 assay. (B and C) Western blot was used to detect the expression of autophagy and apoptosis-related proteins. ***P < 0.001 vs Ctrl group, ###p < 0.001 vs H2O2 group. |
Oxidative Stress Inhibited Mitochondrial Autophagy to Promote Abnormal Proliferation of RA-FLS by Down-Regulation of PINK1/Parkin Pathway
To explore the role of the PINK1/Parkin signaling pathway in oxidative stress mediated mitochondrial autophagy on cell proliferation/apoptosis, RA-FLS cells were stimulated with H2O2 and treated with ROS inhibitor (NAC). We found that oxidative stress inhibited the expression of PINK1 and Parkin, which could reserve by NAC treatment (Figure 4A). Then, we further detected the expression of PINK1 and Parkin in cytoplasm and mitochondria. The Western blot results showed that oxidative stress inhibited the expression of PINK1 and Parkin in mitochondria, which could reserve by NAC treatment (Figure 4B). However, the expression trend of PINK1 and Parkin in cytoplasm is opposite to that in mitochondria, that is, oxidative stress significantly up-regulated PINK1 and Parkin in the cytoplasm, which could reverse by NAC treatment (Figure 4C). These results indicated that oxidative stress inhibited the PINK1/Parkin signaling pathway in mitochondria.
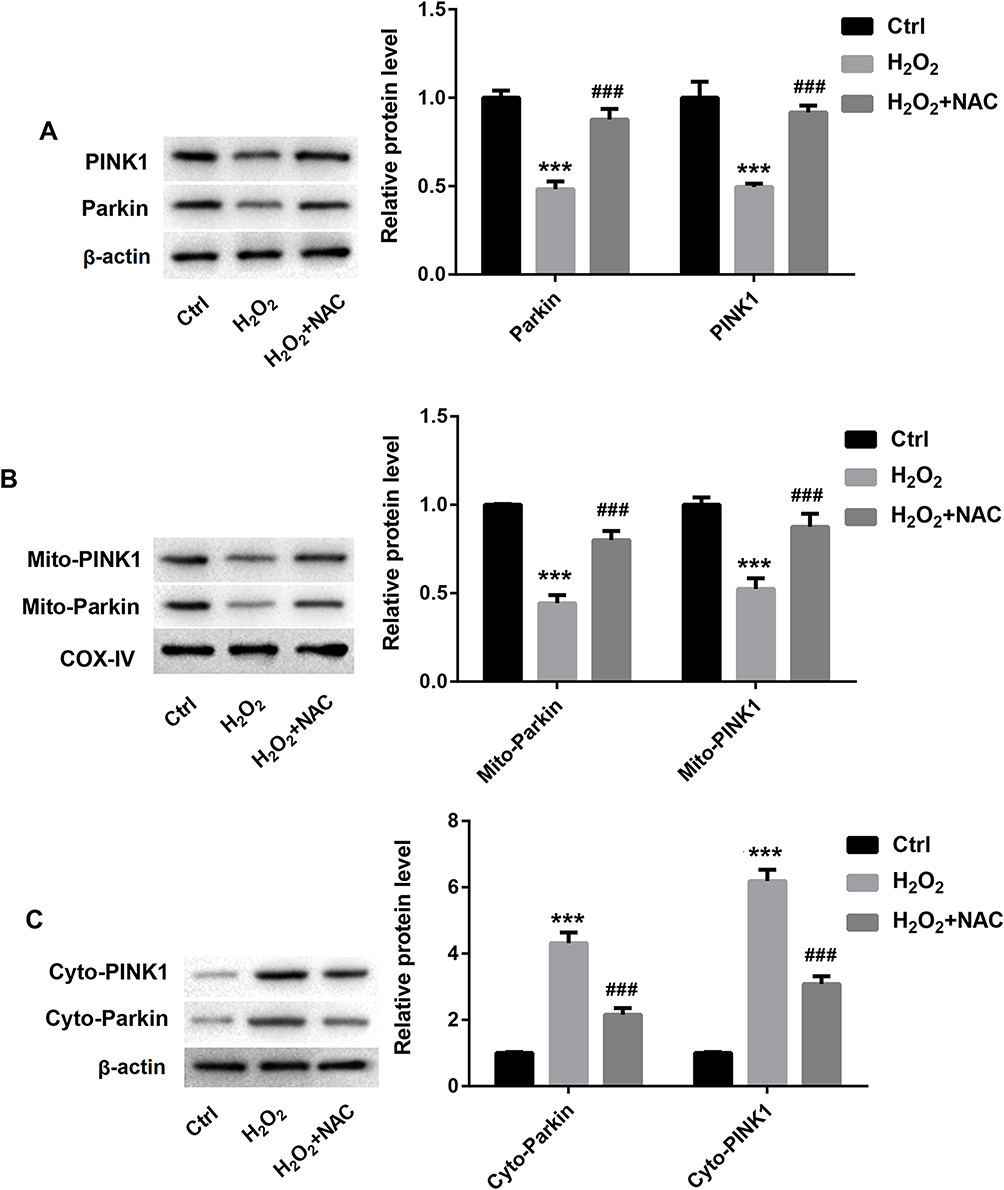 |
