Back to Journals » Drug Design, Development and Therapy » Volume 13
Notopterol-induced apoptosis and differentiation in human acute myeloid leukemia HL-60 cells
Authors Huang Q ![]() , Wang L
, Wang L ![]() , Ran Q, Wang J, Wang C
, Ran Q, Wang J, Wang C ![]() , He H, Li L, Qi H
, He H, Li L, Qi H ![]()
Received 7 October 2018
Accepted for publication 4 April 2019
Published 6 June 2019 Volume 2019:13 Pages 1927—1940
DOI https://doi.org/10.2147/DDDT.S189969
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Tuo Deng
Qinwan Huang,1 Lin Wang,1 Qian Ran,1 Jin Wang,1 Chengqiang Wang,2 Hui He,2 Li Li,2 Hongyi Qi2
1College of Pharmacy, Chengdu University of Traditional Chinese Medicine, Chengdu 611137, People’s Republic of China; 2College of Pharmaceutical Sciences, Southwest University, Chongqing 400716, People’s Republic of China
Purpose: This study aims to observe the effects of notopterol on the apoptosis and differentiation of HL-60 cells and to explore the underlying molecular mechanisms.
Methods: Cell viability was assessed using sulforhodamine B assay. Cell proliferation was determined by the trypan blue dye exclusion test. Colony-forming units were assayed in methylcellulose. Apoptosis assays were carried out by annexin V-fluorescein isothiocyanate(FITC)/propidium iodide (PI) double staining, Hoechst 33342 staining, mitochondrial membrane potential, and Western blot. Wright–Giemsa staining, nitroblue tetrazolium (NBT) reduction assay, CD11b and CD14 and Western blot were detected for induction of differentiation. In addition, cell-cycle phase distribution was analyzed by flow cytometry and Western blot. The combination therapy of notopterol and all-trans retinoic acid (ATRA) on HL-60 cells was examined.
Results: Notopterol obviously inhibited the growth of HL-60 cells with an IC50 value of 40.32 μM and remarkably reduced the number of colonies by 10, 20, and 40 μM. In addtion, notopterol induced the percentage of apoptotic HL-60 cells, reduced the mitochondrial membrane potential, decreased the protein expresstion of Bcl-2 and Mcl-1, and increased the expression of Bax, cleavage of caspase 9, caspase 3, and PARP. As for cell differentiation, notopterol clearly induced chromatin condensation; increased the nucleocytoplasmic ratio, nitroblue tetrazolium-positive cells, expression of CD14 and CD11b, and protein expression of c-Jun and Jun B, and decreased c-myc. Furthermore, notopterol induced the G0/G1 cell-cycle arrest as determined using flow cytometry, which may be related to the regulation of cell-cycle-related proteins p53, CDK2, CDK4, Cyclin D1, Cyclin E, and survivin. The combined use of notopterol and ATRA did not enhance the apoptotic effect as evidenced by cell viability test and Hoechst 33342. However, the combination of notopterol and ATRA enhanced the effect of inducing differentiation when compared with using either notopterol or ATRA alone, which can be evidenced by the increased nucleocytoplasmic ratio, NBT positive cells, and expression of CD14.
Conclusion: This is the first time it has been demonstrated that notopterol could induce apoptosis, differentiation, and G0/G1 arrest in human AML HL-60 cells, suggesting that notopterol has potential therapeutic effects on AML. The combination application of notopterol (20 and 40 μM) and ATRA (2 μM) could augment differentiation of HL-60 cells.
Keywords: notopterol, cell growth, apoptosis, differetiation, cycle arrest, AML
Introduction
Leukemia is a heterogeneous clonal disorder of hemopoietic progenitor cells, which is characterized by abnormal proliferation, clonality and differentiation.1,2 In 2018, there is expected to be 19,520 new cases of acute myeloid leukemia (AML), and 10,670 people died of it.3 At present, the main treatment strategy for leukemia is chemotherapy. In the past 30 years, refinement of supportive treatment has improved the outlook of patients with AML. However, more than half of the young adults and about 90% of the older patients still die of AML.4 Therefore, it is urgent to find other therapeutic strategies for AML treatment.
Notopterygium incisum Ting ex H. T. Chang (N. incisum), known as Qianghuo, is commonly used to treat rheumatism and injury in traditional Chinese medicine. Modern pharmacological studies have confirmed that N. incisum has effects of antipyretic, analgesic, anti-inflammatory, antiviral, anticancer, etc.5,6 Notopterol, one type of coumarin, is an active monomer extracted from N. incisum with antipyretic, analgesic and anti-inflammatory effects, which also has been reported to induce cell-cycle-specific inhibition and apoptosis in MCF-7 cancer cell line.5,7–9 However, there is no report about its effect on leukemia.
Multiple mechanisms of apoptosis are involved in the treatment of cancer, among which the mitochondrial intrinsic apoptotic pathway plays the most pivotal role. Importantly, induced apoptosis by regulating Bcl-2 family members has a major role in the treatment of many drugs on AML.10,11 Recently, tumor differentiation therapy has become a hot spot as all-trans retinoic acid (ATRA) is used in the treatment of AML for inducing differentiation of cancer cells, which has advantages of high efficacy and low toxicity and side effects.12 Cell-cycle proteins are considered as effective targets in cancer therapy.13 Multiple drugs could inhibit HL-60 cells by regulating the expression of cell-cycle proteins and inducing cell-cycle arrest.14,15 In this study, we first characterized the inhibitory effect of notopterol on HL-60 cells which belong to human leukemia cell line that have been used for laboratory research on blood cell formation and physiology. On the one hand, cell viability, proliferation and colony formation assay were performed to determine whether notopterol induces death in HL-60 cells. On the other hand, we explored the mechanism of notopterol-inducing apoptosis by detection of mitochondrial intrinsic apoptotic pathway. Then, we characterized the effect of notopterol on differentiation and cell-cycle arrest of HL-60 cells. We examined the phenotype changes in differentiation, proteins associated with differentiation and cell cycle. In addition, combination therapy of notopterol and ATRA was used to test whether they had beneficial effect on HL-60 cells. Our findings may provide basis for application of notopterol and provide candidate drug for treatment of AML.
