Back to Journals » Journal of Pain Research » Volume 18
Oxymatrine Alleviates Central Post-Stroke Pain in Rats by Reducing the Inflammatory Response
Authors Li HL ![]() , Tan XP, Wang XD, Guo RT, Wang JL
, Tan XP, Wang XD, Guo RT, Wang JL ![]()
Received 29 March 2025
Accepted for publication 5 August 2025
Published 27 August 2025 Volume 2025:18 Pages 4401—4412
DOI https://doi.org/10.2147/JPR.S531280
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor E Alfonso Romero-Sandoval
Hai-Li Li,1– 3 Xing-Ping Tan,1,3 Xiao-Die Wang,1,3 Ren-Tian Guo,1,3 Jiang-Lin Wang1,3,4
1Department of Pain Management, The Affiliated Hospital, Southwest Medical University, Luzhou, People’s Republic of China; 2Department of Anesthesiology, The Affiliated Tianfu Hospital, Southwest Medical University, Meishan, People’s Republic of China; 3Anesthesiology and Critical Care Medicine Key Laboratory of Luzhou, Southwest Medical University, Luzhou, Sichuan, People’s Republic of China; 4Department of Pain Management, The Affiliated Tianfu Hospital, Southwest Medical University, Meishan, People’s Republic of China
Correspondence: Jiang-Lin Wang, Pain Department, The Affiliated Hospital, Southwest Medical University, No. 25 Pacific Street, Luzhou, 646000, People’s Republic of China, Tel +8618090880626, Fax +0830-3165469, Email [email protected]
Background and Purpose: Central post-stroke pain (CPSP) is directly caused by cerebrovascular diseases that affect the central somatosensory system. It is a serious, chronic central neuropathic pain that responds poorly to first-line drugs. Oxymatrine (OMT), a monomer derived from the traditional Chinese medicine Sophora flavescens Ait. exhibits anti-inflammatory, analgesic, and organ- and tissue-protective properties. This study aimed to investigate the therapeutic effects of OMT in a rat model of CPSP.
Methods: Gene Ontology (GO) enrichment analysis was initially employed to elucidate the role of OMT on CPSP target proteins in terms of gene function. Then the CPSP model was induced through focal hemorrhage in the ventral posterolateral nucleus (VPL). Rats were randomly assigned to five groups (n = 10): sham + NaCl, CPSP + NaCl, and CPSP + OMT (100 mg/kg, 200 mg/kg, 400 mg/kg). Daily intraperitoneal injections of NaCl or varying doses of OMT were administered from days 7 to 14 after model establishment, and paw withdrawal mechanical thresholds (PWMT) were assessed. The optimal OMT dose (200 mg/kg) was determined based on PWMT measurements. On day 14, inflammatory markers (IL-1β, IL-6, TNF-α) and alterations in microglia and astrocyte activity at the thalamic lesion site were analyzed.
Results: Elevated levels of IL-1β, IL-6, and TNF-α (P < 0.05) and activation of microglia and astrocytes were observed around the peri-thalamic lesion in CPSP rats. OMT administration significantly reduced mechanical allodynia (P < 0.05), decreased inflammatory cytokine expression (P < 0.05), and inhibited microglial and astrocytic activation.
Conclusion: OMT mitigates central post-stroke pain in rats by attenuating the inflammatory response at the thalamic lesion site.
Keywords: central post-stroke pain, oxymatrine, inflammation, thalamic lesions, mechanical allodynia
Introduction
Stroke is one of the leading causes of disability globally and the second most common cause of death.1 Post-stroke pain (PSP) is a frequent symptom, affecting up to 55% of stroke survivors. Common types of PSP include central PSP (CPSP), complex regional pain syndrome, painful spasticity, and hemiplegic shoulder pain.2,3 Often, multiple pain types coexist. CPSP is estimated to occur in 1–12% of patients with stroke, but up to 63% of these individuals may experience it in combination with other forms of PSP.3 CPSP results from cerebrovascular disease impacting the central somatosensory system, with pain localized to the area of cerebrovascular injury. It is considered a severe, chronic central neuropathic pain disorder.4 CPSP significantly impairs quality of life during post-stroke recovery and is also linked to psychological issues, such as anxiety, depression, and sleep disturbances.5,6 Presently, CPSP shows limited response to first-line clinical treatments, and physical therapy remains underutilized. Thus, exploring potential therapeutic options for CPSP is critical.
The precise pathogenesis of CPSP remains incompletely understood. Central sensitization and thalamic alterations are proposed mechanisms.7–9 Central sensitization results from central nervous system damage, triggering neurochemical, excitotoxic, and inflammatory changes that heighten neuronal excitability, leading to chronic pain. The thalamic changes hypothesis suggests that, as a key pain relay station, the thalamus plays a central role in CPSP, with thalamic injury being a major factor. Disruptions in neurotransmission, loss of inhibitory GABAergic neurons, and activation of distal microglial cells are believed to contribute to these changes following CNS injury.7 Studies have shown that in CPSP rats, microglia and astrocytes around the hematoma are activated,10–12 and the release of pro-inflammatory mediators is heightened at the peri-thalamic lesion sites.11 Thalamic inflammation is crucial in both the onset and progression of CPSP.
Oxymatrine (OMT), with the molecular formula C15H24N2O2 and a molecular mass of 264, is a quinazoline alkaloid primarily extracted from Sophora flavescens Ait., a traditional Chinese herb. It exerts various therapeutic effects, including the regulation of inflammation and oxidative stress, thus offering protection to tissues and organs.13 Long used in traditional Chinese medicine for treating inflammatory diseases, OMT has demonstrated anti-inflammatory effects in rheumatoid arthritis rats through immune cell modulation14 and alleviates lipopolysaccharide-induced mastitis by inhibiting the NF-κB/MAPK signaling pathway.15 Additionally, OMT protects against stroke-related damage, preserving neuronal integrity and reducing brain hypoxic-ischemic injury in animal models of cerebral ischemia-reperfusion by modulating apoptosis and inflammatory responses.16,17 Moreover, OMT exhibits significant analgesic properties, effective not only in inflammatory pain18,19 but also in neuropathic pain. Intraperitoneal OMT injection notably reduced both mechanical and thermal allodynia in sciatica mice.20,21 However, its potential efficacy in treating CPSP remains unexplored. It is hypothesized that OMT alleviates CPSP in rats, with its mechanism of action linked to the reduction of inflammation around the thalamic injury site. This study utilized network pharmacology and animal models to demonstrate that OMT mitigates CPSP by attenuating the inflammatory response.
