Back to Journals » Clinical, Cosmetic and Investigational Dermatology » Volume 19
Neuroimmune Genetic Overlap Between Chronic Pruritic Skin Diseases and Major Depressive Disorder: Insights from Cross-Trait GWAS Analysis
Authors Deng C, Ding Y, Li X, Qiu G, Li Z ![]() , Song Y
, Song Y
Received 12 April 2026
Accepted for publication 13 June 2026
Published 9 July 2026 Volume 2026:19 616258
DOI https://doi.org/10.2147/CCID.S616258
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Michela Starace
ChenWei Deng,1,2 YiFan Ding,2 Xiaojian Li,2 GuiRong Qiu,2,3 Zhiquan Li,1,2 Yu Song1,4
1Department of Dermatology, Longhua Hospital Affiliated to Shanghai University of Traditional Chinese Medicine, Shanghai, People’s Republic of China; 2Clinical Medical College, Jiangxi University of Chinese Medicine, Nanchang, People’s Republic of China; 3Department of Dermatology, Affiliated Hospital of Jiangxi University of Chinese Medicine, Nanchang, People’s Republic of China; 4Department of Dermatology, Longhua Hospital Jiangxi Branch Affiliated to Shanghai University of Traditional Chinese Medicine, Nanchang, People’s Republic of China
Correspondence: Yu Song, Department of Dermatology, Longhua Hospital Affiliated to Shanghai University of Traditional Chinese Medicine, Shanghai, People’s Republic of China, Email [email protected]
Background: Psoriasis (PSO), atopic dermatitis (AD), and urticaria (URT) are chronic inflammatory skin diseases marked by severe pruritus. These conditions are frequently comorbid with psychiatric disorders, especially major depressive disorder (MDD). Despite this clinical overlap, the underlying shared genetic mechanisms driving these comorbidities remain largely unclear.
Methods: We integrated large-scale European-ancestry genome-wide association study (GWAS) summary statistics for PSO, AD, URT, and MDD. Genetic overlap was assessed using linkage disequilibrium score regression (LDSC), high-definition likelihood (HDL), MAGMA-based gene and pathway analyses, PLACO cross-trait pleiotropy analysis, Bayesian colocalization, and summary-data-based Mendelian randomization (SMR). Tissue-specific enrichment and gene expression analyses were performed using GTEx transcriptomic resources.
Results: All three skin diseases showed significant positive genetic correlations with MDD. We identified 70 genome-wide significant pleiotropic SNPs and 13 shared genomic risk loci across the pairwise IAD-MDD analyses, with 11q13.1 and 11q12.2 prominent in AD-MDD. Prioritized genes, including FADS1, SLC22A4, SLC22A5, and FOXP3, showed enrichment in immune-related tissues and brain regions. Functional analyses implicated transcriptional regulation, synaptic signaling, and IgE isotype switching, supporting shared immunological and neurobiological pathways.
Conclusion: This study provides genetic evidence consistent with shared susceptibility between pruritic inflammatory skin diseases and MDD. The findings generate hypotheses about immune-neural pathways underlying skin-psychiatric comorbidity, but they should be interpreted in light of key limitations, including European-only summary statistics, summary-level inference, possible sample overlap, and the absence of an independent replication cohort.
Keywords: psoriasis, atopic dermatitis, major depressive disorder, pleiotropy, GWAS
Introduction
Itch-associated dermatological diseases (IADs), including psoriasis (PSO), atopic dermatitis (AD), and urticaria (URT), are chronic inflammatory skin disorders characterized by intense pruritus. These conditions affect hundreds of millions of individuals worldwide and significantly impair both physical and mental health.1–3 PSO and AD typically have an early onset in adolescence or early adulthood and are characterized by chronic, recurrent courses and poor treatment adherence. URT, on the other hand, often presents with sudden and intense itching, and can also persist chronically, thereby severely compromising quality of life.1–3The FinnGen URT phenotype used in this study captures urticaria as a clinically heterogeneous diagnosis rather than itch severity itself; therefore, we use the IAD framing to denote pruritus-associated inflammatory skin diseases rather than a uniform clinical syndrome.
Extensive clinical observations have shown that patients with IADs frequently experience psychiatric symptoms such as mood disturbances and sleep disorders, particularly major depressive disorder (MDD), with comorbidity rates significantly higher than in the general population.4,5 Traditionally, such comorbidities have been interpreted through the “psychological consequence model”, which posits that chronic inflammation, skin lesions, and social stigma lead to secondary psychological distress. However, recent studies have increasingly supported a “shared etiology model”, suggesting that skin disorders and psychiatric conditions may share overlapping immune, neurological, and metabolic pathways, potentially rooted in common genetic susceptibility.6,7
For instance, pro-inflammatory cytokines such as IL-17 and TNF-α have been shown to activate the NF-κB signaling pathway in both PSO and MDD, contributing to inflammation and neuroregulatory disturbances.8,9 Moreover, emerging evidence from genetic studies has indicated crosstalk between the immune system and the central nervous system in the context of psychiatric disorders. A Mendelian randomization study published in 2022 demonstrated a significant causal effect of AD on MDD.10
These findings point to a potentially shared genetic basis between IADs and MDD. However, a systematic investigation of the genetic architecture underlying this comorbidity is still lacking. In this study, we integrated large-scale genome-wide association study (GWAS) summary statistics from European populations for PSO, AD, URT, and MDD. We employed a comprehensive suite of genetic methods, including linkage disequilibrium score regression (LDSC),11 high-definition likelihood (HDL),12 multi-marker gene-set analysis via MAGMA,13 and cross-trait pleiotropy analysis using the PLACO method.14 Additionally, we incorporated tissue-specific expression data from the GTEx database to explore the biological relevance of shared genes across systemic tissues.15 This study aims to elucidate the genetic mechanisms linking IADs and MDD and to identify potential targets for integrated treatment strategies.
Methods
GWAS Datasets
GWAS summary statistics for PSO, AD, URT, and MDD were obtained from large-scale meta-analyses and publicly available datasets. Data for MDD were derived from the study by Howard et al,16 which included 170,756 cases and 329,443 controls of European ancestry. GWAS summary statistics for the three IADs were retrieved from the FinnGen project. For PSO, data included 12,760 cases and 482,181 controls (GWAS ID: finngen_R12_L12_PSORIASIS); for AD, 26,905 cases and 394,476 controls (GWAS ID: finngen_R11_L12_ATOPIC); and for URT, 12,576 cases and 438,050 controls (GWAS ID: finngen_R11_L12_URTICARIA). All studies adhered to uniform quality control procedures and employed logistic regression models to assess the associations between diseases and SNP genotypes, with principal components included as covariates. All four discovery datasets were restricted to participants of European ancestry. Public download sources and accession identifiers are provided in the Data availability statement.