Materials and methods
Chemicals and antibodies
Notopterol with purity of 98% was purchased from ChromaBio (Chengdu, China), dissolved in dimethyl sulphoxide (DMSO) (Sigma-Aldrich, St. Louis, MO, USA) at 50 mM and stored at −20 °C before use. The antibodies against caspase 3 (1:1,000; cat. no. 96625), cleaved caspase 3 (1:1,000; cat. no. 96645) and Bax (1:1,000; cat. no.147965) were purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA). The antibodies against Cyclin E (1:1,000; cat. no. wl02253), Cyclin D1 (1:1,000; cat. no. wl01435a), CDK2 (1:1,000; cat. no. wl01543), CDK4 (1:1,000; cat. no. wl03343), c-Myc (1:1,000; cat. no. wl01781), PARP (1:1,000; cat. no. wl01932) and survivin (1:1,000; cat. no. wl01684) were purchased from Wanleibio Co., Ltd. (Shenyang, China). The antibodies against c-caspase 9 (1:1,000; cat. no.sc-133109), Bcl-2 (1:1,000; cat. no.sc-509), c-Jun (1:1,000; cat. no.sc-74543), Jun B (1:1,000; cat. no.sc-46), p53 (1:1,000; cat. no.sc-393031) and Mcl-1 (1:1,000; cat. no.sc-74437) were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). The antibody against GAPDH (1:1,000; cat. no.21612) was purchased from SAB (MD, USA). The antibody against β-actin (1:1,000; cat. no. TA-09) was purchased from Zhongshan Golden Bridge Biotechnology (Beijing, China). The antibodies against mouse or rabbit IgG were obtained from Sigma-Aldrich (St. Louis, MO, USA).
Cell culture
AML cell lines HL-60, Kasumi-1, U937 were obtained from the Cell Bank of Chinese Academic of Science (Shanghai, China) and grown in RPMI-1640 medium (Gibco Life Technologies, Carlsbad, CA, USA) supplemented with 10% (v/v) heat-inactivated FBS (Invitrogen, Waltham, MA, USA), 100 μg/mL penicillin (Invitrogen) and 100 μg/mL streptomycin (Invitrogen) in a 5% CO2 humidified incubator at 37°C.
Measurement of cell viability
The cell viability of notopterol was determined by sulforhodamine B (SRB) assay. HL-60 cells, Kasumi-1 and U937 cells (8×104 cells/mL) were inoculated into 96-well plate and administered vehicle (DMSO) or notopterol 1, 2.5, 5, 10, 25, 50 and100 μM, respectively. HL-60 cells were treated with notopterol or ATRA alone or combination of these two drugs as indicated concentration. Every concentration repeated 3 times. Then cells were treated for 72 hrs and subsequently added with 33.3 µL 4°C precooled Trichloroacetic acid(J&K, Beijing, China)solution (30%, w/v) to immobilize cells for 1 hr. Every well was rinsed 5 times with deionized water. 96-well plated was dried at room temperature, and added with 70 μL 0.4% (w/v) sulforhordamine B (SRB) (Sigma, St. Louis, MO, USA) for 30 minutes's dyeing, then washed with 1% (v/v) acetic acid for 4 times. 96-well plate was dried at room temperature, Tris-base alkaline (BBI Life Science, Shanghai, China) solution (10 mM, pH 10.5) was added and oscillated for 20 mins. The OD value was detected at 490 nm with a microplate reader (Biotek, Winooski, VT, USA).
Cell proliferation assay
The proliferation of HL-60 was determined by trypan blue dye exclusion test. Cells (5×104 cells/mL) were inoculated into 6-well plate and administered vehicle or notopterol 0, 10, 20, 40, 60 and 80 μM, respectively. Then cells were treated for 24–120 hrs. Trypan blue (Sigma) staining was performed every day and the number of viable cells using hemocytometer was counted.
Colony formation assay
HL-60 cells (2500 cells/well) were inoculated into 24-well plate in methylcellulose (Methocult H4100, StemCell Technologies Inc, Vancouver, British Columbia, Canada) supplemented with 10% FBS (Invitrogen) and 1% penicillin/streptomycin (Invitrogen) and administered vehicle or notopterol 5, 10, 20 and 40 μM respectively. Then colony-forming cells were evaluated microscopically 14 days after plating.
Cell apoptosis analyzed by annexin V-FITC/PI double-staining assay
Cells were seeded at a concentration of 2×105 cells/mL and incubated for 48 hrs with vehicle or notopterol at various concentrations. Cells were harvested and washed three times with pre-cold PBS. Then, cells were resuspended in the binding buffer containing annexin V-FITC and PI. After incubation for 15 mins at room temperature, BD LSRFortessa Cell Analyzer (BD Biosciences, San Jose, CA, USA) was used to perform flow cytometry analyses.
Hoechst 33342 staining
Cells (2×105 cells/mL) were treated with vehicle or notopterol or ATRA or combination of these two drugs for 48 hrs. Cells were centrifuged and collected in Eppendorf tubes. Then, cells were washed twice using PBS and fixed with 4% paraformaldehyde in 1× PBS for 10 mins and washed with 1× PBS for 3 times. Subsequently, the cells were stained with Hoechst 33342 reagent for 10 mins at room temperature. The cells were washed three times with 1× PBS. Finally, cells were observed and photographed under fluorescence microscope (Leica DM4000B, Wetzlar, Germany).