Materials and Methods
Target Acquisition of Oxymatrine
All OMT targets were retrieved from SwissTargetPrediction (http://www.swisstargetprediction.ch/), SuperPred (https://prediction.charite.de/), and BATMAN-TCM (http://bionet.ncpsb.org.cn/batman-tcm/) databases. The target data were then standardized by cross-referencing with UniProt (https://www.uniprot.org/), removing human gene entries and eliminating duplicate or invalid targets to obtain the standardized gene names.
Target Acquisition is Associated with Central Post-Stroke Pain
Targets related to “stroke” and “central pain” were identified by querying GeneCards (https://www.genecards.org/) and OMIM (https://omim.org/) databases. The target data from these two databases were integrated in Microsoft Excel, with duplicates removed and gene information validated using the UniProt database.
Construction of the Interaction Network of Target Proteins
To further study the protein-protein interactions of Oxymatrine in treating central pain after stroke, the drug-interaction genes were uploaded to the interaction database STRING (https://string-db.org/) to construct a protein interaction network database (PPI). The species was set to “Homo sapiens”, and other parameters were kept at default settings. The results were stored in TSV format and imported into Cytoscape 3.8.2. The network was analyzed, and the results of the analysis were saved. The size and color of the nodes were used to reflect the Degree value. The larger the node, the larger the Degree value. A protein interaction network diagram was made.
Gene Ontology (GO) Enrichment Analysis
The obtained drug components and disease targets were mapped to each other, and a Venn map was created to get the intersection genes. The drug-disease intersection gene was uploaded to the DAVID (https://david.ncifcrf.gov/summary.jsp) database, with the gene identifier selected as OFFICIAL_GENE_SYMBOL, and the species set as Homo Sapiens. Using the function of the DAVID6.8GO gene, ten samples were selected from three aspects: Biological Process (BP), Cellular Component (CC), and Molecular Function (MF), as the main gene function enrichment process of oxymatrine in the treatment of central post-stroke pain.
Databases used in this research were based on publicly available data obtained through legal means, and the research was conducted using anonymized data. This complies with items 1 and 2 of Article 32 of the Measures for Ethical Review of Life Science and Medical Research Involving Human Subjects, dated February 18, 2023, China. Therefore, ethical review is exempt.
Animals
Healthy male Sprague-Dawley (SD) rats, aged 6–8 weeks and weighing 200–220 g, were obtained from the Chongqing Tengxin Huafu Laboratory Animal Sales Company. The rats were housed in the laboratory animal facility of Southwest Medical University under a controlled light-dark cycle. All experiments adhered to the National Institutes of Health Guidelines for the Care and Use of Laboratory Animals (NIH Publication No. 80–23, Revised 1996), as well as the International Association for the Study of Pain’s Ethical Guidelines for Studies Involving Conscious Animals. The study was approved by the Experimental Animal Ethics Committee of Southwestern Medical University (Ethic No.:20220811–017).
Experimental Design and Drug Administration
The experiment was conducted in two phases. First, a rat model of CPSP was established. Rats were randomly assigned to two groups, with 13 animals in each group: sham and CPSP. In the sham group, normal saline was injected into the right ventral posterolateral nucleus (VPL), while type IV collagenase was injected into the right VPL of rats in the CPSP group. On day 7 post-surgery, three rats per group were euthanized for brain tissue hematoxylin and eosin (HE) staining, while the remaining rats (n = 10/group) underwent mechanical pain sensitivity testing. In the second phase, the effects of OMT on CPSP rats were assessed. Rats were randomly assigned to five groups (n = 10): sham + NaCl, CPSP + NaCl, and CPSP + OMT (100 mg/kg, 200 mg/kg, 400 mg/kg). From day 7 to day 14 post-surgery, the baseline mechanical pain threshold was measured daily at 8:00 AM, followed by intraperitoneal administration of NaCl or varying doses of OMT. Paw withdrawal mechanical thresholds (PWMT) were recorded every 30 minutes following OMT administration. The optimal OMT dose for CPSP rats was selected based on PWMT measurements. On day 14, thalamic tissue samples from rats treated with NaCl or the selected OMT dose were analyzed using ELISA (n = 3) to assess levels of IL-1β, IL-6, and TNF-α, and immunofluorescence (n = 3) to evaluate changes in microglia and astrocytes (Figure 1).
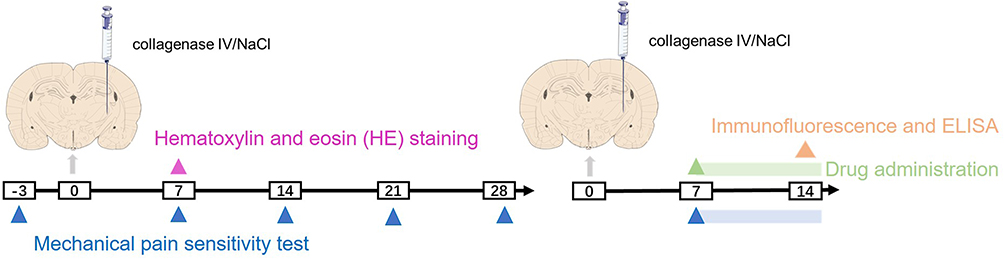 |
Figure 1 Experimental time diagram and design diagram. |
Drugs and Chemicals
OMT, a white crystalline powder with a purity of ≥ 98.0%, was obtained from Gelipu Biotechnology Company, Chengdu. OMT was freshly prepared just before administration and diluted with 0.9% normal saline. Type IV collagenase (Sigma-Aldrich) was dissolved in NaCl to a concentration of 0.025 U /0.25 μL per rat. Sodium pentobarbital, sourced from Sigma-Aldrich, was used for anesthesia, and both OMT and sodium pentobarbital were administered intraperitoneally.
Preparation of the CPSP Animal Model
The CPSP model was established following the protocol outlined by Wasserman et al.22 SD rats weighing 200–220 g were selected for the procedure. After anesthesia with 2% pentobarbital (50 mg/kg), the rats were fixed in place using a brain stereotaxic apparatus. The positioning was adjusted such that the zero point was aligned with the rat’s fontanel, 3.4 mm posterior to the fontanel and 3.5 mm lateral to the midline. A microneedle was inserted through the hole, advancing slowly to a depth of 6.5 mm, targeting the right VPL. After a 5-minute wait, 0.25 μL of type IV collagenase (0.025 U) or normal saline was injected slowly within 15 minutes. The needle was left in place for an additional 5 minutes post-injection to prevent reflux before being withdrawn.
