Back to Journals » ImmunoTargets and Therapy » Volume 15
NUP85 as a Pan-Cancer Immune Biomarker: Integrated Multi Omics and Functional Analyses Reveal Its Role in Tumor Prognosis
Authors Kong W, Hu Y, Zhang K, Zhu Z, Zou Y, Teng L, Wang X, Zhang J ![]()
Received 21 May 2025
Accepted for publication 3 March 2026
Published 17 March 2026 Volume 2026:15 541852
DOI https://doi.org/10.2147/ITT.S541852
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Flavio Salazar-Onfray
Weihao Kong,1,2,* Ying Hu,3,* Kangjie Zhang,2,* Zhipeng Zhu,4,* Yajun Zou,1 Long Teng,1 Xingyu Wang,1 Jianlin Zhang1
1Department of Emergency Surgery, the First Affiliated Hospital of Anhui Medical University, Hefei, Anhui Province, People’s Republic of China; 2West China School of Clinical Medicine, West China Hospital, Sichuan University, Chengdu, Sichuan Province, People’s Republic of China; 3State Key Laboratory of Pharmaceutical Biotechnology, School of Life Sciences, Nanjing University, Nanjing, Jiangsu Province, People’s Republic of China; 4First Clinical Medical College, Anhui Medical University, Hefei, Anhui Province, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Xingyu Wang, Department of Emergency Surgery, the First Affiliated Hospital of Anhui Medical University, Hefei, Anhui Province, People’s Republic of China, Email [email protected] Jianlin Zhang, Department of Emergency Surgery, the First Affiliated Hospital of Anhui Medical University, Hefei, Anhui Province, People’s Republic of China, Email [email protected]
Purpose: NUP85 encodes protein components of the Nup107-160 subunit of the nuclear pore complex, belonging to the Nucleoporins (NUPs) family, potentially implicating its role in human cancer. This study aims to elucidate the potential involvement of NUP85 in cancer pathogenesis.
Methods: Leveraging data from The Cancer Genome Atlas (TCGA), Genotype-Tissue Expression (GTEx), Clinical Proteomic Tumor Analysis Consortium (CPTAC), Cancer Cell Line Encyclopedia (CCLE), Human Protein Atlas (HPA), Gene Expression Profiling Interactive Analysis (GEPIA), CellMiner, and GeneMANIA databases, we investigated the role of NUP85 across various tumors. Correlations between NUP85 expression and pathological stage, histological grade, survival, immune infiltration, tumor mutational burden (TMB), microsatellite instability (MSI), drug resistance, DNA methylation, copy number variation (CNV), and single-cell expression were analyzed. Gene functional enrichment analysis was conducted to explore NUP85-associated pathways. Molecular biology experiments including Western blotting, flow cytometry, trans-well migration, and invasion assays were performed to validate NUP85’s oncogenic role in lung adenocarcinoma (LUAD) and oral squamous cell carcinoma (OSCC) cell lines.
Results: Our findings reveal up-regulated expression of NUP85 in most tumor tissues, with significant correlations observed with pathological stage, survival, immune infiltration, TMB, MSI, drug resistance, DNA methylation, and CNV. Molecular biology experiments confirm NUP85’s tumor-promoting role in LUAD and OSCC cell lines. Single-cell sequencing data suggest elevated NUP85 expression primarily in proliferative T cells (Tprolif).
Conclusion: NUP85 emerges as a potential tumor marker associated with tumor immunity and poor prognosis. These insights offer avenues for the development of novel therapeutic targets and anti-neoplastic drugs.
Keywords: NUP85, pan-cancer analysis, immune, prognosis, tumor marker
Introduction
Cancer is a multi-disease characterized by the uncontrolled proliferation of somatic cells, which can spread to other organs and cause metastasis. Although cell growth, differentiation, and death processes are tightly programmed, some cells, particularly those with somatic and epigenetic mutations, may evade regulation and grow uncontrollably.1 There are many different tumor types, and the majority of tumors have multiple oncogenic alterations with significant degree of intra and inter tumor heterogeneity. The top six cancer types, according to sex, account for more than 50% of new cancer diagnoses and deaths worldwide. The cancer incidence rates from a total of 11.7% estimated cases in 2020 were as follows: breast (11.7%), lung (11.4%), colorectal (10.0%), prostate (7.3%), stomach (5.6%), and liver cancers (4.7%).2 Oncogenic alterations have multiple effects, including increase in proliferative signaling, resistance to cell death, bypassing replicative limit, increase in genome instability.3 Improvements in understanding of the complexity of cancer biology have been instrumental in advancing cancer treatments. Oncologic therapeutics have traversed four overlapping paradigms over the last few centuries: surgery/radio therapy for local control, cytotoxic chemotherapy for systemic micro-metastatic eradication, molecularly targeted agents that silence driver oncogenes with minimal collateral toxicity, and recent immune-centered modalities. Significantly, immune-centered modalities include checkpoint inhibitors, adoptive CAR/TCR-engineered cells, bispecific antibodies, and personalized neo-antigen vaccines that reactivate endogenous or synthetic immunity to achieve treatment-free survivorship in 20–40% of selected advanced solid and hematologic cancers. Future integrative regimens that couple real-time molecular surveillance with CRISPR editing, oncolytic viruses, microbiome tuning and artificial intelligence-driven drug sequencing aspire to improve cancer treatments.4
Nuclear pore complexes (NPCs) are “supramolecular complexes” on the nuclear envelope assembled from multiple copies of approximately 30 different proteins called nucleoporins (Nups). NPCs not only provide aqueous channels for nucleocytoplasmic transport during interphase mediate nucleocytoplasmic transport, but also play a crucial role in genome organization and cellular homeostasis.5 The normal function and localization of proteins are crucial for the normal functioning of cells and good human health. However, the dysregulation of protein nucleocytoplasmic transport by NPCs will gradually increase the basis of human diseases. A number of physiological and cellular pathways implicated in tumorigenesis including RanGap1/RanBP2 signaling axes, GSK3β-Snail, DNA damage response and Wnt/β-Catenin, resistance to apoptosis and chemotherapy, are altered as a result of variation in several NPCs.6,7 Therefore, NPCs play a pivotal role in carcinogenesis. NUP85, also known as FROUNT, encodes the protein components of the Nup107-160 subunit of the nuclear pore complex (NPC). As a crucial component of NPC, the absence of the Nup107-160 complex results in severe defects in NPC assembly, leading to consequential outcomes.8 Studies by Yuya Terashima et al have elucidated that NUP85 binds to the C-terminal of C-C Motif Chemokine Receptor 2 (CCR2), amplifying the chemokine-elicited phosphatidyl-inositol 3-kinase-Rac-lammellipodium protrusion cascade, thereby promoting monocyte/macrophage infiltration.9 This mechanism has been identified to mediate inflammation via NUP85.10,11 Additionally, NUP85 interacts with C-C Motif Chemokine Receptor 5 (CCR5), enhancing CCR5-mediated cell chemotaxis.12 Within the tumor microenvironment, comprising diverse immune cells and other cell types, macrophages are prominent and linked to cancer progression, attributed to mechanisms such as promoting immune evasion.13 Recent investigations have spotlighted the significant impact of NUP85 on tumor growth and metastasis, correlating with prognosis in certain cancers. Notably, NUP85 may facilitate macrophage infiltration by binding to chemokine receptors CCR2 and CCR5, thereby influencing tumorigenesis and progression. In lung cancer, elevated NUP85 expression correlates with poor prognosis. Inhibition of NUP85 expression reduces macrophage accumulation and tumor growth.14 Moreover, in pancreatic cancer, NUP85-targeting nanoparticles effectively suppress retroperitoneal invasion and peritoneal dissemination,15 underscoring NUP85’s pivotal role across various cancers. Despite NUP85’s potential as a biomarker for human malignancies, research remains limited, necessitating comprehensive analysis of its association with pan-cancer.
This study comprehensively examines NUP85’s role in human cancer prognosis and immunology. Through extensive data analyses, we visualize NUP85 expression in pan-cancer and assess its impact on tumor-infiltrating immune cells (TIIC) and related immune markers. Our findings indicate that elevated NUP85 expression correlates with poor prognosis, albeit not universally present across all cancer tissues, suggesting diverse roles in different cancer types. Furthermore, we observe an association between NUP85 and tumor mutation burden and microsatellite instability. Molecular biology experiments based on LUAD and OSCC validate these results. Overall, this study aims to elucidate NUP85’s potential role in human cancer and provide insights for novel tumor treatment strategies.
Materials and Methods
Pan-Cancer Expression Profiling and Clinical Relevance of NUP85
To systematically evaluate the dysregulation of NUP85 across human cancers, we integrated mRNA-seq data from The Cancer Genome Atlas (TCGA) and matched normal tissues from the Genotype-Tissue Expression (GTEx) project. Expression values were uniformly normalized as transcripts per kilobase million (TPM), log2-transformed (log2[TPM+1]), and subjected to quality control (removal of duplicates and missing values). Differential expression between tumor and adjacent/normal tissues was assessed using Wilcoxon rank-sum tests. To complement transcriptomic findings with protein-level evidence, immunohistochemical staining data for NUP85 in normal and malignant tissues were retrieved from the Human Protein Atlas (HPA). Given the emerging role of nucleoporins in cancer progression, we further hypothesized that NUP85 expression might correlate with key clinical outcomes. Using TCGA clinical annotations, we performed univariate Cox regression analyses to evaluate the association between NUP85 expression and overall survival (OS), disease-specific survival (DSS), and progression-free interval (PFI). Survival curves were also generated via median-based stratification of patients into high- and low-expression groups, leveraging the GEPIA platform for consistency with established pan-cancer survival pipelines.
Association of NUP85 with Tumor Immune Microenvironment
To test whether NUP85 expression reflects an immunologically “hot” versus “cold” tumor phenotype—a hallmark of response to immunotherapy—we quantified its correlation with immune cell infiltration across 33 cancer types. Immune scores (ImmuneScore, StromalScore, and ESTIMATEScore) were computed using the ESTIMATE algorithm, while abundances of major immune subsets (eg., CD8⁺ T cells, B cells, macrophages) were inferred via multiple deconvolution methods, including TIMER, CIBERSORT, xCell, MCP-counter, EPIC, IPS, and quanTIseq, as implemented in the IOBR R package. Spearman’s rank correlation was used to assess associations between NUP85 expression and both immune cell fractions and stromal components. Furthermore, given the therapeutic relevance of immune checkpoints, we examined co-expression patterns between NUP85 and canonical immune checkpoints, cytokines/chemokines across TCGA tumors, again using Spearman correlation on log2(TPM+1)-transformed data. To explore NUP85 expression at single-cell resolution across tumor cell types, we leveraged the TISCH database (http://tisch.comp-genomics.org), which integrates scRNA-seq data from diverse cancers.
