Back to Journals » Journal of Pain Research » Volume 13
P2Y2 Receptors Mediate Masseter Muscle Mechanical Hypersensitivity in Rats
Authors Tariba Knežević P ![]() , Vukman R
, Vukman R ![]() , Uhač M
, Uhač M ![]() , Illeš D
, Illeš D ![]() , Kovačević Pavičić D
, Kovačević Pavičić D ![]() , Simonić-Kocijan S
, Simonić-Kocijan S ![]()
Received 24 November 2019
Accepted for publication 7 May 2020
Published 3 June 2020 Volume 2020:13 Pages 1323—1333
DOI https://doi.org/10.2147/JPR.S239831
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor E Alfonso Romero-Sandoval
Petra Tariba Knežević,1,2 Robert Vukman,1,2 Mia Uhač,3 Davor Illeš,4 Daniela Kovačević Pavičić,1,2 Sunčana Simonić-Kocijan1,2
1Department of Prosthodontics, Faculty of Dental Medicine, University of Rijeka, Rijeka, Croatia; 2Department of Prosthodontics, Clinic of Dental Medicine, Clinical Hospital Center Rijeka, Rijeka, Croatia; 3Department of Orthodontics, Clinic of Dental Medicine, Clinical Hospital Center Rijeka, Rijeka, Croatia; 4Department of Removable Prosthodontics, School of Dental Medicine, University of Zagreb, Zagreb, Croatia
Correspondence: Sunčana Simonić-Kocijan
Department of Prosthodontics, Faculty of Dental Medicine, University of Rijeka, Krešimirova 40, Rijeka 51000, Croatia
Tel +385 51 345 633
Fax +385 51 345 630
Email [email protected]
Purpose: P2Y2 receptors (P2Y2Rs) are among the various receptors that play an important role in nociception. The goal of this research was to investigate possible P2Y2R expression changes in the trigeminal ganglion (TRG) in bilateral masseter muscle (MM) hypersensitivity following unilateral MM inflammation. The impact of unilateral intramasseteric administration of P2Y2R antagonist on bilateral MM hypersensitivity was also explored.
Materials and Methods: Bilateral MM hypersensitivity was provoked by unilateral intramasseteric injection of complete Freund’s adjuvant (CFA). The head withdrawal threshold (HWT) was assessed bilaterally 4 days later. Bilateral TRG and MM isolation were followed, and quantitative real-time polymerase chain reaction (qRT-PCR) and histopathological analysis were carried out on these tissues, respectively. The involvement of P2Y2Rs in nocifensive behavior was evaluated by administering two doses of P2Y2R antagonist AR-C118925 (0.2 or 1 mg/100 μL) in inflamed MM 4 days post-CFA administration. Bilateral HWT was assessed at different time points following antagonist injection.
Results: qRT-PCR analysis demonstrated P2Y2R up-regulation in TRG ipsilateral to the site of CFA administration. Compared to the controls, both doses of AR-C118925 injected ipsilateral to the TRG increased the bilateral HWT at 30, 60, 90, and 120 minutes after antagonist administration.
Conclusion: The findings suggest that P2Y2Rs may affect MM inflammatory hypersensitivity owing to its up-regulation in the TRG in MM inflammatory pain states.
Keywords: facial pain, masticatory muscles, temporomandibular disorders, trigeminal ganglion
Plain Language Summary
The pathophysiology of masticatory muscle (MM) pain in temporomandibular disorders (TMDs) remains unclear. Many studies have established the importance of various receptors in the pathophysiology of pain conditions, and P2Y2 receptors (P2Y2Rs) are among them. The goal of this study was to establish whether P2Y2Rs mediate pain transmission from MM in rats. This study involved a behavioral assessment in rats along with laboratory analysis of MM tissue and trigeminal ganglion (TRG) tissue. The TRG mediates pain transmission originating from the MM. Rats were unilaterally injected with complete Freund’s adjuvant (CFA), a reagent that develops inflammation in the MM. A behavioral assessment was performed 4 days later and found bilateral MM mechanical hypersensitivity. Histopathological evaluation confirmed the development of inflammation in CFA-injected MM, and qRT-PCR showed that P2Y2R was up-regulated in the TRG ipsilateral to the CFA injection site. In addition, the effect of two different doses of AR-C118925 on bilateral mechanical hypersensitivity was evaluated. AR-C118925 is a selective antagonist of P2Y2Rs. CFA-injected rats received an injection of AR-C118925 into the same MM, and behavioral assessment followed 15, 30, 60, 90, and 120 minutes after the antagonist injection. Both doses of AR-C118925 resulted in a significant reduction in bilateral mechanical hypersensitivity at 30, 60, 90, and 120 minutes after injection of the antagonist. These results suggest that P2Y2Rs may affect MM inflammatory hypersensitivity because they are up-regulated in TRG in MM inflammatory pain conditions. Their blockage with an antagonist is likely to reduce mechanical hypersensitivity.
