Back to Journals » OncoTargets and Therapy » Volume 12
OX40 (CD134) and OX40 ligand, important immune checkpoints in cancer
Authors Deng J, Zhao S, Zhang X, Jia K, Wang H, Zhou C, He Y ![]()
Received 2 May 2019
Accepted for publication 30 July 2019
Published 6 September 2019 Volume 2019:12 Pages 7347—7353
DOI https://doi.org/10.2147/OTT.S214211
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Gaetano Romano
Juan Deng,1,2 Sha Zhao,1,2 Xiaoshen Zhang,1,2 Keyi Jia,1,2 Hao Wang,1,2 Caicun Zhou,1 Yayi He1
1Department of Medical Oncology, Shanghai Pulmonary Hospital, Tongji University Medical School Cancer Institute, Tongji University School of Medicine, Shanghai 200433, People’s Republic of China; 2Medical School, Tongji University, Shanghai 200092, People’s Republic of China
Correspondence: Caicun Zhou; Yayi He
Department of Medical Oncology, Shanghai Pulmonary Hospital, Tongji University Medical School Cancer Institute, Tongji University School of Medicine, No. 507 Zhengmin Road, Shanghai 200433, People’s Republic of China
Tel +86 216 511 5006
Email [email protected]; [email protected]
Abstract: Immunotherapy has shown promising results in cancer treatment. Research shows that most patients might be resistant to these therapies. So, new immune therapies are needed. OX40 (CD134) and OX40 ligand (OX40L), costimulatory molecules, express on different types of immune cells. The interaction between OX40 and OX40L (OX40/OX40L) induces the expansion and proliferation of T cells and decreases the immunosuppression of regulatory T (Treg) cells to enhance the immune response to the specific antigen. For the important role OX40 takes in the process of immunity, many clinical trials are focusing on OX40 to find out whether it may have active effects in clinical cancer treatment. The results of clinical trials are still not enough. So, we reviewed the OX40 and its ligand (OX40L) function in cancer, clinical trials with OX40/OX40L and the correlation between OX40/OX40L and other immune checkpoints to add more ideas to tumor feasible treatment.
Keywords: cancer, immune checkpoints, OX40/OX40L, immunotherapy
Immunotherapy has shown promising results in cancer treatment,1 cancer immune checkpoint blockades also have got good results.2–5 It was demonstrated that combining cancer vaccines or checkpoint inhibitors with different immunotherapeutic agents could augment the anti-tumor effects and get better results in cancer patients.6,7
Tumor necrosis factor receptor superfamily member 4 (OX40) (CD134) and OX40 ligand (OX40L) (CD134L) (CD252) are on chromosome 1. The OX40 and OX40L could be expressed by endothelial cells, mast cells, activated natural killer (NK) cells, dendritic cells (DCs), B cells, microglial cells, activated T cells and Foxp3+ regulatory T cells.8–10 OX40L could initiate OX40 signals in activated T cells. OX40L on T cells could provide signals via the interactions between T cells and upregulate the anti-apoptotic protein on T cells to enhance T cell survival, cytokine production and induce the CD4 memory T cell expansion.11–16 The co-stimulation in B cells through the OX40/OX40L pathway contributed to CD4 cell generation, survival and T helper 2 (Th2) development.17 OX40/OX40L could promote NK cell activation, cytokine production and cytotoxicity and enhance targeted cells lysis.18,19 Mast cell via the OX40/OX40L pathway could induce T cell proliferation.20,21 OX40 on Treg cells played an important role in Treg cell development and homeostasis.22,23 We made a figure to clarify the function of OX40-OX40L pathway (Figure 1).
 |
Figure 1 OX40–OX40L interaction model.Abbreviations: Th2, T helper 2; NK, natural killer; TCR, T cell receptor; MHC, major histocompatibility complex; APC, antigen presenting cell. |
OX40/OX40L and diseases
Many diseases were associated with OX40/OX40L, so many researchers focused on it to find new way of treatment. The activation of OX40 promoted the generation and expansion of activated T cells and memory T cells, thus aggravating autoimmune diseases like Graves’ disease, autoimmune arthritis and uveitis.24–27 OX40 was critically important in sustaining the anti-viral immune response during the viral infection.19,28–30 OX40–OX40L signaling increased the adaptive immune response to an allograft by promoting effector and memory T cell survival. And blockade of OX40–OX40L interaction could decrease the T cells infiltration in the targeted organs to prevent allograft rejection.31–34 OX40L could promote the inflammatory cells infiltration into lesional tissues, leading to the pathological fibrosis in skin and internal organs. And blocking OX40–OX40L regressed the fibrosis.35,36 OX40–OX40L interaction on immune cells might contribute to idiopathic inflammatory myopathies through different pathways in the inflamed muscle.37 OX40/OX40L pathway was involved in the pathological process of Crohn’s disease (CD). And blockade anti-OX40 might be beneficial for the treatment by controlling the T cell-mediated inflammatory in vivo.38,39 Data implicated that OX40/OX40L participated in pathophysiology of acute myeloid leukemiaand also enhanced NK cell cytotoxicity.18
OX40/OX40L and cancer
OX40 was expressed on the tumor-infiltrating lymphocytes (TIL) in head and neck squamous cell carcinoma, ovarian cancer, gastric cancer, cutaneous squamous cell carcinoma, breast cancer and colorectal cancer.40–45 Agonistic anti-OX40 antibodies had anti-tumor effects.46–52 OX40 triggering regressed Treg cells, allowing DCs to reach the draining lymph nodes and prime the specific CD8 lymphocytes response to the tumor.48,53 Many research focused on the anti-tumor immunotherapy, based on activating costimulatory molecules OX40 and OX40L. Here, we showed some of them (Table 1).