Analysis of the changes in mitochondrial membrane potential (Δψm)
Cells were seeded at a concentration of 3×105 cells/mL and incubated with vehicle or notopterol at various concentrations. After 24-hr treatment, the changes in mitochondrial membrane potential (Δψm) were tested with mitochondrial membrane potential assay kit with JC-1 following the manufacturer’s instruction (Beyotimebio,Shanghai, China). Briefly, collected cells were suspended in 0.5 mL culture medium. After adding 0.5 mL JC-1 staining solution, the cells were incubated in 37°C for 20 mins. The cells were centrifugated (600× g, 4°C) for 4 mins and abandoned the supernatant. Precipitating cells were washed twice with JC-1 staining buffer (1⨰). Then, cells were resuspended with JC-1 staining buffer (1⨰) and analyzed by fluorescence microscope (Leica DM4000B, Wetzlar, Germany).
Wright–Giemsa staining
Cells (2×105 cells/mL) were treated with vehicle or different concentrations of notopterol with or without ATRA (1 μM) for 48 hrs. Then, cells were harvested and the density was adjusted to 20×104cells/mL. Notopterol was given according to the corresponding initial concentration. After 48 hrs, the cells were collected and used Wright–Giemsa (Leagene, Beijing, China) staining according to the manufacturer’s protocol on slides prepared by cytospin to perform morphological assessment. The morphology of cells was observed under a light microscope.
Nitroblue tetrazolium (NBT) reduction assay
For the NBT reduction assay, cells (2×105 cells/mL) were treated with different concentrations of notopterol or ATRA (1 µM, as a positive control) or combination application for 48 hrs. Then, cells were harvested and the density was adjusted to 2×105 cells/mL. Notopterol was given according to the corresponding initial concentration. After 48 hrs, cells were incubated with 1 mg/mL NBT (J&K) solution at 37°C for 30 mins, then the proportion of cells containing the precipitated formazan particles in 300 cells were counted by optical microscope. 12-O-Tetradecanoylphorbol-13-acetate was used to stimulate the formation of formazan.
CD14 and CD11b detected by flow cytometry
For the CD14 or CD11b detection, cells (2×105 cells/mL) were exposed to different concentrations of notopterol or ATRA (1 µM, as a positive control) for 96 hrs. Cells were harvested and washed twice with precold 1×PBS. Then, cells were incubated with direct fluorescein isothiocyanate-labeled anti-CD11b antibody (BD Biosciences) or anti-CD14 antibody (BD Biosciences) in the dark. After 30 mins, cells were washed twice with precold PBS and detected by flow cytometry (BD Biosciences).
Cell-cycle phase distribution determined by flow cytometry
Cells (2×105 cells/mL) were treated with vehicle or different concentrations of notopterol for 48 hrs. Cells were harvested and washed twice with precold PBS and fixed in cold 75% ethanol overnight at 4°C. Next, cells were resuspended in 500 μL PI/RNase staining buffer (Beyotimebio) and detected by flow cytometry (BD Biosciences).
Western blotting analysis
For protein analysis of Bcl-2 family members, caspases, Jun B, c-Jun, protein p53, c-myc, CDK2, CDK4, CyclinD1, CyclinE, survivin and PARP, cells (2×105 cells/mL) were treated with vehicle or different concentrations of notopterol for 48 hrs. Harvested cells were washed with ice-cold PBS for 3 times. RIPA lysis buffer (Cell Signaling Technologies) containing protease inhibitor (Thermo Fisher Scientific, Waltham, MA, US) phenylmethanesulfonyl fluoride (Multi Sciences, Hangzhou, China) was used to extract total protein for 10 mins at 4°C. The supernatant was transferred to another new Eppendorf tube. The protein samples were then separated on 10–15% SDS-PAGE and transferred to polyvinylidene fluoride membrane. The membranes were blocked for 2 hrs in 5% BSA (Sangon Biotech, Shanghai, China) at room temperature, and then incubated with the primary antibodies overnight at 4°C. After incubation, the membranes were washed with Tris-buffered saline containing Tween-20 (TBST). The membranes were then incubated with goat anti-rabbit or goat anti-mouse secondary antibodies for 2 hrs at room temperature. After washing with TBST three times for 10 mins, the immunoreactive complexes were visualized by enhanced chemiluminescence (ECL) detection reagent (GE Healthcare, Uppsala, Sweden). The protein levels were first normalized to β-actin or GAPDH and then normalized to control.
Statistical analysis
Data were expressed as the Mean ± SD. The statistical signifiance of the differences was determined using Student’s t-test or one-way ANOVA in SPSS 17.0 software (SPSS, Inc., Chicago, IL, USA). A value of P<0.05 was considered to be significant. Prism 5.03 (GraphPad Software Inc., San Diego, CA, USA) was used to perform calculations. ImageJ 1.8.0 (National Institutes of Health, Bethesda, MD, USA), Flowjo7.6.2 (Tree Star, Inc., Ashland, OR, USA) and Modfit LT 3.2 (Verity Software House. Inc., Topsham, ME, USA) were used for image analysis.
Results
Notopterol inhibited cell growth of HL-60 cells
To determine the inhibitory effect of notopterol on HL-60, Kasumi-1 and U937 cell growth, we first used SRB assay to evaluate the cell viability. As shown in Figure 1A, notopterol showed greater inhibitory effect on HL-60 cells than Kasumi-1 and U937 with IC50 value of 40.32, 56.68 and 50.69 μM, respectively. Therefore, we chose HL-60 cells for further study. The influence of notopterol on the proliferation of HL-60 cells was determined. Figure 1B showed that 10–80 μM of notopterol inhibited the proliferation of HL-60 cells in a concentration-dependent manner within 120 hrs. HL-60 cells treated with 10, 20 and 40 μM notopterol for 120 hrs presented with significantly reduced cell numbers compared with controls. Notably, 60 and 80 μM of notopterol almost totally suppressed the proliferation of HL-60 cells (P<0.01). To evaluate the inhibitory effect on cell proliferation with a long-term treatment of notopterol, the methylcellulose-based colony formation assay was performed. It can be seen from Figure 1C that the size of colonies decreased in a concentration-dependent manner after HL-60 cells were treated by notopterol (5–40 µM) for 2 weeks. In addition, similar to cell viability results, cells that were treated with notopterol formed less colonies compared with the control group. Quantification results showed that there was 25%, 30%, 60% and 70% decrease in colonies formed with 5 (P<0.05), 10 (P<0.01), 20 (P<0.01) and 40 (P<0.01) µM of notopterol. In summary, the above mentioned results consistently suggest that notopterol exhibits a negative effect on cell survival, proliferation and growth.
