Back to Journals » International Journal of Nanomedicine » Volume 20
Nitric Oxide Therapeutics: New Hopes for More Effective Tuberculosis Treatment Combine with Targeted and Controlled Nanotechnology
Authors Jin X, Wang J ![]() , Ruan Y, Li J, Kong X, Xia J, Yang J, Zhang Q, Liu J, Pi J
, Ruan Y, Li J, Kong X, Xia J, Yang J, Zhang Q, Liu J, Pi J ![]()
Received 31 March 2025
Accepted for publication 9 July 2025
Published 19 July 2025 Volume 2025:20 Pages 9195—9218
DOI https://doi.org/10.2147/IJN.S531255
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. RDK Misra
Xiaoying Jin,1,2,* Jiajun Wang,1,2,* Yongdui Ruan,1,* Jiaxiang Li,1,2 Xinen Kong,1,2 Jiaojiao Xia,3 Jiayi Yang,1,2 Qiao Zhang,3 Juan Liu,1,2 Jiang Pi1,2
1Research Center of Nanotechnology and Application Engineering, School of Medical Technology, The First Dongguan Affiliated Hospital, Guangdong Medical University, Dongguan, People’s Republic of China; 2Dongguan Key Laboratory for Pathogenesis and Experimental Diagnosis of Infectious Diseases, The First Dongguan Affiliated Hospital, School of Medical Technology, Guangdong Medical University, Dongguan, People’s Republic of China; 3Department of Biochemistry and Molecular Biology, School of Basic Medical Sciences, Kunming Medical University, Kunming, Yunnan, 650500, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jiang Pi, Email [email protected] Qiao Zhang, Email [email protected]
Abstract: Tuberculosis (TB) caused by Mycobacterium tuberculosis (Mtb) is one of the most prevalent infectious diseases worldwide. Nitric oxide (NO) is produced by the reaction of arginine and oxygen catalyzed by nitric oxide synthase (NOS) in mammals. Several studies have highlighted the potential therapeutic use of NO for the treatment of various diseases, including infectious diseases. NO plays a direct bactericidal role by damaging the bacterial DNA, proteins, and enzymes. Additionally, it plays a role in modulating immune cell function, contributing to their anti-tuberculosis (anti-TB) effects by regulating macrophage activity. NO has also been shown to eliminate bacterial biofilms, thereby increasing drug sensitivity of drug-resistant bacteria. Therefore, combining NO with antibiotics may be a strategy for treating drug-resistant tuberculosis (DR-TB). However, owing to the limitations of NO, including their short half-life, instability, and cytotoxicity, exogenous supplementation with NO donors has emerged as a promising alternative therapy. Rapid advancements in nanotechnology have led to the development of nanoparticles (NPs) as drug delivery platforms, at the same time, using strategies such as introducing selective organ targeting (SORT) molecules into nanocarrier systems or preparing nanodrugs in inhalable or dry powder inhalation forms can increase the accumulation of nanodrugs in the lungs. Combined with host-directed therapy strategies, this can improve the therapeutic effect on tuberculosis and shorten the treatment time. This review summarizes the biological activities of NO and introduces their applications in the treatment of several major infectious diseases, followed by a systemic analysis of the role and mechanism of action of NO in TB treatment. Moreover, nanotechnology-assisted NO therapeutics are also summarized to explore the potential for more effective Mtb killing based on the advantages of targeted NO release at the infected site and host cells, thus benefiting the development of more effective therapeutics against TB and drug-resistant TB.
Keywords: nitric oxide, nitric oxide donors, nanotechnology, tuberculosis
Introduction
Since the outbreak of the coronavirus disease (COVID-19) pandemic in 2019, increasing attention has been paid to infectious diseases. Tuberculosis (TB) is a deadly disease caused by Mycobacterium tuberculosis (Mtb), and is one of the most threatening infectious diseases. According to the “Global Tuberculosis Report 2024” released by the World Health Organization, the total number of TB patients worldwide in 2023 will reach 10.8 million, making TB the world’s top infectious disease. Traditional treatment methods primarily rely on the use of antibiotics to achieve bactericidal effects. Although antibiotics are effective at killing bacteria, their efficacy against intracellular Mtb is limited when we used them alone, which leads to prolonged treatment durations, severe side effects in clinical settings and the emergence of drug-resistant mutant strains, which poses a significant challenge. While nanotechnology-based nanodrugs demonstrate enhanced therapeutic effects, improved targeting capabilities, and good biosafety and stability. However, the therapeutic effect of standalone drug nano-delivery systems remains insufficient to meet the demand for “eliminating” tuberculosis. Therefore, the search for new anti-TB (anti-tuberculosis) drugs is urgently required.
Nitric oxide (NO) demonstrates clear clinical therapeutic value in cardiovascular diseases (CVDs), tumors, and cancers as a popular gas molecule in current research. Furthermore, studies have indicated that NO can exert direct bactericidal effects or enhance bacterial sensitivity to antibiotics by dispersing or eliminating biofilms, depending on their concentration. The physiological effects of NO are concentration dependent and have a short half-life and instability. Moreover, their production is associated with the concentration and location of the nitric oxide synthase (NOS). Macrophages are the target cells of Mtb, and they can produce inducible nitric oxide synthase (iNOS), which catalyzes the decomposition of L-Arginine (L-Arg) to generate NO. This is particularly true in infectious and inflammatory diseases. In addition, as early as 1997, it was demonstrated that the uninterrupted generation of NO endows macrophages with cytostatic or cytotoxic capabilities against viruses, bacteria, fungi, and tumor cells. These effects can be augmented by other macrophage-derived substances such as acids, hydrogen peroxide, or superoxide.1 Considering that the concentration of endogenous NO is relatively low, and their concentration will be further reduced in many disease states, exogenous supplementation of NO has become a new disease treatment strategy. However, direct NO delivery has many drawbacks including systemic cytotoxicity and low target concentrations. This prompted the search for a suitable NO donor and a class of compounds capable of releasing NO in vivo. NO donors are capable of not only storing and releasing NO, but also improving the pharmacokinetics of NO.2 However, research on the application of NO donor materials is still immature and presents many drawbacks, such as premature leakage of NO and toxic side effects from NO donor metabolites. With the development and maturity of nanotechnology, nano-drug delivery systems have been used for the treatment of diseases. They can encapsulate drugs in nanocarriers (such as liposomes and polymer nanoparticles) to achieve the targeted delivery of drugs, ensure the stability and controlled release of drugs, reduce side effects, and ultimately improve the treatment effect of diseases. This dual-function approach is highly advantageous in the process of resolving the defects of NO donors and achieving a therapeutic effect in diseases.
TB treatment can be performed it in two aspects. On the one hand, it plays a role by regulating the function of macrophages. It has been shown that supplementation of the NO donor SNAP could eliminate Mtb through NO-dependent apoptosis under the regulation of the nicotinamide adenine dinucleotide (NAD+) -dependent deacetylase, sirtuin 7 (SIRT7), in macrophages.3 In addition, NO can also play a role in host targeted treatment of TB by inhibiting ferroptosis of macrophages.4 On the other hand, it directly kills Mtb. This is because NO can disrupt Mtb nucleic acids, lipids and proteins, as well as displace metal cofactors from essential metabolic enzymes.5 From a mechanistic point of view, it has been shown that NO can inhibit the respiration of Mtb and prevent their growth. This is because Mtb SufR contains NO-sensitive 4Fe-4S, and NO treatment leads to active degradation of several iron-sulfur (Fe-S) cluster proteins, thus regulating Mtb metabolism, respiration, and redox balance.6 NO has these two functions at the same time, which undoubtedly show their important potential in anti-TB treatment.
This review provides a comprehensive overview of the biological and regulatory effects of NO in diseases as well as the potential applications of nanotechnology-based NO therapy infectious diseases intervention and treatment. By integrating the primary mechanisms of NO therapy into macrophages, we propose the NO-controlled release of nanomaterials and discuss their applications in the treatment of TB. Finally, we extrapolate that NO therapy holds promise for TB treatment. Additionally, by exploring the design of NO-controlled release nanomaterials, discussing their advantages and disadvantages, and assessing their innovativeness and feasibility, we aim to provide a theoretical foundation for future research.
The Physiological Roles of NO and Their Regulatory Function in Diseases
The Physiological Roles of NO
In the past decade, researchers have developed great interest in antibacterial gas therapy. NO, carbon monoxide (CO), sulfur dioxide (SO2), and hydrogen (H2) are not only known as endogenous signaling molecules, but also play key roles in many pathological processes.7 Among these gases, NO is a hot topic of research at present. Back in the 1980s, Robert Furchgott found that certain substances released by endothelial cells of blood vessels can stimulate endothelial cells and cause smooth muscle relaxation.8 In 1992, NO was called “Molecule of the Year”. Since then, NO has become a universal signal that regulates physiological functions within organisms.9 NO has a short half-life (3–4 s or <5s) and is volatile, and exhibits diverse physiological effects, but these effects are concentration-dependent.10 Furthermore, their diffusion range is 40–200 µm, typically confining their actions to target sites.11 The production, half-life, diffusion distance, and concentration dependence of NO have different physiological roles in the cardiovascular, nervous, and immune systems of the body.
Endogenous NO serves as an important effector and signaling molecule that participates in a variety of physiological processes, including vasodilation, immune response, neurotransmission, apoptosis, and reproduction of bacteria.12 In the cardiovascular system, NO induces vasodilation by activating guanylate cyclase and generating cyclic guanosine monophosphate. In addition, iNOS-mediated massive production of NO is considered to have a protective effect against atherosclerosis by inhibiting platelet aggregation, vascular smooth muscle cell (VSMC) proliferation, VSMC migration, and promotion of endothelial cell survival.13,14 In the nervous system, NO is released as a neurotransmitter by both presynaptic and postsynaptic nerve terminals. It can enhance the signal transmission between neurons, thereby contributing to memory consolidation.15 However, there is evidence indicating that a higher concentration of NO act as a toxic agent can affect neuronal death through multiple mechanisms.16 In the immune system, it has been shown that NO can regulate the function of many immune cells, including T lymphocytes and antigen-presenting cells, rendering them more vigorous in combating infection.17,18 In the reproductive system, NO can affect multiple functions of the reproductive system, such as affecting follicle development and sperm motility.19,20 In terms of their antibacterial properties, NO can enhance the phagocytic function of immune cells to clear bacteria, and it can also regulate the production of cytokines and affect the intensity and type of immune response. In addition, NO acts synergistically with the antimicrobial peptides to enhance the antimicrobial activity of the peptides.21 Finally, NO can kill bacterial cells by directly damaging bacterial DNA and inhibiting its repair, while not affecting normal tissues.22 Moreover, NO can react with reactive oxygen species (ROS), such as O2 and H2O2, to form reactive nitrogen species (RNS), which exhibit stronger bactericidal ability than ROS and can exacerbate lipid peroxidation and damaging bacterial membranes.22,23 In terms of anti-tumor cells, NO can kill tumor cells by inducing the down-regulation of reductase activity, thereby reducing glutathione (GSH) level, inducing mitochondrial dysfunction and activating caspase-3.24 All these reflect the complexity of the physiological function of NO and their great potential as a small molecule for disease treatment (Figure 1).