Quality Control
To ensure the accuracy, directional consistency, and cross-dataset comparability of the GWAS summary statistics, we applied a unified quality-control and harmonization workflow to all datasets. First, we retained only autosomal biallelic SNPs and removed variants with duplicated rsIDs or missing key fields, including chromosome, base-pair position, effect/non-effect allele, beta or log(OR), standard error, P value, and effect allele frequency or minor allele frequency. Second, low-frequency variants with MAF < 0.01 and strand-ambiguous palindromic SNPs (A/T or C/G with allele frequencies close to 0.5) were excluded. Effect alleles were harmonized across FinnGen R11/R12 and the MDD dataset using the effect allele as the reference. When allele directions were reversed, beta coefficients or log(OR) values were strand-flipped to ensure consistent effect directions. Given the high polymorphism, complex linkage disequilibrium structure, and dense immune-related gene content of the MHC region on chromosome 6 (25–35 Mb), which may cause local signal inflation and false-positive associations, SNPs within this region were excluded from the primary analyses. For each trait pair, only the intersecting SNPs that passed quality control, allele harmonization, and reference-panel matching were retained for downstream analyses.
Genome-Wide Genetic Correlation Analysis
To investigate the shared genetic architecture between IADs and MDD, we applied linkage disequilibrium score regression (LDSC).11 In the LDSC analysis, LD scores were constructed using the 1000 Genomes Project Phase 3 European reference panel (GRCh37, baseline-LD version 2.2), and the analysis was restricted to common HapMap3 SNPs.17 Standard errors (SE) were estimated using the block jackknife approach, and LDSC intercepts were used to evaluate potential sample overlap. No substantial overlap was detected among the trait pairs, supporting the robustness of our findings.
To validate the LDSC results, we also employed the high-definition likelihood (HDL) method, which provides more precise estimates of genetic correlation by efficiently leveraging GWAS summary data. HDL reduces variance by approximately 60% compared to LDSC, enhancing accuracy and robustness.12
Gene-Level Exploration
To investigate the shared mechanisms underlying the identified loci, we performed gene-based and gene-set analyses using MAGMA v1.10, and conducted SNP annotation, gene mapping, and risk locus definition through the SNP2GENE module of the FUMA online platform. In the MAGMA gene-based analysis, linkage disequilibrium among genetic markers was accounted for using the 1000 Genomes Project Phase 3 European reference panel. The Bonferroni-corrected significance threshold for gene-based analysis was set at P < 2.73 × 10−6 (0.05/18,345).13 Functional enrichment analysis included gene sets from the c2.all, c5.bp, c5.cc, and c5.mf collections of the MSigDB database.18 Given that a total of 510,003 gene sets were included, the Bonferroni-corrected threshold (P < 0.05/510,003 ≈ 1 × 10−7) was relatively conservative. Therefore, the main text primarily reports pathway results with FDR q < 0.05, while Bonferroni-corrected results were used as a more stringent supplementary criterion.
Pathway enrichment analysis was further performed using the Metascape web tool (metascape.org) to characterize the biological functions of the mapped genes based on MSigDB annotations.19 Tissue-specific enrichment analysis was conducted using gene expression data from 54 tissues in GTEx Analysis Release v8, and tissue expression enrichment results were obtained through FUMA/MAGMA gene-property analysis. In this analysis, the tissue-enrichment t statistic was calculated as the regression coefficient of the tissue expression covariate divided by its standard error (β/SE) in the MAGMA regression model, and the sign of the t value was used to indicate the direction of expression enrichment.
SNP-Level Pleiotropy and Risk Locus Confirmation
We applied the PLACO to systematically identify SNP-level genetic associations shared between multiple IADs and MDD. This method is designed to detect genetic pleiotropy and identify shared genetic variants across phenotypes.20 The genome-wide significance threshold was set at P_PLACO < 5 × 10−8.
Subsequently, PLACO-significant SNPs were functionally annotated and used for risk locus definition through the FUMA platform. Independent significant SNPs were selected using a threshold of P < 5 × 10−8, followed by LD clumping based on the 1000 Genomes Project Phase 3 European reference panel. In FUMA, independent significant SNPs were defined using r2 < 0.6, lead SNPs were defined using r2 < 0.1, and adjacent genomic risk loci were merged if they were located within 250 kb of each other.
Bayesian colocalization analysis was performed using the coloc R package (v5.2.3). For each lead SNP, a ±500 kb genomic window was used as the analysis region. The default prior probabilities were set as p1 = 1 × 10−4, p2 = 1 × 10−4, and p12 = 1 × 10−5. A posterior probability for hypothesis 4 greater than 0.7 (PP.H4 > 0.7) was considered evidence supporting colocalization, indicating that the two traits may share the same causal variant.21
Potential Drug Target Identification in European Populations
We performed summary-data-based Mendelian randomization (SMR) analysis using the 1000 Genomes Project Phase 3 reference panel to integrate GWAS summary statistics with expression quantitative trait locus (eQTL) data.22 We used GTEx v8 eQTL data as the primary reference and selected significant cis-eQTLs as instrumental variables for SMR analysis. The HEIDI test was used to evaluate whether the gene expression and GWAS signals were more consistent with a shared causal variant rather than LD confounding. A P_HEIDI > 0.05 was interpreted as no evidence of significant heterogeneity, although this does not fully exclude the effects of complex LD or multiple causal variants. Because P_SMR < 0.01 represents an exploratory screening threshold, these results are referred to as “eQTL-mediated candidate genes” or “candidate genes for further functional validation”, rather than confirmed causal genes or established therapeutic targets.23,24 Genes passing both SMR and HEIDI tests were considered putative mediators of genetic effects in IAD–MDD comorbidity (Figure 1).