Introduction
Temporomandibular disorders (TMDs) are a set of masticatory muscle and/or temporomandibular joint conditions, and muscle pain is the primary complaint in TMD patients.1,2 TMD afflict 5–12% of the population, primarily the working population between 20 and 40 years of age. This represents a severe public health issue due to the physical and functional limitations, psychosocial discomfort and reduced quality of life reported by these patients.3,4 Previous scientific studies in this area have focused on the relationship between multiple etiological factors and TMD onset. Recent studies have concentrated on the mechanism changes related to pain that are present in TMD, and they have highlighted the crucial role of ion channels and receptors within these pain mechanisms.5–7
Neurons of the trigeminal ganglion (TRG) are a peripheral component of the orofacial pain transmission pathway. These neurons express various receptors such as purinergic receptors, P2R, transient receptor potential vanilloid 1, TRPV1, gamma-aminobutyric acid, GABA(B), voltage-gated potassium channel KCNQ, acid sensing ion channel, ASIC. The importance of various receptors in pain transmission mechanisms has been confirmed in many recent studies that have suggested receptor up-regulation or down-regulation in pain conditions.8–12
The P2Rs include the P2Y receptor (P2YR) and P2X receptor families (P2XR), and both families are involved in pain transmission. The most important ligands for these receptors are uridine triphosphate (UTP) and/or adenosine triphosphate (ATP), which are delivered from intracellular sources during inflammation and in various types of tissue damage. ATP and UTP bind to P2XRs and/or P2Yrs, leading to their activation and subsequent intracellular cascade reactions in different physiological and pathophysiological processes, including pain transmission.13,14 Functions of P2XRs and P2YRs have not been thoroughly investigated. The role of P2XRs remains unclear, and there are even fewer studies concerning the role of P2YRs. The P2YR family includes eight G protein-coupled receptors that play crucial roles in intracellular pathways. Their actions involve G protein binding and subsequent ionic conductance and/or second messenger system activation, resulting in a longer response time compared to P2XRs. The P2YR family also plays an essential part in regulating the activity of membrane voltage-gated ion channels.15–17
Many different cell types express P2Y2 receptors (P2Y2Rs). According to a recent study, the constitutive P2Y2Rs activity produces intracellular Ca2+ tone and suppresses basal lipolysis via an adenylate cyclase-dependent mechanism in human adipocytes.18 Muoboghare et al have suggested that UTP acts via P2Y2Rs to mobilise Ca2+ in human endothelial cells.19 Recent studies have investigated the role of P2Y2Rs in pain mechanisms, but their exact function remains unclear, especially in the fields of orofacial pain and TMD. P2Y2Rs (along with P2Y1 receptors) are the major P2YRs expressed in small-diameter sensory neurons.15 P2Y2R stimulation increases neuronal excitability through Kv7 voltage-gated potassium channel inhibition and TRPV1 facilitation.20 Previous results suggest that P2Y2R inhibition enhances potassium channel expression and subsequent analgesia development in trigeminal neuropathic pain and that P2Y2R activation is related to inflammatory mechanical allodynia.21,22 There is also evidence that P2Y2R take part in the hyperalgesia mechanism by sensitizing ASICs in primary sensory neurons.23 Furthermore, a recent study established the involvement of glial P2Y2Rs in pain.8 Together, these findings suggest that P2Y2 receptors play a very important function in nociception mechanisms.
Several recent studies have reported the involvement of different receptors in inflammatory pain states in the orofacial region. Therefore, we hypothesized that the onset of bilateral masseter muscle (MM) hypersensitivity following unilateral MM inflammation involves, among other receptor changes, P2Y2R expression enhancement in TRG neurons. We also surmised that unilateral administration of a P2Y2R antagonist into the MM could attenuate the bilateral hypersensitivity. The aim of this study was to define possible P2Y2R changes in the TRG during bilateral hypersensitivity following unilateral MM inflammation. The impact of an intramasseteric injection of P2Y2R antagonist on bilateral nocifensive behavior was also investigated.
Materials and Methods
Experimental Animals
Experiments were conducted using 54 Wistar rats (n=54). The animal characteristics, housing protocols, and treatment protocols are described in detail in our previous study.24 Briefly, adult male rats weighing 250–300g were housed in groups of two to three per cage under a 12h/12h light/dark cycle. The rats were housed in a controlled temperature (22±2°C) and humidity (55±10%) environment with food and water access ad libitum. All efforts were made to ensure the minimal number of animals used in the experiment and minimal animal suffering.