 |
Table 1 OX40/OX40L and cancer |
Clinical trials of OX40/OX40L
Based on the role of OX40 and OX40L in the immune system, more and more research focused on its therapeutic effects. Many companies detected the immune checkpoints OX40 and OX40L, searching for the new approaches to treat tumors and autoimmune diseases, many of which are now making great advance in clinical development (Table 2). The results of clinical trials showed the OX40, as a potent immune-stimulating target, played an important role in anti-tumor therapy. The agonist anti-OX40 increased CD4 FoxP3− and CD8 T cells proliferation and the response to the tumor-specific antigen, enhancing both humoral and cellular immunity in cancer treatment.49
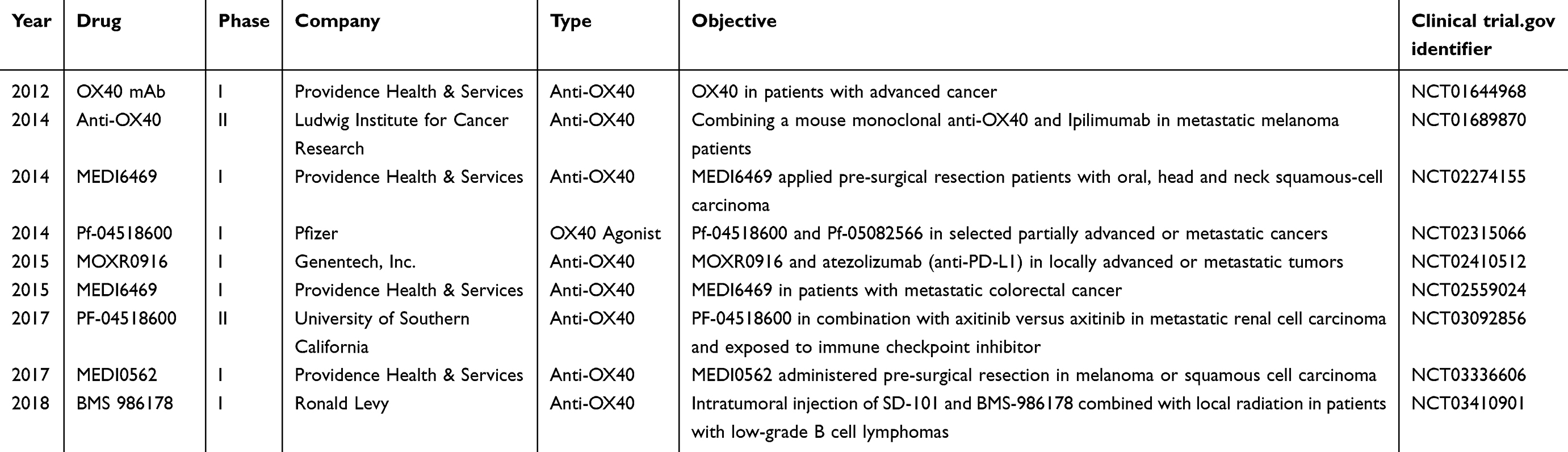 |
Table 2 Clinical trials with anti-OX40 |
Correlation of OX40/OX40L and other immune checkpoints
The results of studies suggested that some diseases were not sensitive to antibody therapy alone. So, it was necessary to study on the relationship between checkpoints to work out more effective treatment. CTLA-4, a molecule on T cells, inhibited the proliferation of T cells and cytokine production, thus limiting the lymphocyte immune reaction.68–72 Anti-CTLA-4 blockade induced the depletion of Treg cells within tumor and activation of Teff cells.71,73–76 Combining agonist anti-OX40 and antagonist anti-CTLA-4 further enhanced CD4 and CD8 T cells responses to antigen, indicating they had synergistic effects in improving tumor regression.77–79 And the cytokine of Th1 and Th2 CD4 T cells increased significantly.64 Whether the combination therapy altered the suppressive function of Treg cells remained deeper exploration.63,64 The combination was still more than the sum of its part.80
Programmed death-1 (PD-1) is a molecule that suppresses the immune reaction, inducing T cell exhaustion and apoptosis. Programmed death-ligand 1 (PD-L1), expressed on tumor cells or other tumor-related immune cells, could suppress anti-tumor immune response.81–84 The function of PD-1 and PD-L1 was affected by the complex immunoregulation. PD-1 blockade had already been used in cancer treatment and got a satisfying result.82,84 It was reported that PD-1 inhibitor added at the initiation of the cancer treatment could reduce the effects of OX40 agonist antibody, for it might cause the antigen-specific CD8+ T cell diminishment.85 And timing of PD-1 blockade using might determine whether it was effective immunotherapy when combined with OX40 therapy.81 In most cases, OX40 agonist and PD-1 blockade had a synergistic effect in disease treatment. OX40, combined with CD27 mediated co-stimulation, could synergize with PD-L1 inhibitor by activating CD8+ T cells.86 Combining OX40 stimulation and PD-L1 blockade could synergistically augment hepatitis B virus (HBV)-specific CD4 T cell responses by promoting Th cells to secrete IFN-γ and IL-21 in patients with HBV infection.87 In some poorly immunogenic tumors, combining PD-1 blockade and OX40 stimulation had an anti-tumor effect by inducing cytotoxic T lymphocyte, increasing the Teff cells and decreasing the immunosuppressive cells, while individual did not.41
