Back to Journals » Infection and Drug Resistance » Volume 11
Novel vancomycin–peptide conjugate as potent antibacterial agent against vancomycin-resistant Staphylococcus aureus
Authors Jelinkova P ![]() , Splichal Z, Jimenez Jimenez AM, Haddad Y
, Splichal Z, Jimenez Jimenez AM, Haddad Y ![]() , Mazumdar A, Sur VP, Milosavljevic V, Kopel P
, Mazumdar A, Sur VP, Milosavljevic V, Kopel P ![]() , Buchtelova H, Guran R
, Buchtelova H, Guran R ![]() , Zitka O
, Zitka O ![]() , Richtera L
, Richtera L ![]() , Hegerova D, Heger Z
, Hegerova D, Heger Z ![]() , Moulick A
, Moulick A ![]() , Adam V
, Adam V ![]()
Received 28 December 2017
Accepted for publication 4 May 2018
Published 12 October 2018 Volume 2018:11 Pages 1807—1817
DOI https://doi.org/10.2147/IDR.S160975
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr Joachim Wink
Pavlina Jelinkova,1 Zbynek Splichal,1,2 Ana Maria Jimenez Jimenez,1,2 Yazan Haddad,1,2 Aninda Mazumdar,1,2 Vishma Pratap Sur,1,2 Vedran Milosavljevic,1,2 Pavel Kopel,1,2 Hana Buchtelova,1 Roman Guran,1,2 Ondrej Zitka,1,2 Lukas Richtera,1,2 Dagmar Hegerova,1,2 Zbynek Heger,1,2 Amitava Moulick,1,2 Vojtech Adam1,2
1Department of Chemistry and Biochemistry, Mendel University in Brno, Zemedelska, Brno, Czech Republic; 2Central European Institute of Technology, Brno University of Technology, Purkynova, Brno, Czech Republic
Background: Increase in vancomycin (Van)-resistant bacterial strains including vancomycin-resistant Staphylococcus aureus (VRSA) and lack of new effective antibiotics have become a formidable health problem.
Materials and methods: We designed a new conjugate composed of Van and a peptide Hecate (Hec; Van/Hec), and its potential antimicrobial activity was evaluated.
Results: Results from disk diffusion test, time-kill assay, determination of minimum inhibitory concentration (MIC), microscopy, and comet assay showed strong antimicrobial effects of Van/Hec against wild-type, methicillin-resistant Staphylococcus aureus (MRSA) and VRSA. Microscopy revealed that the exposure to Van/Hec results in disruption of bacterial cell integrity in all tested strains, which was not observed in case of Van or Hec alone.
Conclusion: Overall, we showed that the preparation of conjugates from antibiotics and biologically active peptides could help us to overcome the limitation of the use of antibiotic in the treatment of infections caused by multidrug-resistant bacteria.
Keywords: vancomycin, antibacterial, Staphylococcus aureus, antibiotic resistance, peptide
Introduction
Bacterial infections have become an important worldwide public health problem and have an increased morbidity, mortality, and cost during a patient’s hospital stay.1 In recent decades, bacterial resistance to antimicrobial agents has been considered as a major problem in veterinary and public health. Because of the extended use or misuse of antibiotics,2 the number of bacteria that is resistant to antimicrobial agents is rapidly increasing.3 Different studies confirmed that the antibiotic resistance in bacterial pathogens can be induced and spread rapidly among bacterial species.4,5
As an aggressive pathogen, Staphylococcus aureus poses a significant public health threat and is becoming increasingly resistant to currently available antibiotics, including vancomycin (Van), the drug of last resort for Gram-positive bacterial infections.6 Despite this, vancomycin-resistant Staphylococcus aureus (VRSA)7,8 is described, complicating the Van application. The first Van-resistant clinical isolate was Enterococcus faecium isolated in 1988.9 Similar strains were later detected in hospitals on the east coast of the USA.10 VRSA was first identified in June 2002.9
From a microbiological perspective, VRSA is associated with a wide array of changes in bacterial physiology. These include an increase in cell wall diameter (Van cannot penetrate the cells), altered autolysis, decreased growth, and alterations in gene transcription.11 The activity of Van can be determined by the substrate specificity of the enzymes that determine the structure of peptidoglycan precursors but not by the affinity for a target enzyme because it does not interact with the biosynthetic enzymes present in cell wall but forms complexes with peptidoglycan precursors.10
Cell-penetrating peptides (CPPs), which facilitate cellular uptake or intake of different molecular equipment, have attracted a great deal of interest.12 Their ability is to form pores into the bacterial cell walls.13 The possibility of conjugation of the peptides with various nanomaterials14 or chemical activators15 offers a wide range of new opportunities for their use in medical applications. Therefore, we used antibacterial peptide Hecate (Hec; Gram+ and Gram- pathogens) for the conjugation with glycopeptide antibiotic Van, which is mostly used as a drug against Gram-positive-resistant bacterial strains.16 Hec (derivative of melittin, a component of honey bee (Apis mellifera) venom) was created from 23 amino acids (FALALKALKKALKKLKKALKKAL) with a high incidence of lysine (positive), leucine and alanine (nonpolar).17 Peptide Hec has an amphipathic α-helix structure, positive net charge, and a high number of hydrophobic amino acids, such as other antibacterial peptides (Ctx-Ha and Hylin-a1).18
Our study is focused on the development of a new conjugate (Van/Hec) using a common antibiotic like Van and the biologically active compounds like peptide Hec to overcome the limitation of the use of antibiotic in the treatment of resistant bacterial infections. The conjugate was tested on the resistant and non-resistant strains of S. aureus using different methods.
Materials and methods
Chemical compounds
All the reagents for peptide synthesis, oligonucleotides, standards, and other chemicals were purchased from Sigma-Aldrich Co. (St Louis, MO, USA) in American Chemical Society (ACS) purity unless noted otherwise.
Synthesis of Hec and conjugation with Van (Van/Hec conjugate)
Synthesis of Hec
Hec was synthesized using standard Fmoc solid phase synthesis on Liberty Blue peptide synthesizer (CEM, Matthews, NC, USA). The sequence and monoisotopic molecular weight of Hec were as follows: FALALKALKKALKKLKKALKKAL and 2535.7 Da, respectively. The purity of the peptide was evaluated using high-performance liquid chromatography with ultraviolet detection (HPLC–UV; ESA Laboratories Inc., Chelmsford, MA, USA). The final purity was >92%. The molecular weight of the peptide was verified by the high-performance liquid chromatography coupled with electrospray ionization quadrupole time-of-flight (HPLC–ESI–Q-TOF; Bruker Daltonik GmbH, Bremen, Germany) technique.
