Back to Journals » Journal of Inflammation Research » Volume 19
PD-L1 as a Potential Inducer for NF-κB Pathway Activation in M1-Type Macrophages During Sepsis: An Integrated Analysis Based on GEO Public Database and Multi-Omics Data
Authors Yuan M ![]() , Jin T, Zhao Q, Tian L
, Jin T, Zhao Q, Tian L ![]()
Received 7 November 2025
Accepted for publication 3 January 2026
Published 9 January 2026 Volume 2026:19 578828
DOI https://doi.org/10.2147/JIR.S578828
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Xin Du
Mu Yuan,* Tong Jin,* Qing Zhao, Lixing Tian
Department of Stem Cell & Regenerative Medicine, State Key Laboratory of Trauma and Chemical Poisoning, Daping Hospital, Army Medical University, Chongqing, 400042, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Lixing Tian, Department of Stem Cell & Regenerative Medicine, State Key Laboratory of Trauma and Chemical Poisoning, Daping Hospital, Army Medical University, Yuzhong District, Chongqing, 400042, People’s Republic of China, Tel +8618584566173, Email [email protected]
Background: M1-type macrophages are a crucial defense line against pathogens during sepsis, and their polarization depends on NF-κB pathway activation. However, the specific molecular regulatory mechanisms of this process are not fully elucidated. Programmed death-ligand 1 (PD-L1), an important immune checkpoint molecule, is often anchored on the cell membrane exerting immunosuppressive effects, but its role in M1-type macrophages remains poorly understood.
Methods: In this study, we preliminarily investigated the potential regulatory effect of PD-L1 on the NF-κB pathway in M1 macrophages by integrating GEO sepsis patient sequencing datasets, RNA-seq data from PD-L1-overexpressing M1 macrophages, immunoprecipitation-mass spectrometry (IP-MS), molecular docking, and co-immunoprecipitation (Co-IP). The Receiver Operating Characteristic (ROC) curve of PD-L1 was used as a prognostic indicator for sepsis. PD-L1 knockout mice were utilized to validate the in vivo regulatory effects of PD-L1.
Results: Analysis of GEO data and peripheral blood mononuclear cells (PBMCs) from sepsis patients revealed significantly elevated PD-L1 expression with a central position among all upregulated genes. Furthermore, this elevated PD-L1 expression was validated in peripheral monocytes from sepsis patients compared to healthy controls. Single-gene GSEA analysis based on GEO data indicated a close association between PD-L1 and NF-κB pathway activation in sepsis patients. RNA-seq analysis of PD-L1-overexpressing M1 macrophages confirmed that PD-L1 significantly activates the NF-κB pathway. IP-MS screening and Co-IP validation identified an interaction between PD-L1 and KLF6, a nuclear factor closely related to the NF-κB pathway. Finally, liver transcriptome data from septic mice confirmed that PD-L1 can also activate the NF-κB pathway in vivo, and ROC curve analysis demonstrated PD-L1’s potential as a prognostic indicator for sepsis.
Conclusion: Based on these results, this study preliminarily reveals a potential activating role of PD-L1 on the NF-κB pathway in M1 macrophages.
Keywords: macrophage, NF-κB signaling pathway, PD-L1
Introduction
Macrophages are vital components of the immune system, belonging to the mononuclear phagocyte system. They exhibit high plasticity and can differentiate into various phenotypes under microenvironmental stimuli to participate in host defense and tissue homeostasis.1,2 M1 polarization is one of their key activated states, often induced by interferon-γ (IFN-γ), lipopolysaccharide (LPS), or TNF-α, characterized by strong pro-inflammatory properties.2,3 This polarization enables macrophages to secrete cytokines like IL-12 and TNF-α, promoting antigen presentation and T cell activation, playing a central role in rapid pathogen clearance and initiating immune defense during early sepsis.1,4,5 However, excessive macrophage polarization can lead to tissue damage or chronic inflammation, making its dynamic regulation crucial for disease treatment and immune balance. The pro-inflammatory function of M1 macrophages is highly dependent on NF-κB pathway activation. This pathway is primarily initiated through Toll-like receptors (eg, TLR4) or cytokine receptors, involving the MyD88-dependent pathway, leading to IκB kinase (IKK) phosphorylation and NF-κB nuclear translocation, subsequently transcribing inflammatory factors (eg, TNF-α, IL-6, iNOS) to drive pathogen clearance and tissue damage.6,7 NF-κB synergizes with transcription factors like interferon regulatory factors (IRFs) to reinforce M1 polarization and maintain inflammatory homeostasis.8,9 However, the upstream signal integration, epigenetic regulation, and interaction with other polarization pathways (eg, JAK/STAT) during its activation are not fully understood.
The programmed death-1 (PD-1)/PD-L1 axis is a well-established negative regulatory pathway for immune homeostasis. PD-L1 is a type I transmembrane glycoprotein primarily expressed on the cell surface of tumor cells, vascular endothelial cells, dendritic cells, and macrophages.10 Its interaction with PD-1 on T cells leads to dephosphorylation of the T cell receptor and co-stimulatory receptor CD28, thereby inhibiting T cell proliferation and activation, ultimately causing T cell exhaustion and an immunosuppressive state. However, numerous studies indicate that immunosuppressive PD-L1 is highly expressed in inflammatory states. Literature reports show significantly high PD-L1 expression on monocytes in sepsis patients, and this marker is an independent risk factor for 28-day mortality.11–14 Furthermore, studies in PD-L1 knockout mice subjected to cecal ligation and puncture (CLP) or hemorrhage+CLP models showed significantly higher survival rates compared to wild-type mice, suggesting PD-L1 may promote the pathological process of sepsis.15,16 Although these studies point to an association between PD-L1 and inflammatory response, the specific mechanisms remain unelucidated, particularly the relationship between PD-L1 and the NF-κB pathway—a key pathway determining polarization—has not been investigated.
In this study, we investigated PD-L1 expression using large-sample GEO datasets and PBMCs from sepsis patients. These findings were further validated in peripheral monocytes from both sepsis patients and healthy controls. Through PD-L1 single-gene GSEA analysis and RNA-seq data, we found a significant association between PD-L1 and NF-κB pathway activation. KLF6 functions as a potent coactivator within the NF-κB signaling pathway. While not directly initiating inflammation, it critically fine-tunes the transcriptional output by enhancing the activity of the p65 subunit, ultimately amplifying the expression of a subset of downstream genes. This regulatory role is particularly vital in the dysregulated immune response characteristic of sepsis.17 Unexpectedly, our preliminary IP-MS screening revealed that PD-L1 can interact with KLF6, further indicating that PD-L1 exerts a positive regulatory effect on the NF-κB pathway. Additionally, ROC curve analysis for sepsis patients and liver RNA-seq data from septic mice indicated that PD-L1 positively regulates the pathological process of sepsis in vivo. In summary, this study preliminarily reveals, from a macro perspective using large-sample, multi-omics data, the regulatory role of PD-L1 on the NF-κB pathway.