4-1BB (CD137), member of the TNFR family enhanced T cell proliferation, effector function and cytokines production, and induced maturation of DC, thus increasing the immune reaction.88–93 Agonistic anti- 4-1 BB increased the TIL within tumor and upregulated the expression of 4-1 BB on the immune cells, augmenting anti-tumor reaction.90,94,95 The costimulatory pathway of OX40–OX40L and 4-1 BB-4-1 BBL functioned independently to enhance immune cells response.88 The combination of OX40 agonist and 4-1BB agonist induced profound expansion of CD8 T cell.96,97 But the response of CD4 T cell to the dual costimulation seemed to be additive instead of synergistic.98 On the whole, the combination therapy could synergistically inhibit cancer by producing more enhanced signals.98,99
Summary
Immune checkpoints play vital roles in cancer treatment. It was proved that the agonist anti-OX40/OX40L could enhance anti-tumor response by promoting the function of immune cells. More and more researchers focused on OX40/OX40L in cancer immunotherapy. But until now, the effects of OX40/OX40L treatment are still limited. Researchers are devoted to combine OX40/OX40L with other immune checkpoints in cancer treatment, which had also made some achievements, but the mechanisms of the synergy between OX40/OX40L and other immune checkpoints still need to be further studied.
Acknowledgment
This study was supported in part by grants from National Natural Science Foundation of China (81802255), Shanghai Pujiang Program (17PJD036), Shanghai Municipal Commission of Health and Family Planning Program (20174Y0131), National Key Research & Development Project (2016YFC0902300), Major Disease Clinical Skills Enhancement Program of three year action plan for promoting clinical skills and clinical innovation in municipal hospitals, Shanghai Shen Kang Hospital Development Center Clinical Research Plan of SHDC (16CR1001A),“Dream Tutor" Outstanding Young Talents Program (fkyq1901), Key Disciplines of Shanghai Pulmonary Hospital (2017ZZ02012), and Shanghai Science and Technology Commission (16JC1405900).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Couzin-Frankel J. Breakthrough of the year 2013. Cancer immunotherapy. Science. 2013;342:1432–1433. doi:10.1126/science.342.6165.1432
2. He Y, Rivard CJ, Rozeboom L, et al. Lymphocyte-activation gene-3, an important immune checkpoint in cancer. Cancer Sci. 2016;107:1193–1197. doi:10.1111/cas.12986
3. He Y, Rozeboom L, Rivard CJ, et al. PD-1, PD-L1 protein expression in non-small cell lung cancer and their relationship with tumor-infiltrating lymphocytes. Med Sci Monit. 2017;23:1208–1216. doi:10.12659/msm.899909
4. He Y, Rozeboom L, Rivard CJ, et al. MHC class II expression in lung cancer. Lung Cancer. 2017;112:75–80. doi:10.1016/j.lungcan.2017.07.030
5. He Y, Yu H, Rozeboom L, et al. LAG-3 protein expression in non-small cell lung cancer and its relationship with PD-1/PD-L1 and tumor-infiltrating lymphocytes. J Thorac Oncol. 2017;12:814–823. doi:10.1016/j.jtho.2017.01.019
6. Bilusic M, Madan RA, Gulley JL. Immunotherapy of prostate cancer: Facts and hopes. Clin Cancer Res. 2017;23:6764–6770. doi:10.1158/1078-0432.ccr-17-0019
7. Dushyanthen S, Teo ZL, Caramia F, et al. Agonist immunotherapy restores T cell function following MEK inhibition improving efficacy in breast cancer. Nat Commun. 2017;8:606. doi:10.1038/s41467-017-00728-9
8. Carboni S, Aboul-Enein F, Waltzinger C, Killeen N, Lassmann H, Peña-Rossi C. CD134 plays a crucial role in the pathogenesis of EAE and is upregulated in the CNS of patients with multiple sclerosis. J Neuroimmunol. 2003;145:1–11.