Mechanical Pain Sensitivity Testing
Mechanical pain sensitivity was assessed using PWMT. All rats were acclimated to the test environment for at least 7 days prior to baseline measurements and for 30 minutes before testing. All tests were performed between 8:00 AM and 12:00 PM. In all behavioral studies, animals were randomly assigned to groups by an investigator blinded to the experimental conditions. Von Frey filaments with forces ranging from 1–26 g were used to assess mechanical sensitivity. Testing began with the 1 g filament, which was applied perpendicularly to the plantar surface of one hind paw, gently bending the filament until a response was elicited. A positive response was defined as rapid paw withdrawal, licking, or shaking. The contralateral paw was tested after a 3–5 second interval. For each paw, the threshold was defined as the minimum force that elicited ≥ 5 positive responses out of 10 consecutive stimuli. If no response was observed at a given force, testing proceeded to higher forces (up to 26 g).
Hematoxylin and Eosin (HE) Staining
On day 7 post-surgery, the rat brains were fixed in 10% neutral formalin for 48 hours, embedded in paraffin, and coronal sections (12 μm thick) were prepared. The sections were dewaxed in xylene, rehydrated through a graded ethanol series, stained with HE, and examined under a Nikon Eclipse Ci-L optical microscope.
ELISA
The thalamic tissue surrounding the hemorrhagic lesion site was collected and homogenized in PBS. The homogenate was then centrifuged, and the supernatant was collected for analysis. Total protein concentration was measured using a BCA protein assay kit (Beyotime, China), and levels of IL-1β (Cat. No. E-EL-R0015c), IL-6 (Cat. No. E-EL-R2856c), and TNF-α (Cat. No. E-EL-R0012c) in the homogenized thalamic tissue were determined via enzyme-linked immunosorbent assay (ELISA) kits (Elabscience, China). The concentrations of each protein of interest were expressed in picograms per milligram of total protein.
Immunofluorescence
Rats were euthanized under 2% pentobarbital anesthesia, followed by cardiac perfusion with normal saline and 4% paraformaldehyde. The brains were extracted and stored overnight at 4°C. Brain tissue was then sectioned into 30 μm-thick slices. The sections were blocked with 1% bovine serum albumin and permeabilized with 0.1% Triton X-100 at room temperature for 2 minutes. The slices were incubated overnight at 4°C with primary antibodies: mouse anti-glial fibrillary acidic protein (GFAP) (1:500; Cell Signaling Technology, CST; #34001S; Lot 6), rabbit anti-IBA-1 (1:100; microglia marker, CST; Cat. No. #17198), and rabbit anti-NeuN (1:500; ab177487; Cat No. GR249899-54). On the following day, Cy3-conjugated AffiniPure Goat Anti-Rabbit IgG (H + L) secondary antibodies (1:500; Proteintech, Cat. No. SA00009-2) were applied and incubated at room temperature for 2–3 hours. Images were captured using a laser scanning confocal fluorescence microscope (Olympus FV1000, Japan). Immunofluorescence intensity was measured at peri-thalamic lesion sites using ImageJ software with a consistent threshold configuration. The extent of immunoreactivity was quantified as the percentage of immunostained area (immunofluorescent-positive surface area/total measured image area × 100%).
Statistical Analysis
Statistical analyses were performed using GraphPad Prism 9.0 for Windows (GraphPad Software, San Diego, CA, USA). Data are presented as mean ± SEM. The paw withdrawal threshold was analyzed using two-way ANOVA, followed by Bonferroni’s multiple comparison test. ELISA and fluorescence data were compared using one-way analysis of variance (ANOVA), followed by the Bonferroni post-hoc test. A P-value of < 0.05 was considered statistically significant.
Results
Cross-Target Genes of Oxymatrine and Central Post-Stroke Pain
Using a Probability > 0 threshold for target inclusion based on SwissTargetPrediction, 31 targets were identified. From the BATMAN-TCM database, 151 targets were included, with a score cutoff > 0.60 (LR = 13.26) and P < 0.05. Eleven targets were obtained from SuperPred, and after removing duplicates, 172 targets were retained. For the terms “stroke” and “central pain” GeneCards and OMIM databases yielded 8899 and 12744 targets, respectively. These gene sets were validated and corrected using the UniProt database. After intersecting the drug targets with those related to CPSP, 123 common target genes were identified, representing the interactive targets of drug treatment for CPSP (Figure 2A). These 123 target genes were imported into the String database for protein-protein interaction analysis. The species was set to Homo sapiens, with a confidence threshold of 0.4, resulting in the construction of a protein-protein interaction network (Figure 2B). Core targets identified included STAT3, NFKB1, MTOR, TLR4, MMP9, HSP90AB1, GSK3B, MMP2, MAPK1, HDAC1, PIK3CA, and AR.
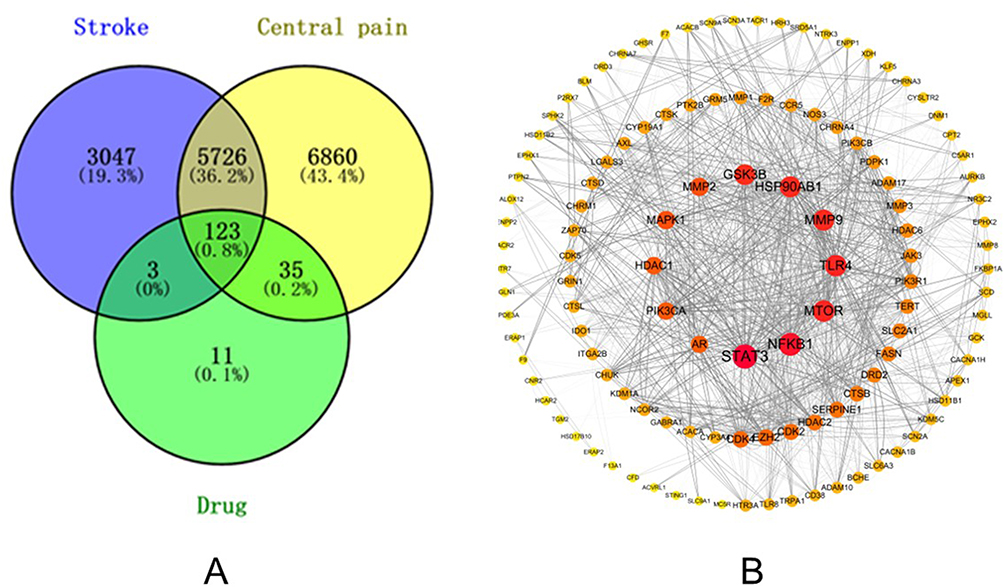 |
Figure 2 (A) Cross-gene between disease and drug. The blue circle represents the target of Stroke, the yellow circle represents the target of Central pain, and the green circle represents the target of OMT. The cross in the middle indicates a potential target for OMT treatment of CPSP. (B) Core Protein-related Interaction Network. It consists of 119 nodes and 1294 edges. The size of the targets is reflected by the degree value and the color, while the thickness of the edges is indicated by the combined score value. |
GO Enrichment Analysis
Drug-disease intersection genes were analyzed through functional enrichment using the DAVID database, focusing on the gene functions of OMT in the treatment of CPSP. The analysis included 209 Gene Ontology (GO) terms, of which 124 were related to biological processes (BP) with a P-value < 0.01. Additionally, 40 cell component (CC) terms and 45 molecular function (MF) terms were identified. BP terms notably involved inflammatory processes (Figure 3).