Materials and Methods
Ethics Statement
The animal experimental protocol was approved by the Laboratory Animal Welfare and Ethics Committee of the Army Medical University (Approval No. AWUWEC20247118). All procedures and animal welfare followed the Army Medical University Ethical Guidelines. The protocol for obtaining samples from sepsis patients and healthy volunteers was approved by the Ethics Committee of the Army Medical Center of PLA (Approval No. 2025318), and informed consent was obtained from participants or their legal guardians. All datasets used in this study were obtained from the public NCBI database and are openly accessible (all involving non-interventional studies). Since the original studies associated with these datasets had obtained ethical approval, this study is exempt from ethical review according to institutional regulations.
GEO Data Acquisition and Processing
The GEO datasets used in this study were obtained from the publicly available NCBI database. Datasets GSE46955, GSE69063, and GSE121349 were used for comparing PD-L1 expression between healthy and sepsis groups and for screening core sepsis genes. Complete MINiML format data including all platforms, samples, and GSE records were downloaded. For unnormalized datasets, log2 transformation was uniformly applied. Data standardization was performed using the normalize.quantiles function from the R preprocessCore package. Probe IDs were converted to gene symbols based on corresponding platform annotation information, probes corresponding to multiple genes were excluded, and average values for genes corresponding to multiple probes were calculated. For different batches within the same dataset and platform, batch effects were removed using the removeBatchEffect function from the R limma package. During integrated analysis of different datasets or platforms, common gene symbols across datasets were first extracted, then different datasets or platforms were marked as different batches, and the removeBatchEffectfunction was similarly used to remove batch effects. Standardization was assessed via boxplots; batch effect removal was evaluated by comparing PCA plots before and after removal. The Wilcoxon test was used for statistical differences between groups. Core gene screening was performed using the cytoHubba plugin in Cytoscape software (v3.10.1). The maximum clique centrality (MCC) algorithm was used to identify central nodes within maximum cliques and generate a centrality ranking list. The top 11 genes based on MCC scores were selected for visualization. Dataset GSE22248 was used to analyze PD-L1 expression in human PBMCs upon LPS stimulation, processed as above. Patient grouping and accession numbers for the used datasets are detailed in Supplemental Table 1, facilitating the lookup of details like age, sex, and disease status for each patient on NCBI.
Single-Gene GSEA Analysis
The integrated, batch-effect-removed GEO datasets were imported into R software (v2024.04.1) for analysis. The specific analysis method is described in reference.18 The code used is provided in Supplemental Method.
Reagents
LPS (Escherichia coli 0111:B4) and thioglycolate were from Sigma‒Aldrich (St. Louis, MO, USA). Recombinant Human IFN-γ (11725-HNAE) was from Sinobiological (Beijing, China). Trypsin, fetal bovine serum (FBS), RPMI 1640 medium, and penicillin‒streptomycin solution were from Gibco (Carlsbad, CA, USA). Anti-GAPDH antibody (#5174), anti-PD-L1 antibody (#13684), and anti-IgG antibody (#3900) were from CST (Danvers, MA, USA). Anti-KLF6 antibody (sc-365633) was from Santa Cruz (Dallas, TX, USA). AP-1 inhibitor PNRI-299 (HY-15131), NF-κB inhibitor PDTC (HY-18738) and STAT3 inhibitor FLLL32 (HY-100544) were obtained from MCE (Monmouth Junction, NJ, USA).
Cell Culture
The THP-1 cell line was obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA). Engineered THP-1 variants—a negative control cell line with baseline PD-L1 expression (PD-L1NC/THP-1) and a PD-L1-overexpressing (PD-L1HI/THP-1) cell line—were established and cultured by Genechem (Shanghai, China). All cells were routinely maintained in RPMI 1640 medium (Gibco, Carlsbad, CA, USA) supplemented with 10% fetal bovine serum (FBS; Gibco, Carlsbad, CA, USA), 100 U/mL penicillin, and 100 µg/mL streptomycin (Thermo Fisher Scientific, Wilmington, DE, USA). To induce differentiation into macrophage-like cells (designated THP-1-M), THP-1 cells were suspended in RPMI 1640 medium with 10% FBS and treated with 100 nM phorbol 12-myristate 13-acetate (PMA; Sigma-Aldrich, St. Louis, MO, USA) for 24–48 h. After PMA treatment, the medium was replaced with fresh PMA-free RPMI 1640/10% FBS, and the cells were allowed to stabilize for 24 h.
Human Peripheral Blood Monocytes Isolation
This study examined PD-L1 expression in monocytes from septic patients by enrolling 8 septic patients and 8 healthy controls. Sepsis diagnosis was confirmed by ICU clinicians based on Sepsis 3.0 criteria. Monocytes were isolated from blood samples and extracted using TRIzol reagent. PD-L1 mRNA levels were assessed via reverse transcription and qRT-PCR. The protocol for differentiating human monocytes into macrophages mirrored the method described for THP-1 cells. The collection and analysis of human samples in this study complied with the principles of the Declaration of Helsinki and were approved by the Ethics Committee of the Army Medical Center of PLA.
Mice
Male C57BL/6 mice (8 weeks old, 20–22 g), including wild-type (PD-L1WT) and PD-L1 knockout (PD-L1KO) strains, were purchased from SPF Biotechnology (Beijing, China) and GemPharmatech (Jiangsu, China), respectively. All mice were housed under specific pathogen-free conditions at 19–23°C, 40–70% humidity, and a 12/12-h light/dark cycle at the Experimental Animal Center of Army Medical University (Chongqing, China; Animal Use License No. SYXK(Yu) 2017–0002), with free access to food and water. Sepsis was induced using the cecal ligation and puncture (CLP) model as previously described.19 PD-L1 knockout and wild-type mice were subjected to CLP, with three mice per group. Following intraperitoneal injection of sodium pentobarbital (100 mg/kg), mice were euthanized by rapid cervical dislocation after a 10-minute interval to ensure deep anesthesia. All animal-related research designs and methods in this study adhered to the ARRIVE guidelines.
qRT‒PCR
Total RNA was extracted using the RNApure High-Purity Total RNA Rapid Extraction Kit (centrifugal column type; Bioteke, Beijing, China). Genomic DNA was removed and first-strand cDNA was synthesized with the PrimeScript™ RT Reagent Kit with gDNA Eraser (Takara, Japan). Each qPCR reaction comprised 1 µL of template, 5 µL of TB Green Premix Ex Taq (Takara, Japan), and 4 µL of a primer–water mixture. All samples were run in technical triplicates to determine mean Ct values. Amplification was carried out on a CFX96 Touch™ Real-Time PCR System (Bio-Rad, USA), and data were analyzed using the 2−ΔΔCt method. Gene expression was normalized to the endogenous reference gene β-actin. The qRT‒PCR primers were listed as follows: Actin-F: 5′-TCATGAAGTGTGACGTGGACATC-3′, Actin-R: 5′-CAGGAGGAGCAATGATCTTGATCT-3′. PD-L1-F: 5′-AGATTGTTGCGGCTGGTCAT-3′, PD-L1-R: 5′-TCAGCTTCTGTATCAGCTGTTC-3′.