9. Ali SA, Ahmad M, Lynam J, et al. Anti-tumour therapeutic efficacy of OX40L in murine tumour model. Vaccine. 2004;22:3585–3594. doi:10.1016/j.vaccine.2004.03.041
10. Kim BS, Kim JY, Kim EJ, et al. Role of thalidomide on the expression of OX40, 4-1BB, and GITR in T cell subsets. Transplant Proc. 2016;48:1270–1274. doi:10.1016/j.transproceed.2015.12.088
11. Soroosh P, Ine S, Sugamura K, Ishii N. OX40-OX40 ligand interaction through T cell-T cell contact contributes to CD4 T cell longevity. J Immunol. 2006;176:5975–5987. doi:10.4049/jimmunol.176.10.5975
12. Rogers PR, Song J, Gramaglia I, Killeen N, Croft M. OX40 promotes Bcl-xL and Bcl-2 expression and is essential for long-term survival of CD4 T cells. Immunity. 2001;15:445–455.
13. Jones RG, Parsons M, Bonnard M, et al. Protein kinase B regulates T lymphocyte survival, nuclear factor kappaB activation, and Bcl-X(L) levels in vivo. J Exp Med. 2000;191:1721–1734. doi:10.1084/jem.191.10.1721
14. Kinnear G, Wood KJ, Marshall D, Jones ND. Anti-OX40 prevents effector T-cell accumulation and CD8+ T-cell mediated skin allograft rejection. Transplantation. 2010;90:1265–1271. doi:10.1097/TP.0b013e3181fe5396
15. Flynn S, Toellner KM, Raykundalia C, Goodall M, Lane P. CD4 T cell cytokine differentiation: the B cell activation molecule, OX40 ligand, instructs CD4 T cells to express interleukin 4 and upregulates expression of the chemokine receptor, Blr-1. J Exp Med. 1998;188:297–304. doi:10.1084/jem.188.2.297
16. Vu MD, Clarkson MR, Yagita H, Turka LA, Sayegh MH, Li XC. Critical, but conditional, role of OX40 in memory T cell-mediated rejection. J Immunol. 2006;176:1394–1401. doi:10.4049/jimmunol.176.3.1394
17. Linton P-J, Bautista B, Biederman E, et al. Costimulation via OX40L expressed by B cells is sufficient to determine the extent of primary CD4 cell expansion and Th2 cytokine secretion in vivo. J Exp Med. 2003;197:875–883. doi:10.1084/jem.20021290
18. Nuebling T, Schumacher CE, Hofmann M, et al. The immune checkpoint modulator OX40 and its ligand OX40L in NK-cell immunosurveillance and acute myeloid leukemia. Cancer Immunol Res. 2018;6:209–221. doi:10.1158/2326-6066.CIR-17-0212
19. Pollmann J, Götz -J-J, Rupp D, et al. Hepatitis C virus-induced natural killer cell proliferation involves monocyte-derived cells and the OX40/OX40L axis. J Hepatol. 2018;68:421–430. doi:10.1016/j.jhep.2017.10.021
20. Turaj AH, Cox KL, Penfold CA, et al. Augmentation of CD134 (OX40)-dependent NK anti-tumour activity is dependent on antibody cross-linking. Sci Rep. 2018;8:2278. doi:10.1038/s41598-018-20656-y
21. Li F, Wang Y, Lin L, et al. Mast cell-derived exosomes promote Th2 cell differentiation via OX40L-OX40 ligation. J Immunol Res. 2016;2016:1–10. doi:10.1155/2016/3623898
22. Takeda I, Ine S, Killeen N, et al. Distinct roles for the OX40-OX40 ligand interaction in regulatory and nonregulatory T cells. J Immunol. 2004;172:3580–3589. doi:10.4049/jimmunol.172.6.3580
23. Piconese S, Pittoni P, Burocchi A, et al. A non-redundant role for OX40 in the competitive fitness of Treg in response to IL-2. Eur J Immunol. 2010;40:2902–2913. doi:10.1002/eji.201040505
24. Wang Q, Shi B-M, Xie F, et al. Enhancement of CD4(+) T cell response and survival via coexpressed OX40/OX40L in Graves’ disease. Mol Cell Endocrinol. 2016;430:115–124. doi:10.1016/j.mce.2016.04.008
25. Jiang J, Liu C, Liu M, et al. OX40 signaling is involved in the autoactivation of CD4(+)CD28(-) T cells and contributes to the pathogenesis of autoimmune arthritis. Arthritis Res Ther. 2017;19:67. doi:10.1186/s13075-017-1261-9
26. Wu X, Rosenbaum JT, Adamus G, et al. Activation of OX40 prolongs and exacerbates autoimmune experimental uveitis. Invest Ophthalmol Vis Sci. 2011;52:8520–8526. doi:10.1167/iovs.11-7664
27. Yoshioka T, Nakajima A, Akiba H, et al. Contribution of OX40/OX40 ligand interaction to the pathogenesis of rheumatoid arthritis. Eur J Immunol. 2000;30:2815–2823. doi:10.1002/1521-4141(200010)30:10<2815::AID-IMMU2815>3.0.CO;2-#
28. Boettler T, Moeckel F, Cheng Y, et al. OX40 facilitates control of a persistent virus infection. PLoS Pathog. 2012;8:e1002913. doi:10.1371/journal.ppat.1002913
29. Tahiliani V, Hutchinson TE. OX40 cooperates with ICOS to amplify follicular th cell development and germinal center reactions during infection. J Immunol. 2017;198:218–228. doi:10.4049/jimmunol.1601356
