Back to Journals » Infection and Drug Resistance » Volume 14
Occurrence of mcr Positive Strains and Molecular Characteristics of Two mcr-1 Positive Salmonella typhimurium and Escherichia coli from a Chinese Women’s and Children’s Hospital
Authors Zheng Z ![]() , Lei Y, Wang Y, Lin C, Lin J
, Lei Y, Wang Y, Lin C, Lin J ![]()
Received 2 June 2021
Accepted for publication 22 July 2021
Published 27 July 2021 Volume 2021:14 Pages 2925—2932
DOI https://doi.org/10.2147/IDR.S322686
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Suresh Antony
Zhenzhu Zheng, Ying Lei, Yinna Wang, Chunli Lin, Jiansheng Lin
Department of Laboratory Medicine, Quanzhou Women’s and Children’s Hospital, Quanzhou, People’s Republic of China
Correspondence: Jiansheng Lin
Department of Laboratory Medicine, Quanzhou Women’s and Children’s Hospital, 700 Fengze Street, Fengze District, Quanzhou, Fujian Province, 350122, People’s Republic of China
Tel +86 0595 22196131
Email [email protected]
Background: The purpose of this study was to evaluate the prevalence of mobile colistin resistance genes (mcr) in Gram-negative bacteria and to analyze the molecular characteristics of mcr-1 positive Salmonella typhimurium strain 75 and Escherichia coli strain 107 from the Quanzhou Women’s and Children’s Hospital in China.
Methods: The genes mcr-1 through mcr-9 were screened via multiplex PCR. Antibiotic susceptibility was detected using a GN11 card with the VITEK-2 compact automated system. Whole genomes were sequenced using PacBio’s single molecule real-time (SMRT) technology.
Results: In this study, mcr-1 was detected in only four strains, with a positivity rate of 0.65% (4/616). All the four strains were resistant to more than three different kinds of antibiotics. The mcr-1 positive S. typhimurium strain 75 harbored IncHI2 plasmid, which carried mcr-1 gene, while the mcr-1 positive E. coli strain 107 contained four plasmids including one mcr-1 harboring IncHI2 plasmid, one IncFII plasmid and two IncI1-I (Alpha) plasmids. Mobile elements carrying mcr-1 in the 75_plasmid and 107_plasmid-1 were located in the IS1086(ISApl1)-IS30A(ISApl1)-mcr-1-hp and IS1086(ISApl1)-mcr-1-hp regions, respectively. Tn6010 carrying drug efflux pump genes was found in 75_plasmid, while cn_31611_IS26 carrying multi-drug resistance (MDR) genes were found in 107_plasmid-1.
Conclusion: This study found that mcr-1 was prevalent at a low frequency in the Quanzhou Women’s and Children’s Hospital. A similar genetic pattern of mcr-1 transmission was found in both E. coli and S. typhimurium.
Keywords: mcr-1, Salmonella typhimurium, Escherichia coli, molecular characteristics
Introduction
Polymyxin is highly active against most gram-negative bacteria. However, its nephrotoxicity and neurotoxicity strongly obstruct further application in the treatment of typical patients.1 The rapid increase of multi-drug resistant (MDR) Gram-negative and the lack of new antibiotics have led to a revival of the clinical use of polymyxin, which is recognized as a last-resort antibiotic for numerous MDR bacterial infections.2,3 Intrinsic resistance to polymyxin originates from the functional expression or mutation of chromosomal genes in Neisseria meningitidis or Pseudomonas aeruginosa.4,5
Since the appearance of the first plasmid-mediated mobile colistin resistance gene (mcr-1) in Enterobacteriaceae,6 this transmissible gene has been found worldwide in various gram-negative bacteria, including E. coli, Klebsiella pneumoniae, Klebsiella oxytoca, Salmonella enterica, and Enterobacter species from animal, meat, food product, environmental, and human sources.7,8 More worrisome is the presence of mcr-1 in Enterobacteriaceae with carbapenem resistance genes, such as blaNDM and blaKPC, which could seriously compromise the treatment of infections caused by these extremely drug-resistant pathogens.9 Salmonella enterica serovar Typhimurium belonged to one of the most important serotypes of Salmonella enterica.10 The multidrug-resistant Salmonella enterica serovar Typhimurium isolate has a high potential to disseminate the mcr-1 gene and further challenge clinical treatment.11,12 Meanwhile, eight new mcr homologues (mcr-2 to mcr-9) have been identified in Enterobacteriaceae.13 E. coli was identified as containing both mcr-1 and mcr-3.19 on a hybrid plasmid, which suggests that the evolution of mcr genes among various plasmids is being driven by mobile elements.14 The phosphoethanolamine (PEA) transferase encoded by mcr can catalyse the attachment of PEA to lipopolysaccharide (LPS)-Lipid A, resulting in resistance to polymyxin.15 Polymyxin resistance has attracted clinical and public health attention.16 The purpose of this study was to evaluate the prevalence of mcr genes in gram-negative bacteria in the Quanzhou Women’s and Children’s Hospital and to analyse the molecular characteristics of mcr-1-positive S. typhimurium strain 75 and E. coli strain 107.