Western Blotting
Cellular proteins were extracted using RIPA buffer (Beyotime, Shanghai, China) supplemented with 1 mM PMSF. Following a 30-minute incubation on ice, lysates were centrifuged at 12,000 × g for 10 min at 4°C. The resulting supernatants were combined with pre-chilled 5× Laemmli buffer and denatured by heating at 100°C for 10 min. Samples were subsequently stored at −80°C until further use. For immunoblotting, proteins were separated on 10% SDS-PAGE gels and transferred to 0.22 μm PVDF membranes (Merck Millipore). Membranes were blocked with QuickBlock™ buffer (Beyotime) for 1 h at room temperature, followed by an overnight incubation with primary antibodies at 4°C. After washing with TBST, membranes were probed with HRP-conjugated secondary antibodies (#7074, CST) for 1 h at room temperature. Signals were detected using Clarity™ ECL substrate (Bio-Rad) and quantified with Image Lab™ software (v6.1).
RNA-Seq
Total RNA was extracted from treated cells and tissues with TRIzol reagent and submitted for RNA sequencing (Majorbio, Shanghai, China) to produce raw sequencing data. Clean reads were mapped to the mouse reference genome (GRCm39) employing the STAR aligner. Transcript quantification metrics—such as read counts, reads per kilobase million (RPM), and fragments per kilobase million (FPKM)—were calculated using the proprietary bioinformatics pipeline of Majorbio. Differential expression analysis was performed with DEGs-seq software to identify significantly differentially expressed genes (DEGs) using stringent cutoffs: |log2(fold change)| ≥ 1.000 and adjusted p-value < 0.001. Enrichment analyses, including pathway and gene set enrichment analysis (GSEA), were subsequently carried out on the Majorbio Cloud Platform using the DEGs and a gene list ranked by log2FC values.
Gene Set Variation Analysis (GSVA)
GSVA was performed to estimate pathway activity levels in an unsupervised manner. The analysis utilized a normalized gene expression matrix as input, along with a curated collection of gene sets from the Molecular Signatures Database (MSigDB). The gsva() function from the GSVA R package was applied with default parameters, using the Gaussian kernel (kcdf = “Gaussian”) for continuous expression data (eg, log2-TPM values). This method transforms the gene-by-sample expression matrix into a gene-set-by-sample enrichment score matrix, representing the relative activation of each pathway per sample. Differential activity analysis of the resulting GSVA scores between experimental groups was conducted using the limma package to identify significantly altered pathways.
IP-MS
Immunoprecipitation-grade lysates were prepared using immunoprecipitation-specific lysis buffer (Beyotime, China). Protein concentration was determined by the BCA method (Beyotime, China). Protein A/G magnetic beads were pre-coupled with either anti-PD-L1 antibody or isotype control antibody, then incubated with the lysate for 16 hours at 4°C. Immune complexes were analyzed by high-resolution LC–MS/MS using a Q Exactive HF–X hybrid quadrupole–Orbitrap mass spectrometer coupled with an EASY–nLC 1200 chromatograph. Target verification was performed via immunoblot analysis.
Molecular Docking
The three-dimensional structures of PD-L1 and KLF6 were predicted using the AlphaFold deep learning framework. Molecular docking simulations were performed with the GRAMM web server (v2.0). Interaction analysis and molecular visualization were conducted in PyMOL (v2.4). A binding topology diagram was generated based on hydrogen bonding and electrostatic interaction profiles.
Statistical Analysis
For the GEO dataset analysis, the Wilcoxon test was used. For bar charts, data are expressed as the mean ± SEM from 3 independent experiments. Two-group continuous variables were compared using Student’s t-test. All statistical analyses were performed using SPSS v16.0 (IBM Corp). and GraphPad Prism v6.0 (GraphPad Software, San Diego, CA). Statistical significance was set at p < 0.05.
Results
PD-L1 Expression is Significantly Increased and Holds a Central Position in Sepsis Patients
To reflect clinical reality, we extracted datasets GSE46955, GSE69063, and GSE121349 from the GEO database and compared PD-L1 expression. Results showed PD-L1 was significantly increased in peripheral blood cells of sepsis patients across all three datasets (Figure 1A). Based on this, we performed differential gene expression (DEG) analysis on each dataset using criteria FC >1.3, p < 0.05, and conducted an intersection analysis of the DEGs from the three datasets, identifying 30 co-upregulated genes (Figure 1B). Further, we performed protein-protein interaction (PPI) analysis on these 30 genes using the STRING online database and imported the results into Cytoscape for core gene screening. Results showed CD274 (the gene encoding PD-L1) ranked 10th, tied with IL18R1, indicating PD-L1 may play an important role in the pathological process of sepsis (Figure 1C and D). We extracted dataset GSE22248 to analyze PD-L1 expression changes in PBMCs upon LPS stimulation, which showed significantly increased PD-L1 expression post-LPS stimulation (Figure 1E). To validate these results and incorporate our focus on the monocyte/macrophage system, we isolated peripheral blood monocytes from 8 healthy volunteers and 8 sepsis patients for testing. Results showed significantly elevated PD-L1 expression in monocytes from sepsis patients (Figure 1F), consistent with literature reports that PD-L1 is primarily expressed on monocytes in sepsis.20 In summary, PD-L1 expression is elevated during sepsis, but its significance remains unclear.
