Back to Journals » Infection and Drug Resistance » Volume 12
Para-aminosalicylic acid increases the susceptibility to isoniazid in clinical isolates of Mycobacterium tuberculosis
Authors Zhang T, Jiang G, Wen S, Huo F, Wang F, Huang H ![]() , Pang Y
, Pang Y ![]()
Received 7 January 2019
Accepted for publication 1 March 2019
Published 11 April 2019 Volume 2019:12 Pages 825—829
DOI https://doi.org/10.2147/IDR.S200697
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sahil Khanna
Tingting Zhang,1,2 Guanglu Jiang,1,2 Shu’an Wen,1,2 Fengmin Huo,1,2 Fen Wang,1,2 Hairong Huang,1,2 Yu Pang1,2
1National Clinical Laboratory on Tuberculosis, Beijing Chest Hospital, Capital Medical University, Beijing Tuberculosis and Thoracic Tumor Institute, Beijing, People’s Republic of China; 2Beijing Key Laboratory on Drug-Resistant Tuberculosis Research, Beijing Chest Hospital, Capital Medical University, Beijing Tuberculosis and Thoracic Tumor Institute, Beijing, People’s Republic of China
Background: The purpose of this work was to assess the activity of para-aminosalicylic acid (PAS) in combination with isoniazid (INH) against clinical isolates of Mycobacterium tuberculosis (MTB).
Materials and methods: A total of 72 MTB isolates with differential in vitro drug susceptibilities were included in this study, comprising 24 pan-susceptible, 24 MDR-TB, and 24 extensively drug-resistant (XDR) isolates. A microplate alamarBlue assay was performed to identify the minimal inhibitory concentrations (MICs) of MTB isolates. Results: The MIC50 of INH was 4 mg/L, and that of PAS was 0.063 mg/L against MTB isolates when single drug used. The combined use of INH and PAS resulted in 16-fold and 8-fold decrease in MIC50 for INH and PAS, respectively. The INH-PAS revealed synergistic activity in 94.4% of the isolates. In addition, there was no significant difference in the FIC index of the INH-PAS combination among individual isolates harboring different susceptibility pattern (P>0.05).
Conclusion: The synergy between INH and PAS is demonstrated using non-multidrug-resistant (non-MDR) and MDR-TB strains, which will provide clinicians with useful hints to reuse this combination for treatment of TB patients in clinical practice.
Keywords: Mycobacterium tuberculosis, isoniazid, para-aminosalicylic acid, synergy
Introduction
Mycobacterium tuberculosis (MTB) remains the deadliest bacterial infectious disease worldwide,1 and is responsible for approximately 10.0 million new cases of active tuberculosis (TB) cases and 1.6 million deaths in 2017.2 Treatment of active pulmonary TB patients infected with drug-susceptible MTB requires the 6-month standard regimen: isoniazid (INH), rifampicin (RIF), ethambutol and pyrazinamide.3 The long-term therapy and occurrence of adverse drug events can adversely impact patient adherence to anti-TB treatment,4 thereby resulting the emergence of drug-resistant MTB bacilli. Recently, the increasing incidence of multidrug-resistant TB (MDR-TB), defined as resistance to both INH and RIF, highlights the urgent need for developing new antimicrobial agents and formulating new regimens against this severe form of drug-resistant TB.4 Although great effort has been made in new drug discovery and development in past decades, its costly and lengthy process allows the repurposing of old drugs as an attractive alternative strategy to provide valuable choice for patients infected with drug-resistant TB.
Para-aminosalicylic acid (PAS) is a second-line anti-TB drug (SLD) that was found to be effective for treatment of TB in the 1940s.5 As a structural analog of p-aminobenzoic acid, it disrupts folate metabolism through competitive binding with dihydrofolate reductase, thereby inhibiting the growth of tubercle bacilli.5 Despite having excellent efficacy against TB in both in vitro experiments and clinical trials, PAS was eventually replaced with a better-tolerated ethambutol due to gastrointestinal disturbance associated with the usage of PAS.5 Later, the global spread of MDR-TB necessitated the repurposing of PAS as an important second-line agent.5 Previous pharmacokinetics data indicated that PAS could increase plasma levels and prolong effective time of INH by retarding the acetylation pathway of INH.6 Therefore, the in vivo interaction between INH and PAS indicates that the coadministration of PAS with INH could bring additional benefit for TB cases, especially for rapid acetylators. More importantly, high-dose INH is endorsed as part of a standardized treatment regimen for patients with MDR-TB.7 Despite having the interesting pharmacokinetic property, the synergistic effect between INH and PAS is another concern with clinical coadministrated usage of these agents, whereas little is known about their interaction during TB therapy. Therefore, the purpose of this work was to assess the activity of PAS in combination with INH against clinical isolates of MTB.