Figure 4 Oxidative stress inhibited PINKl and Parkin expression in mitochondria of RA-FLS. RA-FLS cells were stimulated with H2O2 and ROS inhibitor (NAC). (A) The protein expression of PINK1 and Parkin in RA-FLS. (B and C) The protein expression of PINK1 and Parkin in mitochondria and cytoplasm of RA-FLS. ***P < 0.001 vs Ctrl group, ###p < 0.001 vs H2O2 group. |
Next, RA-FLS cells were stimulated by oxidative stress after PINK1 or Parkin overexpression treatment. The RA-FLS was transfected with PINK1 or Parkin overexpression lentivirus. Figure 5 showed the PINKI and Parkin overexpression efficiency. As shown in Figure 6A, H2O2 treatment increased cell viability, which was reversed by PINK1 or Parkin overexpression treatment. The Western blot results showed that H2O2 treatment inhibited autophagy, which were all reversed by PINK1 or Parkin overexpression treatment (Figure 6B). The autophagy related protein expression results suggested that oxidative stress inhibited autophagy by inhibiting the PINK1/Parkin signal pathway. Then, the Western blot results showed that H2O2 treatment reduced apoptosis-related protein expression, and increased the anti-apoptosis protein expression, which were all reversed by PINK1 or Parkin overexpression treatment (Figure 6C). The apoptosis-related protein expression results suggested that oxidative stress inhibited apoptosis by inhibiting the PINK1/Parkin signal pathway. Lastly, cell viability was used to assess cell proliferation. The cell viability results suggested that oxidative stress promoted cell proliferation by restraining PINK1/Parkin pathway.