30. Boettler T, Choi YS, Salek-Ardakani S, et al. Exogenous OX40 stimulation during lymphocytic choriomeningitis virus infection impairs follicular Th cell differentiation and diverts CD4 T cells into the effector lineage by upregulating Blimp-1. J Immunol. 2013;191:5026–5035. doi:10.4049/jimmunol.1300013
31. Curry AJ, Chikwe J, Smith XG, et al. OX40 (CD134) blockade inhibits the co-stimulatory cascade and promotes heart allograft survival. Transplantation. 2004;78:807–814. doi:10.1097/01.tp.0000131670.99000.54
32. Tripathi T, Yin W, Xue Y, et al. Central roles of OX40L-OX40 interaction in the induction and progression of human T cell-driven acute graft-versus-host disease. ImmunoHorizons. 2019;3:110–120. doi:10.4049/immunohorizons.1900001
33. Li T, Ma R, Zhu J, Wang F, Huang L, Leng X. Blockade of the OX40/OX40L pathway and induction of PD-L1 synergistically protects mouse islet allografts from rejection. Chin Med J. 2014;127:2686–2692.
34. Kinnear G, Wood KJ, Fallah-Arani F, Jones ND. A diametric role for OX40 in the response of effector/memory CD4+ T cells and regulatory T cells to alloantigen. J Immunol. 2013;191:1465–1475. doi:10.4049/jimmunol.1300553
35. Elhai M, Avouac J, Hoffmann-Vold AM, et al. OX40L blockade protects against inflammation-driven fibrosis. Proc Natl Acad Sci U S A. 2016;113:E3901–E3910. doi:10.1073/pnas.1523512113
36. Boleto G, Allanore Y, Avouac J. Targeting costimulatory pathways in systemic sclerosis. Front Immunol. 2018;9:2998. doi:10.3389/fimmu.2018.02998
37. Papadopoulos C, Terzis G, Papadimas GK, Manta P. OX40-OX40L expression in idiopathic inflammatory myopathies. Anal Quant Cytol Histol. 2013;35:17–26.
38. Totsuka T, Kanai T, Uraushihara K, et al. Therapeutic effect of anti-OX40L and anti-TNF-alpha MAbs in a murine model of chronic colitis. Am J Physiol Gastrointest Liver Physiol. 2003;284:G595–G603. doi:10.1152/ajpgi.00450.2002
39. Griseri T, Asquith M, Thompson C, Powrie F. OX40 is required for regulatory T cell-mediated control of colitis. J Exp Med. 2010;207:699–709. doi:10.1084/jem.20091618
40. Bell RB, Leidner RS, Crittenden MR, et al. OX40 signaling in head and neck squamous cell carcinoma: overcoming immunosuppression in the tumor microenvironment. Oral Oncol. 2016;52:1–10. doi:10.1016/j.oraloncology.2015.11.009
41. Guo Z, Wang X, Cheng D, et al. PD-1 blockade and OX40 triggering synergistically protects against tumor growth in a murine model of ovarian cancer. PLoS One. 2014;9:e89350. doi:10.1371/journal.pone.0089350
42. Martins MR, Santos RLD, Jatahy KDN, et al. Could OX40 agonist antibody promote activation of the anti-tumor immune response in gastric cancer? J Surg Oncol. 2018;117:840–844. doi:10.1002/jso.25001
43. Lai C, August S, Albibas A, et al. OX40+ regulatory T cells in cutaneous squamous cell carcinoma suppress effector T-cell responses and associate with metastatic potential. Clin Cancer Res. 2016;22:4236–4248. doi:10.1158/1078-0432.ccr-15-2614
44. Hamidinia M, Ghafourian Boroujerdnia M, Talaiezadeh A, Solgi G, Taghdiri M, Khodadadi A. Concomitant increase of OX40 and FOXP3 transcripts in peripheral blood of patients with breast cancer. Iran J Immunol. 2013;10:22–30. doi:IJIv10i1A3
45. Weixler B, Cremonesi E, Sorge R, et al. OX40 expression enhances the prognostic significance of CD8 positive lymphocyte infiltration in colorectal cancer. Oncotarget. 2015;6:37588–37599. doi:10.18632/oncotarget.5940
46. Zhang P, Tu GH, Wei J, et al. Ligand-Blocking and Membrane-Proximal Domain Targeting Anti-OX40 Antibodies Mediate Potent T Cell-Stimulatory and Anti-Tumor Activity. Cell Rep. 2019;27:3117–3123.e5. doi:10.1016/j.celrep.2019.05.027
47. Aspeslagh S, Postel-Vinay S, Rusakiewicz S, Soria J-C, Zitvogel L, Marabelle A. Rationale for anti-OX40 cancer immunotherapy. Eur J Cancer. 2016;52:50–66. doi:10.1016/j.ejca.2015.08.021
48. Buchan SL, Rogel A, Al-Shamkhani A. The immunobiology of CD27 and OX40 and their potential as targets for cancer immunotherapy. Blood. 2018;131:39–48. doi:10.1182/blood-2017-07-741025
49. Curti BD, Kovacsovics-Bankowski M, Morris N, et al. OX40 is a potent immune-stimulating target in late-stage cancer patients. Cancer Res. 2013;73:7189–7198. doi:10.1158/0008-5472.CAN-12-4174
50. Foote JB, Kok M, Leatherman JM, et al. A STING agonist given with OX40 receptor and PD-L1 modulators primes immunity and reduces tumor growth in tolerized mice. Blood. 2017;5:468–479. doi:10.1182/blood-2017-07-741025; 10.1158/2326-6066.cir-16-0284.