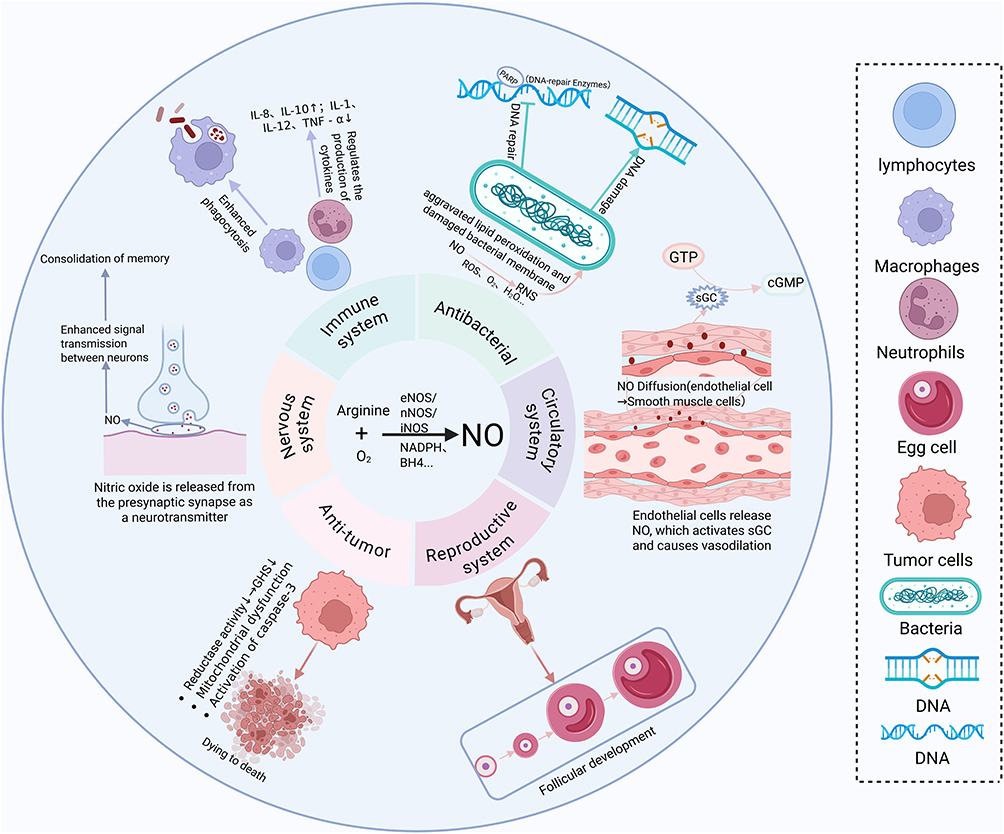 |
Figure 1 Diversity of NO biological activities. Created in BioRender. Liu, J. (2025) https://BioRender.com/rmzey7k. Abbreviations: GTP, Guanosine Triphosphate; cGMP, Cyclic Guanosine Monophosphate; sGC, Soluble Guanylate Cyclase.S. |
NO Plays an Important Role in Several Major Infectious Diseases
Three types of NOS exist in mammals, which play crucial roles in the synthesis of NO from arginine and oxygen, together with several cofactors, including nicotinamide adenine dinucleotide phosphate (NADPH) and tetrahydrobiopterin (BH4).25,26 NO can play various roles in many systems, which are related to the type and location of NOS. Three types of NOS are NOS1 which also known as neuronal NOS (nNOS); NOS2, also called inducible NOS (iNOS); and NOS3, referred to as endothelial NOS (eNOS). NOS1 and NOS3 are classified as constitutive enzymes that are present on the cell membrane, whereas NOS2 is an inducible enzyme that exists in the cytoplasm.27 The specific characteristics of the three enzymes are shown in Table 1.28,29 NO plays various roles in many disease states, and is closely related to the role and distribution of these three NOSs.30 In inflammatory diseases, the expression of iNOS in activated M1 macrophages increases, thereby catalyzing the reaction between L-Arg and oxygen to produce NO and citrulline.31,32 For example, in M1 macrophages infected with Mtb, increased iNOS expression leads to increased NO production, which can exert anti-TB therapeutic effects by directly killing Mtb and regulating macrophage immunity. Furthermore, NO is also widely used in infectious diseases and NO can react with oxygen or superoxide, yielding RNS, which are instrumental in combating infections, including COVID-19 coronavirus and malaria parasites.33
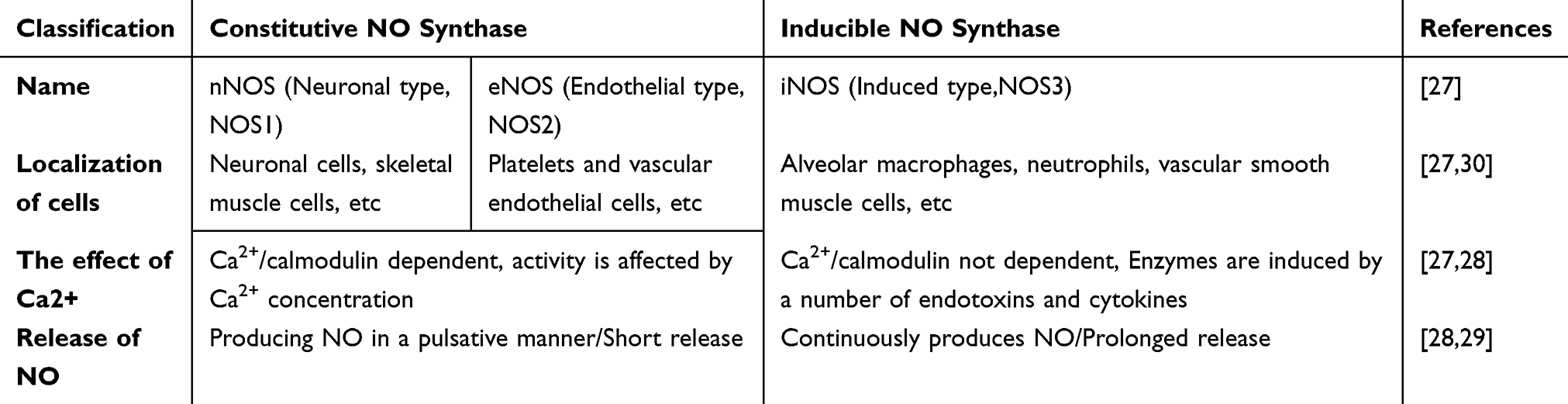 |
Table 1 Classification and Characteristics of NO Synthase |
However, the concentration of endogenous NO is relatively low and direct delivery of NO has many drawbacks, including systemic cytotoxicity and low target concentration, so NO donor can be chosen as an alternative, which is capable of storing and releasing NO, and encapsulating NO donor in nanocarriers enables targeted delivery of drugs, ensures the stability and controlled release of the drugs, reduces side effects, and ultimately improves the therapeutic efficacy of the diseases. Here, we begin by describing the role of NO in several major infectious diseases to set the stage for the later introduction of nanotechnology-assisted NO gas therapy.
Viral Infectious Diseases
Several studies have reported that abnormal NO levels are significant in COVID-19 patients. It has been suggested that low NO levels are closely linked to the ability of SARS-CoV-2 to directly infect endothelial cells, triggering a cytokine storm associated with lung injury. In vitro supplementation with NO or inhaled NO long history as a therapy for hypoxemia.34 Inhaled NO holds promise for COVID-19 prophylaxis and treatment across various stages of the disease. This includes its potential for preventing viral entry, intervening in mild cases, serving as an alternative rescue treatment for moderate to severe cases, and acting as an adjuvant therapy for mechanically ventilated patients.35,36 In a clinical trial, high-dose inhaled NO was proved to be effective in alleviating acute hypoxic respiratory failure caused by COVID-19 by improving arterial oxygenation (PaO2/FiO2).37 All in all, NO plays an important role in improving hypoxia caused in COVID-19. On the other hand, NO has been shown to inhibit SARS-CoV-2 replication by two different mechanisms. First, NO or its derivatives lead to reduced palmitoylation of the de novo expressed spiny (S) protein, which affects the fusion of the S protein with its cognate receptor angiotensin-converting enzyme 2. Second, NO and its derivatives cause a reduction in viral RNA production at early steps of viral replication and Reduction of SARS-CoV-2 protease activity by nitrosating its active site cysteine, which may be attributed to effects on one or both of the cysteine proteases encoded by SARS-CoV-2 Orf1a.33 For this view, in a Phase III clinical trial, exogenous NO supplementation via intranasal administration reduced SARS-CoV-2 RNA load by 93.7 and 99.0% within 24 and 48 hours, respectively.38 Based on the current research progress, it can be seen that NO gas therapy is promising for the treatment of COVID-19.
In addition to this, NO can play an important role in human immunodeficiency virus (HIV) infection. In one study, the authors collected lymph node and blood specimens from 52 HIV-infected patients and measured the expression levels of Arg I and iNOS by immunohistochemistry and fluorescence-based flow cytometry. The results showed that, compared with HIV-uninfected controls, HIV patients had significantly higher levels of Arg I expression in lymph nodes, which correlated positively with viral load, and significantly lower levels of iNOS in the corresponding compartments. This suggests a potential therapeutic strategy to slow the progression of AIDS by modulating the expression of Arg I and iNOS in HIV-infected patients.39 In another clinical trial it was also shown that both HIV infection and a history of TB were associated with lower eNO levels, and that low eNO levels may be a marker of abnormal lung physiology due to perinatally acquired HIV infection and post-tuberculosis lung changes.40 Low levels of eNOS result in insufficient levels of NO produced by endothelial cells to perform their normal immunomodulatory role. Although these results all suggest that HIV infection is associated with reduced levels of NO, the role of NO in the pathogenesis of HIV infection needs to be further investigated.
Parasitic Infectious Diseases
As one of the most successfully adaptive pathogens, parasites infect approximately one - fourth of the global population, posing a severe threat to human health. In mouse models, parasitic infection causes macrophage polarization to the classically activated M1 state, leading to the secretion of inflammatory mediators (such as IL-12 and TNF-α) and the expression of high levels of iNOS and NADPH oxidase (NOX2), which respectively produce NO and ROS, while suppressing or eliminating the parasite.41 At the same time, L-Arg is an important source of NO in macrophages, and parasites, especially Leishmania and Trypanosoma, are highly sensitive to the L-Arg -NO pathway. One reason for this is that NO produced from L-Arg and ROS in M1 macrophages can react to produce various RNS, leading to S-nitrosylation of various proteins, which can lead to death of extracellular parasites.42 To this end, a study in a hamster model of Leishmania protozoa infection significantly reduced parasite load in the spleen by loading exogenous NO donors (DETA/NO precursors) in combination with amphotericin B (AMB) in PLGA nanoparticles for targeted delivery to infected macrophages, and NO nanoparticles alone also exerted antiparasitic effects well. Meanwhile, both AMB and DETA/NO have been reported to trigger iNOS expression in macrophages, which may lead to enhanced intracellular NO.43 Therefore, the present study suggests that NO supplementation through exogenous sources might also play an effective therapeutic role in the fight against parasitic infections.
Bacterial Infectious Diseases
High concentrations of NO can disrupt bacterial proteins and DNA molecules, thus exerting effective antimicrobial activity against bacterial invasion in mammals without causing resistance.44,45 TB is a classic example of bacterial infectious diseases caused by Mtb, and the human body generally mounts a defense mechanism that enables some individuals to remain asymptomatic despite being infected. Approximately 90% of individuals infected with Mtb have no symptoms of TB. In these populations, host immune cells effectively suppressed bacterial replication, but did not completely eliminate Mtb.6,46,47 This safeguard is predicted to play a pivotal role in T cell-derived interferon-γ (IFN-γ), which stimulates the expression of NOS2. Consequently, this leads to the synthesis of NO, a molecule widely acknowledged for its ability to directly suppress Mtb growth, thereby averting subsequent pathological inflammation and playing a crucial role in the prevention.48
The specific mechanisms of NO and other RNS combat Mtb might include damaging bacterial DNA, proteins, and signaling pathways.47,49 For example, in macrophages infected with Mtb, NO treatment leads to differential expression of genes encoding hypothetical proteins (HPs), which likely to contain DNA-binding domains and may play a role in DNA repair.50 In addition to this, there are many studies showing that NO exerts a direct killing effect on Mtb by affecting signaling pathways. Weikert et al demonstrated that in macrophages infected with Bacillus Calmette-Guérin (BCG), surfactant protein A (SP-A) can enhance the signaling function that initiates NO production, thereby augmenting the bactericidal activity of rat macrophages against mycobacteria. The attenuation of this effect following the application of iNOS inhibitors further supports this result.51 The effects of NO on signaling pathways also include irreversible inactivation of cytochrome c oxidase and other respiratory proteins, as well as induction of DosR regulators, through which NO may exert a bacteriostatic effect on Mtb by affecting the bacterial respiratory chain and energy metabolism.52 In the study by Hu Qian et al, it was mentioned that NO is a broad-spectrum antibacterial agent, and its mechanism of action is not affected by bacterial resistance. They summarized the progress in the use of S-nitroso glutathione (GSNO) as a NO donor in antimicrobial therapy, showing that the antibacterial mechanism of NO is different from common antibacterial agents, and demonstrating a promising therapeutic agent for a variety of infectious diseases.53 In addition to the above findings, an observational study based on sputum samples from TB patients showed that Mtb has a biphasic response to NO; that is, at low concentrations of NO (50–100μM), the proportion of liposome-positive cells (%LB+) in sputum increased significantly, which was associated with treatment failure or relapse after anti-TB chemotherapy. However, at high concentrations (250–500μM), NO showed a toxic effect on Mtb, leading to cell membrane damage and bacterial death.54 Moreover, a clinical trial of exhaled NO levels in children with a history of TB and HIV infection also showed that NO levels were lower in such patients.55 It can be seen that the determination of the concentration of NO gas therapy is the key when it is applied to anti-TB treatment. Although many studies at this stage have shown that NO exerts its anti-TB effect mainly through direct bactericidal action, there is also study showing that NO protects mice by inhibiting Mtb replication primarily by inhibiting the neutrophil recruitment cascades, which dependents on the interleukin-1 and 12/15 lipoxygenase.48 We still know very little about the applications of NO in TB, including the research on its specific killing mechanism against Mtb. Therefore, to effectively apply NO in the biomedical field, it is necessary to overcome these problems. Many studies have shown that NO can effectively kill Mtb, but it has been ignored that Mtb may evolve potential adaptive defenses in response to the killing effect of NO. PPE2, a distinct member of the PPE protein family in Mtb, has the ability to restrict the generation of NO by suppressing the transcription of the iNOS gene.56 When Mtb expresses this protein, it can resist the killing effect of NO. This reminds us that when studying the anti-TB therapeutic effect of NO, we should not only pay attention to its bactericidal effect but also to its induced defense effect against Mtb.
There has great potential for the comprehensive treatment of TB. The development of nanomedicines that combine photothermal therapy (PTT) with photodynamic therapy (PDT) is considered a therapeutic strategy for cancer treatment. NO is also known to enhance the PTT and PDT. Based on the above theory, a mitochondria-targeted NO nanogenerator, EArgFe@Ce6, was created, which has superior photothermal ability and can reduce hypoxia by targeting the mitochondria, thereby enhancing PDT. Enhanced PDT can produce large amounts of ROS and further promote NO production from L-Arg. Finally, synergistic photodynamic/gas/photothermal therapy is realized under light irradiation.57 Moreover, the local high temperature of PTT can damage the surrounding healthy tissues, and synergistic NO therapy can enhance the photothermal effect; thus, it can play a role at low temperatures.58 Such a superior synergistic treatment strategy may have great application value in the field of TB treatment in the future.
We can think of NO as a broad-spectrum antimicrobial agent that can also play a role in a variety of other bacterial infectious diseases. NO, as an endogenous inflammation modulator and antimicrobial agent with no apparent resistance, has emerged as an attractive candidate for wound healing therapy.59 In a study on the treatment of infected burn wounds, it was reported that nanomotors loaded with a sulfuric nitric oxide (SNO) donor can self-propel and penetrate deep into the biofilm by generating NO. The locally released NO destroys bacteria within the biofilm, providing a non-antibiotic approach to fighting biofilms without developing resistance.60 Similarly, in diabetic wounds, inflamed wounds provide a suitable environment for bacterial colonization, and infection leads to tissue damage and triggers further inflammation. In a study by Chenxi Tu et al a multifunctional hydrogel with exogenous supplementation of NO was proposed, which down-regulates virulence genes and interferes with bacterial metabolism. This multifunctional hydrogel was able to kill methicillin-resistant Staphylococcus aureus (MRSA), Escherichia coli (E. coli), and Pseudomonas aeruginosa by 94.1%-99.5%, which was effective in treating in vivo infections such as those caused by MRSA and thus significantly facilitated the healing of wounds in the inflammatory phase.61 In addition to its widespread use against wound infections, NO has been extensively studied in other diseases caused by bacterial infections in vivo. For example, recent in vitro data and case reports suggest that inhaled NO (gNO) in gaseous form has antimicrobial activity against non-tuberculous mycobacterial lung disease (NTM). And results from an open-label proof-of-concept trial suggest that gNO does reduce bacterial load in sputum cultures to some extent.62 The downside is that the relatively small number of subjects in this study is not enough to fully confirm the validity of this approach, but it is at least supportive of subsequent research.