All experimental protocols were approved and conducted in accordance with the institutional Ethics Committee, the Croatian national laws and rules (Official Gazette 135/06, 37/13, and 55/13), the European Community Council Directive (86/609/EEC), the Cabinet for Veterinary and Food Safety within the Croatian Ministry of Agriculture, and the Faculty of Medicine Ethics Committee, University of Rijeka.
Prior to any animal manipulation, all rats were housed in the controlled environment for a minimum of 7 days to eliminate stress-related influences.
The experiment was performed in two phases (Figure 1). Phase one involved the induction of unilateral MM inflammation in rats, followed by bilateral nocifensive behavioral measurements (n=16), bilateral TRG and MM tissue isolation, and subsequent quantitative real-time polymerase chain reaction (qRT-PCR) and histopathological examination of these tissues, respectively (Figure 1A). Phase two included examining the effect of unilateral P2Y2R antagonist injection on the nocifensive behavior in sham rats (Figure 1B) and in unilateral MM inflammation-induced rats (Figure 1C) (n=38).
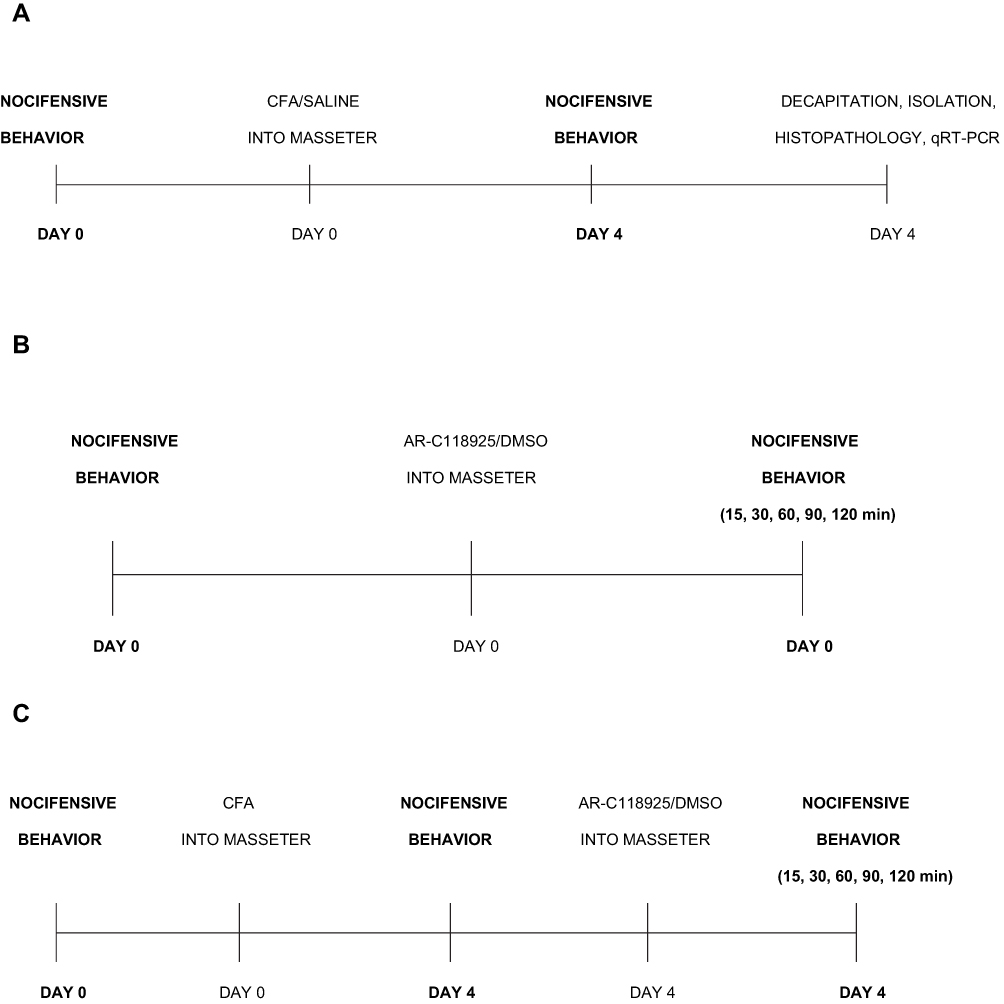 |
Figure 1 Experimental design timelines. (A) Phase one of the experiment – evaluation of CFA injection on nocifensive behavior, masseter muscle (MM) tissue histopathology and TRG tissue qRT-PCR. (B) Phase two – evaluation of AR-C118925/DMSO injection on nocifensive behavior in sham rats. (C) Phase two – evaluation of AR-C118925/DMSO administration on nocifensive behavior in unilateral MM inflammation-induced rats. Abbreviations: CFA, complete Freund’s adjuvant; HWT, head withdrawal threshold; MM, masseter muscle; TRG, trigeminal ganglion; qRT-PCR, quantitative real-time polymerase chain reaction; DMSO, dimethyl sulfoxide.f |
Establishment of Bilateral Hypersensitivity Following Unilateral MM Inflammation
MM inflammation was induced by unilateral injection of complete Freund’s adjuvant (CFA, Sigma F5881, Sigma-Aldrich, Saint Louis, MO, USA; 0.5 mg/mL, heat-killed Mycobacterium tuberculosis suspended in 1:1 oil: saline emulsion), as reported in previous studies.24–26 The protocol details are described in our previous study.24 Briefly, rats were anesthetized with 4% isoflurane (Forane, Abbott Laboratories Ltd, Queenborough, UK) in a gas mixture O2:N2=1:2 to abolish righting and corneal reflexes. They were then injected with 50 μL of CFA (experimental group) or 0.9% saline (control group) in the right MM mid-region. The precise administration site was determined by MM palpation between the zygomatic bone and mandible. After contacting the mandible bone, the needle was placed into the MM mid-region, and the CFA/saline was administered via a 27-gauge needle within 5–10 seconds. All solutions were freshly prepared before administration. The animals were monitored daily for evidence of edema after the CFA/saline injection. MM inflammation development was examined histopathologically, and nocifensive behavior measurements followed to confirm the development of bilateral allodynia. The inflammation and control groups each contained 8 animals (ninflammation=8) (ncontrol=8).
Mechanical Nocifensive Behavior Measurements
A nocifensive behavioral response to the mechanical threshold that previously did not result in a nocifensive response was considered as a sign of mechanical hypersensitivity. The previously described rat head withdrawal threshold (HWT) method was used to evaluate nocifensive behavioral changes.25–28 The method details are reported in our previous study.24 Briefly, a von Frey anesthesiometer (VFA; type 2391, IITC Inc., Woodland Hills, CA, USA) was used bilaterally at different time points to establish the nocifensive response. The lowest force needed to cause active head withdrawal from the 1.0 mm probing tip was considered to be the HWT. The probing tip attached to the VFA was touched against the MM mid-region five times during one minute intervals, and the average value of these five measurements was defined as the HWT value. Before any VFA measurements were conducted, the rats were accustomed to stand uninhibited on the experimenter’s glove. Mechanical thresholds for evoking the head withdrawal responses were measured by bilateral MM probing before and on day 4 after CFA or saline injection, according to the results of a previous study.25 A baseline HWT was assessed prior to the CFA administration. Additional behavioral measurements were carried out after P2Y2R antagonist injection, as described further below. All behavioral measurements were performed by researchers blinded to the study protocol.
Tissue Isolation, Preparation, and Analysis
The final nocifensive behavioral measurements were performed 4 days post-CFA/saline injection, and rat decapitation followed. Bilateral MMs were dissected for histopathological analysis. Bilateral TRGs were harvested for qRT-PCR analysis of P2Y2 mRNA expression.
All tissue samples were frozen in liquid nitrogen and stored at −80°C until processed.
The histopathological MM tissue analysis was performed as reported in our previous study.24 Briefly, the entire both right and left MMs were fixed in 4% paraformaldehyde, embedded in paraffin, cut at 40 μm intervals, and stained with hematoxylin and eosin.