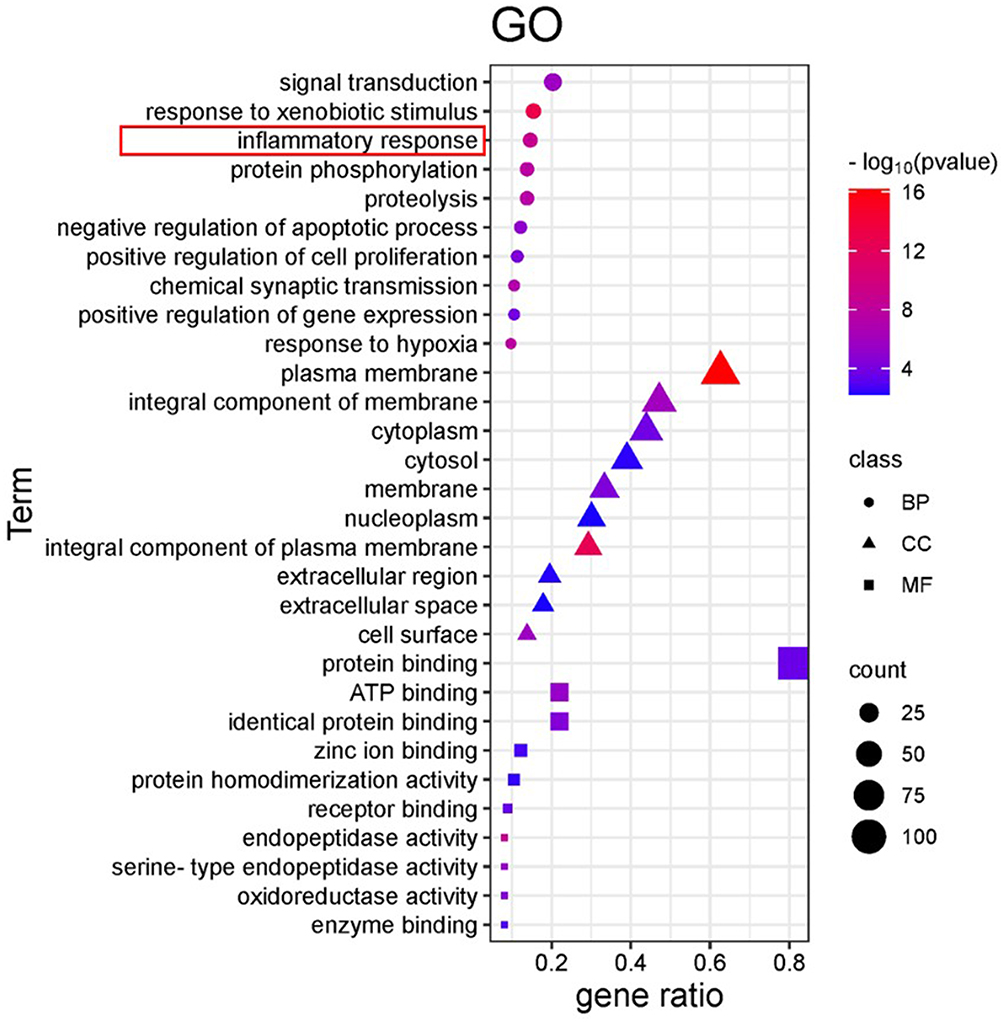 |
Figure 3 Gene Ontology (GO) enrichment analysis. The circle represents Biological Process (BP), the triangle represents Cellular Component (CC), and the square represents Molecular Function (MF). The size of the graph represents the data enriched in the corresponding functional genes, and the P-value decreases gradually from blue to red. |
CPSP Rats Exhibited Thalamic Injury and Mechanical Allodynia
In histological analysis, the cells in the right thalamic region of sham rats appeared well-defined, with intact structures, large round nuclei, and no significant pathological changes. In contrast, seven days post-modeling, rats in the CPSP group exhibited focal necrosis, disordered cell arrangement, cytoplasmic condensation, and nuclear shrinkage in the right thalamic region (Figure 4A). Compared to the sham group, the bilateral plantar PWMT in the CPSP group was significantly reduced from day 7 post-modeling and persisted for at least 28 days (Figure 4B).
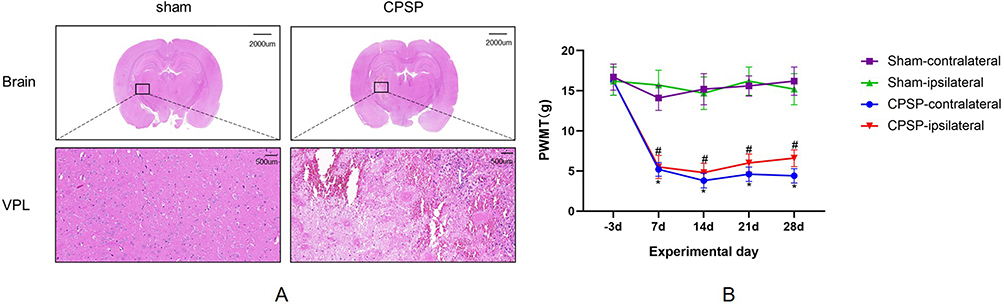 |
Figure 4 (A) Pathological changes of the rat thalamus induced by type IV collagenase. Brain scale =2000um (0.5×), VPL scale =500um (20.0×). VPL is the ventral posterior lateral nucleus. (B) Progression of paw withdrawal mechanical thresholds (PWMT) induced by type IV collagenase. (n=10/group. *P<0.05 CPSP-contralateral vs Sham-contralateral. #P<0.05 CPSP-ipsilateral vs Sham-ipsilateral). |
OMT Treatment Can Relieve Mechanical Allodynia Induced by CPSP
Compared to NaCl intraperitoneal injection, no significant changes in the PWMT of both hind paws were observed in CPSP rats treated with 100 mg/kg OMT (P > 0.05). In contrast, treatment with 200 mg/kg and 400 mg/kg OMT significantly increased the PWMT in both hind paws at 90 and 120 minutes (P < 0.05). No significant difference in PWMT was observed between the 200 mg/kg and 400 mg/kg OMT-treated groups (P > 0.05) (Figure 5).