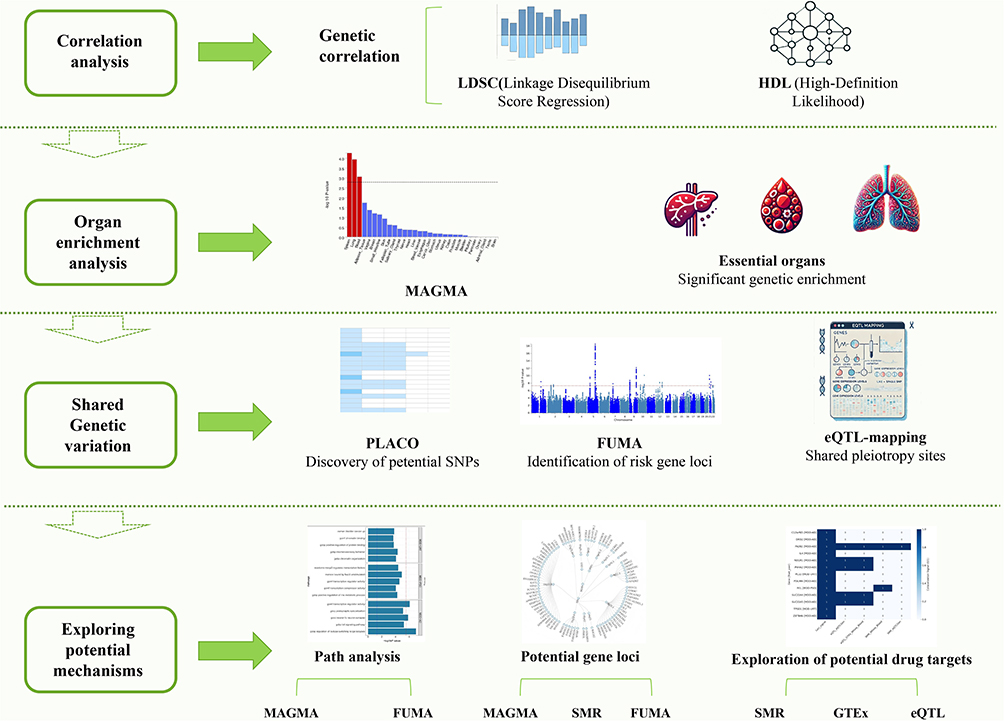 |
Figure 1 Research Flowchart. |
Sample Overlap, MHC Exclusion, and Validation Limitations
The Howard MDD dataset was derived from a meta-analysis of European-ancestry GWAS datasets, including the Psychiatric Genomics Consortium and UK Biobank, whereas the PSO, AD, and URT datasets were obtained from the FinnGen project. Therefore, limited sample overlap between different data sources may theoretically exist and cannot be completely excluded. Because this study was based exclusively on publicly available summary-level data, individual-level cohort identifiers were not accessible, and the exact degree of participant overlap among FinnGen, UK Biobank, and other European GWAS datasets could not be directly quantified.
To indirectly assess potential sample overlap and cross-trait systematic confounding, we used the cross-trait LDSC intercepts and reported the corresponding intercept values in the Results section. The major histocompatibility complex (MHC) region was excluded from the main analyses because of its highly complex linkage disequilibrium structure. In addition, because the publicly available datasets were predominantly derived from individuals of European ancestry and individual-level data were unavailable for leave-one-cohort-out analyses, multi-ancestry validation and leave-one-cohort validation could not be performed. These issues are further acknowledged as study limitations in the Discussion section.
Results
Shared Genetic Architecture Between IADs and MDD
We first evaluated the genetic correlations between PSO, AD, URT, and MDD using LDSC and HDL. The results from both methods were highly consistent (Table 1). Specifically, LDSC revealed significant positive genetic correlations between MDD and each IAD phenotype, including PSO–MDD (r_g = 0.1609, P = 1.55 × 10−5), AD–MDD (r_g = 0.1363, P = 4.94 × 10−5), and URT–MDD (r_g = 0.2434, P = 8.57 × 10−8). Similar positive associations were observed using HDL, with genetic correlations of 0.2289 for PSO–MDD (P = 2.24 × 10−7), 0.1602 for AD–MDD (P = 1.16 × 10−5), and 0.3485 for URT–MDD (P = 2.17 × 10−6). In addition, the cross-trait LDSC intercepts were close to zero for all trait pairs, including PSO–MDD (intercept = 0.0004, SE = 0.0059), AD–MDD (intercept = −0.0037, SE = 0.0057), and URT–MDD (intercept = 0.0006, SE = 0.0049), providing no evidence of substantial sample overlap or cross-trait systematic confounding. These findings suggest a shared genetic architecture between IADs and MDD.
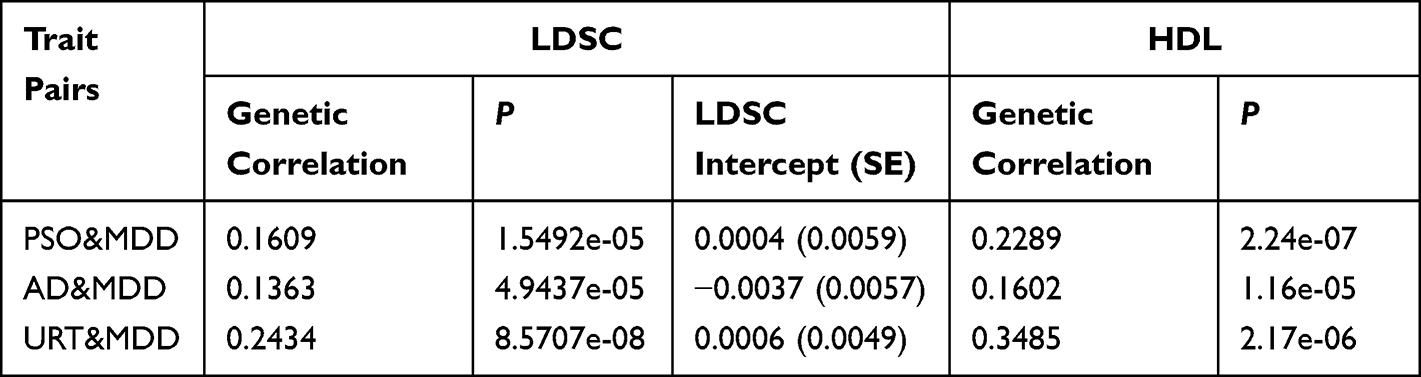 |
Table 1 Genetic Correlation Results for Trait Pairs Based on LDSC and HDL |
Gene-Level Enrichment Analysis via MAGMA
Using the FUMA platform with MAGMA for gene-based analysis, we identified 3727 nominally significant genes (P < 0.05). To avoid overinterpretation of exploratory findings, we focused on the 137 genes that passed the FDR threshold of q < 0.05 as the core gene set for the main interpretation and subsequent functional enrichment analyses (Figure 2 and Table S1–S2 in Supplementary Material).