Quantitative real-time PCR was performed as follows. TRG tissue samples 1 cm2 in size were frozen in 500 μL of RNAlater (Ambion, Austin, TX, USA) at −80°C. The tissue samples were homogenized using a MagNA Lyser instrument (Roche Life Sciences, Mannheim, Germany). Total RNA was isolated using a NucleoSpin RNA kit (Macherey-Nagel, Duren, Germany) according to the manufacturer’s instructions. A Qubit 3.0 instrument and Qubit RNA Broad Range reagent (Life Technologies, Carlsbad, CA, USA) were used for determining the total RNA concentration. Transcription was performed using a High Capacity cDNA kit (Applied Biosystems, Foster City, CA, USA) with 500 ng of total RNA. cDNA libraries were stored at −25°C prior to the quantification process. Random hexamer primers provided with the kit were used for reverse transcription. Specific mRNA expression levels for the P2Y2 gene were defined using a 7500 Fast Real-Time PCR (Applied Biosystems). The primers used for qRT-PCR were from the Taqman Gene Expression Assay (Applied Biosystems) for P2Y2 (Rn00568476_m1) and for the housekeeping genes GAPDH (Rn01775763_g1) and beta-actin (Rn00667869_m1). The P2Y2 mRNA relative expression levels were determined by the 2-ΔΔCt method to compare its expression with those of the housekeeping genes.24,29,30
P2Y2 Receptor Antagonist Administration in Sham Animals
The impact of an intramasseteric injection of the competitive and selective P2Y2R antagonist AR-C118925 (Tocris Bioscience, St Louis, MO, USA) on the baseline HWT was evaluated.24,25 The rats were lightly anesthetized with 2% isoflurane in an O2:N2=1:2 mixture.24 AR-C118925 was then injected at a dose of either 0.2 mg/100 μL (Dlow, n=6) or 1 mg/100 μL (Dhigh, n=6). The volumes and doses of AR-C118925 were defined taking into consideration findings from a recent study.8 The same volume (100 μL) of dimethyl sulfoxide (DMSO)(Sigma-Aldrich) was administered to the control group animals (n=6). Behavioral measurements were performed prior to the AR-C118925/DMSO injection and at 15, 30, 60, 90, and 120 minutes after the AR-C118925/DMSO administration.24,25,31
P2Y2 Receptor Antagonist Administration in the Inflammation Group
The impact of AR-C118925 or DMSO administration into the MM on bilateral HWT in animals with CFA-induced unilateral MM inflammation (the inflammation group) was subsequently estimated.8,24,25 The inflammation induction and antagonist administration protocols are reported in our previous study.24 Briefly, the baseline HWT was evaluated. The rats were subsequently anesthetized with 4% isoflurane and unilaterally injected with CFA. On day 4 after the intramasseteric CFA injection, bilateral HWT was measured to confirm the development of bilateral mechanical allodynia. After the behavioral evaluation, each animal was lightly anesthetized with 2% isoflurane and AR-C118925 was injection at a dose of either 0.2 mg/100 μL (Dlow, n=6) or 1 mg/100 μL (Dhigh, n=6). Control animals were injected with 100 μL of DMSO (n=6). The volumes and doses of AR-C118925 were defined taking into consideration the results of a recent study.8 All solutions were freshly prepared before use. P2Y2 antagonist was injected unilaterally to the mid-region of the right MM, as described above. The HWT was measured 15, 30, 60, 90, and 120 minutes after the AR-C118925/DMSO injection.8,24,25,31
Statistical Analysis
The results were analyzed using SPSS 21.0 software (SPSS Inc., Chicago, USA). The data are presented as the means±standard error of the mean (SEM). The Kolmogorov–Smirnov test was used to test the normality of distribution of all measurements. The independent samples t-test was utilized to establish the differences in nocifensive behavior before and after the CFA/saline injection. The one sample t-test was used to analyze the differences in P2Y2 mRNA expression between the two groups. Mechanical HWT measurements between the inflammation and control groups injected with AR-C118925/DMSO were assessed using mixed ANOVA, with time points as a within-subjects factor and the group as a between-subjects factor. Significance was defined as p< 0.05. The results are presented graphically.
Results
CFA Injection Caused Ipsilateral Inflammation in the Rat MM
The unilateral administration of CFA into the right MM mid-region produced redness and edema around the site of administration. MM contralateral to the site of CFA administration and bilateral MMs in control rats showed no signs of inflammation.
The histological signs of massive granular leukocyte infiltration and the large number of vacuoles present in the CFA-administered MM unequivocally confirmed the development of inflammation (Figure 2A). Neither the contralateral MM in CFA-administered animals (Figure 2B) nor the MMs of control rats showed any histological signs of inflammation (Figure 2C).
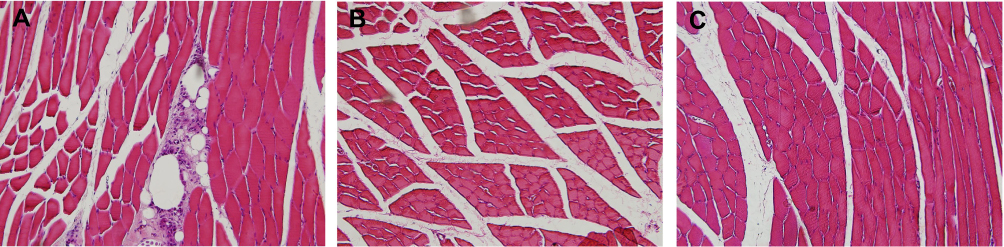 |
Figure 2 Histopathological analysis of the masseter muscle (MM) tissue (hematoxylin and eosin staining, 20x). (A) Complete Freund’s adjuvant (CFA)-injected MM presents signs of inflammation. (B) MM contralateral to CFA injection shows no signs of inflammation. (C) MM of control rats shows no signs of inflammation. Abbreviations: CFA, complete Freund’s adjuvant; MM, masseter muscle. |
MM Inflammatory Hypersensitivity Was Related to Up-Regulated P2Y2 Expression in the TRG
qRT-PCR analysis of the P2Y2 receptors mRNA expression levels in bilateral TRGs isolated on day 4 after unilateral intramasseteric inflammation induction showed statistically significant enhancement of P2Y2 mRNA expression in the TRG ipsilateral to inflamed MM compared to the control group (p=0.011) (Figure 3A). In the contralateral MMs, no significant difference in P2Y2 mRNA expression was detected in comparison to the control animals (p=0.647) (Figure 3B).
