Back to Journals » Cancer Management and Research » Volume 12
NSD2 Promotes Renal Cancer Progression Through Stimulating Akt/Erk Signaling
Authors Han X ![]() , Piao L, Xu X, Luo F, Liu Z, He X
, Piao L, Xu X, Luo F, Liu Z, He X
Received 16 July 2019
Accepted for publication 6 November 2019
Published 16 January 2020 Volume 2020:12 Pages 375—383
DOI https://doi.org/10.2147/CMAR.S222673
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Eileen O'Reilly
Xu Han, 1,* Lianhua Piao, 2,* Xiaoshuang Xu, 2 Fengbao Luo, 1 Zhiwei Liu, 3 Xiaozhou He 1
1Department of Urology, The Third Affiliated Hospital of Soochow University, Changzhou 213003, People’s Republic of China; 2Institute of Bioinformatics and Medical Engineering, Jiangsu University of Technology, Changzhou 213001, People’s Republic of China; 3Department of Orthopaedics, The Third Affiliated Hospital of Soochow University, Changzhou 213003, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Xiaozhou He
Department of Urology, The Third Affiliated Hospital of Soochow University, 185 Juqian Street, Changzhou 213003, Jiangsu Province, People’s Republic of China
Tel +86 519 6887 1251
Email [email protected]
Background: Nuclear receptor suppressor of variegation, enhancer of zeste, and trithorax (SET) domain-containing 2 (NSD2), is a well-known histone lysine methyltransferase (HMTase). The aim of this study was to investigate the biological role of NSD2 in clear cell renal cell carcinoma (ccRCC).
Methods: GEO and OncoLnc databases were used to identify NSD2 expression and estimate its clinical value in ccRCC. Immunohistochemistry (IHC) was applied to further evaluate NSD2 protein level in ccRCC tissues. The expression of NSD2 in different cell lines and the transfection efficiency were determined by quantitative real-time PCR and Western blot analysis. The effect of NSD2 and the underlying mechanism in ccRCC progression were investigated via MTT, flow cytometry, Western blotting and xenograft tumor assays.
Results: NSD2 was over-expressed in both ccRCC tissues and cell lines. NSD2 expression could discriminate ccRCC samples from normal samples, and moreover, high NSD2 expression was characterized with a short overall survival (OS) time. Additionally, knockdown of NSD2 suppressed proliferation and induced apoptosis of cancer cells by inhibiting Akt/Erk signaling and regulating Bcl-2 and Bax expression. Meanwhile, up-regulation of NSD2 contributed to the opposite effects. Silencing of NSD2 reduced xenograft tumor growth in vivo.
Conclusion: NSD2 serves as an oncogenic factor in the progression of ccRCC via activation of Akt/Erk signaling.
Keywords: NSD2, renal cancer, proliferation, apoptosis, Akt/Erk signaling
Introduction
The incidence of kidney and renal pelvis cancer is increasing in the United States, with 65,340 new cases and 14,970 deaths estimated for 2018.1 It is one of top 10 common cancers and ranks the third most prevalent genitourinary cancers. Relying on advanced imaging techniques, the rising number of incidental small renal cancers can be precisely diagnosed in the early stages.2 Currently, the most effective therapeutic method is radical nephrectomy, but clinical prognosis is still less than satisfactory.3 Clear cell renal cell carcinoma (ccRCC) is the most common histological subtype of renal cancers, which is often characterized by 3p loss that harbors mutational inactivation or hypermethylation of the tumor suppressor gene VHL.4 Since ccRCC is not sensitive to conventional radiotherapy and chemotherapy, novel molecular therapeutic targets are required to be applied to clinical therapeutics.
The NSD protein family consisted of three members and each protein contained one conserved SET [Su(var)3–9, Enhancer-of-zeste, Trithorax] domain, several PWWP (proline-tryptophan-tryptophan-proline) and PHD (plant homeo domain) domains.5 Over the past decades, the protein members have been reported to be involved in chromatin integrity and gene expression through mono- or di-methylating lysine 36 of histone H3 (H3K36).6–8 Besides, they were implicated in multiple cellular processes such as cell proliferation, apoptosis and invasion. Especially, NSD2 (WHSC1/MMSET) has been a vital regulator in cancer progression. NSD2 was over-expressed in 10%~20% multiple myeloma (MM) cases.9–11 Knockdown of NSD2 in t(4;14)+ MM cells could reduce cell proliferation, inhibit cell-cycle progression and DNA repair, meanwhile induce cell apoptosis.12–16 In prostate cancer, NSD2 was served as downstream of EZH2 (enhancer of zeste homolog 2) and the oncogenic functions of EZH2 required NSD2 activity.17 NSD2 may mediate NF-κB signaling for cell proliferation and survival via direct interaction with NF-κB.18 At present, the biological function of NSD2 in ccRCC progression remains unknown.