51. Moran AE, Kovacsovics-Bankowski M, Weinberg AD. The TNFRs OX40, 4-1BB, and CD40 as targets for cancer immunotherapy. Curr Opin Immunol. 2013;25:230–237. doi:10.1016/j.coi.2013.01.004
52. Sanmamed MF, Pastor F, Rodriguez A, et al. Agonists of co-stimulation in cancer immunotherapy directed against CD137, OX40, GITR, CD27, CD28, and ICOS. Semin Oncol. 2015;42:640–655. doi:10.1053/j.seminoncol.2015.05.014
53. Weinberg AD, Rivera MM, Prell R, et al. Engagement of the OX-40 receptor in vivo enhances antitumor immunity. J Immunol. 2000;164:2160–2169. doi:10.4049/jimmunol.164.4.2160
54. Kjaergaard J, Tanaka J, Kim JA, Rothchild K, Weinberg A, Shu S. Therapeutic efficacy of OX-40 receptor antibody depends on tumor immunogenicity and anatomic site of tumor growth. Cancer Res. 2000;60:5514–5521.
55. Morris A, Vetto JT, Ramstad T, et al. Induction of anti-mammary cancer immunity by engaging the OX-40 receptor in vivo. Breast Cancer Res Treat. 2001;67:71–80.
56. Petty JK, He K, Corless CL, Vetto JT, Weinberg AD. Survival in human colorectal cancer correlates with expression of the T-cell costimulatory molecule OX-40 (CD134). Am J Surg. 2002;183:512–518. doi:10.1016/s0002-9610(02)00831-0
57. Andarini S, Kikuchi T, Nukiwa M, et al. Adenovirus vector-mediated in vivo gene transfer of OX40 ligand to tumor cells enhances antitumor immunity of tumor-bearing hosts. Cancer Res. 2004;64:3281–3287.
58. Song A, Tang X, Harms KM, Croft M. OX40 and Bcl-xL promote the persistence of CD8 T cells to recall tumor-associated antigen. J Immunol. 2005;175:3534–3541. doi:10.4049/jimmunol.175.6.3534
59. Gough MJ, Ruby CE, Redmond WL, Dhungel B, Brown A, Weinberg AD. OX40 agonist therapy enhances CD8 infiltration and decreases immune suppression in the tumor. Cancer Res. 2008;68:5206–5215. doi:10.1158/0008-5472.CAN-07-6484
60. Pham Minh N, Murata S, Kitamura N, et al. In vivo antitumor function of tumor antigen-specific CTLs generated in the presence of OX40 co-stimulation in vitro. Int J Cancer. 2018;142:2335–2343. doi:10.1002/ijc.31244
61. Assudani DP, Ahmad M, Li G, Rees RC, Ali SA. Immunotherapeutic potential of DISC-HSV and OX40L in cancer. Cancer Immunol Immunother. 2006;55:104–111. doi:10.1007/s00262-005-0004-y
62. Zaini J, Andarini S, Tahara M, et al. OX40 ligand expressed by DCs costimulates NKT and CD4+ Th cell antitumor immunity in mice. J Clin Invest. 2007;117:3330–3338. doi:10.1172/JCI32693
63. Redmond WL, Linch SN, Kasiewicz MJ. Combined targeting of costimulatory (OX40) and coinhibitory (CTLA-4) pathways elicits potent effector T cells capable of driving robust antitumor immunity. Cancer Immunol Res. 2014;2:142–153. doi:10.1158/2326-6066.CIR-13-0031-T
64. Linch SN, Kasiewicz MJ, McNamara MJ, Hilgart-Martiszus IF, Farhad M, Redmond WL. Combination OX40 agonism/CTLA-4 blockade with HER2 vaccination reverses T-cell anergy and promotes survival in tumor-bearing mice. Proc Natl Acad Sci U S A. 2016;113:E319–E327. doi:10.1073/pnas.1510518113
65. Berrong Z, Mkrtichyan M, Ahmad S, et al. Antigen-specific antitumor responses induced by OX40 agonist are enhanced by the IDO inhibitor indoximod. Cancer Immunol Res. 2018;6:201–208. doi:10.1158/2326-6066.CIR-17-0223