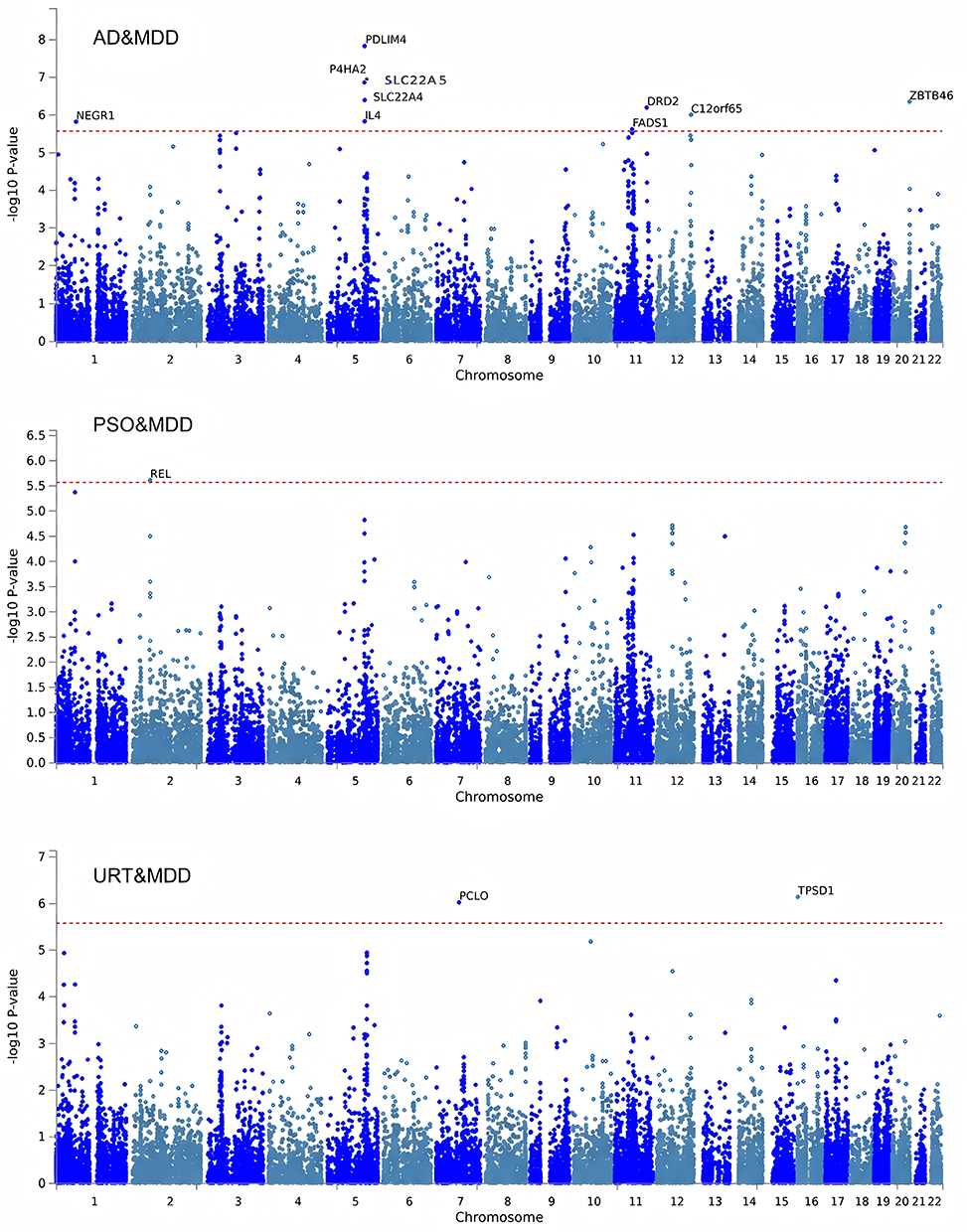 |
Figure 2 Results of Gene Enrichment. |
Further functional annotation revealed that these genes were involved in several key biological pathways, including transcription regulator activity, GOCC: neuron-to-neuron synapse, and GOBP: regulation of isotype switching to IgE isotypes (Figure 3 and Table S3 in Supplementary Material).
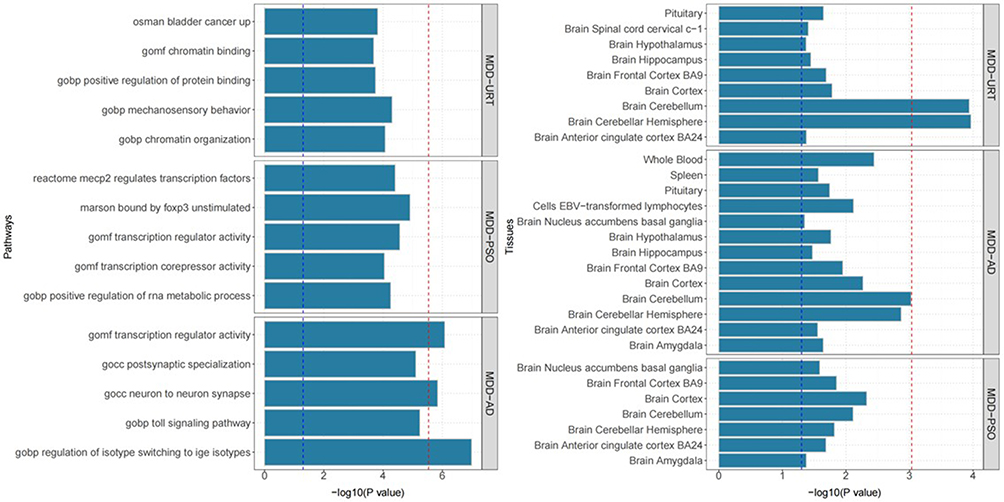 |
Figure 3 MAGMA gene set and tissue specificity of genome-wide pleiotropy results; Shared genes between pruritic skin diseases and MDD are involved in key immune and neural pathways (left) and are highly expressed in both immune tissues and brain regions (right). |
Tissue-specific enrichment analysis demonstrated that the genetic signals shared between MDD and the three IADs were predominantly enriched in central nervous system tissues, such as the cerebral cortex, cerebellar hemisphere, and prefrontal cortex. Notably, genetic signals shared between MDD and AD also showed enrichment in peripheral immune organs, including whole blood and spleen.
SNP-Level Pleiotropy and Risk Locus Analysis
Given the significant shared genetic correlations between IADs and MDD identified through LDSC and HDL, we employed PLACO, a cross-trait pleiotropy detection method, to identify potential pleiotropic loci. A total of 70 novel SNPs (P < 5 × 10−8) exhibiting pleiotropic associations across PSO, AD, and MDD were identified (Table S4 in Supplementary Material).
Based on the PLACO results, we further mapped 13 pleiotropic genomic risk loci using the FUMA platform (Figure 4 and Table S5 in Supplementary Material).