Conjugation of Van and Hec through 1,1′-carbonyldiimidazole (CDI)
The Van solution (0.5 mL, 2.70 mM) was mixed with Hec (0.5 mL, 1.58 mM) and a zero-length cross-linker (CDI – 6.2 µM in 20 µL dimethyl sulfoxide [DMSO]). The mixture was shaken overnight at 25 °C on Biosan Orbital Shaker (Biosan, Riga, Latvia).
Matrix-assisted laser desorption/ionization (MALDI)–time of flight mass spectrometry (TOF MS) analyses of Hec, Van, and Van/Hec
The samples were diluted in 0.1% trifluoroacetic acid (TFA), and 1 µL of each of the samples was spotted in duplicate onto the MALDI target (MTP 384 target polished steel plate; Bruker Daltonics, Bremen, Germany) and air-dried at 25 °C. Then, each spot was overlaid with 1 µL of 2,5-dihydroxybenzoic acid (DHB) or sinapic acid (SA) matrix solution (20 mg/mL) in 50% acetonitrile and 0.1% TFA and air-dried completely prior to the MALDI–TOF MS measurement on ultrafleXtreme (Bruker Daltonics, Bremen, Germany). Reflector positive mode was used for Van and Hec, and in case of Van/Hec analysis, linear positive mode was used. Each spectrum was taken in the m/z range 500–8000 Da; one sample mass spectrum was averaged from 1000 sub-spectra per spot.
Fourier transform infrared spectroscopy (FTIR)
All FTIR spectra were collected using a Nicolet iS10 FTIR spectrometer with a diamond attenuated total reflectance (ATR) attachment (Thermo Fisher Scientific, Waltham, MA, USA). The sample was supplied dropwise (5 μL) on the diamond crystal of the ATR cell first, and then, the film was measured after spontaneous evaporation of the solvent. The infrared (IR) spectra were recorded from 4000 to 650 cm-1 at a resolution of 4 cm-1. Each spectrum was acquired by adding together 64 interferograms. The spectra were acquired at 22°C.
Cultivation of bacterial strains
Bacterial strains (S. aureus NCTC 8511, methicillin-resistant Staphylococcus aureus [MRSA] ST239:SCCmec IIIA, VRSA, Van-resistant Enterococcus faecalis ATCC 51299, Escherichia coli NCTC 13216) were obtained from the Czech Collection of Microorganisms, Faculty of Science, Masaryk University, Brno, Czech Republic. Four human pathogenic MRSA were obtained from England with the cooperation of the University of Cambridge. The composition of cultivation medium was as follows: nutrient broth 13 g/L and 1000 mL sterilized MilliQ water with 18 MΩ. pH of the cultivation medium was adjusted to pH 7.4. Prior to experiments, the bacterial cultures were diluted by the cultivation medium to OD600 nm = 0.5 McFarland.19
Determination of inhibition zones using disk diffusion method
The antimicrobial effect was analyzed using the measurement of inhibition zones by the agar microdilution method. The agar surface of Petri dish was covered with a mixture of 100 µL of overnight bacterial cultures and 3 mL of Luria–Bertani (LB) broth to get the final density of 0.5 McFarland standard. The excess volume of the mixture of the Petri dishes was aspirated. The disks (⌀ 6 mm) were soaked with the appropriate concentrations of the solutions of antibiotics (lincomycin, amoxicillin, tetracycline, penicillin, erythromycin, and Van – antibiotics were tested at concentrations referred to as limits for the emergence of resistance) and antibacterial agents (Van, peptide Hec, and Van/Hec conjugate) by addition of 10 µL of the solutions.20–22 The disks were placed onto the Mueller–Hinton (MH) agar plates. The MH agar plates were protected from possible external contamination and placed in a thermostat (Tuttnauer 2450EL; Tuttnauer Ltd., Beit Shemesh, Israel) at 37 °C for 24 h. After 24 h of incubation, the inhibition zones were measured by caliper and photographed.23
Minimum inhibitory concentration (MIC) determination
The susceptibility of staphylococcal cultures was determined by the standard broth microdilution method (European Committee on Antimicrobial Susceptibility Testing) and detected by the unaided eye. The MIC was defined as the lowest concentration of antimicrobial agent that inhibited the bacterial growth. The MIC of antimicrobial agents was determined by preparation of solutions with increasing concentrations. The different concentrations of solutions were added into the microplate wells and were mixed with bacterial cultures (0.5 McFarland and final dilution 1:100 with the MH medium). The microplate with antimicrobial agents and bacterial cultures was incubated at 37 °C for 24 h. The results were evaluated the next day. The well with the concentration of higher dilution with no bacterial growth was considered the MIC. As positive control, the MH medium with inoculation was used.23
Time-kill analysis
The Eppendorf tubes containing MH broth and antimicrobial agents with the concentration of 5 µM (Van, peptide Hec and Van/Hec conjugate) were inoculated by bacterial cultures (to a density of 0.5 McFarland), adjusted to 1 × 108. Then, the samples were incubated for different time periods (0, 6, 12 and 24 h) in a shaker at 37 °C. Separate samples were prepared for each of the time points to be studied. The Eppendorf tubes with samples were removed from the shaker after a particular time and then serially diluted (10-fold dilutions) in sterile water to measure viable counts. In all, 100 µL of the each of the diluted samples was plated separately on blood agar and then incubated at 37 °C for 24 h. The number of colonies on blood agar plate was counted. The ultimate colony-forming units (CFUs) were the number of colonies multiplied by the dilution ratio.24–27 The experiment was repeated five times.
Cell culture and testing of the cytotoxicity
The PNT1A human cell line established by immortalization of normal adult prostatic epithelial cells, which were bought from Sigma-Aldrich Co., was cultured in Roswell Park Memorial Institute (RPMI)-1640 medium with 10% fetal bovine serum, supplemented with penicillin (100 U/mL) and streptomycin (0.1 mg/mL). The treatment was initiated after the cells reached ~60%–80% confluence. The cell viability was estimated using the MTT assay following the conditions used in our previous study.28
Estimation of hemocompatibility
Fresh human blood was collected from the volunteer with signed informed consent by a venepuncture from an antecubital vein. This study was approved by the ethics committee of the University Hospital Brno (Brno, Czech Republic). Plasma and serum were removed and washed with 150 mM NaCl from the sample by centrifuging at 2000× g for 5 min three times. Red blood cells (RBCs) were diluted with PBS (pH 7.4), and subsequently, Van, Hec, and Van/Hec were added in different concentrations (20, 10, 5, and 2.5 µM) separately into the RBC solution. PBS and 0.1% Triton X-100 were used as a negative control and a positive control, respectively. The samples were mixed and incubated for 1 h at 37 °C. After that, the samples were centrifuged at 3000× g for 10 min, and the absorbance was measured at λ = 540 nm. A percentage of hemolysis was calculated according to the following equation: %hemolysis = [(At - Ac)/(A100% - Ac)] × 100, where At is the absorbance of the supernatant from the samples incubated with the Van, Hec, or Van/Hec; Ac is the absorbance of the supernatant from negative control (PBS, pH 7.4); and A100% is the absorbance of the supernatant of positive control (0.1% Triton X-100), which causes complete lysis of RBCs.