66. Jahan N, Talat H, Curry WT. Agonist OX40 immunotherapy improves survival in glioma-bearing mice and is complementary with vaccination with irradiated GM-CSF-expressing tumor cells. Neuro-oncology. 2018;20:44–54. doi:10.1093/neuonc/nox125
67. Virani NA, Thavathiru E, McKernan P, Moore K, Benbrook DM, Harrison RG. Anti-CD73 and anti-OX40 immunotherapy coupled with a novel biocompatible enzyme prodrug system for the treatment of recurrent, metastatic ovarian cancer. Cancer Lett. 2018;425:174–182. doi:10.1016/j.canlet.2018.03.027
68. Chikuma S. CTLA-4, an essential immune-checkpoint for T-cell activation. Curr Top Microbiol Immunol. 2017;410:99–126. doi:10.1007/82_2017_61
69. Chen X, Shao Q, Hao S, et al. CTLA-4 positive breast cancer cells suppress dendritic cells maturation and function. Oncotarget. 2017;8:13703–13715. doi:10.18632/oncotarget.14626
70. Shi L, Meng T, Zhao Z, et al. CRISPR knock out CTLA-4 enhances the anti-tumor activity of cytotoxic T lymphocytes. Gene. 2017;636:36–41. doi:10.1016/j.gene.2017.09.010
71. van Hooren L, Sandin LC, Moskalev I, et al. Local checkpoint inhibition of CTLA-4 as a monotherapy or in combination with anti-PD1 prevents the growth of murine bladder cancer. Eur J Immunol. 2017;47:385–393. doi:10.1002/eji.201646583
72. Zhao Y, Yang W, Huang Y, Cui R, Li X, Li B. Evolving roles for targeting CTLA-4 in cancer immunotherapy. Cell Physiol Biochem. 2018;47:721–734. doi:10.1159/000490025
73. Selby MJ, Engelhardt JJ, Quigley M, et al. Anti-CTLA-4 antibodies of IgG2a isotype enhance antitumor activity through reduction of intratumoral regulatory T cells. Cancer Immunol Res. 2013;1:32–42. doi:10.1158/2326-6066.cir-13-0013
74. Subrahmanyam PB, Dong Z, Gusenleitner D, et al. Distinct predictive biomarker candidates for response to anti-CTLA-4 and anti-PD-1 immunotherapy in melanoma patients. J Immunother Cancer. 2018;6:18. doi:10.1186/s40425-018-0328-8
75. Wang B, Qin L, Ren M, Sun H. Effects of combination of anti-CTLA-4 and anti-PD-1 on gastric cancer cells proliferation, apoptosis and metastasis. Cell Physiol Biochem. 2018;49:260–270. doi:10.1159/000492876
76. Wei SC, Levine JH, Cogdill AP, et al. Distinct cellular mechanisms underlie anti-CTLA-4 and anti-PD-1 checkpoint blockade. Cell. 2017;170:1120–1133.e17. doi:10.1016/j.cell.2017.07.024
77. Goncalves-Lopes RM, Lima NF, Carvalho KI, Scopel KKG, Kallás EG, Ferreira MU. Surface expression of inhibitory (CTLA-4) and stimulatory (OX40) receptors by CD4(+) regulatory T cell subsets circulating in human malaria. Microbes Infect. 2016;18:639–648. doi:10.1016/j.micinf.2016.06.003
78. Montler R, Bell RB, Thalhofer C, et al. OX40, PD-1 and CTLA-4 are selectively expressed on tumor-infiltrating T cells in head and neck cancer. Clin Transl Immunol. 2016;5:e70. doi:10.1038/cti.2016.16
79. Redmond WL, Linch SN, Kasiewicz MJ. Combined targeting of costimulatory (OX40) and coinhibitory (CTLA-4) pathways elicits potent effector T cells capable of driving robust antitumor immunity. Proc Natl Acad Sci U S A. 2014;2:142–153. doi:10.1073/pnas.1510518113; 10.1158/2326-6066.cir-13-0031-t.