Genomic Instability and Epigenetic Regulation of NUP85
We investigated whether NUP85 dysregulation is linked to underlying genomic or epigenetic alterations. Tumor mutational burden (TMB) was calculated from TCGA somatic mutation data (MAF files) using the maftools R package, defined as total exonic mutations per megabase. Microsatellite instability (MSI) scores were obtained directly from TCGA molecular annotations. Spearman correlation analyses were then performed to evaluate relationships between NUP85 expression and both TMB and MSI—two established biomarkers of genomic instability and potential predictors of immunotherapy response. To explore epigenetic and structural drivers of NUP85 expression, we leveraged the Gene Set Cancer Analysis (GSCA) platform to assess correlations between mRNA levels and copy number variation (CNV) as well as DNA methylation across pan-cancer cohorts. Statistically significant associations (P < 0.05) were visualized using GSCA’s built-in circular plots, with effect size indicated by color intensity.
Functional Context and Drug Sensitivity Implications
To place NUP85 within a broader biological network, we constructed a protein–protein interaction (PPI) network using GeneMANIA, incorporating physical interactions, co-expression, pathway co-membership, and shared domains. Top 20 interacting partners were subjected to Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analyses via the clusterProfiler R package, revealing potential roles in nuclear transport, cell cycle regulation, and RNA processing. Finally, to assess the translational relevance of NUP85 expression, we queried the CellMiner database—which integrates multi-omics and drug sensitivity profiles across >1,000 cancer cell lines—for correlations between NUP85 mRNA levels and responses to 20,000+ compounds. Missing drug response values were imputed using k-nearest neighbors (KNN), and Pearson correlation coefficients were computed; associations with P < 0.05 were considered significant, highlighting candidate therapeutics whose efficacy may depend on NUP85 status.
Cell Lines and Cell Culture
Human lung adenocarcinoma (LUAD) PC-9 cells, human oral squamous cell carcinoma (OSCC) HSC-2 cells, and HEK 293T cells were obtained from Wuhan Procell Life Science & Technology Co., Ltd. (Wuhan, China). All cells were stored in liquid nitrogen and cultured in modified Eagle’s Medium (DMEM, Gibco BRL, USA) supplemented with 1% antibiotics (100 U/mL penicillin and 100 ug/mL streptomycin sulfates, Sigma, USA) and 10% heat-inactivated fetal bovine serum (FBS, Gibco, USA) in a humidified incubator (5% CO2 at 37°C).
NUP85 Short Hairpin RNA Transfection
Short hairpin RNA (shRNA) oligonucleotides targeting NUP85 genes and scrambled shRNA were designed and synthesized by the Gene Pharma Corporation (Shanghai, China) and contained the following sequences:NUP85-shRNA: GGAGATGCTGGTATGTGAAAC. Briefly, Human LUAD cells (PC-9 cells) and Human OSCC cells (HSC-2 cells) in the logarithmic phase were inoculated into 6-well plates with antibiotic-free DMEM. The cell density per well was 6×105 cells. LipofectamineTM2000 kit was used for transfection. Immediately, cells were cultured in a humidified incubator (5% CO2 at 37°C) for 24h and then collected.
Western Blotting
Total proteins were extracted by using a radioimmunoprecipitation assay buffer reagent (RIPA, Beyotime, China) and phenylmethanesulfonyl fluoride (PMSF, Sigma, USA). A Bicinchoninic acid (BCA) protein assay kit (Beyotime, China) was used to measure the protein concentrations. Proteins from each sample were separated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred onto the Polyvinylidene fluoride (PVDF) membrane (Millipore, USA). The lysates of tissues or cells with equal weight were separated as aforementioned. After blocking in the 5% skim milk with tris buffered saline tween (TBST, Boster, China), the transferred membranes were incubated in the primary antibodies at 4°C overnight and then washed by TBST (TBS+Tween) three times for 15 minutes each. Subsequently, the transferred membranes were incubated with secondary antibodies at RT for 1h. After extensive washing in TBST, immunoblotting was visualized using the enhanced chemiluminescence (ECL) development kit (Thermo Scientific, USA) and analyzed with Quantity-one software (Bio-Rad, USA). Antibodies used: rabbit anti-MMP-2, rabbit anti-MMP-9, rabbit anti-Bax, and rabbit antiBcl-2 were purchased from Abcam UK.
Flow Cytometry Assay
For cell cycle assay, cells were first harvested by trypsinization and then fixed in 75% ice-cold ethanol in phosphate-buffered saline. The cells were added Bovine pancreatic RNase (2 mg/mL, Sigma) and propidium iodide (10 mg/mL, Invitrogen). And then, the cells were incubated for 30 min at RT and protected from light meanwhile. The flow cytometer (BD Biosciences, NJ, USA) was used to analyze the cell distribution. For cell apoptosis assay, specific operating procedures refer to the manufacturer’s protocols. Cell apoptosis was then measured by using the flow cytometer (BD Biosciences, NJ, USA). In the graph, the four quadrants respectively stand for necrotic cells, viable cells, early-stage apoptotic cells, and late-stage apoptotic cells.
Transwell Migration and Invasion Assay
Human PC-9 and HSC-2 cells were inoculated into the upper chamber with a serum-free medium. Each well was a density of 2×106 cells. The bottom chamber is filled with 500 µL of 20% fetal bovine serum (FBS) culture medium. After incubating in a 5% (v/v) CO2 incubator at RT for 2 d, removing the non-invasive cells and matrigel in the upper chamber, and then the cells were fixed on the lower surface with 10% neutral buffered formalin solution and 0.1% crystal violet staining. Count invading cells in five randomly selected microscope fields.
Results
Nup85 Expression Was Altered in Cancer
RNA sequencing data sourced from the TCGA database revealed significant differential upregulation of NUP85 mRNA across 20 cancer types (P<0.01), with the exception of downregulation noted in KICH (P<0.01) (Figure 1A). To address the lack of normal samples in certain tumors, we integrated data from the GTEx database, which highlighted a noteworthy upregulation of NUP85 mRNA in 24 tumor types and a downregulation in 6 tumor types (Figure 1B). Discrepancies observed between Figure 1A and B may stem from limited normal samples in the TCGA dataset. Furthermore, analysis from the CPTAC database unveiled a significant increase in NUP85 protein expression in colon cancer, breast cancer, glioblastoma, hepatocellular carcinoma, head and neck squamous cell carcinoma, lung adenocarcinoma, lung squamous cell carcinoma, and renal clear cell carcinoma compared to normal samples (P<0.001) (Figure 1C). Consistent with these findings, data from the DepMap database demonstrated CERES scores predominantly less than −1 across most tumors (Supplementary Figure 1). Moreover, insights from the TISCH database delineate varying expression abundances of NUP85 across distinct cell subpopulations in various tumor types. Notably, in colorectal cancer (CRC), liver hepatocellular carcinoma (LIHC), and skin cutaneous melanoma (SKCM), the highest expression of NUP85 was observed in proliferating T cells (Supplementary Figure 2). Immunohistochemical staining results sourced from the HPA database validated heightened protein levels of NUP85 in various cancer tissues compared to normal tissues (Supplementary Figure 3). Collectively, these findings underscore substantial differences in NUP85 expression levels between tumor tissues and corresponding normal tissues, thereby positioning NUP85 as a promising candidate for a tumor marker.
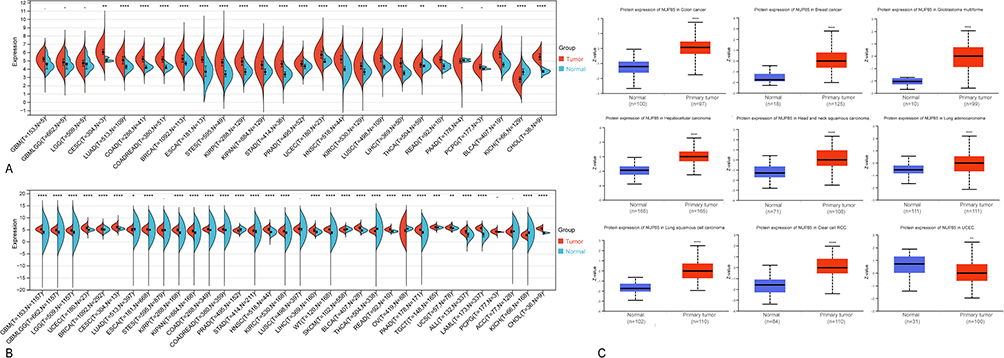 |
Figure 1 The expression level of NUP85 in different tumors and corresponding normal tissues. (A) Expression levels of NUP85 in tumor tissues compared to corresponding normal tissues across 26 cancer types using TCGA data. Blue indicates normal tissue, and yellow represents tumor tissue. Significance levels are denoted as *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001. (B) Differential expression analysis of NUP85 between tumor tissues and normal tissues across 34 cancer types, integrating TCGA and GTEx data. Log2 (TPM +1) scale is utilized. (C) Expression differences of NUP85 protein in normal tissues compared to tumor tissues sourced from the CPTAC Database. Notes: Significance levels are denoted as *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001. |
The Expression Level of NUP85 Was Associated with the Tumor Stage and Histological Grade
The Kruskal–Wallis test served to analyze expression differences of the gene across samples from distinct clinical stages within each tumor type. Significant differences were observed in eight tumor types, namely ACC (P<0.01), BRCA (P<0.05), HNSC (P<0.01), KIPAN (P<0.05), LIHC (P<0.001), LUSC (P<0.05), OV (P<0.01), and SKCM (P<0.01) (Figure 2A).