Methods
Bacterial Strains
A total of 616 clinical gram-negative isolates were collected at the Quanzhou Women’s and Children’s Hospital from January 2018 to April 2019. The specimens were derived from 218 stool, 142 sputum, 102 throat swab, 68 blood, 35 urine, 35 pus, 10 alveolar lavage fluid, 2 cerebrospinal fluid, 2 ascites, and 2 endotracheal intubation tube segment samples. The study was conducted in accordance with the Declaration of Helsinki, and ethical permission for this study was approved by the Quanzhou Women’s and Children’s Hospital ethics committee (2017 Ethical Review No. 11). Guardians provided informed consent on behalf of the minors. Strains were identified using the GN card with the VITEK-2 compact automated system (bioMérieux).
DNA Extraction
The DNA extraction procedure was performed according to previously reported methods.17 After the strain was recovered overnight on LB agar plates, a single colony was picked and resuspended in 100 µL of double distilled water. The suspension was heated at 100°C for 10 min and centrifuged at 5000 rpm for 10 min. DNA was quantified using a UV spectrophotometer, and 1 µL of the supernatant was collected for PCR.
mcr Gene Detection
mcr genes (mcr-1–9) were screened by multiplex PCR using primers as previously described.10 Multiplex PCR reagents were purchased from Nanjing Vazyme Biotechnology Co., Ltd. PCR was conducted by an ABI 7500 instrument, and PCR products were visualised by observing their position after DNA electrophoresis. The PCR products were sent to Fuzhou Boshang Biological Co., Ltd for Sanger sequencing and the sequences were aligned by NCBI BLAST.
Antibiotic Susceptibility Testing
The minimum inhibitory concentration (MIC) of antibiotics was detected using a GN11 card using the VITEK-2 compact automated system (bioMérieux). Antibiotics tested included ampicillin (AMP), ampicillin/sulbactam (SAM), piperacillin/tazobactam (TZP), cefazolin (CZO), cefotetan (CTT), ceftazidime (CAZ), ceftriaxone (CRO), cefepime (FEP), aztreonam (ATM), ertapenem (ETP), imipenem (IMP), amikacin (AMK), gentamicin (KAN), tobramycin (TOB), ciprofloxacin (CIP), levofloxacin (LEV), nitrofurantoin (NIT), and trimethoprim/sulfamethoxazole (SXT). Interpretation of the results was based on the guidelines published by the Clinical and Laboratory Standards Institute (CLSI).18 The MIC of colistin (COL) purchased from Shanghai Jizhi Biochemical Technology Co., Ltd. was determined using a broth microdilution method. Briefly, the COL diluted in multiples was added to a sterilized 96-well polystyrene plate, 100 µL bacterial suspension diluted 1:1000 with MH broth was added into each well and OD600 was determined after it was incubated at 35°C for 20h. ≥4µg/mL was interpreted as resistance in according to the European Committee on Antimicrobial Susceptibility Testing (EUCAST) breakpoint standard.19 E. coli ATCC8739 and S. typhimurium ATCC14028 were selected as quality control strains.
