Back to Journals » OncoTargets and Therapy » Volume 11
Overexpression of long noncoding RNA LINC00882 is associated with poor prognosis in hepatocellular carcinoma
Authors Zhu L ![]() , Huang F
, Huang F ![]() , Wan T, Xu H
, Wan T, Xu H ![]() , Zhao Q
, Zhao Q
Received 11 April 2018
Accepted for publication 17 May 2018
Published 13 September 2018 Volume 2018:11 Pages 5209—5217
DOI https://doi.org/10.2147/OTT.S170825
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Sanjeev K. Srivastava
Lei Zhu,1 Feizhou Huang,1 Tao Wan,1 Hongbo Xu,1 Qian Zhao2,3
1Department of Hepatobiliary and Pancreatic Surgery, the Third Xiangya Hospital of Central South University, Changsha, Hunan 410013, People’s Republic of China; 2Department of Adult Stem Cells, Institute of Reproduction and Stem Cell Engineering, Central South University, Changsha, Hunan 410078, People’s Republic of China; 3Institute of Transformation Medicine Affiliated to Hunan Yearth Biotechnology Co., Ltd., Changsha, Hunan 410200, People’s Republic of China
Introduction: Hepatocellular carcinoma (HCC) is the third major cause of malignant tumor-related death worldwide because it is initially diagnosed in the advanced stage and its therapeutic outcomes are usually poor. Based on this, it is urgent to identify effective early diagnosis biomarkers and new therapeutic targets to promote HCC treatment. Long noncoding RNAs (lncRNAs) have been reported as promising biomarkers for tumor diagnosis and treatment.
Materials and methods: In this study, we profiled expression patterns and dysregulation of lncRNAs in HCC tissues by analyzing two datasets GSE55191 and GSE64631 from Gene Expression Omnibus database firstly, each of which contains expression profiles of 3 primary HCC tissues and 3 normal liver tissues, respectively.
Results: LncRNA 882 (LINC00882) is one of the lncRNAs that was significantly upregulated in HCC tissues compared with normal liver tissues. We verified the upregulation of LINC00882 in HCC by using two separate cohorts that contained 85 HCC tissues and paired adjacent noncancerous tissues and 86 HCC tissues and 89 independent noncancerous liver tissues, respectively.
Discussion: We found that upregulation of LINC00882 is correlated with poorer prognosis of HCC patients. In vitro cell experiments demonstrated that knockdown of LINC00882 inhibits proliferation, migration and invasion of HCC cell lines.
Conclusion: These results indicated that LINC00882 promotes HCC progression and could be a potential prognostic biomarker and therapeutic target for HCC.
Keywords: hepatocellular carcinoma, long noncoding RNAs, LINC00882, proliferation, migration
Introduction
Hepatocellular carcinoma (HCC) is the most prevalent subtype of liver cancer, which is associated with different risk factors, including chronic infections with hepatitis B virus (HBV) or hepatitis C virus, alcohol addiction, cirrhosis, and non-alcoholic fatty liver disease.1–3 Certain cellular situations such as tumor microenvironment, hypoxia, oxidative stress and inflammation also contribute to the onset and development of HCC.1
Accumulating evidence indicated that long noncoding RNAs (lncRNAs) have participated in a lot of biological processes, such as cell proliferation and cell migration, and played important roles in cancer development, progression, and recurrence.4–7 Potential molecular mechanisms of lncRNAs in tumor progression are currently of great concern. Considering the key functional roles of lncRNAs in tumorigenesis, discovery of critical lncRNAs in different cancer types will be valuable for developing lncRNA-based diagnostic and therapeutic strategies in the future.
In the recent years, some lncRNAs have been shown to be dysregulated in HCC and act as important regulators of HCC development, partially achieved by their spatiotemporally specific expression patterns in different cell types.8,9 Up to now, more than 10 lncRNAs have been shown to be increased in HCC.10 On the contrary, some lncRNAs are decreased in HCC. It was believed that lncRNAs play critical roles in HCC progression and may potentially serve as biomarkers for diagnosis and treatment of HCC. However, the functional roles of lncRNAs have not been fully clarified.
In this research, we profiled lncRNA expression patterns and dysregulation in HCC firstly by analyzing two datasets, GSE55191 and GSE64631, from Gene Expression Omnibus (GEO) database, each of which contains expression profiles of 3 primary HCC specimen and 3 normal liver tissues, respectively. LncRNA 882 (LINC00882) is among the lncRNAs that were detected to be significantly upregulated in HCC tissues. We verified the overexpression of LINC00882 in HCC by using two separate cohorts that contained 85 HCC tissues and paired adjacent noncancerous tissues and 86 HCC tissues and 89 independent noncancerous liver tissues, respectively. We further found that upregulation of LINC00882 is correlated with poorer prognosis of HCC patients. In vitro cell assays demonstrated that knockdown of LINC00882 inhibits proliferation, migration, and invasion of HCC cell lines. These data indicated that LINC00882 promotes HCC progression and is a potential prognostic biomarker and therapeutic target for HCC.