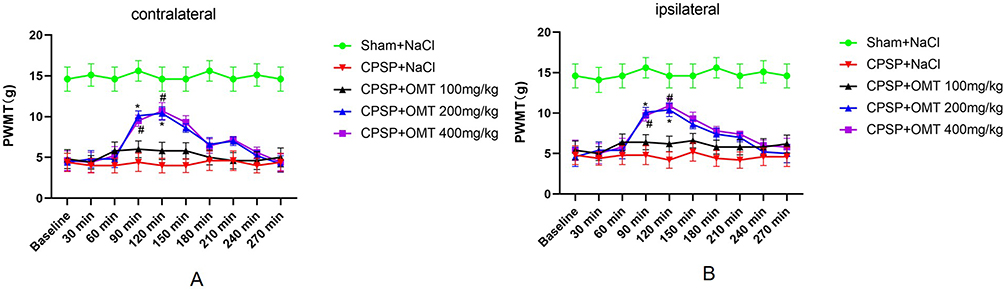 |
Figure 5 Effect of Drug Administration on mechanical pain threshold in rats (The 14th Day). (A) Changes of paw withdrawal mechanical thresholds (PWMT) on the contralateral side of thalamic injection; (B) Changes of paw withdrawal mechanical thresholds (PWMT) on the ipsilateral side of thalamic injection. (n=10/group. *P < 0.05 CPSP+OMT 200 mg/kg vs CPSP +NaCl. #P< 0.05 CPSP+OMT 400mg/kg vs CPSP +NaCl). The data on days 8 to 14 are similar, and the trends are repetitive. |
OMT Treatment Decreased the Expression of Inflammatory Cytokines IL-1β, IL-6, and TNF-α in Peri-Thalamic Lesion Sites of CPSP Rats
In comparison to Sham rats, the expression levels of inflammatory factors IL-1β, IL-6, and TNF-α in the peri-thalamic lesion areas of CPSP rats were significantly elevated (P < 0.05). Treatment with 200 mg/kg OMT significantly reduced the expression of these inflammatory factors in the peri-thalamic lesion sites compared to NaCl treatment (P < 0.05) (Figure 6).
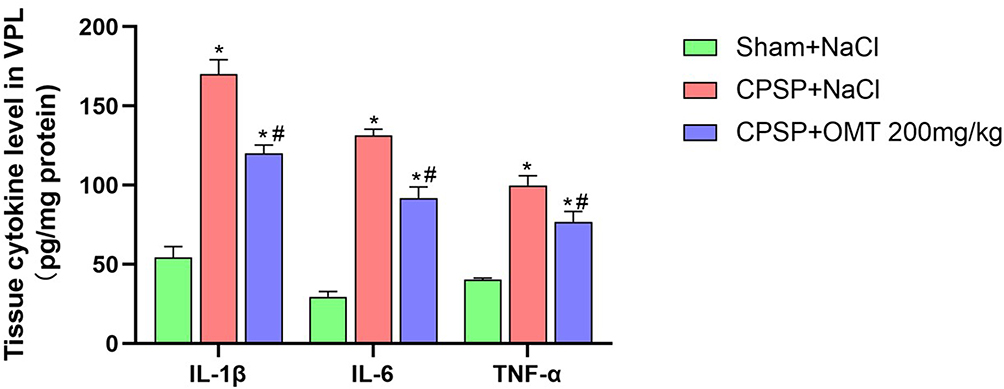 |
Figure 6 Expression of inflammatory factors around the thalamic-injection site of rats. (n=3/group. *P < 0.05 vs Sham +NaCl. #P< 0.05 vs CPSP +NaCl). |
OMT Treatment Inhibited the Activation of Microglia and Astrocytes in Peri-Thalamic Lesion Sites of CPSP Rats
CPSP induced increased expression of Iba-1 and GFAP in the peri-thalamic lesion sites compared to Sham rats (P < 0.05). However, treatment with 200 mg/kg OMT significantly reduced the expression of both Iba-1 and GFAP (P < 0.05) (Figure 7).
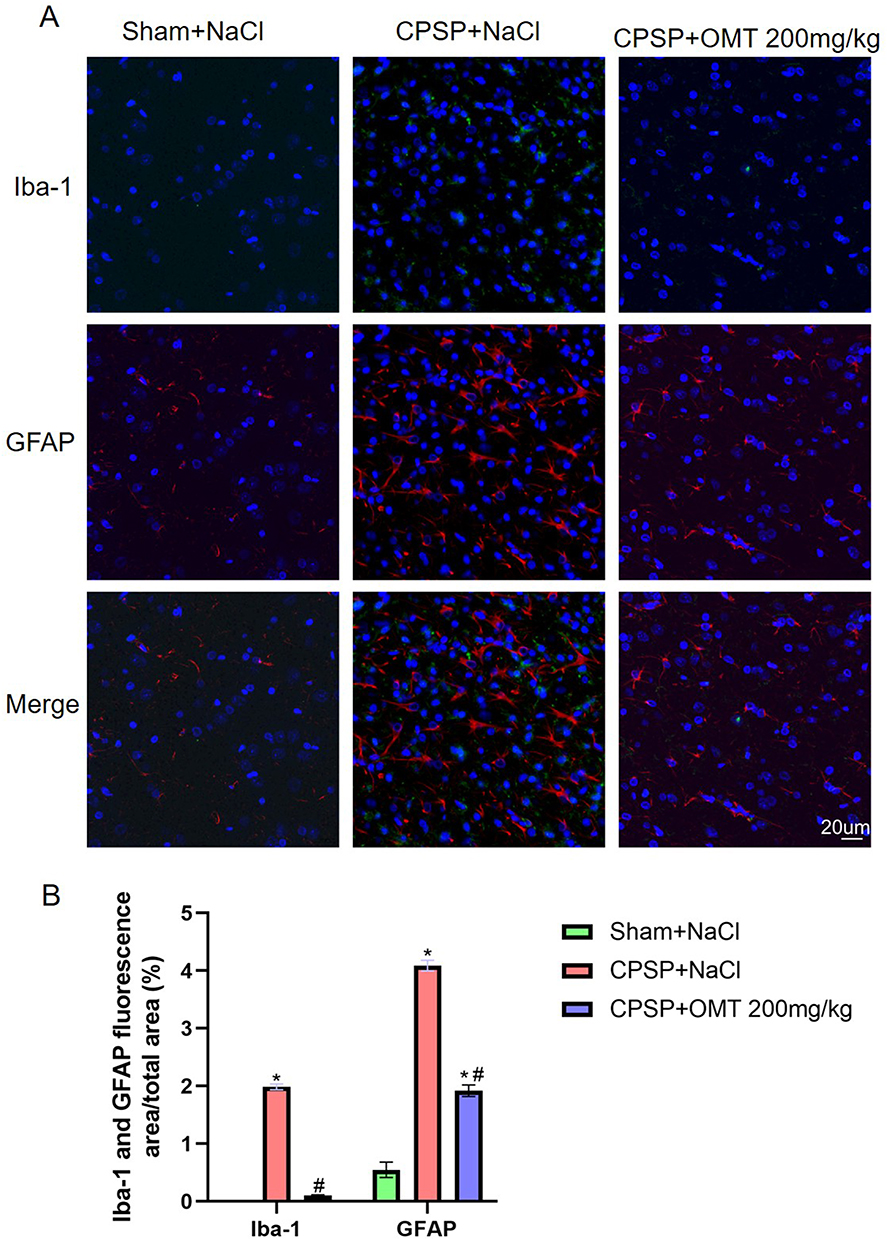 |
Figure 7 Changes of microglia and astrocytes around the thalamic-injection site of rats. (A) Representative immunofluorescence image of microglia and astrocytes around the thalamic-injection site of rats. GFAP (red) labels astrocytes, IBA1 (green) marks microglia, and NeuN (blue) identifies neurons. Scale =20μm (40.0×). (B) The proportion of Iba-1-positive area and GFAP-positive area around the thalamic-injection site of rats. (n=3/group. *P < 0.05 vs Sham +NaCl. #P< 0.05 vs CPSP +NaCl). |
Discussion
This study demonstrates that (1) network pharmacology predicts that OMT’s mechanism in treating CPSP is primarily linked to modulating the inflammatory response. (2) Hemorrhagic stroke can induce thalamic injury and mechanical allodynia. (3) Increased expression of inflammatory factors IL-1β, IL-6, and TNF-α, alongside the activation of microglia and astrocytes, was observed at the peri-thalamic lesion sites in CPSP rats. (4) OMT treatment alleviates mechanical pain in CPSP, reduces the expression of inflammatory markers IL-1β, IL-6, and TNF-α, and inhibits microglial and astrocytic activation at the peri-thalamic lesion sites.
Pain is one of the most debilitating consequences following a stroke. PSP resulting directly from brain injury is classified as CPSP. The thalamus serves as a key relay station for pain transmission, receiving both upstream and downstream inputs, and plays a critical role in modulating nociceptive information.23 Clinical studies have identified specific thalamic regions, particularly the VPL, as being closely linked to the onset of CPSP.24–26 Wasserman et al first established a rat model of stroke pain following thalamic hemorrhage by injecting type IV collagenase into the VPL.22 This model is advantageous due to its high reproducibility and its ability to mimic the development of central pain symptoms in the thalamus. In our study, the injection of type IV collagenase into the VPL resulted in pathological damage at the injection site, confirmed by HE staining. A significant reduction in the PWMT of both hind paws was observed on day 7 post-surgery (Figure 4), aligning with previous studies and the typical characteristics of CPSP animal models.27–29
