Back to Journals » Journal of Inflammation Research » Volume 18
Nsun2 Knockdown Alleviates Asthma Progression by Inhibiting Hnrnpk Expression Through Decreased 5-Methylcytosine
Received 15 April 2025
Accepted for publication 9 August 2025
Published 21 August 2025 Volume 2025:18 Pages 11401—11413
DOI https://doi.org/10.2147/JIR.S531754
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Tara Strutt
Fen Wu, Jijing Zhao
Department of Respiratory, The First People’s Hospital of Jiande, Hangzhou, Zhejiang, People’s Republic of China
Correspondence: Fen Wu, Department of Respiratory, The First People’s Hospital of Jiande, No. 599 Yanzhou Avenue, Xin’anjiang Street, Hangzhou, Zhejiang, 311600, People’s Republic of China, Email [email protected]
Introduction: Asthma is a chronic inflammatory airway disease. 5-methylcytosine (m5C), an RNA modification, plays an unclear role in asthma pathogenesis. This study aimed to investigate the mechanism of m5C modification in asthma development.
Methods: Ovalbumin (OVA)-challenged BALB/c mice and PDGF-BB-stimulated airway smooth muscle cells (ASMCs) were used as in vivo and in vivo asthma models, respectively. Lung pathology was assessed through hematoxylin-eosin staining, inflammatory cell counts, and cytokine analysis in bronchoalveolar lavage fluid (BALF). Colony formation assays, ELISA, and transwell assays were conducted to evaluate cell proliferation, migration, invasion, and inflammatory responses in ASMCs. Mechanistic studies included methylated RNA immunoprecipitation (MeRIP), RNA immunoprecipitation (RIP), and dual-luciferase reporter assays.
Results: The asthma mouse model showed exacerbated lung injury and inflammation, accompanied by increased m5C levels and Nsun2 expression. Nsun2 knockdown suppressed PDGF-BB-induced ASMC proliferation, migration, invasion, and inflammatory responses through reduced Hnrnpk mRNA stability mediated by decreased m5C modification. Hnrnpk overexpression reversed the inhibitory effects of Nsun2 knockdown in PDGF-BB-treated ASMCs. Moreover, Nsun2 knockdown ameliorated airway inflammation and lung injury in asthmatic mice.
Conclusion: Nsun2 knockdown alleviated asthma by downregulating Hnrnpk expression through m5C-dependent regulation, providing novel mechanistic insights and potential therapeutic targets for asthma.
Keywords: asthma, m5C, Nsun2, Hnrnpk
Introduction
Asthma is characterized by airway inflammation and bronchoconstriction.1 Traditionally, asthma has been defined as a single disease entity based on its clinical phenotype. However, emerging evidence highlights that asthma constitutes a heterogeneous syndrome driven by distinct molecular endotypes. These endotypes elicit shared pathological features through divergent mechanisms, such as Th2-high and neutrophilic inflammation, among others.2,3 Approximately 300 million individuals worldwide suffer from asthma, with over 400,000 annual deaths attributed to this condition, making it a critical public health concern.4 Asthma development involves multiple factors, including genetic predisposition, environmental exposures, and immune system abnormalities.5 Genetic susceptibility combined with external triggers like allergens and air pollutants contributes to the onset and progression of chronic airway inflammation.6,7 The hallmark features of asthma include recurrent episodes of wheezing, coughing, chest tightness, and shortness of breath, which typically worsen at night or early morning and can be triggered by specific stimuli.8 Current asthma treatment primarily relies on pharmacological interventions and supportive care. Although existing treatment regimens effectively control symptoms in most patients, a significant proportion still fail to achieve satisfactory outcomes.9 Therefore, elucidating novel mechanisms underlying asthma pathophysiology is essential for developing more effective therapeutic strategies.
Epigenetic modifications refer to alterations in gene expression that occur without changes to the DNA sequence. These modifications respond to environmental cues and are heritable to daughter cells, playing critical roles in organismal development, cellular differentiation, and disease pathogenesis.10 5-methylcytosine (m5C) is a widespread RNA epigenetic modification present in tRNAs, rRNAs, and mRNAs. M5C modifications regulate gene expression through multiple mechanisms, including modulation of RNA structural stability, translational efficiency, and splicing dynamics.11 Specifically, m5C modifications exert regulatory effects by recruiting effector molecules via specific RNA-binding proteins, thereby modulating the functional fate of target RNAs. Moreover, these modifications are intricately linked to RNA editing processes, degradation pathways, and intracellular trafficking mechanisms, forming a complex regulatory network for RNA homeostasis.12,13 In recent years, the pathological significance of m5C modifications in human diseases has gained increasing attention. Numerous studies have demonstrated that dysregulated m5C modification levels contribute to various disease states, including malignant tumors and neurological disorders.14,15 For example, abnormal expression of m5C writers or erasers in specific cancer types can lead to oncogenic activation or tumor suppressor gene silencing, facilitating tumorigenesis.16,17 In neurological diseases, perturbations in m5C modification patterns may disrupt neuronal function, resulting in cognitive impairments or abnormal neurobehavioral manifestations.18 Despite accumulating evidence on m5C modifications in diverse pathophysiological contexts, their specific roles in asthma remain poorly understood. Further elucidation of m5C modification mechanisms in asthma may uncover novel therapeutic targets and drive innovative strategies for prevention and treatment.
In this study, we focused on a well-established experimental model of allergic asthma induced by ovalbumin (OVA) sensitization and challenge in rats, which represents a specific endotype involving Th2-driven immune responses. We investigated the mechanism by which m5C modification regulates asthma development through both in vivo and in vivo experiments. This study not only aims to deepen the understanding of asthma pathophysiology but also provides scientific evidence for developing innovative treatment strategies based on epigenetic regulation.
Methods
Animals
BALB/c mice (6- to 8-week-old, 18–22 g) were housed at 20–22°C under a 12-hour light/dark cycle. Mice were randomly divided into the control group, the asthma group, the asthma + shNC group, and the asthma + shNsun2 group, with six mice per group. Mice in the asthma + shNC group and the asthma + shNsun2 group were administered short hairpin RNA (shRNA) targeting Nsun2 (shNsun2) or a non-targeting shRNA negative control (shNC) via tail vein injection one day prior to asthma mouse model construction.
Establishment of Asthma Mouse Model
Asthma was induced in mice using OVA. Mice were sensitized by intraperitoneal injections of 100 μL of a saline solution containing 20 μg OVA and 2 mg aluminum hydroxide gel on days 0, 7, and 14. Subsequently, mice received daily nebulized inhalation of 5% OVA solution once a day for 30 minutes, for seven consecutive days. Mice in the control group received intraperitoneal injections of an equal volume of saline on the first 20 days and were exposed to aerosolized 0.9% NaCl from day 21 onward.