In conclusion, We think that NO has great application prospects in the treatment of bacterial infectious diseases, which can be achieved by delivering NO donors via nanocarriers or inhaling NO gas directly, thus exerting direct bactericidal effects or modulating host immunity.
Other Infectious Diseases
NO synthesized by iNOS has also been implicated in the control of virulent rickettsiae in several cell types in other infectious diseases. Specifically, NO significantly reduces rickettsial adhesion by destroying bacterial energy. In addition, NO-treated rickettsiae are unable to synthesize proteins or replicate in infected cells. These data suggest that NO is a potent innate immune anti-Rickettsia effector.63 In terms of antifungal infection, a study was conducted by loading NO and the antifungal drug clotrimazole (Clo) onto nanoparticles, where NO can effectively inhibit the formation of fungal hyphae, have a dissipative effect on mature biofilm, and can further bind with Clo to completely eradicate the fungus within the biofilm. Of course, this synergistic effect can also effectively eliminate Candida albicans in the planktonic state, and ultimately effectively cure Candida albicans-induced vaginal infection in mice.64
NO is an important endogenous small-molecule mediator that plays an important role in regulating the occurrence and development of many infectious diseases. Moreover, there are clinical trials related to the therapeutic application of NO inhalation therapy in a variety of infectious diseases, such as severe malaria and COVID-19.37,65,66 These clinical trials further illustrate the potential of NO gas therapy for the treatment of TB. Controlling the concentration of NO is the key to the treatment of many diseases. It can be reduced using NOS inhibitors or increased by the exogenous delivery of NO donors to regulate the treatment process of many diseases. Controlling the concentration and location of NO under pathological conditions will be an important strategy for disease treatment.
Potential Application of Nanotechnology-Based NO Therapy in the Treatment of TB
Types of NO Donors, and Their Advantages and Disadvantages in Disease Intervention and Treatment
NO has strong therapeutic potential, but it has high reactivity and high concentration-dependent physiological effects in air; therefore, it is not convenient to use NO gas directly in clinical trials. Therefore, many NO donors have been developed that can reversibly store and release NO under specific circumstances, including diazeniumdiolates (NONOates), S-nitrosothiols, organic nitrates, nitrates/nitrites, and metal-nitroso complexes.67,68 (Figure 2) This article describes the characteristics of several common NO donors, each of which has advantages and disadvantages in the therapeutic application of diseases (Table 2).
 |
Table 2 Advantages and Disadvantages of NO Donors and Their Potential Delivery Systems |
 |
Figure 2 Diagrams of the chemical structures of six common NO donors. Created in BioRender. Liu, J. (2025) https://BioRender.com/g8a1ra8. |
NONOates
NONOates are a series of compounds possessing an X-[N(O)NO] − functional parent nuclear structural unit. NONOate is an effective substance that imitates the transient biosynthesis of NO. Because NONOates produce NO at different rates, NONOates are valuable in mimicking the low, transient synthesis of NO by eNOS and nNOS.69 Moreover, NONOates do not produce by-products other than the original amine after NO release.70 However, in the process of NO release, if the quantity and rate of NO release cannot be accurately regulated, such spontaneous systemic release may give rise to significant off-target effects. In particular, at high concentrations, this can result in adverse effects in diverse cell types.69 Despite the promising application of NONOates, their clinical use has been limited.
S-Nitrosothiols
S-Nitrosothiols are derivatives of NO, formed by the S-nitrosation of thiols or cysteine residues of proteins, and have the general formula RSNO.85,86 The S-NO bond of RSNO is destroyed under the stimulation of ultraviolet light, heat, metal ions, etc, resulting in the production of NO.71–73 In preclinical investigations, no adverse effects associated with RSNO have been recorded. Both in vitro and in vivo experiments demonstrated that certain RSNOs displayed relatively minor cytotoxicity when administered at concentrations relevant to pharmaceutical applications. The presence of flexible metabolic pathways reduces the likelihood of tolerance induction after long-term use. Consequently, RSNO has emerged as an especially auspicious category of NO donors for in vivo use.67,87 However, the stability of RSNO is insufficient to meet clinical requirements. Additionally, the hydrophilic nature of RSNO and the lability of the S-NO bond during the formulation process augment the complexity of incorporating RSNO, thereby constraining their concomitant use with delivery systems.67 Therefore, in order to use RSNO as one widely used NO donors, it is essential to solve their instability.
Organic Nitrates
Organic nitrates, which assume the nitrite ester form and possess the nitroxide functional group (-ONO2), represent the most prevalent NO-releasing pharmaceuticals.74 Among these, glyceryl nitrate (GTN) is the most extensively investigated NO donor and has been used to alleviate acute angina pectoris episodes for over 150 years.71,75 Nevertheless, prolonged administration of nitroglycerin might result in nitrate tolerance or attenuation of the natriuretic property of such organic nitrates. It can also augment oxidative stress, give rise to vascular and intravascular adverse effects, and precipitate endothelial dysfunction.76 Therefore, nitrate tolerance, oxidative stress, and endothelial dysfunction are major limiting factors for the use of these organic nitrates as NO donors. But by loading them into drug delivery systems and using them synergistically with other therapeutic options, such as drugs containing gold nanoparticles, it may be possible to reduce the use of such drugs and mitigate side effects.77
Nitrate/Nitrite
Oral microbiota can reduce nitrate to nitrite, resulting in the production of NO.88 The NO metabolite nitrite is acknowledged as a particular NO donor within hypoxic/acidic circumstances.78 However, a significant disadvantage of inorganic nitrite treatment might be the generation of nitrosamines or the consequent occurrence of methemoglobinemia.79,80 Furthermore, nitrite serves as a physiologically significant NO donor when present at low concentrations; however, it exhibits toxicity at elevated levels. The build-up of nitrite gives rise to the overproduction of NO and the induction of nitrosative stress.89 Inorganic nitrate and nitrite treatment seemingly constitutes an economical and efficacious cardiovascular therapeutic approach, with relatively fewer adverse effects in comparison to organic nitrate treatment. Nevertheless, additional research is essential to establish the effectiveness of this treatment modality in diverse cardiovascular ailments, including regulation of dosage and treatment duration.
Metal−Nitrosyl Complexes
The metal nitroso complex encompasses at least one NO functional group that is directly affixed to the metal center, which is designated as the metal nitroso group.71,72 Sodium nitroprusside (SNP) represents the first metal nitroso complex to be identified and has been the subject of extensive research.90,91 Although the precise mechanism by which NO is released from SNP remains incompletely elucidated, it has been unequivocally demonstrated that the release of NO necessitates either light irradiation or single-electron reduction irradiation.81 However, the dissociation of SNP, which leads to the release of cyanide, gives rise to cytotoxicity.73 There is toxicity resulting from the dissociation of cyanide during sodium nitroprusside administration, which leads to the recommended level.82 Consequently, SNP ought to be administered in a minimal dose, the dissociation of cyanide must be monitored and regulated continuously to the release of cyanide, gives rise to cytotoxicity.
Arginine
Arginine is an important precursor for NO production in mammals.92 Systemic administration of arginine has been reported for many years to release NO in humans.93 In mammals, arginine can be decomposed by three variants of NOS to generate NO and citrulline. Subsequently, citrulline can synthesize arginine via the citrulline-NO cycle, thereby facilitating efficient NO synthesis.84,94,95 Since arginine can produce abundant NO without toxic byproducts and arginine supplementation has been shown to be a safe therapeutic option.83 There is clinical study have shown that supplementation with L-Arg and vitamin C(VC) plays a synergistic role in improving the bioavailability of NO to improve the sequelae of acute COVID-19.96 M1 macrophages are known to be rich in iNOS, which can efficiently utilize L-Arg to produce NO.97 Therefore, arginine supplementation is likely to be an effective strategy for the treatment of macrophage-related diseases such as TB. However, arginine, as a “new NO donor”, requires the participation of NOS, oxygen, and other cofactors to decompose NO, limiting its clinical application. Arginine is a basic raw material used for protein synthesis. It can be taken up by bacteria, that is, it has a nutritional effect on bacteria.84 In the future, if arginine is to be widely used as a NO donor in clinical practice, it must be delivered to the environment that is conducive to its decomposition to produce NO through targeted delivery systems, and its nutritional support role should be considered during antibacterial or antiviral therapy.
Despite the significant application prospects of NO donors, several shortcomings remain to be overcome. These include toxicity, storage issues arising from photochemical and temperature instability, challenges in modulating long-term release, and resistance to certain compounds. By comparing the advantages and disadvantages of several commonly used NO donors in the treatment of diseases, we can better choose NO donors for the treatment of target diseases and improve their therapeutic effects.
Nanotechnology-Assisted NO Therapeutics
To augment the stability and adjust the NO release patterns, small-molecule NO donors were conjugated with the polymeric scaffolds through covalent bonds. This endows them with multifunctional integration, protracted release periods, and enhanced therapeutic efficacies.68 The NO donor and polymeric scaffolds constitute a NO delivery system, which can release NO in a microenvironmental or exogenous stimulus-controlled release mode, such as pH response, light response and temperature response. This helps to improve the storage stability of NO donors and reduce the systemic toxicity caused by accidental NO exposure.98–102 In addition, this controlled drug delivery system (DDS) requires relatively low drug doses.103 This review briefly introduces several commonly loaded biomaterials and their common preparation methods and expounds the principles of drugs released by delivery systems and their application in the treatment of diseases (Figure 3).
 |
Figure 3 Common controllable nitric oxide-delivering platforms for biomedical application. (a) 1. Nanoparticles were synthesized using microfluidic technology. 2.The drug is released in response to X-rays and near infrared light (NIR). (b) 1. Liposomes were synthesized using membrane hydration method. 2.The vaccine was prepared by adding adjuvant to the liposome loaded with mRNA and inoculated. 3.In the acidic environment of viral infection, liposomes release mRNA in response to pH. 4.Viral mRNA is expressed into viral proteins, which are recognized by the immune system and produce specific antibodies to play an antiviral role. (c) 1. The functional groups interact with each other to synthesize hydrogels. 2.As wound dressings, drug-loaded hydrogels can exert anti-infective and pro-angiogenic effects. 3.Hydrogels can release drugs in response to pH, temperature, electricity, and so on, depending on the composition of the synthesis. Created in BioRender. Liu, J. (2025) https://BioRender.com/p3uitxd. |
Nanoparticles
In terms of biomedical applications, nanomaterials can be used for the delivery of drugs, bioimaging, and biosensors.104 Nanoparticles are the preferred drug carriers for biomedical applications owing to their small size (less than 100 nm in one dimension) and display distinctive physicochemical and biological characteristics (for example, augmented reaction zones and the capacity to traverse cell and tissue barriers).103 Numerous approaches to nanoparticle synthesis, such as emulsification, solvent diffusion, and emulsification–reverse salting-out methods.105 Among the preparation methods of nanoparticle drug delivery systems, microfluidics have emerged as an advanced method, which will facilitate the large-scale production and translation of nanoparticles for clinical applications.106 To load a drug in a nanoparticle, the drug can either be adsorbed or predominantly attached to the nanocarrier surface, or they may be encapsulated within.103 In the treatment of diseases, several investigations have covalently conjugated the NO donor GSNO with polydopamine nanoparticles (PDA-GSNO NPs). Such nanoparticles enable near-infrared (NIR)-regulated NO release and PTT, thereby augmenting in vivo wound healing efficacy.107 Another type of nanomaterial, based on scintillation nanoparticles (SCNP) and the UV-responsive NO donor Rusin black salt (RBS), can generate both NO and O2−· upon X-ray irradiation, leading to the formation of ONOO−. This strategy can effectively enhance the efficiency of radiotherapy and, consequently, elevate the level of cancer treatment.108 These findings indicate that the combination of phototherapy and nanoparticles can effectively control the production and release of NO, thereby improving the therapeutic effect of the disease. And on the other hand, many metallic nanoparticles have therapeutic effects of their own. For example, a silver nanoparticle based on green chemistry has recently been reported to be very effective against Mycobacterium spp. and multi-drug resistant (MDR) strains, including a wide range of strains such as Acinetobacter baumannii isolated from patient samples.109 What’s more, our group has designed a kind of zinc oxide-selenium nanoparticles (ZnO-Se NPs), and it was found that this kind of metal nanoparticles had a strong killing effect on extracellular Mtb, including BCG and virulent H37Rv. Moreover, ZnO-Se NPs could inhibit intracellular Mtb growth and exert host cell immunosuppressive effects by promoting M1 polarization to increase the production of the fungicide NO.110 These findings further demonstrate the promising application of nanoparticles in the treatment of diseases.
Liposomes
Liposomes represent a closed phospholipid bilayer spherical vesicle system, which is composed of amphiphilic molecules, including phospholipids and steroids (eg, cholesterol) or other surfactants, and they have the capacity to carry hydrophobic/hydrophilic molecules.111 Liposomes usually have a size range of 80–300 nm, which form spontaneously when certain lipids are dispersed in aqueous media where liposomes can be prepared.112,113 Specifically, liposomes can be generally classified into solvent-free methods and solvent replacement methods. Ultimately, by repeatedly passing the crude heterogeneous lipid mixture through a polycarbonate membrane with minute pores, uniformly distributed small-molecule liposomes with diameters corresponding to the pore size can be acquired. Liposomes possess the capacity to enclose hydrophilic substances within their lumen and hydrophobic substances within their membrane.114 The liberation of a drug from liposomes is contingent upon the liposome composition, osmotic gradient, and ambient environment. For example, due to the oxidation of unsaturated fatty acid chains by ROS or an acidic infection environment, lead to the disintegration of liposomes and the release of liposome-loaded drugs.115 In the domain of disease treatment, liposomes, by virtue of their favorable biocompatibility, versatile structural modification, and adjustable properties, are extensively employed as carriers to enhance the efficacy of drug therapy. Notably, in recent years, against the backdrop of COVID-19, liposomes have been developed as a multifunctional vaccine adjuvant delivery system (VADS), which has substantially augmented the effectiveness of vaccination.116 This reflects the flexible loading capacity of liposomes as a drug delivery system, which has great application prospects in disease treatment.