Materials and methods
Materials
A total of 72 MTB isolates from Beijing Chest Hospital were included in this study, comprising 24 pan-susceptible, 24 MDR-TB, and 24 extensively drug-resistant (XDR) isolates. The MTB isolates were routinely cultured for diagnosis purpose rather than for this study. The drug susceptibility for isoniazid, rifampicin, ethambutol, streptomycin, amikacin (AMK) and levofloxacin (LVX) was determined with the absolute concentration method as previously described.8 Mycobacterium species identification was performed by sequencing of multiple genes, including 16S rRNA, hsp65, rpoB, and 16S-23S rRNA internal transcribed spacer sequence.9 MDR-TB is defined as MTB isolate resistant to at least INH and RIF; XDR-TB is defined as MTB strain resistant to INH, RIF, LVX and AMK. A strain of H37Rv (ATCC 27294) was used as the control. Stock solutions of the isolates were stored in Middlebrook 7H9 broth containing 10% glycerol at −70°C until assay. Prior to conducting phenotypic drug susceptibility testing, the strains were subcultured on on Löwenstein–Jensen (L-J) slant for 4 weeks at 37°C.
Antibacterial activity assay
The antimicrobial agents used in study were ordered from Sigma-Aldrich (St Louis, MO, USA). A microplate alamarBlue assay was performed to identify the minimal inhibitory concentrations (MICs) of MTB isolates as previously described.10 Briefly, the inoculum was prepared with the 4-week-old cultures on L–J medium. The turbidity of the cultures was adjusted to a 1.0 McFarland standard, and diluted 1:20 in Middlebrook 7H9 broth. Then 100 μL of dilution was added into the wells of the 96-well plate containing two-fold serial dilutions of each tested compound. After 7 days of incubation at 37°C, 70 μL of alamarBlue solution was pipetted to each well, incubated for 24 hours at 37°C, and assessed for color change. The MIC was defined as the lowest drug concentration that prevented a color change from blue to pink. The reference strain, MTB ATCC 27294, was used as a control in all MIC determinations. All the experiments were performed in triplicate.
Combination testing
A checkerboard assay was employed to assess whether drug combinations act synergistically, indifferently, or antagonistically.10 Fractional inhibitory concentration index (FICI) was calculated by use of the following formula: the MIC of drug A tested in combination divided by the MIC of drug A alone plus the MIC of drug B tested in combination divided by the MIC of drug B alone. FICI values were interpreted as follows: FICI ≤0.5, synergy; 0.5<FICI≤2, indifference; FICI >2, antagonism.
Data analysis
The Pearson chi-squared test was used to compare the proportions of synergistic effect in MTB isolates stratified by drug susceptibility profiles and genetic mutation types. Two-sided P-values of <0.05 was considered statistically significant. All the statistical analyses were done using SPSS version 17.0 software (SPSS Inc., Chicago, IL, USA).
Results and discussion
A total of 72 MTB isolates with differential in vitro drug susceptibilities were included in this study. The detailed drug susceptibility profiles are given in Table 1. Out of 24 MDR-TB isolates, 7 (29.2%) were resistant only to INH and RIF. In addition, 4 (16.7%) were resistant to EMB, 11 (45.8%) were resistant to STR, and 3 (12.5%) were resistant to LVX. For 24 XDR-TB isolates, streptomycin (STR) resistance was the most frequently detected resistance, accounting for 66.7% (16/24) of XDR-TB isolates tested. There were 9 isolates (37.5%) that were resistance to all the six anti-TB drugs.