One hour following the final aerosol treatment, mice were euthanized by intraperitoneal administration of 1.5% sodium pentobarbital. The trachea and thoracic cavity were exposed by opening the thoracic cavity, and the trachea was ligated and cannulated. Bronchoalveolar lavage was performed using precooled PBS with three successive lavages. Bronchoalveolar lavage fluid (BALF) was collected, centrifuged, and the supernatant stored at −20°C for subsequent analysis. Subsequently, the mice were dissected, and the left lung tissue was removed and fixed in 4% paraformaldehyde for histopathological evaluation. The right lung tissue was harvested and stored at −80°C for future experimental use. All animal experiments adhered to the ARRIVE guidelines for reporting in vivo experiments.
Hematoxylin Eosin (HE) Staining
HE staining was performed to evaluate pathological changes in lung tissues using an HE staining kit (Beyotime, Shanghai, China). The lung tissues were fixed in 4% paraformaldehyde, dehydrated with graded ethanol, cleared in xylene, embedded in paraffin, and sectioned into 5-μm-thick slices. The tissue sections were stained with hematoxylin and eosin according to the manufacturer’s instructions. Following staining, the paraffin sections were mounted with neutral gum and examined under a light microscope.
BALF Analysis
The numbers of neutrophilic granulocytes, eosinophilic granulocytes, and lymphocytes among total cells in BALF were determined using a hemacytometer.
ELISA Assay
The levels of IL-1β, IL-6, TNF-α and MCP-1 in BALF were measured using corresponding ELISA kits (Beyotime). The assays were performed in accordance with the manufacturer’s instructions, and absorbance was read using a microplate reader.
AHR Assay
AHR was assessed using an animal pulmonary function instrument (Buxco Electronics). Mice were anesthetized with 1% sodium pentobarbital and then connected to a mechanical ventilator. Following stabilization, an ascending series of methacholine concentrations (3.125–50 mg/mL) was delivered via a nebulizer into the trachea. Baseline airway resistance was first measured using nebulized PBS. The resistance index (RI) of both total lung and airway was calculated according to the manufacturer’s instructions.
Dot Blot Assay
The m5C levels in mouse lung tissues were detected using a dot blot assay. Total RNA was extracted using Trizol reagent (Thermo Scientific, Waltham, MA, USA). Following denaturation, RNA samples were spotted onto Hybond-N⁺ membranes, which were then subjected to UV cross-linking for 20 min. The membranes were blocked with 5% skim milk solution at room temperature for 1 h and subsequently incubated overnight at 4 °C with an anti-m5C antibody. Afterward, the membranes were incubated with horseradish peroxidase-conjugated secondary antibody (1:10,000 dilution; ab6721, Abcam, Cambridge, MA, USA) for 1 h at room temperature. Following three washes with 1× TBST, specific signals were visualized using an ECL detection reagent (Beyotime). To normalize RNA loading, membranes were stained with 0.02% methylene blue (Sigma-Aldrich, St. Louis, MO, USA) in 0.3 M sodium acetate (pH 5.2) for 15 min as an internal control.
Quantitative Real-Time PCR (qPCR)
Total RNA was extracted from lung tissues and mouse airway smooth muscle cells (ASMCs) using Trizol reagent (Thermo Scientific, Waltham, MA, USA). RNA samples were reverse-transcribed into cDNA using the Hifair® Advanced Fast 1st Strand cDNA Synthesis Kit (Yeasen, Shanghai, China). qPCR was carried out using SYBR Green Master Mix (Vazyme, Nanjing, China) on a real-time PCR system. The relative mRNA expression levels were calculated using the 2−ΔΔCt method, with GAPDH serving as the internal reference gene. The sequences of the qPCR primers used in this study are listed below: Nsun2, 5′-AGGTGGCTATCCCGAGATCG-3’ (sense) and 5’-GACTCCATGAATTGGTCCCATT-3’ (antisense); Nsun3, 5’-CAATATGCCATCCTCTTCAACCG-3’ (sense) and 5’-AGGACTGTGTGATAGCCCCTC-3’ (antisense); Nsun4, 5’-TGGGATAGTGTGAGTGCTAAGC-3’ (sense) and 5’-AAGCATCGAAGATTTGGGCTG-3’ (antisense); Nsun5, 5’-ACCTGAAGCAGTTGTACGCTC-3’ (sense) and 5’-CCCCTTCCCCAGCAATAATTC-3’ (antisense); Nsun6, 5’-AAGACAACAGGGTGAAGTGATTG-3’ (sense) and 5’-TCCATCAAATTCTTTGGCTCCTT-3’ (antisense); Nsun7, 5’-TCTCAAGGTGGTCTACCGAAA-3’ (sense) and 5’-TTCATTGCGTGTGTTAGCTGT-3’ (antisense).
Cell Culture
ASMCs were isolated from mouse bronchi. The primary bronchi were dissected, cut into small tissue fragments, and incubated in a digestion solution containing 15 mg collagenase, 1 mg proteinase, 10 mg BSA, and 10 mg trypsin inhibitor. Following enzymatic digestion, the tissue samples were transferred to Hanks’ Ca²⁺-free solution and gently agitated for 20 min.19 The dissociated ASMCs were collected and cultured in RPMI-1640 medium (Gibco, Grand Island, NY, USA) supplemented with 10% FBS (Gibco) and 1% penicillin/streptomycin at 37 °C in a humidified atmosphere containing 5% CO2. To mimic airway remodeling under asthmatic conditions in vitro, ASMCs were stimulated with 50 ng/mL PDGF-BB for 24 h.
Cell Transfection
Hnrnpk overexpression plasmids (pcDNA3.1-Hnrnpk), empty vector (pcDNA3.1), shNsun2, and shNC were synthesized by GeneScript (Nanjing, China). ASMCs in the logarithmic growth phase were seeded into 6-well plates and transfected with the respective constructs using Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. Following a 48-hour transfection period, cells were harvested for further analysis.
Colony Formation Assay
The colony formation assay was performed to assess the proliferative capacity of ASMCs. Cells in the logarithmic growth phase were seeded into 6-well plates at a density of 500 cells per well and cultured in DMEM for 14 days. Following incubation, colonies were fixed with 4% paraformaldehyde for 15 min and stained with 0.1% crystal violet solution for 30 min. The number of visible colonies was then counted under a light microscope.