 |
Figure 5 PINKI and Parkin overexpression efficiency. (A and B) The RA-FLS was transfected with PINK1 overexpression lentivirus or control lentivirus. The mRNA and protein expression of PINK1 were detected by RT-qPCR and Western blot to demonstrate the overexpression efficiency. (C and D) The RA-FLS was transfected with Parkin overexpression lentivirus or control lentivirus. The mRNA and protein expression of Parkin were detected by RT-qPCR and Western blot to demonstrate the overexpression efficiency. ***P < 0.001 vs NC group. |
 |
Figure 6 Oxidative stress inhibited mitochondrial autophagy to promote abnormal proliferation of RA-FLS by down-regulation of the PINK1/Parkin pathway. RA-FLS cells were overexpressed with PINK1 and Parkin, and then stimulated with H2O2. (A) The RA-FLS cell viability was detected by CCK-8 assay. (B and C) Western blot was used to detect the expression of autophagy and apoptosis-related proteins. ***P < 0.001 vs Ctrl group, ##p < 0.01, ###p < 0.001 vs H2O2 + NC group. |
Oxidative Stress Promoted the Pathogenesis of RA by Inhibiting Mitochondrial Autophagy
Through the above in vitro cell experiments, we proved that oxidative stress inhibited mitochondrial autophagy to promote abnormal proliferation of RA-FLS by down-regulation of the PINK1/Parkin pathway. Finally, we used ROS inhibitor (NAC) and mitochondrial autophagy activator (FCCP) in rats to explore whether oxidative stress participated in the development of RA by inhibiting mitochondrial autophagy. Adjuvant arthritis (AA) rat models were treated with NAC or FCCP, respectively. After treatment with NAC or FCCP, the arthritis index of rats in the NAC or FCCP group were both significantly reduced (Figure 7A). H&E staining results of rat joint synovium displayed the significantly proliferated synovial cells in the model group, which was effectively reduced in NAC and FCCP treatment group (Figure 7B). Later, we isolated synovial cells from synovial tissue and detected their viability with a CCK8 kit (Figure 7C). The results showed that RA-FLS in the model group had higher cell activity, while the RA-FLS activity in the NAC and FCCP treatment group were significantly reduced, suggesting that NAC and FCCP treatment could effectively inhibit RA-FLS proliferation. Moreover, we used the TUNEL kit to detect the apoptotic cells in the synovial tissue of rats in each group. The results showed that the proportion of TUNEL positive cells in model group was significantly lower than that in control group, but significantly higher in the NAC and FCCP treatment group. TUNEL-positive cells represent apoptotic cells, and the TUNEL results showed that NAC and FCCP treatment could effectively promote the apoptosis of synovial cells (Figure 7D and E). These in vivo results demonstrated that oxidative stress promoted the pathogenesis of RA by inhibiting mitochondrial autophagy.
 |
Figure 7 Oxidative stress promoted the pathogenesis of RA by inhibiting mitochondrial autophagy. SD rats were randomly divided into 4 groups: Normal control group, AA model group, NAC treatment group, and FCCP treatment group. (A) Arthritis index was calculated; (B) H&E staining was used to evaluate the pathology of synovial tissue, scale bar=200µm. (C) Synovial cells were isolated and cell viability was detected by CCK8. (D and E) TUNEL was used to detect the percentage of apoptosis of synovial cells in synovial tissue, scale bar=100µm. ***P < 0.001 vs Normal group, ###p < 0.001 vs model group. |
Discussion
FLS are the major population in the synovium and play a critical role in arthritis pathogenesis.14 FLS show aggressive/transformational phenotype. In an environment rich in oxygen-free radicals, nitric oxide and cytokines, FLS’s autonomous survival ability is similar to the phenotype of some tumors.4,15 In this study, the synovial cells were isolated from RA patients, and the Vimentin and CD68 protein immunohistochemistry results proved that we had successfully isolated synovial cells.
ROS plays a key role in many biological processes.16 Researches showed that 5 μM H2O2 could increase cell activity to 111%.7,17 Here, we used 5 μM H2O2 to stimulate oxidative stress in RA-FLS cells. Consistent with the reported literature,17–19 our study found that oxidative stress significantly induced RA-FLS proliferation, and increased ROS level and mitochondrial membrane potential. Dysfunctional autophagy is associated with the development of RA.20 Here, we found that oxidative stress inhibited mitochondrial autophagy in RA-FLS. Studies have shown that acute dissipation of mitochondrial membrane potential by mitochondrial uncouplers (FCCP) can initiate mitophagy.21 One of the reasons for FLS abnormal proliferation in RA is the imbalance between FLS cell apoptosis and autophagy.22 Thus, we used FCCP to activate mitochondrial autophagy and found that FCCP reversed the effects of H2O2 on FLS cell autophagy, proliferation and apoptosis. At the same time, the animal experiment results showed that after intra-articular injection of FCCP in AA rats, the joint index and synovial hyperplasia of rats decreased significantly. These results indicated that oxidative stress affected the proliferation of RA-FLS cells by regulating mitochondrial autophagy.