In the present study, we found that the expression of NSD2 was elevated both in ccRCC tissues and cell lines. Furthermore, NSD2 might be a promising biomarker applied to clinical diagnosis and prognosis analysis. In addition, NSD2 promoted cell proliferation and suppressed cell apoptosis through motivating Akt/Erk signaling, accompanied with modulating Bcl-2 and Bax expression. Depletion of NSD2 reduced xenograft tumor growth in vivo. Altogether, the findings indicated that NSD2 played an important role in facilitating ccRCC tumorigenesis.
Materials and Methods
GEO and OncoLnc Databases Reanalysis
The original NSD2 expression data was retrieved from GEO database (von Roemeling Renal, http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE53757). The expression of NSD2 mRNA between normal and ccRCC tissues was reanalyzed. ROC curve and AUC were performed to evaluate the sensitivity and specificity of NSD2 expression to diagnose ccRCC. Besides, the original survival and expression data were retrieved from OncoLnc and the overall survival (OS) time was reanalyzed.
Tissue Samples and Immunohistochemistry (IHC)
Between July 2018 and December 2018, a total of 30 specimens from 30 patients who had never accepted chemotherapy, radiotherapy or biotherapy were collected in the Department of Urology, the Third Affiliated Hospital of Soochow University. The patients were informed of the study and asked to sign informed consent. Application of tissues was approved by the Ethics Committee of the Third Affiliated Hospital of Soochow University. The specimens were pathologically confirmed as ccRCC according to the WHO for the classification criteria.
The NSD2 protein level in ccRCC and adjacent normal tissues were evaluated by IHC. The paraffin-embedded specimens were deparaffinized, rehydrated, and treated in antigen retrieval buffer at 96°C for 20 mins. Then, the samples were incubated with anti-NSD2 (Abcam, dilution: 1:200) primary antibody at room temperature for 1 hr. The DAB kit (MXB Biotechnologies, China) was used for detecting immune complexes. One-renal cancer pathologist performed semi-quantitative analysis of NSD2 staining using a 4-grade scale was defined as follows: Sections with no labeling or labeled cells <5% were scored as 0. Sections with 5–30% of labeled cells were scored as +1, with 31–70% of labeled cells as +2, and with >70% of labeled cells as +3.
Cell Lines and Cell Culture
The human normal cortex/proximal tubule epithelial cell line (HK-2) and three ccRCC cell lines (786-O, ACHN and Caki-1) were obtained from the Chinese Academy of Sciences Cell Bank (Shanghai, China). Three cell lines (HK-2, 786-O and ACHN) and one-cell line (Caki-1) were cultured in RPMI 1640 Medium (Gibco, USA) and McCoy’s 5a (Modified) Medium (Gibco, USA) respectively, supplemented with 10% fetal bovine serum (BI, Israel), 100 U/mL penicillin and 100μg/mL streptomycin (Gibco, USA) at 37°C in a 5% CO2 incubator.
Cell Transfection
The small interfering RNA targeting NSD2 (siNSD2) and negative control (siNC) were synthesized by GenePharma Company (Shanghai, China). Human NSD2 cDNA (NM_133330.2) was PCR-amplified, and then subcloned into a plasmid vector (Invitrogen, USA). 18 h before transfection, cancer cells suspended in medium were added to a 6-well plate. After reaching 50% confluence, the cells were transfected using Lipofectamine 2000 (Invitrogen, USA) in accordance with the manufacturer’s protocols. The efficiency of transfection was detected by qRT-PCR after 48 h and Western blot analysis after 72 h.
The lentiviral vector inhibiting NSD2 expression was named LV-shNSD2 from GenePharma Company (Shanghai, China). The GFP vector (LV-shNC) was used for control. 786-O cells were transfected with LV-shNSD2 or LV-shNC in the presence of polybrene (Solarbio, China). After 48 hrs, fresh medium containing puromycin (Merck, USA) was used for the selection of stable clones. The efficiency of transfection was detected through a fluorescence microscope (Olympus, Japan).
RNA Isolation and Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
Total RNA was isolated using Ultrapure RNA Kit (DNase I) (CWBIO, China) according to the manufacturer’s instructions. The cDNA was synthesized with the PrimeScript RT reagent Kit with gDNA Eraser (Takara, Japan). qRT-PCR was performed using the TB Green Premix Ex Taq II (Takara, Japan) in ABI 7500 Real-Time PCR instrument. GAPDH was acted as an internal control. The primer sequences used for qRT-PCR were recorded in Table 1. The relative mRNA levels were normalized to GAPDH and calculated by the 2−ΔΔCt method.