Transwell Assay
Cell migration and invasion were evaluated using Transwell chambers (Corning, NY, USA). For the migration assay, cells were seeded into the upper chamber and incubated for 24 h. Following incubation, cells that had migrated to the lower surface of the membrane were washed with PBS, fixed with 4% paraformaldehyde for 15 min, and stained with 0.1% crystal violet for 30 min. For the invasion assay, the upper surface of the Transwell membranes was pre-coated with a 1:6 dilution of Matrigel (Corning) to form a basement membrane barrier. Subsequently, cells resuspended in 100 μL of serum-free medium were added to the upper chamber, while 600 μL of complete medium was placed in the lower chamber as a chemoattractant. After incubation, non-migrated or non-invaded cells on the upper side of the membrane were removed, and the migrated or invaded cells were visualized under a microscope and quantified.
Bioinformatic Analysis
The GeneMANIA database (http://genemania.org) was utilized to predict potential downstream target genes regulated by Nsun2. The possible m5C modification sites within Hnrnpk were predicted using the RNA m5Cfinder database (http://www.rnanut.net/rnam5cfinder/).
Methylated RNA Immunoprecipitation (MeRIP)
MeRIP was performed to detect m5C methylation levels on Hnrnpk using an m5C MeRIP kit (Cloudseq, Shanghai, China). Total RNA was extracted from ASMCs and fragmented using a PCR thermocycler. PGM beads were pre-incubated with either anti-m5C antibody or control anti-IgG antibody for 1 h at room temperature. Following incubation, the beads were washed with IP buffer and then incubated with fragmented RNA samples for 1 h at 4 °C. Afterward, the RNA–bead complexes were washed again with IP buffer. Bound RNA was eluted using nuclease-free water and purified according to the manufacturer’s instructions. The enrichment of m5C on Hnrnpk was subsequently analyzed by qPCR.
RIP
The interaction between Nsun2 and Hnrnpk was evaluated using an RIP assay kit (Beyotime, Shanghai, China). Protein A/G agarose beads were washed with wash buffer and pre-incubated with anti-Nsun2, anti-Hnrnpk, or control anti-IgG antibody for 30 min at room temperature. ASMCs were lysed in lysis buffer on ice, and the cell lysates were centrifuged at 14,000 × g for 10 min at 4 °C to collect the supernatant. The supernatant was then incubated with the antibody-bound protein A/G agarose beads for 4 h at 4 °C under gentle rotation. Following immunoprecipitation, the RNA–protein complexes were eluted and purified according to the manufacturer’s instructions, and the enriched RNA was analyzed by qPCR.
Dual Luciferase Report
To assess the regulatory effect of m5C modification on Nsun2 expression, wild-type (WT) and mutant (Mut) Nsun2 sequences containing predicted m5C sites were cloned into the pGL3-basic vector to generate luciferase reporter plasmids. ASMCs were co-transfected with the luciferase reporter plasmids and either shNC (negative control) or shNsun2 using Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA), according to the manufacturer’s instructions. Luciferase activity was measured using a Dual-Luciferase Reporter Assay System (Promega, San Luis Obispo, CA, USA). Renilla luciferase was used as an internal control to normalize firefly luciferase signals.
RNA Stability Assay
Evaluation of Hnrnpk mRNA stability in ASMCs was detected by qPCR at 1, 4, 8, 12 h following 5 μg/mL actinomycin D (ACD; Merck, Darmstadt, Germany) treatment.
Statistical Analysis
All data were analyzed using SPSS 22.0 software. Results are expressed as mean ± standard deviation (SD) from at least three independent experiments. Comparisons between two groups were performed using Student’s t-test, while one-way analysis of variance (ANOVA) followed by Tukey’s post hoc test was used for comparisons among multiple groups. A value of P < 0.05 was considered statistically significant.
Results
OVA Induction Increases Lung Injury and Levels of Inflammatory Cytokines in Mice
Mice were administered OVA to induce asthma-like symptoms. Compared to the control group, histological analysis via HE staining revealed that mice in the asthma group showed significantly thickened alveolar walls and marked infiltration of inflammatory cells (Figure 1A). The numbers of eosinophilic granulocytes, neutrophilic granulocytes, and lymphocytes in BALF were significantly elevated in the asthma group (Figure 1B–D). Moreover, OVA exposure significantly increased the levels of IL-1β, IL-6, TNF-α, and MCP-1 in BALF of asthmatic mice (Figure 1E–H). Additionally, the RI in the asthma group after methacholine challenge was markedly higher than that in the control group (Figure 1I). Collectively, these results indicated that OVA-induced asthma exacerbated lung injury and inflammatory cytokine production in mice.
 |
Figure 1 OVA induction increased lung injury and levels of inflammatory cytokines in mice. (A) Lung injury of mice was evaluated by HE staining (20×). (B–D) The number of eosinophilic granulocytes, neutrophil granulocytes and lymphocytes in BALF of mice. (E–H) The levels of IL-1β, IL-6, TNF-α and MCP-1 in BALF of mice were measured using ELISA assay. (I) AHR of mice was evaluated by detection of lung RI. |
Nsun2 Knockdown Inhibits ASMCs Proliferation, Migration, Invasion and Inflammation
Subsequently, we examined the m5C levels in BALF of mice. We found that m5C modification was significantly elevated in the asthma model group compared to controls (Figure 2A). Furthermore, we assessed the expression levels of m5C-related enzymes and observed that only Nsun2 was upregulated in the asthma group (Figure 2B). To further investigate the functional role of Nsun2 in asthma, mouse ASMCs were treated with PDGF-BB to simulate airway remodeling in vitro. Our results showed that PDGF-BB treatment led to a significant upregulation of Nsun2 expression in ASMCs (Figure 3A). qPCR analysis confirmed that Nsun2 expression was effectively reduced following transfection with shNsun2 (Figure 3B). We found that PDGF-BB significantly promoted ASMC proliferation, an effect that was markedly suppressed by Nsun2 knockdown, as indicated by both colony formation and CCK-8 assays (Figure 3C and D). Moreover, Nsun2 silencing significantly attenuated the PDGF-BB-induced increases in IL-1β, IL-6, and TNF-α levels in ASMCs (Figure 3E–G). In addition, Transwell assays revealed that PDGF-BB enhanced the migratory and invasive capacities of ASMCs, which were significantly inhibited by Nsun2 knockdown (Figure 3H–K). Collectively, these findings indicate that Nsun2 and m5C modification are upregulated in the asthma mouse model, and that Nsun2 knockdown suppresses ASMC proliferation, migration, invasion, and inflammatory cytokine production.