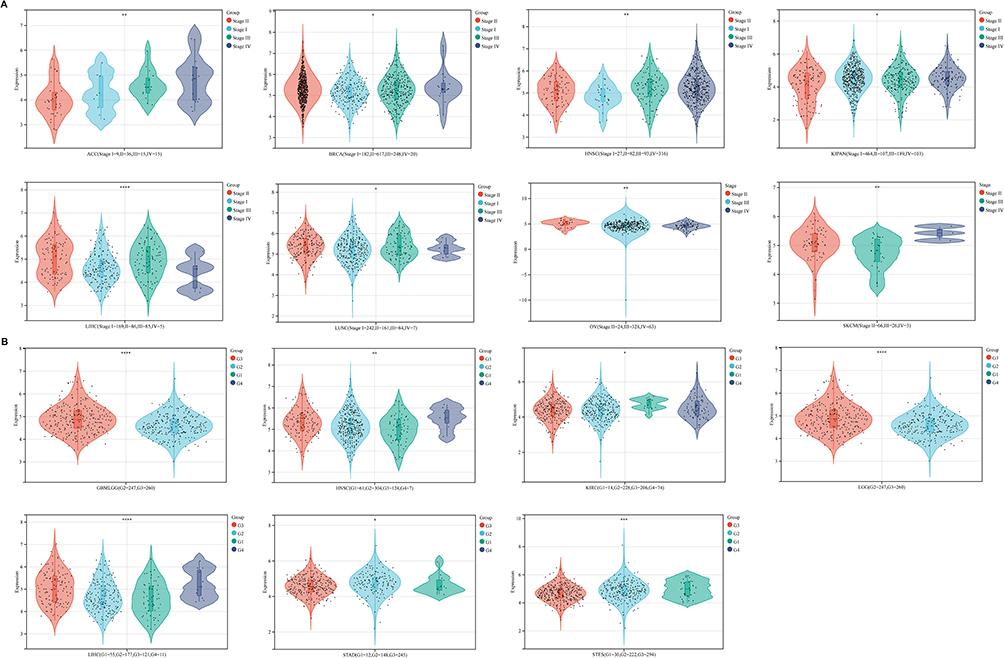 |
Figure 2 Differential analysis of NUP85 mRNA expression across various pathological stages and histological grades. (A) Differential analysis of NUP85 mRNA expression across various pathological stages. (B) Differential analysis of NUP85 mRNA expression across various histological grades. Notes: Significance levels are denoted as *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001. |
To delve deeper, we utilized the “single gene analysis” module of the GEPIA database, focusing on “NUP85”, and explored its correlation with the pathological stage of cancer using the “pathological staging map” module. Log2 (TPM+1) was adopted as the logarithmic scale. Supplementary Figure 4A illustrates a significant correlation between NUP85 expression and the pathological stage of pan-cancer (P<0.001).
Further exploration revealed significant correlations between NUP85 expression and Histological grade in GBMLGG (P<0.001), HNSC (P<0.01), KIRC (P<0.05), LGG (P<0.001), LIHC (P<0.001), STAD (P<0.05), and STES (P<0.001) (Figure 2B). These findings shed light on the potential involvement of NUP85 in cancer progression and grading across diverse tumor types.
High Expression of NUP85 Was Associated with a Poor Prognosis
mRNA expression data of NUP85 and corresponding patient survival information were extracted from the TCGA database. Univariate Cox regression analysis, facilitated by the survival package, unveiled significant insights, as illustrated in Figure 3A. Notably, the expression of NUP85 exhibited a notable risk ratio (HR>1, P<0.05) across 9 cancer types: GBMLGG, KIPAN, ACC, LGG, KICH, KIRC, LIHC, PRAD, and Uveal Melanoma (UVM). Elevated NUP85 expression emerged as a high-risk factor for these cancers, signifying a worse prognosis for patients with higher NUP85 expression levels.
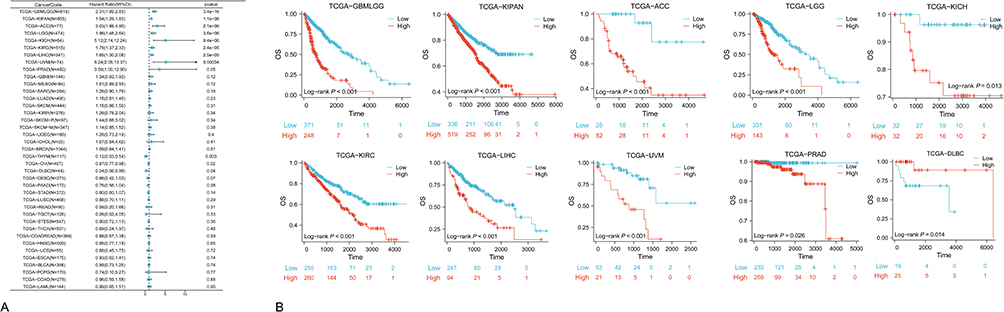 |
Figure 3 OS Analysis based on TCGA data. (A) Forest plot obtained from univariate cox regression analysis depicting the correlation between NUP85 expression and overall survival (OS). (B) Kaplan-Meier survival curves comparing high and low NUP85 expression groups in 10 cancer types (P<0.05). |
Further survival analyses were conducted for each specific cancer. Employing the best cut-off value for NUP85 expression, patients were segregated into high and low NUP85 expression groups. Figure 3B underscores a correlation between high NUP85 expression and poorer cancer prognosis across GBMLGG, KIPAN, ACC, LGG, KICH, KIRC, LIHC, UVM, and PRAD. Survival analysis conducted via the GEPIA website, dividing all cancer patients based on median NUP85 expression levels into high (4749 patients) and low (4747 patients) expression groups, revealed a significant difference in survival prognosis between the two groups (P=1.3e-14). Patients with elevated NUP85 expression exhibited significantly worse outcomes compared to those with lower expression levels, as depicted in the Kaplan-Meier survival curve (Supplementary Figure 4B).
Similar analyses were replicated using Disease-Specific Survival (DSS) data, yielding parallel results. Univariate Cox analysis (Figure 4A) reiterated significant risk ratios (HR>1, P<0.05) for NUP85 expression in GBMLGG, KICH, LGG, ACC, KIPAN, KIRC, LIHC, UVM, and SKCM. Corresponding survival analysis (Figure 4B) affirmed the association between high NUP85 expression and poorer prognosis across the aforementioned cancer types, along with PRAD. The results from the GEPIA website indicate that patients with expression levels of NUP85 mRNA above the median value have significantly worse DFS compared to patients with expression levels below the median of NUP85 (Supplementary Figure 4C).
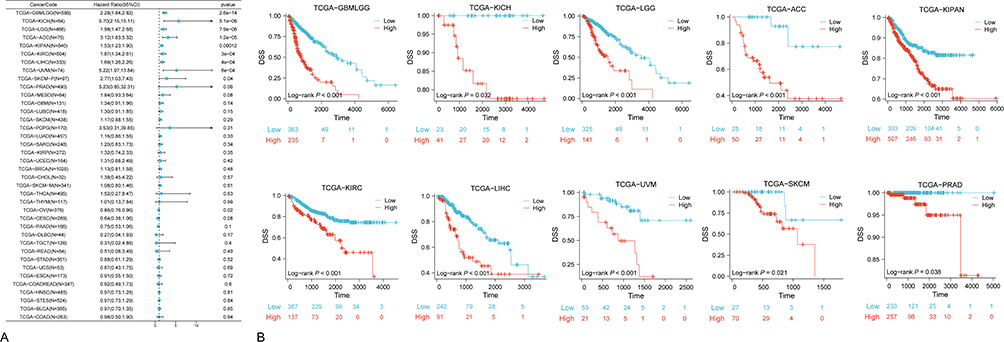 |
Figure 4 DSS Analysis based on TCGA data. (A) Forest plot obtained from univariate cox regression analysis reflecting the correlation between NUP85 expression and disease-specific survival (DSS). (B) Kaplan-Meier survival curves comparing high and low NUP85 expression groups in 10 cancer types (P<0.05). |
Additionally, considering Progression-Free Interval (PFI) as the survival outcome, univariate analysis (Figure 5A) disclosed significant associations (P<0.05) with GBMLGG, PRAD, LIHC, LGG, UVM, KIPAN, KICH, ACC, and PCPG. Subsequent survival analysis (Figure 5B) reiterated the impact of NUP85 expression on PFI, with patients exhibiting high NUP85 mRNA levels differing significantly in PFI compared to those with low expression across GBMLGG, PRAD, LIHC, LGG, UVM, KIPAN, KICH, ACC, PCPG, and KIRC.
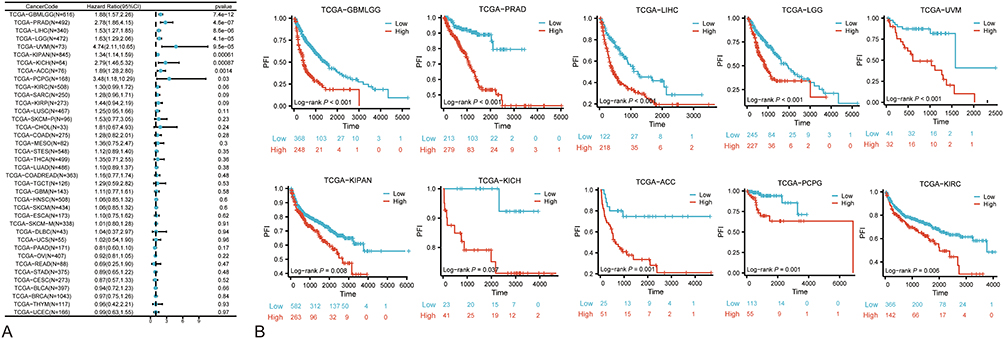 |
Figure 5 PFI Analysis based on TCGA data. (A) Forest plot obtained from univariate cox regression analysis depicting the correlation between NUP85 expression and progression-free interval (PFI). (B) Kaplan-Meier survival curves comparing high and low NUP85 expression groups in 10 cancer types (P<0.05). |
NUP85 Expression Level Correlated with the Tumor Immune Microenvironment
Tumor progression is intricately shaped by the co-evolution between tumor cells and their microenvironment, with the immune microenvironment (TME) playing a pivotal role. Thus, we embarked on a preliminary exploration of the interaction between NUP85 and the immune microenvironment. Leveraging the IOBR package, we analyzed the correlation between NUP85 and various immune infiltration markers employing algorithms such as TIMER, ESTIMATE, MCP, EPIC, IPS, Quantiseq, Cibersort, and xCell.