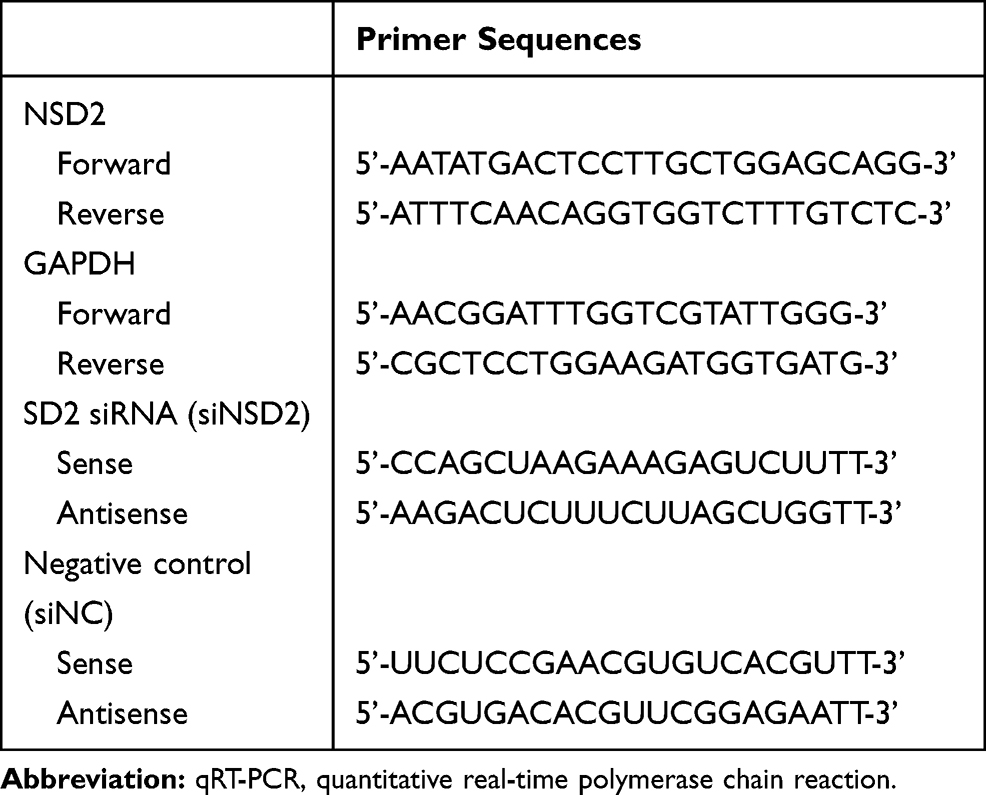 |
Table 1 All Primers for qRT-PCR of NSD2 and GAPDH |
Cell Viability Assay
The cells were seeded at 2x103 cells per well in 96-well plates and cultured for 24, 48, 72 and 96 h after transfection. Cell viability was measured by using 20 μL of 5 mg/mL MTT dye (Beyotime, China) for 4 hrs at 37°C, then removed the supernatant and followed by adding 200 μL dimethyl sulphoxide (Sigma-Aldrich, USA). Absorbance of each well was detected at 570 nm through a microplate reader (ThermoFisher, USA).
Cell Apoptosis Assay
The cells cultured for 72 h after transfection were collected and washed twice with cold PBS. Then, the transfected cells were stained with 5ul Annexin V-FITC and 10ul Propidium Iodide (Multisciences Biotech, China) in the dark for 15 mins at room temperature. Cell apoptosis was assessed by flow cytometry (BD Biosciences, USA) and the data was analyzed by FlowJo software.
Western Blot Analysis
Total protein from transfected cells was obtained using Whole Cell Lysis Assay (Keygen Biotech, China). Protein samples were subjected to SDS–PAGE and transferred to PVDF membranes (Millipore, USA). After blocking with 5% skim milk for 1 h at room temperature, the membranes were incubated with the primary antibody as follows: monoclonal antibodies against NSD2 (Abcam, UK), Akt, p-Akt (S473), Erk1/2, p-Erk1/2 (Thr202/Tyr204), Bcl-2, Bax (Cell Signaling Technology, USA) and GAPDH (Keygen Biotech, China). After washing thrice with TBST, the membranes were incubated with corresponding secondary antibodies (Keygen Biotech, China). Signals were detected using an ECL system (CLiNX, China) and quantified by ImageJ software.
Xenograft Tumor Assay
All animal experiments were conducted according to the National Institute of Health Guide for the Care and Use of Laboratory Animals and were approved by the ethics committee of Soochow University. Ten BALB/C nude male mice (6-week old) were purchased from the Shanghai SLAC Laboratory Animals. 786-O (5 × 106) cells which have been stably infected with LV-shNSD2 or LV-shNC were inoculated subcutaneously together with Matrigel (Corning, USA) into the flank of nude mice. After 7 days of implantation of tumor cells, tumor size was measured every 3 days and tumor volume was calculated with the following formula: (length × width2)/2. At the end of experiments, the mice were sacrificed and tumors were dissected and weighed.