Microscopy of antibacterial agents against bacteria in ambient light and live/dead cell assay
The microscopic assay for the evaluation of live/dead bacterial cells was performed using an inverted Olympus IX 71S8F-3 fluorescence microscope (Olympus Corporation, Tokyo, Japan) equipped with Olympus UIS2 series objective LUCPlanFLN 40× (N.A. 0.6, WD 2.7–4 mm, F.N. 22) and a mercury arc lamp X-cite 12 (120 W; Lumen Dynamics, Mississauga, ON, Canada). Two fluorescent dyes were used for the live/dead cell assay: propidium iodide (PI) for staining of cells with damaged membranes and SYTO9 (Thermo Fisher Scientific) for permeating both intact and damaged membranes of the cells.29
Single-cell gel electrophoresis (comet assay)
The human cells were plated at a density of 106 cells/well in six-well dishes (The bacterial cells were diluted at a density 0.5 McFarland.) and treated with Van, peptide Hec, and Van/Hec for 2 h. The applied concentration of Van, peptide Hec, and Van/Hec was 5 µM. As a control, 250 µM H2O2 was used. After harvesting (in the case of the bacterial cells, the lysozyme was added for destroying of bacterial cell wall in the concentration of 20 µg/mL; the samples were shaken together for 1 h, and after lysis reaction, the steps of the methods were followed as for the human cells), ~10 μL of the cell suspension was mixed with 75 μL of 0.8% low-melting point agarose (CLP, San Diego, CA, USA) and layered on one end of a frosted plain glass slide. Then, it was covered with a layer of the low-melting agarose (100 μL). After solidification of the gel, the slides were immersed in a lysing solution (2.5 M NaCl, 100 mM Na2EDTA, 10 mM Tris, pH 10 containing 1% Triton X-100 and 10% DMSO) for 2 h incubation at 4 °C. A cold alkaline electrophoresis buffer was poured into the chamber and incubated for 30 min at 4 °C. The electrophoresis was carried at 4 °C for 30 min at 1.25 V/cm and 300 mA. The slides were neutralized (0.4 M Tris, pH 7.5) and then stained with ethidium bromide (EtBr; 2 µg/mL). The cells were analyzed using a fluorescence microscope (Olympus IX 71S8F-3; Olympus Corporation) and classified according to the shape of the fluorescence of the comet tail (0 [no visible tail] to 4 [significant DNA in the tail]).
Statistical analysis
Software Statistica (data analysis software system), version 10.0 (StatSoft, Tulsa, OK, USA) was used for data processing. The general regression model was used to analyze the differences between the measured values. To reveal differences between the cell lines, Tukey’s post hoc test within homogenous groups was used. Unless noted otherwise, p < 0.05 was considered as significant.
Molecular characterization of VRSA
Preparation of samples for isolation of bacterial DNA
The harvested bacterial pellets were resuspended in 400 µL of lysis buffer (6 M guanidine hydrochloride, 0.1 M sodium acetate). Then, the lysates were incubated for 30 min on a roller incubator at 25 °C. Subsequently, DNA was isolated from the lysates using the MagNA Pure Compact Nucleic Acid Isolation Kit following the manufacturer’s instructions on MagNA Pure Compact Instrument (Hoffman-La Roche Ltd., Basel, Switzerland).
Detection of vanA gene
Polymerase chain reaction (PCR) with the following set of primers designed in Primer3 online tool was used for detection of vanA gene: vanA fw 5′-CCATACAAGGTCTGTTTGAATTG-3′ and vanA rw 5′-GGCCATCATACGGGGATAAC-3′. PCR amplification was performed with Mastercycler® nexus (Eppendorf, Hamburg, Germany) in a volume of 20 µL. PCR reaction mix includes the following components: 0.1 μg bacterial DNA; 1.5 mM MgCl2; 200 μM each of dATP, dTTP, dGTP, and dCTP; 0.5 μM of each primer; and 1 unit Taq polymerase (NEB, Ipswich, MA, USA). The PCR program was carried out as follows: initial denaturation at 94°C for 2 min, 40 cycles of denaturation at 94 °C for 20 s, annealing at 55 °C for 20 s, and extension at 68 °C for 45 s, with the final extension at 68 °C for 5 min. After PCR, 650 bp amplicons were detected by gel electrophoresis and EtBr staining.
Results and discussion
The main objectives were to synthesize Van/Hec conjugate and test the antimicrobial effects against S. aureus and its resistant counterparts – MRSA, VRSA, and vancomycin-resistant Enterococcus faecalis (VRE). Individual parts of designated conjugate showed different antimicrobial activities depending on the susceptibility of bacterial strains. Van as an antibiotic has a good antimicrobial activity against S. aureus and MRSA but not against VRSA. The high resistance of VRSA toward Van limits its application.30 On the other hand, it has been reported that Hec has antimicrobial activity.31 Despite this, as most of CPPs, Hec has drawbacks resulting from pronounced unwanted toxicity in eukaryotic cells.32 We produced Van/Hec (Figure 1A) using Van and Hec as building blocks, which were conjugated through zero-length cross-linker. The Van/Hec was produced to increase the antimicrobial activity of Van through a CPP-based penetration of VRSA’s cell wall and to decrease the toxicity of the naked and linear CPP molecule. The conjugation site is most likely the carboxyl group of Van, which is similar to the product of Yarlagadda et al.33 The Van/Hec was characterized using MALDI–TOF MS (Figure 1B) and FTIR (Figures 1C and S1). The IR spectra of Van/Hec conjugate exhibit the presence of vibrations associated with both of the constituents. The identification of the newly formed (presumed) amide bond between Van and Hec is extremely hard because of the considerable amount of very similar functional groups (amide groups, hydroxyl groups). Despite this, the spectra are consistent with the expected product – Van/Hec, showing the vibrations attributable to either Van or Hec.