Sophora flavescens Ait., a traditional Chinese medicinal herb, is known for its anti-arrhythmic, anti-tumor, hepatoprotective, anti-fibrotic, anti-inflammatory, and anti-sensitivity properties.30 OMT, a key component of Sophora flavescens, possesses a wide array of beneficial pharmacological effects. Network pharmacology employs multidisciplinary approaches to uncover the principles and mechanisms underlying interactions between drugs and biological systems by analyzing, visualizing, and interpreting biological network data.31 GO enrichment analysis revealed that the biological processes associated with OMT’s treatment of CPSP are closely linked to inflammation (Figure 3), a finding that was corroborated through animal experiments.
OMT has an LD50 of 898.22 mg/kg (95% confidence interval 832.46‐963.98).14 Previous research has reported that 120 mg/kg OMT alleviates neuronal damage in the cerebral cortex and hippocampal CA3 regions caused by ischemia and hypoxia in neonatal rats.16 In this study, the analgesic effects of intraperitoneal OMT were investigated using concentration gradients of 100 mg/kg, 200 mg/kg, and 400 mg/kg in CPSP rats. PWMT was assessed daily from day 7 to day 14, showing minimal day-to-day variation and repeating trends. For clarity, only data from the 14th day post-operation are presented (Figure 5). The results indicated no significant change in PWMT in both hind paws of CPSP rats treated with 100 mg/kg OMT compared to NaCl injection (P > 0.05). In contrast, 200 mg/kg and 400 mg/kg OMT intraperitoneal injections increased PWMT in both hind paws of CPSP rats at 90–120 minutes post-injection (Figure 5). No significant toxicity or adverse effects were observed during the experiment, except for a transient quiet state in a few rats following the 400 mg/kg OMT injection. Given that the 400 mg/kg dose did not significantly enhance PWMT compared to 200 mg/kg (P > 0.05), 200 mg/kg was determined to be the optimal dose for CPSP rats. Both 200 mg/kg and 400 mg/kg OMT led to a return to baseline PWMT levels 270 minutes after injection (Figure 5). This likely reflects the relatively short duration of OMT’s effect, with a half-life of approximately 3.44 hours,17 and may also be linked to the persistent nature of CPSP pain. Currently, amitriptyline is the first-line treatment for CPSP, but its effects are often incomplete, and high doses are poorly tolerated by patients with stroke.32,33 Calcium channel modulators, such as pregabalin and gabapentin, have emerged as potential treatments for CPSP. In a placebo-controlled, double-blind study of 219 patients with CPSP, pregabalin did not significantly reduce the mean pain score.34 In clinical practice, treating CPSP often involves trial and error until effective pain relief is achieved, often requiring a combination of several drugs.35 OMT may hold promise as an adjunctive treatment for CPSP.
