Back to Journals » Pharmacogenomics and Personalized Medicine » Volume 13
Pharmacogenetics of Pediatric Asthma: Current Perspectives
Authors Perez-Garcia J ![]() , Espuela-Ortiz A
, Espuela-Ortiz A ![]() , Lorenzo-Diaz F, Pino-Yanes M
, Lorenzo-Diaz F, Pino-Yanes M ![]()
Received 20 December 2019
Accepted for publication 3 March 2020
Published 18 March 2020 Volume 2020:13 Pages 89—103
DOI https://doi.org/10.2147/PGPM.S201276
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Martin Bluth
Javier Perez-Garcia,1 Antonio Espuela-Ortiz,1 Fabian Lorenzo-Diaz,1,2 Maria Pino-Yanes1,3,4
1Genomics and Health Group, Department of Biochemistry, Microbiology, Cell Biology and Genetics, Universidad de La Laguna, San Cristóbal de La Laguna, Santa Cruz de Tenerife, Spain; 2Instituto Universitario de Enfermedades Tropicales y Salud Pública de Canarias (IUETSPC), Universidad de La Laguna, San Cristóbal de La Laguna, Santa Cruz de Tenerife, Spain; 3CIBER de Enfermedades Respiratorias, Instituto de Salud Carlos III, Madrid, Comunidad de Madrid, Spain; 4Instituto de Tecnologías Biomédicas (ITB), Universidad de La Laguna, San Cristóbal de La Laguna, Santa Cruz de Tenerife, Spain
Correspondence: Maria Pino-Yanes
Genomics and Health Group, Department of Biochemistry, Microbiology, Cell Biology and Genetics, Universidad de La Laguna, Apartado 456, San Cristóbal de La Laguna 38200, Santa Cruz de Tenerife, Spain
Tel +34 922316 502 – 6343
Email [email protected]
Abstract: Asthma is a chronic respiratory disease that affects 339 million people worldwide and has a considerable impact on the pediatric population. Asthma symptoms can be controlled by pharmacological treatment. However, some patients do not respond to therapy and continue suffering from symptoms, which impair the quality of life of patients and limit their daily activity. Genetic variation has been shown to have a role in treatment response. The aim of this review is to update the main findings described in pharmacogenetic studies of pediatric asthma published from January 1, 2018 to December 31, 2019. During this period, the response to short-acting beta-agonists and inhaled corticosteroids in childhood asthma has been evaluated by eleven candidate-gene studies, one meta-analysis of a candidate gene, and six pharmacogenomic studies. The findings have allowed validating the association of genes previously related to asthma treatment response (ADRB2, GSDMB, FCER2, VEGFA, SPAT2SL, ASB3, and COL2A1), and identifying novel associations (PRKG1, DNAH5, IL1RL1, CRISPLD2, MMP9, APOBEC3B-APOBEC3C, EDDM3B, and BBS9). However, some results are not consistent across studies, highlighting the need to conduct larger studies in diverse populations with more homogeneous definitions of treatment response. Once stronger evidence was established, genetic variants will have the potential to be applied in clinical practice as biomarkers of treatment response enhancing asthma management and improving the quality of life of asthma patients.
Keywords: treatment response, short-acting beta-agonists, inhaled corticosteroids, candidate-gene studies, genome-wide association studies, whole-genome sequencing
Introduction
Asthma is a heterogeneous respiratory disease characterized by chronic airway inflammation, reversible airflow obstruction, and airway hyperresponsiveness.1 Asthma symptoms include wheeze, dyspnea, chest tightness, and cough. These symptoms may resolve spontaneously, but most patients need a pharmacological treatment to control them.2 However, pharmacotherapy is not always effective, and some patients may develop worsening episodes of the baseline disease situation, known as exacerbations. These acute episodes are responsible for limitations in patient’s daily activity and may cause disability, the need for intubation, and can even be life-threatening.1,2
It has been estimated that 339 million people worldwide have asthma, and 420 thousand deaths per year are linked to this disease.1 Moreover, although asthma affects people of all ages, it has a towering impact on the pediatric population, being the most prevalent chronic disease in children.3 The global prevalence of pediatric asthma is about 11.5%, but there is a large variability of prevalence between regions, reaching values of 18.8% and 24.3% in Latin America and Oceania, respectively.4 Despite the scarce mortality of childhood asthma (global death rate: 0.0–0.7/100,000), children or young adults represent a considerable fraction of people who die from asthma. As an example, in the United Kingdom between 2012 and 2013, from a total of 194 deaths related to asthma, 28 of them occurred to people under 20 years old (14.4%).5
In order to enhance asthma management in terms of better diagnosis and more precise treatment, different disease subtypes sharing some common clinical characteristics, triggers, or type of inflammatory processes have been identified.6 Although to date no consistent findings have been discovered,7 the relationship between asthma phenotypes and treatment response may be used to select the most appropriate therapy for each patient.2,6 Nonetheless, despite following asthma guidelines,2 non-response to the pharmacological treatment is not uncommon in pediatric nor adult asthma.8 Thus, there is a need to establish new biomarkers that could allow improving disease management.
Genetic variants are known to have a role in asthma treatment response with different medications given that these traits have been shown to be heritable.9 During the last decades, different approaches have been developed to identify genetic markers associated with complex diseases, most of them based on identifying variants affecting only one base pair, which are called single-nucleotide polymorphisms (SNPs).10 Candidate-gene studies have been performed to identify genetic variants in specific genes previously associated with the disease and the pharmacological treatment.11 Genome-wide association studies (GWAS) have emerged as a pharmacogenomic approach to identify novel genetic markers without a prior hypothesis of the mechanisms involved in treatment response. GWAS data can be obtained both with genotyping microarrays and by means of high-throughput DNA sequencing. The microarrays used for genome-wide genotyping focus on certain SNPs that are representative of the variation across the genome, taking advantage of the linkage disequilibrium (LD) between variants. Thus, genotyping data allows subsequent inference of genetic variants throughout the whole genome.10 Furthermore, the more recent development of high-throughput DNA sequencing has allowed performing whole-genome sequencing (WGS), attaining a higher resolution of the genomic variation than genotyping arrays.12
The evidence of the role of genetic variants on asthma treatment response has been summarized in some recent reviews.13–18 Thereby, this review aims to update the current findings described in pharmacogenetic studies of pediatric asthma published from 2018 through 2019.
Methods
A literature research was performed in PubMed using different keywords combinations (Table S1). The inclusion criteria of the studies were: 1) studies published in PubMed from January 1, 2018, to December 31, 2019, 2) studies focused on pharmacogenetics of asthma, and 3) papers written in English. The exclusion criteria were: 1) studies focused only on adulthood asthma, 2) animal or in-vitro model studies, 3) studies not assessing asthma treatment response, and 4) reviews, editorials, opinion articles or case studies. A stepwise evaluation of the studies according to the eligibility criteria was conducted based on the title, the abstract, and finally the full text. At least two different authors independently evaluated each paper.
Short-Acting Beta-Agonists (SABA)
SABA are one of the main reliever medications in children and adolescents with asthma due to its rapid effect relieving asthma symptoms.2 The mechanism of action of these drugs consists in the activation of the β2 adrenergic receptor, which results in the relaxation of the bronchial smooth muscle and the consequent bronchodilation.19 Unlike long-acting beta-agonists (LABA), SABA have both rapid-onset bronchodilation and a shorter half-life.19 Genetic variants contribute to differences in SABA response in children with asthma.20,21 To study the genes involved in SABA response, one of the parameters most frequently analyzed as an outcome has been the bronchodilator response (BDR), based on changes in lung function after the administration of SABA.22 Also, lung function parameters such as forced expiratory volume in the first second (FEV1), forced vital capacity (FVC), and the ratio between them (FEV1/FVC), have been used to evaluate treatment response.