Whole Genome Sequencing (WGS)
S. typhimurium strain 75 and E. coli strain 107 (MIC of COL ≥8 µg/mL) were selected for whole genome sequencing. DNA extraction was carried out according to the instructions of the bacterial genomic DNA extraction kit (TIANGEN Biochemical Technology Co., Ltd). Genomes were sequenced using PacBio’s SMRT third-generation sequencing technology, using the HGAP 4 process from PacBio’s official analysis software SMRT Link (V6.0.0.47841) for de novo genome assembly. Library construction and sequencing was performed by Tianjin Biochip Bio Co., Ltd. Multi-locus sequence typing (MLST), plasmid type, plasmid MLST, antibiotic resistance genes/chromosomal point mutations, and mobile genetic elements were analysed using MLST 2.0, PlasmidFinder 2.1, pMLST 2.0, ResFinder 4.1, and MobileElementFinder, respectively.20–23 Circular imaging and comparisons between multiple plasmids were conducted using the BLAST ring image generator (BRIG).24
Nucleotide Sequence
The complete sequences of 75_chromosome, 75_plasmid, 107_chromosome, 107_plasmid-1, 107_plasmid-2, 107_plasmid-3, 107_plasmid-4 were deposited in GenBank under the accession numbers CP075372, CP075373, CP075374, CP075375, CP075376, CP075377, CP075378, respectively, which will be made public on May 24, 2021.
Results
mcr Gene Screening and Drug Susceptibility results of mcr-Positive Strains
The mcr-1 gene was only detected in four strains, with a positivity rate of 0.65% (4/616). Two S. typhimurium strains and one E. coli strain were found in the gastrointestinal tract of three different patients, and one E. coli strain was found in the urine of a single patient. The age of the patients ranged between 1.3 and 2.7 years. All four patients recovered after treatment. Additional clinical data of the four patients with mcr-1-positive S. typhimurium or E. coli infections are shown in Table 1. Drug susceptibility results indicated that the S. typhimurium strain 74, S. typhimurium strain 75, E. coli strain 107, and E. coli strain 332 were multi-resistant, and their profiles appear in Table 2.
 |
Table 1 Clinical Data of Four Patients Isolated E. coli or S. typhimurium with mcr-1 Positive |
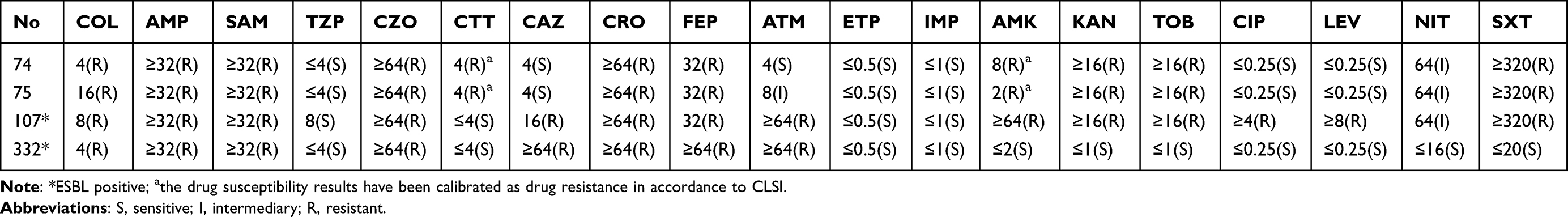 |
Table 2 MIC of mcr-1 Positive S. typhimurium and E. coli Strains |
Genetic Characteristics of Strains and Plasmids Bearing mcr-1
The genome of the S. typhimurium strain 75, which belongs to the ST34 strain type, consisted of a 4.88 Mb chromosome which contained no mutations of the acrB, parC and 16S_rrsD genes. It also contained a circular 0.24 Mb mcr-1-positive 75_plasmid encoding the IncHI2 replication protein as well as drug resistance genes such as oqxA, oqxB, FosA3, mcr-1, aac(3)-IV, aadA8b, aadA2, aadA1, sul2, sul3, sul1, dfrA12, cmlA1, floR, and blaCTX-M-14 (Table 3). The genome of the E. coli strain 107 belonged to an unknown strain type and consisted of a 4.88 Mb chromosome with mutations such as parC:p.S80I, parE:p.S458A, gyrA:p.S83L, and gyrA:p.D87N. It also contained four plasmids, identified here as 107_plasmid 1–4. 107_plasmid-1 is 0.27 Mb, circular, mcr-1 positive, and encodes the IncHI2 replication protein, as well as the drug resistance genes dfrA27, sul2, sul1, aph(3ʹ)-Ia, aph(4)-Ia, aadA16, aac(3)-IV, fosA3, mph(A), ARR-3, floR, blaCTX-M-14, and qacE. 107_plasmid-2 is an IncFII type plasmid and carries the drug resistance genes rmtB, blaTEM-214, blaTEM-141, blaCTX-M-55, blaTEM-206, and blaTEM-1B. Both 107_plasmid-3 and −4 encode the IncI1-I(Alpha) replication protein (Table 3).