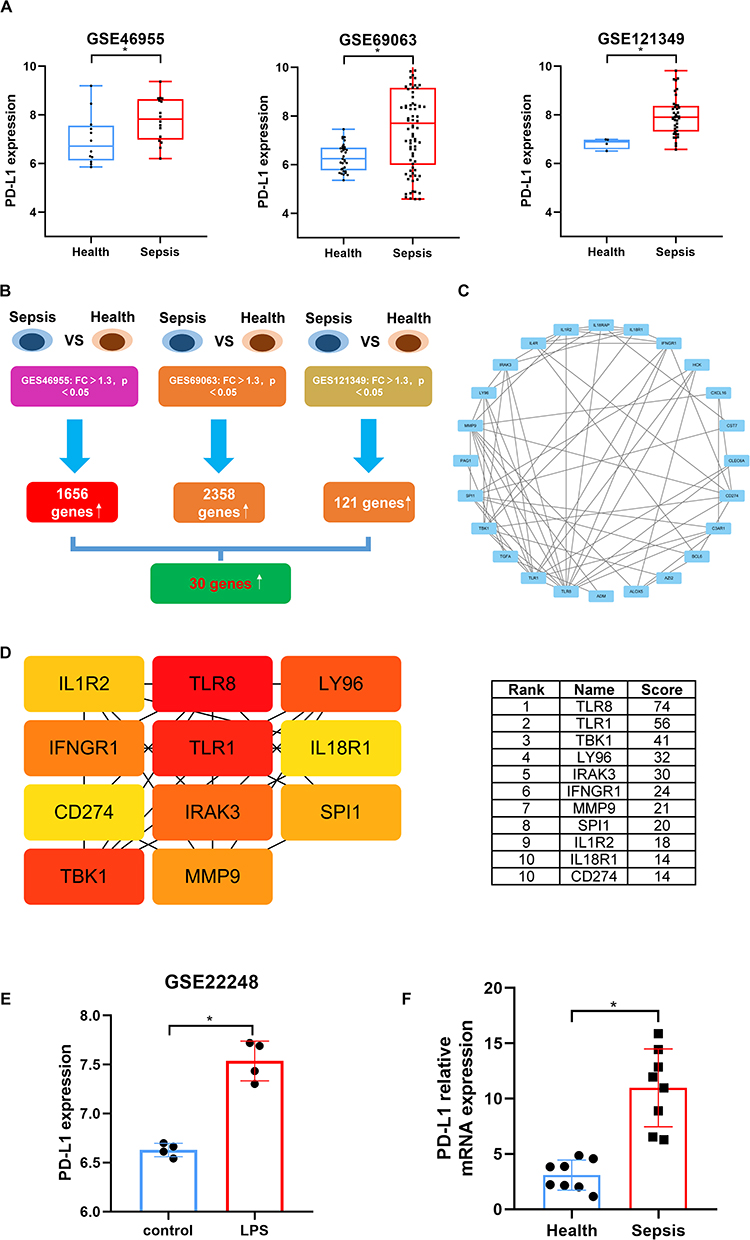 |
Figure 1 PD-L1 expression is upregulated and holds a central position in sepsis. (A) Comparison of PD-L1 mRNA expression levels between healthy controls and septic patients in three independent GEO datasets (GSE46955, GSE69063, GSE121349). Data are presented as box plots; *p < 0.05, Wilcoxon test. (B) Venn diagram showing the overlap of significantly upregulated differentially expressed genes (DEGs, FC > 1.3, p < 0.05) from the three GEO datasets in (A), identifying 30 common upregulated genes. (C) Protein-protein interaction (PPI) network of the 30 common upregulated genes from (B), constructed using the STRING database and visualized with Cytoscape. (D) Identification of top 11 hub genes from the PPI network in (C) using the CytoHubba plugin (MCC algorithm). The color of nodes correspond to their MCC score ranking. CD274 (PD-L1) is ranked 10th. (E) Analysis of PD-L1 expression in human peripheral blood mononuclear cells (PBMCs) from the GSE22248 dataset following LPS stimulation. *p < 0.05, Wilcoxon test. (F) Validation of PD-L1 mRNA expression in monocytes isolated from peripheral blood of 8 healthy volunteers and 8 septic patients. Data are presented as mean ± SEM; *p < 0.05, Student’s t-test. |
Single-Gene GSEA Analysis Based on Sepsis Patients Shows an Association Between PD-L1 and NF-κB Pathway Activation
To study the activation of the NF-κB pathway by PD-L1 using GEO datasets, we first performed batch-effect removal and standardization on the aforementioned datasets. Boxplots and PCA plots before and after batch removal indicated the processed datasets met the requirements for subsequent analysis (Figure 2A and B). The processed dataset is detailed in Supplemental Table 2. Using this curated dataset, we conducted PD-L1 single-gene GSEA to analyze pathways positively correlated with PD-L1 expression changes. The ridge plot of changing pathways showed the NF-κB pathway ranked 10th among the top 20 pathways (Figure 2C). All changing pathways are detailed in Supplemental Table 3. Extracting the GSEA plot specifically for the NF-κB pathway revealed a positive correlation between PD-L1 and its activation (Figure 2D). Furthermore, we generated a differential gene map associated with PD-L1 high/low expression based on the single-gene analysis, showing that genes closely associated with high PD-L1 expression included classic M1 polarization factors regulated by the NF-κB pathway, such as IL-1 and TNF (Figure 2E). All differential genes are detailed in Supplemental Table 4. These results indicate a significant correlation between PD-L1 expression and NF-κB pathway activation in sepsis, but this effect needs validation in PD-L1-overexpressing cells.
 |
Figure 2 Single-gene GSEA analysis links PD-L1 to NF-κB pathway activation in sepsis patients. (A) Box plots and (B) Principal Component Analysis (PCA) plots demonstrating the successful normalization and batch effect removal of the integrated GEO datasets (GSE46955, GSE69063, GSE121349) before analysis. (C) Ridge plot of the top 20 Hallmark pathways positively correlated with PD-L1 expression from the single-gene GSEA analysis. The NF-κB pathway ranks 10th. (D) GSEA enrichment plot for the NF-κB signaling pathway gene set, showing a positive correlation with PD-L1 expression level. (E) Heatmap of genes differentially expressed between PD-L1 High and PD-L1 Low groups from the integrated sepsis cohort. Key NF-κB pathway-related genes (eg, IL1, TNF) are among the most significantly upregulated genes in the PD-L1 High group. |
PD-L1 Promotes NF-κB Pathway Activation in THP-1-Derived Macrophages Stimulated with LPS+IFN-γ
To simulate the pathological changes in monocytes/macrophages of sepsis patients, we used the human acute monocytic leukemia cell line THP-1, induced into macrophages (THP-1-M) for subsequent studies. We used the classic M1 polarization induction method—LPS+IFN-γ—to stimulate negative control cells (PD-L1NC/THP-1-M) and PD-L1-overexpressing cells (PD-L1HI/THP-1-M), followed by RNA-seq analysis. Based on the RNA-seq data, we also performed PD-L1 single-gene GSEA analysis. HALLMARK enrichment showed that PD-L1 overexpression significantly induced inflammatory response and activated the NF-κB pathway (Figure 3A). Extracting the GSEA plots for the inflammatory response and NF-κB pathway separately showed a significant positive correlation between PD-L1 overexpression and these effects (Figure 3B). Additionally, we extracted the GSEA analysis details for the inflammatory response and NF-κB pathways, revealing that among the upregulated genes associated with both upon PD-L1 overexpression, a multitude of pro-inflammatory factors were enriched (Figure 3C). All enriched genes are detailed in Supplemental Table 5. GSVA (Gene Set Variation Analysis) is a non-parametric and unsupervised algorithm. Unlike GSEA, GSVA does not require pre-defined sample groups. Instead, it calculates an enrichment score for specific gene sets in each individual sample, thereby quantifying gene set enrichment results. This approach provides enhanced biological relevance and interpretability for analyzing differentially expressed gene sets. To validate the regulatory effect of PD-L1 overexpression on NF-κB through a more unsupervised algorithm, we performed GSVA on the aforementioned RNA-seq data. The results demonstrated that PD-L1 overexpression indeed enhanced the activation of the NF-κB pathway (Figure 3D). These results indicate that PD-L1 overexpression significantly activates the NF-κB pathway and promotes macrophage polarization towards the M1 type, consistent with the GEO dataset results.