Hydrogel
Hydrogels are a class of substances that possess 3D polymer networks that can swell in water or biological fluids and remain undissolved owing to chemical or physical crosslinks.117–119 In the preparation of hydrogels, one of the simplest and most prevalent methods involves mixing all precursors in an aqueous solution. When these mixtures are injected, a hydrogel can be conveniently formed in situ because of the reaction between the functional groups of the precursors.120 The hydrogel can release drugs in response to pH, temperature, and electricity by changing their composition and synthesis method. For example, a conductive polymer hydrogel (CPH) can be synthesized by adding a conducting polymer, poly(3,4-ethylenedioxythiophene), to a hydrogel to release proteins in response to electrical stimulation.121–123 Hydrogels possess favorable adhesion, biocompatibility, and flow characteristics, making them excellent materials for wound dressings. Zheng et al designed a hydrogel loaded with dual NO donors, namely GSNO and binary L-Arginine (bArg) (referred to as CAT/bArg/GSNO). This hydrogel has the potential to promote angiogenesis and exhibits antibacterial properties in chronic wounds, thus serving as a promising dressing for wound repair.124 The favorable biocompatibility, predictable degradation rate, adaptable mechanical properties, and excellent elasticity of hydrogels render them outstanding materials for biomedical applications.
For the selection of an NO donor delivery system, the first consideration is the cytotoxicity caused by NO leakage. Therefore, the stability of the delivery system must be good. Compared to the above three delivery systems, hydrogels are not an appropriate choice because of their low mechanical strength and ease of rupture, leading to leakage of internal NO donors and further release of NO. Secondly, under physiological conditions, arginine in macrophages slowly releases NO through the iNOS pathway; therefore, it is better to deliver materials that meet this property for better antibacterial effects. Compared to nanoparticles and liposomes, liposomes can release NO more slowly and prolong the action time. Although nanoparticles can be loaded with more drugs, liposomes can be loaded with both hydrophobic and hydrophilic drugs, which is the best choice for simultaneous loading of a hydrophilic NO donor and hydrophobic RIF (rifampicin) at the same time. In addition, liposomes have good biocompatibility and can enter macrophages to exploit the advantages of host immunotherapy for TB. The rapid clearance of liposomes in vivo has hindered their clinical application. Drug delivery materials do not have only one characteristic. For example, some current drug delivery systems synthesize lipid nanoparticles(LNPs) by combining the advantages of liposomes and inorganic/organic nanoparticles into one system, in which the diverse nanoparticles compensate for the poor stability and rapid elimination of liposomes from the blood circulation.125 With the increasing diversification of medical material delivery systems, the strategy of loading NO donors into the delivery system for targeted therapy effectively avoids some of the defects of the NO donor itself, but there is still much room for further development. Future research is mainly focused on continuously improving the stability of the drug delivery system to ensure the storage stability of the loaded drug and to reduce the chance of underdosing at the target site and toxic side effects caused by accidental drug leakage. This is also the direction of our efforts to find more targeted molecules and to modify them into delivery systems.
Potential Applications in TB Treatment
Based on the above studies on NO donors and nanodrug delivery systems, it can be seen that nanotechnology-assisted NO gas-based therapies have great promise in the treatment of TB. Specifically, it can be analyzed in terms of the mechanism of NO against Mtb and the advantages of the new nanomedicine are expected to solve the problem of insufficient treatment for TB at this stage.
Anti-TB Mechanisms Associated with NO
Macrophages are the principal target cells of Mtb infection and have developed a variety of defence strategies to counteract the infection caused by Mtb. Specifically, upon Mtb infection, macrophages perceive diverse Mtb pathogen-related molecular patterns and respond by initiating antimicrobial pathways, among which are the induction of toxic antimicrobial effectors, such as NO and ROS.126 The function of NO in vivo is highly contingent upon the concentration generated within a specific context, as well as the location and timing of its their synthesis (Figure 4). Consequently, a thorough understanding of NO metabolism within the macrophages of TB patients is necessary to clarify how NO gas therapy can be utilized in the treatment of TB.
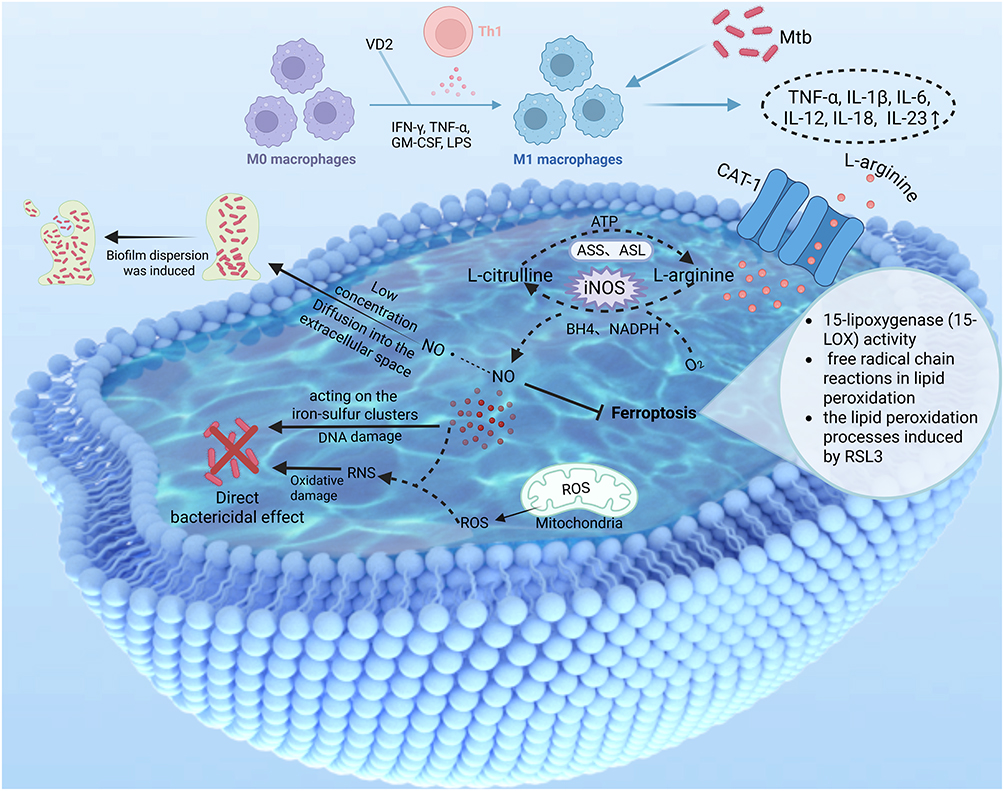 |
Figure 4 Major anti-Mtb mechanisms of NO in M1 macrophages. Created in BioRender. Liu, J. (2025) https://BioRender.com/gcfqdzr. Abbreviations: VD2, Vitamin D2; GM-CSF, Granulocyte-Macrophage Colony-Stimulating Factor. |
Anti-TB Mechanism of iNOS
Interferon-γ (IFN-γ), with microorganisms or their products, such as lipopolysaccharides (LPS), can contact with macrophages to trigger the classical activation of macrophages into M1, which directly inhibits Mtb replication.127–132 Activated M1 macrophages possess a wide range of antimicrobial capabilities, characterized by the catalytic reaction of arginine and oxygen to citrulline and NO via iNOS located in the cytoplasm.31,133,134 In vitro experiments have demonstrated that M1 macrophages rely more on CAT-1 for the uptake of L-Arg.135 Although human macrophages express iNOS at much lower levels than rodent macrophages, patients with inflammation or infection have elevated levels and produce NO.32 And it has been shown that the rate of intracellular bacterial growth in macrophages varies markedly from macrophage to macrophage, and that this heterogeneity is driven by changes in the activity of intercellular iNOS, which is why some macrophages are able to kill Mtb intracellularly, whereas others are unable to control bacterial replication.136 It is evident that the expression of iNOS within macrophages is important for the fight against TB. On what factors iNOS expression is affected in macrophages, it has been reported in the literature that the transcription factor YY1 binds to the iNOS promoter region, and YY1 overexpression significantly enhances the iNOS promoter activity, thus promoting the transcription and expression of the iNOS gene, and ultimately the highly expressed iNOS promotes the production of NO, which exerts a direct bactericidal effect to clear intracellular Mtb.137 Mtb has also adopted defense mechanisms against iNOS-mediated host resistance to Mtb, such as a significant reduction in the level of amyloid β(A4)-precursor-like protein 2 (Aplp2) in TB patients, Mtb-infected macrophages, and mice, accompanied by a decrease in the expression of iNOS. In response to this defense mechanism, it has been shown that the induction of Aplp2 expression, which increases the expression level of iNOS and promotes Mtb clearance through NO-mediated bactericidal action, could ultimately play a key role in protection against Mycobacterium infection.138 It can be seen that iNOS still has great application prospects in anti-TB treatment, and future research can induce its expression in macrophages infected with Mtb at elevated levels through various pathways, so as to kill Mtb more effectively.
Direct Anti-TB Mechanisms of NO
Activated macrophages generate ROS in the mitochondria.139 Once NO permeates the phagosome membrane, it spontaneously reacts with ROS within the phagocytic cavity, giving rise to RNS. These highly reactive intermediates carry out their microbicidal function through oxidative damage to membrane lipids, DNA, thiols, and tyrosine residues. NO can display its toxicity by directly acting on the iron-sulfur clusters of central metabolic enzymes and affecting bacterial signaling pathways, such as the bacterial respiratory chain.126 Addressing DR-TB is critical for achieving the overarching goal of eliminating TB. DR-TB can be caused by Mtb resistance driven by infection with drug-resistant bacteria in the environment and the misuse of antibiotics.140 Therefore, prevention of DR-TB prevents the generation of drug-resistant Mtb. When Mtb forms a biofilm, it can lead to an increase in the dose of antibiotic chemotherapy, resulting in drug resistance. Interestingly, at lower concentrations, reports indicated that NO can inhibit the formation and dispersion of biofilms by activating the functions of effector proteins and inducing the breakdown of biofilms through the cyclic di-GMP pathway. Thereby diminishing bacterial tolerance to antibiotic interventions141 and increasing bacterial sensitivity to antibiotics,142 which can significantly reduce the amount of antibiotics used. Therefore, NO gas therapy in combination with antibiotics may solve the difficult problem of DR-TB treatment. Nevertheless, to manifest antimicrobial activity via these pathways, NO must be continuously synthesized. Therefore, a restricted supply of L-Arg impedes NO formation. Fortunately, citrulline, the end product of the NO pathway, can be used to synthesize arginine through the citrulline-NO cycle, which is regulated by arginine-succinate synthase (ASS) and arginine-succinate esterase (ASL).94 Given that arginine can produce NO in macrophages to play an anti-Mtb role, and this bactericidal effect is limited by the availability of arginine, exogenous supplementation of arginine will help M1 macrophages to exert an immune response, which may be an effective anti-TB strategy (Figure 4).
Mechanisms of NO-Related Host-Targeted Therapy in Anti-TB
The host immune system plays a key role in Mtb infection, and host-directed therapy (HDT) has emerged as an effective strategy for the treatment of TB, including drug-resistant TB (DR-TB).143 Ferroptosis is the consequence of an uncontrolled state of three processes, iron metabolism, lipid peroxidation, and thiol regulation, which are frequently observed in a pro-inflammatory context.144 Ferroptosis serves as a principal means of necrosis during Mtb infection and represents a target for host-directed TB treatment.145 Activated M1 macrophages generate nitric oxide radical (NO•) via the iNOS pathway. NO• can inhibit ferroptosis as it suppresses 15-lipoxygenase (15-LOX) activity, free radical chain reactions in lipid peroxidation, the lipid peroxidation processes induced by RSL3 and so on (Figure 4).4,127 Furthermore, over two decades ago, it was demonstrated that NO• functions as an inhibitor of 15-LOX-catalyzed oxidation. This implies that iNOS/NO• might substitute GPX4 to safeguard against ferroptosis.146 So as for how NO regulates host macrophages to exert anti-TB effect, we hypothesized that ferroptosis may be an effective way. Further studies are needed to determine whether NO inhibits ferroptosis in Mtb-infected macrophages.
In the complex treatment of TB, NO exhibits complexity, including the expression of iNOS, which is influenced by multiple factors. On the one hand, NO and ROS cooperate to produce RNS, which play a direct killing effect on Mtb. Low NO concentrations can induce the spread of bacterial biofilms, thereby improving the sensitivity of drug-resistant bacteria. On the other hand, NO can indirectly kill Mtb by regulating macrophage activity, including inhibiting ferroptosis. In addition, we cannot ignore the duality of NO, that is, to ensure their killing activity, but also to avoid their cytotoxicity. We have not yet seen any reports of dose thresholds or therapeutic windows for NO in TB treatment, but it is known that the antimicrobial effect can be exerted only when the NO concentration reaches the 250–500 μM level.54,147 So determination of the therapeutic concentration and duration of NO will be the first step in conducting our research efforts. It’s really not that difficult to design materials containing different concentrations of NO to perform cytotoxicity experiments at different time points, such as 12 hours, 24 hours, 48 hours or even longer time points. Under the condition of high cell survival rate (eg, more than 80%), we can initially determine the corresponding NO concentration and time of action, which can be used as a dosage threshold and a therapeutic window, and then carry out the later experiments to evaluate the therapeutic effect. If a good therapeutic effect can be achieved, then it indicates that the NO concentration and treatment time we choose are appropriate. Of course, a combination of experiments with different concentrations and time points would have to be performed, with the aim of screening out the best NO treatment concentration and time. We think that this may solve the difficulty of concentration dependence of NO gas treatment and achieve the effect of anti-Mtb without toxicity to cells.