Statistical Analysis
All above data were obtained from at least three independent experiments and presented as means and standard deviation (SD). Statistical analyses were performed using SPSS 19.0 software and GraphPad Prism 5.0. Comparisons between different groups were made via the two-tailed Student’s t-test and Mann–Whitney U-test. Survival curve was drawn using Kaplan-Meier method and Log-rank test. P < 0.05 was considered statistically significant.
Results
NSD2 Expression and Impact on Diagnosis and Outcome of Patients with ccRCC
The RNA sequencing data of ccRCC database (GSE53757) was reanalyzed, which contained 72 ccRCC samples and 72-paired adjacent normal samples. The results suggested that NSD2 mRNA expression in tumor tissues was higher than in adjacent normal tissues with fold change=1.587 (8.802±0.285 vs 8.136±0.425, Figure 1A and B). Furthermore, ROC curve analysis showed the diagnostic value of NSD2 expression, which could be used for discriminating cancer specimens from normal specimens (AUC=0.9026, Figure 1C). Next, the overall survival (OS) data downloaded from OncoLnc suggested that ccRCC patients with higher NSD2 expression had a shorter OS time (Figure 1D).
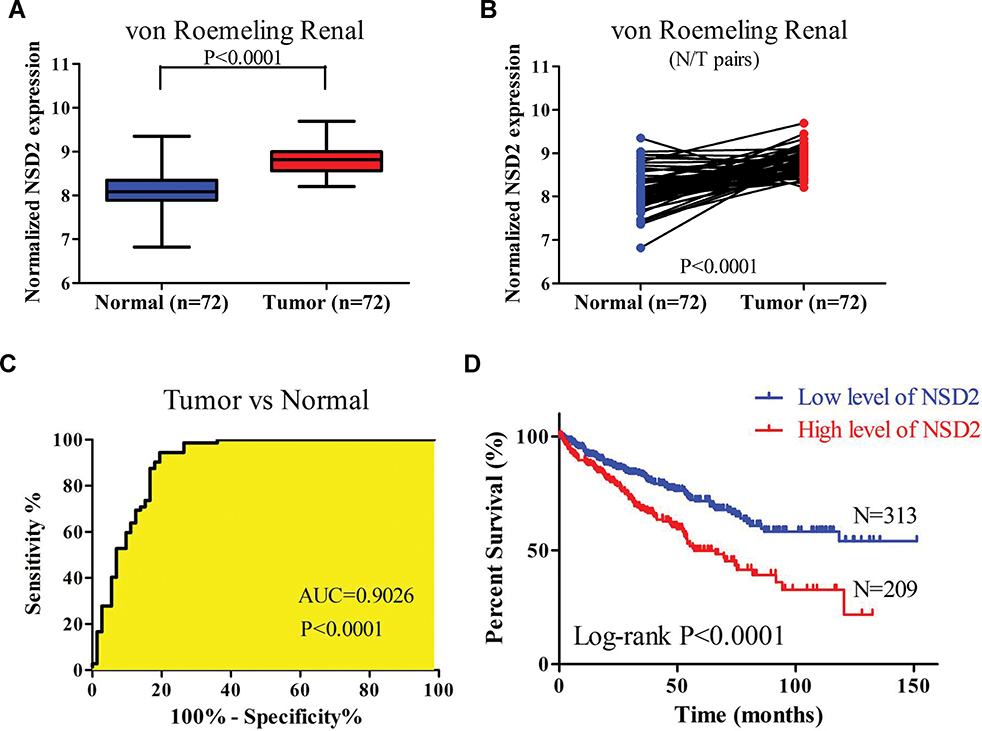 |
Figure 1 NSD2 expression and impact on diagnosis and outcome of patients with ccRCC. (A, B) NSD2 expression is up-regulated in ccRCC samples (N=72) compared with paired adjacent normal samples (N=72) from the GSE53757 database (p<0.0001). (C) ROC curve analysis using NSD2 expression in ccRCC samples and normal samples (AUC=0.9026, p<0.0001). (D) NSD2 expression is negatively correlated with overall survival (OS) time in ccRCC cases from OncoLnc (N=522, p<0.0001). Abbreviations: ccRCC, clear cell renal cell carcinoma; ROC, receiver operating characteristic; AUC, area under ROC curve. |
NSD2 Is Over-Expressed in ccRCC Tissues by Immunohistochemistry (IHC)
Immunohistochemistry (IHC) was conducted to determine the expression level of NSD2 protein in ccRCC and adjacent normal tissues. Representative results with 4-grade scale (IHC score 0, +1, +2 and +3) were shown (Figure 2A–D). Intriguingly, the data showed that NSD2 expression in ccRCC specimens was obviously higher than in adjacent normal specimens (Figure 2E). Collectively, the data suggested that NSD2 might be a vital promoter in the carcinogenesis of ccRCC.