Candidate-Gene Studies of SABA Response
ADRB2 gene has been the most studied candidate gene of SABA response.16,18,23 It encodes the β2 adrenergic receptor, which is implicated in the bronchodilation process and is the biological target of β2 adrenergic agonists.19 In fact, in the reviewed period four candidate-gene studies have assessed the effect of the SNPs rs1042713 (arginine to glycine [Arg16Gly]) and rs1042714 (glutamic acid to glutamine [Glu27Gln]) on changes in lung function after SABA administration in European24,25 and Egyptian26,27 populations (Table 1). However, the results were not entirely consistent across studies. Scaparrotta et al24 identified that G allele carriers for the SNP rs1042713 had a lower post-bronchodilator FEV1 (β for the G allele: −0.244, p-value=0.02) while Jovicic et al25 reported the opposite effect (p-value=0.044). Additionally, when both SNPs were analyzed as a haplotype, inconsistent results were also found since the AC haplotype was associated with a worse response to SABA in European children25 (p-value=0.026) while in Egyptian children the same haplotype was associated with a better response (p-value=0.027).27 All these inconsistencies might be due to the heterogeneity in statistical approaches. Scaparrotta et al applied linear regression models adjusted for confounders, while the other studies used parametric/non-parametric tests that do not allow adjusting for covariates. On the other hand, none of these studies found an association of the SNP rs1042714 with SABA response.24–27
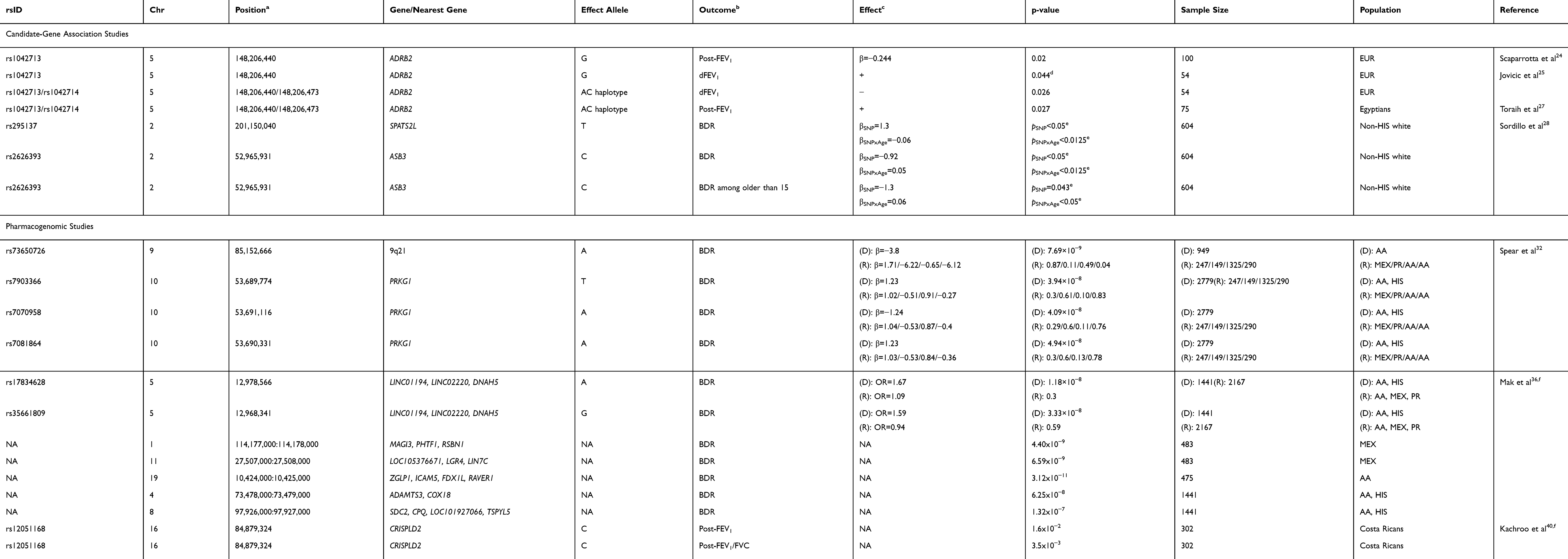 |
Table 1 Summary of the Genetic Associations with Short-Acting Beta Agonist Response Described in Pharmacogenetic Studies |
Sordillo et al28 conducted a longitudinal study in non-Hispanic white children (n=604) that were followed up 4–18 years. Four SNPs suggestively associated with SABA response by previous GWAS21,29,30 (rs11252394, rs6988229, rs295137, and rs2626393) were studied for association with asthma treatment response and age interaction analyzing BDR after albuterol administration as outcome. The SNP rs295137 near SPATS2L was associated with an increase in BDR (β for the T allele: 1.3, p-value<0.05) consistently with the results previously reported.29 This effect was progressively mitigated for each year of aging (β: −0.06, p-value<0.0125). In contrast, the SNP rs2626393 near ASB3 was found to be associated with a decrease in BDR (β for the C allele: −0.92, p-value<0.05), similarly to the findings of a previous GWAS for which this SNP is a proxy,30 and also its effect was reduced by aging (β: 0.05, p-value<0.0125). This SNP is in high LD with rs350729, a polymorphism near ASB3 previously associated with BDR, which has been identified as an expression quantitative trait locus (eQTL) of 52 genes that interact with ADRB2 and ASB3.30 In addition, these variants were also studied only with the data collected from 15 years of age and a similar effect of the rs2626393 remained significant at these older ages (p-value<0.05).28 The association of both genes and SABA response is plausible since SPATS2L appears to regulate the expression of the β2 adrenergic receptor29 and ASB3 regulates the muscle cells differentiation process and its expression is modified by β2 agonists.31
Pharmacogenomic Studies of SABA Response
During the reviewed period, three pharmacogenomic studies of SABA response have been performed, one based on genotyping array data and two based on WGS (Table 1). A GWAS performed by Spear et al32 identified a population-specific genome-wide significant variant (rs73650726 on 9q21) in African Americans (n=949), which was associated with a decrease of BDR after albuterol administration (β for the A allele: −3.8, p-value=7.69 × 10−9). This SNP does not map to any gene, but it is in high LD (r2≥0.8) with other SNPs located within enhancer histone marks in lung tissue.33 Furthermore, a meta-GWAS across African Americans and Latinos (n=2,779) allowed the identification of three SNPs in the PRKG1 gene related to SABA response: rs7903366 (β for the T allele: 1.23, p-value=3.94 × 10−8), rs7070958 (β for the A allele: −1.24, p-value=4.09 × 10−8), and rs7081864 (β for the A allele: 1.23, p-value=4.94 × 10−8). Nevertheless, these SNPs were assessed for replication in three independent African American and Latino cohorts and none of them demonstrated consistent effects. The PRKG1 gene encodes an isoform of the protein kinase cGMP-dependent, which is a key mediator of the nitric oxide (NO) pathway. NO is involved in several pathophysiological processes in the airways related to asthma status,34 and it is also involved in the bronchodilator effect on the bronchial smooth muscle of β2 agonist drugs such as albuterol.35
In 2018, Mak et al36 conducted a GWAS of SABA response based on WGS data in childhood asthma analyzing the extreme of the distribution in BDR after albuterol administration as outcome. A trans-ethnic meta-analysis in African American and Latino children (n=1,441) identified 27 SNPs suggestively associated (p-value<7.06 × 10−6) with BDR of which three SNPs were significantly associated after Bonferroni correction (p-value<3.53 × 10−7) and two reached the commonly used genome-wide significance threshold for GWAS (p-value<5 × 10−8). These two SNPs were located on chromosome band 5p15.2 near DNAH5, and both of them were associated with a better response to albuterol (rs17834628, odds ratio [OR] for the A allele: 1.67, 95% confidence interval [CI]: 1.29–2.16, p-value=1.18 × 10−8 and rs35661809, OR for the G allele: 1.59, 95% CI: 1.20–2.10, p-value=3.33 × 10−8). Despite the associations of these SNPs reached genome-wide significance, none of them replicated in five independent African American and Latino studies, analyzed separately or meta-analyzed. These findings are biologically plausible since genetic variants on DNAH5 have been previously associated with lung function in chronic obstructive pulmonary disease (COPD)37 and immunoglobulin E (IgE) levels.38 Moreover, combined effects of common and rare variants on BDR were identified in three population-specific loci (two in Mexicans and one in African Americans) and in two loci shared among African American and Latino populations which reached genome-wide significance (Table 1).36 Interestingly, one of the loci identified may have a functional role in response to SABA since it is located near ADAMTS3/COX18, a locus previously associated with the response to β-adrenergic drugs in animal models to treat cardiovascular diseases.39