 |
Figure 3 PD-L1 overexpression promotes NF-κB pathway activation and inflammatory response in M1 macrophages in vitro. (A) PD-L1HI/THP-1-M and PD-L1NC/THP-1-M cells were treated with LPS (100 ng/mL) +IFN-γ (20 ng/mL) for 12 h before RNA-seq analysis (n=3). HALLMARK pathway enrichment analysis from single-gene GSEA based on RNA-seq data of PD-L1-overexpressing M1 macrophages (PD-L1HI/THP-1-M+LPS+IFN-γ) versus control (PD-L1NC/THP-1-M+LPS+IFN-γ). Inflammatory response and TNFA signaling via NF-κB are among the top enriched pathways. (B) GSEA enrichment plots for the HALLMARK inflammatory response and HALLMARK TNF-α signaling via NF-κB gene sets, confirming positive enrichment in PD-L1 overexpressing cells. (C) Tables listing representative core enriched genes (leading edge subset) from the GSEA for the inflammatory response and TNF-α signaling via NF-κB pathways, showing numerous pro-inflammatory cytokines. (D) GSVA analysis based on RNA-seq data of PD-L1-overexpressing M1 macrophages (PD-L1HI/THP-1-M+LPS+IFN-γ) versus control (PD-L1NC/THP-1-M+LPS+IFN-γ). |
PD-L1 and NF-κB Form a Positive Feedback Loop in M1 Macrophages
To further elucidate the complete regulatory circuit of PD-L1 in the inflammatory response during sepsis, this study explored its upstream expression regulatory mechanisms. Based on literature reports, the expression of PD-L1 in tumor cells is regulated by inflammatory-related transcription factors such as AP-1, NF-κB, and STAT3.21 Building on this, our study found that in LPS+IFN-γ-induced M1-type macrophages, using corresponding inhibitors (PNRI-299, PDTC, FLLL32) to block the activity of AP-1, NF-κB, or STAT3 significantly reduced the mRNA and protein expression levels of PD-L1 (Figure 4A and B). These results confirm that in inflammatory macrophages, PD-L1 expression is also directly driven by these key inflammatory signaling pathways. Integrating our previous findings—that PD-L1 can activate the NF-κB pathway—we reveal a PD-L1/NF-κB positive feedback regulatory mechanism existing in M1-type macrophages: initial septic signals induce PD-L1 expression by activating pathways such as AP-1, NF-κB, and STAT3. The highly expressed PD-L1, in turn, amplifies the activation of NF-κB signaling, which then feedback to maintain and enhance the transcriptional expression of PD-L1. This self-reinforcing loop may be a key mechanism underlying the uncontrolled inflammation during sepsis.
 |
Figure 4 PD-L1 and NF-κB Form a Positive Feedback Loop in M1 Macrophages. THP-1-M were stimulated with LPS (100 ng/mL) + IFN-γ (20 ng/mL) in the presence or absence of specific inhibitors for AP-1 (PNRI-299, 10 μM), NF-κB (PDTC, 20 μM), or STAT3 (FLLL32, 5 μM) for 12 h (n=3). (A) The PD-L1 mRNA levels were determined by qRT-PCR. (B) PD-L1 protein levels were analyzed by Western blotting. Data are presented as mean ± SEM; *p < 0.05, Student’s t-test. |
PD-L1 Binds to KLF6, Closely Associated with the NF-κB Pathway, in THP-1-M Cells Stimulated with LPS+IFN-γ
To investigate the role of PD-L1 in macrophage inflammatory response from a protein interaction perspective, we performed IP-MS screening on THP-1-M cells treated with PBS or LPS+IFN-γ. Results showed 208 proteins bound to PD-L1 only upon LPS+IFN-γ stimulation. Among the top-ranked proteins, KLF6 attracted our attention due to its close association with the NF-κB pathway (Figure 5A). Analysis based on GEO datasets (GSE134374) showed a linear correlation between PD-L1 and KLF6 expression in sepsis patients, and single-gene GSEA analysis based on KLF6 confirmed its close association with NF-κB pathway activation in sepsis (Figure 5B). Furthermore, we obtained full-length models of PD-L1 and KLF6 using AlphaFold and performed molecular docking using the GRAMM docking service. Results indicated a strong potential for binding, and Co-IP validated their physical interaction (Figure 5C and D). Based on the docking results, we identified 15 potential binding pairs between them, including 3 stronger salt bridges and 12 hydrogen bonds (Figure 5E), laying the foundation for future research into the biological effects of PD-L1-KLF6 binding. In summary, IP-MS screening identified that PD-L1 can bind to KLF6, which is closely associated with the NF-κB pathway, providing insight into how PD-L1 might regulate NF-κB pathway activation.
 |
Figure 5 PD-L1 interacts with the NF-κB-associated transcription factor KLF6. (A) Data and MS/MS spectra from IP-MS analysis showing proteins specifically binding to PD-L1 in THP-1-M cells stimulated with LPS (100 ng/mL) +IFN-γ (20 ng/mL) for 12 h compared to PBS control. KLF6 is a significantly enriched binding partner. (B) (Left) Correlation analysis between PD-L1 and KLF6 expression in the integrated GEO sepsis cohort (GSE134374). (Right) GSEA enrichment plot showing that KLF6 expression is positively correlated with NF-κB pathway activation in sepsis. (C) Predicted three-dimensional structure of the PD-L1/KLF6 complex generated by molecular docking using GRAMM (v2.0), with an energy score of −583. (D) Co-IP validation of the physical interaction between PD-L1 and KLF6 in THP-1-M cells stimulated with LPS+IFN-γ. (E) Schematic diagram of the predicted interaction sites between PD-L1 and KLF6, detailing 3 salt bridges and 12 hydrogen bonds. |
In vivo Studies Confirm PD-L1 Also Positively Regulates NF-κB Pathway Activation
We first validated the relationship between PD-L1 and sepsis prognosis using the GSE65682 dataset via ROC curve analysis. We performed Bootstrap resampling validation (n=2000) on the GSE65682 dataset using R software, confirming the robustness of the results. An AUC value > 0.8 indicates that PD-L1 expression level can serve as a reliable biomarker for prognostic assessment in sepsis patients, with potential clinical application value (Figure 6A). To more directly reflect the regulatory effect of PD-L1 on NF-κB pathway activation in vivo, we constructed PD-L1 knockout mice (PD-L1KO/mice) and corresponding wild-type mice (PD-L1WT/mice) and subjected them to the classic sepsis model—CLP. Since the liver is one of the most important immune organs, we first analyzed pathway changes before and after PD-L1 knockout using RNA-seq. Analysis of the overall table and the extracted GSEA table showed that PD-L1 knockout significantly inhibited TNF-mediated inflammatory pathway and NF-κB pathway activation (Figure 6B and C). Simultaneously, we extracted the GSEA analysis details for the TNF-mediated inflammatory pathway and the NF-κB pathway, showing that among the downregulated genes associated with both upon PD-L1 knockout, numerous pro-inflammatory factors were enriched (Figure 6D). All enriched genes are detailed in Supplemental Table 6. Notably, studies have reported significantly higher survival rates in PD-L1 knockout mice subjected to CLP or hemorrhage+CLP compared to wild-type mice, suggesting PD-L1 may promote the pathological process of sepsis,15,16 which is consistent with our results. Integrating the above data, we validated the positive correlation between PD-L1 and inflammatory response/NF-κB pathway activation in the context of in vivo PD-L1 knockout, and also macroscopically verified PD-L1 as an observable molecule for sepsis prognosis.