Results from the TIMER algorithm revealed a significant correlation between NUP85 mRNA and six types of immune cells (Figure 6A), notably prominent in KICH, LUSC, and THYM (Figure 6B). Additionally, we evaluated the relationship between NUP85 expression and tumor immune infiltration using Stromal Score, Immune Score, and ESTIMATE Score. Figure 7 delineates the top 3 tumors exhibiting the most significant correlation with NUP85 expression across these scores: BRCA, STES, and LUSC (Stromal Score); GBMLGG, STES, and LUSC (Immune Score); and STES, STAD, and LUSC (ESTIMATE Score). Moreover, analysis results from MCP, EPIC, IPS, Quantiseq, Cibersort, and xCell algorithms further corroborated a correlation between NUP85 mRNA and multiple immune cells (Figure 8).
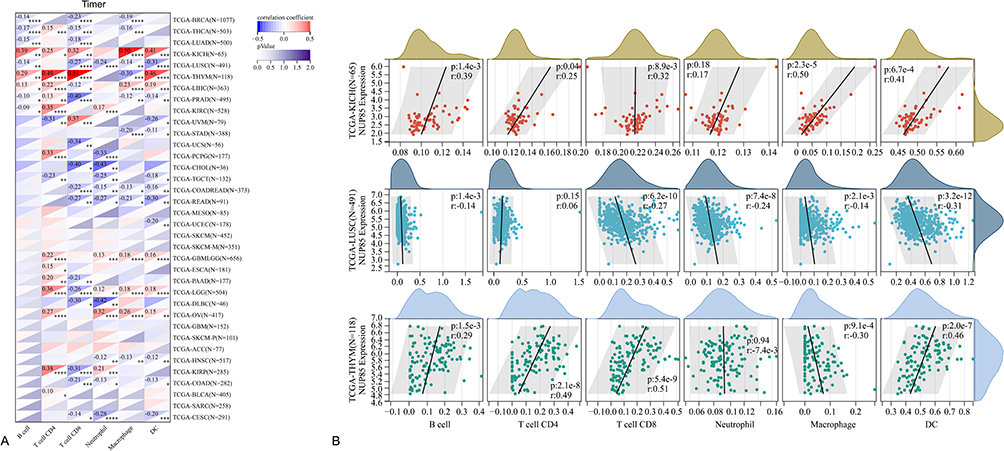 |
Figure 6 Analysis of the correlation between NUP85 expression levels and infiltration abundance of six main immune cell types (B cells, CD4+ T cells, CD8+ T cells, dendritic cells, macrophages, and neutrophils) using the TIMER method. (A) Heatmap of correlations between NUP85 and six immune cell types in the Timer algorithm. (B) Top three tumor types showing the strongest correlations with immune cells and their corresponding scatter plots. Notes: Significance levels are denoted as *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001. |
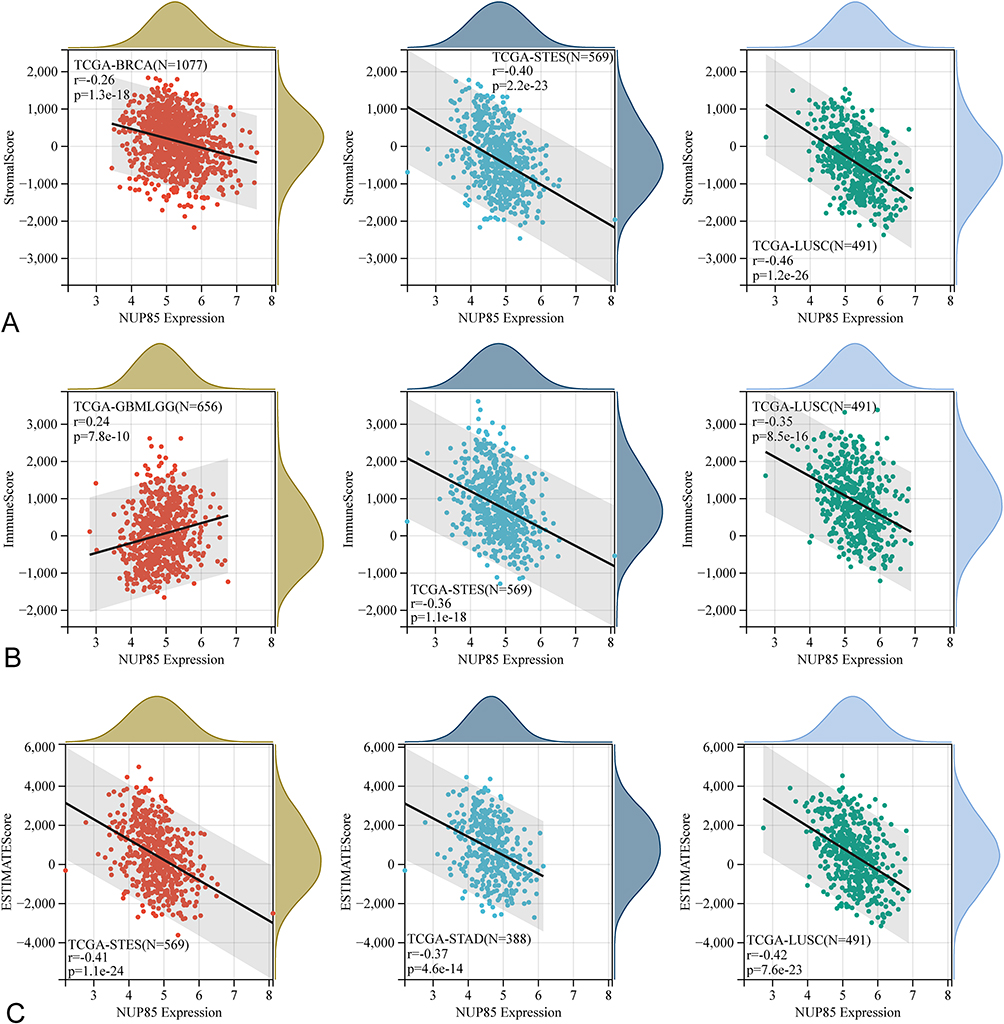 |
Figure 7 The correlation between NUP85 expression and three immune scores using the ESTIMATE method. The top 3 tumors exhibiting the most significant correlation between immune infiltration degree and NUP85 expression are LUSC, BRCA, and STES for Stromal Score; LUSC, GBMLGG, and STES for Immune Score; LUSC, STAD, and STES for ESTIMATE Score. (A) Scatter plots showing the correlation between NUP85 and stromal score for the top three tumors with the strongest association. (B) Scatter plots showing the correlation between NUP85 and immune score for the top three tumors with the strongest association. (C) Scatter plots showing the correlation between NUP85 and ESTIMATE score for the top three tumors with the strongest association. |
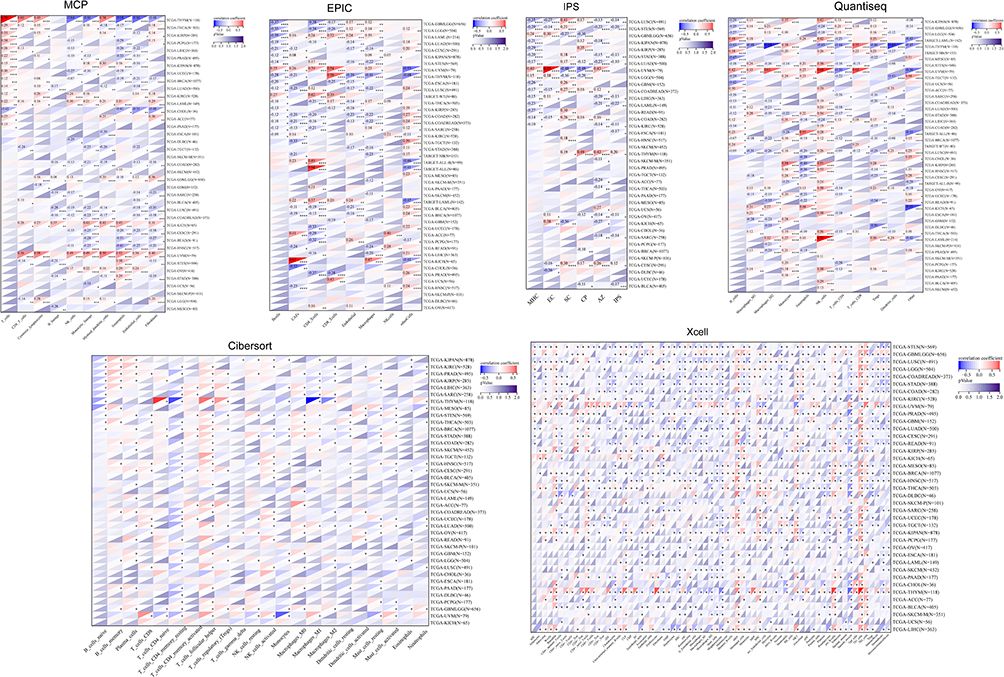 |
Figure 8 The correlation between NUP85 expression and multiple immune cells using various methods including MCP, EPIC, IPS, Quantiseq, Cibersort, and xCell. Notes: Significance levels are denoted as *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001. |
Furthermore, leveraging RNA sequencing data from the TCGA database, we examined the relationship between NUP85 expression and the expression of immune checkpoint genes (supplementary Figure 7A). Results underscored significant correlations between NUP85 expression and certain immune checkpoint genes across different tumors (P<0.05). For instance, in ACC, BLCA, BRCA, GBM, HNSC, KICH, KIRP, NUP85 expression were positively correlated with the TNFRSF14 and LAG3 expression (P<0.05). Similarly, we observed that NUP85 is significantly associated with multiple cytokines/chemokines and immune-related pathways (supplementary Figure 7B and C). Finally, our analysis of immunotherapy cohorts revealed that NUP85 demonstrates notable value in predicting both treatment response and patient prognosis (supplementary Figure 8).
NUP85 Expression Level Correlated with Tumor Mutational Burden
MSI, denoting microsatellite instability, signifies alterations in microsatellite (MS) sequence length due to insertion or deletion mutations during DNA replication, intimately linked with tumor mutagenesis. Leveraging MSI scores from the TCGA database, we analyzed their correlation with NUP85 expression data. Figure 9A elucidates a significant positive correlation between NUP85 expression and MSI in LUAD, BRCA, STES, SARC, STAD, PRAD, HNSC, LUSC, LIHC, THCA, PAAD, BLCA, CHOL, and DLBC. Conversely, in GBMLGG, COAD, and COADREAD, NUP85 expression exhibited a negative correlation with MSI. These findings were statistically significant (P < 0.05).