Materials and methods
Cell culture and gene silencing
The HCC cell lines HCCLM3 and Hep3B were obtained from ATCC and were cultured under the condition of 5% CO2 at 37°C with DMEM/F12 medium supplemented with 1% antibiotics (100 U/mL penicillin and 100 μg/mL streptomycin sulfates) and 10% fetal bovine serum (FBS; Thermo Fisher Scientific, Waltham, MA, USA).
Three siRNA sequences targeting LINC00882 were designed and synthesized. Transient transfection was carried out in Hep3B, and the one with the best interference efficiency (siRNA-314) was selected to synthesize short hairpin RNA (shRNA) and lentiviral packaging. The sequence information was listed as follows: siRNA-314: 5′-GCTGAGATGTCACATGGCA-3′, siRNA-404: 5′-CCTGCCATCTTATGCAGAA-3′, siRNA-542: 5′-GATCTAATTCAAGTGCCAA-3′.
About 2 × 105 HCC cells were seeded into 6-well plates 24 hours before infection performance. The medium was replaced with the virus-containing supernatant, which was supplemented with 8 μg/mL polybrene (Genomeditech, Shanghai, People’s Republic of China). The medium was replaced with fresh medium again 8 hours after infection. The infected cells were selected using 1 μg/mL puromycin to obtain LINC00882-silenced HCC cells.
Vector construction and lentivirus packaging
The LINC00882 amplification fragment was cloned into the lentivirus vector PGMLV-PA6 (Genomeditech). The specific primer sequences for PCR amplification were as follows: ccgGAATTCAGGCTCGGCGTTGGCCTCC (EcoR I), ccgGGATCCCTGAATTGCTTTTATTGTTTTGTG (Bamh I). 293FT cells were co-transfected with LINC00882, VSVG and PGMLV using the FuGene transfection reagent (Hoffman-La Roche Ltd., Basel, Switzerland) for lentivirus packaging. The medium was harvested 48 hours later, and the supernatant was filtered to remove cell debris and used for infection.
In situ hybridization
Expressions of LINC00882 in tissue microarrays containing HCC specimen and noncancerous liver specimen were detected by in situ hybridization. Three probes targeting different LINC00882 regions (Ribo™ lncRNA FISH Probe Mix; RiboBio, Guangzhou, People’s Republic of China) were synthesized and labeled with DIG-dUTP at the 3′ end. GAPDH probes (lnc110102 h-18S FISH Probe Mix; RIBO Biotech, Guangzhou, People’s Republic of China) were used as controls. A scoring criterion, which counts both the staining intensity and the number of positively stained cells, was used for semi-quantification of in situ hybridization. The scoring was graded as 0 (negative), 1 (<10% positive), 2 (10%–50% positive) or 3 (>50% positive) according to the positively stained proportion and staining intensity of cells. The final scores ranged from 0 to 1, which were regarded as low expression, and scores 2–3 were regarded as high expression. Five representative visual fields were selected randomly by two pathologists to score.
RNA isolation and qRT-PCR
Total RNAs were extracted from the cells using TRIzol reagent (Invitrogen, Carlsbad, CA, USA). One microgram of total RNA from the samples was reverse transcribed using a Reverse Transcription Kit (Bio-Rad, Hercules, CA, USA). Quantitative real-time PCR was performed to detect the relative mRNA levels of LINC00882 and various cancer-related genes using SYBR Green Supermix (Bio-Rad) in the CFX96 Real-Time PCR Detection System (Bio-Rad) according to the manufacturer’s instructions. All tests were carried out in 3 independent experiments. The primers are listed in Table 1. The quantification analysis for relative mRNA levels of target genes was performed using the relative quantification comparative Ct method.
 | Table 1 Primer sequence information used in qRT-PCR |
Western blotting
Western blot analysis was performed to detect the protein expression of proliferation markers and invasive markers according to the methods described in the literature.11
Cell proliferation, migration and invasion assays
Cell proliferation was analyzed using a Cell-Light™ EdU Cell Proliferation Detection kit (RiboBio). Control or LINC00882 loss-of-function stable cell models, which were labeled with 5-ethynyl-2′-deoxyuridine (EdU), were seeded at 1.5 × 105 cells/well in 24-well plates and then fixed with 4% paraformaldehyde. After DNA staining, cellular immunostaining was photographed using an epifluorescence microscope (Leica Microsystems, Wetzlar, Germany).
Wound healing assay was used to assess the migration ability of HCC cells. Cells were seeded in 6-well culture plates and grown to 90% confluence. Vertical wounds in the cell monolayer were created by a 10 μL tip and washed three times with PBS. Gap width was measured by microscopy at the designated time points.
Transwell assay was performed to evaluate the invasion ability of HCC cells. A total of 1 × 105 cells in 100 μL of 0.1% serum medium were placed in the upper of transwell cell culture chambers, which were pre-coated with Matrigel (1:8 dilution; BD Biosciences, San Jose, CA, USA), and medium containing 10% FBS was added to the lower chamber. After 24 hours of incubation at 37°C, the invasive cells were fixed with methanol and stained with 0.5% crystal violet. Numbers of invasive cells were counted and photographed using a phase contrast microscope.