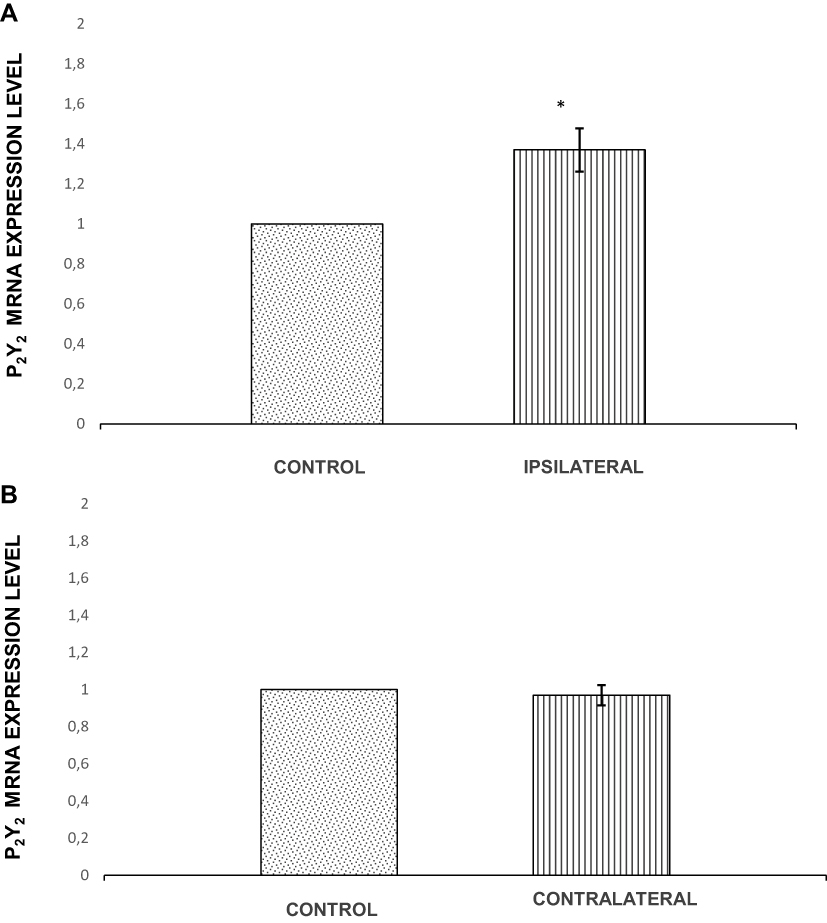 |
Figure 3 Relative levels of P2Y2 mRNA expression in the trigeminal ganglion (TRG). (A) Relative levels of P2Y2 mRNA expression in the TRG ipsilateral to complete Freund’s adjuvant (CFA) injection. (B) Relative levels of P2Y2 mRNA expression relative levels in the TRG contralateral to CFA administration. Note: *p˂0.05. Abbreviations: TRG, trigeminal ganglion; CFA, complete Freund’s adjuvant. |
Unilateral Intramasseteric Administration of AR-C118925 Had No Effect on Baseline HWT in Sham Animals
To rule out the possible effect of the antagonist injection on the baseline HWT, AR-C118925 or DMSO were unilaterally injected into the MMs of sham rats. The tested doses of AR-C118925 (Dlowand Dhigh) and DMSO were unable to provoke any changes to the baseline HWT in sham rats at any observed time point (15, 30, 60, 90, and 120 after injection) (Figure 4A and B). Neither ipsilateral nor contralateral differences were established between groups at any observed time point (Figure 4A and B).
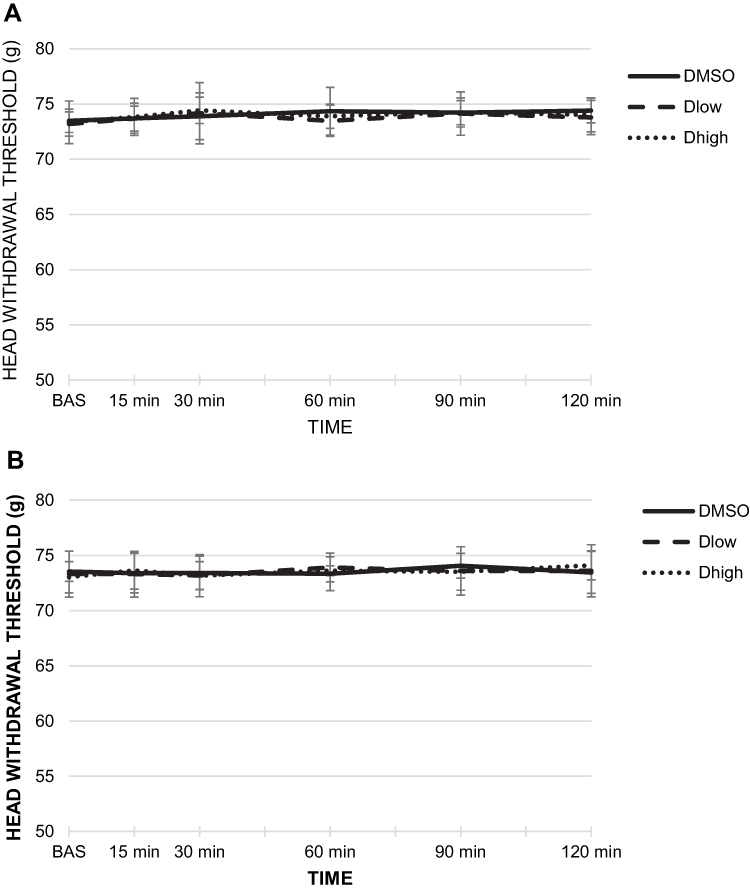 |
Figure 4 The effect of unilateral intramasseteric administration of AR-C118925 or dimethyl sulfoxide (DMSO) on baseline head withdrawal threshold (HWT). (A) The effect on baseline HWT of AR-C118925/DMSO ipsilateral injection. (B) The effect on baseline HWT of AR-C118925/DMSO contralateral injection. Abbreviations: DMSO, dimethyl sulfoxide; HWT, head withdrawal threshold. |
Unilateral Intramasseteric Administration of AR-C118925 Alleviated Bilateral MM Hypersensitivity in Inflammation Group of Animals
AR-C118925 was unilaterally injected into the MM to examine the relation between P2Y2R changes and bilateral MM hypersensitivity following unilateral MM inflammation.
All animals that received unilateral CFA injection (inflammation group) showed a significant bilateral reduction in the baseline HWT values 4 days after the CFA injection. This finding reaffirmed the development of bilateral hypersensitivity (p≤0.002) (Figure 5A and B).