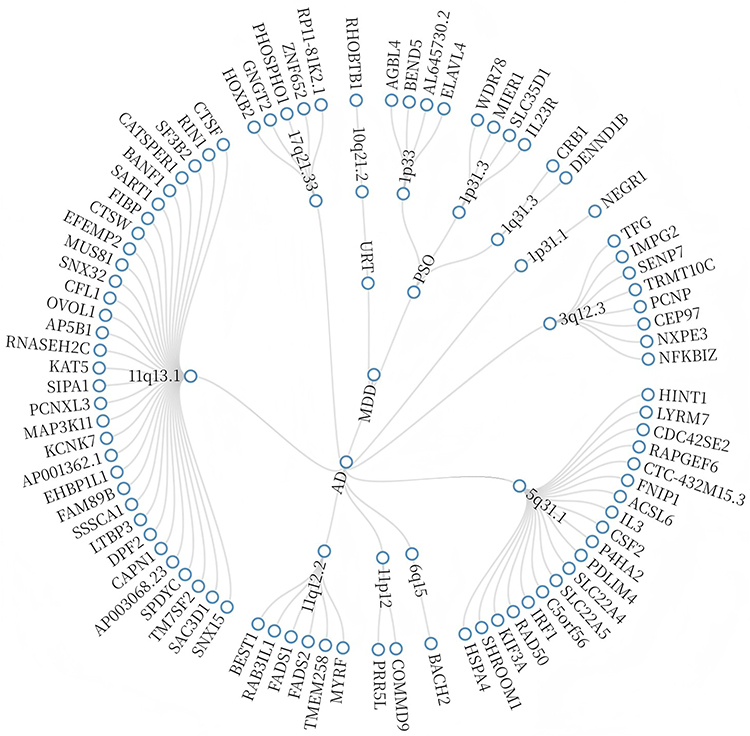 |
Figure 4 Shared Polytropic Regions Between Different Combinations. |
Bayesian colocalization analysis identified only three loci, represented by rs12130046, rs2488389, and rs9825208, with PP.H4 > 0.7. These results provide supportive evidence that the corresponding trait pairs may share causal variants at these loci (Figure 5 and Table S6 in Supplementary Material). Specifically: 10q21.2 was identified as the only shared locus between MDD and URT, 1p33 and 1p31.3 were shared between MDD and PSO, The largest number of shared loci was observed between AD and MDD, including 11q13.1 and 11q12.2 (Table S7 in Supplementary Material).
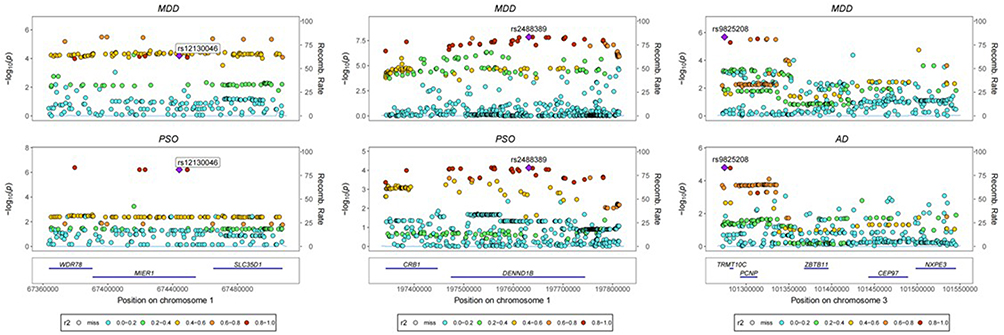 |
Figure 5 Colocalization Analysis Results. |
Potential Drug Targets in European Populations
Using the summary-data-based Mendelian randomization (SMR) approach, we identified 1411 putative candidate drug targets under complex genetic regulation (pSMR < 0.01, pHEIDI > 0.05) (Table S8 in Supplementary Material).
By integrating PLACO-identified pleiotropic signals with the results from FUMA, MAGMA, and SMR analyses, we further prioritized a subset of functionally relevant genes with multi-tissue specificity (Table S9 in Supplementary Material). Notably, several shared genes between AD and MDD—such as FADS1, SLC22A4, and SLC22A5—exhibited significant genetic signals across multiple tissues, highlighting their potential as cross-disease therapeutic targets (Figure 6).
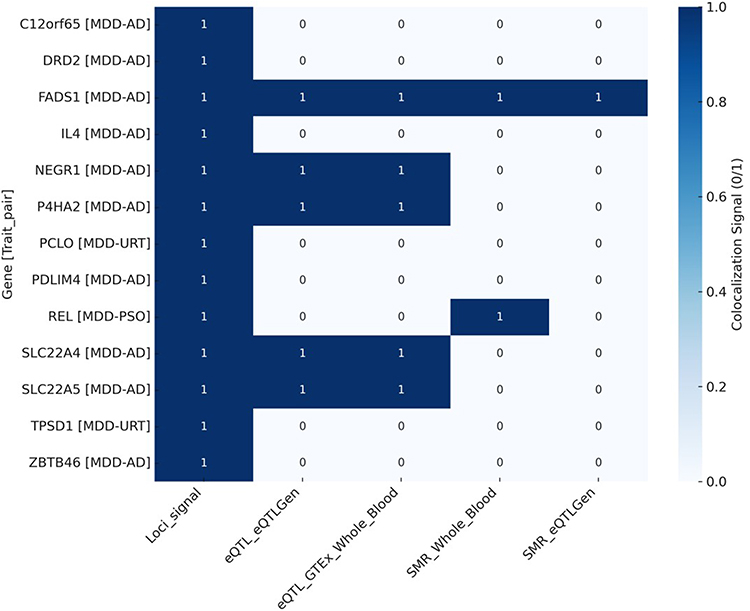 |
Figure 6 Candidate drug targets in the European population; The heatmap shows which genes have significant colocalization signals (1 = significant, 0 = not significant) across different methods. Genes like FADS1, NEGR1, and SLC22A4 are significant in blood-related datasets, suggesting they may be involved in both depression and skin disease. |
Discussion
Clinically, patients with IADs frequently present with varying degrees of emotional disturbances, depression, and sleep disorders, which not only complicate disease control but also substantially diminish quality of life and social functioning. Traditionally, these psychiatric manifestations have been viewed as secondary psychological responses to chronic pruritus, persistent inflammation, or disfigurement. However, the genetic evidence presented in this study suggests that psychiatric symptoms in IADs may not be purely reactive, but instead reflect a systemic pathological phenotype underpinned by a shared genetic basis.
At the genome-wide level, we observed significant genetic correlations between PSO, AD, URT, and MDD using both LDSC and HDL methods. The strongest correlation was observed between URT and MDD (HDL rg = 0.3485, P = 2.17×10−6), suggesting a strong genetic coupling between chronic pruritus and mood regulatory pathways. These findings align with previous epidemiological observations of high depression comorbidity and poor sleep quality among patients with dermatological disorders and underscore the need for a genetic investigation into the IAD–MDD comorbidity.
Tissue-specific enrichment analysis revealed that the shared genetic signals between IADs and MDD are primarily enriched in peripheral immune-related tissues (eg, whole blood, spleen) and central nervous system regions (eg, prefrontal cortex, cerebellum, hippocampus), suggesting potential crosstalk via the “peripheral–central immune–brain axis”. These findings support clinical observations of IAD–MDD comorbidity and provide empirical tissue-level evidence.
In the context of AD–MDD comorbidity, the enrichment in spleen and whole blood was particularly notable, indicating a central mediating role of the peripheral immune system. Numerous studies have confirmed elevated levels of classical proinflammatory cytokines in the peripheral blood of AD patients, including TNF-α, IL-6, and IL-1β, which not only sustain cutaneous inflammation but also affect central nervous system function through multiple pathways.25,26 These cytokines can cross the blood–brain barrier (BBB), activate microglia, and trigger neuroinflammatory cascades, thereby disrupting neurotransmitter metabolism, axon development, synaptic plasticity, and neural network stability—mechanisms strongly implicated in depression and sleep disturbances.27,28 These processes are well aligned with established MDD pathophysiology, particularly in the context of chronic inflammation. The spleen, as a core organ of the peripheral immune system, has recently been recognized as a key regulator of immune–nervous system interactions. In addition to cytokine synthesis and immune cell maturation and trafficking, it is anatomically and functionally connected to the central nervous system via the splenic nerve.29 Evidence from animal stress models supports the biological plausibility of a spleen-brain axis,30,31 but these models should be viewed as mechanistic context rather than direct genetic validation of the human GWAS findings.