The first WGS family-based analysis of lung function in Latino children with asthma (n=302) was recently conducted by Kachroo et al,40 analyzing not only baseline but also post-bronchodilator lung function measurements. In this study, the SNP rs12051168 from the CRISPLD2 gene was suggestively associated with the baseline FEV1/FVC ratio (p-value=9.1 × 10−6) and it was nominally associated with post-bronchodilator measurements of FEV1 (p-value=1.6 × 10−2) and FEV1/FVC (p-value=3.5 × 10−3). Interestingly, CRISPLD2 encodes a secreted lipopolysaccharide-binding protein that protects against bacterial infections41 and has a role in lung development.42 Also, this gene has been previously nominally associated with ICS response and BDR,43 and its gene expression levels have been found to be changed by indacaterol.44
Inhaled Corticosteroids (ICS)
Based on its efficacy, effectiveness, safety, and cost, ICS are the preferred treatment to control asthma symptoms as well as to prevent the development of asthma exacerbations.2 According to the clinical features of the patient and the degree of asthma severity, ICS may be used at different doses or may be combined with other pharmacological therapies such as LABA or leukotriene receptors antagonists (LTRA).2 Corticosteroids contribute to control asthma symptoms due to its anti–inflammatory and immunosuppressive effects as a result of the interaction with the glucocorticoid receptor (GR).45 Nonetheless, many factors such as an incorrect inhaler technique,46 a poor adherence,47 comorbidities,48 lung microbiome dysbiosis,49–52 and genetic variants,13,17 may contribute to a loss in asthma control and to the development of asthma exacerbations despite ICS-contained therapies. The identification of these factors is essential to differentiate children with difficult-to-treat asthma from those that are resistant to pharmacological therapies.53 Treatment response to ICS is usually evaluated by the development of asthma exacerbations while on ICS treatment, by clinical measurements (eg fractional exhaled nitric oxide [FENO], FEV1, and FEV1/FVC), and by the assessment of the asthma control test (ACT) score.
Candidate-Gene Studies of ICS Response
Several candidate-gene studies have been performed to evaluate the influence of genetic variants on treatment response to ICS (Table 2).26,54-59 Similarly to the response to SABA, the ADRB2 gene is one of the most extensively analyzed in pharmacogenetic studies of ICS.16,18,23 In 2018, Sood et al54 studied two SNPs (rs1042713 and rs1042714) from the ADRB2 gene in a European American and African American cohort (n=373) of children with asthma under ICS treatment. In this study, treatment response was defined as the prescription of systemic corticosteroids due to asthma exacerbations in the last 12 months. Asthma individuals with CC genotype at the SNP rs1042714 had an increased risk of asthma exacerbation despite ICS treatment in comparison with heterozygous individuals (CG; p-value=0.04) and homozygous individuals (GG; p-value=0.013).54 In the same year, Alghobashy et al26 also evaluated these two SNPs in Egyptian children with asthma (n=52) treated with ICS in combination with LABA and assessed lung function measurement (FEV1 and FEV1/FVC) as the primary outcome of treatment response. Worse treatment response to a combination of ICS plus LABA was reported for homozygous GG at rs1042713 (p-value<0.05) and homozygous GG at rs1042714 (p-value<0.05) individuals. Given that all the patients were treated with ICS plus LABA, this study design does not allow assessing whether the effect of the SNP conditions the response to ICS or LABA. However, no effect of rs1042713 in FEV1 or FEV1/FVC was found in a meta-analysis in five mixed pediatric populations under ICS and LABA treatment performed by Wang et al.60
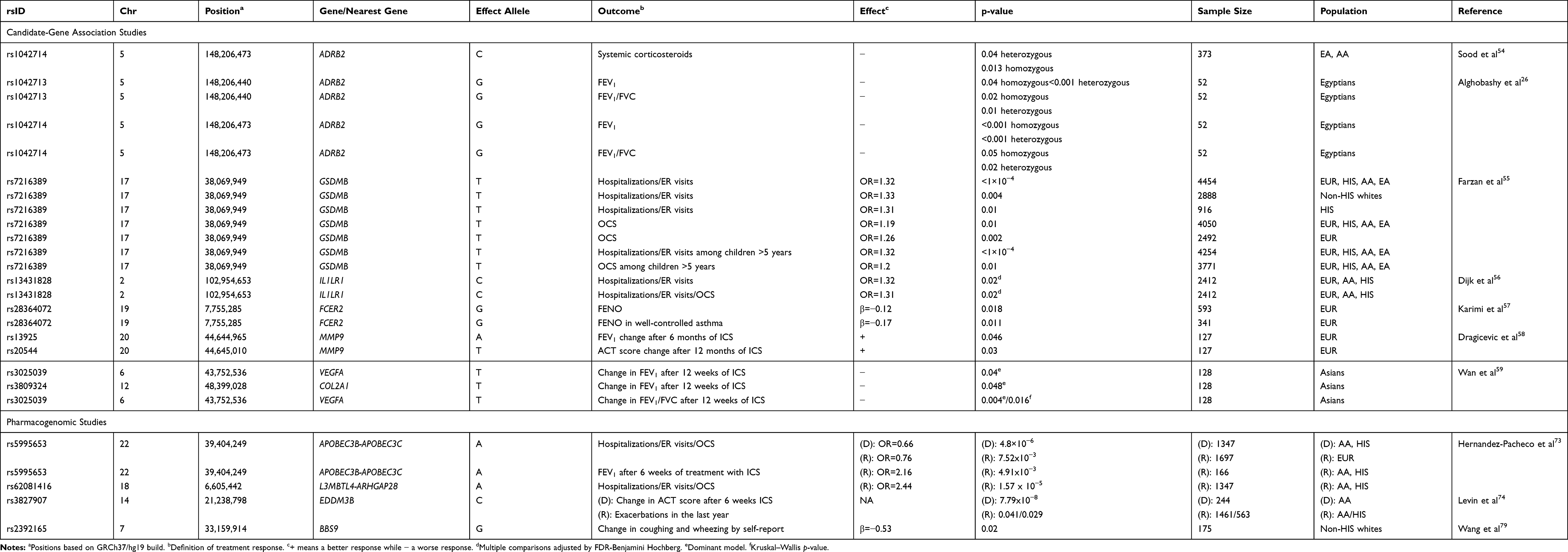 |
Table 2 Summary of the Genetic Associations with Inhaled Corticosteroids Response Described in Pharmacogenetic Studies |
In 2019, Farzan et al55 assessed the association of a polymorphism in the GSDMB gene (rs7216389) at the locus 17q21 with ICS response by means of a meta-analysis (n=4,454) of different ethnic populations, including Europeans (n=2,888), Hispanics/Latinos (n=916), African Americans (n=468), and East Asians (n=182). This locus has not only shown a consistent relationship with childhood-onset asthma61,62 but has also been associated with ICS response in asthma.63 They measured treatment response according to the development of asthma exacerbations, defined as 1) oral corticosteroid (OCS) prescription and 2) hospitalizations/emergency room visits (ER) due to asthma. The SNP rs7216389 was associated with a worse response to ICS for both outcomes (OR for the T allele: 1.19, 95% CI: 1.04–1.36, p-value=0.01 for OCS prescription, and OR: 1.32, 95% CI: 1.17–1.49, p-value=<1 × 10–4 for hospitalizations/ER visits). Moreover, stratified analyses by age and ethnicity showed similar results, except for pre-school children (2–4 years of age) (p-value>0.05), and OCS prescription in Hispanics/Latinos (p-value>0.05).