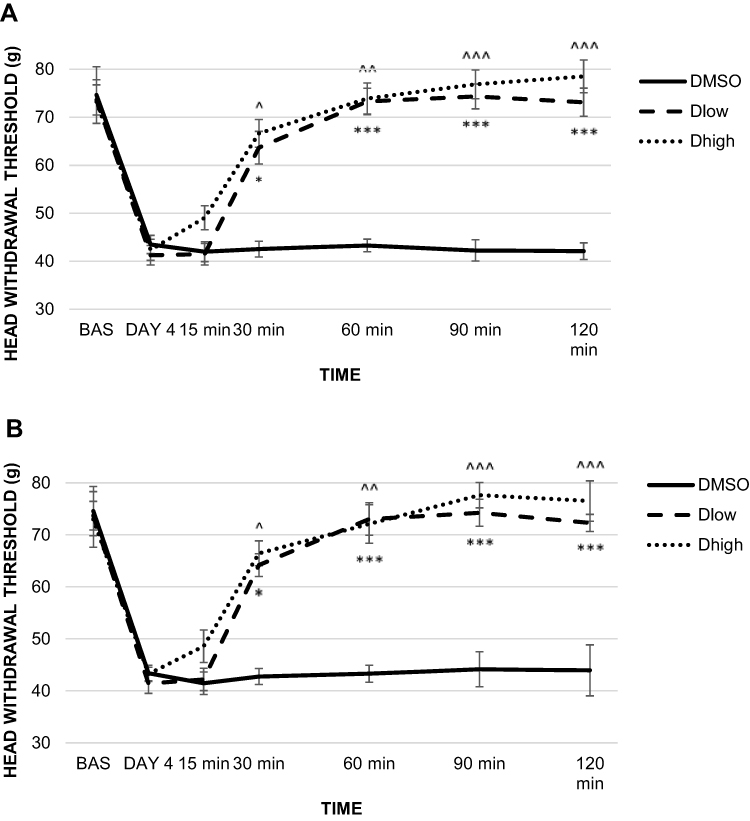 |
Figure 5 The effect of unilateral administration of AR-C118925 or dimethyl sulfoxide (DMSO) on head withdrawal threshold (HWT) in rats injected with complete Freund’s adjuvant (CFA). (A) The effect on HWT of ipsilateral CFA and AR-C118925/DMSO injection. (B) The effect on HWT of contralateral CFA and AR-C118925/DMSO injection. Notes: *p˂0.05, ***p≤0.001 for Dlow group, ^p˂0.05, ^^p≤0.01, ^^^p≤0.001 for Dhigh group. Abbreviations: DMSO, dimethyl sulfoxide; HWT, head withdrawal threshold; CFA, complete Freund’s adjuvant; Dlow, 0.2 mg/100 μL dose of AR-C118925; Dhigh, 1 mg/100 μL dose of AR-C118925. |
AR-C118925 injected ipsilateral to the CFA injection site resulted in a statistically significant increase in ipsilateral HWT in both the Dlow and Dhigh inflammation groups compared to the DMSO group at 30, 60, 90, and 120 minutes after the AR-C118925/DMSO injection (p≤0.001). HWT measurement values obtained 15 minutes after antagonist administration showed no significant pain attenuation in the Dlow and Dhigh groups (Figure 5A).
Contralateral to CFA and AR-C118925/DMSO administration site, the statistical test revealed significant HWT increase in Dlow and Dhigh inflammation groups compared to controls 30, 60, 90, and 120 minutes after AR-C118925/DMSO injection (p≤0.001). No significant HWT increase was observed 15 minutes after antagonist injection in both the Dlow and Dhigh groups (Figure 5B).
Discussion
The role of P2Y2Rs in bilateral muscular orofacial pain is poorly understood. Furthermore, the mechanism involved in pain transmission and the expression pattern of pain receptors in neurons in the dorsal root ganglion (DRG) and the TRG differ according to pain location.31,33 Therefore, this experimental design was established to assess the possible changes in P2Y2R expression in bilateral MM inflammatory hypersensitivity in the TRG and to test the effect of selective and competitive P2Y2R antagonist AR-C118925 on the nocifensive behavior induced by unilateral injection of CFA into the MM.
Some studies have suggested the involvement of P2Y2Rs in trigeminal pain, although their role is not yet clear. Ando et al have reported that the administration of UTP, a potent P2Y2R agonist, resulted in mechanical allodynia attenuation in a bilateral neuropathic pain model, but it had only ipsilateral analgesic effect in plantar acute pain model.34 Conversely, another study found that the administration of P2Y2R antagonist AR-C118925 was followed by a complete blockage of satellite glial cell activation and had a long-lasting analgesic effect in facial allodynia induced by CFA administration into the temporomandibular joint.8 A recent study on chronic constriction injury of the infraorbital nerve as a neuropathic trigeminal pain model showed dose- and time-dependent analgesic effects of P2Y2R antagonists.21
Several studies have reported P2Y2R expression changes in inflammatory pain states that affect different body parts, but no study has investigated TRG P2Y2R expression levels in an inflammatory MM pain model. Malin et al found that P2Y2Rs are the only Gq P2YR that are up-regulated in DRG neurons in inflammatory conditions.35 Magni et al established an in vivo inflammatory temporomandibular joint pain model that resulted in satellite glial cell activation and enhanced P2Y2R expression ipsilateral to the inflammation site.8 As mentioned above, the expression patterns of pain receptors in the TRG and DRG and the pain transmission pathways seems to vary depending on the site of pain onset.32,33 However, no study has yet investigated the expression of P2Y2Rs in the TRG using an inflammatory MM model. Our qRT-PCR analysis revealed a statistically significant up-regulation of P2Y2R mRNA expression in the TRG ipsilateral to MM inflammation, suggesting a direct relationship between MM inflammation and ipsilateral increased P2Y2R expression (ie ipsilateral hypersensitivity and increased P2Y2R expression are positively correlated). The lack of P2Y2R mRNA expression changes contralateral to the MM inflammation suggests that P2Y2Rs are not likely to have a direct impact on the development of peripheral non-inflammatory pain.
Behavioral measurements were performed before and after CFA and AR-C118925 were injected to assess possible changes in pain sensation. In accordance with recent studies, AR-C118925 was chosen for blockage of the up-regulated P2Y2Rs.8,18,19,36 The results of the current study showed attenuation of bilateral hypersensitivity after AR-C118925 was administered ipsilateral to the inflammation. This suggests that the analgesic effect of AR-C118925 on ipsilateral MM hypersensitivity may be mediated by its effect on P2Y2Rs. A possible contralateral hypersensitivity mechanism implicates P2Y2R up-regulation ipsilateral to the site of inflammation, followed by a second messengers cascade and activation of certain central nervous system (CNS) structures, resulting in contralateral hypersensitivity. As previous studies suggest the importance of satellite glial cells in pain transmission, neuron-glia interaction may also be taken into consideration in explaining a contralateral hypersensitivity mechanism.37,38 Neuron-glia interaction in the TRG possibly has a crucial impact on neuronal and glial function within CNS structures in increased and abnormal afferent activity in orofacial inflammatory nociception processes. There is evidence that non-synaptically released chemical mediators from neurons and satellite glia may induce chronic pain via autocrine and/or paracrine mechanisms and that augmented excitability of primary afferent neurons results in changes in central pain-signaling neurons (central sensitization).37 Our results show that AR-C118925 injection ipsilateral to inflammation leads to the inhibition of contralateral hypersensitivity.