 |
Table 3 Phenotypic and Molecular Features of mcr-1 Positive S. typhimurium Strain 75 and E. coli Strain 107 |
A BLAST search indicated that the 75_plasmid showed an overall query coverage (97–99%) and nucleotide similarity (99.98%) to several plasmids, such as pGDP37-4 (GenBank no. MK673548.1), pS438 (CP061125.1), and pSH16G0648 (MH522418.1). In addition, the size and backbone structure of these plasmids were quite similar (Figure 1A). The 107_plasmid-1 showed an overall query coverage (96%) and nucleotide similarity (99.95–99.99%) to several plasmids, such as pWF5-29 (GenBank no. MG385063.1), pXGE1mcr (KY990887.1), and pLD22-1-MCR1 (CP047877.1). The size and backbone structure of these plasmids were also quite similar (Figure 1B). Further research shows that a 102,172–103,380 region sequence structure encoding hypothetical protein (hp) was inserted in 75_plasmid, and a 251,336–253,752 region sequence structure encoding hp, y4hO, and y4hP was inserted 107_plasmid-1 (Figure 1). Mobile elements carrying mcr-1 sequences of the IS1086(ISApl1)-IS30A(ISApl1)-mcr-1-hp and IS1086(ISApl1)-mcr-1-hp regions were identified in the 75_plasmid and 107_plasmid-1, respectively (Figure 1). MobileElementFinder analysis showed that the 72,498–79,343 region of the 75_plasmid belonged to a composite transposon of Tn6010, while the 251,336–253,752 and 77,044–108,655 regions of the 107_plasmid-1 belonged to the IS682 and cn_31611_IS26 regions, respectively.
 |
Figure 1 Circular representation of the studied plasmids. GC content and GC Skew were represented on the distance scale (in kbp) on the inner map. Each plasmid was compared to its most closely-related plasmid. The red arc around the map indicated ORFs. Certain important genes were also indicated on the ring. The hp is short for hypothetical protein. (A) 75_Plasmid alignment map; (B) 107_Plasmid-1 alignment map. |
Discussions
In this study, the mcr-1 gene was detected in only four strains, with a positivity rate of 0.65% (4/616), which was similar to the 0.62% rate reported by Rong Fan et al.25 This indicated that there was no detection of any mcr types other than mcr-1 in Quanzhou. CRO was used to treat diarrhoea infection in patient #107 but bacterial resistance may have led to treatment failure, resulting in prolonged hospital stay (10 days).
mcr-1 was found to frequently coexist with other drug resistance genes (ie genes responsible for resistance to carbapenems26,27 and extended-spectrum β-lactam drugs),28,29 indicating the possibility of the emergence of bacteria with MDR.30 Drug susceptibility results of this study indicated that all the four strains were resistant to more than three different groups of drugs. All mcr-1-positive strains were resistant to COL. It was previously reported that mcr-1 appeared in carbapenem-resistant Enterobacteriaceae.31 Fortunately, we found that the four strains were sensitive to ETP and IMP. Further analysis showed that the S. typhimurium strain 75 and E. coli strain 107 were susceptible to carbapenem, and this was consistent with the absence of carbapenem-resistant genes in both the strains.