In bone metabolic disorders disease, PINK1/Parkin-mediated mitophagy plays a protective role in RA.23,24 Any deficiency of Parkin or PINK1 will damage mitochondrial autophagy, thereby increasing oxidative stress.25 In this paper, we found that oxidative stress inhibited the PINK1 and Parkin expression in mitochondria, suggesting damaged mitochondrial autophagy by H2O2 treatment, which was consistent with some existing reports.26,27 Then, NAC was used to inhibit ROS levels. The animal experiment results showed that after intra-articular injection of NAC in AA rats, the joint index and synovial hyperplasia of rats decreased significantly, which corresponds to Batooei’s clinical research report.28 The effect of NAC on PINK1 and Parkin expression is as expected, that is, NAC treatment up-regulated the PINK1 and Parkin expression in mitochondria. In addition, PINK1 or Parkin overexpression reversed the role of H2O2 treatment on proliferation and apoptosis of RA-FLS. This result proved that oxidative stress inhibited mitochondrial autophagy to promote abnormal proliferation of RA-FLS by down-regulation of the PINK1/Parkin pathway.
In conclusion, oxidative stress plays a key role in promoting the abnormal proliferation of RA-FLS, but its specific mechanism is still unclear. Therefore, our study provides a new theoretical basis for the mechanism of abnormal proliferation of RA-FLS.
Funding
This study was supported by 2018 Natural Science Foundation of Anhui Province (No. 1808085MH243).
Disclosure
All authors declare no conflicts of interest in this work.
References
1. Weyand CM, Goronzy JJ. The immunology of rheumatoid arthritis. Nat Immunol. 2021;22:10–18.
2. Lu Y, Hao C, Yu S, et al. Cationic amino acid transporter-1 (CAT-1) promotes fibroblast-like synoviocyte proliferation and cytokine secretion by taking up L-arginine in rheumatoid arthritis. Arthritis Res Ther. 2022;24:234.
3. Nygaard G, Firestein GS. Restoring synovial homeostasis in rheumatoid arthritis by targeting fibroblast-like synoviocytes. Nat Rev Rheumatol. 2020;16:316–333.
4. Bartok B, Firestein GS. Fibroblast-like synoviocytes: key effector cells in rheumatoid arthritis. Immunol Rev. 2010;233:233–255.
5. Phull AR, Nasir B, Haq IU, Kim SJ. Oxidative stress, consequences and ROS mediated cellular signaling in rheumatoid arthritis. Chem Biol Interact. 2018;281:121–136.
6. Pradhan A, Bagchi A, De S, et al. Role of redox imbalance and cytokines in mediating oxidative damage and disease progression of patients with rheumatoid arthritis. Free Radic Res. 2019;53:768–779.
7. Zhang J, Song X, Cao W, et al. Autophagy and mitochondrial dysfunction in adjuvant-arthritis rats treatment with resveratrol. Sci Rep. 2016;6:32928.
8. Ciccarone F, Castelli S, Ciriolo MR. Oxidative stress-driven autophagy acROSs onset and therapeutic outcome in hepatocellular carcinoma. Oxid Med Cell Longev. 2019;2019:6050123.
9. Yao RQ, Ren C, Xia ZF, Yao YM. Organelle-specific autophagy in inflammatory diseases: a potential therapeutic target underlying the quality control of multiple organelles. Autophagy. 2021;17:385–401.
10. Gladkova C, Maslen SL, Skehel JM, Komander D. Mechanism of parkin activation by PINK1. Nature. 2018;559:410–414.
11. Gan ZY, Callegari S. Activation mechanism of PINK1. Nature. 2022;602:328–335.
12. Yun HR, Jo YH, Kim J, Shin Y, Kim SS, Choi TG. Roles of autophagy in oxidative stress. Int J Mol Sci. 2020;1:21.
13. Liu X, Hussain R. Mitochondrial-endoplasmic reticulum communication-mediated oxidative stress and autophagy. Biomed Res Int. 2022;2022:6459585.
