Back to Journals » Infection and Drug Resistance » Volume 11
Novel mechanism responsible for high-level macrolide resistance in Moraxella catarrhalis
Authors Kasai A, Ohta A, Maeda Y, Yamada K ![]() , Tao K, Saito R
, Tao K, Saito R ![]()
Received 28 July 2018
Accepted for publication 2 October 2018
Published 1 November 2018 Volume 2018:11 Pages 2137—2140
DOI https://doi.org/10.2147/IDR.S181714
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Suresh Antony
Ayako Kasai,1 Ayaka Ohta,1 Yuina Maeda,1 Kageto Yamada,2 Kazuyuki Tao,3 Ryoichi Saito1
1Department of Molecular Microbiology, Graduate School of Medical and Dental Sciences, Tokyo Medical and Dental University, Bunkyo-ku, Tokyo 113-8510, Japan; 2Department of Clinical Laboratory, Toho University Medical Center Omori Hospital, Ota-ku, Tokyo 143-8541, Japan; 3Radioisotope Center, University of Tokyo, Bunkyo-ku, Tokyo 113-0032, Japan
Background: High-level macrolide-resistant Moraxella catarrhalis strains have been isolated; however, the underlying mechanism has not been well elucidated. We investigated the role of mutations in the 23S rRNA gene and the L4 and L22 ribosomal proteins using spontaneous erythromycin-resistant mutants and transformants.
Materials and methods: The erythromycin-susceptible M. catarrhalis ATCC25238 and clinical isolate Mc19 were used as parental strains. To obtain spontaneous erythromycin-resistant mutants, in vitro stepwise selection was performed using brain–heart infusion agar plates containing various concentrations of erythromycin. The role of the mutations identified in the spontaneous mutants was validated using transformation experiments.
Results: We obtained two spontaneous mutants with high-level resistance to erythromycin, S25-32-af10 and S19-256-af10, from ATCC25238 and Mc19, respectively. S25-32-af10 exhibited mutations of Q61R in L4 and Insertion98SRADRIS in L22. S19-256-af10 exhibited three C2611T-mutated alleles in the 23S rRNA gene and G65A in L4. Transformants with single mutations identified in S25-32-af10 or S19-256-af10 showed higher erythromycin and azithromycin minimum inhibitory concentrations (MICs) than those of each parental strain. However, transformants with multiple mutations identified in S25-32-af10 or S19-256-af10 showed macrolide MICs similar to those of each parental strain.
Conclusion: Our results provide the first evidence suggesting that Q61R in L4 and Insertion98SRADRIS in L22 are involved in the synergistic acquisition of high-level resistance to both 14- and 15-member macrolides, and that C2611T in the 23S rRNA gene and G65A in L4 also synergistically contribute toward conferring high-level 14-member macrolide resistance to M. catarrhalis.
Keywords: macrolide, Moraxella catarrhalis, ribosomal protein, 23S rRNA gene
Introduction
Moraxella catarrhalis is a common pathogen involved in lower and upper respiratory tract infections in both children and elderly people.1 The 14-membered macrolide clarithromycin and 15-membered macrolide azithromycin are used for pharmacological treatment of M. catarrhalis infections.2,3 However, M. catarrhalis clinical isolates with high-level resistance against these macrolides have been reported in both Japan4,5 and China.6
The mechanisms underlying macrolide resistance in the diplococci Neisseria gonorrhoeae and Streptococcus pneumoniae are associated with active efflux, drug inactivation, and changes in the ribosome target sites.7 Our previous studies have shown that an A2058T mutation in three or four alleles of the 23S rRNA gene of M. catarrhalis confers high-level macrolide resistance.4,5 Liu et al also demonstrated that high-level macrolide-resistant strains possessed a number of mutations in the 23S rRNA gene6 and that several genes, excluding the 23S rRNA gene and the L4 and L22 ribosomal protein-encoding genes, may be related to macrolide resistance in M. catarrhalis.8 However, their role in macrolide resistance remains unclear because molecular functional analysis has not been performed for several of the genes identified in those studies.