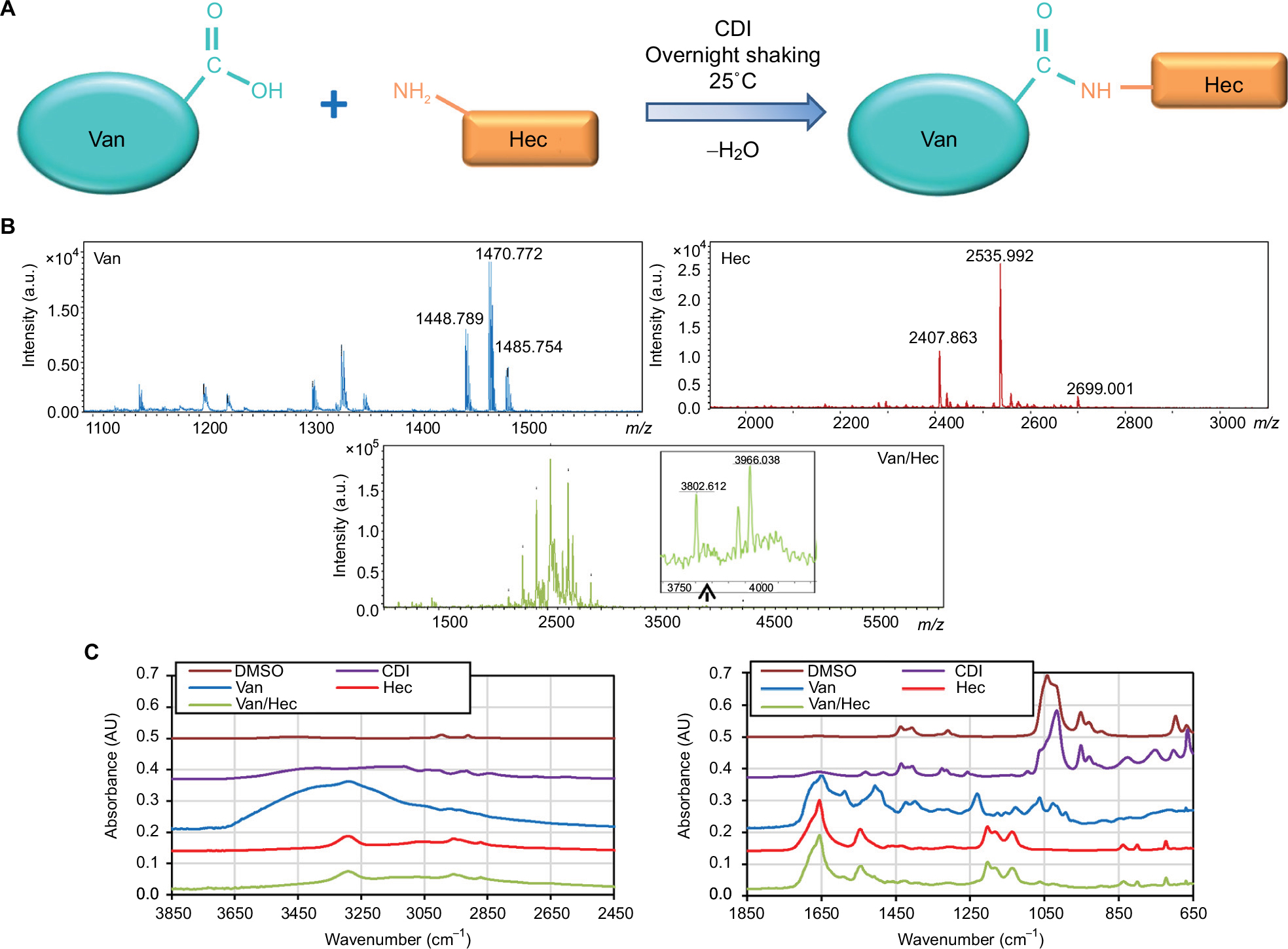 | Figure 1 Van/Hec Cross-linking and characterization Notes: (A) Scheme of preparation of Van/Hec through zero-length cross-linker (CDI). (B) Representation of MALDI–TOF MS of Van, Hec, and Van/Hec. As matrix, a DHB was used at a concentration of 20 mg/mL in 50% acetonitrile and 0.1% TFA. Linear positive mode was used. In total, 320 mass spectra were averaged per one sample spot. (C) FTIR spectrum of all individual components used for Van/Hec conjugation and completed Van/Hec. Abbreviations: Van, vancomycin; Hec, Hecate; CDI, 1,1′-carbonyldiimidazole; MALDI, matrix-assisted laser desorption/ionization; TOF MS, time of flight mass spectrometry; DHB, 2,5-dihydroxybenzoic acid; TFA, trifluoroacetic acid; FTIR, Fourier transform infrared spectroscopy; a.u., arbitrary unit; DMSO, dimethyl sulfoxide. |
Van resistance results in a pronounced cross-resistance to other antibiotics
In order to obtain information about the possible Van-inducted cross-resistance, we decided to test the susceptibility of established strains to selected conventional antibiotics (Figure S2A). The analysis of inhibition zones of applied antibiotics (lincomycin, amoxicillin, tetracycline, penicillin, erythromycin, and Van) revealed their pronounced antimicrobial activity only in S. aureus with inhibition zones between 16 and 34 nm (Figure S2Aa). In MRSA, the antibacterial effect of the antibiotics significantly decreased and only tetracycline (32 mm), Van, and amoxicillin (16 mm) retained their antimicrobial properties (Figure S2Ab). Finally, the testing of antibiotics in VRSA revealed the total absence of inhibition zones, but we can see a partial inhibition zone after the application of tetracycline on VRSA (Figure S2Ac). The values of inhibition zones are summarized in Table 1. Overall, it has to be noted that the VRSA owns superior properties, which predispose it as an exceptional model for studying novel antimicrobial agents. MICs for the selected antibiotics were studied by several other researchers,34–39 which are in good agreement with our results.
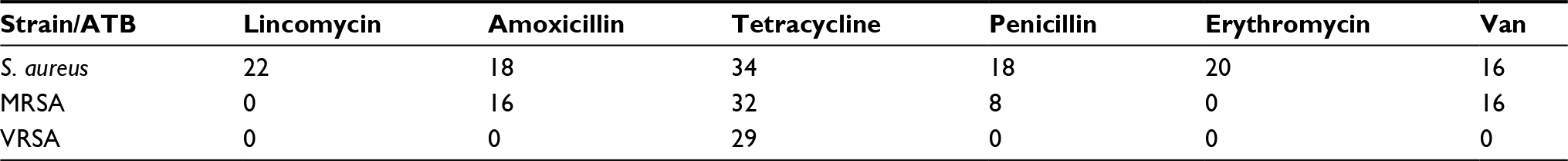 | Table 1 Inhibition zones (mm) showing significant cross-resistance of VRSA strain Note: In case of tetracycline, VRSA showed a partial inhibition zone. Abbreviations: VRSA, vancomycin-resistant Staphylococcus aureus; Van, vancomycin; S. aureus, Staphylococcus aureus; MRSA, methicillin-resistant Staphylococcus aureus. |
At first, we investigated the susceptibility of VRSA to Van, peptide Hec, and Van/Hec and then compared the data with the susceptibility of MRSA and S. aureus. As we expected, the inhibition zones showed that Van had the strongest antimicrobial activity in S. aureus (16 mm) and MRSA (16 mm; Figure S2Ba and b), while for VRSA, the application of Van showed no observable antimicrobial activity (Figure S2Bc). Small inhibition zones (8 mm) were found in the case of peptide Hec against S. aureus and MRSA strains (Figure S2Ba and b) but not in the case of VRSA (Figure S2Bc). However, a strong effect of Van/Hec was found for all the tested bacteria. The diameters of inhibition zones were 25, 24 and 12 mm for S. aureus, MRSA, and VRSA, respectively (Figure S2C). To validate the results, we determined the MICs of antimicrobial agents by broth microdilution method (Table 2). The MICs of Van for S. aureus, MRSA, and VRSA were 0.6, 5, and > 80 μM, respectively. All used bacterial strains in the study had a peptide Hec MIC higher than 80 μM, and MICs of Van/Hec for S. aureus, MRSA, and VRSA were 0.5, 3.5, and 5 μM respectively (Table 2). We also used VRE for testing the susceptibility against these agents. Only the Van/Hec conjugate had antimicrobial activity against VRE (Figure S3). The obtained results confirmed the successful preparation of Van/Hec as antimicrobial agents and led us to conclude that Van resistance can be suppressed by Van/Hec.