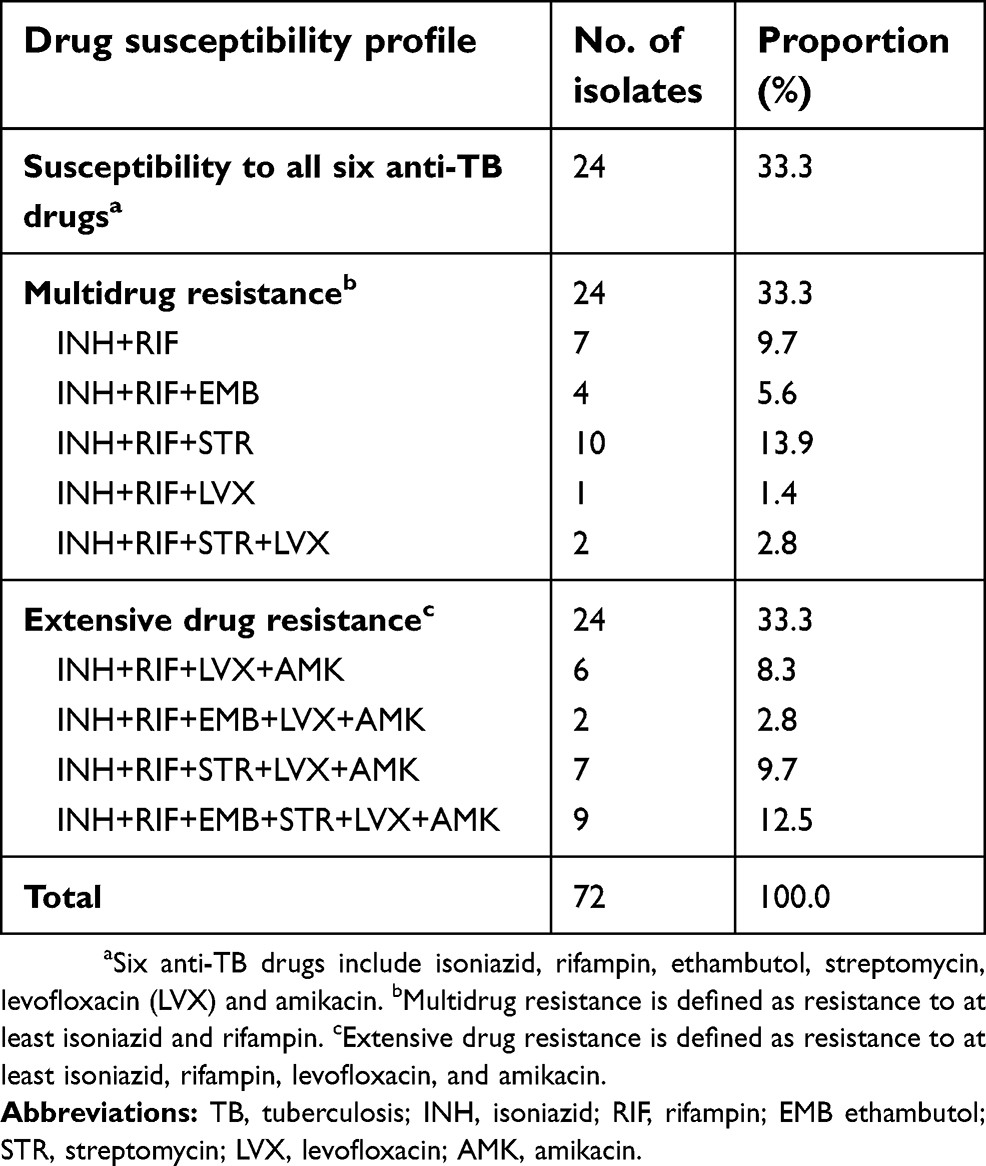 | Table 1 Drug susceptibility profiles of M. tuberculosis isolates enrolled in this study |
The comparative data obtained by using the x/y quotients for the determination of the interactions between INH and PAS against 72 MTB isolates are summarized in Table 2. By the microdilution method, the MIC50 of INH was 4 mg/L, and that of PAS was 0.063 mg/L when single drug used. The combined use of INH and PAS resulted in 16-fold and 8-fold decrease in MIC50 for INH (MIC50=0.25 mg/L) and PAS (MIC50=0.008 mg/L), respectively. As a consequence, the INH-PAS revealed synergistic activity in 94.4% (68/72) of the isolates, whereas indifference and antagonism were detected in only three (4.2%) and one (1.4%) strains, respectively. In addition, we also analyzed the proportion of strains exhibiting synergy with the INH-PAS combination stratified to drug resistant profiles. As shown in Table 2, synergistic activity was identified in 24 (24/24, 100.0%) Pan-S, 23 (23/24, 95.8%) MDR-TB and 21 (21/24, 87.5%) XDR-TB isolates. There was no significant difference in the FICI of the INH-PAS combination among individual isolates harboring a different susceptibility pattern (P>0.05).