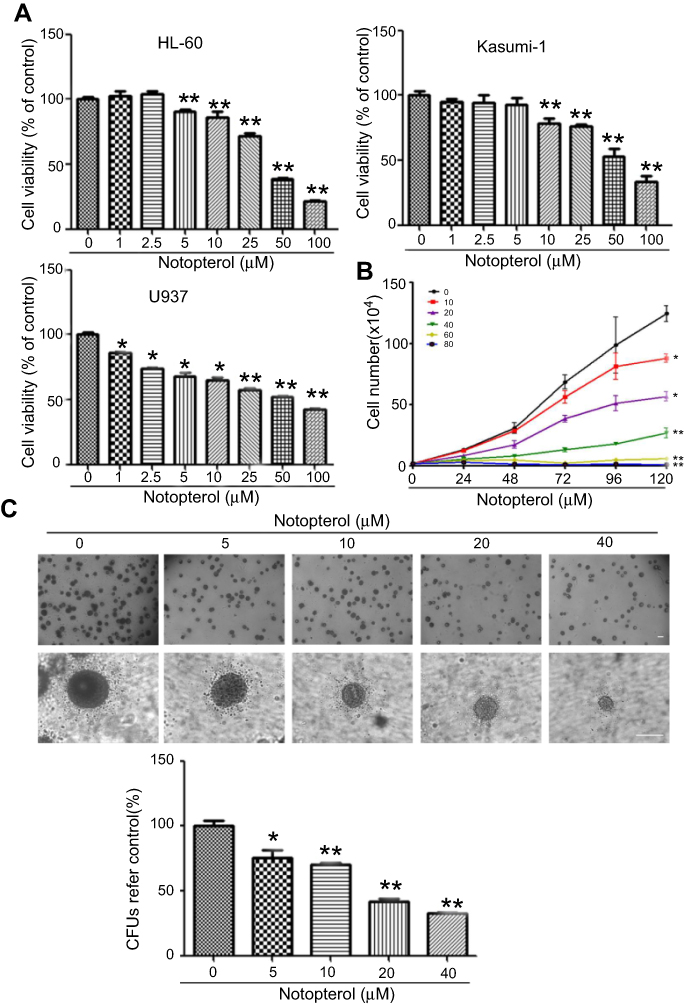 | Figure 1 Notopterol inhibited cell growth in HL-60 cells. (A) Cell viability. HL-60, U937 and Kasumi-1 cells were treated with notopterol as indicated for 72 hrs. The cell viability was assessed by sulforhodamine B (Mean ± SD, n=3). (B) Proliferation. Exponentially growing cells were treated with different concentrations of notopterol (0, 10, 20, 40, 60 and 80 μM) for 120 hrs. Cell proliferation was assessed using trypan blue exclusion assay (Mean± SD, n=3). (C) Colony formation. Cells were cloned in in methylcellulose and treated with different concentrations of notopterol (0, 5, 10, 20 and 40 μM) for 2 weeks and photographed by microscope. The scale bar is 100 μm. The colony images were a representative of three independent experiments. *P<0.05, **P<0.01 as compared to the control. CFUs represents Colony-Forming Units. |
Notopterol-induced apoptosis of HL-60 cells
To characterize the effect of notopterol on cell death of HL-60 cells, we first examined the apoptosis rate of cells treated with notopterol by annexin V-FITC and PI staining by flow cytometry analysis. As shown in Figure 2A, flow cytometry analysis showed that notopterol promoted cell apoptosis in a dose-dependent manner. After being treated with notopterol for 48 hrs, 5.79%±0.72%, 8.42%±0.92%, 12.67%±3.09%, 19.38%±4.09% and 37.97±3.17% apoptotic cells were found in HL-60 cells treated with 5 µM, 10 µM, 20 µM (P<0.01), 40 µM (P<0.01) and 60 µM (P<0.01), which were significantly higher than that of control cells (4.57%±1.16%). Then, we further characterized the apoptosis by Hoechst 33342 staining. It can be seen from Figure 2B that the proportion of cells with nuclear fragmentation and condensation was obviously increased after notopterol treatment. These data suggest that notopterol induces cell apoptosis.
 | Figure 2 Notopterol-induced apoptosis in HL-60 cells. (A) Effect of notopterol on apoptosis. Cells were treated with different concentrations of notopterol (0, 5, 10, 20, 40 and 60 μM) for 48 hrs. Apoptotic cells were quantified by flow cytometry after staining with fluorescein isothiocyanate (FITC)-conjugated Annexin V and propidium iodide(PI) (Mean± SD, n=3). (B) Hoechst 33342 staining assay. Cells were treated with different concentrations of notopterol (0, 5, 10, 20, 40 and 60 μM) for 48 hrs and stained by Hoechst 33342, and then were observed under microscope (Mean± SD, n=3). Apoptotic cells were quantified. The scale bar is 100 μm. The images were a representative of three independent experiments. **P<0.01 as compared to the control. |
Notopterol-induced apoptosis is mediated by activation of the mitochondrial intrinsic apoptotic pathway
To further explore the underlying mechanism of apoptosis induced by notopterol, we first examined whether notopterol induces apoptosis through the mitochondrial intrinsic apoptotic pathway by immunofluorescence staining. Mitochondrial membrane potential was examined by staining with JC-1, a lipophilic, cationic dye that exhibits a fluorescence emission shift upon aggregation from 530 nm (green) to 590 nm (red). In normal cells with a high mitochondrial membrane potential, JC-1 enters the mitochondrial matrix in a potential-dependent manner and forms aggregates. However, in apoptotic cells, mitochondrial transmembrane potential depolarizes, then JC- 1 was released from the mitochondria. Consequently, it reverses to a monomeric form that emits green fluorescence. As shown in Figure 3A, there was a significant increase in green fluorescence and decrease in red fluorescence in the cells exposed to notopterol (5–60 µM) when compared with control, indicating collapse of mitochondrial membrane. Then, we examined the expression levels of proteins associated with mitochondrial intrinsic apoptotic pathway, including Bcl-2, Bax, caspase-3 and −9, PARP and Mcl-1. As shown in Figure 3B, notopterol increased the expression of Bax and decreased the expression of Bcl-2 and Mcl-1 in HL-60 cells after 48-hr treatment. Moreover, notopterol also promoted the cleavage of caspase 9, caspase 3 and PARP. Taking these results into account, we hypothesize that mitochondria plays a role in notopterol-induced apoptosis.