A recent study by Dijk et al56 also evaluated the role of a gene identified by previous GWAS of asthma,14,64 IL1RL1, on the response to ICS measured as asthma exacerbations (OCS use and Hospitalizations/ER visits). For that purpose, a study focused on Europeans (n=720) was analyzed as discovery, and replication was assessed in Hispanics/Latinos (n=876), African Americans (n=525), and Europeans (n=301). This gene encodes the interleukin 1 receptor-like 1 (IL1RL1), which acts as a decoy receptor for interleukin 33 (IL33), and it is known that the IL33/IL1RL1 signaling pathway is related to the type 2 inflammation.65 For the six SNPs analyzed, a meta-analysis across the 2,412 samples allowed finding consistent effects for the C allele of the SNP rs13431828 for ER visits/hospitalizations (OR=1.32, 95% CI: 1.08–1.62, adjusted p-value=0.02) and risk of exacerbations (OR=1.31, 95% CI: 1.07–1.59, adjusted p-value=0.02).
Karimi et al57 evaluated a SNP located in the FCER2 gene (rs28364072) and its relationship with FENO in European children with asthma under ICS treatment (n=593). This gene encodes CD23, the low-affinity Fc receptor for IgE which is involved in the regulation of IgE,66 and besides that, the G allele of the SNP rs28364072 has been previously associated with low-response to ICS in childhood asthma.67–69 However, Karimi et al identified the opposite effect, since carriers of the G allele had a lower concentration of FENO, suggesting a better ICS response (β for log-transformed FENO levels: −0.12, 95% CI: −0.23, −0.02, p-value=0.018). Nevertheless, when stratified analyses based on asthma control status were performed, this effect was not observed in patients with partially controlled and uncontrolled asthma.57
Some studies have evaluated the effect of genetic variants not only on ICS response but also on LTRA. Dragicevic et al58 conducted a randomized longitudinal study in European children with asthma (n=127) who were treated with ICS or LTRA and evaluated the effect of two MMP9 gene SNPs on treatment response. This gene encodes matrix metallopeptidase-9 (MMP-9), which is involved in airway remodeling and inflammation. MMP-9 expression levels are reduced in central airways and alveolar parenchyma of asthma patients compared to healthy controls.70 Carriers of the A allele of the SNP rs13925 showed an improvement in FEV1after 6 months of treatment (p-value=0.046), but the long-term effect after 12 months was not significant (p>0.05). Besides, carriers of the T allele of rs20544 demonstrated a higher increase in ACT score after 12 months (p-value=0.03).58 However, stratified analysis by pharmacological treatment did not show any genetic association either with ICS or LTRA (p-value>0.05). Thereby, the randomization of the evaluated medications limited the assessment of treatment response in this study. In addition, Wan et al59 evaluated the effect of 27 SNPs in three genes (VEGFA, TBX21, and COL2A1) previously reported to be associated with asthma in a study of Asian children with asthma (n=128). These patients were treated with ICS during the last 12 weeks and LTRA as required to prevent asthma exacerbations. Carriers of the T allele of the SNP rs3025039, located within VEGFA, had lower increments of FEV1 (p-value=0.04) and FEV1/FVC (p-value=0.004) than non-carriers after ICS treatment. VEGFA encodes the vascular endothelial growth factor A, which is implicated in both physiological and pathological angiogenesis and it has also been observed to be overexpressed in asthma patients.71 Likewise, for the SNP rs3809324 in COL2A1, carriers of the T allele showed similar outcomes, with a lower improvement of FEV1 after ICS treatment (p-value=0.048).59 COL2A1 encodes the α-1 chain of type II collagen which is a structural component of the extracellular matrix, and it has been previously associated with lung function in adulthood asthma patients under fluticasone treatment.72
Pharmacogenomic Studies of ICS Response
In 2019, two GWAS have assessed ICS response in pediatric asthma (Table 2).73,74 Hernandez-Pacheco et al73 conducted the largest meta-GWAS of ICS response to date in Hispanic/Latino and African American asthma patients (n=1,347). The definition of treatment response was the presence of asthma exacerbations during the last 12 months, considering as exacerbations ER visits, hospitalizations, or the administration of OCS because of asthma. Fifteen independent SNPs were suggestively associated with ICS response (p-value≤5 × 10−6) and one of them replicated in Europeans (n=1,697) at a nominal significance level (rs5995653, p-value=0.008). This SNP located in the APOBEC3B-APOBEC3C intergenic region demonstrated a protective effect for asthma exacerbations in a meta-analysis across Hispanics/Latinos, African Americans, and Europeans (OR for the A allele: 0.70, 95% CI: 0.61–0.81, p-value=3.31 × 10−7). The same SNP was also associated with improvement of FEV1 after ICS treatment (OR for the A allele: 2.16, 95% CI: 1.26–3.70, p-value=4.91 × 10−3). APOBEC3B and APOBEC3C encode two subunits of the apolipoprotein B mRNA‐editing catalytic polypeptide 3 (APOBEC3), which is involved in RNA editing and participate in the innate immunity against viral infections.75,76
Levin et al74 performed a GWAS in African American adults with asthma (n=244) who were under ICS treatment for 6 weeks controlling for adherence and examined the association of genetic variants with ICS response. In this case, the definition of the treatment response was measured by changes in ACT score after 6 weeks with ICS therapy and the predictors were the combined effect of the SNP and SNP x ICS treatment interaction and relevant covariates. Establishing a threshold on p-value<5.0 × 10−7, three SNPs (rs3827907, rs73906251 and 2629529) were found to be associated with asthma control status mediated by ICS treatment. Nevertheless, only EDDM3B rs3827907 remained genome-wide significant after Bonferroni correction for multiple testing (p-value=7.79 × 10−8). These three SNPs were attempted to be replicated in independent samples of African American (n=803) and European American (n=98) adults, and in two studies focused on Hispanic/Latino (n=1,461) and African American (n=563) pediatric populations. In these validation cohorts, the definition of ICS treatment response was the presence of asthma exacerbations during the last year. The rs3827907 from EDDM3B nominally replicated in the pediatric studies focused on African Americans (p-value=0.041) and Hispanics/Latinos (p-value=0.029). Furthermore, the SNP rs3827907 was found to be an eQTL associated with a decreased gene expression of RNASE2 (p-value=6.10 × 10−4) and RNASE1 (p-value=7.92 × 10−4) both in asthma and healthy individuals. Interestingly, RNASE2 encodes the eosinophil-derived neurotoxin (EDN), a protein found in eosinophil cytoplasmic granules and that has been suggested to be a biomarker of eosinophilic inflammation.77,78
Interestingly, Wang et al conducted a pharmacogenomic study of placebo response in non-Hispanic white children with asthma and attempted to evaluate the top hits on patients under pharmacological treatment (budesonide or nedocromil).79 A significant interaction was found between the SNP rs2392165 near BBS9 with ICS use on wheezing/coughing (p-valueinteraction= 1.48 × 10−7), being associated with better ICS response (β for the G allele: −0.53, p-value=0.02). BBS9 is implicated in lung development and ciliary function.79
Discussion
Between 2018 and 2019, the pharmacogenetic studies conducted in pediatric asthma have been mainly focused on therapeutic response to ICS and SABA albeit some of them have studied simultaneously the response to ICS and LABA/LTRA. Altogether, eleven candidate-gene studies,24–28,55–59 a meta-analysis of a candidate gene,60 and six pharmacogenomic studies (one of them in adults but including replication in children) have been performed.32,36,40,73,74,79