In this study, we focused on the 23S rRNA gene and the L4 and L22 ribosomal proteins, established as the major targets of macrolide resistance in bacteria,7 and examined their involvement in the acquisition of high-level macrolide resistance by analyzing in vitro erythromycin-resistant spontaneous mutants and transformants.
Materials and methods
To generate M. catarrhalis erythromycin-resistant spontaneous mutants, pre-cultured erythromycin-susceptible reference strain ATCC25238 and clinical isolate Mc19 (minimum inhibitory concentration [MIC] of erythromycin for both =0.06 mg/L) were suspended in Tris–EDTA (TE) buffer to a concentration equivalent to a McFarland standard of 4.0, and a 0.2 mL aliquot of the suspension was spread on brain–heart infusion (BHI; Oxoid, Basingstoke, UK) agar plates containing 0.25, 0.5, or 1 mg/L of erythromycin. Each plate was incubated at 37°C under 5% CO2 until colonies became visible. A single colony of each strain was plated on another BHI agar plate with the same concentration of erythromycin and this step was then repeated at least three times. Subsequently, a single colony of each strain was harvested on to a BHI agar plate containing a twofold higher concentration of erythromycin than the previous passage. The erythromycin concentration was gradually increased until no colony growth was observed.
The erythromycin, clarithromycin, azithromycin, and clindamycin MICs were determined using the E-test (bioMérieux Japan, Tokyo, Japan) according to the manufacturer’s recommendations. Interpretation of erythromycin, clarithromycin, and azithromycin breakpoints was based on the European Committee on Antimicrobial Susceptibility Testing (EUCAST) Version 8.1 criteria (http://www.eucast.org/clinical_breakpoints/). Interactive criteria for clindamycin followed Clinical and Laboratory Standards Institute (CLSI) document M45-A2.9
PCR amplification and DNA sequencing of the 23S rRNA gene and L4 and L22 ribosomal protein genes were performed as previously described.4,5 Sequences from in vitro erythromycin-resistant spontaneous mutants were compared to those of the parental strains.
Transformation experiments were performed as per our previous study,4 with some modifications. In brief, PCR fragments with mutations in the macrolide-resistance targets were amplified as previously described using the following primer sets: 23SrRNA-F3 (5′-CTCTGGGGATAACAGGCTGA-3′) and 23SrRNA-R3 (5′-CAACCGAAACACCAGAGGTT-3′) for the C2611T substitution, L4-24F and L4-589R4 for both the G65A and Q61R substitutions, and L22-32F and L22-325R4 for insertion of the 98SRADRIS mutation. Parental strains ATCC25238 or Mc19 were suspended in TE buffer, centrifuged, and then inoculated on a BHI agar plate with approximately 1 µg of amplified PCR fragment. After incubation for 7 hours, the harvested colonies were plated on a BHI agar plate containing 0.25, 1, and 4 mg/L of erythromycin. For grown transformant candidates, the presence of mutations in macrolide-resistance targets and the MICs for each antibiotic were determined as described above.
Efflux activity was examined using two efflux pump inhibitors (EPIs), phenylalanine arginine β-naphthylamide (PAβN; Sigma-Aldrich Co., St Louis, MO, USA) and carbonyl cyanide m-chlorophenylhydrazone (CCCP; Nacalai Tesque, Kyoto, Japan), as previously described.10 Overexpression was defined as greater than a two-fold decrease in erythromycin MICs with or without the EPIs. PAβN contributes toward increasing antibiotic levels in the bacterial cells by enhancing the permeability of cell membranes. CCCP is an uncoupler that can block up the energy resources of efflux pumps.