 | Figure 3 Notopterol-induced apoptosis is mediated by activation of the mitochondrial intrinsic apoptotic pathway. (A) Effect of notopterol on mitochondrial memberane potential (Mean ± SD, n=3). Cells were treated with different concentrations of notopterol (0, 5, 10, 20, 40 and 60 μM) for 24 hrs and then incubated with the membrane potential indicator, JC-1. Intracellular red and green fluorescence of JC-1 was determined by an inverted fluorescent microscope. The intracellular red and green fluorescence of JC-1 was quantified using Image J software. (B) Western blot detection of apoptosis-associated protein expression. Cells were treated with different concentrations of notopterol (0, 5, 10, 20 and 40 μM) for 48 hrs and then harvested. β-actin served as an internal reference. The scale bar is 100 μm. Expression fold vs control was shown under the corresponding band. The images were a representative of three independent experiments. *P<0.05 as compared to the control. |
Notopterol-induced differentiation of HL-60 cells
To examine the effect of notopterol on the differentiation of HL-60 cells, we first detected the morphologic changes of cells treated with notopterol by Wright–Giemsa staining. As shown in Figure 4A, notopterol (10, 20 and 40 µM) clearly induced the chromatin condensation and increased the nucleocytoplasmic ratio in HL-60 cells. Then, the NBT assay was performed to detect the production of oxidative bursts during the cell differentiation (Figure 4B). NBT-positive cells were concentration-dependently increased after treated by notopterol (5 to 40 µM) (Figure 4B). To further characterize the differentiation mediated by notopterol, the levels of CD11b and CD14, markers of myeloid differentiation, were measured by flow cytometry. An increase in the presence of CD14-positive cells could be detected in the notopterol-treated (10, 20 and 40 µM) HL-60 cells for 96 hrs, evident by the observed increase in the mean fluorescent intensity when compared to untreated control (Figure 5A). The increase in the expression of CD11b also was observed in HL-60 cells treated with notopterol at 40 µM. Moreover, in protein level, notopterol (40 µM) time-dependently increased the expression of c-Jun from 6 hrs to 48 hrs (Figure 5B). The expression of Jun B was also observed with an increase at 12 hrs after notopterol (40 µM) treatment (Figure 5B). In addition, notopterol (5 to 40 µM) dose-dependently inhibited the expression of c-myc at 48 hrs (Figure 5C). The above results suggest notopterol exerts inhibitory effect partly via induction of cell differentiation.
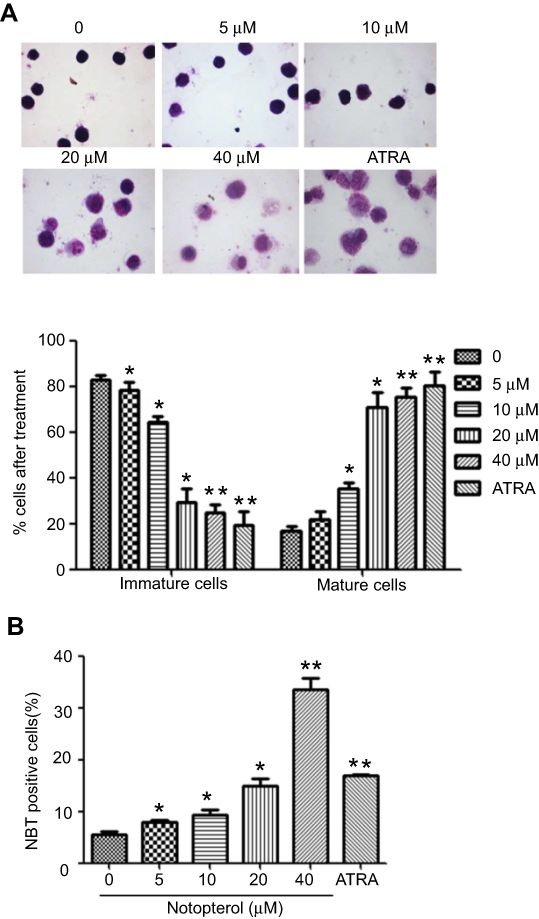 | Figure 4 Notopterol altered the morphology of HL-60 cells. (A) Giemsa staining assay (Mean ± SD, n=3). Cells were treated with different concentrations of notopterol (0, 5, 10, 20 and 40 μM) for 96 hrs and subjected to Giemsa staining. The magnification of the images shown in the panels is ×100. The percentage of immature and mature cells was quantified. (B) Nitroblue tetrazolium (NBT) reduction assay (Mean ± SD, n=3). Cells were treated with different concentrations of notopterol (0, 5, 10, 20 and 40 μM) for 96 hrs, NBT reduction experiment was performed to count the positive cell rate. All-trans retinoic acid (ARTA) is used as positive drug. The images display a representative experiment from three independent experiments. *P<0.05, **P<0.01 as compared to the control. |
 | Figure 5 Notopterol-induced cell differentiation in HL-60 cells. (A) Effect of notopterol on the expression of CD14 and CD11b (Mean ± SD n=3). Cells were given different concentrations of notopterol (0, 5, 10, 20 and 40 μM) for 96 hrs, and the expression levels of CD14 and CD11b were detected by flow cytometry. The mean fluorescence intensity was shown within the picture. Quantitative measurement of CD11b and CD14 signals by the flow cytometry was analyzed. The images display a representative experiment from two independent experiments. (B) Western blot detection of c-jun and Jun B protein expression. Cells were treated with 40 μM notopterol for different time. (C) Western blot detection of c-myc protein expression. Cells were treated with different concentrations of notopterol (0, 5, 10, 20 and 40 μM) for 48 hrs, GAPDH served as an internal reference. Expression fold vs control was shown under the corresponding band. *P<0.05 as compared to the control. FITC refers to fluorescein isothiocyanate. |