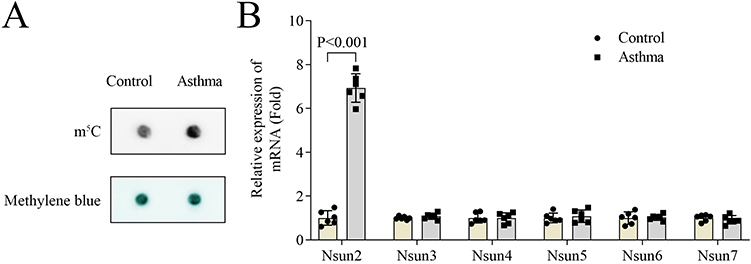 |
Figure 2 m5C level and Nsun2 expression were increased in asthma mouse model. (A) m5C levels of lung tissues were detected by dot blot assay. (B) qPCR was performed to measure the expression of m5C-related enzymes in lung tissues of mice. |
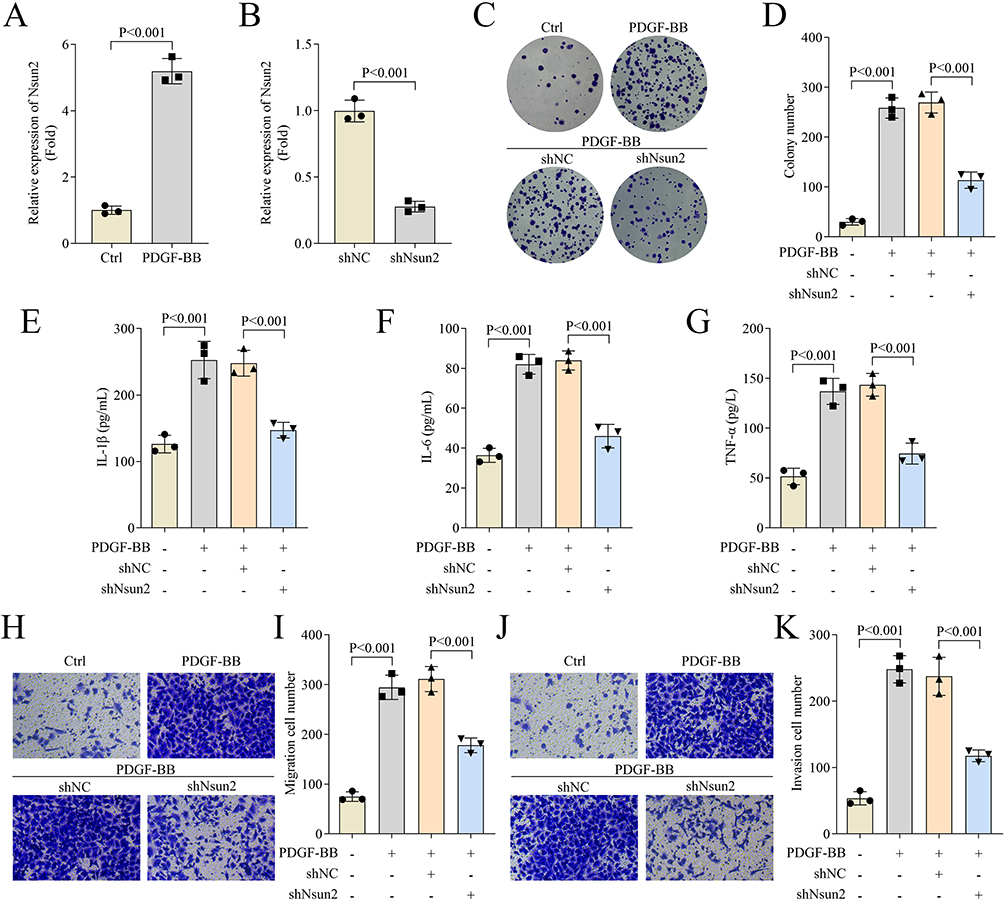 |
Figure 3 Nsun2 knockdown inhibited ASMCs proliferation, migration, invasion and inflammation. (A and B) Nsun2 expression of ASMCs was measured by qPCR. (C and D) Cell proliferation of ASMCs was evaluated by colony formation assay. (E–G) The levels of IL-1β, IL-6 and TNF-α in ASMCs were detected by ELISA assay. (H–K) Cell migration and invasion of ASMCs were evaluated by transwell assay. The magnification of Figures H and J is 10×. |
Nsun2 Knockdown Suppresses the Stability of Hnrnpk by Decreasing Hnrnpk m5C Modification
Next, we explored the mechanism by which Nsun2 mediates asthma progression. Bioinformatic analysis predicted several downstream target genes regulated by Nsun2, as shown in Figure 4A, among which Hnrnpk was identified as a potential candidate. Further investigation revealed that Nsun2 knockdown significantly reduced both the expression level and m5C modification level of Hnrnpk (Figure 4B and C). RIP assay confirmed a direct interaction between Nsun2 and Hnrnpk, suggesting that Hnrnpk may be a functional target of Nsun2 (Figure 4D). Based on these findings, we predicted several potential m5C modification sites within Hnrnpk. The top two candidate sites were selected and designated as site 1# and site #2 (Figure 4E). Dual-luciferase reporter assays showed that Nsun2 knockdown significantly decreased the luciferase activity of the WT site 1#, but had no effect on WT site #2 (Figure 4F). Moreover, RNA stability assays demonstrated that Nsun2 knockdown significantly reduced the stability of Hnrnpk mRNA (Figure 4G). Taken together, these results indicate that Nsun2 knockdown suppresses Hnrnpk mRNA stability through the reduction of its m5C modification.