Statistical analysis
All experiments were independently repeated three times. Statistical analysis was conducted using SPSS v17.0 (SPSS, Chicago, IL, USA) and presented with Graphpad Prism software. Differences between two independent groups were evaluated by Student’s t-tests, and differences for multiple comparisons were evaluated by one-way ANOVA. Overall survival (OS) was calculated using the Kaplan–Meier method; p ≤ 0.05 was regarded as statistically significant. Data are presented as mean ± SEs.
Results
Mining of dysregulated lncRNAs in HCC tissues
In order to identify novel lncRNAs differentially expressed between HCC cancer tissues and noncancerous liver tissues, we explored two GEO datasets, GSE55191 and GSE64631, each of which contains lncRNA expression profiles of 3 primary HCC specimen and 3 noncancerous liver tissues. By combination of lncRNA expression signatures from these two GEO datasets, some lncRNAs were revealed as dysregulated, most of which are not well identified and functionally unknown, such as LINC00882 (Table 2). LINC00882 was negatively associated with OS in chromophobe renal cell carcinoma.12 Altered expression of LINC00882 was also observed in lung tissues of patients with chronic obstructive pulmonary disease.13 To investigate the role of lncRNAs in HCC progression, we focused on LINC00882, which was consistently identified as one of the most upregulated lncRNAs in both datasets (Figure 1A, p < 0.01 in both datasets with 3.2-fold and 2.6-fold, respectively) and functionally unclear in human cancer.
 | Table 2 Results of differential expression of LINC00882 |
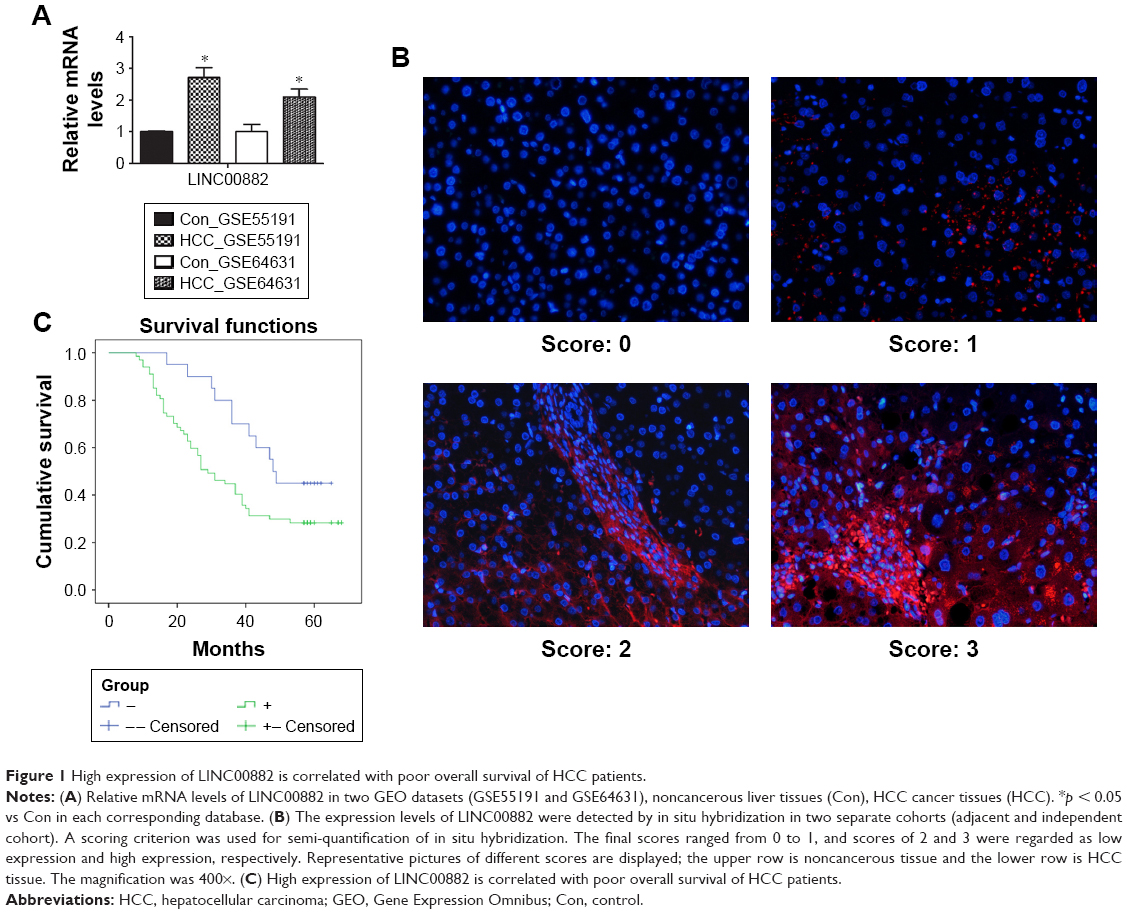 | Figure 1 High expression of LINC00882 is correlated with poor overall survival of HCC patients. |
To confirm the upregulation of LINC00882 expression in HCC tissues as identified in the GEO datasets GSE55191 and GSE64631, we further detected the expression levels of LINC00882 by in situ hybridization in two separate cohorts, which included 85 HCC tissues and paired adjacent noncancerous tissues and 86 HCC tissues and 89 independent noncancerous liver tissues, respectively. In consistency with the GEO datasets, the in situ hybridization data showed that the transcript level of LINC00882 in HCC tissues was remarkably upregulated compared to that in the adjacent or independent noncancerous liver tissues according to the result of the scores (Figure 1B).