During this period, candidate-gene studies reported the association of genes previously described in pharmacogenetic studies of pediatric asthma with treatment response (ADRB2, GSDMB, FCER2, and VEGFA).13,16-18 Additionally, other candidate genes have been related for the first time to this phenotype in children (SPATS2L, ASB3, IL1RL1, MMP9, and COL2A1). Although several findings support previous evidence of some of these genes in asthma treatment response, inconsistent results across the literature or even across the studies included in this review have been found. For instance, although ADRB2 is one of the most studied candidate genes so far, its role in asthma treatment response is still not clear.23 In fact, conflicting results were reported for variants of this gene within the period reviewed, which could be due to the small sample sizes (48±27 individuals), differences in statistical approaches, and the lack of multiple comparison adjustments. Moreover, most of the studies of ADRB2 have focused on the same polymorphisms within this gene.24–27,54 Recently, GWAS have strongly associated different SNPs within some of these candidate genes (ADRB2, IL1RL1, and FCER2) with related phenotypes such as lung function, suggesting that future studies could focus on other genetic variants that might have a role in treatment response.80–83
The application of pharmacogenomic approaches without a prior hypothesis has provided new insights, enabling researchers to discover novel associations of genetic variants across the whole genome with the treatment response in pediatric asthma (PRKG1, DNAH5, CRISPLD2, APOBEC3B-APOBEC3C, EDDM3B, and BBS9). Moreover, the relationship of these genes with asthma treatment response may be plausible since other studies have also reported them with asthma-related phenotypes, such as lung function (PRKG1,82 DNAH5,83 and BBS984), allergic sensitization (DNAH538), asthma susceptibility (PRKG185), and childhood-onset asthma (CRISPLD286). Nonetheless, the replication of pharmacogenetic findings is still scarce. Hernandez-Pacheco et al did not find any consistent association of 22 SNPs associated with treatment response by previous GWAS.73 Similarly, Spear et al and Mak et al failed to replicate the association of SNPs associated with BDR by previous candidate-gene association studies or GWAS.32,36 Differences in the statistical power of candidate-gene studies and GWAS and the analysis of different racial/ethnic groups could explain the inconsistent results among studies.
It should also be noted that albeit childhood-onset and adult-onset asthma have been identified as different phenotypes and usually do not share the same genetic components, some similarities have been identified.62 Interestingly, a GWAS reported the association of a SNP within EDDM3B with ICS response in adults as discovery, and this finding was replicated in childhood asthma.74 Since this gene is related to eosinophilic inflammation,74,77,78 this feature may be common in both phenotypes and may condition treatment response to ICS in both age groups. Moreover, some of the candidate genes associated during this period with treatment response in childhood asthma had been previously reported with ICS response in asthma adult patients (COL2A172) and with bronchodilator response both in children and adults (ASB330 and SPAT2SL29). On the other hand, two SNPs from GLCCI1, a gene extensively studied in pediatric pharmacogenetic studies of asthma,87 were associated with ICS response in adult asthma as well.88,89 Nevertheless, the effect of the SNPs within ASB3 and SPAT2SL on SABA response was found to be mitigated by aging.28 Therefore, it is important to continue investigating whether the genetic component of asthma treatment response is shared between adults and children.
Pharmacogenetic studies are aimed to establish genetic markers that may be applied to the clinical practice to improve asthma management. The implementation of genetic variants described in pharmacogenetic studies, such as the SNP rs295137 near SPATS2L, into clinical predictive models of ER management failure in children with moderate-to-severe asthma has demonstrated to improve their sensibility and specificity, compared to models incorporating only clinical information.90 Nevertheless, given the discrepancies found in the literature, the small effect size of polymorphisms associated with asthma treatment response, and the lack of replication across populations, the clinical relevance of genetic variants remains unreliable.
To improve future pharmacogenetic studies, standardized methods and further analyses are required. Notably, the studies conducted in 2018 and 2019 focused on different populations, had heterogeneous eligibility criteria (Table S2) and analyzed different outcomes that may reflect different underlying mechanisms of treatment response. Studies focused on SABA response used lung function measurements (mainly BDR) as a treatment response measure. Nonetheless, most studies that assessed ICS response analyzed asthma exacerbations as the main outcome (46% of the studies), but they also examined other measurements of treatment response such as lung function measurements (27%), FENO (9%), ACT score (9%), or self-reported asthma symptoms (9%). Since asthma is a multifactorial disease, different phenotypes with different underlying etiology may not share the same genetic architecture and the effect of genetic variants may even be conditioned by ethnicity. Moreover, some limitations are shared among the assessed studies, especially in those focused on ICS response: the analysis of self-reported exacerbations as the main outcome, the concomitant use of other medications, or the lack of information about the type of ICS and dosage. To overcome the limitations, studies could analyze cohorts that represent more homogeneous populations in terms of asthma phenotype and demographic features, but trying to reach the largest sample sizes as well. Furthermore, collaboration among researchers is needed to validate the reported results and to evaluate them in different populations. The establishment of the Pharmacogenomics in Childhood Asthma (PiCA) consortium has allowed in the last years performing multiethnic pharmacogenetic analyses in pediatric populations.91 Moreover, the integration of clinical data with the knowledge coming from multiple omics will help to understand in detail how genetic variants are implicated in different pathways that condition treatment response in asthma patients.
In conclusion, pharmacogenetic studies of pediatric asthma have recently moved from candidate-gene approaches to the study of the whole genome variation and have improved the representation of different racial/ethnic groups. However, stronger evidence is needed to move forward towards the application of genetic markers in clinical practice. Once genetic variants were established as biomarkers of treatment response, precision medicine based on pharmacogenetics would be feasible in clinical practice. This will improve the safety and effectiveness of asthma therapy and will avoid using a specific medication in potentially non-responder patients, diminishing not only the costs associated with the treatment but also potential side-effects.
Author Contributions
All the authors were involved in the conception and design of the review, the acquisition and interpretation of the data, the drafting or revision of the manuscript, and the approval for the final version. All the authors agree to be accountable for all the aspects of the work.
Funding
Supported by grant SAF2017-83417R by the Spanish Ministry of Economy, Industry, and Competitiveness, the State Research Agency, and the European Regional Development Funds from the European Union (MINECO/AEI/FEDER, UE) to MP-Y and FL-D. AE-O received salary support from grant SAF2017-83417R by the Spanish Ministry of Economy, Industry, and Competitiveness. MP-Y was also supported by the Ramón y Cajal Program by the Spanish Ministry of Economy, Industry, and Competitiveness (RYC-2015-17205).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Global Asthma Network. The Global Asthma Report 2018. Auckland, New Zealand; 2018.