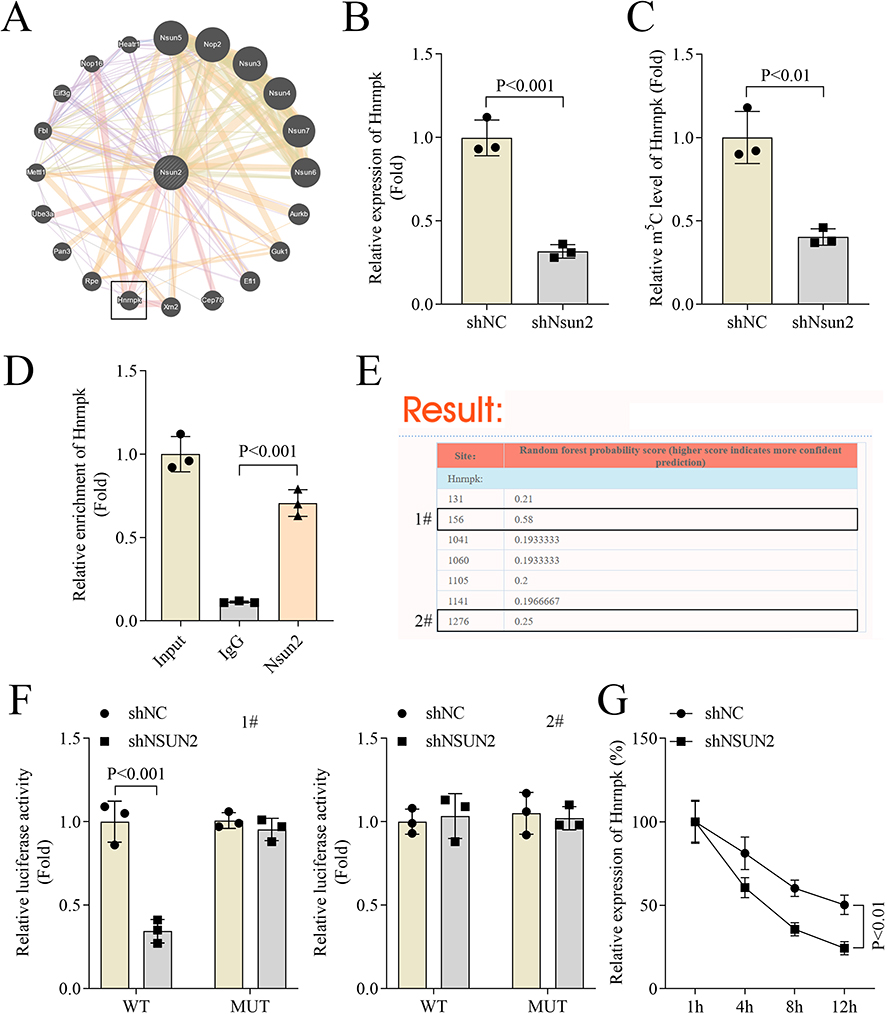 |
Figure 4 Nsun2 knockdown suppressed the stability of Hnrnpk by decreasing Hnrnpk m5C modification. (A) GENEMANIA database was performed to predict the genes regulated by Nsun2. (B) Hnrnpkexpression of ASMCs was measured by qPCR. (C) m5C levels of Hnrnpk was measured by MeRIP. (D) The interaction between Nsun2 and Hnrnpk was investigated by RIP. (E) Prediction of the potential m5C sites of Hnrnpk were preformed using the RNAm5Cfinder database. (F) The luciferase activity of Hnrnpk was measured by dual luciferase activity. (G) qPCR was performed to evaluate the stability of Hnrnpk mRNA in ASMCs at 1, 4, 8 and 12 h following ACD treatment. 1# and 2# represented two m5C methylation sites of Hnrnpk. |
Hnrnpk Overexpression Restores Cell Proliferation, Migration, Invasion and Inflammation Inhibited by Nsun2 Knockdown in PDGF-BB-Treated-ASMCs
Subsequently, we investigated the functional role of Hnrnpk in asthma through rescue experiments. Results showed that Hnrnpk overexpression significantly increased Hnrnpk mRNA expression in ASMCs (Figure 5A). Notably, Hnrnpk overexpression partially reversed the inhibitory effects of Nsun2 knockdown on cell proliferation in PDGF-BB-treated ASMCs, as shown by colony formation and CCK-8 assays (Figure 5B and C). Moreover, the PDGF-BB-treated ASMCs exhibited reduced levels of inflammatory cytokines IL-1β, IL-6, and TNF-α following Nsun2 knockdown, and these levels were partially restored upon Hnrnpk overexpression (Figure 5D–E). In addition, Transwell assays demonstrated that Hnrnpk overexpression enhanced the migratory and invasive capacities of PDGF-BB-treated ASMCs that had been suppressed by Nsun2 knockdown (Figure 5G–J). Collectively, these findings indicate that Hnrnpk overexpression effectively rescues cell proliferation, migration, invasion, and inflammatory responses that are inhibited by Nsun2 knockdown in PDGF-BB-treated ASMCs.
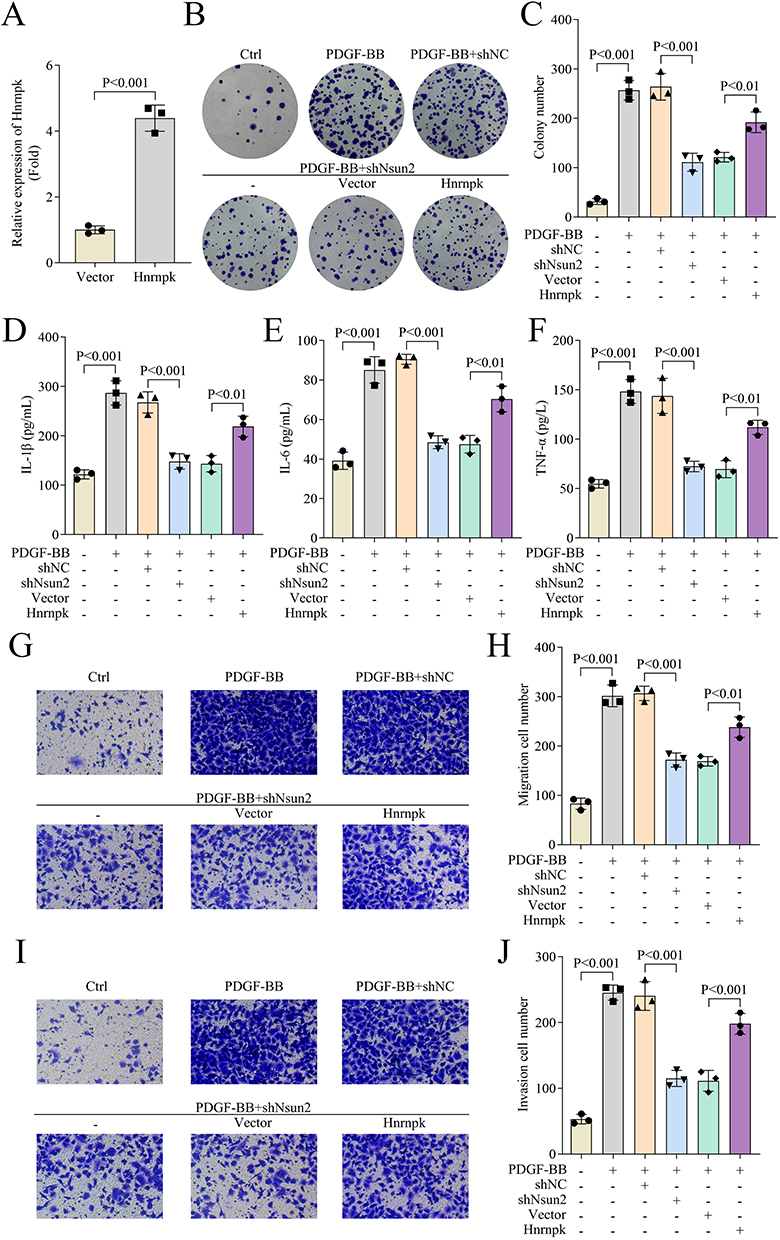 |
Figure 5 Hnrnpk overexpression restored cell proliferation, migration, invasion and inflammation inhibited by Nsun2 knockdown in PDGF-BB-treated-ASMCs. (A) The expression of Hnrnpk in ASMCs was measured by qPCR. (B and C) Cell proliferation of ASMCs was evaluated by colony formation assay. (D–F) The levels of IL-1β, IL-6 and TNF-α in ASMCs were detected by ELISA assay. (G–J) Cell migration and invasion of ASMCs were evaluated by transwell assay. The magnification of Figures G and I is 10×. |
Nsun2 Knockdown Improves Lung Injury and Decreases the Levels of Inflammatory Cytokines in Asthma Mouse Model
Finally, the in vivo role of Nsun2 in asthma was evaluated using an established asthma mouse model. Histological analysis by HE staining revealed that Nsun2 knockdown significantly alleviated bronchial inflammatory infiltration and reduced alveolar wall thickening compared to the asthma group (Figure 6A). Compared to the asthma group, Nsun2 knockdown markedly decreased the numbers of eosinophilic granulocytes, neutrophilic granulocytes, and lymphocytes in BALF (Figure 6B–D). Furthermore, the elevated levels of inflammatory cytokines IL-1β, IL-6, TNF-α, and MCP-1 in BALF from asthmatic mice were significantly reduced following Nsun2 knockdown (Figure 6E–H). In addition, the increased RI observed in the asthma group after methacholine challenge was notably attenuated by Nsun2 knockdown (Figure 6I). Collectively, these findings demonstrate that Nsun2 knockdown ameliorates lung injury and suppresses inflammatory cytokine production in the asthma mouse model.