14. Bustamante MF, Garcia-Carbonell R, Whisenant KD, Guma M. Fibroblast-like synoviocyte metabolism in the pathogenesis of rheumatoid arthritis. Arthritis Res Ther. 2017;19:110.
15. Bottini N, Firestein GS. Duality of fibroblast-like synoviocytes in RA: passive responders and imprinted aggressors. Nat Rev Rheumatol. 2013;9:24–33.
16. Zhang L, Wang K, Lei Y, Li Q, Nice EC, Huang C. Redox signaling: potential arbitrator of autophagy and apoptosis in therapeutic response. Free Radic Biol Med. 2015;89:452–465.
17. Cao W, Zhang J, Wang G, Lu J, Wang T, Chen X. Reducing-autophagy derived mitochondrial dysfunction during resveratrol promotes fibroblast-like synovial cell apoptosis. Anatomical Record. 2018;301:1179–1188.
18. Yang J, Huang Y, Liu M, et al. N-acetylcysteine inhibits the proliferation of hydrogen peroxide treated fibroblast-like synoviocytes in rats with adjuvant arthritis (AA) via blocking Nrf2/Keap1 pathway. Chine J Cel Mol Immunol. 2021;37:687–692.
19. Lv X, Huang J, Wang H. MiR-30a-3p ameliorates oxidative stress in rheumatoid arthritis synovial fibroblasts via activation of Nrf2-ARE signaling pathway. Immunol Lett. 2021;232:1–8.
20. Ding JT, Hong FF, Yang SL. Roles of autophagy in rheumatoid arthritis. Clin Exp Rheumatol. 2022;1:485.
21. Georgakopoulos ND, Wells G, Campanella M. The pharmacological regulation of cellular mitophagy. Nat Chem Biol. 2017;13:136–146.
22. Misra S, Bagchi A, Sarkar A, et al. Methotrexate and theaflavin-3, 3’-digallate synergistically restore the balance between apoptosis and autophagy in synovial fibroblast of RA: an ex vivo approach with cultured human RA FLS. Inflammopharmacology. 2021;29:1427–1442.
23. Wang S, Deng Z, Ma Y, et al. The role of autophagy and mitophagy in bone metabolic disorders. Int J Biol Sci. 2020;16:2675–2691.
24. Jiang JM, Mo ML, Long XP, Xie LH. MiR-144-3p induced by SP1 promotes IL-1β-induced pyroptosis in chondrocytes via PTEN/PINK1/Parkin axis. Autoimmunity. 2022;55:21–31.
25. Barodia SK, Creed RB, Goldberg MS. Parkin and PINK1 functions in oxidative stress and neurodegeneration. Brain Res Bull. 2017;133:51–59.
26. Cao S, Wang C, Yan J, Li X, Wen J, Hu C. Curcumin ameliorates oxidative stress-induced intestinal barrier injury and mitochondrial damage by promoting Parkin dependent mitophagy through AMPK-TFEB signal pathway. Free Radic Biol Med. 2020;147:8–22.
27. Zhang H, Liu X, Fan Y, et al. l-Arginine alleviates hydrogen peroxide-induced oxidative damage in ovine intestinal epithelial cells by regulating apoptosis, mitochondrial function, and autophagy. J Nutr. 2021;151:1038–1046.
28. Batooei M, Tahamoli-Roudsari A, Basiri Z, et al. Evaluating the effect of oral N-acetylcysteine as an Adjuvant treatment on clinical outcomes of patients with rheumatoid arthritis: a randomized, double blind clinical trial. Rev Recent Clin Trials. 2018;13:132–138.
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Norisoboldine Reduces Arthritis Severity by Attenuating Inflammation, Oxidative Stress, and Extracellular Matrix Degradation in a Rat Model of Rheumatoid Arthritis
Wang Y, Zhao X, Wang J, Zhu X
Journal of Inflammation Research 2024, 17:8839-8852
Published Date: 15 November 2024