Results and discussion
M. catarrhalis with high-level macrolide resistance is extremely rare,4–6,11,12 and little is known regarding the underlying resistance mechanism(s). Therefore, we investigated the mechanism related to novel mutations in the 23S rRNA and L4 and L22 ribosomal proteins. In this study, we acquired two spontaneous mutants, S25-32-af10 (derived from 25238) and S19-256-af10 (derived from Mc19), with high-level erythromycin resistance (MIC >256 mg/L) by erythromycin passage (Table 1). S25-32-af10 showed high-level resistance to clarithromycin and azithromycin, but low-level resistance to clindamycin, similar to the parental strain. S19-256-af10 demonstrated high-level resistance to clarithromycin, but low-level resistance to azithromycin and clindamycin. In addition, S25-32-af10 harbored a Q61R substitution and a seven-amino acid insertion (Insertion98SRADRIS) in L4 and L22, respectively (Table 1). In contrast, S19-256-af10 exhibited C2611T mutations in three 23S rRNA alleles, as well as a G65A substitution in L4. However, mutants of these strains did not contain a A2058T mutation in the 23S rRNA gene, which is known to cause high-level macrolide resistance.4 Analysis of the efflux system demonstrated that the erythromycin MICs of S25-32-af10 and S19-256-af10 remained unchanged with or without EPI (data not shown). Stepwise azithromycin selection of S. pneumoniae strains revealed that mutations in either the 23S rRNA alleles or ribosomal protein L4 were responsible for macrolide resistance.13 In addition, in Campylobacter jejuni, a combination of amino acid mutations in L4 and L22 resulted in the development of high-level erythromycin resistance.14 Furthermore, in the Gram-negative diplococcus N. gonorrhoeae, the mutation of three or four alleles at position 2,611 of the 23S rRNA gene was responsible for moderate levels of macrolide resistance.15 Collectively, these results strongly suggest that high-level macrolide resistance in S25-32-af10 and S19-256-af10 resulted from multiple mutations in L4, L22, or the 23S rRNA gene.
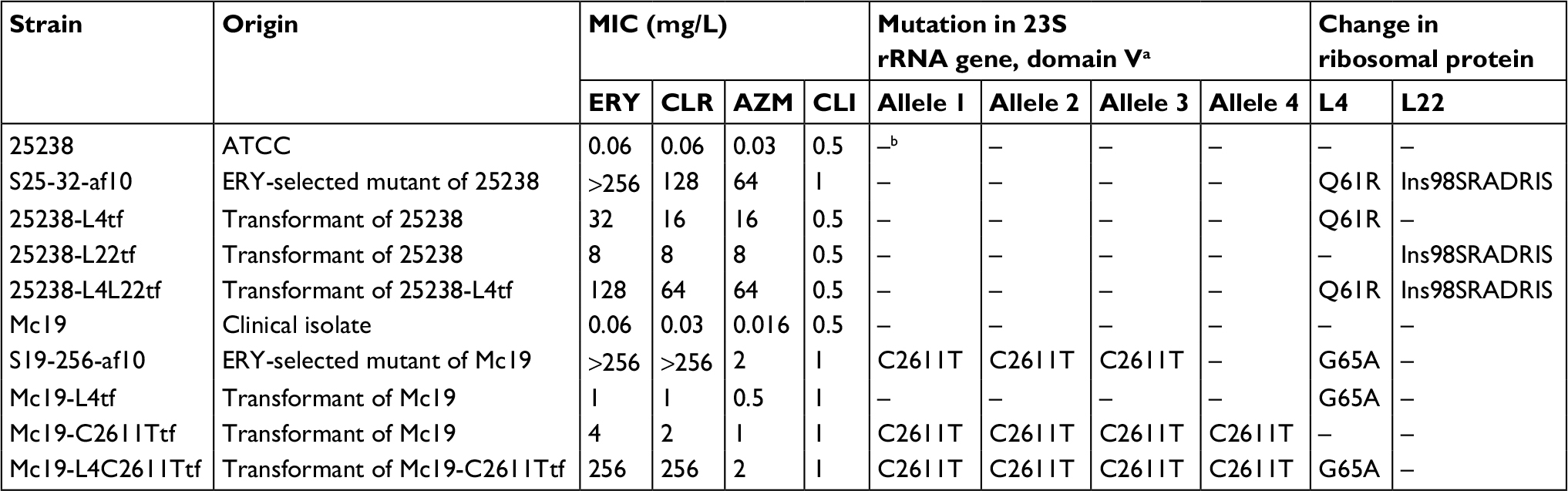 | Table 1 MICs and mechanisms underlying macrolide resistance in parent strains, in vitro erythromycin-resistant spontaneous mutants, and transformants of Moraxella catarrhalis Notes: aEscherichia coli numbering (GenBank accession number V00331); bno mutation detected. Abbreviations: ATCC, American Type Culture Collection; MIC, minimum inhibitory concentration; ERY, erythromycin; CLR, clarithromycin; AZM, azithromycin; CLI, clindamycin; Ins, insert. |