The S. typhimurium monophasic pandemic sequence ST34 clone has attracted global attention because of its rapidly increasing resistance to MDR. This clone was associated with resistant-pattern ASSuT (AMP, streptomycin, sulfamethoxazole, and tetracycline).32 The ST34 phenotype-associated drug resistance genes blaTEM-1B, aph(3”)-Ib, aph(6)-Id, sul2, and tet(B) were also found in S. typhimurium strain 75. It may be that unknown mutations in acrB, parC, and 16S_rrsD do not confer quinolone resistance, which was consistent with the sensitivity of this strain to CIP and LEV. The detection of ST34 strains carrying the mcr-1 gene in diarrhoea suggests that the Chinese children were suffering from the disease caused by this pathogen.33,34 The presence of the COL resistance genes (mcr-1, mcr-3, and mcr-5) combined with the MDR phenotype in ST34 had spread across different countries, with most mcr-1 genes from the ST34 isolates detected in the plasmid type IncHI2, followed by IncI2 and IncX4.35 The mcr-1-positive 75_plasmid was circular and encodes the IncHI2 replication protein. Several genes encoding components involved in antitoxin systems as well as colicin, tellurite, and heavy metal tolerance likely play critical roles in the environmental stability of IncHI2 plasmids in the ST34 clone.36 Therefore, the ST34 strain is more likely to acquire mcr-bearing plasmids. All strains harbouring the mcr-1 gene were reported to carry the MDR plasmid. The 75_plasmid also contained drug resistance genes, which mediated the emergence of MDR. Here, we reported for the first time that mcr-1 and aadA8b coexist in the IncHI2 plasmid of the ST34 strain. This finding was important because this genetic combination may increase the spread risk of aminoglycoside resistance.
The most common mcr-1-positive E. coli strain ST10 has been frequently detected in China.37 The genetic makeup of mcr-1-positive E. coli was highly diverse.38 Therefore, it was unsurprising that we found an unknown sequence type in the E. coli strain 107. The 107_chromosome of E. coli strain 107 in this study contained mutation types such as parC:p.S80I, parE:p.S458A, gyrA:p.S83L, gyrA:p.D87N, which confered quinolone resistance.39 In 90% of E. coli cases, the mcr-1 gene was related to IncX4, IncHI2, and IncI2 mobilizable plasmids, which spread easily worldwide.40–42 The mcr-1-positive 107_plasmid-1 was circular and encoded the IncHI2 replication protein. Two mcr-1–bearing plasmids (IncI2 and IncHI2) were observed in the same strain.43 The 107_plasmid-1 and 2 both carried numerous drug resistance genes, which may increase the risk of the development of pan-drug resistance.
BLASTp analysis showed that the hp inserted in the 75_plasmid was an IS4-like element ISVsa5 family transposase reported to inactivate the mcr-1 gene by insertion in S. typhimurium;44 The hp, y4hO, and y4hP that were inserted into the 107_plasmid-1 belonged respectively to IS66 transposase TnpA, TnpB, and IS66-like element ISEc23 family transposase, which were present in the other mcr-1-positive plasmids. MobileElementFinder analysis showed that the 72,498–79,343 region belonged to a composite transposon of Tn6010, including the efflux pump genes oqxA and oqxB. These genes were similar to those of the oqxAB-carrying plasmids pHXY0908 (from chickens) and pHK0653 (from a human patient) in S. typhimurium.45 Four mobile element structures carrying mcr-1 in Enterobacteriaceae have been reported: (1) with both ends containing the ISApl1 transposon Tn6330 (ISApl1–mcr-1–orf–ISApl1 structure); (2) with a single upstream ISApl1 (ISApl1–mcr-1–orf structure); (3) with a single downstream ISApl1 (mcr-1–orf–ISApl1 structure); (4) with sequences lacking ISApl1 altogether (mcr-1–orf structure).46,47 Of note, the sequence region of IS1086(ISApl1)-IS30A(ISApl1)-mcr-1-hp in 75_plasmid and the sequence region of IS1086(ISApl1)-mcr-1-hp in 107_plasmid-1 both belonged to upstream ISApl1 element, which was observed in 78% of IncHI2 plasmids in E. coli.40 Cn_31611_IS26 (IS6 family) included 11 drug resistance genes (dfrA27, sul2, sul1, aph(4)-Ia, aadA16, aac(3)-IV, mcr-1, mph(A), ARR-3, floR, and qacE), which may be acquired from a Tn region containing several drug resistance genes.
In conclusion, this study revealed that the mcr-1 gene had low prevalence in the Quanzhou Women’s and Children’s Hospital. The same genetic strategy for mcr-1 transmission was found in both E. coli and S. typhimurium. mcr-1 transmission should attract the attention of the public health sector, which should adopt urgent methods such as strictly controlling the large-scale use of polymyxin in agriculture and animal husbandry to control its spread.