Further gene set enrichment analyses revealed widespread and significant pathway-level commonalities between IADs and MDD. Notably, pathways such as transcription regulator activity (GOMF_TRANSCRIPTION_REGULATOR_ACTIVITY), neuron-to-neuron synaptic structure (GOCC_NEURON_TO_NEURON_SYNAPSE), and IgE isotype class switching (GOBP_REGULATION_OF_ISOTYPE_SWITCHING_TO_IGE_ISOTYPES) were highly enriched. The transcription regulator pathway was significantly enriched in both MDD and AD/PSO, implicating shared transcription factors such as FOXP3, STAT6, and CREB1. The gene set MARSON_BOUND_BY_FOXP3_UNSTIMULATED also showed enrichment in both MDD and PSO.
FOXP3, the master transcription factor of regulatory T cells (Tregs), has been causally implicated in anxiety- and depression-like behaviors. In DEREG mice lacking FOXP3⁺ Tregs, anxiety- and depression-like behaviors are accompanied by NLRP3 inflammasome activation in the brain, while restoration of FOXP3⁺ cells reverses these phenotypes and reduces hippocampal IL‑1β expression.32 Human studies also support a link between FOXP3⁺ Tregs and MDD. One study reported significantly increased CD25⁺FOXP3⁺ Treg subpopulations and elevated chemokine receptor expression (eg, CCR6, CCR2) in MDD patients, suggesting abnormal migration and activity.33 Another study found peripheral expansion of FOXP3⁺ Tregs in patients with severe AD, with levels positively correlating with disease severity.34 Therefore, our interpretation is not that FOXP3 changes occur in one uniform direction across disorders, but that dysregulated immune tolerance and Treg-related transcriptional programs may contribute to shared vulnerability.
The enrichment of the GOCC_NEURON_TO_NEURON_SYNAPSE pathway reflects the potential involvement of central synaptic structure and signal transmission in the comorbidity mechanism. Itch perception is not confined to the periphery but involves ascending synaptic pathways projecting to emotion-related brain regions. GRPR⁺ spinal neurons transmit glutamatergic signals to the paraventricular nucleus–amygdala circuit; optogenetic inhibition of this pathway reduces itch perception but increases pain sensitivity.35 Sun & Chen et al previously identified GRPR⁺ dorsal horn neurons as itch-specific without affecting pain transmission, confirming the existence of a distinct itch pathway.36 Moreover, descending dopaminergic pathways enhance spinal GRPR signaling via DRD1 activation and glutamate/GRP release. Interrupting this pathway has been proposed as a therapeutic approach for chronic itch.37
Enrichment of IgE isotype switching pathways suggests that allergic immunity may contribute to shared biology between cutaneous inflammation and psychiatric symptoms. Clinical studies of anti-IgE therapy in chronic spontaneous urticaria have reported improvements in skin symptoms and psychological scores.38,39 These treatment observations are clinically relevant, but they do not prove that the pleiotropic genetic signals identified here act through the same therapeutic pathway.
Pleiotropy analysis using PLACO identified 70 genome-wide significant loci (P_PLACO < 5 x 10^-8) across pairwise analyses of PSO, AD and MDD. Notably, 10q21.2 (PCLO) was the locus highlighted in the MDD-URT analysis. PCLO encodes the presynaptic active zone protein Piccolo and has been implicated in synaptic plasticity and stress-related phenotypes.40 SNP rs2522833 in PCLO has been associated with reduced gray matter volume in the left temporal pole of first-episode, drug-naïve MDD patients carrying the C allele.41
In the MDD–AD shared loci, 11q13.1 and 11q12.2 were particularly notable for their enrichment in immune-related genes (eg, FADS1, IL4) and association with skin barrier function and inflammatory response, suggesting a central regulatory hub connecting skin inflammation and mood regulation. Through integrative filtering via FUMA, MAGMA, and SMR, we identified several candidate genes for downstream functional validation. Among these, FADS1, SLC22A4, and SLC22A5 showed significant cross-tissue genetic signals.
FADS1 encodes delta-5 fatty acid desaturase and participates in omega-3 and omega-6 polyunsaturated fatty acid metabolism. Altered fatty acid metabolism may influence epidermal lipid composition, inflammatory mediator production, and neural membrane biology. However, prior work has reported no consistent association between FADS1–FADS2 cluster SNPs and major depression or suicidal behavior. Accordingly, FADS1 should be viewed as a biologically plausible and genetically prioritized candidate in the present cross-trait framework, rather than a confirmed depression-risk gene.42–44
SLC22A4 (OCTN1) and SLC22A5 (OCTN2), as organic cation transporters for carnitine and choline, are important regulators of immune cell metabolism, cellular energy supply, and antioxidant responses. Their expression in AD keratinocytes correlates with IL‑4 and TNF‑α levels, implicating them in local immune regulation and skin inflammation.45 These genes are also expressed in brain tissues and have been associated with glial activity and emotional disorders, suggesting they may serve as mechanistic links in the AD–MDD comorbidity.
Study Strengths and Limitations
In this study, we applied five complementary genetic methods (LDSC, HDL, MAGMA, PLACO, SMR) to systematically assess the pleiotropic genetic mechanisms underlying PSO, AD, URT, and MDD at genome-wide, SNP, and tissue levels. We identified multiple biologically relevant shared loci and demonstrated significant enrichment in central nervous and peripheral immune tissues, supporting systemic mechanisms underlying skin–mental comorbidity.
Nevertheless, this study has several limitations. First, the GWAS datasets were restricted to European-ancestry participants, limiting generalizability to other populations. Second, the PSO dataset came from FinnGen R12 whereas AD and URT came from FinnGen R11, which may introduce release-specific differences despite harmonization. Third, summary-level analyses cannot fully exclude linkage disequilibrium confounding, sample overlap, or residual heterogeneity across contributing cohorts. Fourth, the exploratory SMR threshold requires caution because candidate genes may not remain significant under stricter genome-wide correction. Finally, these findings overlap conceptually with prior work on AD-MDD and related immune-psychiatric genetics, and independent replication plus experimental validation will be needed before causal mechanisms or treatment implications can be established.