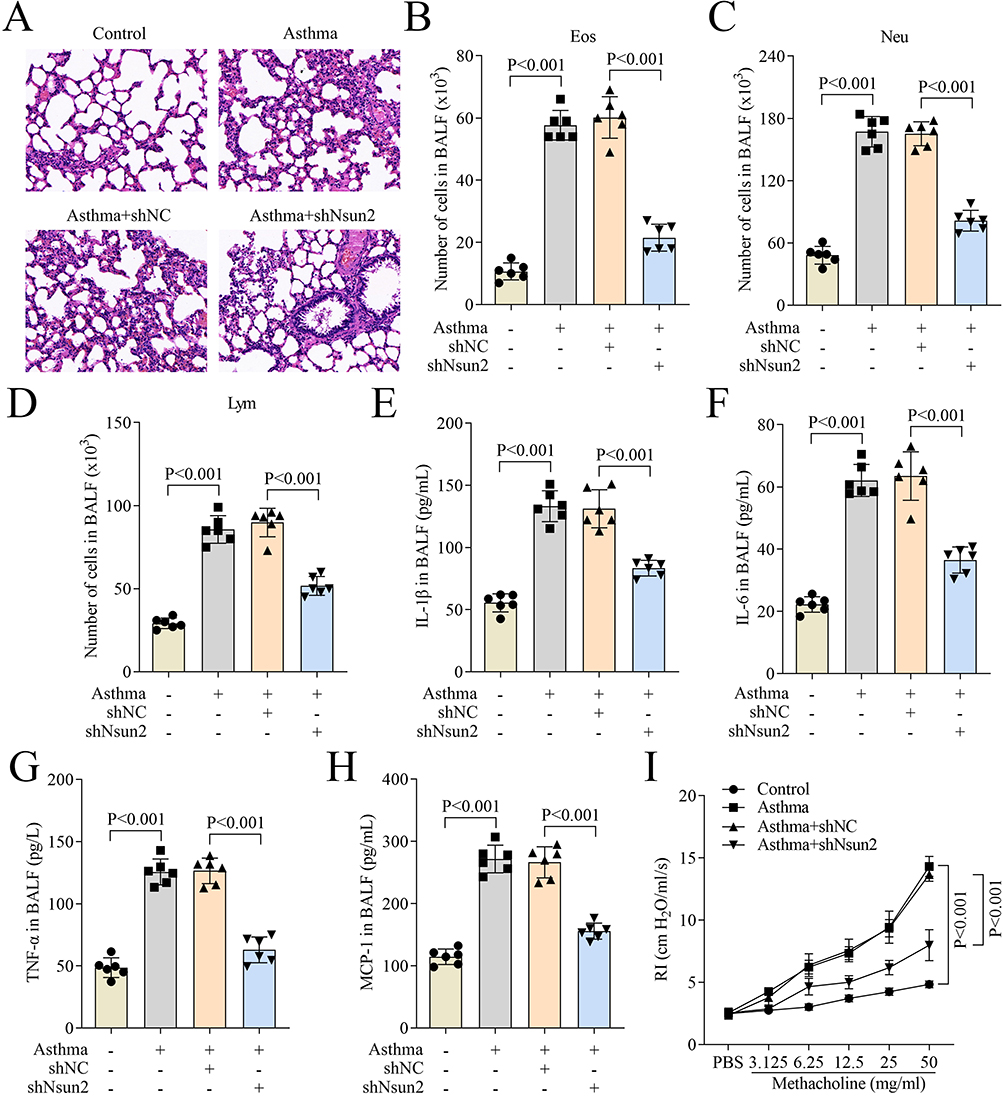 |
Figure 6 Nsun2 knockdown improved lung injury and decreases the levels of inflammatory cytokines in asthma mouse. (A) Lung injury of mice was evaluated by HE staining (20×). (B–D) The number of eosinophilic granulocytes, neutrophil granulocytes and lymphocytes in BALF of mice. (E–H) The levels of IL-1β, IL-6, TNF-α and MCP-1 in BALF of mice were measured using ELISA assay. (I) AHR of mice was evaluated by detection of lung RI. |
Discussion
m5C modification is a novel RNA modification that regulates gene expression by modulating RNA stability and translation efficiency, thereby playing a critical role in diverse biological processes.20 Although m5C modification has been implicated in the progression of multiple diseases, its mechanisms of action in asthma remain poorly understood. In this study, we demonstrated that m5C modification and Nsun2 expression were significantly elevated in the asthma mouse model, suggesting that elevated levels of m5C and Nsun2 may contribute to asthma pathogenesis. These findings offer novel insights into the potential pathogenic role of m5C modification in asthma.