Taking together, our results indicate that P2Y2Rs are important in hypersensitivity onset that occurs both ipsilateral and contralateral to the site of inflammation. Our findings show that inflammation can trigger up-regulated P2Y2R expression in primary neurons ipsilateral, but not contralateral to the inflammation. The increased P2Y2R expression can be directly related to ipsilateral hypersensitivity. It is possible that a neuronal cascade induces a contralateral hypersensitivity response that is not directly connected to changes in P2Y2R expression in primary neurons.
There are some limitations of this study. The role of P2Y2Rs was investigated only at TRG level solely (and not at the level of MM or CNS structures). In addition, the P2Y2R expression levels were assessed in the entire TRG, while only a limited number of neurons innervating the MM were influenced by the CFA injection.
Conclusions
Our findings indicate that P2Y2Rs may functionally influence MM hypersensitivity. P2Y2R up-regulation in TRG neurons ipsilateral to MM inflammation appears to be involved in ipsilateral inflammatory MM hypersensitivity. By contrast, contralateral hypersensitivity shows a non-inflammatory phenomenon that is not likely to be directly related to changes in P2Y2R expression.
Data Sharing Statement
The data and materials, as well as the raw data, are available by contacting the corresponding author.
Acknowledgments
The authors thank members of the Department of Pharmacology and the Department of Anatomy at the Faculty of Medicine of the University of Rijeka for their technical support, assistance in rat housing and surgery, and aid in the histopathological analysis. We particularly thank associate professor Kristina Pilipović for writing assistance and suggestions.
Author Contributions
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; gave final approval of the version to be published; and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. McNeill C, Danzig WM, Farrar WB, et al. Position paper of the American Academy of Craniomandibular Disorders. Craniomandibular (TMJ) disorders – the state of art. J Prosthet Dent. 1980;44(4):434–437. doi:10.1016/0022-3913(80)90104-3
2. Dworkin SF, LeResche L. Research diagnostic criteria for temporomandibular disorders: review, criteria, examinations and specifications, critique. J Craniomandib Disord. 1992;6(4):301–355.
3. Bäck K, Hakeberg M, Wide U, Hange D, Dahlström L. Orofacial pain and its relationship with oral health-related quality of life and psychological distress in middle-aged women. Acta Odontol Scand. 2019;1–7.
4. Haviv Y, Zini A, Etzioni Y, et al. The impact of chronic orofacial pain on daily life: the vulnerable patient and disruptive pain. Oral Surg Oral Med Oral Pathol Oral Radiol. 2017;123(1):58–66. doi:10.1016/j.oooo.2016.08.016
5. Quinelato V, Bonato LL, Vieira AR, Granjeiro JM, Tesch R, Casado PL. Association between polymorphisms in the genes of estrogen receptors and the presence of temporomandibular disorders and chronic arthralgia. J Oral Maxillofac Surg. 2018;76(2):
6. McIlwrath SL, Nesemeier R, Ma F, Oz HS, Zhang L, Westlund KN. Inflammatory ‘double hit’ model of temporomandibular joint disorder with elevated CCL2, CXCL9, CXCL10, RANTES and behavioral hypersensitivity in TNFR1/R2-/- mice. Eur J Pain. 2017;21(7):1209–1223. doi:10.1002/ejp.1021
7. Nascimento TD, Yang N, Salman D, et al. µ-opioid activity in chronic TMD pain is associated with COMT polymorphism. J Dent Res. 2019;98(12):1324–1331. doi:10.1177/0022034519871938
8. Magni G, Merli D, Verderio C, Abbracchio MP, Ceruti S. P2Y2 receptor antagonists as anti-allodynic agents in acute and sub-chronic trigeminal sensitization: role of satellite glial cells. Glia. 2015;63(7):1256–1269. doi:10.1002/glia.22819
9. Sato M, Sato T, Yajima T, Shimazaki K, Ichikawa H. The transient receptor potential cation channel subfamily V members 1 and 2, P2X purinoceptor 3 and calcitonin gene-related peptide in sensory neurons of the rat trigeminal ganglion, innervating the periosteum, masseter muscle and facial skin. Arch Oral Biol. 2018;96:66–73. doi:10.1016/j.archoralbio.2018.08.012
10. Xu XX, Cao Y, Ding TT, Fu KY, Li Y, Xie QF. Role of TRPV1 and ASIC3 channels in experimental occlusal interference-induced hyperalgesia in rat masseter muscle. Eur J Pain. 2016;20(4):552–563. doi:10.1002/ejp.758
11. Bereiter DA, Thompson R, Rahman M. Sex differences in estradiol secretion by trigeminal brainstem neurons. Front Integr Neurosci. 2019;13:3. doi:10.3389/fnint.2019.00003
12. Ling J, Erol F, Viatchenko-Karpinski V, Kanda H, Gu JG. Orofacial neuropathic pain induced by oxaliplatin: downregulation of KCNQ2 channels in V2 trigeminal ganglion neurons and treatment by the KCNQ2 channel potentiator retigabine. Mol Pain. 2017;13:1744806917724715. doi:10.1177/1744806917724715
13. Lazarowski ER, Boucher RC, Harden TK. Mechanisms of release of nucleotides and integration of their action as P2X- and P2Y-receptor activating molecules. Mol Pharmacol. 2003;64(4):785–795. doi:10.1124/mol.64.4.785
14. Hockley JR, Tranter MM, McGuire C, et al. P2Y receptors sensitize mouse and human colonic nociceptors. J Neurosci. 2016;36(8):2364–2376. doi:10.1523/JNEUROSCI.3369-15.2016