 |
Figure 2 NSD2 is over-expressed in ccRCC tissues by immunohistochemistry (IHC). (A–D) IHC staining (4-grade score 0, +1, +2, +3) for NSD2 protein in ccRCC tissues and adjacent normal tissues. (E) NSD2 protein level in ccRCC specimens was higher than in normal specimens (*p=0.0003; Mann–Whitney U-test). |
NSD2 Expression Is Elevated in ccRCC Cell Lines
To measure the expression of NSD2 in different cell lines, qRT-PCR and Western blotting were conducted. NSD2 mRNA expression was increased in three examined ccRCC cell lines (786-O, ACHN and Caki-1) compared with normal cortex/proximal tubule epithelial cell line (HK-2) (Figure 3A). Interestingly, Western blot analysis indicated that NSD2 protein expression was merely up-regulated in 786-O and ACHN cells, but not altered in Caki-1 cells (Figure 3B). To investigate the biological roles of NSD2 in ccRCC progression, the siRNA and cDNA were designed for functional experiments. The results showed that NSD2 was greatly knocked down after treating siNSD2 in 786-O and ACHN cells (Figure 3C and D), and over-expressed after transfecting cDNA in Caki-1 cells (Figure 3E and F).
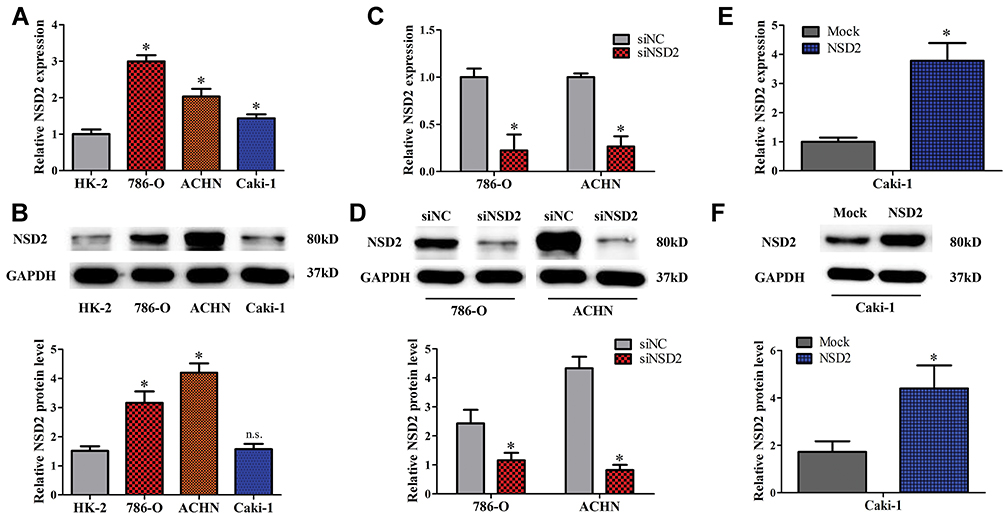 |
Figure 3 NSD2 expression is elevated in ccRCC cell lines. (A) The expression of NSD2 mRNA is increased in three-ccRCC cell lines (786-O, ACHN and Caki-1) compared with normal cortex/proximal tubule epithelial cell line (HK-2). (B) The protein level of NSD2 is up-regulated in two-ccRCC cell lines (786-O and ACHN), but not obviously altered in Caki-1 cells. (C, D) The knockdown efficiency of siNSD2 is confirmed by qRT-PCR and Western blotting in 786-O and ACHN cells. (E, F) The transfection efficiency of cDNA is confirmed in Caki-1 cells. *p<0.05, n.s. p>0.05. Abbreviations: NC, negative control; qRT-PCR, quantitative real-time polymerase chain reaction; n.s., no statistical significance. |
NSD2 Promotes Proliferation and Inhibits Apoptosis in ccRCC Cells
To identify the effect of NSD2 on cell proliferation in ccRCC, siNSD2 and siNC were used to treat ccRCC cells for 24, 48, 72 and 96 h. The MTT assay revealed that down-regulation of NSD2 could reduce cell viability of 786-O and ACHN cells (Figure 4A). Since decreased cell viability was observed in NSD2 depleted cells, it was hypothesized that NSD2 could involve in the process of cell apoptosis. The flow cytometry assay was performed to explore the role of NSD2 in modulating cell apoptosis. The data indicated that suppression of NSD2 could result in a significant increase of apoptotic cells in 786-O and ACHN cells (Figure 4B). Inversely, enhanced NSD2 could promote proliferation and inhibit apoptosis in Caki-1 cells (Figure 4C and D).