In our current study, we observed increased inflammatory infiltration in lung tissues, as well as elevated numbers of eosinophilic granulocytes, neutrophil granulocytes, and lymphocytes in the BALF of the asthma mouse model. Airway inflammation represents one of the most prominent pathological features of asthma and, over time, contributes to the development of airway remodeling.21 The accumulation of eosinophils, neutrophils, and lymphocytes can exacerbate airway inflammation and is closely associated with disease severity and the frequency of asthma exacerbations.22 A similar pattern of airway inflammation, accompanied by increased eosinophils and basophils, is also observed in allergic rhinitis, an allergen-induced inflammatory disorder.23 Notably, Nsun2 knockdown effectively alleviated both airway inflammation and inflammatory cell infiltration in the asthma mouse model. In patients with asthma, PDGF-BB has been shown to induce ASMC proliferation, which contributes to airway remodeling.24 In our study, we found that Nsun2 knockdown significantly inhibited PDGF-BB-induced ASMC proliferation, migration, invasion, and inflammatory cytokine production. These findings suggest that Nsun2 deficiency exerts anti-inflammatory effects in asthma. Collectively, our results imply that targeting Nsun2 expression may represent a promising therapeutic strategy for asthma. Nsun2 is a key m5C methyltransferase that is frequently overexpressed in various cancers, where it promotes tumor progression, as seen in colorectal cancer and osteosarcoma.25,26 However, research on other functional roles of Nsun2 remains limited. Notably, Luo et al27 first demonstrated that Nsun2 deficiency protects vascular endothelial cells from inflammatory injury. Moreover, Miao et al28 revealed that Nsun2 mediates the progression of hyperhomocysteinemia-aggravated abdominal aortic aneurysm (AAA), promoting inflammatory cell infiltration in this context. These studies indicate that Nsun2 may function as a pro-inflammatory regulator; however, the precise mechanisms by which Nsun2 modulates inflammatory responses remain to be fully elucidated.
Additionally, our findings revealed that Nsun2 knockdown suppressed Hnrnpk expression by reducing m5C modification on Hnrnpk. Hnrnpk is an RNA-binding protein belonging to the heterogeneous nuclear ribonucleoprotein family and plays essential roles in multiple stages of gene expression, including transcription, RNA splicing, mRNA stability regulation, and translation.29 Although Hnrnpk has been extensively studied in cancer research—where it is overexpressed in various malignancies and associated with tumorigenesis—its role in asthma remains poorly understood. There is growing evidence that Hnrnpk regulates malignant phenotypes such as invasiveness, proliferation, and clonogenicity.30 For instance, Zhu et al31 demonstrated that the interaction between PROX1 and Hnrnpk inhibits Hnrnpk ubiquitination, thereby activating the WNT signaling pathway to promote breast cancer invasion and metastasis. Moreover, Hnrnpk has been shown to facilitate NLRP3 inflammasome activation in Raw264.7 macrophages, suggesting its potential as a therapeutic target for chronic kidney disease.32 Recently, Wang et al33 constructed a ceRNA network related to severe eosinophilic asthma and identified Hnrnpk as a potential regulator of this condition. However, the precise mechanisms by which Hnrnpk contributes to asthma pathogenesis remain to be fully elucidated. In our study, we demonstrated that Nsun2 modulates Hnrnpk expression in ASMCs through m5C modification. Furthermore, Hnrnpk overexpression partially reversed the suppression of cell proliferation, migration, invasion, and inflammatory cytokine production induced by Nsun2 knockdown in PDGF-BB-treated ASMCs. These results highlight a functional link between Nsun2-mediated m5C modification and Hnrnpk activity in asthma and suggest that targeting this regulatory axis may offer novel therapeutic strategies for asthma management.
Despite the promising findings of this study, several limitations must be acknowledged. The mechanistic insights into Nsun2-mediated m5C modification were primarily derived from in vitro cell models and an OVA-induced mouse model of asthma, which may not fully replicate the complexity and heterogeneity of human asthma pathophysiology. Moreover, although Hnrnpk was identified as a downstream target of Nsun2, the detailed molecular pathways linking Hnrnpk to ASMC function and inflammation remain to be further elucidated. Future studies should validate the clinical relevance of this regulatory axis by examining Nsun2/Hnrnpk expression and m5C profiles in human asthmatic airways. Additionally, the therapeutic potential of pharmacological Nsun2 inhibition should be explored across diverse asthma endotypes to assess its translational applicability.
Taken together, our findings demonstrate that Nsun2 knockdown alleviates asthma progression by downregulating Hnrnpk through m5C modification reduction. These results not only identify Nsun2 and Hnrnpk as potential molecular targets for precision therapy in asthma but also provide mechanistic insights into the role of RNA epigenetic modifications in Th2-driven airway inflammatory diseases. Furthermore, this regulatory axis may represent a novel therapeutic strategy for allergic rhinitis and other related disorders characterized by chronic Th2-mediated inflammation.
Data Sharing Statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Ethical Approval
This study was approved by the Ethics Committee of MDKN Biotechnology Co., Lt. (MDKN-2024-310). All animal experiments comply with the ARRIVE guidelines. All animal experiments comply with the National Standards for Laboratory Animal Welfare issued by the Chinese government (GB/T 35892-2018) and the Guide for the Care and Use of Laboratory Animals (National Research Council, 8th Edition, 2011).
Funding
The authors declare that no funds, grants, or other support were received during the preparation of this manuscript.
Disclosure
The authors have no relevant financial or non-financial interests to disclose for this work.