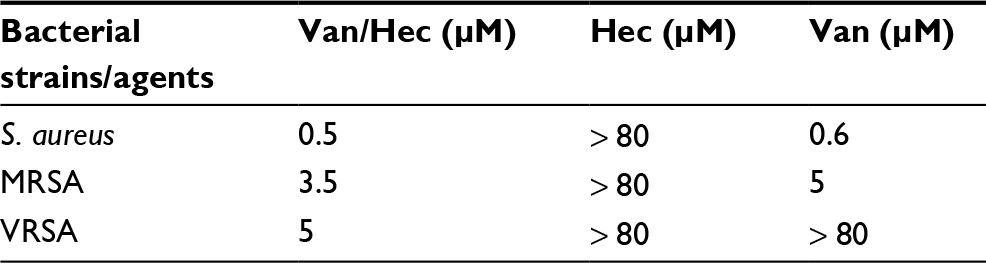 | Table 2 Comparison of antimicrobial agents using MICs determined by broth microdilution method. Abbreviations: MIC, minimum inhibitory concentration; Van, vancomycin; Hec, Hecate; S. aureus, Staphylococcus aureus; MRSA, methicillin-resistant Staphylococcus aureus. |
Influence of Van/Hec on growth properties of bacteria and its undesired toxicity in eukaryotic cells (hemolysis assay)
The effect of Van/Hec was investigated by time-kill assay against S. aureus, MRSA, and VRSA (Figure 2A and B). Van/Hec significantly inhibited bacterial growth of all bacterial strains when compared with the growth control. S. aureus, MRSA, and VRSA showed similar colony count reductions after 24 h of Van/Hec treatment (control 8 and after treatment 2 log10 in CFU/mL). The reduction in bacterial growth was > 3 log10 CFU/mL for all bacterial strains after treatment with Van/Hec, indicative of a bactericidal effect. Van inhibited the growth of S. aureus and MRSA, but not in the case of VRSA. We can also conclude that Hec did not have any beneficial antibacterial effect, or it is necessary to use a higher concentration (MIC > 80 µM). However, after the application of Van/Hec, the bacterial growth started to decrease even in the case of VRSA (Figure 2A and B). This confirms our presumptions that Van/Hec should have better antibacterial activity in comparison with its individual parts or other antibiotics. Finally, the antibacterial effect of Van, peptide Hec, and Van/Hec was confirmed by time-kill analysis against four pathogenic MRSA isolates, and the effect of the agents was similar to that of MRSA from the Czech collection of microorganisms (Figure S4). The effect of the agents has also been established on VRE. The percentage of viable bacterial cells after treatment with Van/Hec confirmed its antimicrobial activity (Figure S3). These results show a significant impact of Van/Hec on weakening the bacterial resistance. The cross-resistance, which is caused by the reduction of membrane permeability to antibiotics and drug inactivation or rapid efflux of the antibiotic, can be avoided using Van/Hec, which provides easier penetration and delivery of the antibiotics when conjugated to CPP.40,41 An experiment was also carried out to check the activity of the conjugate against Gram-negative bacterium E. coli (NCTC 13216). The viability of E. coli cells was studied after 24 h of the treatment with the conjugate using Multiskan EX (Thermo Fisher Scientific). The experiment was repeated five times. The result (Figure S5) indicated that 10 µM of the conjugate was able to kill almost 98% of the cells.
 | Figure 2 Antibacterial activity and cytotoxic analysis. Notes: (A) Time-kill analysis was performed by inoculating ~108 CFU containing 5 µM antibacterial agents onto blood agar. Aliquots were removed at 0, 6, 12, 24 h after inoculation at 37 °C and plated on blood agar plates for CFU calculation. Bactericidal activity was defined as a 3 log10 reduction, and bacteriostatic activity was defined as the maintenance of, or a 3 log10 reduction of, the total number of CFU/mL in the original inoculum. Results are presented in relative units compared with controls (0 h). Data represent the mean ± SD of the mean of three individual experiments. *p < 0.01 or **p < 0.001. (B) Typical photographs of VRSA colonies (CFU/mL) after treatment with Van/Hec conjugate onto the blood agar plates. (C) Cell viability (MTT) assay using nonmalignant PNT1A epithelial cells showing increase in biocompatibility of Van/Hec compared to that of Hec. Values are expressed as the mean of three independent replicates (n = 3). Asterisks indicate significant differences (p < 0.05) compared to the untreated cells. (D) Hemolysis assay of Van, Hec and Van/Hec using red blood human cells. (E) Ambient microscopy of PNT1A cells exposed to Van, Hec, and Van/Hec showing relatively a high amount of apoptotic features in Hec-treated cells. The length of scale bar is 20 µm. Abbreviations: CFU, colony-forming unit; VRSA, vancomycin-resistant Staphylococcus aureus; Van, vancomycin; Hec, Hecate; S. aureus, Staphylococcus aureus; MRSA, methicillin-resistant Staphylococcus aureus. |
Furthermore, cytotoxicity of Van, Hec, and Van/Hec was evaluated using nonmalignant epithelial cell PNT1A (Figure 2C). The cell viability after the application of Van was not found for all the applied concentrations. In contrast, Hec caused significant toxic effects within the higher applied concentrations. However, similar results close to Van were shown after the applications of Van/Hec to eukaryotic cells; the toxicity was estimated to be ~10% for higher concentrations. Similarly, ambient microscopy revealed a significant increase in apoptotic features of Hec-treated cells compared to other treated cells (Figure 2E).