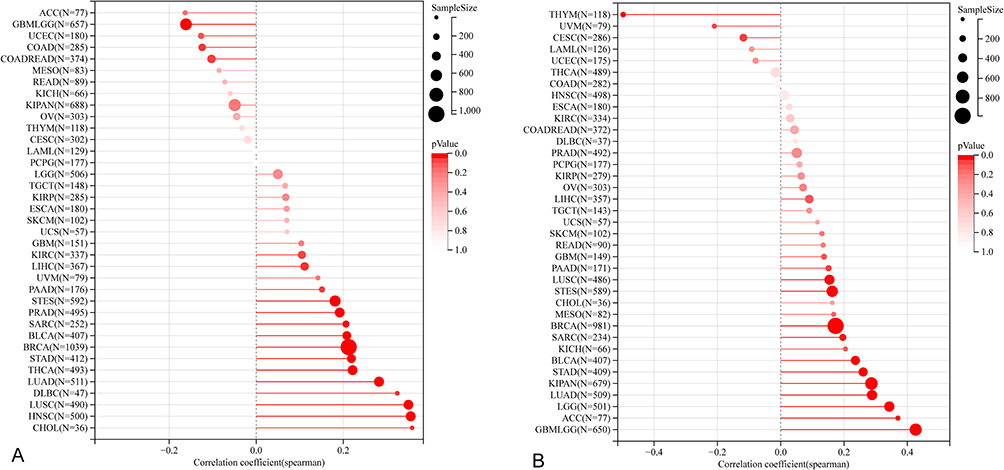 |
Figure 9 The correlation of NUP85 with MSI, and TMB. (A) Correlation analysis between NUP85 expression and MSI. (B) Correlation analysis between NUP85 expression and TMB. Notes: Black values represent the range, and red curves indicate the correlation coefficient. |
Tumor mutation constitutes a pivotal aspect of tumorigenesis. To delve deeper into the potential role of NUP85 in cancer, we delved into tumor mutation data. Somatic cell data (MAF data) spanning human pan-cancer were retrieved from the TCGA database, utilizing the number of mutations in exons to gauge TMB levels. As depicted in Figure 9B, NUP85 expression displayed a significant positive correlation with TMB levels in GBMLGG, LGG, LUAD, BRCA, STES, SARC, KIPAN, STAD, LUSC, BLCA, and ACC. Conversely, NUP85 expression exhibited a negative correlation with TMB levels in THYM and CESC.
NUP85 and Drug Resistance in Cancer
We conducted drug sensitivity analysis utilizing the CellMiner database. The mRNA expression level of NUP85 exhibited significant positive correlations with Nelarabine, Vorinostat, Fludarabine, Chelerythrine, Belinostat, Amonafide, 8-Chloro-adenosine, Cytarabine, Cladribine, AT-13387, and Hydroxyurea. Conversely, it demonstrated negative correlations with Mithramycin, Eribulin mesylate, Okadaic acid, Depsipeptide, and Actinomycin D (Figure 10). These findings shed light on potential associations between NUP85 expression and drug sensitivity, providing valuable insights for further investigation.
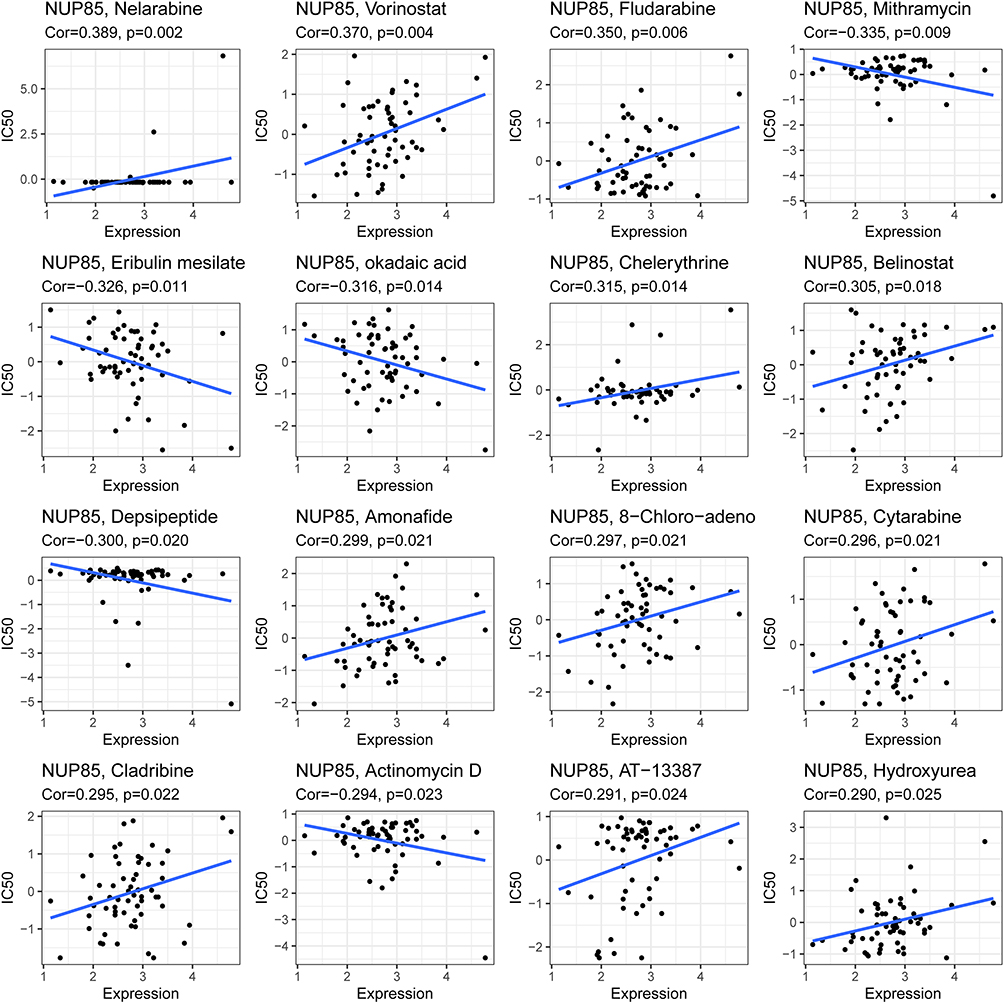 |
Figure 10 NUP85 and drug resistance in cancer. |
NUP85 Expression Level Was Associated with Gene Copy Number Variation and Methylation Level
On the GSCA platform, we assessed the correlation between CNV and NUP85 expression. Across 24 cancers, with the exception of CHOL, DLBC, GBM, LAML, LGG, PCPG, THCA, THYM, and UVM, NUP85 mRNA levels exhibited a positive correlation with CNV (Figure 11A). Notably, Figure 11B–D highlights the top three cancers (OV, LUSC, and BRCA) with the highest correlation scores.
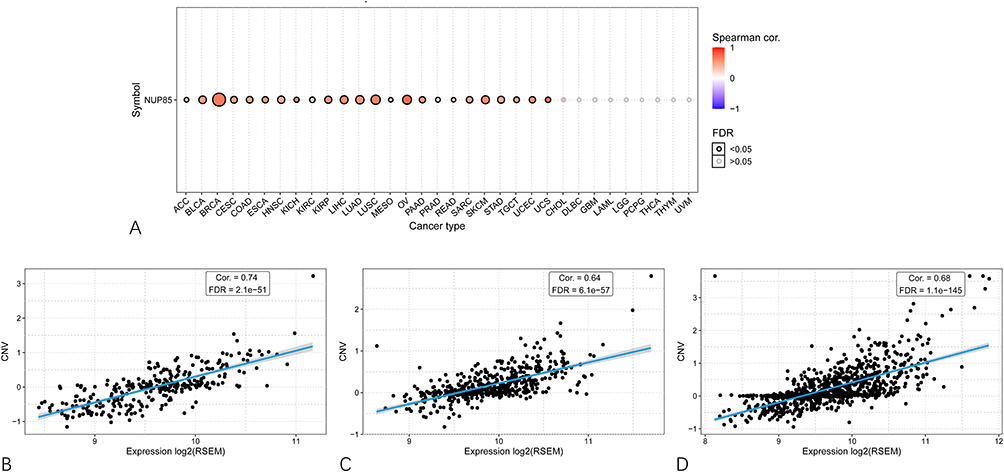 |
Figure 11 Association of NUP85 expression level with gene copy number variation. (A) Spearman correlation analysis between NUP85 mRNA expression and CNV in pan-cancer. Red indicates a positive correlation. (B–D) Top three cancer types (OV for (B) LUSC for (C) BRCA for (D) with the highest correlation scores between NUP85 expression and CNV. |
Furthermore, given the association between NUP85 expression and certain DNA methylation genes, we proceeded to explore the correlation between methylation and NUP85 expression. In 26 cancers, excluding CHOL, DLBC, GBM, LAML, OV, and THYM, NUP85 mRNA levels demonstrated a negative correlation with methylation (Figure 12A). Figure 12B–D showcases the top three cancers (LGG, PRAD, and BRCA) with the highest correlation scores. These findings underscore the intricate relationship between NUP85 expression, CNV, and methylation across various cancer types.