High expression of LINC00882 in HCC is correlated with tumor recurrence
The HCC patients were divided into two groups according to relative mRNA levels of LINC00882 (Table 3). By incorporating the LINC00882 expression data with the clinicopathological parameters of the corresponding HCC patients, we performed single-factor statistical analysis to explore the correlation of LINC00882 expression level with various pathological factors in HCC, including sex, tumor numbers, clinical stage, recurrence, cirrhosis, age, tumor size and HBV infection. While expression level of LINC00882 was not significantly correlated with sex, tumor numbers, clinical stage, age, tumor size or HBV infection, high LINC00882 expression was found to be positively correlated with recurrence status (p = 0.04) and cirrhosis (p = 0.043) of HCC patients (Table 4), which indicated that LINC00882 could stimulate biological processes involved in HCC progression, such as angiogenesis and metastasis.
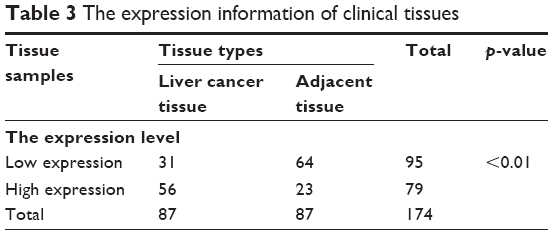 | Table 3 The expression information of clinical tissues |
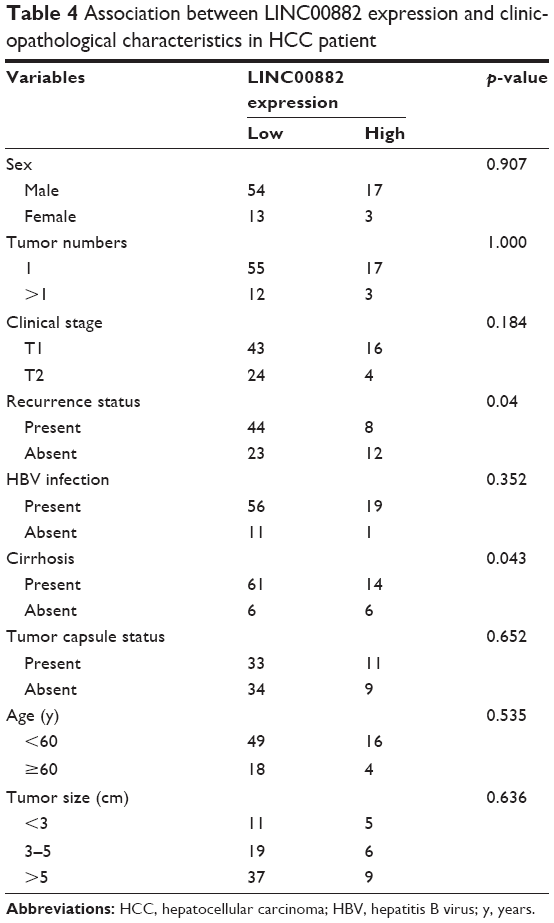 | Table 4 Association between LINC00882 expression and clinicopathological characteristics in HCC patient |
Upregulation of LINC00882 predicts poor prognosis of HCC patients
Further analysis of the correlation between the LINC00882 expression and clinical outcomes of HCC showed that the median OS time was 29 and 48 months in HCC patients with high and low expression of LINC00882, respectively (Tables 5 and 6, p = 0.043). High expression of LINC00882 predicted poor OS of HCC patients, which indicated that LINC00882 could be a potential prognosis biomarker for HCC (Figure 1C). Besides, COX regression analysis showed that the level of LincRNA00881 expression, recurrence and tumor size (<3 cm/>5 cm) were the risk factors affecting the prognosis of liver cancer patients (Table 7, p < 0.05).