The hemolytic effects of Van were negligible and similar to negative control – PBS (Figure 2D). On the contrary, Hec alone induced a higher level of hemolysis (~15% after treatment with 20 µM). Such hemolytic effects are well known and described for Hec and are one of the major complications of its application in medicine. Despite this, Sanches et al16 described that the coupling of gallic acid at N-terminus of the peptide decreases its activity. This could be the possible explanation, not only for decreasing cytotoxicity in epithelial cells but also for decreased hemotoxicity. Our results showed that the conjugation pronouncedly enhances the biocompatibility of Van/Hec and its antimicrobial effects.
Microscopic estimation of live/dead cells and damage to DNA (bacterial and human cells)
The antibacterial effect of Van, peptide Hec, and Van/Hec after applications for S. aureus, MRSA, and VRSA were confirmed under ambient light illumination by an optical microscope (Figure 3). Decreasing numbers of all bacterial strains after treatment with agents were shown in comparison with those of the control groups. Van/Hec was found to rupture the cell wall of S. aureus, MRSA and VRSA.
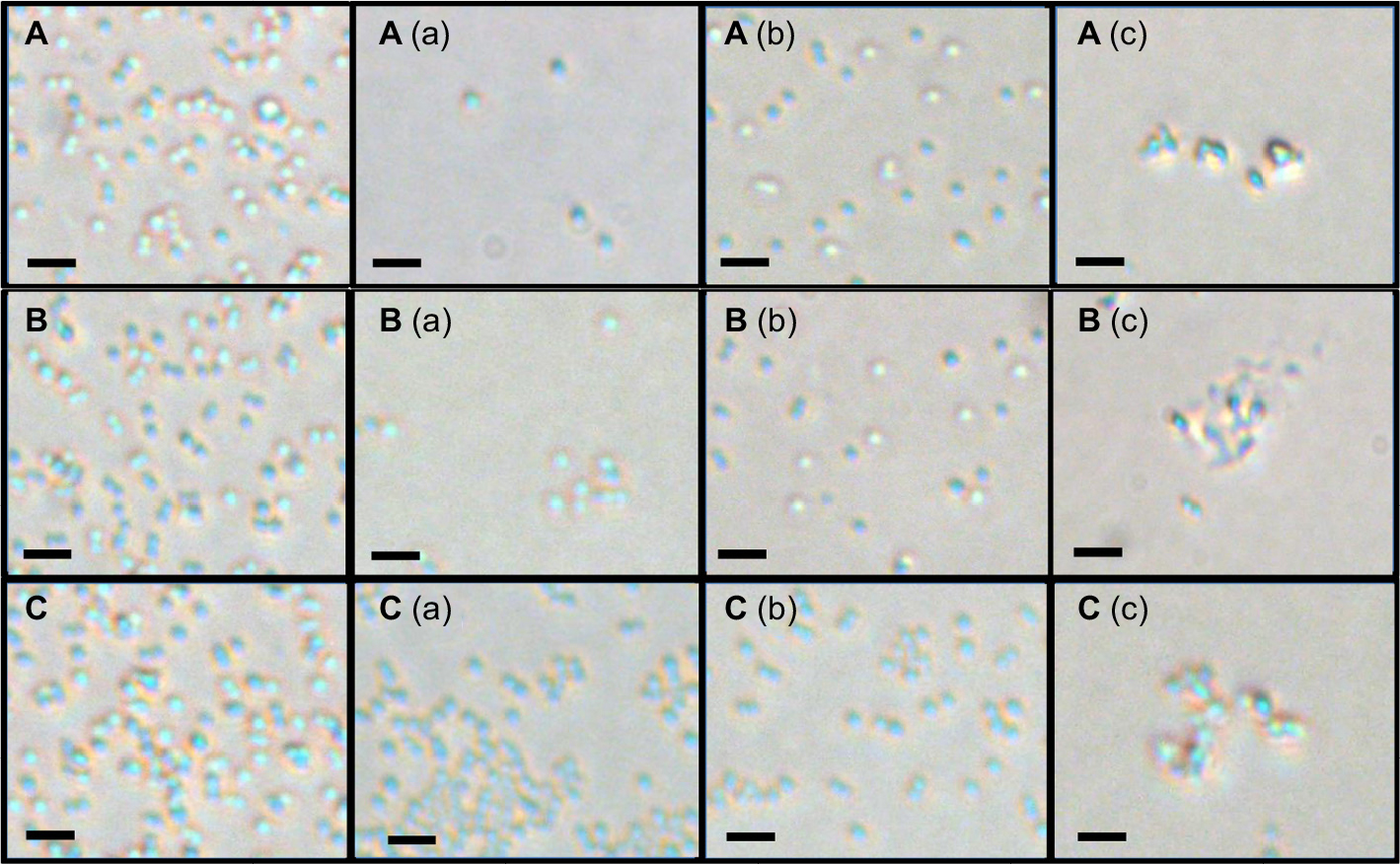 | Figure 3 Bacterial cells. Note: (A) S. aureus, (B) MRSA, and (C) VRSA after applications of 5 µM antibacterial agents: (a) Van, (b) peptide Hec, and (c) Van/Hec conjugate using optical microscope in ambient light. Scale bar is 2 µm; magnification is 600¥. Abbreviations: S. aureus, Staphylococcus aureus; MRSA, methicillin-resistant Staphylococcus aureus; VRSA, vancomycin-resistant Staphylococcus aureus; Van, vancomycin; Hec, Hecate. |
The viability of the tested bacterial strains after the addition of the antimicrobial agents was further investigated using live/dead assay by fluorescence microscopy. To quantify live and dead bacterial cells, two fluorescent dyes were used. SYTO9 indicates live bacteria in green fluorescent color because it can permeate cell membrane, while PI showed dead bacteria in red fluorescent color with destroyed membrane.29 The bacterial strains without the application of Van, Hec, and Van/Hec were used as the control group. The staining of live cells (control group) can be observed by SYTO9 as green color. After the treatment of Van, Hec, and Van/Hec with bacterial strains, a decrease in cell viability can be seen as a red color (PI). It was confirmed that the combination of Van and peptide Hec after application on the bacteria (Figure S6) contributes to the inhibition of the growth of the bacterial cells and even for resistant strains of bacteria to Van (Figure 4A).