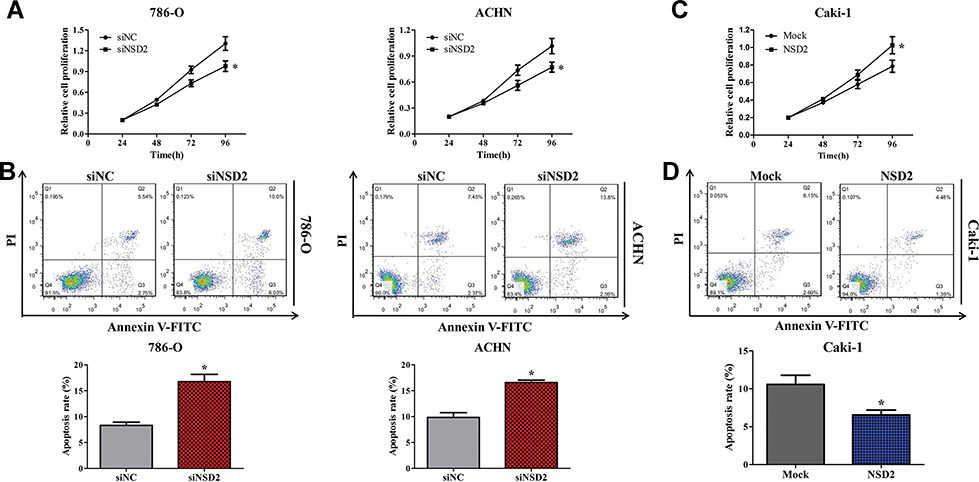 |
Figure 4 NSD2 promotes proliferation and inhibits apoptosis in ccRCC cells. (A) Down-regulation of NSD2 reduces cell viability of 786-O and ACHN cells. (B) Suppression of NSD2 induces cell apoptosis of 786-O and ACHN cells. (C-D) Elevation of NSD2 promotes proliferation and inhibit apoptosis in Caki-1 cells. *p<0.05. Abbreviations: MTT, 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide; FITC, fluorescein isothiocyanate; PI, propidium iodide. |
NSD2 Activates Akt/Erk Signaling and Regulates Bcl-2 and Bax Expression
Previously, it was reported that NSD2 regulated osteosarcoma progression and chemosensitivity through Erk and Akt pathways.19 Meanwhile, the anti-apoptotic Bcl-2 and pro-apoptotic Bax, both belong to the Bcl-2 protein family, were confirmed to act as key regulators in ccRCC development.20,21 Here, Western blot analysis revealed that silencing of NSD2 could repress the expression of p-Akt, p-Erk1/2, Bcl-2, and enhance Bax at protein level, but not alter total Akt and Erk1/2 expression in 786-O and ACHN cells (Figure 5A). Contrarily, over-expression of NSD2 increased p-Akt, p-Erk1/2, Bcl-2 protein, and decreased Bax protein in Caki-1 cells (Figure 5B). The findings illustrated that NSD2 suppressed cell apoptosis through motivating Akt/Erk pathways and mediating Bcl-2 and Bax expression in ccRCC.
 |
Figure 5 NSD2 activates Akt/Erk signaling and regulates Bcl-2 and Bax expression. (A) Silencing of NSD2 represses p-Akt, p-Erk1/2, Bcl-2 expression, and enhances Bax expression, but not alters total Akt and Erk1/2 expression in 786-O and ACHN cells. (B) Over-expression of NSD2 increases p-Akt, p-Erk1/2 and Bcl-2 protein, and decreases Bax protein in Caki-1 cells. *p<0.05. Abbreviations: p-Akt, phospho-Akt; p-Erk1/2, phospho-Erk1/2. |
Knockdown of NSD2 Reduces the Formation of Xenograft Tumors in vivo
To determine the in vivo effect of NSD2 on tumor growth, ten-nude mice were carried out to establish xenograft models. The lentiviral vector (LV-shNSD2) and negative control (LV-shNC) were constructed to stably transfect 786-O cells (Figure 6A), and the transfected cells were subsequently implanted into nude mice. The results showed that knockdown of NSD2 would reduce tumor growth in vivo, compared with control group (Figure 6B), as manifested by decreased tumor volume and weight (Figure 6C and D).
 |
Figure 6 Knockdown of NSD2 reduces the formation of xenograft tumors in vivo. (A) The fluorescence microscope is used to detect the efficiency of LV-shNC and LV-shNSD2 transfection. (B) Subcutaneous tumors formed in nude mice by 786-O cells stably inhibition of NSD2 or control. (C) Tumor formation growth curves after injection of transfected cells. (D) The mean tumor weights of each group. *p<0.05. Abbreviation: LV, lentiviral vector. |
Discussion
Renal cell carcinoma (RCC) has been a severe problem threatening public health around the world, which accounts for approximately 2.2% of new cancer cases and 1.8% of cancer deaths.22 On account of few effective therapeutic strategies, the 5-year survival rate of RCC remains at merely 55%.23 The most frequently occurring subtype is clear cell renal cell carcinoma (ccRCC), which is responsible for 75–80% of RCC.24 Hence, it is imperative to discover new reliable biomarkers and novel therapeutic targets in ccRCC.