Notopterol induces G0/G1 phase arrest associated with cell-cycle proteins in HL-60 cells
To further explore whether the inhibitory effect of notopterol on HL-60 cells is due to its induction of cell-cycle arrest, high-resolution flow cytometry analysis of PI-stained nuclei was performed. As shown in Figure 6A, when compared with control group, the exposure of HL-60 cells to notopterol remarkably resulted in the accumulation of cells in G1 phase from 25.67±3.77 to 31.76±1.24, 35.44±4.09, 44.50±6.91 and 51.99±3.64 at 0, 5, 10 (P<0.05), 20 (P<0.05), 40 (P<0.01) µM, respectively. Furthermore, the protein expression levels of the molecules involved in the G0/G1-phase arrest were analyzed by western blot. As shown in Figure 6B, the expression levels of CDK2, CDK4, CyclinD1, CyclinE and survivin were decreased in a concentration-dependent manner with elevated p53 expression. These results demonstrate that notopterol inhibits the growth of HL-60 cells probably via arresting inducing G0/G1 phase arrest.
 | Figure 6 Notopterol-induced cell cycle arrest in HL-60 cells. (A) Effect of notopterol on cell cycle (Mean± SD, n=3). Cells were treated with different concentrations of notopterol (0, 5, 10, 20 and 40 μM) for 48 hrs. The images display a representative experiment from three independent experiments. Cells were quantitatively measured by flow cytometry. *P<0.05, **P<0.01 as compared to the control. (B) Western blot detection of cell-cycle-associated protein expression. Cells were treated with different concentrations of notopterol (0, 5, 10, 20 and 40 μM) for 48 hrs. β-actin served as an internal reference. Expression fold vs control was shown under the corresponding band. PI refers to propidium iodide. |
The effect of combination therapy notopterol and ATRA on HL-60 cells
First, we examined whether the combination use of notopterol and ATRA had beneficial effect on cell death. As shown in Figure 7A, notopterol can dose-dependently inhibited the cell viability of HL-60 (P<0.01). 1 and 2 μM of ATRA-treated cells were inhibited (P<0.01). However, the combined use of notopterol and ATRA did not enhance this inhibition. In Figure 7B, 20 or 40 μM of notopterol could increase the apoptotic rate, while 2 μM of ATRA did not induce apoptotis of HL-60. Likewise, the combination of notopterol and ATRA did not enhance the apoptotic effect. Altogether, we can draw a preliminary conclusion that the combination of notopterol and ATRA(1, 2 μM) has no synergistic effect on apoptosis and viability of HL-60 cells. Then, we tested its effect on differentiation. In Figure 8A, notopterol (20, 40 μM), ATRA (1 μM) could induce the chromatin condensation and increased the nucleocytoplasmic ratio separately. The combination of these two drugs (40 μM notopterol +1 μM ATRA, P<0.01) could obviously enhance the effect when compared with using either ATRA or notopterol alone. Moreover, in Figure 8B, the combination use (40 μM notopterol + 1 μM ATRA, P<0.01) also increased the NBT-positive cells, as compared with ATRA alone. In addition, according to previous study, we detected the expression of cell surface maker CD14 (Figure 8C). It showed the combination application (40 μM notopterol +1 μM ATRA, P< 0.01) enhanced the expression level of CD14, as compared with notopterol or ATRA alone. Therefore, the above results can indicate the combination of notopterol and ATRA has synergistic effect on differentiation of HL-60 cells.