 | Table 2 The distribution of synergistic effect between INH and PAS against MTB isolates |
INH remains one of the most potent anti-TB agents used in the treatment of tuberculosis.4 The increasing prevalence of INH resistance, especially in a setting with a high burden of TB, impairs the efficacy of INH against MTB in clinical practice.11 In this study, our in vitro data demonstrated that the INH-PAS combination revealed synergistic activity in most of clinical MTB isolates. In other words, the presence of PAS significantly increases the susceptibility to INH in clinical MTB isolates. This synergistic effect may have several explanations. First, recent findings clearly demonstrate that PAS has a specific role for interference of folate metabolism, thereby inhibiting the biosynthesis of nucleic acid.5 We hypothesize that the supplement of PAS at nonlethal dose would perturb the production of the target gene of INH encoding an enoyl-acyl carrier protein reductase, and further decrease the MIC level of INH in inhibition of MTB growth. Second, while folate metabolism is a principal target of PAS action in MTB, there is strong evidence that PAS can disrupt with mycobacterial iron acquisition.5 In addition, it was found that the decreased iron supplement has a positive impact on the effect of anti-TB treatment by INH,12 indicating that MTB showed an enhancement in INH susceptibility when cultivated under iron-limiting conditions. Therefore, the capability of PAS in the iron acquisition for MTB may be another important explanation for the synergistic effect. Besides, we found that there were four isolates exhibiting indifference or antagonism with the INH-PAS combination, which were all from MDR- or XDR-TB groups. A previous study by Velayati and colleagues demonstrated that the thickness of cell for drug-resistant TB was significantly thicker than that for susceptible isolates.13 Despite lack of experimental evidence, we hypothesize that a lack of synergistic effect between INH and PAS may be due to the potential increase in the cell wall thickness for these isolates.
The synergy between PAS and INH demonstrated that INH could lower the MIC of PAS for these MTB isolates. Although the exact reason remains unclear, one possible explanation for this observation may be due to the bactericidal mechanism of INH that inhibits cell wall lipid synthesis.14 There is no doubt that the in vitro addition of INH will increase permeability of the bacterial envelope, thus increasing penetration of PAS into bacilli and contributing the enhanced susceptibility of MTB to PAS. Our findings have several important implications for TB treatment with INH. On the one hand, the existing evidence indicates that high-dose INH is effective in the presence of the inhA promoter mutation while in absence of the katG mutation.15 Therefore, the combined use of PAS and INH may bring additional benefit for treating TB patients infected with mutants of low level INH resistance. Consistent with our observation, a clinical trial from Ukraine has demonstrated that the regimens with PAS for previously treated TB cases resulted in faster time to sputum conversion and better clinical outcomes than the regimens without PAS.16 On the other hand, patients can be divided into slow or rapid inactivators on the basis of the rate of acetylation of INH,17 which is associated with the diversity of serum INH levels. The rapid acetylators may experience treatment failure due to low INH levels.17 Previous studies have confirmed that the coadministration of PAS with INH could increase plasma levels of INH.6 In addition to the above-mentioned advantage, our finding on the synergy between INH and PAS suggests that this anti-TB drug combination would improve the effectiveness of INH against MTB for the rapid acetylators. Further clinical trials are urgently needed to confirm the in vitro potency of INH-PAS combination.
In conclusion, our data demonstrate that PAS shows the synergistic effect against non-MDR, MDR and XDR-TB when in combination with INH. Although the exact explanation for this synergy remains unclear, our results will provide clinicians with useful hints to reuse this combination for treatment of TB patients in clinical practice.