Pain not only induces changes within neurons themselves but also activates various non-neuronal cell types in the pain circuit, including glial cells and immune cells, which contribute to neuroinflammation in both the central and peripheral nervous systems.36 A key feature of neuroinflammation is the activation of glial cells, such as microglia and astrocytes, which release pro-inflammatory cytokines and chemokines. Glial activation is a central characteristic of neuroinflammation and plays a pivotal role in the development of chronic pain.37,38 Pro-inflammatory cytokines, including IL-1β, TNF-α, and IL-6, derived from microglia, are strongly associated with pain hypersensitivity.39 Echeverry et al demonstrated that neutralizing these pro-inflammatory cytokines with a mixture of anti-IL-1β, TNF-α, and IL-6 antibodies significantly reduced tactile and cold allodynia in rats with partial sciatic nerve injury, observed two weeks post-injury.40 These findings align with our hypothesis that hemorrhagic stroke in the thalamus activates microglia and astrocytes around the lesion site, increasing the levels of IL-1β, TNF-α, and IL-6, which subsequently lead to mechanical allodynia. Localized inflammation interacts bidirectionally with damaged nerve cells, driving chronic pain. Our results showed that OMT inhibits the activation of microglia and astrocytes induced by stroke injury and reduces the release of pro-inflammatory cytokines IL-1β, TNF-α, and IL-6 (Figures 6 and 7). The core targets predicted by network pharmacology offer insights into the potential neuroprotective mechanisms underlying OMT’s therapeutic effects (Figure 2B). A key signaling pathway in the inflammatory response is the overactivation of the Toll-like receptor 4 (TLR4)/high mobility group box 1 (HMGB1) axis. When astrocytes and microglia are activated, the released HMGB1 further stimulates microglia, creating a negative feedback loop through the HMGB1/TLR4/NF-κB pathway. OMT targets the TLR4 receptor, alleviating the inflammatory response induced by stroke damage. Additionally, OMT reduces oxidative stress and cell apoptosis by inhibiting the EGFR/PI3K/Akt/mTOR signaling pathway and STAT3.17 However, the exact mechanisms through which OMT modulates CPSP require further investigation.
This study has several limitations. While network pharmacology and in vivo experiments demonstrated that OMT attenuates CPSP in rats by suppressing neuroinflammation, the specific inflammatory pathways involved remain unidentified. Additionally, our investigation was limited to assessing the effects of a single OMT dose (200 mg/kg), which was behaviorally optimized, on inflammatory factors, microglia, and astrocytes around the thalamic injection site. Moreover, Moreover, the relationship between behavioral outcomes observed with different drug doses and molecular responses remains unclear and warrants further research.
Conclusion
In conclusion, network pharmacology revealed that the therapeutic mechanism of OMT in treating CPSP is closely linked to its modulation of inflammatory responses. In vivo experiments further clarified that the alleviation of mechanical allodynia induced by CPSP following OMT treatment is associated with a reduction in the expression of inflammatory factors IL-1β, IL-6, and TNF-α, as well as the inhibition of microglial and astrocytic activation at the peri-thalamic lesion sites in CPSP rats. The anti-inflammatory properties of OMT suggest its potential as a valuable treatment for CPSP.
Acknowledgments
This work was supported by the Sichuan Science and Technology Program (No. 2023YFS0253) and the Open Project of Sichuan Provincial Key Laboratory of Nuclear Medicine and Molecular Imaging in 2020 (No. HYX20008).
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare that they have no competing interests in this work.
References
1. Westerlind E, Singh R, Persson HC, Sunnerhagen KS. Experienced pain after stroke: a cross-sectional 5-year follow-up study. BMC Neurol. 2020;20(1):4. doi:10.1186/s12883-019-1584-z
2. Treister AK, Hatch MN, Cramer SC, Chang EY. Demystifying poststroke pain: from etiology to treatment. PM R. 2017;9(1):63–75. doi:10.1016/j.pmrj.2016.05.015
3. Klit H, Finnerup NB, Andersen G, Jensen TS. Central poststroke pain: a population-based study. Pain. 2011;152(4):818–824. doi:10.1016/j.pain.2010.12.030
4. Oh H, Seo W. A comprehensive review of central post-stroke pain. Pain Manag Nurs. 2015;16(5):804–818. doi:10.1016/j.pmn.2015.03.002
5. Şahin-Onat Ş, Ünsal-Delialioğlu S, Kulaklı F, Özel S. The effects of central post-stroke pain on quality of life and depression in patients with stroke. J Phys Ther Sci. 2016;28(1):96–101. doi:10.1589/jpts.28.96
6. Infantino R, Schiano C, Luongo L, et al. MED1/BDNF/TrkB pathway is involved in thalamic hemorrhage-induced pain and depression by regulating microglia. Neurobiol Dis. 2022;164:105611. doi:10.1016/j.nbd.2022.105611
7. Klit H, Finnerup NB, Jensen TS. Central post-stroke pain: clinical characteristics, pathophysiology, and management. Lancet Neurol. 2009;8(9):857–868. doi:10.1016/S1474-4422(09)70176-0
8. Liampas A, Velidakis N, Georgiou T, et al. Prevalence and management challenges in central post-stroke neuropathic pain: a systematic review and meta-analysis. Adv Ther. 2020;37(7):3278–3291. doi:10.1007/s12325-020-01388-w
9. Nicholson BD. Evaluation and treatment of central pain syndromes. Neurology. 2004;62(5 Suppl 2):S30–S36. doi:10.1212/wnl.62.5_suppl_2.s30
10. Shih HC, Kuan YH, Shyu BC. Targeting brain-derived neurotrophic factor in the medial thalamus for the treatment of central poststroke pain in a rodent model. Pain. 2017;158(7):1302–1313. doi:10.1097/j.pain.0000000000000915
11. Yang F, Luo WJ, Sun W, et al. SDF1-CXCR4 signaling maintains central post-stroke pain through mediation of glial-neuronal interactions. Front Mol Neurosci. 2017;10:226. doi:10.3389/fnmol.2017.00226