 | Figure 7 The combination therapy of notopterol and all-trans retinoic acid (ATRA) on cell viability and apoptosis of HL-60 cells. (A) Cell viability. HL-60 cells were treated with notopterol (10, 20 and 40 μM) alone, ATRA (1, 2 μM) alone and the combination for 72 hrs. The cell viability was assessed by sulforhodamine B (n=3). (B) Hoechst 33342 staining assay. Cells were treated with different concentrations of notopterol (20, 40 μM) alone, ATRA (2 μM) alone and combination of notopterol and ATRA for 48 hrs (Mean ± SD, n=3). The scale bar is 100 μm. **P<0.01 as compared to the control. |
 | Figure 8 The combination therapy of notopterol and all-trans retinoic acid (ATRA) on cell differentiation of HL-60 cells. (A)Giemsa staining assay (Mean ± SD, n=3). Cells were treated with different concentrations of notopterol (20 and 40 μM) with or without ATRA (1 μM) for 96 hrs and subjected to Giemsa staining. The magnification of the images shown in the panels is ×100. The percentage of immature and mature cells was quantified. (B) Nitroblue tetreazolium (NBT) reduction assay (Mean± SD, n=3). Cells were treated with different concentrations of notopterol (20, 40 μM) with or without ATRA (1 μM) for 96 hrs, NBT reduction experiment was performed to count the positive cell rate. The images display a representative experiment from three independent experiments. (C) The expression of CD14. Cells were given different concentrations of notopterol (20, 40 μM) with or without ATRA (2 μM) for 96 hrs, the expression levels of CD14 were detected by flow cytometry. The mean fluorescence intensity was shown within the picture. *P<0.05, **P<0.01 as compared to the control. #P<0.05, ##P<0.01 as compared to cells treated with either ATRA or notopterol alone. FITC refers to fluorescein isothiocyanate. |
Discussion
At present, although chemotherapy is the most commonly used treatment, it also kills normal cells, causing many side effects. Therefore, it is urgent to develop novel alternative therapeutic strategies to overcome these problems. Studies showed notopterol, a main component existing in N. incisum, had antipyretic, analgesic and anti-inflammatory effects; however, there no reports about its effect on AML, let alone its application in clinics. In our preliminary investigation, through screening the active ingredients of traditional Chinese medicine, it was found that notopterol remarkably induced cell death of HL-60 cells U937, and Kasumi-1 cells. Interestingly, the inhibitory effect of notopterol on HL-60 cells was stronger than that on U937 and Kasumi-1 cells. Thus, we characterized the effect and molecular mechanisms of notopterol in this study to explore its potential therapeutic benefits for HL-60 cells. The results revealed that notopterol significantly inhibited cell growth as evidenced by cell viability, proliferation and colony formation tests. Many drugs have been reported to induce cancer cell apoptosis. Disruption of mitochondrial membrane potential is closely related to early stage of apoptosis. Bcl-2 protein family, including proapoptotic proteins Bad, Bid, Bax and antiapoptotic proteins Bcl-2, Bcl-x and Mcl-1, regulates the intrinsic mitochondrial pathway in apoptosis by mediating the release of Cyt-c from mitochondria to cytosol to activate downstream procaspase-9 and leading to the activation of caspase 3 and PARP.16–18 The analysis of flow cytometry combined with Hoechst 33342 staining confirmed notopterol-induced apoptosis in HL-60 cells. Then, results revealed that notopterol reduced mitochondrial membrane potential in concentration-dependent manner; increased the expression of Bcl-2, cleaved caspase 9, cleaved caspase 3 and cleaved PARP and decreased the expression of Mcl-1 and Bax. These results revealed that notopterol induced HL-60 cell apoptosis partly through the mitochondrial intrinsic pathway. Inducing cell differentiation is an effective therapy for AML,19 and C-myc is highly expressed in undifferentiated conditions and abolished when differentiation was induced by chemical inducers in HL-60 cells.20–22 Transcription factors c-Jun and Jun B that play important roles in myeloid differentiation can inhibit leukemic self-renewal capacity and initiate the myelomonocytic differentiation.10,23 Our results found that notopterol-treated HL-60 cells exhibited typical morphological changes, increased the percentage of NBT-positive cells and expression of myeloid differentiation surface marker CD14 and CD11b and protein expression of c-Jun, Jun B and c-myc, which confirmed the induction of differentiation. Arresting cell cycle is an important target for treating tumor. With an agreement of our assumption, results revealed the G0/G1 phase arrest in HL-60 cells following notopterol treatment. Different CDKs and cyclins are expressed in different cell-cycle period by forming CDK kinase complex and regulating the cell cycle by corresponding function.24,25 For G1 phase to S phase transition, all cells must activate CDK4/6, which combines with CyclinD1. CDK4/6 neutralizes p27Kip1 and induces E2F-dependent transcription of cyclins, resulting in the activation of CDK2, consequently let cells enter into S phase. CDK2 combines with Cyclin (E1, E2) and Cyclin A, which can be suppressed by p53.26,27 The expressions of CDK2, CDK4, CyclinD1 and CyclinE were decreased with the increasing expression of p53 following notopterol treatment. Overexpression of survivin leads to an increase in S phase, which protects cells from G1 phase blockade, eventually leading to the activation of CDK2/Cyclin E complex.28 Our results found that the expression of survivin was also repressed by notopterol. ATRA together with chemotherapy used as the first-line treatment for the newly diagnosed acute promyelocytic leukemia (APL) patients. However, APL patients treated with ATRA alone easily relapse. Nowadays, efforts have focused on the search of agents that can enhance ATRA’s effect on AML cells. Our results suggested that combination of notopterol and ATRA could not increase cell death. Interestingly, inducing differentiation effect was obviously enhanced when cells were treated with both notopterol and ATRA as evidenced by increasing nucleocytoplasmic ratio, NBT-positive cells and expression level of CD14.
Conclusion
In summary, we found notopterol effectively inhibited the proliferation, promoted the apoptosis and induced the differentiation and G0/G1 arrest in HL-60 cells. To the best of our knowledge, this is the first study to reveal that notopterol has the inhibitory effect on HL-60 cells, which could provide a basis to further evaluate the potential applications of notopterol as a promising drug for the treatment of AML. Moreover, combination therapy of notopterol and ATRA is worth for further research.
Consent for publication
All authors have provided consent for publication.
Acknowlegments
This work was supported by the Chongqing Research Program of Basic Research and Frontier Technology (cstc2017jcyjAX0299 and cstc2018jcyjA1295), the TCM Scientific Program Grant (ZY201702120) from Chongqing Health and Family Planning Commission, and the Key Project at Central Government Level: The Ability Establishment of Sustainable Use for Valuable Chinese Medicine Resources (2060302).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Estey E, Döhner H. Acute myeloid leukaemia. Lancet. 2006;368(9550):1894–1907. doi:10.1016/S0140-6736(06)69780-8
2. Döhner H, WeisdorfLongo DL, Weisdorf DJ, et al. Acute myeloid leukemia. N Engl J Med. 2015;373(12):1136–1152. doi:10.1056/NEJMra1406184
3. Siegel RL, Miller KD, Jemal A. Cancer statistics. CA Cancer J Clin. 2018;68(1):7–30. doi:10.3322/caac.21442
4. Ferrara F, Schiffer C. Acute myeloid leukaemia in adults. Lancet. 2013;381(9865):484–495. doi:10.1016/S0140-6736(12)61727-9
5. Li Y, Zheng X, Meng X. Determination of notopterol concentrations and pharmacokinetic in rat plasma by high-performance liquid chromatography. J Nanchang Univ: Med Ed. 2012;52(7):8–11.