 |
Figure 6 PD-L1 positively regulates NF-κB pathway activation in vivo. (A) ROC curve analysis of PD-L1 expression for predicting sepsis prognosis using the GSE65682 dataset. The AUC value is greater than 0.8. Bootstrap resampling (n=2000) was used. (B) GSEA enrichment plot showing significant inhibition of the TNF signaling pathway and NF-κB signaling pathway in liver tissue from PD-L1 knockout (PD-L1KO) septic mice (n=3) compared to wild-type (PD-L1WT) septic mice (n=3) subjected to CLP after 12 h (a negative NES value indicates inhibition). (C) GSEA enrichment plot showing significant inhibition of the TNF signaling pathway and NF-κB signaling pathway in liver tissue from PD-L1KO septic mice compared to PD-L1WT septic mice. (D) Tables listing representative core enriched genes (leading edge subset) from the GSEA for the TNF and NF-κB signaling pathways, showing downregulation of pro-inflammatory genes in PD-L1KO mice. |
Discussion
M1 macrophage polarization plays a central pro-inflammatory role in inflammatory diseases, significantly exacerbating damage in sepsis. Sepsis is host immune dysregulation in response to infection leading to life-threatening organ dysfunction.22 M1 macrophages secrete large amounts of pro-inflammatory factors like TNF-α and IL-6, triggering a systemic inflammatory storm and causing widespread tissue damage.23 Specific damages include: destruction of alveolar structure in acute lung injury, increasing inflammatory exudation and hypoxia risk;24 exacerbation of cardiomyocyte necrosis and functional failure in myocardial injury via M1 macrophage-derived exosomes, worsening cardiac function indicators;23 promotion of liver injury and acute kidney injury through pathways like heparin-binding protein, amplifying the inflammatory cascade.25 These damages significantly increase the risk of multiple organ failure and mortality.26 In summary, M1 polarization drives a vicious cycle of inflammatory damage in sepsis, hindering tissue repair processes.
The dysregulated mechanism of M1 macrophage polarization is closely related to aberrant activation of the NF-κB signaling pathway. In inflammatory diseases like sepsis, pathogen-associated molecular patterns (eg, LPS) activate the downstream MyD88-dependent pathway via Toll-like receptors, triggering IKK complex phosphorylation, leading to IκB degradation. This process promotes nuclear translocation of NF-κB dimers (eg, p50/p65), which, upon binding DNA, initiate transcription of pro-inflammatory genes (eg, TNF-α, IL-6, iNOS).27 Studies show that proteins like MMP-9 and SENP3 can drive macrophage polarization towards M1 by activating the NF-κB pathway in sepsis, exacerbating the inflammatory cascade.28 Furthermore, NF-κB acts synergistically with the MAPK pathway to promote inflammatory cytokine release, forming a positive feedback loop.27 However, NF-κB regulation is highly complex: sustained activation leads to uncontrolled M1 polarization, inhibits anti-inflammatory M2 conversion, and aggravates tissue damage;29 yet, this pathway is indispensable for immune defense, and excessive inhibition may impair pathogen clearance.30 Current key bottlenecks include: the activation thresholds and interaction networks of NF-κB in different organ microenvironments are not systematically elucidated;31 fine-tuning regulatory mechanisms like histone modifications and non-coding RNAs (eg, miR-155) on NF-κB require deeper investigation;32,33 reliable models for the time-dependent switching patterns of NF-κB signaling across different stages of sepsis are lacking.34 These mechanistic gaps limit clinical intervention efficacy. For example, NF-κB inhibitors (eg, PDTC) can reduce inflammation in animal models, but human clinical trials often fail due to off-target effects and immunosuppression risks.35 Similarly, TLR4-targeting antibodies, while blocking NF-κB activation, have not significantly improved sepsis patient survival, suggesting microenvironmental interaction networks are far more complex than current understanding.27 Therefore, elucidating the multi-dimensional regulatory mechanisms of the NF-κB pathway is key to overcoming clinical translation challenges.
PD-L1 is the primary ligand for the immune checkpoint molecule PD-1, playing a key role in maintaining immune homeostasis by inhibiting T cell activation and cytokine secretion. In inflammatory responses, PD-L1 prevents excessive immune activation and tissue damage, but its aberrant expression is also associated with immune escape. As an important expressing cell of PD-L1, macrophage regulation is closely related to the inflammatory microenvironment. Inflammatory factors in tumor or infection microenvironments can significantly induce PD-L1 expression on macrophages. IFN-γ secreted by activated T cells upregulates PD-L1 via the JAK-STAT pathway; IL-10 enhances its expression via the Akt/PI3K pathway, and IL-27 via the STAT3 pathway.36 Under hypoxia, HIF-1α directly binds the PD-L1 promoter region, promoting expression and inhibiting T cell activation.37 In chronic inflammation (eg, arthritis), the PD-L1/PD-1 pathway inhibits effector T cell function, promoting inflammation resolution and maintaining self-tolerance.38,39 Beyond immunosuppression, PD-L1 has also been reported to have a positive regulatory role in inflammation. PD-L1 knockout significantly improves survival in mice subjected to CLP and hemorrhage+CLP sepsis models, but the specific molecular mechanisms are unclear.15,16 Although studies show PD-L1 can activate interferon signaling pathways and transcription factors (eg, SPI1/MAFB) to promote pro-inflammatory cytokine secretion in human pluripotent stem cell (hPSC)-derived macrophages,40 this has not been validated in sepsis models. Additionally, PD-L1 can exacerbate lung injury via the HIF-1α pathway, but the focus was primarily on lung epithelial and vascular endothelial cells rather than immune cells.41 As mentioned, the crucial role of macrophages in sepsis pathology is well-established. However, the regulatory mechanisms of PD-L1 within macrophages during sepsis are not fully elucidated, especially its relationship with the NF-κB pathway requires urgent investigation.
This study preliminarily explored the regulatory role of PD-L1 on the NF-κB pathway during M1 macrophage polarization in sepsis through integrated multi-omics analysis and experimental validation. First, based on multi-dataset GEO analysis and PBMC detection in sepsis patients, we found significantly upregulated PD-L1 expression during sepsis (Figure 1A and F), and it held a central position in the co-expression network (Figure 1C and D), suggesting its potential involvement in the disease process. Further single-gene GSEA analysis showed a significant positive correlation between PD-L1 expression and NF-κB signaling pathway activation (Figure 2). To validate this correlation, we found that PD-L1 overexpression significantly enhanced the enrichment of NF-κB pathway-related gene sets in LPS+IFN-γ-stimulated M1 macrophages (Figure 3). Furthermore, our inhibitor assays demonstrated the existence of a positive feedback loop involving PD-L1/NF-κB in M1-type macrophages (Figure 4), providing new insights into how PD-L1 could amplify the pro-inflammatory response. Regarding the molecular mechanism, IP-MS screening and Co-IP experiments confirmed a direct interaction between PD-L1 and the NF-κB-related transcription factor KLF6 (Figure 5), suggesting its potential involvement in signal transduction through protein interaction. In vivo experiments further showed that NF-κB pathway activation was significantly inhibited in the liver tissue of PD-L1 knockout septic mice (Figure 6B–D), and clinical data analysis indicated that PD-L1 expression has good predictive value for sepsis prognosis (AUC > 0.8; Figure 6A). In summary, this study preliminarily elucidates the role of PD-L1 as a potential activator of the NF-κB pathway in M1 macrophages from the perspectives of clinical association, cellular phenotype, molecular interaction, and whole-animal level. Another study reported that after the NF-κB signaling pathway is activated by inflammatory factors (eg, TNF-α) in macrophages, it can directly upregulate PD-L1 transcription by binding to its gene promoter.42 Combined with our finding that PD-L1 promotes NF-κB activation, we speculate a positive feedback mechanism exists between them. This bidirectional regulation could lead to sustained amplification of inflammatory signals, thereby exacerbating the inflammatory response and pathological damage.