2. Global Inititative for Asthma. Global Strategy for Asthma Management and Prevention; 2019.
3. Asher I, Pearce N. Global burden of asthma among children. Int J Tuberc Lung Dis. 2014;18(11):1269–1278. doi:10.5588/ijtld.14.0170
4. Mallol J, Crane J, von Mutius E, Odhiambo J, Keil U, Stewart A. The international study of asthma and allergies in childhood (ISAAC) Phase three: a global synthesis. Allergol Immunopathol (Madr). 2013;41(2):73–85. doi:10.1016/j.aller.2012.03.001
5. Royal College of Physicians. Why Asthma Still Kills: The National Review of Asthma Deaths (NRAD); 2014.
6. Wenzel SE. Asthma phenotypes: the evolution from clinical to molecular approaches. Nat Med. 2012;18(5):716–725. doi:10.1038/nm.2678
7. Anderson GP. Endotyping asthma: new insights into key pathogenic mechanisms in a complex, heterogeneous disease. Lancet. 2008;372(9643):1107–1119. doi:10.1016/S0140-6736(08)61452-X
8. Dharmage SC, Perret JL, Custovic A. Epidemiology of asthma in children and adults. Front Pediatr. 2019;7:246. doi:10.3389/fped.2019.00246
9. Park H-W, Tantisira KG, Weiss ST. Pharmacogenomics in asthma therapy: where are we and where do we go? Annu Rev Pharmacol Toxicol. 2015;55(1):129–147. doi:10.1146/annurev-pharmtox-010814-124543
10. Hardy J, Singleton A. Genomewide association studies and human disease. N Engl J Med. 2009;360(17):1759–1768. doi:10.1056/NEJMra0808700
11. Kwon JM, Goate AM. The candidate gene approach. Alcohol Res Heal. 2000;24(3):164–168.
12. Campbell CD, Mohajeri K, Malig M, et al. Whole-genome sequencing of individuals from a founder population identifies candidate genes for asthma. PLoS One. 2014;9(8):e104396. doi:10.1371/journal.pone.0104396
13. Keskin O, Farzan N, Birben E, et al. Genetic associations of the response to inhaled corticosteroids in asthma: a systematic review. Clin Transl Allergy. 2019;9:2. doi:10.1186/s13601-018-0239-2
14. Hernandez-Pacheco N, Pino-Yanes M, Flores C. Genomic predictors of asthma phenotypes and treatment response. Front Pediatr. 2019;7:6. doi:10.3389/fped.2019.00006
15. Slob EMA, Vijverberg SJH, Palmer CNA, et al. Pharmacogenetics of inhaled long-acting beta2-agonists in asthma: a systematic review. Pediatr Allergy Immunol. 2018;29(7):705–714. doi:10.1111/pai.12956
16. Herrera-Luis E, Hernandez-Pacheco N, Vijverberg SJ, Flores C, Pino-Yanes M. Role of genomics in asthma exacerbations. Curr Opin Pulm Med. 2019;25(1):101–112. doi:10.1097/MCP.0000000000000533
17. Farzan N, Vijverberg SJ, Kabesch M, Sterk PJ, Maitland-van der Zee AH. The use of pharmacogenomics, epigenomics, and transcriptomics to improve childhood asthma management: where do we stand? Pediatr Pulmonol. 2018;53(6):836–845. doi:10.1002/ppul.v53.6
18. García-Menaya JM, Cordobés-Durán C, García-Martín E, Agúndez JAG. Pharmacogenetic factors affecting asthma treatment response. potential implications for drug therapy. Front Pharmacol. 2019;10:520. doi:10.3389/fphar.2019.00520
19. Johnson M. Beta2-adrenoceptors: mechanisms of action of beta2-agonists. Paediatr Respir Rev. 2001;2(1):57–62. doi:10.1053/prrv.2000.0102
20. Drake KA, Torgerson DG, Gignoux CR, et al. A genome-wide association study of bronchodilator response in Latinos implicates rare variants. J Allergy Clin Immunol. 2014;133(2):370–378. doi:10.1016/j.jaci.2013.06.043
21. Duan QL, Lasky-Su J, Himes BE, et al. A genome-wide association study of bronchodilator response in asthmatics. Pharmacogenomics J. 2014;14(1):41–47. doi:10.1038/tpj.2013.5
22. Pellegrino R, Viegi G, Brusasco V, et al. Interpretative strategies for lung function tests. Eur Respir J. 2005;26(5):948–968. doi:10.1183/09031936.05.00035205
23. Litonjua AA, Gong L, Duan QL, et al. Very important pharmacogene summary ADRB2. Pharmacogenet Genomics. 2010;20(1):64–69. doi:10.1097/FPC.0b013e328333dae6
24. Scaparrotta A, Franzago M, Marcovecchio ML, et al. Role of THRB, ARG1, and ADRB2 genetic variants on bronchodilators response in asthmatic children. J Aerosol Med Pulm Drug Deliv. 2019;32(3):164–173. doi:10.1089/jamp.2018.1493
25. Jovicic N, Babic T, Dragicevic S, Nestorovic B, Nikolic A. ADRB2 gene polymorphisms and salbutamol responsiveness in Serbian children with asthma. Balk J Med Genet. 2018;21(1):33–38. doi:10.2478/bjmg-2018-0007
26. Alghobashy AA, Elsharawy SA, Alkholy UM, et al. B 2 adrenergic receptor gene polymorphism effect on childhood asthma severity and response to treatment. Pediatr Res. 2018;83(3):597–605. doi:10.1038/pr.2017.304
27. Toraih EA, Hussein MH, Ibrahim A, et al. Beta2-adrenergic receptor variants in children and adolescents with bronchial asthma. Front Biosci (Elite Ed). 2019;11:61–78.