6. Azietaku J, Ma H, Yu X, et al. A review of the ethnopharmacology, phytochemistry and pharmacology of notopterygium incisum. J Ethnopharmacol. 2017;202:241–255. doi:10.1016/j.jep.2017.03.022
7. Wu S, Pang F, Wen Y, Zhang H-F, Zhao Z, Hu J-F. Antiproliferative and apoptotic activities of linear furocoumarins from notopterygium incisum on cancer cell lines. Planta Med. 2010;76(1):82–85. doi:10.1055/s-0029-1185971
8. Zhang P, Yang X. [Studies on chemical constituents in roots and rhizomes of notopterygium incisum]. Zhongguo Zhong Yao Za Zhi. 2008;33(24):2918–2921.
9. Okuyama E, Nishimura S, Ohmori S, Ozaki Y, Satake M, Yamazaki M. Analgesic component of Notopterygium incisum Ting. Chem Pharm Bull (Tokyo). 1993;41(5):926–929.
10. Wang C, He H, Dou G, et al. Ginsenoside 20(S)-Rh2 induces apoptosis and differentiation of acute myeloid leukemia cells: role of orphan nuclear receptor Nur77. J Agric Food Chem. 2017;65(35):7687–7697. doi:10.1021/acs.jafc.7b02299
11. Vakamullu S, Arepalli S, Velatooru L, et al. In vitro apoptotic mechanism of a novel synthetic Quinazolinyl derivative: induces caspase-dependent intrinsic pathway on THP-1, leukemia cell line. Chem Biol Interact. 2018;280:117–127. doi:10.1016/j.cbi.2017.12.015
12. Testa U, Lo-Coco F. Targeting of leukemia-initiating cells in acute promyelocytic leukemia. Stem Cell Invest. 2015;2:8.
13. Otto T, Sicinski P. Cell cycle proteins as promising targets in cancer therapy. Nat Rev Cancer. 2017;17(2):93–115. doi:10.1038/nrc.2016.138
14. Omer F, Hashim N, Ibrahim M, et al. Beta-mangostin from Cratoxylum arborescens activates the intrinsic apoptosis pathway through reactive oxygen species with downregulation of the HSP70 gene in the HL60 cells associated with a G/G cell-cycle arrest. Tumour Biol. 2017;39(11):1010428317731451. doi:10.1177/1010428317731451
15. Husain I, Bala K, Wani A, Makhdoomi U, Malik F, Sharma A. Arginase purified from endophytic Pseudomonas aeruginosa IH2: induce apoptosis through both cell cycle arrest and MMP loss in human leukemic HL-60 cells. Chem Biol Interact. 2017;274:35–49. doi:10.1016/j.cbi.2017.07.001
16. Deng J. How to unleash mitochondrial apoptotic blockades to kill cancers? Acta Pharm Sin B. 2017;7(1):18–26. doi:10.1016/j.apsb.2016.08.005
17. Bhola P, Letai A. Mitochondria-judges and executioners of cell death sentences. Mol Cell. 2016;61(5):695–704. doi:10.1016/j.molcel.2016.02.019
18. Ulukaya E, Acilan C, Yilmaz Y. Apoptosis: why and how does it occur in biology? Cell Biochem Funct. 2011;29(6):468–480. doi:10.1002/cbf.1774
19. Wang R, Xia L, Gabrilove J, Waxman S, Jing Y. Sorafenib inhibition of Mcl-1 accelerates ATRA-induced apoptosis in differentiation-responsive AML cells. Clinl Cancer Res Off J Am Asso Cancer Res. 2016;22(5):1211–1221. doi:10.1158/1078-0432.CCR-15-0663
20. Grosso L, Pitot H. Transcriptional regulation of c-myc during chemically induced differentiation of HL-60 cultures. Cancer Res. 1985;45(2):847–850.
21. Filmus J, Buick RN. Relationship of c-myc expression to differentiation and proliferation of HL-60 cells. Cancer Res. 1985;45(2):822.
22. Holt J, Redner R, Nienhuis A. An oligomer complementary to c-myc mRNA inhibits proliferation of HL-60 promyelocytic cells and induces differentiation. Mol Cell Biol. 1988;8(2):963–973.
23. Ghaffarizadeh A, Flann N, Podgorski G. Multistable switches and their role in cellular differentiation networks. BMC Bioinformatics. 2014;15(Suppl 7):S7. doi:10.1186/1471-2105-15-S7-S7
24. Massagué J. G1 cell-cycle control and cancer. Nature. 2004;432(7015):298. doi:10.1038/nature03094
25. Feng Y, Xu X, Zhang Y, et al. HPIP is upregulated in colorectal cancer and regulates colorectal cancer cell proliferation, apoptosis and invasion. Sci Rep. 2015;5:9429. doi:10.1038/srep09429
26. Massagué J. G1 cell-cycle control and cancer. Nature. 2004;432(7015):298–306. doi:10.1038/nature03094
27. Zhang Y, Shen Q, Li J. Traditional Chinese medicine targeting apoptotic mechanisms for esophageal cancer therapy. Acta Pharmacol Sin. 2016;37(3):295–302. doi:10.1038/aps.2015.116
28. Suzuki A, Hayashida M, Ito TH, et al. Survivin initiates cell cycle entry by the competitive interaction with Cdk4/p16(INK4a) and Cdk2/cyclin E complex activation. Oncogene. 2000;19(29):3225–3234. doi:10.1038/sj.onc.1203665
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.