Increasing evidence has revealed that NSD2 contributed to cancer pathogenesis. NSD2 mRNA was amplified in 13 cancer types compared with their normal counterparts according to expression profile analysis.25 Besides, up-regulation of NSD2 was related to higher grade in oligodendroglioma, breast, prostate and head and neck cancers.26 However, the biological role of NSD2 in ccRCC progression remains to be elucidated. Here, we found that NSD2 expression in ccRCC tissues was higher than in normal tissues. Furthermore, NSD2 expression could discriminate ccRCC samples from normal samples and higher NSD2 expression exhibited shorter OS time. The findings indicated that NSD2 might play an oncogenic role in ccRCC tumorigenesis.
The molecular mechanism of how NSD2 exerts biological functions on ccRCC is undetermined. It is well-known that Akt and Erk signaling pathways involve in cell proliferation and apoptosis. Additionally, Bcl-2 and Bax protein were both implicated in cancer progression and their abnormal expression was frequently detected in ccRCC.27 Here, down-regulation of NSD2 repressed proliferation and induced apoptosis in 786-O and ACHN cells, accompanied with decreased Akt and Erk1/2 phosphorylation, Bcl-2 protein, and increased Bax expression. Whereas, over-expression of NSD2 exhibited the opposite effects in Caki-1 cells. In addition, depletion of NSD2 reduced xenograft tumor growth in vivo. Thus, the results demonstrated that NSD2 would facilitate ccRCC carcinogenesis via stimulation of Akt/Erk signaling and regulation of Bcl-2 and Bax expression.
Conclusions
In summary, NSD2 expression level was up-regulated and NSD2 functioned as tumor promoter in ccRCC progression. Therefore, we proposed NSD2 as a potential therapeutic target for ccRCC treatment.
Acknowledgment
This research was supported by Changzhou Sci&Tech Program (Grant No. CJ20180064).
Disclosure
The authors declare that they have no conflicts of interest in this work.
References
1. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2018. CA Cancer J Clin. 2018;68(1):7–30. doi:10.3322/caac.21442
2. Znaor A, Lortet-Tieulent J, Laversanne M, Jemal A, Bray F. International variations and trends in renal cell carcinoma incidence and mortality. Eur Urol. 2015;67(3):519–530. doi:10.1016/j.eururo.2014.10.002
3. Heuer R, Gill IS, Guazzoni G, et al. A critical analysis of the actual role of minimally invasive surgery and active surveillance for kidney cancer. Eur Urol. 2010;57(2):223–232. doi:10.1016/j.eururo.2009.10.023
4. Cancer Genome Atlas Research Network. Comprehensive molecular characterization of clear cell renal cell carcinoma. Nature. 2013;499(7456):43–49. doi:10.1038/nature12222
5. Han X, Piao L, Zhuang Q, Yuan X, Liu Z, He X. The role of histone lysine methyltransferase NSD3 in cancer. Onco Targets Ther. 2018;11:3847–3852. doi:10.2147/OTT
6. Wagner EJ, Carpenter PB. Understanding the language of Lys36 methylation at histone H3. Nat Rev Mol Cell Biol. 2012;13(2):115–126. doi:10.1038/nrm3274
7. Vougiouklakis T, Hamamoto R, Nakamura Y, Saloura V. The NSD family of protein methyltransferases in human cancer. Epigenomics. 2015;7(5):863–874. doi:10.2217/epi.15.32
8. Lucio-Eterovic AK, Singh MM, Gardner JE, Veerappan CS, Rice JC, Carpenter PB. Role for the nuclear receptor-binding SET domain protein 1 (NSD1) methyltransferase in coordinating lysine 36 methylation at histone 3 with RNA polymerase II function. Proc Natl Acad Sci U S A. 2010;107(39):16952–16957. doi:10.1073/pnas.1002653107
9. Chesi M, Nardini E, Lim RS, Smith KD, Kuehl WM, Bergsagel PL. The t(4;14) translocation in myeloma dysregulates both FGFR3 and a novel gene, MMSET, resulting in IgH/MMSET hybrid transcripts. Blood. 1998;92(9):3025–3034. doi:10.1182/blood.V92.9.3025