Although this study preliminarily reveals that PD-L1 may influence M1 macrophage polarization by regulating the NF-κB pathway in sepsis, several limitations remain. First, due to the high cost and stability challenges of single-cell sequencing technology, we were unable to deeply resolve the PD-L1 expression profile and heterogeneity of monocyte/macrophage subsets in peripheral blood and major organs during sepsis at the single-cell level. Second, although bioinformatic and omics analyses suggested NF-κB pathway activation, key molecular events such as phosphorylation and nuclear translocation of this pathway were not directly validated in vitro. Furthermore, due to the significant heterogeneity of the screened inflammatory factors, this study did not functionally focus on or clinically validate key specific factors. Mechanistically, although an interaction between PD-L1 and KLF6 was found, the downstream molecular events regulating the NF-κB pathway following this interaction remain to be elucidated, and validation using gene knockdown or blockade experiments targeting key nodes like KLF6 is lacking. Finally, in vivo experiments were based solely on liver transcriptome data; future work requires systematic validation using macrophage-specific PD-L1 knockout mice regarding survival analysis, multi-organ function assessment, and dynamic changes in inflammatory factors. In conclusion, this study remains at the stage of macroscopic correlation exploration, and more in-depth mechanistic details and translational value need to be expanded in subsequent work.
Conclusion
This study demonstrates that PD-L1 expression is significantly upregulated during sepsis and shows a positive correlation with the activation of the NF-κB signaling pathway. Preliminary experiments suggest a potential interaction between PD-L1 and the transcription factor KLF6, which might be one mechanism underlying its potential role in M1 macrophage polarization. Multi-omics analysis and in vivo and in vitro experimental results indicate that PD-L1 could serve as a potential regulatory molecule in the pathological process of sepsis and related inflammatory responses, with its expression level showing certain prognostic value. This study provides preliminary clues and directions for further exploration of the non-classical functions of PD-L1 in immune regulation during sepsis.
Data Sharing Statement
The datasets used and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Ethics Approval and Consent to Participate
The animal study was approved by the Laboratory Animal Welfare and Ethics Committee of the Army Medical University (no. AWUWEC20247118). All animal treatment operations were executed according to the Army Medical University Ethical Guidelines for Animal Experiments. Patient sample acquisition was approved by the Ethics Committee of the Army Medical Center of PLA (NO. 2025318). Informed consent was obtained directly from all healthy volunteers. For patients with sepsis, informed consent was obtained from their legal guardians, as the majority of these patients were in a comatose state upon enrollment and were thus unable to provide consent themselves.
Author Contributions
All authors took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work. Mu Yuan: Validation, Investigation, Formal analysis, Data curation. Tong Jin: Validation, Investigation, Formal analysis, Data curation. Qing Zhao: Investigation. Lixing Tian: Writing – original draft, Validation, Supervision, Methodology, Investigation, Formal analysis, Data curation, Funding acquisition.
Funding
This work was supported by grants from the National Natural Science Foundation of China (82402548), Basic Research Projects of the Army Medical Center (ZXJCYJ202519) and Postdoctoral Innovative Research Program of the Army Medical Center (ZXBSH023).
Disclosure
The authors have no conflicts of interest to declare.
References
1. Boutilier AJ, Elsawa SF. Macrophage polarization states in the tumor microenvironment. Int J Mol Sci. 2021;22(13):6995. doi:10.3390/ijms22136995
2. Deng L, Jian Z, Xu T, et al. Macrophage polarization: an important candidate regulator for lung diseases. Molecules. 2023;28(5):2379. doi:10.3390/molecules28052379
3. Platanitis E, Decker T. Regulatory networks involving STATs, IRFs, and NFκB in inflammation. Front Immunol. 2018;9:2542. doi:10.3389/fimmu.2018.02542
4. Atri C, Guerfali FZ, Laouini D. Role of human macrophage polarization in inflammation during infectious diseases. Int J Mol Sci. 2018;19(6):1801. doi:10.3390/ijms19061801
5. Pérez S, Rius-Pérez S. Macrophage polarization and reprogramming in acute inflammation: a redox perspective. Antioxidants. 2022;11(7):1394. doi:10.3390/antiox11071394
6. Toledo B, Zhu Chen L, Paniagua-Sancho M, et al. Deciphering the performance of macrophages in tumour microenvironment: a call for precision immunotherapy. J Hematol Oncol. 2024;17:44. doi:10.1186/s13045-024-01559-0
7. Li H, Li Y, Luo S, et al. The roles and mechanisms of the NF-κB signaling pathway in tendon disorders. Front Vet Sci. 2024;11:1382239. doi:10.3389/fvets.2024.1382239
8. Ni R, Jiang L, Zhang C, et al. Biologic mechanisms of macrophage phenotypes responding to infection and the novel therapies to moderate inflammation. Int J Mol Sci. 2023;24(9):8358. doi:10.3390/ijms24098358
9. Mussbacher M, Derler M, Basílio J, et al. NF-κB in monocytes and macrophages - an inflammatory master regulator in multitalented immune cells. Front Immunol. 2023;14:1134661. doi:10.3389/fimmu.2023.1134661
10. Liu D, Wen C, Chen L, et al. The emerging roles of PD-L1 subcellular localization in tumor immune evasion. Biochem Pharmacol. 2024;220:115984. doi:10.1016/j.bcp.2023.115984
11. Chang K, Svabek C, Vazquez-Guillamet C, et al. Targeting the programmed cell death 1: programmed cell death ligand 1 pathway reverses T cell exhaustion in patients with sepsis. Crit Care. 2014;18:R3. doi:10.1186/cc13176
12. da Mota NVF, Brunialti MKC, Santos SS, et al. Immunophenotyping of monocytes during human sepsis shows impairment in antigen presentation: a shift toward nonclassical differentiation and upregulation of FCγRi-receptor. Shock. 2018;50:293–16. doi:10.1097/SHK.0000000000001078