Potential Applications
In the treatment of TB with traditional antibiotics, including RIF and isoniazid (INH), Mtb can develop drug resistance, which is an inescapable result of drug use.148 In addition, long administration times, serious side effects, and poor patient compliance lead to further drug resistance and treatment failure, eventually leading to multidrug-resistant tuberculosis (MDR-TB) and extensively drug-resistant tuberculosis (XDR-TB).149 For this reason, the WHO has underlined the need to invest in the research and development of novel TB medicines.150
Considering that NO can exert anti-Mtb effects, it may be the main component of new anti-TB drugs. With the excavation of exogenous NO or NO donors and the design of drug delivery materials such as nanomaterials, the combination of an NO donor and drug delivery platform, and their delivery to the target site through targeted action, can not only ensure the sterilization effect, but also have good biological safety, which will be the design strategy for new anti-TB drugs. Compared with traditional NO donors, arginine may be a better choice because it is the main source of NO in mammals and can be produced by the iNOS pathway in Mtb-infected macrophages, making it a potential source of NO for anti-TB treatment.126 In addition, LNPs have been widely studied as drug carriers, are regarded as the most advanced nucleic acid delivery technology, and have been proven to be very successful as a delivery platform for COVID-19 mRNA vaccines.151,152 The structure of LNPs, which have an aqueous core and a lipid bilayer structure, can encapsulate drugs with different properties, such as arginine and RIF, for delivery. In addition, LNPs have the advantages of high encapsulation efficiency, low toxicity, enhanced cellular uptake, high stability, and long circulation time.153–155 Moreover, their stability and tissue uptake can be improved by changing the lipid composition and ratio.153,156 This suggests that arginine can be effectively loaded into LNPs by changing the lipid composition.
Considering the concentration dependence of the physiological functions of NO, the targeting and controllable release of drugs is essential. The targeting of the drug, because Mtb mainly infects and settles macrophages, refers to targeting macrophages. Strategies for targeting macrophages infected with Mtb can be divided into two types: passive and active. The former is mainly because PEGylated liposomes and other types of nanoparticles can cross the endothelial barrier near the site of infection within minutes after injection by enhancing the permeation and retention (EPR) effect and accumulating at the site of inflammation.157 However, such targeting is still not as good as the latter. Currently, active targeting of macrophages can specifically recognize macrophage surface receptors (eg, mannose and scavenger receptors) through surface modifications (eg, antibodies and ligands). For example, mannose-modified nanomaterials can target macrophages by binding to the mannose receptor on their surface, thus entering macrophages to exert intracellular killing effects of Mtb.158–161 In addition, modification with the demonstration of tuftsin peptide on the nanocarrier can achieve enhanced localization and active targeting of macrophages.162 Moreover, hyaluronic acid-modified materials that can bind to the highly expressed CD44 receptor on the surface of macrophages are also a good strategy.163 Therefore, whether in the TB model or the osteoarthritis model, it is speculated that after ligand modification on the surface of nanoparticles, nanoparticles can actively target Mtb infected macrophages with high expression of corresponding receptors. Through various pathways mediated by ligand-receptor binding, they enter macrophages to exert intracellular killing effects on Mtb. However, when selecting a surface-modified ligand, it is necessary to consider whether their connection with the drug delivery system is stable and whether its specificity for targeting macrophages is high. The three ligand modified nanomaterials mentioned above were compared because there is a highly expressed mannose receptor (CD206) on the surface of macrophages with high affinity,161 while tuftsin peptide can interact with Fc and neuropilin-1 (Nrp1) receptors on macrophages with good specificity.164,165 However, in contrast to mannose modification targeting a single well-defined receptor, the receptors on which tuftsin peptides act are less prevalent and well-defined. The CD44 receptor targeted by hyaluronic acid is expressed on a variety of tumor cells,166 which results in less specific targeting to macrophages than mannose modification and tuftsin peptide modification. In addition, mannose can be modified by covalent conjugation to lipid components such as PEGylated lipids,161 which greatly improves the stability of the modification. Tuftsin is a tetrapeptide composed of threonine (Thr), lysine (Lys), proline (Pro) and arginine (Arg), its structure lacks certain stability. Therefore, based on the comparison of targeting specificity and modification stability, mannose-modified LNPs may play a better role in the treatment of TB. Controlled release of NO can also be achieved directly by altering the lipid components. Based on the development of various stimulus-responsive delivery platforms, stimulus-responsive liposomes can release the embedded drug in response to the presence of exogenous or endogenous stimulants when exposed to the target site.167 Traditional thermosensitive liposomes (TTSL) contain phospholipid components that can change the physical state of lipids from a solid gel ordered phase to a highly permeable liquid crystal phase at the phase transition temperature (Tm), leading to the osmotic release of drugs through the membrane.168 In addition, when liposomes contain DOPE, a pH-sensitive lipid component that can become unstable in response to an acidic environment in infected cells and release the drug through enhanced fusion of liposomes with endosomal membranes.169 In the study of Zimei Wu et al, a liposome containing PEG-benzaldehyde hydrazone lipid (PEGB-Hz-CHEMS) was reported to release drugs in pancreatic cancer cell model and animal model in response to pH.170 The hydrazone bond and ester bond are sensitive to pH, which are easy to hydrolyze and break under acidic conditions, further promoting the structural disruption of the LNPs and the release of the drug, but relatively stable under neutral or basic conditions. Therefore, it can be hypothesized that LNPs containing hydrazone or ester bond lipid components can release the loaded drug in Mtb infected macrophages through pH response. In addition, one study has reported that AuNP can rapidly convert light energy into heat energy under NIR laser irradiation by combining heat sensitive-based liposome (HSL) and gold nanoparticles in breast tumor models, so that the local temperature is increased above the phase transition temperature of HSL, and the lipid membrane of HSL changes from a gel state to a liquid crystal state, and the membrane permeability is significantly increased. Rapid release of the encapsulated drug.171 Therefore, such treatment strategy may be very suitable for the treatment of cutaneous TB. Another recent study showed that when liposomes loaded with L-Arg reach inflammatory sites (such as atherosclerotic plaques) or acidic environments within macrophages (pH 6.5 to 5.0), the charge and conformation of the liposome membrane change, which promotes the release of L-Arg. At the same time, NOS in macrophages catalyzes the production of NO from L-Arg, which consumes L-Arg and creates a concentration gradient that promotes the sustained release of L-Arg in liposomes to replenish the substrate.172 This pH-sensitive and enzyme-catalyzed liposome may also be a good choice for targeting macrophages infected with Mtb for the treatment of TB. At present, there are few reports on the application of LNPs with controlled drug release in the treatment of TB, but based on their application in the treatment of tumors and other inflammatory diseases, it can be concluded that LNPs may be a feasible strategy for the treatment of TB. In conclusion, LNPs, which combine the advantages of nanoparticles and liposomes, may be an excellent choice for arginine delivery platforms through surface modification and modification of liposome composition. By targeting macrophages and releasing arginine in response, it produces NO and exerts an anti-MTB effect, which may represent an innovative strategy for the development of anti-TB drugs.
Outlook
Although TB is curable, approximately 25000 people die every week.173 DR-TB emerges when Mtb acquires resistance to any anti-TB drug. Although bedaquiline (Bdq) and delamanid (Dlm) are used to treat DR-TB, DR-TB remains a global crisis because of the increasing incidence of drug-resistant strains.174 NO exerts antibacterial effects at high concentrations through destructive effects on proteins and DNA molecules.175 Moreover, it can reduce the tolerance of bacteria to antibiotic intervention by eliminating biofilms. It possesses strong antimicrobial efficacy against bacterial intrusions in mammals without inducing drug resistance. There is currently extensive research on the use of NO gas therapy in infectious diseases, including a number of clinical trials, suggesting that it is a therapeutic approach with high potential for clinical translation. As more NO donors are identified and drug delivery systems continue to improve, NO gas therapy will become more widely used in the treatment of diseases including DR-TB.
With the introduction of novel antimicrobial agents and the extensive utilization of repurposed drugs to boost new treatment regimens, remarkable advancements have been made in reducing the toxicity related to DR-TB treatment, shortening the treatment duration, and enhancing DR-TB treatment results.174 In view of the great anti-infective potential of NO, we chose NO gas therapy for anti-TB treatment. As a potential NO donor, biocompatible L-Arg can spontaneously produce NO, which is catalyzed by NOS. This compensates for some of the defects of traditional NO donors, and the presence of iNOS in macrophages, which plays a major role in anti-TB treatment, makes us more firmly choose arginine as the NO donor for anti-TB treatment. Of course, we cannot ignore that arginine, as a nutrient, will be taken up by Mtb to support their growth. Therefore, we need to design a targeted delivery system to deliver arginine to the cytoplasm of macrophages rather than to the phagolysosomes, so as to ensure that arginine is catalyzed by iNOS to produce NO as much as possible. It is possible that DOPE, a lipid component, can be added to the LNPs, which can promote the endosomal escape of the LNPs from co-localization with lysosomes176,177 and thus away from most of the intracellular Mtb. The controlled release of drugs and stability of circulation still need to be ensured using drug delivery systems. With the development of nanotechnology, nanomaterial-based drug delivery has become a popular approach to maximize the therapeutic potential of drugs. Among them, LNPs can improve the biocompatibility of materials compared to nanoparticles alone, have the ability to slow drug release, and can be loaded with both hydrophilic and hydrophobic drugs, which are important for the delivery of NO donors in anti-TB treatment. Therefore, we believe that LNPs are expected to be introduced into NO gas treatment of anti-TB through the simultaneous delivery of drugs, such as arginine and RIF, which may provide a novel strategy for the treatment of TB and even solve the problem of DR-TB. In addition, after determining that NO gas therapy has great potential for anti-TB treatment, we can not only increase the concentration of NO in macrophages by exogenous supplementation of NO donors but also stimulate macrophages or genetically modify macrophages to improve their ability to produce NO. In a study by Sheng et al, it was reported that in macrophages overexpressing zinc-finger DHHC domain palmitoyl-transferase 2 (ZDHHC2), the expression of iNOS and the concentration of NO were increased.178 In addition, it has been reported that the loss of the IL-4Rα signaling pathway in B cells increases the production of NO in macrophages by regulating the inflammatory response of macrophages.179 In summary, it is not difficult to understand that NO, an inflammatory molecule in macrophages, may increase the concentration of NO when the expression of inflammatory cytokines in treated macrophages increases. Therefore, when the expression of inflammatory cytokines in macrophages increases, it is necessary to examine changes in the expression of iNOS and NO, which may provide a new strategy for NO anti-TB treatment. NO gas therapy is a promising anti-TB strategy; however, it is a complex process. This is not only due to the concentration dependence and cytotoxicity of the physiological action of NO but also involves the complex regulatory mechanisms of the immune system, including the regulation of macrophages. Moreover, Mtb is a bacterium that is susceptible to immune evasion. Therefore, in this multidisciplinary research context, the combination of nanotechnology, immunology, and microbiology will facilitate the application of NO gas therapy in the treatment of TB. The exploration of these future research directions is expected to provide a more solid theoretical basis and practical guidance for NO gas therapy in the treatment of TB, including DR-TB, and consequently, may have a positive impact on improving the treatment effect of TB.
Summary
The broad biological role of NO makes NO-based therapies attractive for treating various diseases, especially for infectious diseases. In the context of TB, in infected Mtb macrophages, NO produced via the iNOS pathway enhances the bactericidal activity of macrophages and directly kills Mtb through various signaling pathways. Additionally, the ability of NO to enhance the efficacy of antibiotics against resistant strains by eliminating bacterial biofilms further emphasizes its therapeutic potential. However, the short half-life, instability, and toxicity of NO present significant challenges for their clinical application, thereby limiting their direct use. This has driven research on NO donors, which despite their promise, suffer from instability and harmful by-products. Arginine, a precursor for endogenous NO production, offers a promising alternative as a stable, non-toxic NO donor. To enhance their therapeutic potential, drug delivery systems, especially those using nanotechnology, can improve their stability and controlled release. In conclusion, NO-based therapies, particularly those that leverage nanotechnology, hold significant promise in improving TB treatment outcomes, warranting further research and experimentation.
Consent for Publication
All authors in the paper agree to be published.
Acknowledgments
We thank Biorender and 3ds Max for their assistance in creating all the figures.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work. Xiaoying Jin, Jiajun Wang, and Yongdui Ruan share the first authorship.
Funding
This work was supported by National Natural Science Foundation of China (82272348, 82404964, 82270013, 82300016), Guangdong Basic and Applied Basic Research Foundation (2023A1515030195, 2023A1515110502, 2023A1515140072), High Talent Project of Guangdong Province (2021QN02Y720), Key Project of Universities in Guangdong Province (2022ZDZX2021), Innovation Team Project of Universities in Guangdong Province (2022KCXTD010), Science and Technology Project of Dongguan (20231800940512 and 20211800905542), Discipline Construction Project of Guangdong Medical University (4SG24010G), Project of Songshan Lake Innovation Center of Medicine & Engineering of Guangdong Medical University (4SG22317P), Medical-Industrial Integration Project of Songshan Lake (4SG22309P), Construction Project of Nanotechnology and Application Engineering Research Center of Guangdong Medical University (4SG24179G) and Doctoral Initial Funding of Guangdong Medical University (4SG23258G, 4SG23229G).
Disclosure
The authors declare no competing interests.
References
1. MacMicking J, wen XQ, Nathan C. Nitric oxide and macrophage function. Annu Rev Immunol. 1997;15(1):323–350. doi:10.1146/annurev.immunol.15.1.323
2. Andrabi SM, Sharma NS, Karan A, et al. Nitric oxide: physiological functions, delivery, and biomedical applications. Adv Sci. 2023;10(30):2303259. doi:10.1002/advs.202303259
3. Zhang S, Liu Y, Zhou X, et al. Sirtuin 7 regulates nitric oxide production and apoptosis to promote mycobacterial clearance in macrophages. Front Immunol. 2021:12. doi:10.3389/fimmu.2021.779235.