Acknowledgments
We thank the patients and colleagues in our department.
Funding
This research was partially funded by Quanzhou Science and Technology Project (no:2018Z159).
Disclosure
The authors declare that they have no conflict of interest.
References
1. Fekety FR. The treatment of gram-negative bacillary infections with colistin. The toxicity and efficacy of large doses in forty-eight patients. Ann Intern Med. 1962;57:214. doi:10.7326/0003-4819-57-2-214
2. Li J, Nation RL, Turnidge JD. Colistin: the re-emerging antibiotic for multidrug-resistant gram-negative bacterial infections. Lancet Infect Dis. 2006;6(9):589–601. doi:10.1016/S1473-3099(06)70580-1
3. Biswas S, Brunel J-M, Dubus J-C, Reynaud-Gaubert M, Rolain J-M. Colistin: an update on the antibiotic of the 21st century. Expert Rev Anti Infect Ther. 2012;10(8):917–934. doi:10.1586/eri.12.78
4. Wanty C, Anandan A, Piek, S, et al.The structure of the neisserial lipooligosaccharide phosphoethanolamine transferase A (LptA) required for resistance to polymyxin. J Mol Biol. 2013;425(18):3389–3402. doi:10.1016/j.jmb.2013.06.029
5. McPhee JB, Lewenza S, Hancock REW. Cationic antimicrobial peptides activate a two-component regulatory system, PmrA-PmrB, that regulates resistance to polymyxin B and cationic antimicrobial peptides in Pseudomonas aeruginosa. Mol Microbiol. 2003;50(1):205–217. doi:10.1046/j.1365-2958.2003.03673.x
6. Liu -Y-Y, Wang Y, Walsh TR. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: a microbiological and molecular biological study. Lancet Infect Dis. 2016;16(2):161–168. doi:10.1016/S1473-3099(15)00424-7
7. Anyanwu MU, Jaja IF, Nwobi OC. Occurrence and characteristics of Mobile Colistin Resistance (mcr) gene-containing isolates from the environment: a review. Int J Environ Res Public Health. 2020;17(3):1028. doi:10.3390/ijerph17031028
8. Nang SC, Li J, Velkov T. The rise and spread of mcr plasmid-mediated polymyxin resistance. Crit Rev Microbiol. 2019;45(2):131–161. doi:10.1080/1040841X.2018.1492902
9. Wang Y, Tian G-B, Zhang R. Prevalence, risk factors, outcomes, and molecular epidemiology of mcr-1-positive Enterobacteriaceae in patients and healthy adults from China: an epidemiological and clinical study. Lancet Infect Dis. 2017;17(4):390–399. doi:10.1016/S1473-3099(16)30527-8
10. Lamas A, Miranda JM, Regal P, Vázquez B, Franco CM, Cepeda A. A comprehensive review of non-enterica subspecies of Salmonella enterica. Microbiol Res. 2018;206:60–73. doi:10.1016/j.micres.2017.09.010
11. Li Y, Zhang Y, Chen M. Plasmid-borne colistin resistance gene mcr-1 in a multidrug resistant Salmonella enterica serovar Typhimurium isolate from an infant with acute diarrhea in China. Int J Infect Dis. 2021;103:13–18. doi:10.1016/j.ijid.2020.11.150
12. Wang Z, Xu H, Tang Y, Li Q, Jiao X. A multidrug-resistant monophasic Salmonella typhimurium co-harboring mcr-1, fosA3, bla CTX-M-14 in a transferable IncHI2 plasmid from a healthy catering worker in China. Infect Drug Resist. 2020;13:3569–3574. doi:10.2147/IDR.S272272
13. El-Sayed Ahmed MAE-G, Zhong -L-L, Shen C, Yang Y, Doi Y, Tian G-B. Colistin and its role in the era of antibiotic resistance: an extended review (2000–2019). Emerg Microbes Infect. 2020;9(1):868–885. doi:10.1080/22221751.2020.1754133
14. Li R, Zhang P, Yang X. Identification of a novel hybrid plasmid coproducing MCR-1 and MCR-3 variant from an Escherichia coli strain. J Antimicrob Chemother. 2019;74(6):1517–1520. doi:10.1093/jac/dkz058
15. Sun J, Zhang H, Liu Y-H, Feng Y. Towards understanding MCR-like colistin resistance. Trends Microbiol. 2018;26(9):794–808. doi:10.1016/j.tim.2018.02.006
16. Srinivas P, Rivard K. Polymyxin resistance in gram-negative pathogens. Curr Infect Dis Rep. 2017;19(11):38. doi:10.1007/s11908-017-0596-3
17. He Q-W, Xu X-H, Lan F-J. Molecular characteristic of mcr-1 producing Escherichia coli in a Chinese university hospital. Ann Clin Microbiol Antimicrob. 2017;16(1):32. doi:10.1186/s12941-017-0207-z