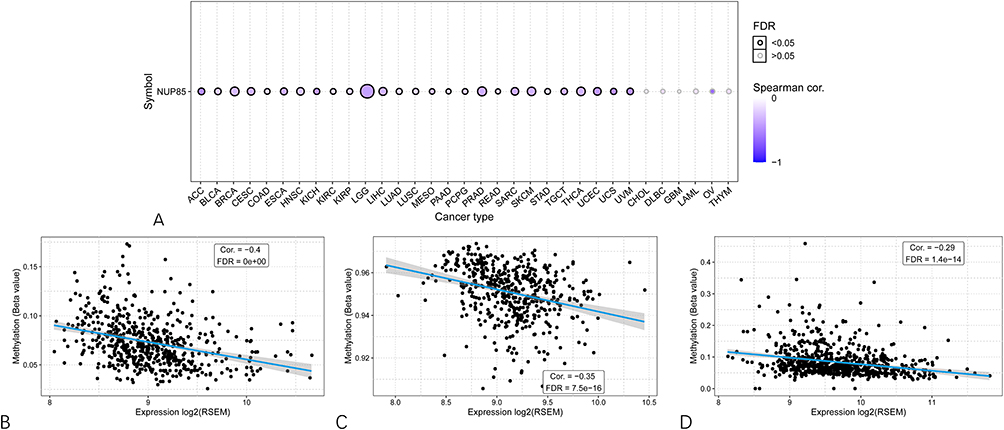 |
Figure 12 Association of NUP85 expression level with methylation level. (A) Spearman correlation analysis between NUP85 mRNA expression and methylation degree in pan-cancer. Blue indicates a negative correlation. (B–D) Top three cancer types (LGG for (B) PRAD for (C) BRCA for (D) with the highest correlation scores between NUP85 expression and methylation degree. |
PPI Network and Gene Enrichment Analysis of NUP85 in Cancer
We utilized the GeneMANIA online platform to construct a protein interaction network associated with NUP85, identifying 20 genes closely linked to NUP85 across six forms of interaction (Physical Interactions, Co-expression, Predicted, Co-localization, Genetic Interactions, Pathway, and Shared protein domains) (Figure 13A). Subsequently, we conducted a functional enrichment analysis of these genes (Figure 13B). The results of GO enrichment analysis revealed significant enrichment across 5 Biological Processes (Regulation of Glycolytic Process, ncRNA Export from Nucleus, tRNA Transport, tRNA-Containing Ribonucleoprotein Complex Export from Nucleus, and tRNA Transport from Nucleus), 5 Cellular Components (Centromeric Region, Kinetochore, Nuclear Pore, Host, and Nuclear Pore Outer Ring), and 5 Molecular Functions (Structural Constituent of Nuclear Pore, Signal Sequence Binding, Nuclear Localization Sequence Binding, tRNA (Guanine) Methyltransferase Activity, and Toxic Substance Binding). Furthermore, KEGG enrichment analysis indicated significant enrichment in pathways such as RNA Transport, Amyotrophic Lateral Sclerosis, mTOR Signaling Pathway, Viral Protein Interaction with Cytokine and Cytokine Receptor, and Thyroid Hormone Synthesis.
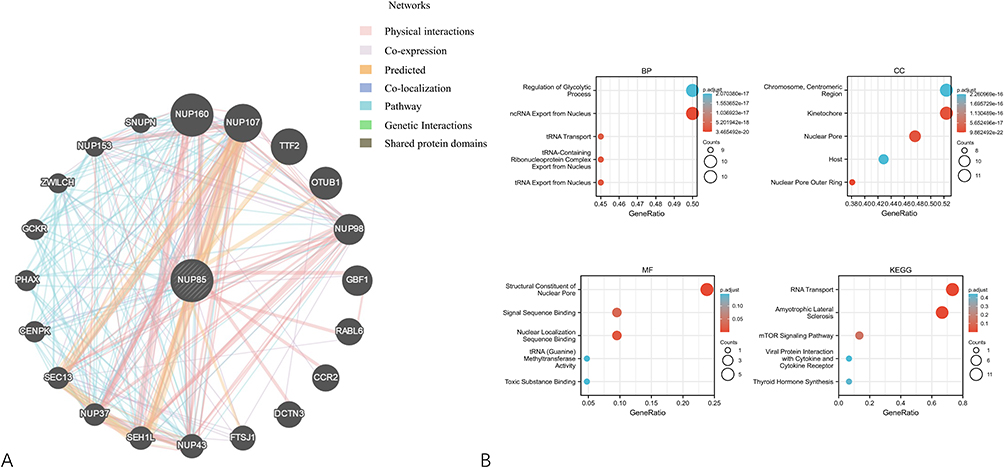 |
Figure 13 Functional enrichment analysis of NUP85. (A) Protein-protein interaction (PPI) network based on 20 NUP85-related genes. (B) Functional enrichment analysis including GO enrichment analysis (MF, BP, CC) and KEGG enrichment analysis. |
Moreover, leveraging pan-cancer expression matrices from TCGA and CCLE databases, we conducted correlation analysis between NUP85 and various genes. Based on different gene names and the magnitude of correlation, we performed GSEA analysis on NUP85. The results demonstrated significant enrichment of NUP85 in E2F Targets, G2M Checkpoint, and Myc Targets (Top 3) in both the TCGA and CCLE datasets (Supplementary Figure 5). These findings illuminate the diverse molecular mechanisms and pathways associated with NUP85 across different cancer types.
NUP85 Expression Levels Were Elevated in PC-9 Cells and HSC-2 Cells
We first analyzed the expression level of NUP85 in tumor cells and 293T cells. Western blotting results showed that the expression level of NUP85 in PC-9 cells and HSC-2 cells was higher than that in 293T cells (Supplementary Figure 6A and B).
NUP85 Inhibited Apoptosis and Promoted Proliferation in PC-9 and HSC-2 Cells
In vitro, we knocked down the expression of NUP85 in PC-9 and HSC-2 cells by transfecting shRNA (Supplementary Figure 6C and D) to further study the role of NUP85 in LUAD and OSCC. The results of flow cytometry showed that silencing NUP85 significantly promoted the apoptosis of PC-9 and HSC-2 cells (Supplementary Figure 6E and F). The results of Western blotting (Supplementary Figure 6G and H) validate the above conclusions. The shRNA of NUP85 inhibited the expression of Bcl-2 (B-cell lymphoma-2) and promoted the expression of Bax (Bcl-2-associated X) in PC-9 and HSC-2 cells. In addition, flow cytometry showed that PC-9 and HSC-2 cells transfected with NUP85-shRNA increased G0 / G1 phase, S phase, and G2 / M phase arrest (Supplementary Figure 6I and J). At the same time, the results of Western blotting (Supplementary Figure 6K and L) showed that the expression of Cyclin A2 protein and P-Rb protein decreased in PC-9 and HSC-2 cells transfected with NUP85-shRNA. Based on the above results, we found that NUP85 could inhibit apoptosis and promote cell proliferation in PC-9 and HSC-2 cells.
NUP85 Promoted Invasion and Migration of PC-9 and HSC-2 Cells
We further observed the effect of NUP85 on the invasion and migration of PC-9 and HSC-2 cells. Transwell and wound healing results (Supplementary Figure 6M and N) showed that the silencing of NUP85 could inhibit the invasion and migration of cells. The results of Western blotting (Supplementary Figure 6O and P) showed that the protein expression of MMP2 and MMP9 decreased in PC-9 and HSC-2 cells transfected with NUP85-shRNA. To sum up, we can conclude that NUP85 may promote the migration and invasion of tumor cells in vitro.
Discussion
The nuclear pore complex (NPC) is a massive protein assembly that regulates the transport of macromolecules between the nucleus and the cytoplasm, and it has been shown to be a key player in the regulation of important nuclear processes such as chromatin organization, gene expression, and DNA repair.16 NPC and nuclear transport receptors are important components of the nuclear transport system (NTS). Recent studies suggest that the nuclear transport factor has become a powerful target for targeted therapy of cancer.17 NUP85 is a member of the Nucleoporins (NUPs) family. NUPs are involved in a variety of basic cellular processes, such as cell differentiation, gene regulation, chromatin tissue/modification, DNA repair, mitosis, and reprogramming.18–20 On the one hand, NUPs play an important role in various biological processes, on the other hand, it also has a close relationship with cancer.21,22 Some NUPs, such as NUP62, NUP93, and NUP88, were thought to promote the proliferation, migration, and invasion of tumor cells.23–25 A study has shown that tumor cell metastasis is inhibited by targeting the mechanical transduction and focal adhesion defects required for tumor cell metastasis by targeting the expression of NUP210.26 Another study has shown that targeting inhibition of NUP214 expression by miR-133b in tumor cells leads to delayed mitosis, which in turn enhances chromosome abnormalities and apoptosis.27 Our study chose NUP85 as the object of study. Current studies have shown that NUP85 is highly expressed in tumor tissues, which mainly mediates the response of macrophages by binding to the C-terminal of CCR2, CCR5, thus giving full play to the tumor-promoting activity of macrophages.14 Therefore, targeted inhibition of NUP85 expression is considered to be an effective cancer treatment and has been verified in pancreatic cancer-related studies.15,28 However, the role of NUP85 in different cancers and whether it can be used as an effective tumor marker is unclear. Therefore, the pan-cancer analysis of NUP85 is necessary, which can help us to explore the role of NUP85 in different types of cancer and provide a basis for further elucidating the specific molecular mechanism of NUP85 in cancer.
To address this knowledge gap, we leveraged large-scale multi-omics resources to conduct a systematic investigation of NUP85 across diverse cancer types. The Cancer Genome Atlas (TCGA) remains a cornerstone of pan-cancer multi-omics research and has facilitated numerous biomarker discoveries—such as SHTN1 in bladder cancer, the exploration of oncogenic signaling pathways in oral squamous cell carcinoma (OSCC), and the identification of immune biomarkers in ovarian cancer.29–31 Building on this established paradigm, our study integrates multiple complementary approaches—including differential expression analysis (limma), tumor microenvironment characterization, functional enrichment (KEGG/GO), protein–protein interaction (PPI) network construction, survival modeling (Kaplan–Meier), and single-cell RNA-seq validation—to comprehensively delineate the multifaceted roles of NUP85 across cancer types. Nevertheless, bulk transcriptomic resources like TCGA carry inherent limitations, including variable tumor purity, contamination from stromal and immune cells, and technical batch effects. These factors can obscure cell-type-specific gene functions and pathway activities, necessitating cautious interpretation of findings derived from bulk data.32,33 To address these constraints, emerging computational strategies offer promising solutions. In particular, generative adversarial networks (GANs) have been adapted to synthesize realistic transcriptomes, impute missing data, and deconvolve bulk signals into cell-type-resolved profiles.34,35 Applied to NUP85—a nucleoporin implicated in both nuclear transport and immune modulation—such models could leverage paired bulk and single-cell datasets to infer its context-dependent expression or simulate perturbations (eg., knockdown) in silico. This may clarify whether NUP85’s dual roles arise from distinct cellular niches or reflect intrinsic multifunctionality within individual cells, a distinction largely inaccessible to conventional bulk analyses. While beyond the scope of this work, integrating GANs with spatial transcriptomics and machine learning–based drug response prediction represents a compelling avenue for future mechanistic and translational exploration.