4. Mikulska-Ruminska K, Anthonymuthu T, Levkina A, et al. NO● represses the oxygenation of arachidonoyl PE by 15LOX/PEBP1: mechanism and role in ferroptosis. IJMS. 2021;22(10):5253. doi:10.3390/ijms22105253
5. Samanovic MI, Tu S, Novák O, et al. Proteasomal control of cytokinin synthesis protects mycobacterium tuberculosis against nitric oxide. Mol Cell. 2015;57(6):984–994. doi:10.1016/j.molcel.2015.01.024
6. Anand K, Tripathi A, Shukla K, et al. Mycobacterium tuberculosis SufR responds to nitric oxide via its 4Fe–4S cluster and regulates Fe–S cluster biogenesis for persistence in mice. Redox Biol. 2021;46:102062. doi:10.1016/j.redox.2021.102062
7. Wang TY, Zhu XY, Wu FG. Antibacterial gas therapy: strategies, advances, and prospects. Bioactive Mat. 2023;23:129–155. doi:10.1016/j.bioactmat.2022.10.008
8. Palmer RMJ, Ferrige AG, Moncada S. Nitric oxide release accounts for the biological activity of endothelium-derived relaxing factor. Nature. 1987;327(6122):524–526. doi:10.1038/327524a0
9. Culotta E, Koshland DE. NO news is good news: a startlingly simple molecule unites neuroscience, physiology, and immunology and revises scientists’ understanding of how cells communicate and defend themselves. Science. 1992;258(5090):1862–1865. doi:10.1126/science.1361684
10. Kim JO, Noh JK, Thapa RK, et al. Nitric oxide-releasing chitosan film for enhanced antibacterial and in vivo wound-healing efficacy. Int J Biol Macromol. 2015;79:217–225. doi:10.1016/j.ijbiomac.2015.04.073
11. Luo Z, Zhou Y, Yang T, Gao Y, Kumar P, Chandrawati R. Ceria nanoparticles as an unexpected catalyst to generate nitric oxide from S ‐nitrosoglutathione. Small. 2022;18(11):2105762. doi:10.1002/smll.202105762
12. O’Dell TJ, Hawkins RD, Kandel ER, Arancio O. Tests of the roles of two diffusible substances in long-term potentiation: evidence for nitric oxide as a possible early retrograde messenger. Proc Natl Acad Sci USA. 1991;88(24):11285–11289. doi:10.1073/pnas.88.24.11285
13. Kibbe M. Inducible nitric oxide synthase and vascular injury. Cardiovasc Res. 1999;43(3):650–657. doi:10.1016/S0008-6363(99)00130-3
14. Vidanapathirana AK, Psaltis PJ, Bursill CA, Abell AD, Nicholls SJ. Cardiovascular bioimaging of nitric oxide: Achievements, challenges, and the future. Med Res Rev. 2021;41(1):435–463. doi:10.1002/med.21736
15. Picón-Pagès P, Garcia-Buendia J, Muñoz FJ. Functions and dysfunctions of nitric oxide in brain. Biochimica Et Biophysica Acta. 2019;1865(8):1949–1967. doi:10.1016/j.bbadis.2018.11.007
16. Brown GC. Nitric oxide and neuronal death. Nitric Oxide. 2010;23(3):153–165. doi:10.1016/j.niox.2010.06.001
17. Zhou X, Meng Z, She J, et al. Near-infrared light-responsive nitric oxide delivery platform for enhanced radioimmunotherapy. Nano-Micro Lett. 2020;12(1):100. doi:10.1007/s40820-020-00431-3
18. Bogdan C. Nitric oxide and the immune response. Nat Immunol. 2001;2(10):907–916. doi:10.1038/ni1001-907
19. Luo Y, Zhu Y, Basang W, Wang X, Li C, Zhou X. Roles of nitric oxide in the regulation of reproduction: a review. Front Endocrinol. 2021;12:752410. doi:10.3389/fendo.2021.752410
20. Wang J, He Q, Yan X, Cai Y, Chen J. Effect of exogenous nitric oxide on sperm motility in vitro. Biol Res. 2014;47(1):44. doi:10.1186/0717-6287-47-44
21. Fei Y, Wu J, An HW, et al. Identification of new nitric oxide-donating peptides with dual biofilm eradication and antibacterial activities for intervention of device-related infections. J Med Chem. 2020;63(17):9127–9135. doi:10.1021/acs.jmedchem.9b01832
22. Yuan Z, Lin C, He Y, et al. Near-infrared light-triggered nitric-oxide-enhanced photodynamic therapy and low-temperature photothermal therapy for biofilm elimination. ACS Nano. 2020;14(3):3546–3562. doi:10.1021/acsnano.9b09871
23. Gao Q, Zhang X, Yin W, et al. Functionalized MoS2 nanovehicle with near‐infrared laser‐mediated nitric oxide release and photothermal activities for advanced bacteria‐infected wound therapy. Small. 2018;14(45):1802290. doi:10.1002/smll.201802290
24. Yang HZ, Chen JJ, Zhang L, et al. A dual responsive nitric oxide / β-lapachone co-delivery platform for redox imbalance-enhanced tumor therapy. Eur J Pharmaceut Biopharmaceut. 2024;201:114348. doi:10.1016/j.ejpb.2024.114348
25. Lundberg JO, Gladwin MT, Weitzberg E. Strategies to increase nitric oxide signalling in cardiovascular disease. Nat Rev Drug Discov. 2015;14(9):623–641. doi:10.1038/nrd4623
26. Lundberg JO, Weitzberg E. Nitric oxide signaling in health and disease. Cell. 2022;185(16):2853–2878. doi:10.1016/j.cell.2022.06.010
27. Villanueva C, Giulivi C. Subcellular and cellular locations of nitric oxide synthase isoforms as determinants of health and disease. Free Rad Biol Med. 2010;49(3):307–316. doi:10.1016/j.freeradbiomed.2010.04.004
28. Pautz A, Art J, Hahn S, Nowag S, Voss C, Kleinert H. Regulation of the expression of inducible nitric oxide synthase. Nitric Oxide. 2010;23(2):75–93. doi:10.1016/j.niox.2010.04.007
29. Barraud N, Kelso M, Rice S, Kjelleberg S. Nitric oxide: a key mediator of biofilm dispersal with applications in infectious diseases. CPD. 2014;21(1):31–42. doi:10.2174/1381612820666140905112822
30. Olthof BMJ, Gartside SE, Rees A. Puncta of neuronal nitric oxide synthase (nNOS) mediate NMDA receptor signaling in the auditory midbrain. J Neurosci. 2019;39(5):876–887. doi:10.1523/JNEUROSCI.1918-18.2018
31. Villalta SA, Nguyen HX, Deng B, Gotoh T, Tidball JG. Shifts in macrophage phenotypes and macrophage competition for arginine metabolism affect the severity of muscle pathology in muscular dystrophy. Human Mol Genet. 2008;18(3):482–496. doi:10.1093/hmg/ddn376
32. Sweet MJ, Ramnath D, Singhal A, Kapetanovic R. Inducible antibacterial responses in macrophages. Nat Rev Immunol. 2025;25(2):92–107. doi:10.1038/s41577-024-01080-y
33. Wang Z, Jin A, Yang Z, Huang W. Advanced nitric oxide generating nanomedicine for therapeutic applications. ACS Nano. 2023;17(10):8935–8965. doi:10.1021/acsnano.3c02303
34. Mandal SM. Nitric oxide mediated hypoxia dynamics in COVID-19. Nitric Oxide. 2023;133:18–21. doi:10.1016/j.niox.2023.02.002
35. Rajendran R, Chathambath A, Al-Sehemi AG, et al. Critical role of nitric oxide in impeding COVID-19 transmission and prevention: a promising possibility. Environ Sci Pollut Res. 2022;29(26):38657–38672. doi:10.1007/s11356-022-19148-4
36. Fang W, Jiang J, Su L, et al. The role of NO in COVID-19 and potential therapeutic strategies. Free Rad Biol Med. 2021;163:153–162. doi:10.1016/j.freeradbiomed.2020.12.008
37. Di Fenza R, Shetty NS, Gianni S, et al. High-dose inhaled nitric oxide in acute hypoxemic respiratory failure due to COVID-19: a multicenter Phase II trial. Am J Respir Crit Care Med. 2023;208(12):1293–1304. doi:10.1164/rccm.202304-0637OC
38. Tandon M, Wu W, Moore K, et al. SARS-CoV-2 accelerated clearance using a novel nitric oxide nasal spray (NONS) treatment: a randomized trial. Lancet Regional Health. 2022;3:100036. doi:10.1016/j.lansea.2022.100036
39. Zhang N, Deng J, Wu F, Lu X, Huang L, Zhao M. Expression of arginase I and inducible nitric oxide synthase in the peripheral blood and lymph nodes of HIV-positive patients. Mol Med Rep. 2016;13(1):731–743. doi:10.3892/mmr.2015.4601
40. Sovershaeva E, Kranzer K, Mchugh G, et al. History of tuberculosis is associated with lower exhaled nitric oxide levels in HIV-infected children. AIDS. 2019;33(11):1711–1718. doi:10.1097/QAD.0000000000002265
41. Yue CJ, Kai ZJ, Pan W. Immunometabolism: towards a better understanding the mechanism of parasitic infection and immunity. Front Immunol. 2021;12:661241. doi:10.3389/fimmu.2021.661241
42. Holzmuller P, Geiger A, Nzoumbou-Boko R, et al. Trypanosomatid infections: how do parasites and their excreted–secreted factors modulate the inducible metabolism of l-arginine in macrophages? Front Immunol. 2018;9:778. doi:10.3389/fimmu.2018.00778
43. Pandya S, Verma RK, Khare P, et al. Supplementation of host response by targeting nitric oxide to the macrophage cytosol is efficacious in the hamster model of visceral leishmaniasis and adds to efficacy of amphotericin B. Int J Parasitol. 2016;6(2):125–132. doi:10.1016/j.ijpddr.2016.01.001
44. Yu YL, Wu JJ, Lin CC, et al. Elimination of methicillin-resistant Staphylococcus aureus biofilms on titanium implants via photothermally-triggered nitric oxide and immunotherapy for enhanced osseointegration. Military Med Res. 2023;10(1):21. doi:10.1186/s40779-023-00454-y
45. Hossain S, Nisbett LM, Boon EM. Discovery of two bacterial nitric oxide-responsive proteins and their roles in bacterial biofilm regulation. Acc Chem Res. 2017;50(7):1633–1639. doi:10.1021/acs.accounts.7b00095
46. Mishra BB, Rathinam VAK, Martens GW, et al. Nitric oxide controls the immunopathology of tuberculosis by inhibiting NLRP3 inflammasome–dependent processing of IL-1β. Nat Immunol. 2013;14(1):52–60. doi:10.1038/ni.2474
47. Chan ED, Chan J, Schluger NW. What is the role of nitric oxide in murine and human host defense against tuberculosis?: current knowledge. Am J Respir Cell Mol Biol. 2001;25(5):606–612. doi:10.1165/ajrcmb.25.5.4487
48. Mishra BB, Lovewell RR, Olive AJ, et al. Nitric oxide prevents a pathogen-permissive granulocytic inflammation during tuberculosis. Nat Microbiol. 2017;2(7):17072. doi:10.1038/nmicrobiol.2017.72
49. Jamaati H, Mortaz E, Pajouhi Z, et al. Nitric oxide in the pathogenesis and treatment of tuberculosis. Front Microbiol. 2017;8:2008. doi:10.3389/fmicb.2017.02008
50. Namouchi A, Gómez-Muñoz M, Frye SA, et al. The Mycobacterium tuberculosis transcriptional landscape under genotoxic stress. BMC Genomics. 2016;17(1):791. doi:10.1186/s12864-016-3132-1
51. Weikert LF, Lopez JP, Abdolrasulnia R, Chroneos ZC, Shepherd VL. Surfactant protein A enhances mycobacterial killing by rat macrophages through a nitric oxide-dependent pathway. Ame J Physiol Lung Cell Mol Physiol. 2000;279(2):L216–L223. doi:10.1152/ajplung.2000.279.2.L216
52. Voskuil MI, Bartek IL, Visconti K, Schoolnik GK. The response of mycobacterium tuberculosis to reactive oxygen and nitrogen species. Front Microbio. 2011;2. doi:10.3389/fmicb.2011.00105
53. Qian H, Ye Z, Pi L, Ao J. Roles and current applications of S-nitrosoglutathione in anti-infective biomaterials. Mate Today Bio. 2022;16:100419. doi:10.1016/j.mtbio.2022.100419
54. Tarekegn BG, Tientcheu LD, Decker J, et al. Host and pathogen factors that influence variability of Mycobacterium tuberculosis lipid body content in sputum from patients with tuberculosis: an observational study. Lancet Microbe. 2024;5(9):100885. doi:10.1016/S2666-5247(24)00108-3
55. History of tuberculosis is associated with lower exhaled nitric oxide levels in HIV-infected children - PubMed. Available from: https://pubmed.ncbi.nlm.nih.gov/31107249/.
56. Bhat KH, Srivastava S, Kotturu SK, Ghosh S, Mukhopadhyay S. The PPE2 protein of Mycobacterium tuberculosis translocates to host nucleus and inhibits nitric oxide production. Sci Rep. 2017;7(1):39706. doi:10.1038/srep39706
57. Shi H, Xiong CF, Zhang LJ, et al. Light-triggered nitric oxide nanogenerator with high l-arginine loading for synergistic photodynamic/gas/photothermal therapy. Adv Healthc Mater. 2023;12(20):e2300012. doi:10.1002/adhm.202300012
58. Near-infrared light-triggered nitric-oxide-enhanced photodynamic therapy and low-temperature photothermal therapy for biofilm elimination - PubMed. Available from: https://pubmed.ncbi.nlm.nih.gov/32069025/.