18. CLSI. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically; Approved Standard.
19. EUCAST. Breakpoints tables for interpretation of MICs and zone diameters, version 6.0. 2016.
20. Zankari E, Hasman H, Cosentino S. Identification of acquired antimicrobial resistance genes. J Antimicrob Chemother. 2012;67(11):2640–2644. doi:10.1093/jac/dks261
21. Carattoli A, Zankari E, García-Fernández A. In silico detection and typing of plasmids using plasmidfinder and plasmid multilocus sequence typing. Antimicrob Agents Chemother. 2014;58(7):3895–3903. doi:10.1128/AAC.02412-14
22. Rodríguez I, Thomas K, Van Essen A. Chromosomal location of blaCTX-M genes in clinical isolates of Escherichia coli from Germany, The Netherlands and the UK. Int J Antimicrob Agents. 2014;43(6):553–557. doi:10.1016/j.ijantimicag.2014.02.019
23. Johansson MHK, Bortolaia V, Tansirichaiya S, Aarestrup FM, Roberts AP, Petersen TN. Detection of mobile genetic elements associated with antibiotic resistance in Salmonella enterica using a newly developed web tool: MobileElementFinder. J Antimicrob Chemother. 2021;76(1):101–109. doi:10.1093/jac/dkaa390
24. Alikhan N-F, Petty NK, Ben Zakour NL, Beatson SA. BLAST Ring Image Generator (BRIG): simple prokaryote genome comparisons. BMC Genom. 2011;12:402. doi:10.1186/1471-2164-12-402
25. Fan R, Li C, Duan R. Retrospective screening and analysis of mcr-1 and blaNDM in gram-negative bacteria in China, 2010–2019. Front Microbiol. 2020;11:121. doi:10.3389/fmicb.2020.00121
26. Poirel L, Kieffer N, Liassine N, Thanh D, Nordmann P. Plasmid-mediated carbapenem and colistin resistance in a clinical isolate of Escherichia coli. Lancet Infect Dis. 2016;16(3):281. doi:10.1016/S1473-3099(16)00006-2
27. Delgado-Blas JF, Ovejero CM, Abadia-Patiño L, Gonzalez-Zorn B. Coexistence of mcr-1 and bla NDM-1 in Escherichia coli from Venezuela. Antimicrob Agents Chemother. 2016;60(10):6356–6358. doi:10.1128/AAC.01319-16
28. Falgenhauer L, Waezsada S-E, Yao Y. Colistin resistance gene mcr-1 in extended-spectrum β-lactamase-producing and carbapenemase-producing gram-negative bacteria in Germany. Lancet Infect Dis. 2016;16(3):282–283. doi:10.1016/S1473-3099(16)00009-8
29. Liu G, Ali T, Gao J. Co-occurrence of plasmid-Mediated Colistin Resistance (mcr-1) and extended-spectrum β-lactamase encoding genes in Escherichia coli from bovine mastitic milk in China. Microb Drug Resist. 2020;26(6):685–696. doi:10.1089/mdr.2019.0333
30. Shafiq M, Huang J, Ur Rahman S. High incidence of multidrug-resistant Escherichia coli coharboring mcr-1 and bla CTX-M-15 recovered from pigs. Infect Drug Resist. 2019;12:2135–2149. doi:10.2147/IDR.S209473
31. Li Y, Sun QL, Shen Y, et al.Rapid increase in prevalence of Carbapenem-Resistant Enterobacteriaceae (CRE) and emergence of colistin resistance gene mcr-1 in CRE in a hospital in Henan, China. J Clin Microbiol. 2018;56(4):e01932–01917.