To confirm this hypothesis, we next performed transformation experiments (Table 1). The transformants 25238-L4tf harboring a Q61R mutation in L4 and 25238-L22tf with an insertion (Insertion98SRADRIS) in L22 showed moderate resistance to erythromycin, clarithromycin, and azithromycin. In contrast, 25238-L4L22tf containing both mutations showed macrolide MIC profiles very similar to spontaneous mutant S25-32-af10, indicating that although Q61R in L4 or the seven-amino acid insertion in L22 independently cause the acquisition of moderate-level resistance against both 14- and 15-member macrolides, the mutations synergistically contribute toward conferring high-level resistance. Conversely, the transformants Mc19-L4tf harboring a G65A substitution in L4 and Mc19-C2611Ttf with four C2611T-mutated alleles exhibited low-level resistance to erythromycin, clarithromycin, and azithromycin. The G65A substitution in Mc19-L4tf was identified in our previous study using clinical isolates.5 A high-level erythromycin-resistant transformant, Mc19-C2611TL4tf, harboring both a G65A substitution and four C2611T-mutated alleles in the 23S rRNA had MIC profiles nearly identical to spontaneous mutant S19-256-af10, indicating that these mutations synergistically cause high-level 14-member macrolide resistance. Coupling our results with previous findings,7,13–15 it becomes clear that a mechanism other than A2058T substitutions in the 23S rRNA gene is involved in the development of high-level macrolide resistance in M. catarrhalis. However, we focused only on mutations in the 23S rRNA gene and the ribosomal proteins L4 and L22, known as the key targets for macrolide resistance in the Gram-negative diplococci M. catarrhalis and N. gonorrhoeae; therefore, a genome-wide study of our spontaneous erythromycin-resistant mutants may be required for more in-depth analysis of the mechanisms underlying macrolide resistance.
Conclusion
To the best of our knowledge, we provide the first evidence indicating that 1) amino acid mutations in both L4 (Q61R) and L22 (Insertion98SRADRIS) are involved in the synergistic acquisition of high-level resistance against both 14- and 15-member macrolides and 2) two mechanisms, C2611T substitutions in the 23S rRNA gene and an amino acid substitution in L4 (G65A), also synergistically contribute toward the development of high-level resistance to 14-member macrolides in M. catarrhalis. Understanding the mechanisms underlying macrolide resistance can aid in the development of treatments against high-level macrolide-resistant M. catarrhalis infections and the prevention of their spread. In addition, our findings emphasize the need for continuous surveillance of macrolide resistance in clinical isolates of M. catarrhalis in future studies.
Accession numbers
Nucleotide sequences of the ATCC 25238 strain and erythromycin-resistant spontaneous mutants obtained in this study were deposited in GenBank/EMBL/DDBJ, and their accession numbers are LC386088 to LC386093 and LC386096 to LC386102, respectively.
Acknowledgment
Ayako Kasai’s present address is at Department of Clinical Laboratory, Tokyo Metropolitan Tama Medical Center, Fuchu-shi, Tokyo 183-8524, Japan.