28. Sordillo JE, McGeachie M, Lutz SM, et al. Longitudinal analysis of bronchodilator response in asthmatics and effect modification of age-related trends by genotype. Pediatr Pulmonol. 2019;54(2):158–164. doi:10.1002/ppul.v54.2
29. Himes BE, Jiang X, Hu R, et al. Genome-wide association analysis in asthma subjects identifies SPATS2L as a novel bronchodilator response gene. PLoS Genet. 2012;8(7):e1002824. doi:10.1371/journal.pgen.1002824
30. Israel E, Lasky-Su J, Markezich A, et al. Genome-wide association study of short-acting β2-agonists. A novel genome-wide significant locus on chromosome 2 near ASB3. Am J Respir Crit Care Med. 2015;191(5):530–537. doi:10.1164/rccm.201408-1426OC
31. McDaneld TG, Hancock DL, Moody DE. Altered mRNA abundance of ASB15 and four other genes in skeletal muscle following administration of β-adrenergic receptor agonists. Physiol Genomics. 2004;16(2):275–283. doi:10.1152/physiolgenomics.00127.2003
32. Spear ML, Hu D, Pino-Yanes M, et al. A genome-wide association and admixture mapping study of bronchodilator drug response in African Americans with asthma. Pharmacogenomics J. 2019;19(3):249–259. doi:10.1038/s41397-018-0042-4
33. Howie BN, Donnelly P, Marchini J. A flexible and accurate genotype imputation method for the next generation of genome-wide association studies. PLoS Genet. 2009;5(6):e1000529. doi:10.1371/journal.pgen.1000529
34. Ricciardolo FLM. Multiple roles of nitric oxide in the airways. Thorax. 2003;58(2):175–182. doi:10.1136/thorax.58.2.175
35. Kanazawa H, Hirata K, Yoshikawa J. Nitrogen oxides reduce albuterol-induced bronchodilation in patients with bronchial asthma. Respiration. 2002;69(6):490–495. doi:10.1159/000066466
36. Mak ACY, White MJ, Eckalbar WL, et al. Whole-Genome sequencing of pharmacogenetic drug response in racially diverse children with asthma. Am J Respir Crit Care Med. 2018;197(12):1552–1564. doi:10.1164/rccm.201712-2529OC
37. Lee JH, McDonald ML, Cho MH, et al. DNAH5 is associated with total lung capacity in chronic obstructive pulmonary disease. Respir Res. 2014;15:97. doi:10.1186/s12931-014-0097-y
38. Ramasamy A, Curjuric I, Coin LJ, et al. A genome-wide meta-analysis of genetic variants associated with allergic rhinitis and grass sensitization and their interaction with birth order. J Allergy Clin Immunol. 2011;128(5):996–1005. doi:10.1016/j.jaci.2011.08.030
39. Hersch M, Peter B, Kang HM, et al. Mapping genetic variants associated with beta-adrenergic responses in inbred mice. PLoS One. 2012;7(7):e41032. doi:10.1371/journal.pone.0041032
40. Kachroo P, Hecker J, Chawes B, et al. Whole genome sequencing identifies CRISPLD2 as a lung function gene in children with asthma. Chest. 2019;156(6):1068–1079. doi:10.1016/j.chest.2019.08.2202
41. Zhang H, Kho AT, Wu Q, et al. CRISPLD2 (LGL1) inhibits proinflammatory mediators in human fetal, adult, and COPD lung fibroblasts and epithelial cells. Physiol Rep. 2016;4(17):e12942. doi:10.14814/phy2.12942
42. Zhang H, Sweezey NB, Kaplan F. LGL1 modulates proliferation, apoptosis, and migration of human fetal lung fibroblasts. Am J Physiol Lung Cell Mol Physiol. 2015;308(4):L391–L402. doi:10.1152/ajplung.00119.2014
43. Himes BE, Jiang X, Wagner P, et al. RNA-Seq transcriptome profiling identifies CRISPLD2 as a glucocorticoid responsive gene that modulates cytokine function in airway smooth muscle cells. PLoS One. 2014;9(6):e99625. doi:10.1371/journal.pone.0099625
44. Yan D, Hamed O, Joshi T, et al. Analysis of the indacaterol-regulated transcriptome in human airway epithelial cells implicates gene expression changes in the adverse and therapeutic effects of β2-adrenoceptor agonists. J Pharmacol Exp Ther. 2018;366(1):220–236. doi:10.1124/jpet.118.249292
45. Keenan CR, Radojicic D, Li M, Radwan A, Stewart AG. Heterogeneity in mechanisms influencing glucocorticoid sensitivity: the need for a systems biology approach to treatment of glucocorticoid-resistant inflammation. Pharmacol Ther. 2015;150:81–93. doi:10.1016/j.pharmthera.2015.01.006
46. Lavorini F, Magnan A, Christophe Dubus J, et al. Effect of incorrect use of dry powder inhalers on management of patients with asthma and COPD. Respir Med. 2008;102(4):593–604. doi:10.1016/j.rmed.2007.11.003
47. Laforest L, Belhassen M, Devouassoux G, Didier A, Ginoux M, Van Ganse E. Long-term inhaled corticosteroid adherence in asthma patients with short-term adherence. J Allergy Clin Immunol Pract. 2016;4(5):890–899.e2. doi:10.1016/j.jaip.2016.07.008
48. Ramratnam SK, Bacharier LB, Guilbert TW. Severe asthma in children. J Allergy Clin Immunol Pract. 2017;5(4):889–898. doi:10.1016/j.jaip.2017.04.031
49. Huang YJ, Nariya S, Harris JM, et al. The airway microbiome in patients with severe asthma: associations with disease features and severity. J Allergy Clin Immunol. 2015;136(4):874–884. doi:10.1016/j.jaci.2015.05.044
50. Durack J, Lynch SV, Nariya S, et al. Features of the bronchial bacterial microbiome associated with atopy, asthma, and responsiveness to inhaled corticosteroid treatment. J Allergy Clin Immunol. 2017;140(1):63–75. doi:10.1016/j.jaci.2016.08.055
51. Denner DR, Sangwan N, Becker JB, et al. Corticosteroid therapy and airflow obstruction influence the bronchial microbiome, which is distinct from that of bronchoalveolar lavage in asthmatic airways. J Allergy Clin Immunol. 2016;137(5):1398–1405.e3. doi:10.1016/j.jaci.2015.10.017
52. Goleva E, Jackson LP, Harris JK, et al. The effects of airway microbiome on corticosteroid responsiveness in asthma. Am J Respir Crit Care Med. 2013;188(10):1193–1201. doi:10.1164/rccm.201304-0775OC
53. Licari A, Brambilla I, Marseglia A, De Filippo M, Paganelli V, Marseglia GL. Difficult vs. severe asthma: definition and limits of asthma control in the pediatric population. Front Pediatr. 2018;6:170. doi:10.3389/fped.2018.00170
54. Sood N, Connolly JJ, Mentch FD, et al. Leveraging electronic health records to assess the role of ADRB2 single nucleotide polymorphisms in predicting exacerbation frequency in asthma patients. Pharmacogenet Genomics. 2018;28(11):256–259. doi:10.1097/FPC.0000000000000355
55. Farzan N, Vijverberg SJ, Hernandez-Pacheco N, et al. 17q21 variant increases the risk of exacerbations in asthmatic children despite inhaled corticosteroids use. Allergy Eur J Allergy Clin Immunol. 2018;73(10):2083–2088. doi:10.1111/all.2018.73.issue-10
56. Dijk FN, Vijverberg SJ, Hernandez‐Pacheco N, et al. IL1RL1 gene variations are associated with asthma exacerbations in children and adolescents using inhaled corticosteroids. Allergy. 2019. doi:10.1111/all.14125
57. Karimi L, Vijverberg SJH, Farzan N, Ghanbari M, Verhamme KMC, Maitland‐van der Zee AH. FCER2 T2206C variant associated with FENO levels in asthmatic children using inhaled corticosteroids: the PACMAN study Clin Exp Allergy. 2019;49(11):1429–1436. doi:10.1111/cea.13460
58. Dragicevic S, Kosnik M, Divac Rankov A, et al. The variants in the 3′ untranslated region of the matrix metalloproteinase 9 gene as modulators of treatment outcome in children with asthma. Lung. 2018;196(3):297–303. doi:10.1007/s00408-018-0113-y
59. Wan Z, Tang Y, Song Q, et al. Gene polymorphisms in VEGFA and COL2A1 are associated with response to inhaled corticosteroids in children with asthma. Pharmacogenomics. 2019;20(13):947–955. doi:10.2217/pgs-2019-0036
60. Wang X, Li Q, Liu R, et al. ADRB2 Arg16Gly polymorphism and pulmonary function response of inhaled corticosteroids plus long-acting beta agonists for asthma treatment: a systematic review and meta-analysis. Can Respir J. 2018;2018:5712805.