59. Malone‐Povolny MJ, Maloney SE, Schoenfisch MH. Nitric oxide therapy for diabetic wound healing. Adv Healthcare Mat. 2019;8(12):1801210. doi:10.1002/adhm.201801210
60. Peng J, Xie S, Huang K, et al. Nitric oxide-propelled nanomotors for bacterial biofilm elimination and endotoxin removal to treat infected burn wounds. J Mater Chem B. 2022;10(22):4189–4202. doi:10.1039/D2TB00555G
61. Tu C, Lu H, Zhou T, et al. Promoting the healing of infected diabetic wound by an anti-bacterial and nano-enzyme-containing hydrogel with inflammation-suppressing, ROS-scavenging, oxygen and nitric oxide-generating properties. Biomaterials. 2022;286:121597. doi:10.1016/j.biomaterials.2022.121597
62. Flume PA, Garcia BA, Wilson D, Steed L, Dorman SE, Winthrop K. Inhaled nitric oxide for adults with pulmonary non-tuberculous mycobacterial infection. Resp Med. 2023;206:107069. doi:10.1016/j.rmed.2022.107069
63. Fitzsimmons LF, Clark TR, Hackstadt T. Nitric Oxide Inhibition of Rickettsia rickettsii. Infect Immun. 2021;89(12):e00371–21. doi:10.1128/IAI.00371-21
64. Li G, Lv K, Pan X, et al. Dynamic nitric oxide/drug codelivery system based on polyrotaxane architecture for effective treatment of Candida albicans infection. Acta Biomaterialia. 2023;155:618–634. doi:10.1016/j.actbio.2022.11.006
65. Bangirana P, Conroy AL, Opoka RO, et al. Inhaled nitric oxide and cognition in pediatric severe malaria: a randomized double-blind placebo controlled trial. PLoS One. 2018;13(1):e0191550. doi:10.1371/journal.pone.0191550
66. Safaee Fakhr B, Wiegand SB, Pinciroli R, et al. High concentrations of nitric oxide inhalation therapy in pregnant patients with severe coronavirus disease 2019 (COVID-19). Obstetrics Gynecol. 2020;136(6):1109–1113. doi:10.1097/AOG.0000000000004128
67. Wu W, Gaucher C, Diab R, et al. Time lasting S-nitrosoglutathione polymeric nanoparticles delay cellular protein S-nitrosation. Eur J Pharmaceut Biopharmaceut. 2015;89:1–8. doi:10.1016/j.ejpb.2014.11.005
68. Cheng J, He K, Shen Z, Zhang G, Yu Y, Hu J. Nitric oxide (NO)-releasing macromolecules: rational design and biomedical applications. Front Chem. 2019;7:530. doi:10.3389/fchem.2019.00530
69. Li B, Ming Y, Liu Y, et al. recent developments in pharmacological effect, mechanism and application prospect of diazeniumdiolates. Front Pharmacol. 2020;11:923. doi:10.3389/fphar.2020.00923
70. Sadrearhami Z, Nguyen TK, Namivandi-Zangeneh R, Jung K, Wong EHH, Boyer C. Recent advances in nitric oxide delivery for antimicrobial applications using polymer-based systems. J Mater Chem B. 2018;6(19):2945–2959. doi:10.1039/C8TB00299A
71. Wang PG, Xian M, Tang X, et al. Nitric oxide donors: chemical activities and biological applications. Chem Rev. 2002;102(4):1091–1134. doi:10.1021/cr000040l
72. Bhowmik R, Roy M. Recent advances on the development of NO-releasing molecules (NORMs) for biomedical applications. Eur J Med Chem. 2024;268:116217. doi:10.1016/j.ejmech.2024.116217
73. Huerta S, Chilka S, Bonavida B. Nitric oxide donors: novel cancer therapeutics. Int J Oncol. 2008;33(5):909–927.
74. Thatcher GRJ, Nicolescu AC, Bennett BM, Toader V. Nitrates and no release: contemporary aspects in biological and medicinal chemistry. Free Rad Biol Med. 2004;37(8):1122–1143. doi:10.1016/j.freeradbiomed.2004.06.013
75. Huang Z, Fu J, Zhang Y. Nitric oxide donor-based cancer therapy: advances and prospects. J Med Chem. 2017;60(18):7617–7635. doi:10.1021/acs.jmedchem.6b01672
76. Daiber A, Münzel T. Organic nitrate therapy, nitrate tolerance, and nitrate-induced endothelial dysfunction: emphasis on redox biology and oxidative stress. Antioxidants Redox Signal. 2015;23(11):899–942. doi:10.1089/ars.2015.6376
77. Di Bella D, Ferreira JPS, Rdno S, et al. Gold nanoparticles reduce inflammation in cerebral microvessels of mice with sepsis. J Nanobiotechnol. 2021;19(1):52. doi:10.1186/s12951-021-00796-6
78. Cauwels A, Brouckaert P. Nitrite regulation of shock. Cardiovasc Res. 2011;89(3):553–559. doi:10.1093/cvr/cvq317
79. Gilchrist M, Shore AC, Benjamin N. Inorganic nitrate and nitrite and control of blood pressure. Cardiovasc Res. 2011;89(3):492–498. doi:10.1093/cvr/cvq309
80. Gilchrist M, Winyard PG, Benjamin N. Dietary nitrate – good or bad? Nitric Oxide. 2010;22(2):104–109. doi:10.1016/j.niox.2009.10.005
81. Bates JN, Baker MT, Guerra R, Harrison DG. Nitric oxide generation from nitroprusside by vascular tissue. Evidence that reduction of the nitroprusside anion and cyanide loss are required. Biochem Pharmacol. 1991;42 Suppl:S157–165. doi:10.1016/0006-2952(91)90406-u
82. Udeh CI, Ting M, Arango M, Mick S. Delayed presentation of nitroprusside-induced cyanide toxicity. Annals Thoracic Surg. 2015;99(4):1432–1434. doi:10.1016/j.athoracsur.2014.05.097
83. Viribay A, Burgos J, Fernández-Landa J, Seco-Calvo J, Mielgo-Ayuso J. Effects of arginine supplementation on athletic performance based on energy metabolism: a systematic review and meta-analysis. Nutrients. 2020;12(5):1300. doi:10.3390/nu12051300
84. Zhang K, Mishra A, Jagannath C. New insight into arginine and tryptophan metabolism in macrophage activation during tuberculosis. Front Immunol. 2024;15:1363938. doi:10.3389/fimmu.2024.1363938
85. Ferro A. S-nitrosothiols as nitric oxide-donors: chemistry, biology and possible future therapeutic applications. CMC. 2004;11(20):2679–2690. doi:10.2174/0929867043364397
86. Hogg N. The biochemistry and physiology of S-nitrosothiols. Annu Rev Pharmacol Toxicol. 2002;42(1):585–600. doi:10.1146/annurev.pharmtox.42.092501.104328
87. Belder AJD, MacAllister R, Radomski MW, Moncada S, Vallance PJT. Effects of S-nitroso-glutathione in the human forearm circulation: evidence for selective inhibition of platelet activation. Cardiovasc Res. 1994;28(5):691–694. doi:10.1093/cvr/28.5.691
88. Rosier BT, Johnston W, Carda-Diéguez M, et al. Nitrate reduction capacity of the oral microbiota is impaired in periodontitis: potential implications for systemic nitric oxide availability. Int J Oral Sci. 2024;16(1):1. doi:10.1038/s41368-023-00266-9
89. Jensen FB, Hansen MN. Differential uptake and metabolism of nitrite in normoxic and hypoxic goldfish. Aquatic Toxicol. 2011;101(2):318–325. doi:10.1016/j.aquatox.2010.11.007
90. Lehnert N, Kim E, Dong HT, et al. The biologically relevant coordination chemistry of iron and nitric oxide: electronic structure and reactivity. Chem Rev. 2021;121(24):14682–14905. doi:10.1021/acs.chemrev.1c00253
91. Olabe J. Coordination chemistry of nitric oxide and biological signaling. Scirevfew. 2020;2(1):64–99. doi:10.52712/sciencereviews.v2i1.33
92. Kurhaluk N. The effectiveness of L-arginine in clinical conditions associated with hypoxia. IJMS. 2023;24(9):8205. doi:10.3390/ijms24098205
93. Hishikawa K, Nakaki T, Tsuda M, et al. Effect of systemic L-arginine administration on hemodynamics and nitric oxide release in man. Jpn Heart J. 1992;33(1):41–48. doi:10.1536/ihj.33.41
94. Rath M, Mã¼ller I, Kropf P, Closs EI, Munder M. Metabolism via arginase or nitric oxide synthase: two competing arginine pathways in macrophages. Front Immunol. 2014;5. doi:10.3389/fimmu.2014.00532
95. Stuehr DJ. Structure-function aspects in the nitric oxide synthases. Annu Rev Pharmacol Toxicol. 1997;37:339–359. doi:10.1146/annurev.pharmtox.37.1.339
96. Izzo R, Trimarco V, Mone P, et al. Combining L-Arginine with vitamin C improves long-COVID symptoms: the Lincoln survey. Pharmacol Res. 2022;183:106360. doi:10.1016/j.phrs.2022.106360
97. Galván-Peña S, Oâ€TMNeill LAJ. Metabolic reprograming in macrophage polarization. Front Immunol. 2014;5. doi:10.3389/fimmu.2014.00420
98. Fraix A, Sortino S. Photoactivable platforms for nitric oxide delivery with fluorescence imaging. Chem Asian J. 2015;10(5):1116–1125. doi:10.1002/asia.201403398
99. Bonavida B, Baritaki S, Huerta-Yepez S, Vega MI, Chatterjee D, Yeung K. Novel therapeutic applications of nitric oxide donors in cancer: roles in chemo- and immunosensitization to apoptosis and inhibition of metastases. Nitric Oxide. 2008;19(2):152–157. doi:10.1016/j.niox.2008.04.018
100. Oliveira C, Benfeito S, Fernandes C, Cagide F, Silva T, Borges F. NO and HNO donors, nitrones, and nitroxides: past, present, and future. Med Res Rev. 2018;38(4):1159–1187. doi:10.1002/med.21461
101. Choi HW, Kim J, Kim J, et al. Light-induced acid generation on a gatekeeper for smart nitric oxide delivery. ACS Nano. 2016;10(4):4199–4208. doi:10.1021/acsnano.5b07483
102. Tan L, Huang R, Li X, Liu S, Shen YM. Controllable release of nitric oxide and doxorubicin from engineered nanospheres for synergistic tumor therapy. Acta Biomaterialia. 2017;57:498–510. doi:10.1016/j.actbio.2017.05.019
103. Wilczewska AZ, Niemirowicz K, Markiewicz KH, Car H. Nanoparticles as drug delivery systems. Pharmacol Rep. 2012;64(5):1020–1037. doi:10.1016/s1734-1140(12)70901-5
104. Xu C, Zhang YS, Begin S, Thanh NTK. Introduction to advanced functional nanomaterials for biomedical applications. Nanoscale. 2022;14(20):7441–7443. doi:10.1039/D2NR90077G
105. Vauthier C, Bouchemal K. Methods for the preparation and manufacture of polymeric nanoparticles. Pharm Res. 2009;26(5):1025–1058. doi:10.1007/s11095-008-9800-3
106. Martins JP, Torrieri G, Santos HA. The importance of microfluidics for the preparation of nanoparticles as advanced drug delivery systems. Expert Opinion Drug Delivery. 2018;15(5):469–479. doi:10.1080/17425247.2018.1446936
107. Wang W, Sheng H, Cao D, et al. S-nitrosoglutathione functionalized polydopamine nanoparticles incorporated into chitosan/gelatin hydrogel films with NIR-controlled photothermal/NO-releasing therapy for enhanced wound healing. Int J Biol Macromol. 2022;200:77–86. doi:10.1016/j.ijbiomac.2021.12.125
108. Du Z, Zhang X, Guo Z, et al. X‐ray‐controlled generation of peroxynitrite based on nanosized LiLuF4:Ce3+ scintillators and their applications for radiosensitization. Adv Mat. 2018;30(43):1804046. doi:10.1002/adma.201804046
109. Aman S, Kaur N, Mittal D, et al. Novel biocompatible green silver nanoparticles efficiently eliminates multidrug resistant nosocomial pathogens and mycobacterium species. Indian J Microbiol. 2023;63(1):73–83. doi:10.1007/s12088-023-01061-0
110. Lin W, Fan S, Liao K, et al. Engineering zinc oxide hybrid selenium nanoparticles for synergetic anti-tuberculosis treatment by combining Mycobacterium tuberculosis killings and host cell immunological inhibition. Front Cell Infect Microbiol. 2023;12:1074533. doi:10.3389/fcimb.2022.1074533
111. Zhang W, Jiang Y, He Y, et al. Lipid carriers for mRNA delivery. Acta Pharmaceutica Sinica B. 2023;13(10):4105–4126. doi:10.1016/j.apsb.2022.11.026
112. Silva R, Ferreira H, Cavaco-Paulo A. Sonoproduction of liposomes and protein particles as templates for delivery purposes. Biomacromolecules. 2011;12(10):3353–3368. doi:10.1021/bm200658b
113. Sunderland CJ, Steiert M, Talmadge JE, Derfus AM, Barry SE. Targeted nanoparticles for detecting and treating cancer. Drug Develop Res. 2006;67(1):70–93. doi:10.1002/ddr.20069
114. Rideau E, Dimova R, Schwille P, Wurm FR, Landfester K. Liposomes and polymersomes: a comparative review towards cell mimicking. Chem Soc Rev. 2018;47(23):8572–8610. doi:10.1039/C8CS00162F
115. Zhao Y, Hou X, Chai J, et al. Stapled liposomes enhance cross-priming of radio-immunotherapy. Adv Mater. 2022;34(3):e2107161. doi:10.1002/adma.202107161
116. Wang N, Chen M, Wang T. Liposomes used as a vaccine adjuvant-delivery system: from basics to clinical immunization. J Controlled Release. 2019;303:130–150. doi:10.1016/j.jconrel.2019.04.025
117. Huang Q, Zou Y, Arno MC, et al. Hydrogel scaffolds for differentiation of adipose-derived stem cells. Chem Soc Rev. 2017;46(20):6255–6275. doi:10.1039/C6CS00052E
118. Serra L, Doménech J, Peppas NA. Drug transport mechanisms and release kinetics from molecularly designed poly(acrylic acid-g-ethylene glycol) hydrogels. Biomaterials. 2006;27(31):5440–5451. doi:10.1016/j.biomaterials.2006.06.011
119. Peppas NA, Khare AR. Preparation, structure and diffusional behavior of hydrogels in controlled release. Adv Drug Delivery Rev. 1993;11(1–2):1–35. doi:10.1016/0169-409X(93)90025-Y
120. Yang JA, Yeom J, Hwang BW, Hoffman AS, Hahn SK. In situ-forming injectable hydrogels for regenerative medicine. Progress Polymer Sci. 2014;39(12):1973–1986. doi:10.1016/j.progpolymsci.2014.07.006
121. Cheah E, Bansal M, Nguyen L, et al. Electrically responsive release of proteins from conducting polymer hydrogels. Acta Biomaterialia. 2023;158:87–100. doi:10.1016/j.actbio.2023.01.013
122. Tang Q, Lu B, He J, et al. Exosomes-loaded thermosensitive hydrogels for corneal epithelium and stroma regeneration. Biomaterials. 2022;280:121320. doi:10.1016/j.biomaterials.2021.121320
123. Han Z, Wang P, Mao G, et al. Dual pH-responsive hydrogel actuator for lipophilic drug delivery. ACS Appl Mater Interfaces. 2020;12(10):12010–12017. doi:10.1021/acsami.9b21713
124. Zheng Y, Yang D, Gao B, et al. A DNA-inspired injectable adhesive hydrogel with dual nitric oxide donors to promote angiogenesis for enhanced wound healing. Acta Biomaterialia. 2024;176:128–143. doi:10.1016/j.actbio.2024.01.026
125. Meng Y, Niu X, Li G. Liposome nanoparticles as a novel drug delivery system for therapeutic and diagnostic applications. CDD. 2023;20(1):41–56. doi:10.2174/1567201819666220324093821
126. Weiss G, Schaible UE. Macrophage defense mechanisms against intracellular bacteria. Immunol Rev. 2015;264(1):182–203. doi:10.1111/imr.12266
127. Kapralov AA, Yang Q, Dar HH, et al. Redox lipid reprogramming commands susceptibility of macrophages and microglia to ferroptotic death. Nat Chem Biol. 2020;16(3):278–290. doi:10.1038/s41589-019-0462-8
128. Schneider BE, Korbel D, Hagens K, et al. A role for IL‐18 in protective immunity against Mycobacterium tuberculosis. Eur J Immunol. 2010;40(2):396–405. doi:10.1002/eji.200939583
129. Deretic V, Saitoh T, Akira S. Autophagy in infection, inflammation and immunity. Nat Rev Immunol. 2013;13(10):722–737. doi:10.1038/nri3532
130. Mosser DM. The many faces of macrophage activation. J Leukoc Biol. 2003;73(2):209–212. doi:10.1189/jlb.0602325
131. MacMicking JD, Taylor GA, McKinney JD. Immune Control of Tuberculosis by IFN-γ-Inducible LRG-47. Science. 2003;302(5645):654–659. doi:10.1126/science.1088063
132. Nussler AK, Billiar TR, Liu ZZ, Morris SM. Coinduction of nitric oxide synthase and argininosuccinate synthetase in a murine macrophage cell line. Implications for regulation of nitric oxide production. J Biol Chem. 1994;269(2):1257–1261.