32. Antunes P, Mourao J, Pestana N, Peixe L. Leakage of emerging clinically relevant multidrug-resistant Salmonella clones from pig farms. J Antimicrob Chemother. 2011;66(9):2028–2032. doi:10.1093/jac/dkr228
33. Lu X, Zeng M, Xu J. Epidemiologic and genomic insights on mcr-1-harbouring Salmonella from diarrhoeal outpatients in Shanghai, China, 2006–2016. EBioMedicine. 2019;42:133–144. doi:10.1016/j.ebiom.2019.03.006
34. Lu J, Quan J, Zhao D, Wang Y, Yu Y, Zhu J. Prevalence and molecular characteristics of mcr-1 gene in Salmonella typhimurium in a tertiary hospital of Zhejiang Province</em>. Infect Drug Resist. 2018;12:105–110. doi:10.2147/IDR.S190269
35. Biswas S, Li Y, Elbediwi M, Yue M. Emergence and dissemination of mcr-carrying clinically relevant salmonella typhimurium monophasic clone ST34. Microorganisms. 2019;7(9):298. doi:10.3390/microorganisms7090298
36. Johnson TJ, Wannemeuhler YM, Scaccianoce JA, Johnson SJ, Nolan LK. Complete DNA sequence, comparative genomics, and prevalence of an IncHI2 plasmid occurring among extraintestinal pathogenic Escherichia coli isolates. Antimicrob Agents Chemother. 2006;50(11):3929–3933. doi:10.1128/AAC.00569-06
37. Shen Y, Wu Z, Wang Y, et al.Heterogeneous and flexible transmission of mcr-1 in hospital-associated Escherichia coli. mBio. 2018;9(4):e00943–00918.
38. Liu F, Zhang R, Yang Y, et al.Occurrence and molecular characteristics of mcr-1-positive Escherichia coli from healthy meat ducks in Shandong Province of China. Animals (Basel). 2020;10(8):1299.
39. Yang F, Zhang S, Shang X, Wang L, Li H, Wang X. Characteristics of quinolone-resistant Escherichia coli isolated from bovine mastitis in China. J Dairy Sci. 2018;101(7):6244–6252. doi:10.3168/jds.2017-14156
40. Matamoros S, Van Hattem JM, Arcilla MS, et al.Global phylogenetic analysis of Escherichia coli and plasmids carrying the mcr-1 gene indicates bacterial diversity but plasmid restriction. Sci Rep. 2017;7(1):15364. doi:10.1038/s41598-017-15539-7
41. Elbediwi M, Li Y, Paudyal N, et al.Global burden of colistin-resistant bacteria: mobilized colistin resistance genes study (1980–2018). Microorganisms. 2019;7(10):461. doi:10.3390/microorganisms7100461
42. Li R, Xie M, Zhang J, et al.Genetic characterization of mcr-1-bearing plasmids to depict molecular mechanisms underlying dissemination of the colistin resistance determinant. J Antimicrob Chemother. 2017;72(2):393–401. doi:10.1093/jac/dkw411
43. Wu R, Yi L-X, Yu L-F. Fitness advantage of mcr-1–bearing IncI2 and IncX4 plasmids in vitro. Front Microbiol. 2018;9:331. doi:10.3389/fmicb.2018.00331
44. Fan J, Zhang L, He J. Plasmid dynamics of mcr-1-positive Salmonella spp. in a general hospital in China. Front Microbiol. 2020;11:604710. doi:10.3389/fmicb.2020.604710
45. Wang J, Zhi C-P, Chen X-J. Characterization of oqxAB in Escherichia coli isolates from animals, retail meat, and human patients in Guangzhou, China. Front Microbiol. 2017;8:1982. doi:10.3389/fmicb.2017.01982
46. Pham thanh D, Thanh Tuyen H, Nguyen Thi Nguyen T. Inducible colistin resistance via a disrupted plasmid-borne mcr-1 gene in a 2008 Vietnamese Shigella sonnei isolate. J Antimicrob Chemother. 2016;71(8):2314–2317. doi:10.1093/jac/dkw173
47. Snesrud E, He S, Chandler M. A model for transposition of the colistin resistance gene mcr-1 by IS Apl1. Antimicrob Agents Chemother. 2016;60(11):6973–6976. doi:10.1128/AAC.01457-16
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.