Consistent with these methodological considerations, our integrative analyses yielded robust evidence linking NUP85 to adverse clinical outcomes and immune modulation in the tumor microenvironment. The current research focuses on conducting a comprehensive pan-cancer analysis of NUP85 to delve into its expression patterns across various cancer types. Our findings reveal a strong association between NUP85 overexpression and poor prognosis among cancer patients. Notably, the expression level of NUP85 correlates closely with the abundance of immune cell infiltration within tumors. Furthermore, NUP85 exhibits heightened expression across multiple tumor types and demonstrates significant correlations with immune markers, microsatellite instability (MSI), tumor mutational burden (TMB), and drug resistance. These results underscore the pivotal role of NUP85 in tumorigenesis and cancer progression, particularly highlighting its involvement in tumor immunity and mutation processes. The findings shed light on the potential of NUP85 as a prognostic marker and therapeutic target in cancer management.
Given the centrality of immune interactions in shaping tumor behavior, we next examined how NUP85 might interface with immune regulatory mechanisms within the TME. Mounting evidence underscores the immune system as a pivotal determinant in both tumor initiation and progression. The dynamic crosstalk between cancer cells and the tumor microenvironment (TME) has not only reshaped our understanding of oncogenesis but also paved the way for innovative immunotherapeutic strategies, particularly with the advent of immune checkpoint inhibitors.36,37 Emerging studies further reveal that dysregulation of nucleocytoplasmic transport—a fundamental cellular process mediated by nuclear pore complexes (NPCs)—can profoundly impair anti-tumor immunity. For instance, certain viral proteins subvert host interferon responses by blocking the nuclear import of key immune transcription factors such as STAT1 and IRF7.38 In the context of cancer, aberrant nucleocytoplasmic shuttling of NF-κB, a master regulator of inflammation and immune activation, has been shown to disrupt immune homeostasis and foster a pro-tumorigenic milieu.39 Collectively, these findings highlight an underappreciated yet critical role for nucleocytoplasmic transport machinery in modulating immune evasion. In this framework, our integrative analyses position NUP85—a core nucleoporin—not merely as a structural NPC component but as a potential regulator of the immune TME. We demonstrate that elevated NUP85 expression is significantly associated with increased macrophage infiltration in liver hepatocellular carcinoma (LIHC) (Figure 6A) and consistently correlates with poor clinical outcomes across multiple cancer types (Figures 3–5). This observation resonates with the work of Terashima et al,14 who reported that FROUNT–CCR2-driven accumulation of tumor-associated macrophages in the TME promotes cancer progression. Building on these convergent lines of evidence, we propose a mechanistic hypothesis: NUP85 overexpression in tumor cells may enhance macrophage recruitment or drive their polarization toward an immunosuppressive, pro-tumorigenic phenotype—potentially by facilitating the nuclear transport of immune-modulatory transcription factors or through context-dependent alterations in nucleoporin composition. Importantly, when evaluated across multiple immune-classified patient cohorts, NUP85 expression remains significantly predictive of both therapeutic response and overall survival, providing additional—albeit indirect—support for its functional relevance within the tumor immune landscape.
To translate these bioinformatic insights into biologically actionable knowledge, we performed experimental validation in representative cancer models. Through systematic analysis, we found that NUP85 is not only a potential tumor marker but also has a significant correlation with a variety of potential biomarkers. Common potential biomarkers are tumor mutation burden (TMB), microsatellite instability (MSI), and immune check molecule.40–42 Our research has demonstrated a significant relationship between NUP85 expression and various cancer biomarkers. Additionally, employing molecular biology techniques, we investigated the role of NUP85 in LUAD and OSCC. Western blotting confirmed up-regulated NUP85 expression in both LUAD and OSCC tissues. Knockout of the NUP85 gene in lung adenocarcinoma cells and oral squamous cell carcinoma cells revealed an increase in G0/G1 phase cells and apoptosis rate, indicating NUP85’s role in promoting proliferation and inhibiting apoptosis. Transwell assays further validated NUP85’s promotion of invasion and migration in these cancer cells. These experiments corroborate our bioinformatics analysis findings related to NUP85 and ensure the reliability of our research. Moving forward, similar molecular biological verifications will be conducted across additional cancer types.
Altogether, our findings converge on a model in which NUP85 exerts both canonical nuclear and non-canonical immune-modulatory functions that collectively fuel oncogenesis. Our study presents the first systematic pan-cancer analysis of NUP85, showing its consistent upregulation across cancers and strong links to poor prognosis, immune infiltration, genomic instability, and drug resistance. These findings extend prior work implicating NUP85 (FROUNT) in tumor-promoting inflammation—particularly its role in enhancing CCR2/CCR5-mediated macrophage recruitment, which aligns with our observation of elevated macrophage infiltration in LIHC. Beyond this known extranuclear function, we uncover novel nuclear roles: functional and PPI analyses tie NUP85 to RNA/tRNA export and kinetochore assembly as part of the Nup107–160 complex. GSEA further links it to E2F targets, G2M checkpoint, and Myc signaling, suggesting involvement in cell cycle dysregulation. Together, these results reveal a dual functionality—NUP85 may act both as a canonical nucleoporin and a cytoplasmic immune modulator—potentially explaining its broad oncogenic impact. However, despite our comprehensive approach utilizing multiple databases, our study has several limitations. Firstly, while we validated NUP85’s tumor-promoting role in LUAD and OSCC through molecular biology methods, our functional validation was limited to these two cancer types. Thus, further in vitro or in vivo studies using diverse cancer cell lines and clinical samples are warranted to confirm its role in tumorigenesis and development, and to support its potential as a pan-cancer biomarker with caution in generalizing the findings to all cancers. Secondly, the racial composition in the databases primarily comprises white and black individuals, necessitating validation across other racial groups. Thirdly, the integration of TCGA tumor data with GTEx normal tissues—though widely adopted—introduces potential batch effects due to differences in sample processing, sequencing platforms, and tissue sourcing. Although we applied uniform normalization (log2[TPM+1]), residual technical variation may influence differential expression estimates. Fourthly, while DepMap/CERES scores provide valuable insights into gene essentiality across cancer cell lines, they reflect in vitro genetic dependencies that may not fully recapitulate in vivo tumor biology. CERES corrects for copy-number-driven fitness effects but cannot account for microenvironmental influences—such as immune surveillance, stromal interactions, or spatial heterogeneity—that critically shape NUP85’s role in actual tumors. The fact that NUP85 shows strong CERES depletion (scores < –1) in many lineages supports its functional importance, yet therapeutic targeting would require validation in immunocompetent or patient-derived models where immune and stromal components are preserved. Fifth, while our results suggest NUP85’s impact on immune cell infiltration and patient prognosis, direct evidence for its influence on immune-related pathways and patient outcomes remains to be established. Finally, the absence of clinical trials utilizing anti-NUP85 therapeutic monoclonal antibodies precludes determining their efficacy in cancer models.
Conclusion
Our pioneering pan-cancer analysis of NUP85 has unveiled its association with multiple tumor markers, including immune cell infiltration, drug resistance, and immune markers. Future research will delve deeper into the specific mechanisms underlying NUP85’s tumor-immune interaction and facilitate the development and testing of novel anti-tumor drugs targeting NUP85.
Abbreviations
NUPs, Nucleoporins; TCGA, The Cancer Genome Atlas; GTEx, Genotype-Tissue Expression; CPTAC, Clinical Proteomic Tumor Analysis Consortium; CCLE, Cancer Cell Line Encyclopedia; HPA, Human Protein Atlas; GEPIA, Gene Expression Profiling Interactive Analysis; TMB, tumor mutational burden; MSI, microsatellite instability; CNV, copy number variation; LUAD, lung adenocarcinoma; OSCC, oral squamous cell carcinoma; CCR2, C-C Motif Chemokine Receptor 2; TIIC, tumor-infiltrating immune cells; OS, overall survival; DSS, disease specific survival; PFI, Progression free interval; HR, Hazard Ratio; DFS, disease-free survival; GSCA, Gene Set Cancer Analysis; TCGA-ACC, Adrenocortical carcinoma; TCGA-BLCA, Bladder Urothelial Carcinoma; TCGA-BRCA, Breast invasive carcinoma; TCGA-CESC, Cervical squamous cell carcinoma and endocervical adenocarcinoma; TCGA-CHOL, Cholangiocarcinoma; TCGA-COAD, Colon adenocarcinoma; TCGA-COADREAD, Colon adenocarcinoma/Rectum adenocarcinoma Esophageal carcinoma; TCGA-DLBC, Lymphoid Neoplasm Diffuse Large B-cell Lymphoma; TCGA-ESCA, Esophageal carcinoma; TCGA-FPPP, FFPE Pilot Phase II; TCGA-GBM, Glioblastoma multiforme; TCGA-GBMLGG, Glioma; TCGA-HNSC, Head and Neck squamous cell carcinoma; TCGA-KICH, Kidney Chromophobe; TCGA-KIPAN, Pan-kidney cohort (KICH+KIRC+KIRP); TCGA-KIRC, Kidney renal clear cell carcinoma; TCGA-KIRP, Kidney renal papillary cell carcinoma; TCGA-LAML, Acute Myeloid Leukemia; TCGA-LGG, Brain Lower Grade Glioma; TCGA-LIHC, Liver hepatocellular carcinoma; TCGA-LUAD, Lung adenocarcinoma; TCGA-LUSC, Lung squamous cell carcinoma; TCGA-MESO, Mesothelioma; TCGA-OV, Ovarian serous cystadenocarcinoma; TCGA-PAAD, Pancreatic adenocarcinoma; TCGA-PCPG, Pheochromocytoma and Paraganglioma; TCGA-PRAD, Prostate adenocarcinoma; TCGA-READ, Rectum adenocarcinoma; TCGA-SARC, Sarcoma; TCGA-STAD, Stomach adenocarcinoma; TCGA-SKCM, Skin Cutaneous Melanoma; TCGA-STES, Stomach and Esophageal carcinoma; TCGA-TGCT, Testicular Germ Cell Tumors; TCGA-THCA, Thyroid carcinoma; TCGA-THYM, Thymoma; TCGA-UCEC, Uterine Corpus Endometrial Carcinoma; TCGA-UCS, Uterine Carcinosarcoma; TCGA-UVM, Uveal Melanoma; TARGET-OS, Osteosarcoma; TARGET-ALL, Acute Lymphoblastic Leukemia; TARGET-NB, Neuroblastoma; TARGET-WT, High-Risk Wilms Tumor.
Data Sharing Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding author (Jianlin Zhang).
Ethics Statement
Our study was approved by the Ethics Committee of the First Affiliated Hospital of Anhui Medical University (PJ2025-11-10).