61. Moffatt MF, Gut IG, Demenais F, et al. A large-scale, consortium-based genomewide association study of asthma. N Engl J Med. 2010;363(13):1211–1221. doi:10.1056/NEJMoa0906312
62. Ferreira MAR, Mathur R, Vonk JM, et al. Genetic architectures of childhood- and adult-onset asthma are partly distinct. Am J Hum Genet. 2019;104(4):665–684. doi:10.1016/j.ajhg.2019.02.022
63. Berce V, Kozmus CEP, Potočnik U. Association among ORMDL3 gene expression, 17q21 polymorphism and response to treatment with inhaled corticosteroids in children with asthma. Pharmacogenomics J. 2013;13(6):523–529. doi:10.1038/tpj.2012.36
64. Vicente CT, Revez JA, Ferreira MAR. Lessons from ten years of genome-wide association studies of asthma. Clin Transl Immunol. 2017;6(12):e165. doi:10.1038/cti.2017.54
65. Grotenboer NS, Ketelaar ME, Koppelman GH, Nawijn MC. Decoding asthma: translating genetic variation in IL33 and IL1RL1 into disease pathophysiology. J Allergy Clin Immunol. 2013;131(3):856–865. doi:10.1016/j.jaci.2012.11.028
66. Chan MA, Gigliotti NM, Aubin BG, Rosenwasser LJ. FCER2 (CD23) asthma-related SNP yields increased IgE binding and Egr-1 expression in human B cells. Am J Respir Cell Mol Biol. 2013;50(2):263–269.
67. Rogers AJ, Tantisira KG, Fuhlbrigge AL, et al. Predictors of poor response during asthma therapy differ with definition of outcome. Pharmacogenomics. 2009;10(8):1231–1242. doi:10.2217/pgs.09.86
68. Tantisira KG, Silverman ES, Mariani TJ, et al. FCER2: a pharmacogenetic basis for severe exacerbations in children with asthma. J Allergy Clin Immunol. 2007;120(6):1285–1291. doi:10.1016/j.jaci.2007.09.005
69. Koster ES, Maitland-van Der Zee AH, Tavendale R, et al. FCER2 T2206C variant associated with chronic symptoms and exacerbations in steroid-treated asthmatic children. Allergy Eur J Allergy Clin Immunol. 2011;66(12):1546–1552. doi:10.1111/j.1398-9995.2011.02701.x
70. Weitoft M, Andersson C, Andersson-Sjöland A, et al. Controlled and uncontrolled asthma display distinct alveolar tissue matrix compositions. Respir Res. 2014;15:67. doi:10.1186/1465-9921-15-67
71. Lee CG, Link H, Baluk P, et al. Vascular endothelial growth factor (VEGF) induces remodeling and enhances TH2-mediated sensitization and inflammation in the lung. Nat Med. 2004;10(10):1095–1103. doi:10.1038/nm1105
72. Mougey EB, Chen C, Tantisira KG, et al. Pharmacogenetics of asthma controller treatment. Pharmacogenomics J. 2013;13(3):242–250. doi:10.1038/tpj.2012.5
73. Hernandez-Pacheco N, Farzan N, Francis B, et al. Genome-wide association study of inhaled corticosteroid response in admixed children with asthma. Clin Exp Allergy. 2019;49(6):789–798. doi:10.1111/cea.13354
74. Levin AM, Gui H, Hernandez-Pacheco N, et al. Integrative approach identifies corticosteroid response variant in diverse populations with asthma. J Allergy Clin Immunol. 2019;143(5):1791–1802. doi:10.1016/j.jaci.2018.09.034
75. Janahi EM, McGarvey MJ. The inhibition of hepatitis B virus by APOBEC cytidine deaminases. J Viral Hepat. 2013;20(12):821–828. doi:10.1111/jvh.2013.20.issue-12
76. Desimmie BA, Delviks-Frankenberrry KA, Burdick RC, Qi D, Izumi T, Pathak VK. Multiple APOBEC3 restriction factors for HIV-1 and one vif to rule them all. J Mol Biol. 2014;426(6):1220–1245. doi:10.1016/j.jmb.2013.10.033
77. Walsh GM. Eosinophil granule proteins and their role in disease. Curr Opin Hematol. 2001;8(1):28–33. doi:10.1097/00062752-200101000-00006
78. Gon Y, Ito R, Hattori T, et al. Serum eosinophil-derived neurotoxin: correlation with persistent airflow limitation in adults with house-dust mite allergic asthma. Allergy Asthma Proc. 2015;36(6):e113–e120. doi:10.2500/aap.2015.36.3884
79. Wang RS, Croteau-Chonka DC, Silverman EK, Loscalzo J, Weiss ST, Hall KT. Pharmacogenomics and Placebo response in a randomized clinical trial in asthma. Clin Pharmacol Ther. 2019;106(6):1261–1267. doi:10.1002/cpt.1646
80. Wain LV, Shrine N, Artigas MS, et al. Genome-wide association analyses for lung function and chronic obstructive pulmonary disease identify new loci and potential druggable targets. Nat Genet. 2017;49(3):416–425. doi:10.1038/ng.3787
81. Shrine N, Guyatt AL, Erzurumluoglu AM, et al. New genetic signals for lung function highlight pathways and chronic obstructive pulmonary disease associations across multiple ancestries. Nat Genet. 2019;51(3):481–493. doi:10.1038/s41588-018-0321-7
82. Kichaev G, Bhatia G, Loh PR, et al. Leveraging polygenic functional enrichment to improve GWAS power. Am J Hum Genet. 2019;104(1):65–75. doi:10.1016/j.ajhg.2018.11.008
83. Lutz SM, Cho MH, Young K, et al. A genome-wide association study identifies risk loci for spirometric measures among smokers of European and African ancestry. BMC Genet. 2015;16:138. doi:10.1186/s12863-015-0299-4
84. Wain LV, Shrine N, Miller S, et al. Novel insights into the genetics of smoking behaviour, lung function, and chronic obstructive pulmonary disease (UK BiLEVE): A genetic association study in UK Biobank. Lancet Respir Med. 2015;3(10):769–781. doi:10.1016/S2213-2600(15)00283-0
85. Ferreira MAR, Matheson MC, Duffy DL, et al. Identification of IL6R and chromosome 11q13.5 as risk loci for asthma. Lancet. 2011;378(9795):1006–1014. doi:10.1016/S0140-6736(11)60874-X
86. Almoguera B, Vazquez L, Mentch F, et al. Identification of four novel loci in asthma in European American and African American Populations. Am J Respir Crit Care Med. 2017;195(4):456–463. doi:10.1164/rccm.201604-0861OC
87. Tantisira KG, Lasky-Su J, Harada M, et al. Genomewide association between GLCCI1 and response to glucocorticoid therapy in asthma. N Engl J Med. 2011;365(13):1173–1183. doi:10.1056/NEJMoa0911353
88. Salhi M, Lahmar O, Salah MO. et al. GLCCI1 and STIP1 variants are associated with asthma susceptibility and inhaled corticosteroid response in a Tunisian population. J Asthma;2019. 1–10. doi:10.1080/02770903.2019.1666867
89. Rijavec M, Žavbi M, Lopert A, Fležar M, Korošec P. GLCCI1 polymorphism rs37973 and response to treatment of asthma with inhaled corticosteroids. J Investig Allergol Clin Immunol. 2018;28(3):165–171. doi:10.18176/jiaci
90. Tse SM, Krajinovic M, Chauhan BF, et al. Genetic determinants of acute asthma therapy response in children with moderate-to-severe asthma exacerbations. Pediatr Pulmonol. 2019;54(4):378–385. doi:10.1002/ppul.v54.4
91. Farzan N, Vijverberg SJ, Andiappan AK, et al. Rationale and design of the multiethnic pharmacogenomics in childhood asthma consortium. Pharmacogenomics. 2017;18(10):931–943. doi:10.2217/pgs-2017-0035
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.