Author contributions
All authors contributed toward data analysis, drafting and critically revising the paper, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
Murphy TF, Parameswaran GI. Moraxella catarrhalis, a human respiratory tract pathogen. Clin Infect Dis. 2009;49(1):124–131. | ||
Coco A, Vernacchio L, Horst M, Anderson A. Management of acute otitis media after publication of the 2004 AAP and AAFP clinical practice guideline. Pediatrics. 2010;125(2):214–220. | ||
Seemungal TA, Wilkinson TM, Hurst JR, Perera WR, Sapsford RJ, Wedzicha JA. Long-term erythromycin therapy is associated with decreased chronic obstructive pulmonary disease exacerbations. Am J Respir Crit Care Med. 2008;178(11):1139–1147. | ||
Saito R, Nonaka S, Nishiyama H, Okamura N. Molecular mechanism of macrolide-lincosamide resistance in Moraxella catarrhalis. J Med Microbiol. 2012;61(Pt 10):1435–1438. | ||
Kasai A, Ogihara S, Yamada K, Tanimichi Y, Nishiyama H, Saito R. Prevalence and molecular analysis of macrolide-resistant Moraxella catarrhalis clinical isolates in Japan, following emergence of the highly macrolide-resistant strain NSH1 in 2011. J Med Microbiol. 2015;64(7):708–713. | ||
Liu Y, Zhao C, Zhang F, et al. High prevalence and molecular analysis of macrolide-nonsusceptible Moraxella catarrhalis isolated from nasopharynx of healthy children in China. Microb Drug Resist. 2012;18(4):417–426. | ||
Roberts MC. Update on macrolide-lincosamide-streptogramin, ketolide, and oxazolidinone resistance genes. FEMS Microbiol Lett. 2008;282(2):147–159. | ||
Liu YL, Li DF, Xu HP, Chen H, Chen M, Wang H. Use of next generation sequence to investigate potential novel macrolide resistance mechanisms in a population of Moraxella catarrhalis isolates. Sci Rep. 2016;6:35711. | ||
Clinical and Laboratory Standards Institute. M45-A2. Methods for Antimicrobial Dilution and Disk Susceptibility Testing for Infrequently Isolated or Fastidious Bacteria; Approved Guideline – Second Edition. Wayne, PA: CLSI; 2010. | ||
Nonaka S, Matsuzaki K, Kazama T, et al. Antimicrobial susceptibility and mechanisms of high-level macrolide resistance in clinical isolates of Moraxella nonliquefaciens. J Med Microbiol. 2014;63(Pt 2):242–247. | ||
Flamm RK, Sader HS, Farrell DJ, Jones RN. Macrolide and tetracycline resistance among Moraxella catarrhalis isolates from 2009 to 2011. Diagn Microbiol Infect Dis. 2012;74(2):198–200. | ||
Iwata S, Sato Y, Toyonaga Y, Hanaki H, Sunakawa K. Genetic analysis of a pediatric clinical isolate of Moraxella catarrhalis with resistance to macrolides and quinolones. J Infect Chemother. 2015;21(4):308–311. | ||
Tait-Kamradt A, Davies T, Cronan M, Jacobs MR, Appelbaum PC, Sutcliffe J. Mutations in 23S rRNA and ribosomal protein L4 account for resistance in pneumococcal strains selected in vitro by macrolide passage. Antimicrob Agents Chemother. 2000;44(8):2118–2125. | ||
Hao H, Yuan Z, Shen Z, et al. Mutational and transcriptomic changes involved in the development of macrolide resistance in Campylobacter jejuni. Antimicrob Agents Chemother. 2013;57(3):1369–1378. | ||
Ng LK, Martin I, Liu G, Bryden L. Mutation in 23S rRNA associated with macrolide resistance in Neisseria gonorrhoeae. Antimicrob Agents Chemother. 2002;46(9):3020–3025. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.