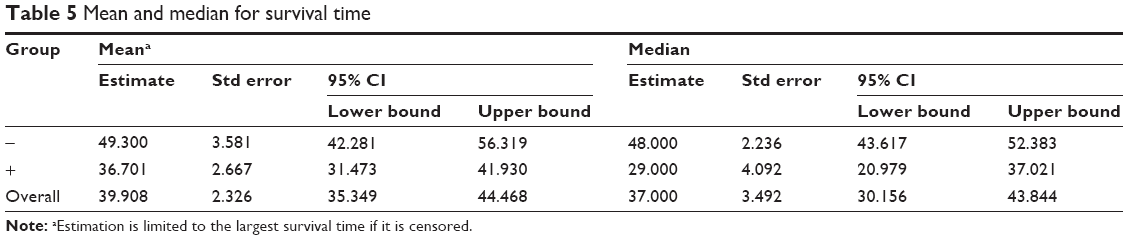 | Table 5 Mean and median for survival time |
 | Table 6 Overall comparisons |
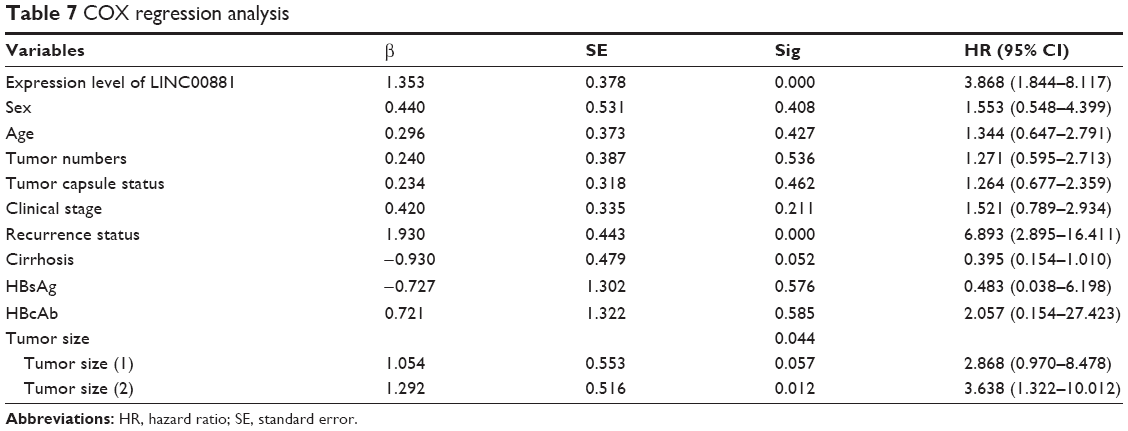 | Table 7 COX regression analysis |
LINC00882 knockdown reduces abilities of proliferation, migration and invasion of HCC cells
To investigate the role of LINC00882 in HCC carcinogenesis, the endogenous mRNA levels of LINC00882 were detected in 3 HCC cell lines and normal hepatocytes L02. It was found that LINC00882 was upregulated in 3 HCC cells compared with L02 (Figure 2A). Then, LINC00882 was knocked down by infecting HCC cell lines with lentivirus shRNAs targeting LINC00882. Silencing of LINC00882 in the cell lines was confirmed by qRT-PCR (Figure 2B). It was worth mentioning that knockdown of LINC00882 resulted in expression suppression of growth markers and matrix metalloproteinases (MMPs) 2 and 9 at both mRNA and protein levels, which suggested that LINC00882 could play important roles in proliferation and metastasis of HCC cells (Figure 2C and D).14,15
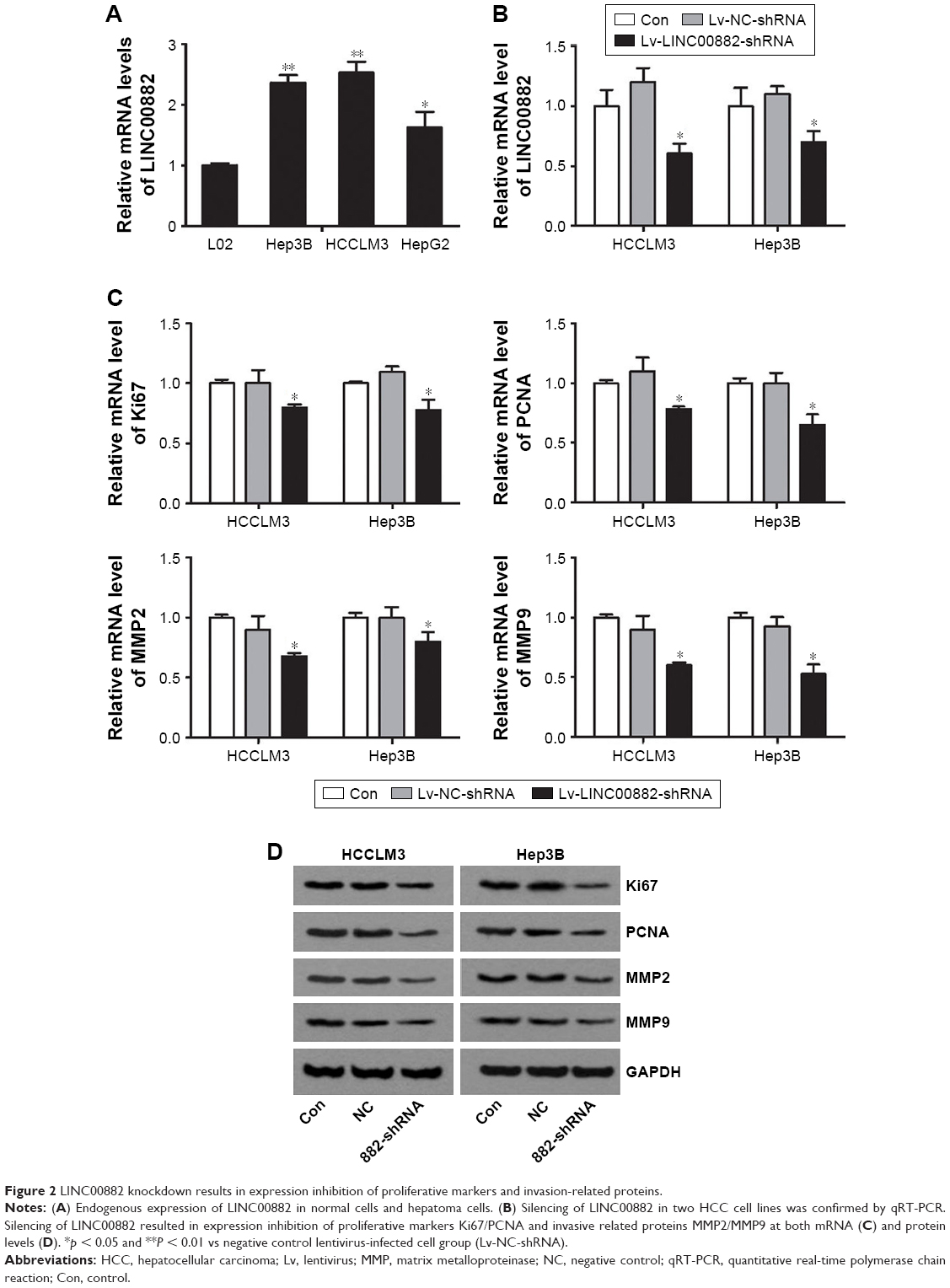 | Figure 2 LINC00882 knockdown results in expression inhibition of proliferative markers and invasion-related proteins. |
In order to confirm the regulation of LINC00882 on HCC proliferation and metastasis from the cellular functional level, EdU, migration and invasion assays were performed. LINC00882-silenced HCC cells exhibited decreased EdU staining, which suggested decrease in cell proliferation rate (Figure 3A and C). An altered transwell assay was used to assess the invasion ability of HCC cells in which chambers were pre-coated with Matrigel to mimic tissue barriers in vivo. Knockdown of LINC00882 resulted in remarkable decrease of number of HCC cells passing through the membrane (Figure 3B and C), which indicated damaged invasion ability of LINC00882-silenced HCC cells. The wound healing assay indicated that the migration ability of HCC cells was suppressed by LINC00882 silencing (Figure 3D). In conclusion, knockdown of LINC00882 led to suppression of proliferation, migration and invasion of HCC cells.