80. Linch SN, Redmond WL. Combined OX40 ligation plus CTLA-4 blockade: more than the sum of its parts. Oncoimmunology. 2014;3:e28245. doi:10.4161/onci.28245
81. Messenheimer DJ, Jensen SM, Afentoulis ME, et al. Timing of PD-1 blockade is critical to effective combination immunotherapy with anti-OX40. Clin Cancer Res. 2017;23:6165–6177. doi:10.1158/1078-0432.CCR-16-2677
82. Chamoto K, Al-Habsi M, Honjo T. Role of PD-1 in immunity and diseases. Curr Top Microbiol Immunol. 2017;410:75–97. doi:10.1007/82_2017_67
83. Kuol N, Stojanovska L, Nurgali K, Apostolopoulos V. PD-1/PD-L1 in disease. Immunotherapy. 2018;10:149–160. doi:10.2217/imt-2017-0120
84. Salmaninejad A, Khoramshahi V, Azani A, et al. PD-1 and cancer: molecular mechanisms and polymorphisms. Immunogenetics. 2018;70:73–86. doi:10.1007/s00251-017-1015-5
85. Shrimali RK, Ahmad S, Verma V, et al. Concurrent PD-1 blockade negates the effects of OX40 agonist antibody in combination immunotherapy through inducing T-cell apoptosis. Cancer Immunol Res. 2017;5:755–766. doi:10.1158/2326-6066.CIR-17-0292
86. Buchan S, Manzo T, Flutter B, et al. OX40- and CD27-mediated costimulation synergizes with anti-PD-L1 blockade by forcing exhausted CD8+ T cells to exit quiescence. J Immunol. 2015;194:125–133. doi:10.4049/jimmunol.1401644
87. Jacobi FJ, Wild K, Smits M, et al. OX40 stimulation and PD-L1 blockade synergistically augment HBV-specific CD4 T cells in patients with HBeAg-negative infection. J Hepatol. 2019;70:1103–1113. doi:10.1016/j.jhep.2019.02.016
88. Dawicki W, Bertram EM, Sharpe AH, Watts TH. 4-1BB and OX40 act independently to facilitate robust CD8 and CD4 recall responses. J Immunol. 2004;173:5944–5951. doi:10.4049/jimmunol.173.10.5944
89. Cannons JL, Lau P, Ghumman B, et al. 4-1BB ligand induces cell division, sustains survival, and enhances effector function of CD4 and CD8 T cells with similar efficacy. J Immunol. 2001;167:1313–1324. doi:10.4049/jimmunol.167.3.1313
90. Chester C, Ambulkar S, Kohrt HE. 4-1BB agonism: adding the accelerator to cancer immunotherapy. Cancer Immunol Immunother. 2016;65:1243–1248. doi:10.1007/s00262-016-1829-2
91. Chester C, Sanmamed MF, Wang J, Melero I. Immunotherapy targeting 4-1BB: mechanistic rationale, clinical results, and future strategies. J Biol Chem. 2018;131:49–57. doi:10.1074/jbc.M117.814905; 10.1182/blood-2017-06-741041.
92. Lu Y, Li C, Du S, et al. 4-1BB signaling promotes alveolar macrophages-mediated pro-fibrotic responses and crystalline silica-induced pulmonary fibrosis in mice. Front Immunol. 2018;9:1848. doi:10.3389/fimmu.2018.01848
93. Oh HS, Choi BK, Kim YH, et al. 4-1BB signaling enhances primary and secondary population expansion of CD8+ T cells by maximizing autocrine IL-2/IL-2 receptor signaling. PLoS One. 2015;10:e0126765. doi:10.1371/journal.pone.0126765
94. Bartkowiak T, Curran MA. 4-1BB agonists: multi-potent potentiators of tumor immunity. Front Oncol. 2015;5:117. doi:10.3389/fonc.2015.00117
95. Sakellariou-Thompson D, Forget M-A, Creasy C, et al. 4-1BB agonist focuses CD8(+) tumor-infiltrating T-cell growth into a distinct repertoire capable of tumor recognition in pancreatic cancer. Clin Cancer Res. 2017;23:7263–7275. doi:10.1158/1078-0432.ccr-17-0831
96. Munks MW, Mourich DV, Mittler RS, Weinberg AD, Hill AB. 4-1BB and OX40 stimulation enhance CD8 and CD4 T-cell responses to a DNA prime, poxvirus boost vaccine. Immunology. 2004;112:559–566. doi:10.1111/j.1365-2567.2004.01917.x
97. Hendriks J, Xiao Y, Rossen JWA, et al. During viral infection of the respiratory tract, CD27, 4-1BB, and OX40 collectively determine formation of CD8+ memory T cells and their capacity for secondary expansion. J Immunol. 2005;175:1665–1676. doi:10.4049/jimmunol.175.3.1665
98. Lee SJ, Myers L, Muralimohan G, et al. 4-1BB and OX40 dual costimulation synergistically stimulate primary specific CD8 T cells for robust effector function. J Immunol. 2004;173:3002–3012. doi:10.4049/jimmunol.173.5.3002
99. Lee SW, Park Y, Song A, Cheroutre H, Kwon BS, Croft M. Functional dichotomy between OX40 and 4-1BB in modulating effector CD8 T cell responses. J Immunol. 2006;177:4464–4472. doi:10.4049/jimmunol.177.7.4464
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.