12. Hiraga SI, Itokazu T, Hoshiko M, Takaya H, Nishibe M, Yamashita T. Microglial depletion under thalamic hemorrhage ameliorates mechanical allodynia and suppresses aberrant axonal sprouting. JCI Insight. 2020;5(3):e131801. doi:10.1172/jci.insight.131801
13. Lan X, Zhao J, Zhang Y, Chen Y, Liu Y, Xu F. Oxymatrine exerts organ- and tissue-protective effects by regulating inflammation, oxidative stress, apoptosis, and fibrosis: from bench to bedside. Pharmacol Res. 2020;151:104541. doi:10.1016/j.phrs.2019.104541
14. Ma A, Yang Y, Wang Q, Wang Y, Wen J, Zhang Y. Anti-inflammatory effects of oxymatrine on rheumatoid arthritis in rats via regulating the imbalance between Treg and Th17 cells. Mol Med Rep. 2017;15(6):3615–3622. doi:10.3892/mmr.2017.6484
15. Yang Z, Yin R, Cong Y, et al. Oxymatrine lightened the inflammatory response of LPS-induced mastitis in mice through affecting NF-κB and MAPKs signaling pathways. Inflammation. 2014;37(6):2047–2055. doi:10.1007/s10753-014-9937-7
16. Liu Y, Wang H, Liu N, et al. Oxymatrine protects neonatal rat against hypoxic-ischemic brain damage via PI3K/Akt/GSK3β pathway. Life Sci. 2020;254:116444. doi:10.1016/j.lfs.2019.04.070
17. Dhurandhar Y, Tomar S, Das A, et al. Unlocking the potential of oxymatrine: a comprehensive review of its neuroprotective mechanisms and therapeutic prospects in neurological disorders. ACS Chem Neurosci. 2024;15(23):4245–4257. doi:10.1021/acschemneuro.4c00338
18. Zhu T, Zhou D, Zhang Z, et al. Analgesic and antipruritic effects of oxymatrine sustained-release microgel cream in a mouse model of inflammatory itch and pain. Eur J Pharm Sci. 2020;141:105110. doi:10.1016/j.ejps.2019.105110
19. Zhou J, Yang G, Jin S, Tao L, Yu J, Jiang Y. Oxymatrine-carbenoxolone sodium inclusion compound induces antinociception and increases the expression of GABA(A)alpha1 receptors in mice. Eur J Pharmacol. 2010;626(2–3):244–249. doi:10.1016/j.ejphar.2009.09.065
20. Wang H, Li Y, Dun L, et al. Antinociceptive effects of oxymatrine from Sophora flavescens, through regulation of NR2B-containing NMDA receptor-ERK/CREB signaling in a mice model of neuropathic pain. Phytomedicine. 2013;20(11):1039–1045. doi:10.1016/j.phymed.2013.04.012
21. Liu HY, Li YX, Hao YJ, et al. Effects of oxymatrine on the neuropathic pain induced by chronic constriction injury in mice. CNS Neurosci Ther. 2012;18(12):1030–1032. doi:10.1111/cns.12026
22. Wasserman JK, Koeberle PD. Development and characterization of a hemorrhagic rat model of central post-stroke pain. Neuroscience. 2009;161(1):173–183. doi:10.1016/j.neuroscience.2009.03.042
23. Lenz FA, Dougherty PM, Meeker TJ, Saffer MI, Oishi K. Neuroscience of the human thalamus related to acute pain and chronic “thalamic” pain. J Neurophysiol. 2024;132(6):1756–1778. doi:10.1152/jn.00065.2024
24. Krause T, Brunecker P, Pittl S, et al. Thalamic sensory strokes with and without pain: differences in lesion patterns in the ventral posterior thalamus. J Neurol Neurosurg Psychiatry. 2012;83(8):776–784. doi:10.1136/jnnp-2011-301936
25. Sprenger T, Seifert CL, Valet M, et al. Assessing the risk of central post-stroke pain of thalamic origin by lesion mapping. Brain. 2012;135(Pt 8):2536–2545. doi:10.1093/brain/aws153
26. Vartiainen N, Perchet C, Magnin M, et al. Thalamic pain: anatomical and physiological indices of prediction. Brain. 2016;139(Pt 3):708–722. doi:10.1093/brain/awv389
27. Lu J, Guo X, Yan M, et al. P2X4R contributes to central disinhibition via TNF-α/TNFR1/GABAaR pathway in post-stroke pain rats. J Pain. 2021;22(8):968–980. doi:10.1016/j.jpain.2021.02.013
28. Chen X, Li Z, Zhang B, et al. Alleviation of mechanical allodynia by 14,15-epoxyeicosatrienoic acid in a central poststroke pain model: possible role of allopregnanolone and δ-subunit-containing gamma-aminobutyric acid a receptors. J Pain. 2019;20(5):577–591. doi:10.1016/j.jpain.2018.11.006
29. Fu G, Du S, Huang T, et al. FTO (Fat-mass and obesity-associated protein) participates in hemorrhage-induced thalamic pain by stabilizing toll-like receptor 4 expression in thalamic neurons. Stroke. 2021;52(7):2393–2403. doi:10.1161/STROKEAHA.121.034173
30. Huan DQ, Hop NQ, Son NT. Oxymatrine: a current overview of its health benefits. Fitoterapia. 2023;168:105565. doi:10.1016/j.fitote.2023.105565
31. Li ZY, Dai YX, Wu ZM, et al. Network pharmacology analysis and animal experiment validation of neuroinflammation inhibition by total ginsenoside in treating CSM. Phytomedicine. 2024;126:155073. doi:10.1016/j.phymed.2023.155073
32. Kim JS. Pharmacological management of central post-stroke pain: a practical guide. CNS Drugs. 2014;28(9):787–797. doi:10.1007/s40263-014-0194-y
33. Kumar G, Soni CR. Central post-stroke pain: current evidence. J Neurol Sci. 2009;284(1–2):10–17. doi:10.1016/j.jns.2009.04.030
34. Kim JS, Bashford G, Murphy KT, Martin A, Dror V, Cheung R. Safety and efficacy of pregabalin in patients with central post-stroke pain. Pain. 2011;152(5):1018–1023. doi:10.1016/j.pain.2010.12.023
35. Tamasauskas A, Silva-Passadouro B, Fallon N, et al. Management of central poststroke pain: systematic review and meta-analysis. J Pain. 2025;26:104666. doi:10.1016/j.jpain.2024.104666
36. Li L, Li T, Qu X, Sun G, Fu Q, Han G. Stress/cell death pathways, neuroinflammation, and neuropathic pain. Immunol Rev. 2024;321(1):33–51. doi:10.1111/imr.13275
37. Ji RR, Nackley A, Huh Y, Terrando N, Maixner W. Neuroinflammation and central sensitization in chronic and widespread pain. Anesthesiology. 2018;129(2):343–366. doi:10.1097/ALN.0000000000002130
38. Guillot X, Semerano L, Decker P, Falgarone G, Boissier MC. Pain and immunity. Joint Bone Spine. 2012;79(3):228–236. doi:10.1016/j.jbspin.2011.10.008
39. Atta AA, Ibrahim WW, Mohamed AF, Abdelkader NF. Microglia polarization in nociplastic pain: mechanisms and perspectives. Inflammopharmacology. 2023;31(3):1053–1067. doi:10.1007/s10787-023-01216-x
40. Echeverry S, Shi XQ, Yang M, et al. Spinal microglia are required for long-term maintenance of neuropathic pain. Pain. 2017;158(9):1792–1801. doi:10.1097/j.pain.0000000000000982
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.