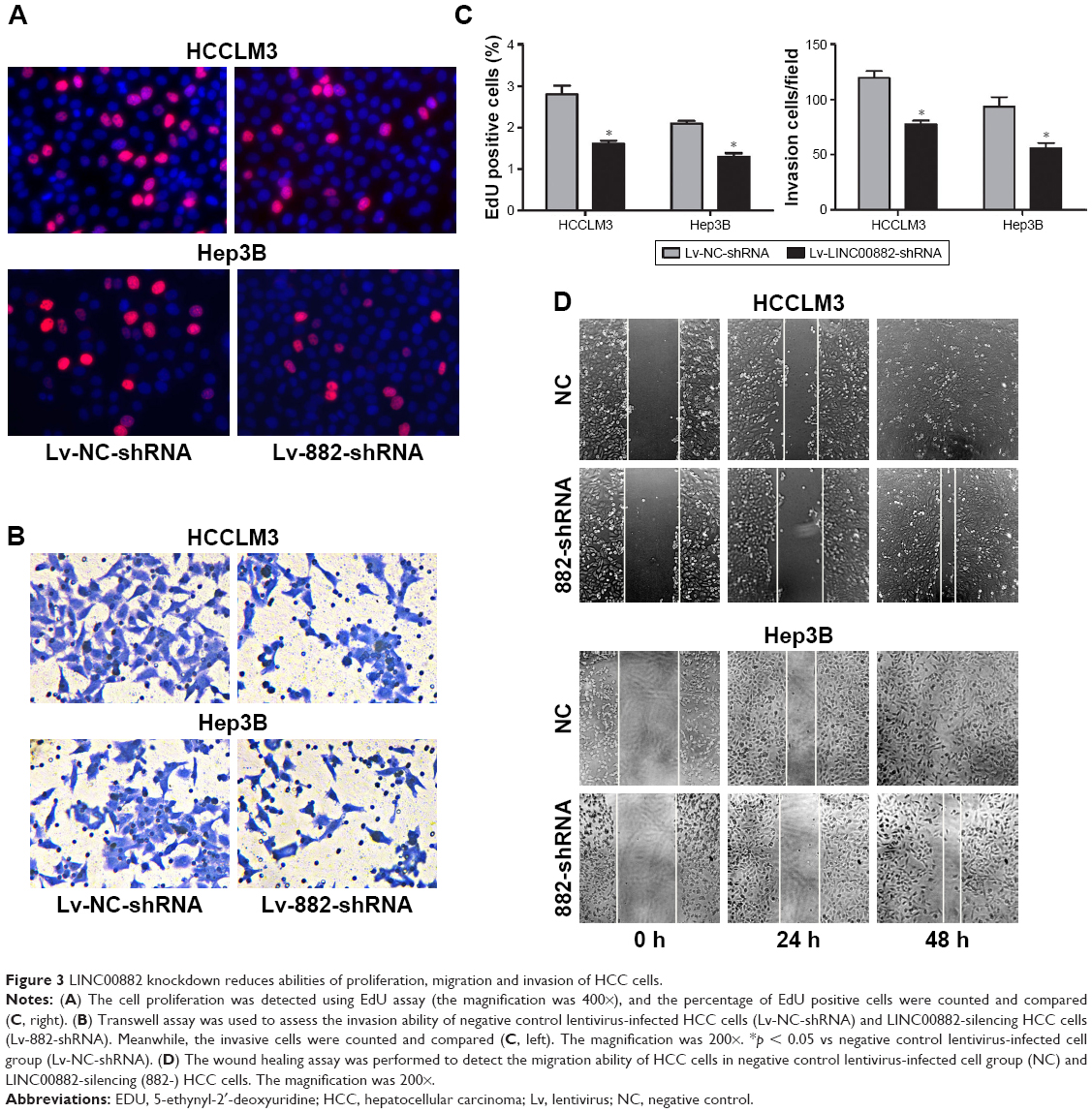 | Figure 3 LINC00882 knockdown reduces abilities of proliferation, migration and invasion of HCC cells. |
Discussion
In the recent years, accumulating studies have indicated that lncRNAs, including HULC, lncRNA-hPVT1, UFC1, H19, HOTAIR and MVIH, are dysregulated in HCC and play crucial roles in biological processes involved in HCC progression. These lncRNAs affect proliferation, migration and invasion of HCC cells by regulating variety of cellular processes, including cell cycling, epithelial-to-mesenchymal transition, angiogenesis and metastasis. In the present study, we identified differentially expressed lncRNAs in HCC by mining two GEO datasets. We found that 10 lncRNAs were significantly differentially expressed in HCC tissues as compared with noncancerous liver tissues. Most of these dysregulated lncRNAs are not functionally characterized. More efforts are warranted to explore the roles and molecular mechanisms of these lncRNAs in HCC carcinogenesis.
As one of the most profoundly dysregulated lncRNAs in both GEO datasets, involvement of LINC00882 in HCC was not previously reported. Therefore, we chose, in this study, to further elucidate the roles of LINC00882 in HCC carcinogenesis. We first validated upregulation of LINC00882 in HCC by analyzing two separate cohorts containing 85 HCC tissues and paired adjacent noncancerous tissues and 86 HCC tissues and 89 independent noncancerous liver tissues, respectively. Moreover, we found that high expression of LINC00882 was associated with tumor recurrence and poor prognosis of HCC patients, which indicated that LINC00882 may promote HCC carcinogenesis. Functions of LINC00882 in HCC carcinogenesis were