15. Molliver DC, Cook SP, Carlsten JA, Wright DE, McCleskey EW. ATP and UTP excite sensory neurons and induce CREB phosphorylation through the metabotropic receptor, P2Y2. Eur J Neurosci. 2002;16(10):1850–1860. doi:10.1046/j.1460-9568.2002.02253.x
16. Jacobson KA, Balasubramanian R, Deflorian F, Gao ZG. G protein coupled adenosine (P1) and P2Y receptors: ligand design and receptor interactions. Purinergic Signal. 2012;8(3):419–436. doi:10.1007/s11302-012-9294-7
17. Abbracchio MP, Ceruti S. Roles of P2 receptors in glial cells: focus on astrocytes. Purinergic Signal. 2006;2(4):595–604. doi:10.1007/s11302-006-9016-0
18. Ali SB, Turner JJO, Fountain SJ. Constitutive P2Y2 receptor activity regulates basal lipolysis in human adipocytes. J Cell Sci. 2018;131(22):jcs221994. doi:10.1242/jcs.221994
19. Muoboghare MO, Drummond RM, Kennedy C. Characterisation of P2Y2 receptors in human vascular endothelial cells using AR-C118925XX, a competitive and selective P2Y2 antagonist. Br J Pharmacol. 2019;176(16):2894–2904. doi:10.1111/bph.14715
20. Yousuf A, Klinger F, Schicker K, Boehm S. Nucleotides control the excitability of sensory neurons via two P2Y receptors and a bifurcated signaling cascade. Pain. 2011;152(8):1899–1908. doi:10.1016/j.pain.2011.04.016
21. Li N, Lu Z, Yu L, Burnstock G, Deng X, Ma B. Inhibition of G protein coupled P2Y2 receptor induced analgesia in a rat model of trigeminal neuropathic pain. Mol Pain. 2014;10:21. doi:10.1186/1744-8069-10-21
22. Zhu H, Yu Y, Zheng L, et al. Chronic inflammatory pain upregulates expression of P2Y2 receptor in small-diameter sensory neurons. Metab Brain Dis. 2015;30(6):1349–1358. doi:10.1007/s11011-015-9695-8
23. Ren C, Gan X, Wu J, Qiu CY, Hu WP. Enhancement of acid-sensing ion channel activity by metabotropic P2Y UTP receptors in primary sensory neurons. Purinergic Signal. 2016;12(1):69–78. doi:10.1007/s11302-015-9479-y
24. Tariba Knežević P, Vukman R, Antonić R, Kovač Z, Uhač I, Simonić-Kocijan S. The role of P2X3 receptors in bilateral masseter muscle allodynia in rats. Croat Med J. 2016;57(6):530–539. doi:10.3325/cmj.2016.57.530
25. Simonić-Kocijan S, Liu W, Wu Y, Uhac I, Wang KW, Wang K. TRPV1 channel mediated bilateral allodynia induced by unilateral masseter muscle inflammation in rats. Mol Pain. 2013;9:68. doi:10.1186/1744-8069-9-68
26. Ferrara-Jr JI, de Souza ET, Franciosi AC, Toniolo EF, Dale CS. Photobiomodulation-induced analgesia in experimental temporomandibular disorder involves central inhibition of fractalkine. Lasers Med Sci. 2019;34(9):1841–1847. doi:10.1007/s10103-019-02785-6
27. Yan SD, Yang GJ, Mo SY, Liu Y, Xie QF. Effect of long-term resistance exercise on masseter muscle mechanical hyperalgesia in rats.[Article in Chinese]. Beijing Da XueXueBao Yi Xue Ban. 2019;51(1):21–27.
28. Wu YW, Hao T, Kou XX, Gan YH, Ma XC. Synovial TRPV1 is upregulated by 17-β-estradiol and involved in allodynia of inflamed temporomandibular joints in female rats. Arch Oral Biol. 2015;60(9):1310–1318. doi:10.1016/j.archoralbio.2015.05.011
29. Bhattacharya A, Vavra V, Svobodova I, Bendova Z, Vereb G, Zemkova H. Potentiation of inhibitory synaptic transmission by extracellular ATP in rat suprachiasmatic nuclei. J Neurosci. 2013;33(18):8035–8044. doi:10.1523/JNEUROSCI.4682-12.2013
30. Deiteren A, van der Linden L, de Wit A, et al. P2X3 receptors mediate visceral hypersensitivity during acute chemically induced colitis and in the post-inflammatory phase via different mechanisms of sensitization. PLoS One. 2015;10(4):e0123810. doi:10.1371/journal.pone.0123810
31. Ro JY, Lee JS, Zhang Y. Activation of TRPV1 and TRPA1 leads to muscle nociception and mechanical hyperalgesia. Pain. 2009;144(3):270–277. doi:10.1016/j.pain.2009.04.021
32. Harriott AM, Dessem D, Gold MS. Inflammation increases the excitability of masseter muscle afferents. Neuroscience. 2006;141(1):433–442. doi:10.1016/j.neuroscience.2006.03.049
33. Ambavalanar R, Moritani M, Dessem D. Trigeminal P2X3 receptor expression differs from dorsal root ganglion and is modulated by deep tissue inflammation. Pain. 2005;17(3):280–291. doi:10.1016/j.pain.2005.06.029
34. Ando RD, Mehesz B, Gyires K, Illes P, Sperlagh B. A comparative analysis of the activity of ligands acting at P2X and P2Y receptor subtypes in models of neuropathic, acute and inflammatory pain. Br J Pharmacol. 2012;159(5):1106–1117. doi:10.1111/j.1476-5381.2009.00596.x
35. Malin SA, Davis BM, Koerber HR, Reynolds IJ, Albers KM, Molliver DC. Thermal nociception and TRPV1 function are attenuated in mice lacking the nucleotide receptor P2Y2. Pain. 2008;138(3):484–496. doi:10.1016/j.pain.2008.01.026
36. Kindon N, Davis A, Dougall I, et al. From UTP to AR-C118925, the discovery of a potent non nucleotide antagonist of the P2Y2 receptor. Bioorg Med Chem Lett. 2017;27(21):4849–4853. doi:10.1016/j.bmcl.2017.09.043
37. Takeda M, Takahashi M, Matsumoto S. Contribution of the activation of satellite glia in sensory ganglia to pathological pain. Neurosci Biobehav Rev. 2009;33(6):784–792. doi:10.1016/j.neubiorev.2008.12.005
38. Sessle BJ. Peripheral and central mechanisms of orofacial inflammatory pain. Int Rev Neurobiol. 2011;97:179–206.
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.