Conclusion
In conclusion, this study used multiple genetic analysis methods to show that pruritic skin diseases—such as PSO, AD, and URT—have significant genetic links with MDD. We identified shared risk genes like FADS1, SLC22A4, SLC22A5, and FOXP3, and found that these genetic signals are mainly active in the immune and nervous systems. This suggests that the immune–neural pathway may play an important role in connecting skin inflammation with depression. These findings offer new genetic evidence for the comorbidity of skin diseases and mental disorders, and may help guide future treatment strategies.
Data Sharing Statement
All GWAS summary statistics analyzed in this study are publicly available. Summary statistics for psoriasis, atopic dermatitis, and urticaria were obtained from the FinnGen project. The corresponding FinnGen GWAS IDs were finngen_R12_L12_PSORIASIS, finngen_R11_L12_ATOPIC, and finngen_R11_L12_URTICARIA. FinnGen summary statistics can be accessed through the FinnGen public browser and release-specific manifest files: R12 manifest, https://storage.googleapis.com/finngen-public-data-r12/summary_stats/finngen_R12_manifest.tsv; R11 manifest, https://storage.googleapis.com/finngen-public-data-r11/summary_stats/finngen_R11_manifest.tsv. Summary statistics for major depressive disorder were obtained from the Howard et al. GWAS dataset deposited in OpenGWAS under accession ieu-b-102: https://opengwas.io/datasets/ieu-b-102.
Ethical Approval
In accordance with Article 32 of the Measures for Ethical Review of Life Science and Medical Research Involving Human Beings issued by the National Science and Technology Ethics Committee of the People’s Republic of China, this study was exempted from ethical review because the data analyzed pose no harm to human participants, do not involve sensitive personal information or commercial interests, and were obtained from open and legally accessible databases.
Acknowledgment
The authors affirm that they have no commercial or financial affiliations that could be perceived as potential conflicts of interest. This includes any relationships or financial interests that could influence or bias the study’s results and conclusions.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
The research received funding from the Shanghai Municipal Science and Technology Commission under the Shanghai 2023 “Science and Technology Innovation Action Plan” (Grant No. 23Y31920304), entitled “Study on the Safety and Skin Flora of Xia’s Dermatological Yangxue Runzao Method in Maintaining the Remission Stage of Moderate-to-Severe Atopic Dermatitis”; and from the Joint Research Institute of Shanghai Pharmaceuticals Holding Co., Ltd. and Shanghai University of Traditional Chinese Medicine (Grant No. E4-H2002103), entitled “Clinical Efficacy of Yunpi Huashi Qingfei Biscuits Based on the Theory of ‘Medicine and Food Homology’ in Preventing Relapse During Remission in Children with Atopic Dermatitis.”
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Li X, Wu Y, Chen S, et al. CD8 + T cells may mediate the effect of gut microbiota on psoriasis: evidence from two-step mendelian randomization and Bayesian weighting. Arch Dermatol Res. 2025;317(1):370. doi:10.1007/s00403-025-03857-0
2. David Boothe W, Tarbox JA, Tarbox MB. Atopic Dermatitis: pathophysiology. Adv Exp Med Biol. 2017;1027:21–13. doi:10.1007/978-3-319-64804-0_3
3. Szymanski K, Schaefer P. Urticaria and Angioedema. Prim Care. 2023;50(2):237–252. doi:10.1016/j.pop.2022.11.003
4. Chen YH, Wang WM, Li IH, Kao HH, Yeh CB, Kao LT. Major depressive disorder increased risk of psoriasis: a propensity score matched cohort study. J Affect Disord. 2021;278:407–412. doi:10.1016/j.jad.2020.09.108
5. Yang T, Huang X, Xu J, et al. Explore the underlying mechanism between atopic dermatitis and major depressive disorder. Front Genet. 2021;12:640951. doi:10.3389/fgene.2021.640951
6. Amanat M, Salehi M, Rezaei N. Neurological and psychiatric disorders in psoriasis. Rev Neurosci. 2018;29(7):805–813. doi:10.1515/revneuro-2017-0108
7. Lewinson RT, Vallerand IA, Lowerison MW, et al. Depression is associated with an increased risk of psoriatic arthritis among patients with psoriasis: a population-based study. J Invest Dermatol. 2017;137(4):828–835. doi:10.1016/j.jid.2016.11.032
8. Liu F, Jia Y, Zhao L, et al. Escin ameliorates CUMS-induced depressive-like behavior via BDNF/TrkB/CREB and TLR4/MyD88/NF-κB signaling pathways in rats. Eur J Pharmacol. 2024;984:177063. doi:10.1016/j.ejphar.2024.177063
9. Pan Y, You Y, Sun L, et al. The STING antagonist H-151 ameliorates psoriasis via suppression of STING/NF-κB-mediated inflammation. Br J Pharmacol. 2021;178(24):4907–4922. doi:10.1111/bph.15673
10. Qi HJ, Li LF. Association of atopic dermatitis with depression and suicide: a two-sample mendelian randomization study. Biomed Res Int. 2022;2022:4084121. doi:10.1155/2022/4084121
11. Bulik-Sullivan B, K FH, Anttila V, et al. An atlas of genetic correlations across human diseases and traits. Nat Genet. 2015;47(11):1236–1241. doi:10.1038/ng.3406
12. Ning Z, Pawitan Y, Shen X. High-definition likelihood inference of genetic correlations across human complex traits. Nat Genet. 2020;52(8):859–864. doi:10.1038/s41588-020-0653-y
13. de Leeuw, Mooij JM, Christiaan A, et al. MAGMA: generalized gene-set analysis of GWAS data. PLOS Comput Biol. 2015;11(4):e1004219. doi:10.1371/journal.pcbi.1004219
14. Lu H, Qiao J, Shao Z, Wang T, Huang S, Zeng P. A comprehensive gene-centric pleiotropic association analysis for 14 psychiatric disorders with GWAS summary statistics. BMC Med. 2021;19(1):314. doi:10.1186/s12916-021-02186-z
15. Consortium G, Thomas J, Salvatore M. The Genotype-Tissue Expression (GTEx) project. Nat Genet. 2013;45(6):580–585. doi:10.1038/ng.2653
16. Howard DM, Adams MJ, Clarke TK, et al. Genome-wide meta-analysis of depression identifies 102 independent variants and highlights the importance of the prefrontal brain regions. Nat Neurosci. 2019;22(3):343–352. doi:10.1038/s41593-018-0326-7
17. Cui G, Li S, Ye H, et al. Gut microbiome and frailty: insight from genetic correlation and mendelian randomization. Gut Microbes. 2023;15(2):2282795. doi:10.1080/19490976.2023.2282795
18. Subramanian A, Tamayo P, K MV, et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc Natl Acad Sci U S A. 2005;102(43):15545–15550. doi:10.1073/pnas.0506580102
19. Zhou Y, Zhou B, Pache L, et al. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat Commun. 2019;10(1):1523. doi:10.1038/s41467-019-09234-6
20. Hu YQ, Jin XJ, Lei SF, et al. Inflammatory bowel disease and osteoporosis: common genetic effects, pleiotropy, and causality. Hum Immunol. 2024;85(5):110856. doi:10.1016/j.humimm.2024.110856
21. Lin J, Zhou J, Xu Y. Potential drug targets for multiple sclerosis identified through Mendelian randomization analysis. Brain. 2023;146(8):3364–3372. doi:10.1093/brain/awad070
22. Wu J, Mei Y, Li X, et al. PRCP is a promising drug target for intracranial aneurysm rupture supported via multi-omics analysis. Stroke Vasc Neurol. 2024;2024:1.