 | Figure 4 Representative live/dead cell assay micrographs of VRSA. Notes: (Aa) Negative control, (Ab) Van, (Ac) peptide Hec, and (Ad) Van/Hec by using two fluorescent dyes (PI/SYTO9). The length of scale bar is 100 µm. Single-cell gel electrophoresis assay showing DNA damage of VRSA after applications of 5 µM antibacterial agents. (Ba) Negative control, (Bb) positive control – H2O2 (150 µM), (Bc) Van, (Bd) peptide Hec, and (Be) Van/Hec. Scale bar is 2 µm. Comet formed 2 h after treatment of bacteria with agents indicating damage of bacterial DNA (Bd and Be) and normal bacterial DNA (Ba, Bc, and Bd), which remained intact after SCGE assay. (Ca) Comet grade and (Cb) index of damage. |
DNA damage of VRSA after treatment with antibacterial agents (positive control – H2O2 150 µM, Van, peptide Hec, and Van/Hec) was shown by the single-cell gel electrophoresis assay (comet assay). Comet tails were formed 2 h after treatment of bacteria with agents indicating bacterial DNA damage. Normal bacterial DNA without treatment has been depicted as a negative control, which remained intact after the single cell gel electrophoresis (SCGE) assay. Damage of bacterial DNA was found in the case of each antibacterial agent for S. aureus, MRSA, and VRSA (Figures 4B and S7). Comet grade and index of damage for all bacterial strains were the largest (grade 4) and most visible after treatment with Van/Hec (Figures 4C and S7C and D).
Comet assay has been also done for testing genotoxicity in the human cells – PNT1A (Figure S8). The human cells were treated for 2 h with antibacterial agents and positive control (H2O2). Insignificant genotoxicity was found after treatment with the antibacterial agents in comparison with positive control (H2O2). Our results demonstrate that our designed Van/Hec is highly biocompatible with eukaryotic organisms. Therefore, the Van/Hec could be considered as a promising antimicrobial agent toward VRSA.
Confirmation of presence of vanA gene in VRSA
In our study, we focused on confirmation of the presence of vanA gene coding the ligase responsible for the synthesis of D-Ala-D-Lac dipeptide that seems to be essential for the development of high-level Van resistance in VRSA strains.42 We verified the presence of vanA gene in the VRSA strain (Figure S9). This result is in compliance with the data from MIC that showed high-level Van resistance of tested strain (MIC > 80 μM) typical for the presence of vanA gene cluster.9
Conclusion
In the present experiment, we developed a new potential antimicrobial agent by conjugating a common antibiotic (Van) and a CPP peptide (Hec) linked through a zero-length linker (CDI) to overcome the limitation of the use of antibiotics in the treatment of the resistant and non-resistant S. aureus strains, including MRSA and VRSA. Results from disk diffusion test, time-kill assay, determination of MIC, microscopy, and comet assay showed strong antimicrobial effects of Van/Hec conjugate against wild-type, methicillin- and Van-resistant staphylococcal strains (MRSA and VRSA). Microscopic results revealed that the exposure to Van/Hec conjugate caused disruption of bacterial cell integrity in all the tested strains, which was not found in case of Van and Hec alone. The conjugate has been found to be nontoxic, which was confirmed by different assays on nonmalignant epithelial cells and human RBCs. Overall, Van/Hec conjugate appears to be a promising antibacterial agent, which can help us to overcome the limitation of the use of antibiotic for the treatment of the infections caused by resistant Staphylococcal strains including MRSA and VRSA.
Acknowledgments
This work was supported by Internal Grant Agency of Mendel University in Brno (IGA IP_10/2017) and CEITEC 2020 (LQ 1601) from the Ministry of Education, Youth and Sports of the Czech Republic under the National Sustainability Programme II. The authors wish to express special gratitude to Dr. Mark Holmes and his work groups from Veterinary Medicine Department of University of Cambridge (UK) for providing resistant bacterial strains and technical help. The founding sponsors had no role in the design of the study; collection, analyses, or interpretation of data; writing of the manuscript, and decision to publish the results.
Disclosure
The authors report no conflicts of interest in this work.
References
Tan RM, Liu JL, Li ML, Huang J, Sun JY, Qu HP. Epidemiology and antimicrobial resistance among commonly encountered bacteria associated with infections and colonization in intensive care units in a university-affiliated hospital in Shanghai. J Microbiol Immunol Infect. 2014;47(2):87–94. | ||
McNeece G, Naughton V, Woodward MJ, Dooley JSG, Naughton PJ. Array based detection of antibiotic resistance genes in Gram negative bacteria isolated from retail poultry meat in the UK and Ireland. Int J Food Microbiol. 2014;179:24–32. | ||
Juknius T, Tamulevicius T, Grazuleviciute I, Klimiene I, Matusevicius AP, Tamulevicius S. In-situ measurements of bacteria resistance to antimicrobial agents employing leaky mode sub-wavelength diffraction grating. Sens Actuator B Chem. 2014;204:799–806. | ||
Wang RX, Wang AL, Wang JY. Antibiotic resistance monitoring in heterotrophic bacteria from anthropogenic-polluted seawater and the intestines of oyster Crassostrea hongkongensis. Ecotoxicol Environ Saf. 2014;109:27–31. | ||
Wang HH, Schaffner DW. Antibiotic resistance: how much do we know and where do we go from here? Appl Environ Microbiol. 2011;77(20):7093–7095. | ||
Cui LZ, Iwamoto A, Lian JQ, et al. Novel mechanism of antibiotic resistance originating in vancomycin-intermediate Staphylococcus aureus. Antimicrob Agents Chemother. 2006;50(2):428–438. | ||
Chang S, Sievert DM, Hageman JC, et al; Vancomycin-Resistant Staphylococcus aureus Investigative Team. Infection with vancomycin-resistant Staphylococcus aureus containing the vanA resistance gene. N Engl J Med. 2003;348(14):1342–1347. | ||
Chung DR, Baek JY, Kim HA, et al. First report of vancomycin-intermediate resistance in sequence type 72 community genotype methicillin-resistant Staphylococcus aureus. J Clin Microbiol. 2012;50(7):2513–2514. | ||
Weigel LM, Clewell DB, Gill SR, et al. Genetic analysis of a high-level vancomycin-resistant isolate of Staphylococcus aureus. Science. 2003;302(5650):1569–1571. | ||
Courvalin P. Vancomycin resistance in gram-positive cocci. Clin Infect Dis. 2006;42(suppl 1):S25–S34. | ||