13. Lu Y, Wang G, Li C. Expression of peripheral monocytic programmed death ligand-1 in severe sepsis combined with HBV-related cirrhosis. A pilot observational study. Cent Eur J Immunol. 2021;46:217–224. doi:10.5114/ceji.2021.108179
14. Tai H, Xing H, Xiang D, et al. Monocyte programmed death ligand-1, a predicator for 28-day mortality in septic patients. Am J Med Sci. 2018;355:362–367. doi:10.1016/j.amjms.2017.12.008
15. Rossi AL, Le M, Chung CS, et al. A novel role for programmed cell death receptor ligand 2 in sepsis-induced hepatic dysfunction. Am J Physiol Gastrointest Liver Physiol. 2018;316:G106–G114. doi:10.1152/ajpgi.00204.2018
16. Lomas-Neira J, Monaghan SF, Huang X, et al. Novel role for PD-1: PD-L1 as mediator of pulmonary vascular endothelial cell functions in pathogenesis of indirect ARDS in mice. Front Immunol. 2019;9:3030. doi:10.3389/fimmu.2018.03030
17. Zhang Y, Lei CQ, Hu YH, et al. Krüppel-like factor 6 is a co-activator of NF-κB that mediates p65-dependent transcription of selected downstream genes. J Biol Chem. 2014;289:12876–12885. doi:10.1074/jbc.M113.535831
18. Subramanian A, Tamayo P, Mootha VK, et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc Natl Acad Sci U S A. 2005;102:15545–15550. doi:10.1073/pnas.0506580102
19. Hou JC, Chen QX, Wu XL, et al. S1PR3 signaling drives bacterial killing and is required for survival in bacterial sepsis. Am J Respir Crit Care Med. 2017;196:1559–1570. doi:10.1164/rccm.201701-0241OC
20. Zhang Y, Li J, Lou J, et al. Upregulation of programmed death-1 on T cells and programmed death ligand-1 on monocytes in septic shock patients. Crit Care. 2011;15:R70. doi:10.1186/cc10059
21. Kalantari Khandani N, Ghahremanloo A, Hashemy SI. Role of tumor microenvironment in the regulation of PD-L1: a novel role in resistance to cancer immunotherapy. J Cell Physiol. 2020;235:6496–6506. doi:10.1002/jcp.29671
22. Chen Y, Yang L, Li X. Advances in Mesenchymal stem cells regulating macrophage polarization and treatment of sepsis-induced liver injury. Front Immunol. 2023;14:1238972. doi:10.3389/fimmu.2023.1238972
23. Bi C, Wang D, Hao B, et al. Snhg14/miR-181a-5p axis-mediated “M1” macrophages aggravate LPS-induced myocardial cell injury. Heliyon. 2024;10:e37104. doi:10.1016/j.heliyon.2024.e37104
24. Wang Z, Wang Z. The role of macrophages polarization in sepsis-induced acute lung injury. Front Immunol. 2023;14:1209438. doi:10.3389/fimmu.2023.1209438
25. Xing L, Zhongqian L, Chunmei S, et al. Activation of M1 macrophages in sepsis-induced acute kidney injury in response to heparin-binding protein. PLoS One. 2018;13:e0196423. doi:10.1371/journal.pone.0196423
26. Rudd KE, Johnson SC, Agesa KM, et al. Global, regional, and national sepsis incidence and mortality, 1990–2017: analysis for the Global Burden of Disease Study. Lancet. 2020;395:200–211. doi:10.1016/S0140-6736(19)32989-7
27. Yang Z, Gao Y, Zhao L, et al. Molecular mechanisms of Sepsis attacking the immune system and solid organs. Front Med. 2024;11:1429370. doi:10.3389/fmed.2024.1429370
28. Yi L, Chen Y, Zhang Y, et al. Deleting fibroblast growth factor 2 in macrophages aggravates septic acute lung injury by increasing M1 polarization and inflammatory cytokine secretion. Mol Biomed. 2024;5:50. doi:10.1186/s43556-024-00203-0
29. Cornice J, Verzella D, Arboretto P, et al. NF-κB: governing macrophages in cancer. Genes. 2024;15(2):197. doi:10.3390/genes15020197
30. Wang W, He Z. Gasdermins in sepsis. Front Immunol. 2023;14:1203687. doi:10.3389/fimmu.2023.1203687
31. Ge Z, Chen Y, Ma L, et al. Macrophage polarization and its impact on idiopathic pulmonary fibrosis. Front Immunol. 2024;15:1444964. doi:10.3389/fimmu.2024.1444964
32. Li Y, Ai S, Li Y, et al. The role of natural products targeting macrophage polarization in sepsis-induced lung injury. Chin Med. 2025;20:19. doi:10.1186/s13020-025-01067-4
33. Xie D, Ouyang S. The role and mechanisms of macrophage polarization and hepatocyte pyroptosis in acute liver failure. Front Immunol. 2023;14:1279264. doi:10.3389/fimmu.2023.1279264
34. Qin F, Tan H, Yang Y, et al. Upregulation of Cullin1 neddylation promotes glycolysis and M1 polarization of macrophage via NF-κB p65 pathway in sepsis. Funct Integr Genomics. 2024;24:204. doi:10.1007/s10142-024-01483-z
35. Wang Y, Lu X, Lu J, et al. The role of macrophage polarization in tendon healing and therapeutic strategies: insights from animal models. Front Bioeng Biotechnol. 2024;12:1366398. doi:10.3389/fbioe.2024.1366398
36. Goldman N, Lomakova YD, Londregan J, et al. High macrophage PD-L1 expression not responsible for T cell suppression. Cell Immunol. 2018;324:50–58. doi:10.1016/j.cellimm.2017.12.013
37. Noman MZ, Desantis G, Janji B, et al. PD-L1 is a novel direct target of HIF-1α, and its blockade under hypoxia enhanced MDSC-mediated T cell activation. J Exp Med. 2014;211:781. doi:10.1084/jem.20131916
38. Canavan M, Floudas A, Veale DJ, et al. The PD-1: PD-L1 axis in Inflammatory Arthritis. BMC Rheumatol. 2021;5:1. doi:10.1186/s41927-020-00171-2
39. Boussiotis VA. Molecular and biochemical aspects of the PD-1 checkpoint pathway. N Engl J Med. 2016;375:1767–1778. doi:10.1056/NEJMra1514296
40. Cao H, Xiang Y, Zhang S, et al. PD-L1 regulates inflammatory programs of macrophages from human pluripotent stem cells. Life Sci Alliance. 2023;7(2):e202302461. doi:10.26508/lsa.202302461
41. Zhao S, Gao J, Li J, et al. PD-L1 regulates inflammation in LPS-induced lung epithelial cells and vascular endothelial cells by interacting with the HIF-1α signaling pathway. Inflammation. 2021;44:1969–1981. doi:10.1007/s10753-021-01474-3
42. Antonangeli F, Natalini A, Garassino MC, et al. Regulation of PD-L1 expression by NF-κB in cancer. Front Immunol. 2020;11:584626. doi:10.3389/fimmu.2020.584626
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
MiR-126-5p Down-Regulation Alleviates the Inflammatory Response of Allergic Rhinitis in Children via Inhibiting HIPK2/NF-κB Signaling Pathway

Jing Y, Xie J, Long S, Huang M
Journal of Inflammation Research 2025, 18:3981-3992
Published Date: 17 March 2025
Kushe Tincture Ameliorates DNCB-Induced Atopic Dermatitis by Affecting NF-Kb/JAK-STAT3 Pathway: Bioinformatics Analysis and Animal Experiment Verification
Feng H, Zhou Y, Ren X, Zhu X, Duan X, Qu S, Zhu B, Li Y, Qin Y
Journal of Inflammation Research 2025, 18:15293-15308
Published Date: 4 November 2025