then investigated by in vitro functional experiments. Indeed, in vitro cell assays, including cell proliferation assay, wound healing assay and transwell assay, demonstrated that knockdown of LINC00882 distinctly suppressed the proliferation, migration and invasion abilities of HCC cells, suggesting that LINC00882 is critical to tumor growth and metastasis of HCC. In consistency with these results, expressions of proliferation markers Ki-61/PCNA and invasion markers MMP2/MMP9 were suppressed by knockdown of LINC00882, which supported the conclusion that LINC00882 promotes HCC progression.
LncRNAs compete for miRNAs with specific mRNAs, which share common micRNA response elements and therefore regulate expression of these mRNAs. In a future study, we will focus on elucidation of the molecular mechanisms of LINC00882 and biological processes regulated by this lncRNA in HCC carcinogenesis. Bioinformatical tools will be employed to explore potential mRNAs and miRNAs, which are regulated by LINC00882. Briefly, we first plan to identify candidate mRNAs regulated by LINC00882 by evaluating the correlation of LINC00882 mRNA levels in HCC tissues and adjacent tissues. We will then perform GO and KEGG pathway analysis for the coding genes corresponding to these mRNAs to explore the functional categories and signaling pathways involved in HCC carcinogenesis. Potential miRNAs interacting with LINC00882 can be predicted by starBase v2.0 and miRcode,16,17 and experimentally validated mRNAs interacting with these candidate miRNAs will be obtained from miRTarBase.18 Maximal information coefficient algorithms are used to identify the robustness of pair-wise relationships of miRNA–lncRNA and miRNA–mRNA.19 Last, we will verify regulation of these bioinformatically predicted target miRNAs and mRNAs by LINC00882 and their roles in HCC carcinogenesis by in vitro and in vivo functional experiments.