References
1. Habib N, Pasha MA, Tang DD. Current understanding of asthma pathogenesis and biomarkers. Cells. 2022;11(17):2764. doi:10.3390/cells11172764
2. Maspero J, Adir Y, Al-Ahmad M, et al. Type 2 inflammation in asthma and other airway diseases. ERJ Open Res. 2022;8(3):00576–2021. doi:10.1183/23120541.00576-2021
3. Ray A, Kolls JK. Neutrophilic inflammation in asthma and association with disease severity. Trends Immunol. 2017;38(12):942–954. doi:10.1016/j.it.2017.07.003
4. Kuruvilla ME, Vanijcharoenkarn K, Shih JA, et al. Epidemiology and risk factors for asthma. Respir Med. 2019;149:16–22. doi:10.1016/j.rmed.2019.01.014
5. Schoettler N, Strek ME. Recent advances in severe asthma: from phenotypes to personalized medicine. Chest. 2020;157(3):516–528. doi:10.1016/j.chest.2019.10.009
6. Komlosi ZI, van de Veen W, Kovács N, et al. Cellular and molecular mechanisms of allergic asthma. Mol Aspects Med. 2022;85:100995. doi:10.1016/j.mam.2021.100995
7. Aghapour M, Ubags ND, Bruder D, et al. Role of air pollutants in airway epithelial barrier dysfunction in asthma and COPD. Eur Respir Rev. 2022;31(163):210112. doi:10.1183/16000617.0112-2021
8. Chung KF, Dixey P, Abubakar-Waziri H, et al. Characteristics, phenotypes, mechanisms and management of severe asthma. Chin Med J. 2022;135(10):1141–1155. doi:10.1097/CM9.0000000000001990
9. Thomas D, McDonald VM, Pavord ID, et al. Asthma remission: what is it and how can it be achieved? Eur Respir J. 2022;60(5):2102583. doi:10.1183/13993003.02583-2021
10. Fitz-James MH, Cavalli G. Molecular mechanisms of transgenerational epigenetic inheritance. Nat Rev Genet. 2022;23(6):325–341. doi:10.1038/s41576-021-00438-5
11. Bohnsack KE, Hobartner C, Bohnsack MT. Eukaryotic 5-methylcytosine (m(5)C) RNA methyltransferases: mechanisms, cellular functions, and links to disease. Genes. 2019;10(2):102. doi:10.3390/genes10020102
12. Chen D, Gu X, Nurzat Y, et al. Writers, readers, and erasers RNA modifications and drug resistance in cancer. Mol Cancer. 2024;23(1):178. doi:10.1186/s12943-024-02089-6
13. Liu WW, Zheng S-Q, Li T, et al. RNA modifications in cellular metabolism: implications for metabolism-targeted therapy and immunotherapy. Signal Transduct Target Ther. 2024;9(1):70. doi:10.1038/s41392-024-01777-5
14. Li M, Tao Z, Zhao Y, et al. 5-methylcytosine RNA methyltransferases and their potential roles in cancer. J Transl Med. 2022;20(1):214. doi:10.1186/s12967-022-03427-2
15. Teng S, Han C, Zhou J, et al. m(5)C RNA methylation: a potential mechanism for infectious Alzheimer’s disease. Front Cell Dev Biol. 2024;12:1440143. doi:10.3389/fcell.2024.1440143
16. Hu Y, Chen C, Tong X, et al. NSUN2 modified by SUMO-2/3 promotes gastric cancer progression and regulates mRNA m5C methylation. Cell Death Dis. 2021;12(9):842. doi:10.1038/s41419-021-04127-3
17. Yu G, Bao J, Zhan M, et al. Comprehensive analysis of m5C methylation regulatory genes and tumor microenvironment in prostate cancer. Front Immunol. 2022;13:914577. doi:10.3389/fimmu.2022.914577
18. Lu Y, Yang L, Feng Q, et al. RNA 5-methylcytosine modification: regulatory molecules, biological functions, and human diseases. Genomics Proteomics Bioinf. 2024;22(5). doi:10.1093/gpbjnl/qzae063
19. Matthews RM, Bradley E, Hollywood MA, et al. Modulation of fast sodium current in airway smooth muscle cells by exchange protein directly activated by cAMP. Am J Physiol Cell Physiol. 2024;326(1):C1–C9. doi:10.1152/ajpcell.00417.2023
20. Gu X, Ma X, Chen C, et al. Vital roles of m(5)C RNA modification in cancer and immune cell biology. Front Immunol. 2023;14:1207371. doi:10.3389/fimmu.2023.1207371
21. Russell RJ, Boulet L-P, Brightling CE, et al. The airway epithelium: an orchestrator of inflammation, a key structural barrier and a therapeutic target in severe asthma. Eur Respir J. 2024;63(4):2301397. doi:10.1183/13993003.01397-2023
22. Gauthier M, Kale SL, Oriss TB, et al. CCL5 is a potential bridge between type 1 and type 2 inflammation in asthma. J Allergy Clin Immunol. 2023;152(1):94–106.e12. doi:10.1016/j.jaci.2023.02.028
23. Cergan R, Berghi O, Dumitru M, et al. Interleukin 8 molecular interplay in allergic rhinitis and chronic rhinosinusitis with nasal polyps: a scoping review. Life. 2025;15(3):469. doi:10.3390/life15030469
24. Listyoko AS, Okazaki R, Harada T, et al. beta-tocotrienol decreases PDGF-BB-induced proliferation and migration of human airway smooth muscle cells by inhibiting RhoA and reducing ROS production. Pharmaceuticals. 2024;17(6):712. doi:10.3390/ph17060712
25. Chen B, Deng Y, Hong Y, et al. Metabolic recoding of NSUN2-mediated m(5)Cm 5 C modification promotes the progression of colorectal cancer via the NSUN2/YBX1/m(5)C-ENO1NSUN2/YBX1/m 5 C-ENO1 positive feedback loop. Adv Sci. 2024;11(28):e2309840. doi:10.1002/advs.202309840
26. Yang M, Wei R, Zhang S, et al. NSUN2 promotes osteosarcoma progression by enhancing the stability of FABP5 mRNA via m(5)C methylation. Cell Death Dis. 2023;14(2):125. doi:10.1038/s41419-023-05646-x
27. Luo Y, Feng J, Xu Q, et al. NSun2 Deficiency Protects Endothelium From Inflammation via mRNA Methylation of ICAM-1. Circ Res. 2016;118(6):944–956. doi:10.1161/CIRCRESAHA.115.307674
28. Miao Y, Zhao Y, Han L, et al. NSun2 regulates aneurysm formation by promoting autotaxin expression and T cell recruitment. Cell Mol Life Sci. 2021;78(4):1709–1727. doi:10.1007/s00018-020-03607-7
29. Li Y, Wang H, Wan J, et al. The hnRNPK/A1/R/U complex regulates gene transcription and translation and is a favorable prognostic biomarker for human colorectal adenocarcinoma. Front Oncol. 2022;12:845931. doi:10.3389/fonc.2022.845931
30. Lee WJ, Shin CH, Ji H, et al. hnRNPK-regulated LINC00263 promotes malignant phenotypes through miR-147a/CAPN2. Cell Death Dis. 2021;12(4):290. doi:10.1038/s41419-021-03575-1
31. Zhu L, Tian Q, Gao H, et al. PROX1 promotes breast cancer invasion and metastasis through WNT/beta-catenin pathway via interacting with hnRNPK. Int J Biol Sci. 2022;18(5):2032–2046. doi:10.7150/ijbs.68960
32. Feng J, Li H, Li J, et al. hnRNPK knockdown alleviates NLRP3 inflammasome priming by repressing FLIP expression in Raw264.7 macrophages. Redox Rep. 2020;25(1):104–111. doi:10.1080/13510002.2020.1857157
33. Wang H, Zhang Z, Ma Y, et al. Construction of severe eosinophilic asthma related competing endogenous rna network by weighted gene co-expression network analysis. Front Pharmacol. 2022;13:852536. doi:10.3389/fphar.2022.852536
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.