Alexander EL, Gardete S, Bar HY, Wells MT, Tomasz A, Rhee KY. Intermediate-type vancomycin resistance (VISA) in genetically-distinct Staphylococcus aureus isolates is linked to specific, reversible metabolic alterations. PLoS One. 2014;9(5):1–9. | ||
Zhu WL, Lan HL, Park IS, et al. Design and mechanism of action of a novel bacteria-selective antimicrobial peptide from the cell-penetrating peptide Pep-1. Biochem Biophys Res Commun. 2006;349(2):769–774. | ||
Groh T, Hrabeta J, Khalil MA, Doktorova H, Eckschlager T, Stiborova M. The synergistic effects of DNA-damaging drugs cisplatin and etoposide with a histone deacetylase inhibitor valproate in high-risk neuroblastoma cells. Int J Oncol. 2015;47(1):343–352. | ||
Piecyk K, Jankowska-Anyszka M. Chemical conjugation of an mRNA cap analogue with a cell-penetrating peptide as a potential membrane permeable translation inhibitor. Tetrahedron Lett. 2014;55(3):606–609. | ||
Montenegro J, Matile S. Anionic activators for differential sensing with cell-penetrating peptides. Chem Asian J. 2011;6(2):681–689. | ||
Sanches PRS, Carneiro BM, Batista MN, et al. A conjugate of the lytic peptide Hecate and gallic acid: structure, activity against cervical cancer, and toxicity. Amino Acids. 2015;47(7):1433–1443. | ||
Rivero-Muller A, Vuorenoja S, Tuominen M, et al. Use of hecate-chorionic gonadotropin beta conjugate in therapy of lutenizing hormone receptor expressing gonadal somatic cell tumors. Mol Cell Endocrinol. 2007;269(1–2):17–25. | ||
Vicente EF, Basso LGM, Cespedes GF, et al. Dynamics and conformational studies of TOAC spin labeled analogues of Ctx(Ile(21))-Ha peptide from Hypsiboas albopunctatus. PLoS One. 2013;8(4):1–12. | ||
Chudobova D, Cihalova K, Dostalova S, et al. Comparison of the effects of silver phosphate and selenium nanoparticles on Staphylococcus aureus growth reveals potential for selenium particles to prevent infection. FEMS Microbiol Lett. 2014;351(2):195–201. | ||
Wright SJL. A simple agar plate method, using micro-algae, for herbicide bio-assay or detection. Bull Environ Contam Toxicol. 1975;14(1):65–70. | ||
Valgas C, de Souza SM, Smania EFA, Smania A. Screening methods to determine antibacterial activity of natural products. Braz J Microbiol. 2007;38(2):369–380. | ||
Wang HL, Hesseltine CW, Ellis JJ. Antibacterial activity produced by molds commonly used in oriental food fermentations. Mycologia. 1972;64(1):218–221. | ||
Balouiri M, Sadiki M, Ibnsouda SK. Methods for in vitro evaluating antimicrobial activity: a review. J Pharm Anal. 2016;6(2):71–79. | ||
Sakoulas G, Moise-Broder PA, Schentag J, Forrest A, Moellering RC, Eliopoulos GM. Relationship of MIC and bactericidal activity to efficacy of vancomycin for treatment of methicillin-resistant Staphylococcus aureus bacteremia. J Clin Microbiol. 2004;42(6):2398–2402. | ||
Petersen PJ, Jones CH, Bradford PA. In vitro antibacterial activities of tigecycline and comparative agents by time-kill kinetic studies in fresh Mueller-Hinton broth. Diagn Microbiol Infect Dis. 2007;59(3):347–349. | ||
Pankey GA, Ashcraft DS. In vitro antibacterial activity of tigecycline against resistant Gram-negative bacilli and enterococci by time-kill assay. Diagn Microbiol Infect Dis. 2009;64(3):300–304. | ||
Petersen PJ, Wang TZ, Dushin RG, Bradford PA. Comparative in vitro activities of AC98-6446, a novel semisynthetic glycopeptide derivative of the natural product mannopeptimycin alpha, and other antimicrobial agents against gram-positive clinical isolates. Antimicrob Agents Chemother. 2004;48(3):739–746. | ||
Heger Z, Rodrigo MAM, Michalek P, et al. Sarcosine up-regulates expression of genes involved in cell cycle progression of metastatic models of prostate cancer. PLoS One. 2016;11(11):1–20. | ||
Berney M, Hammes F, Bosshard F, Weilenmann HU, Egli T. Assessment and interpretation of bacterial viability by using the LIVE/DEAD BacLight kit in combination with flow cytometry. Appl Environ Microbiol. 2007;73(10):3283–3290. | ||
Hiramatsu K, Kayayama Y, Matsuo M, et al. Vancomycin-intermediate resistance in Staphylococcus aureus. J Glob Antimicrob Resist. 2014;2(4):213–224. | ||
Reinhardt A, Neundorf I. Design and application of antimicrobial peptide conjugates. Int J Mol Sci. 2016;17(5):701. | ||
Leuschner C, Hansel W. Membrane disrupting lytic peptides for cancer treatments. Curr Pharm Des. 2004;10(19):2299–2310. | ||
Yarlagadda V, Konai MM, Manjunath GB, Ghosh C, Haldar J. Tackling vancomycin-resistant bacteria with ‘lipophilic-vancomycin-carbohydrate conjugates’. J Antibiot. 2015;68(5):302–312. | ||
Suller MTE, Russell AD. Triclosan and antibiotic resistance in Staphylococcus aureus. J Antimicrob Chemother. 2000;46(1):11–18. | ||
Sabath LD, Garner C, Wilcox C, Finland M. Susceptibility of Staphylococcus aureus and Staphylococcus epidermidis to 65 antibiotics. Antimicrob Agents Chemother. 1976;9(6):962–969. | ||
Ligozzi M, Bernini C, Bonora MG, de Fatima M, Zuliani J, Fontana R. Evaluation of the VITEK 2 system for identification and antimicrobial susceptibility testing of medically relevant gram-positive cocci. J Clin Microbiol. 2002;40(5):1681–1686. | ||
Andrews JM. Determination of minimum inhibitory concentrations. J Antimicrob Chemother. 2001;48(suppl 1):5–16. | ||
Abb J. In vitro activity of linezolid, quinupristin-dalfopristin, vancomycin, teicoplanin, moxifloxacin and mupirocin against methicillin-resistant Staphylococcus aureus: comparative evaluation by the E test and a broth microdilution method. Diagn Microbiol Infect Dis. 2002;43(4):319–321. | ||
Goudarzi M, Goudarzi H, Figueiredo AMS, et al. Molecular characterization of methicillin resistant Staphylococcus aureus strains isolated from intensive care units in Iran: ST22-SCCmec IV/t790 emerges as the major clone. PLoS One. 2016;11(5):1–13. | ||
Baker-Austin C, Wright MS, Stepanauskas R, McArthur JV. Co-selection of antibiotic and metal resistance. Trends Microbiol. 2006;14(4):176–182. | ||
Hallbrink M, Oehlke J, Papsdorf G, Bienert M. Uptake of cell-penetrating peptides is dependent on peptide-to-cell ratio rather than on peptide concentration. Biochim Biophys Acta. 2004;1667(2):222–228. | ||
Perichon B, Courvalin P. VanA-type vancomycin-resistant Staphylococcus aureus. Antimicrob Agents Chemother. 2009;53(11):4580–4587. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.