133. Liu Y, Xu R, Gu H, et al. Metabolic reprogramming in macrophage responses. Biomark Res. 2021;9(1):1. doi:10.1186/s40364-020-00251-y
134. Martí ILíndez AA, Reith W. Arginine-dependent immune responses. Cell Mol Life Sci. 2021;78(13):5303–5324. doi:10.1007/s00018-021-03828-4
135. McNeill E, Stylianou E, Crabtree MJ, et al. Regulation of mycobacterial infection by macrophage Gch1 and tetrahydrobiopterin. Nat Commun. 2018;9(1):5409. doi:10.1038/s41467-018-07714-9
136. Rutschmann O, Toniolo C, McKinney JD. Preexisting heterogeneity of inducible nitric oxide synthase expression drives differential growth of mycobacterium tuberculosis in macrophages. mBio. 2022;13(5):e02251–22. doi:10.1128/mbio.02251-22
137. Fei Q, Zhang J, Chen L, et al. Modulating ferroptosis and mycobactericidal activity in lung epithelial cells via YY1/iNOS pathway. Life Sci. 2024;358:123131. doi:10.1016/j.lfs.2024.123131
138. Chen J, Tang F, Li H, et al. Mycobacterium tuberculosis suppresses APLP2 expression to enhance its survival in macrophage. Int Immunopharmacol. 2023;124:111058. doi:10.1016/j.intimp.2023.111058
139. Tur J, Pereira-Lopes S, Vico T, et al. Mitofusin 2 in macrophages links mitochondrial ROS production, cytokine release, phagocytosis, autophagy, and bactericidal activity. Cell Rep. 2020;32(8):108079. doi:10.1016/j.celrep.2020.108079
140. Liebenberg D, Gordhan BG, Kana BD. Drug resistant tuberculosis: implications for transmission, diagnosis, and disease management. Front Cell Infect Microbiol. 2022;12:943545. doi:10.3389/fcimb.2022.943545
141. Barraud N, Schleheck D, Klebensberger J, et al. Nitric oxide signaling in Pseudomonas aeruginosa biofilms mediates phosphodiesterase activity, decreased cyclic Di-GMP levels, and enhanced dispersal. J Bacteriol. 2009;191(23):7333–7342. doi:10.1128/JB.00975-09
142. Choi M, Hasan N, Cao J, Lee J, Hlaing SP, Yoo JW. Chitosan-based nitric oxide-releasing dressing for anti-biofilm and in vivo healing activities in MRSA biofilm-infected wounds. Int J Biol Macromol. 2020;142:680–692. doi:10.1016/j.ijbiomac.2019.10.009
143. Kiran D, Podell BK, Chambers M, Basaraba RJ. Host-directed therapy targeting the Mycobacterium tuberculosis granuloma: a review. Semin Immunopathol. 2016;38(2):167–183. doi:10.1007/s00281-015-0537-x
144. Stockwell BR. Ferroptosis turns 10: emerging mechanisms, physiological functions, and therapeutic applications. Cell. 2022;185(14):2401–2421. doi:10.1016/j.cell.2022.06.003
145. Amaral EP, Costa DL, Namasivayam S, et al. A major role for ferroptosis in Mycobacterium tuberculosis –induced cell death and tissue necrosis. J Exp Med. 2019;216(3):556–570. doi:10.1084/jem.20181776
146. O’Donnell VB, Taylor KB, Parthasarathy S, et al. 15-lipoxygenase catalytically consumes nitric oxide and impairs activation of guanylate cyclase. J Biol Chem. 1999;274(29):20083–20091. doi:10.1074/jbc.274.29.20083
147. Zea AH, Aiyar A, Tate D. Dual effect of interferon (IFNγ)-induced nitric oxide on tumorigenesis and intracellular bacteria. Vitamins & Hormones. 2014;96:299–321. doi:10.1016/B978-0-12-800254-4.00012-X
148. Goossens SN, Sampson SL, Van Rie A. Mechanisms of drug-induced tolerance in Mycobacterium tuberculosis. Clin Microbiol Rev. 2020;34(1):e00141–20. doi:10.1128/CMR.00141-20
149. Singh V, Chibale K. Strategies to combat multi-drug resistance in tuberculosis. Acc Chem Res. 2021;54(10):2361–2376. doi:10.1021/acs.accounts.0c00878
150. Tacconelli E, Carrara E, Savoldi A, et al. Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect Dis. 2018;18(3):318–327. doi:10.1016/S1473-3099(17)30753-3
151. Eygeris Y, Gupta M, Kim J, Sahay G. Chemistry of lipid nanoparticles for RNA delivery. Acc Chem Res. 2022;55(1):2–12. doi:10.1021/acs.accounts.1c00544
152. Wang Z, Ma W, Fu X, Qi Y, Zhao Y, Zhang S. Development and applications of mRNA treatment based on lipid nanoparticles. Biotechnol Adv. 2023;65:108130. doi:10.1016/j.biotechadv.2023.108130
153. Hald Albertsen C, Kulkarni JA, Witzigmann D, Lind M, Petersson K, Simonsen JB. The role of lipid components in lipid nanoparticles for vaccines and gene therapy. Adv Drug Delivery Rev. 2022;188:114416. doi:10.1016/j.addr.2022.114416
154. Zhang L, Chan JM, Gu FX, et al. Self-assembled lipid−polymer hybrid nanoparticles: a robust drug delivery platform. ACS Nano. 2008;2(8):1696–1702. doi:10.1021/nn800275r
155. Meng X, Zhang Z, Tong J, Sun H, Fawcett JP, Gu J. The biological fate of the polymer nanocarrier material monomethoxy poly(ethylene glycol)-block-poly(d,l-lactic acid) in rat. Acta Pharmaceutica Sinica B. 2021;11(4):1003–1009. doi:10.1016/j.apsb.2021.02.018
156. Barbieri BD, Peeler DJ, Samnuan K, et al. The role of helper lipids in optimising nanoparticle formulations of self-amplifying RNA. J Controlled Release. 2024;374:280–292. doi:10.1016/j.jconrel.2024.08.016
157. Fenaroli F, Repnik U, Xu Y, et al. Enhanced permeability and retention-like extravasation of nanoparticles from the vasculature into tuberculosis granulomas in zebrafish and mouse models. ACS Nano. 2018;12(8):8646–8661. doi:10.1021/acsnano.8b04433
158. Pi J, Chen D, Wang J, et al. Macrophage targeted graphene oxide nanosystem synergize antibiotic killing and host immune defense for Tuberculosis Therapy. Pharmacol Res. 2024;208:107379. doi:10.1016/j.phrs.2024.107379
159. Fan S, Zhao D, Wang J, et al. Photothermal and host immune activated therapy of cutaneous tuberculosis using macrophage targeted mesoporous polydopamine nanoparticles. Mat Today Bio. 2024;28:101232. doi:10.1016/j.mtbio.2024.101232
160. Shen L, Liao K, Yang E, et al. Macrophage targeted iron oxide nanodecoys augment innate immunological and drug killings for more effective Mycobacterium Tuberculosis clearance. J Nanobiotechnol. 2023;21(1):369. doi:10.1186/s12951-023-02103-x
161. Pi J, Shen L, Shen H, et al. Mannosylated graphene oxide as macrophage-targeted delivery system for enhanced intracellular M.tuberculosis killing efficiency. Mat Sci Eng. 2019;103:109777. doi:10.1016/j.msec.2019.109777
162. Jain S, Tran TH, Amiji M. Macrophage repolarization with targeted alginate nanoparticles containing IL-10 plasmid DNA for the treatment of experimental arthritis. Biomaterials. 2015;61:162–177. doi:10.1016/j.biomaterials.2015.05.028
163. Zhou K, Yang C, Shi K, et al. Activated macrophage membrane-coated nanoparticles relieve osteoarthritis-induced synovitis and joint damage. Biomaterials. 2023;295:122036. doi:10.1016/j.biomaterials.2023.122036
164. Liang DS, Wen ZJ, Wang JH, et al. Legumain protease-sheddable PEGylated, tuftsin-modified nanoparticles for selective targeting to tumour-associated macrophages. J Drug Targeting. 2022;30(1):82–93. doi:10.1080/1061186X.2021.1906886
165. Nissen JC, Selwood DL, Tsirka SE. Tuftsin signals through its receptor neuropilin‐1 via the transforming growth factor beta pathway. J Neurochem. 2013;127(3):394–402. doi:10.1111/jnc.12404
166. Xu H, Niu M, Yuan X, Wu K, Liu A. CD44 as a tumor biomarker and therapeutic target. Exp Hematol Oncol. 2020;9(1):36. doi:10.1186/s40164-020-00192-0
167. Abri Aghdam M, Bagheri R, Mosafer J, et al. Recent advances on thermosensitive and pH-sensitive liposomes employed in controlled release. J Control Release. 2019;315:1–22. doi:10.1016/j.jconrel.2019.09.018
168. Kim KY. Nanotechnology platforms and physiological challenges for cancer therapeutics. Nanomedicine. 2007;3(2):103–110. doi:10.1016/j.nano.2006.12.002
169. Litzinger DC, Huang L. Phosphatidylethanolamine liposomes: drug delivery, gene transfer and immunodiagnostic applications. Biochim Biophys Acta. 1992;1113(2):201–227. doi:10.1016/0304-4157(92)90039-d
170. Kanamala M, Palmer BD, Ghandehari H, Wilson WR, Wu Z. PEG-benzaldehyde-hydrazone-lipid based PEG-sheddable pH-sensitive liposomes: abilities for endosomal escape and long circulation. Pharm Res. 2018;35(8):154. doi:10.1007/s11095-018-2429-y
171. Hosoya H, Dobroff AS, Driessen WHP, et al. Integrated nanotechnology platform for tumor-targeted multimodal imaging and therapeutic cargo release. Proc Natl Acad Sci USA. 2016;113(7):1877–1882. doi:10.1073/pnas.1525796113
172. Peng Y, Feng W, Huang H, Chen Y, Yang S. Macrophage-targeting Antisenescence nanomedicine enables in-Situ NO induction for Gaseous and antioxidative atherosclerosis intervention. Bioactive Mat. 2025;48:294–312. doi:10.1016/j.bioactmat.2025.02.025
173. The Lancet Respiratory Medicine null. Ending tuberculosis: ways forward. Lancet Respir Med. 2024;12(4):255. doi:10.1016/S2213-2600(24)00081-X.
174. Dookie N, Ngema SL, Perumal R, Naicker N, Padayatchi N, Naidoo K. The changing paradigm of drug-resistant tuberculosis treatment: successes, pitfalls, and future perspectives. Clin Microbiol Rev. 2022;35(4):e0018019. doi:10.1128/cmr.00180-19
175. Opoku‐Damoah Y, Zhang R, Ta HT, Xu ZP. Therapeutic gas‐releasing nanomedicines with controlled release: advances and perspectives. Exploration. 2022;2(5):20210181. doi:10.1002/EXP.20210181
176. Bae Y, Jung MK, Song SJ, et al. Functional nanosome for enhanced mitochondria-targeted gene delivery and expression. Mitochondrion. 2017;37:27–40. doi:10.1016/j.mito.2017.06.005
177. Nakamura T, Kuroi M, Harashima H. Influence of endosomal escape and degradation of α-galactosylceramide loaded liposomes on CD1d antigen presentation. Mol Pharmaceutics. 2015;12(8):2791–2799. doi:10.1021/mp500704e
178. Sheng J, You X, Nie D, et al. ZDHHC2 promoted antimycobacterial responses by selective autophagic degradation of B-RAF and C-RAF in macrophages. Sci Adv. 2025;11(4):eadq7706. doi:10.1126/sciadv.adq7706
179. Parihar SP, Ozturk M, Höft MA, et al. IL-4-responsive B cells are detrimental during chronic tuberculosis infection in mice. Front Immunol. 2021;12:611673. doi:10.3389/fimmu.2021.611673
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.