Acknowledgments
This work was supported by Beijing Talents Foundation (2017000021223ZK39), the Beijing Municipal Admini-stration of Hospitals’ Youth Programme (QML20-171601), Beijing Municipal Administration of Hospitals’ Ascent Plan (DFL20181602), Beijing Municipal Administration of Hospitals Clinical Medicine Development of Special Funding Support (ZYLX201809). The authors thank the staff from the National Clinical Laboratory on Tuberculosis for their technical assistance.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Rustomjee R, Diacon AH, Allen J, et al. Early bactericidal activity and pharmacokinetics of the diarylquinoline TMC207 in treatment of pulmonary tuberculosis. Antimicrob Agents Chemother. 2008;52(8):2831–2835. doi:10.1128/AAC.01204-07
2.
3. Sotgiu G, Nahid P, Loddenkemper R, Abubakar I, Miravitlles M, Migliori GB. The ERS-endorsed official ATS/CDC/IDSA clinical practice guidelines on treatment of drug-susceptible tuberculosis. Eur Respir J. 2016;48(4):963–971. doi:10.1183/13993003.01356-2016
4. van Den Boogaard J, Kibiki GS, Kisanga ER, Boeree MJ, Aarnoutse RE. New drugs against tuberculosis: problems, progress, and evaluation of agents in clinical development. Antimicrob Agents Chemother. 2009;53(3):849–862. doi:10.1128/AAC.00749-08
5. Minato Y, Thiede JM, Kordus SL, McKlveen EJ, Turman BJ, Baughn AD. Mycobacterium tuberculosis folate metabolism and the mechanistic basis for para-aminosalicylic acid susceptibility and resistance. Antimicrob Agents Chemother. 2015;59(9):5097–5106. doi:10.1128/AAC.00647-15
6. Jennne JW. Partial purification and properties of the isoniazid transacetylase in human liver. Its relationship to the acetylation of p-aminosalicylic acid. J Clin Invest. 1965;44(12):1992–2002. doi:10.1172/JCI105306
7.
8. Zhang L, Pang Y, Yu X, et al. Linezolid in the treatment of extensively drug-resistant tuberculosis. Infection. 2014;42(4):705–711. doi:10.1007/s15010-014-0632-2
9. Pang Y, Tan Y, Chen J, et al. Diversity of nontuberculous mycobacteria in eastern and southern China: a cross-sectional study. Eur Respir J. 2017;49(3). doi:10.1183/13993003.01429-2016.
10. Zou L, Liu M, Wang Y, Lu J, Pang Y. Determination of in vitro synergy between linezolid and other antimicrobial agents against Mycobacterium tuberculosis isolates. Tuberculosis (Edinb). 2015;95(6):839–842. doi:10.1016/j.tube.2015.07.003
11. Gandhi NR, Nunn P, Dheda K, et al. Multidrug-resistant and extensively drug-resistant tuberculosis: a threat to global control of tuberculosis. Lancet. 2010;375(9728):1830–1843. doi:10.1016/S0140-6736(10)60410-2
12. Lounis N, Maslo C, Truffot-Pernot C, Grosset J, Boelaert RJ. Impact of iron loading on the activity of isoniazid or ethambutol in the treatment of murine tuberculosis. Int J Tuberc Lung Dis. 2003;7(6):575–579.
13. Velayati AA, Farnia P, Ibrahim TA, et al. Differences in cell wall thickness between resistant and nonresistant strains of Mycobacterium tuberculosis: using transmission electron microscopy. Chemotherapy. 2009;55:303–307. doi:10.1159/000226425
14. Zhang Y, Yew WW. Mechanisms of drug resistance in Mycobacterium tuberculosis: update 2015. Int J Tuberc Lung Dis. 2015;19(11):1276–1289. doi:10.5588/ijtld.15.0389
15. Xu Y, Wu J, Liao S, Sun Z. Treating tuberculosis with high doses of anti-TB drugs: mechanisms and outcomes. Ann Clin Microbiol Antimicrob. 2017;16(1):67. doi:10.1186/s12941-017-0239-4
16. Byalik IB, Tsygankova LM, Davidenko VV, Slouch IV. Results of PAS application in complex chemotherapy of patients with destructive, ineffectively treated previously resistant pulmonary tuberculosis. Ukr Pulmonol J. 2006;1(1):56–59.
17. Mitchison D, Davies G. The chemotherapy of tuberculosis: past, present and future. Int J Tuberc Lung Dis. 2012;16(6):724–732. doi:10.5588/ijtld.12.0083
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.