10. Stec I, Wright TJ, van Ommen GJ, et al. WHSC1, a 90 kb SET domain-containing gene, expressed in early development and homologous to a Drosophila dysmorphy gene maps in the Wolf–hirschhorn syndrome critical region and is fused to IgH in t(4;14) multiple myeloma. Hum Mol Genet. 1998;7(7):1071–1082. doi:10.1093/hmg/7.7.1071
11. Finelli P, Fabris S, Zagano S, et al. Detection of t(4;14)(p16.3;q32) chromosomal translocation in multiple myeloma by double-color fluorescent in situ hybridization. Blood. 1999;94(2):724–732. doi:10.1182/blood.V94.2.724
12. Lauring J, Abukhdeir AM, Konishi H, et al. The multiple myeloma associated MMSET gene contributes to cellular adhesion, clonogenic growth, and tumorigenicity. Blood. 2008;111(2):856–864. doi:10.1182/blood-2007-05-088674
13. Brito JL, Walker B, Jenner M, et al. MMSET deregulation affects cell cycle progression and adhesion regulons in t(4;14) myeloma plasma cells. Haematologica. 2009;94(1):78–86. doi:10.3324/haematol.13426
14. Martinez-Garcia E, Popovic R, Min DJ, et al. The MMSET histone methyl transferase switches global histone methylation and alters gene expression in t(4;14) multiple myeloma cells. Blood. 2011;117(1):211–220. doi:10.1182/blood-2010-07-298349
15. Huang Z, Wu H, Chuai S, et al. NSD2 is recruited through its PHD domain to oncogenic gene loci to drive multiple myeloma. Cancer Res. 2013;73(20):6277–6288. doi:10.1158/0008-5472.CAN-13-1000
16. Shah MY, Martinez-Garcia E, Phillip JM, et al. MMSET/WHSC1 enhances DNA damage repair leading to an increase in resistance to chemotherapeutic agents. Oncogene. 2016;35(45):5905–5915. doi:10.1038/onc.2016.116
17. Asangani IA, Ateeq B, Cao Q, et al. Characterization of the EZH2-MMSET histone methyltransferase regulatory axis in cancer. Mol Cell. 2013;49(1):80–93. doi:10.1016/j.molcel.2012.10.008
18. Yang P, Guo L, Duan ZJ, et al. Histone methyltransferase NSD2/MMSET mediates constitutive NF-κB signaling for cancer cell proliferation, survival, and tumor growth via a feed-forward loop. Mol Cell Biol. 2012;32(15):3121–3131. doi:10.1128/MCB.00204-12
19. He C, Liu C, Wang L, Sun Y, Jiang Y, Hao Y. Histone methyltransferase NSD2 regulates apoptosis and chemosensitivity in osteosarcoma. Cell Death Dis. 2019;10(2):65–77. doi:10.1038/s41419-019-1347-1
20. Fang Z, Tang Y, Jiao W, et al. Nitidine chloride induces apoptosis and inhibits tumor cell proliferation via suppressing ERK signaling pathway in renal cancer. Food Chem Toxicol. 2014;66:210–216. doi:10.1016/j.fct.2014.01.049
21. Yuan SY, Cheng CL, Wang SS, et al. Escin induces apoptosis in human renal cancer cells through G2/M arrest and reactive oxygen species-modulated mitochondrial pathways. Oncol Rep. 2017;37(2):1002–1010. doi:10.3892/or.2017.5348
22. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. doi:10.3322/caac.v68.6
23. Ljungberg B, Cowan NC, Hanbury DC, et al. EAU guidelines on renal cell carcinoma: the 2010 update. Eur Urol. 2010;58(3):398–406. doi:10.1016/j.eururo.2010.06.032
24. Srigley JR, Delahunt B, Eble JN, et al. The international society of urological pathology (ISUP) vancouver classification of renal neoplasia. Am J Surg Pathol. 2013;37(10):1469–1489. doi:10.1097/PAS.0b013e318299f2d1
25. Toyokawa G, Cho HS, Masuda K, et al. Histone lysine methyltransferase Wolf-Hirschhorn syndrome candidate 1 is involved in human carcinogenesis through regulation of the Wnt pathway. Neoplasia. 2011;13(10):887–898. doi:10.1593/neo.11048
26. Saloura V, Cho HS, Kiyotani K, et al. WHSC1 promotes oncogenesis through regulation of NIMA-related-kinase-7 in squamous cell carcinoma of the head and neck. Mol Cancer Res. 2015;13(2):293–304. doi:10.1158/1541-7786.MCR-14-0292-T
27. Deng W, Han W, Fan T, et al. Scutellarin inhibits human renal cancer cell proliferation and migration via upregulation of PTEN. Biomed Pharmacother. 2018;107:1505–1513. doi:10.1016/j.biopha.2018.08.127
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.