In conclusion, our findings suggested that LINC00882 may be crucial for HCC cancerogenesis by promoting proliferation, migration and invasion of HCC cells, and high expression of LINC00882 predicts poor OS of HCC patients. More efforts are warranted to unravel molecular mechanisms of LINC00882 and biological processes regulated by this lncRNA in HCC carcinogenesis and to explore the potential of LINC00882 serving as prognostic biomarker and therapeutic target for HCC in the future.
Conclusion
These data indicated that LINC00882 promotes HCC progression and could be a potential prognostic biomarker and therapeutic target for HCC.
Acknowledgments
This work was supported by the Fundamental Research Funds for Central Universities of Central South University (grant no. 2016zzts160), the New Xiangya Talent Project of the Third Xiangya Hospital of Central South University (JY201519) and Provincial Natural Science Foundation of Hunan (2017JJ3419) to Hongbo Xu.
Disclosure
The authors report no conflicts of interest in this work.
References
Aravalli RN, Cressman EN, Steer CJ. Cellular and molecular mechanisms of hepatocellular carcinoma: an update. Arch Toxicol. 2013;87(2):227–247. | ||
Gomaa AI, Khan SA, Toledano MB, Waked I, Taylor-Robinson SD. Hepatocellular carcinoma: epidemiology, risk factors and pathogenesis. World J Gastroenterol. 2008;14(27):4300–4308. | ||
Song P, Feng X, Zhang K, et al. Screening for and surveillance of high-risk patients with HBV-related chronic liver disease: promoting the early detection of hepatocellular carcinoma in China. Biosci Trends. 2013;7(1):1–6. | ||
Cheetham SW, Gruhl F, Mattick JS, Dinger ME. Long noncoding RNAs and the genetics of cancer. Br J Cancer. 2013;108(12):2419–2425. | ||
Li G, Zhang H, Wan X, et al. Long noncoding RNA plays a key role in metastasis and prognosis of hepatocellular carcinoma. Biomed Res Int. 2014;2014:780521. | ||
Tripathi V, Shen Z, Chakraborty A, et al. Long noncoding RNA MALAT1 controls cell cycle progression by regulating the expression of oncogenic transcription factor B-MYB. PLoS Genet. 2013;9(3):e1003368. | ||
Zhang F, Zhang L, Zhang C. Long noncoding RNAs and tumorigenesis: genetic associations, molecular mechanisms, and therapeutic strategies. Tumour Biol. 2016;37(1):163–175. | ||
Dinger ME, Amaral PP, Mercer TR, et al. Long noncoding RNAs in mouse embryonic stem cell pluripotency and differentiation. Genome Res. 2008;18(9):1433–1445. | ||
Wang KC, Chang HY. Molecular mechanisms of long noncoding RNAs. Mol Cell. 2011;43(6):904–914. | ||
Liu YR, Tang RX, Huang WT, et al. Long noncoding RNAs in hepatocellular carcinoma: Novel insights into their mechanism. World J Hepatol. 2015;7(28):2781–2791. | ||
He S, Zhang J, Zhang W, Chen F, Luo R. FOXA1 inhibits hepatocellular carcinoma progression by suppressing PIK3R1 expression in male patients. J Exp Clin Cancer Res. 2017;36(1):175. | ||
He HT, Xu M, Kuang Y, Han XY, Wang MQ, Yang Q. Biomarker and competing endogenous RNA potential of tumor-specific long noncoding RNA in chromophobe renal cell carcinoma. Onco Targets Therapy. 2016;9:6399–6406. | ||
Perry MM, Tsitsiou E, Austin PJ, et al. Role of non-coding RNAs in maintaining primary airway smooth muscle cells. Respir Res. 2014;15:58. | ||
Stroescu C, Dragnea A, Ivanov B, et al. Expression of p53, Bcl-2, VEGF, Ki67 and PCNA and prognostic significance in hepatocellular carcinoma. J Gastrointestin Liver Dis. 2008;17(4):411–417. | ||
Verma RP, Hansch C. Matrix metalloproteinases (MMPs): chemical-biological functions and (Q)SARs. Bioorg Med Chem. 2007;15(6):2223–2268. | ||
Jeggari A, Marks DS, Larsson E. miRcode: a map of putative microRNA target sites in the long non-coding transcriptome. Bioinformatics. 2012;28(15):2062–2063. | ||
Li JH, Liu S, Zhou H, Qu LH, Yang JH. starBase v2.0: decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA interaction networks from large-scale CLIP-Seq data. Nucleic Acids Res. 2014;42(Database issue):D92–D97. | ||
Hsu SD, Tseng YT, Shrestha S, et al. miRTarBase update 2014: an information resource for experimentally validated miRNA-target interactions. Nucleic Acids Res. 2014;42(Database issue):D78–D85. | ||
Reshef DN, Reshef YA, Finucane HK, et al. Detecting novel associations in large data sets. Science. 2011;334(6062):1518–1524. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.