Acknowledgments
We would like to express our gratitude to the TCGA, GTEx, CPTAC, CCLE, GEPIA, Cellminer, GSCA, GeneMANIA, and BEST databases for providing data.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by The University Natural Science Research Project of Anhui Province (2023AH053322), and Basic and Clinical Collaboration Enhancement Program Foundation of Anhui Medical University (2023xkjT038).
Disclosure
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Oketch DJA, Giulietti M, Piva F. Copy number variations in pancreatic cancer: from biological significance to clinical utility. Int J Mol Sci. 2023;25(1):391. doi:10.3390/ijms25010391
2. El-Saadony MT, Alkafaas SS, Saad AM, et al. Medicinal plants: nutritional, immunological and therapeutic role in treating cancer-related malnutrition: a comprehensive review. Can Cell Inter. 2025;25(1):266. doi:10.1186/s12935-025-03720-2
3. Augimeri G, Bonofiglio D. The mediterranean diet as a source of natural compounds: does it represent a protective choice against cancer? Pharmaceuticals. 2021;14(9). doi:10.3390/ph14090920
4. Sonkin D, Thomas A, Teicher BA. Cancer treatments: past, present, and future. Cancer Genetics. 2024;286-287:18–19. doi:10.1016/j.cancergen.2024.06.002
5. Wang X, Wang L, Zhou L, et al. NUP62 alleviates senescence and promotes the stemness of human dental pulp stem cells via NSD2-dependent epigenetic reprogramming. Int J Oral Sci. 2025;17(1):34. doi:10.1038/s41368-025-00362-y
6. Khakwani M, Ji X-Y, Khattak S, Sun Y-C, Yao K, Zhang L. Targeting colorectal cancer at the level of nuclear pore complex. J Adv Res. 2025;70:423–444. doi:10.1016/j.jare.2024.06.009
7. Roy A, Narayan G. Oncogenic potential of nucleoporins in non-hematological cancers: recent update beyond chromosome translocation and gene fusion. J Cancer Res Clin Oncol. 2019;145(12):2901–2910. doi:10.1007/s00432-019-03063-2
8. Harel A, Orjalo AV, Vincent T, et al. Removal of a single pore subcomplex results in vertebrate nuclei devoid of nuclear pores. Molecular Cell. 2003;11(4):853–864. doi:10.1016/S1097-2765(03)00116-3
9. Terashima Y, Onai N, Murai M, et al. Pivotal function for cytoplasmic protein FROUNT in CCR2-mediated monocyte chemotaxis. Nat Immunol. 2005;6(8):827–835. doi:10.1038/ni1222
10. Satoh M, Akatsu T, Ishkawa Y, Minami Y, Nakamura M. A novel activator of C-C chemokine, FROUNT, is expressed with C-C chemokine receptor 2 and its ligand in failing human heart. J Card Fail. 2007;13(2):114–119. doi:10.1016/j.cardfail.2006.11.003
11. Belema-Bedada F, Uchida S, Martire A, Kostin S, Braun T. Efficient homing of multipotent adult mesenchymal stem cells depends on FROUNT-mediated clustering of CCR2. Cell Stem Cell. 2008;2(6):566–575. doi:10.1016/j.stem.2008.03.003
12. Toda E, Terashima Y, Sato T, Hirose K, Kanegasaki S, Matsushima K. FROUNT is a common regulator of CCR2 and CCR5 signaling to control directional migration. J Immunol. 2009;183(10):6387–6394. doi:10.4049/jimmunol.0803469
13. Ostuni R, Kratochvill F, Murray PJ, Natoli G. Macrophages and cancer: from mechanisms to therapeutic implications. Trends Immunol. 2015;36(4):229–239. doi:10.1016/j.it.2015.02.004
14. Terashima Y, Toda E, Itakura M, et al. Targeting FROUNT with disulfiram suppresses macrophage accumulation and its tumor-promoting properties. Nat Commun. 2020;11(1):609. doi:10.1038/s41467-020-14338-5
15. Taniuchi K, Yawata T, Tsuboi M, Ueba T, Saibara T. Efficient delivery of small interfering RNAs targeting particular mRNAs into pancreatic cancer cells inhibits invasiveness and metastasis of pancreatic tumors. Oncotarget. 2019;10(30):2869–2886. doi:10.18632/oncotarget.26880
16. Fernandez-Martinez J, Rout MP. One ring to rule them all? Structural and functional diversity in the nuclear pore complex. Trends Biochem Sci. 2021;46(7):595–607. doi:10.1016/j.tibs.2021.01.003
17. Beck M, Schirmacher P, Singer S. Alterations of the nuclear transport system in hepatocellular carcinoma – new basis for therapeutic strategies. J Hepatol. 2017;67(5):1051–1061. doi:10.1016/j.jhep.2017.06.021
18. Köhler A, Hurt E. Gene regulation by nucleoporins and links to cancer. Molecular Cell. 2010;38(1):6–15. doi:10.1016/j.molcel.2010.01.040
19. Raices M, D’Angelo MA. Nuclear pore complex composition: a new regulator of tissue-specific and developmental functions. Nat Rev Mol Cell Biol. 2012;13(11):687–699. doi:10.1038/nrm3461
20. Nofrini V, Di Giacomo D, Mecucci C. Nucleoporin genes in human diseases. Eur J Human Genetics. 2016;24(10):1388–1395. doi:10.1038/ejhg.2016.25
21. Xu S, Powers MA. Nuclear pore proteins and cancer. Semin Cell Dev Biol. 2009;20(5):620–630. doi:10.1016/j.semcdb.2009.03.003
22. Simon DN, Rout MP. Cancer and the nuclear pore complex. Adv Exp Med Biol. 2014;773:285–307.
23. Hazawa M, Lin D-C, Kobayashi A, et al. ROCK -dependent phosphorylation of NUP 62 regulates p63 nuclear transport and squamous cell carcinoma proliferation. EMBO Rep. 2018;19(1):73–88. doi:10.15252/embr.201744523
24. Bersini S, Lytle NK, Schulte R, Huang L, Wahl GM, Hetzer MW. Nup93 regulates breast tumor growth by modulating cell proliferation and actin cytoskeleton remodeling. Life Sci Alliance. 2020;3(1):e201900623. doi:10.26508/lsa.201900623
25. Makise M, Uchimura R, Higashi K, et al. Overexpression of the nucleoporin Nup88 stimulates migration and invasion of HeLa cells. Histochem Cell Biol. 2021;156(5):409–421. doi:10.1007/s00418-021-02020-w
26. Amin R, Shukla A, Zhu JJ, et al. Nuclear pore protein NUP210 depletion suppresses metastasis through heterochromatin-mediated disruption of tumor cell mechanical response. Nat Commun. 2021;12(1):7216. doi:10.1038/s41467-021-27451-w
27. Bhattacharjya S, Roy KS, Ganguly A, et al. Inhibition of nucleoporin member Nup214 expression by miR-133b perturbs mitotic timing and leads to cell death. Mol Cancer. 2015;14(1):42. doi:10.1186/s12943-015-0299-z
28. Sadoughi F, Mansournia MA, Mirhashemi SM. The potential role of chitosan-based nanoparticles as drug delivery systems in pancreatic cancer. IUBMB Life. 2020;72(5):872–883. doi:10.1002/iub.2252
29. Wang Y, Wang J, Zeng T, Qi J. Data-mining-based biomarker evaluation and experimental validation of SHTN1 for bladder cancer. Cancer Genetics. 2024;288-289:43–53. doi:10.1016/j.cancergen.2024.09.002
30. Gao X, Liu -D-D, Liu J-Z, Wang R. TCGA-based analysis of oncogenic signaling pathways underlying oral squamous cell carcinoma. Oncol Transl Med. 2024;10(2):87–92. doi:10.1097/ot9.0000000000000025
31. Li S, Yao J, Zhang S, et al. Prognostic value of tumor-microenvironment-associated genes in ovarian cancer. BIO Integration. 2023;4(3):84–96. doi:10.15212/bioi-2022-0008
32. Liu H, Li Y, Karsidag M, Tu T, Wang P. Technical and biological biases in bulk transcriptomic data mining for cancer research. J Cancer. 2025;16(1):34–43. doi:10.7150/jca.100922
33. Liu H, Guo Z, Wang P. Genetic expression in cancer research: challenges and complexity. Gene Rep. 2024;37:102042. doi:10.1016/j.genrep.2024.102042
34. Nair R, Mohan DD, Frank S, Setlur S, Govindaraju V, Ramanathan M. Generative adversarial networks for modelling clinical biomarker profiles with race/ethnicity. Br J Clin Pharmaco. 2023;89(5):1588–1600. doi:10.1111/bcp.15623
35. Ge Q, Huang X, Fang S, et al. Conditional generative adversarial networks for individualized treatment effect estimation and treatment selection. Front Genetics. 2020;11:585804. doi:10.3389/fgene.2020.585804
36. Chen DS, Mellman I. Elements of cancer immunity and the cancer-immune set point. Nature. 2017;541(7637):321–330.
37. Chiossone L, Dumas P-Y, Vienne M, Vivier E. Natural killer cells and other innate lymphoid cells in cancer. Nat Rev Immunol. 2018;18(11):671–688. doi:10.1038/s41577-018-0061-z
38. Xue M, Huang C, Jiang X, et al. Human bocavirus NP1 antagonizes host type I interferon response through repressing the nuclear transport of STAT1. Virulence. 2025;16(1):2570000. doi:10.1080/21505594.2025.2570000
39. Alhallaq AS, Sultan NS. Decoding NF-κB: nucleocytoplasmic shuttling dynamics, synthetic modulation and post-therapeutic behavior in cancer. Mole Biol Rep. 2025;52(1):804. doi:10.1007/s11033-025-10917-1
40. Anglim PP, Alonzo TA, Laird-Offringa IA. DNA methylation-based biomarkers for early detection of non-small cell lung cancer: an update. Mol Cancer. 2008;7(1):81. doi:10.1186/1476-4598-7-81
41. Baretti M, Le DT. DNA mismatch repair in cancer. Pharmacol Ther. 2018;189:45–62. doi:10.1016/j.pharmthera.2018.04.004
42. Chan TA, Yarchoan M, Jaffee E, et al. Development of tumor mutation burden as an immunotherapy biomarker: utility for the oncology clinic. Ann Oncol. 2019;30(1):44–56. doi:10.1093/annonc/mdy495
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.