23. Sun X, Chen B, Qi Y, et al. Multi-omics Mendelian randomization integrating GWAS, eQTL and pQTL data revealed GSTM4 as a potential drug target for migraine. J Headache Pain. 2024;25(1):117. doi:10.1186/s10194-024-01828-w
24. X DX, L CD, M CH, et al. DNA methylation biomarkers and myopia: a multi-omics study integrating GWAS, mQTL and eQTL data. Clin Clin Epigenet. 2024;16(1):157. doi:10.1186/s13148-024-01772-1
25. Cesare AD, Meglio PD, Nestle FO. A role for Th17 cells in the immunopathogenesis of atopic dermatitis? J Invest Dermatol. 2008;128(11):2569–2571. doi:10.1038/jid.2008.283
26. Raap U, Wichmann K, Bruder M, et al. Correlation of IL-31 serum levels with severity of atopic dermatitis. J Allergy Clin Immunol. 2008;122(2):421–423. doi:10.1016/j.jaci.2008.05.047
27. Dantzer R, O’Connor JC, Freund GG, Johnson RW, Kelley KW. From inflammation to sickness and depression: when the immune system subjugates the brain. Nat Rev Neurosci. 2008;9(1):46–56. doi:10.1038/nrn2297
28. Miller AH, Raison CL. The role of inflammation in depression: from evolutionary imperative to modern treatment target. Nat Rev Immunol. 2016;16(1):22–34. doi:10.1038/nri.2015.5
29. Ma L, Zhang J, Fujita Y, et al. Effects of spleen nerve denervation on depression-like phenotype, systemic inflammation, and abnormal composition of gut microbiota in mice after administration of lipopolysaccharide: a role of brain-spleen axis. J Affect Disord. 2022;317:156–165. PMID: 36037991. doi:10.1016/j.jad.2022.08.087
30. Powell ND, Sloan EK, Bailey MT, et al. Social stress up-regulates inflammatory gene expression in the leukocyte transcriptome via β-adrenergic induction of myelopoiesis. Proc Natl Acad Sci U S A. 2013;110(41):16574–16579. doi:10.1073/pnas.1310655110
31. Wohleb ES, McKim DB, Shea DT, et al. Re-establishment of anxiety in stress-sensitized mice is caused by monocyte trafficking from the spleen to the brain. Biol Psychiatry. 2014;75(12):970–981. PMID: 24439304; PMCID: PMC4084643. doi:10.1016/j.biopsych.2013.11.029
32. Yang EJ, Rahim MA, Griggs E, Iban-Arias R, Pasinetti GM. Transient anxiety-and depression-like behaviors are linked to the depletion of Foxp3-expressing cells via inflammasome in the brain. PNAS Nexus. 2023;2(8):pgad251. PMID: 37614669; PMCID: PMC10443660. doi:10.1093/pnasnexus/pgad251
33. Alvarez-Mon MA, Gomez-Lahoz AM, Orozco A, et al. Blunted expansion of regulatory T lymphocytes is associated with increased bacterial translocation in patients with major depressive disorder. Front Psychiatry. 2021;11:591962. PMID: 33488424; PMCID: PMC7820111. doi:10.3389/fpsyt.2020.591962
34. Roesner LM, Floess S, Witte T, Olek S, Huehn J, Werfel T. Foxp3(+) regulatory T cells are expanded in severe atopic dermatitis patients. Allergy. 2015;70(12):1656–1660. PMID: 26228301. doi:10.1111/all.12712
35. Sun YG, Zhao ZQ, Meng XL, Yin J, Liu XY, Chen ZF. Cellular basis of itch sensation. Science. 2009;325(5947):1531–1534. doi:10.1126/science.1174868
36. Sun YG, Chen ZF. A gastrin-releasing peptide receptor mediates the itch sensation in the spinal cord. Nature. 2007;448(7154):700–703. doi:10.1038/nature06029
37. Zhang ZJ, Shao HY, Liu C, et al. Descending dopaminergic pathway facilitates itch signal processing via activating spinal GRPR+ neurons. EMBO Rep. 2023;24(10):e56098. doi:10.15252/embr.202256098
38. Can PK, Etikan P, Degirmentepe EN, Kocaturk E. Depression scores change significantly after omalizumab treatment in patients with chronic spontaneous urticaria. Asian Pac J Allergy Immunol. 2024;42(2):132–137. PMID: 33638627. doi:10.12932/AP-180920-0965
39. Tan MG, Bailey AMJ, Dorus B, Kirchhof MG. Clinical impacts of omalizumab on the psychiatric comorbidities of chronic spontaneous urticaria: a systematic review and meta-analysis. J Drugs Dermatol. 2024;23(4):e116–e117. doi:10.36849/JDD.7919
40. Sullivan PF, Agrawal A, Bulik CM, et al. Psychiatric genomics consortium. psychiatric genomics: an update and an agenda. Am J Psychiatry. 2018;175(1):15–27. PMID: 28969442; PMCID: PMC5756100. doi:10.1176/appi.ajp.2017.17030283
41. Igata R, Katsuki A, Kakeda S, et al. PCLO rs2522833-mediated gray matter volume reduction in patients with drug-naive, first-episode major depressive disorder. Transl Psychiatry. 2017;7(5):e1140. doi:10.1038/tp.2017.100
42. Balić A, Vlašić D, Žužul K, Marinović B, Bukvić Mokos Z. Omega-3 versus omega-6 polyunsaturated fatty acids in the prevention and treatment of inflammatory skin diseases. Int J Mol Sci. 2020;21(3):741. PMID: 31979308; PMCID: PMC7037798. doi:10.3390/ijms21030741
43. Lin JY, Ma LJ, Yuan JP, Yu P, Bai BX. Causal effects of fatty acids on atopic dermatitis: a Mendelian randomization study. Front Nutr. 2023;10:1083455. PMID: 36908902; PMCID: PMC9996175. doi:10.3389/fnut.2023.1083455
44. Sublette ME, Vaquero C, Baca-Garcia E, et al. Lack of association of SNPs from the FADS1-FADS2 gene cluster with major depression or suicidal behavior. Psychiatr Genet. 2016;26(2):81–86. PMID: 26513616; PMCID: PMC4764474. doi:10.1097/YPG.0000000000000111
45. Tamai I. Pharmacological and pathophysiological roles of carnitine/organic cation transporters (OCTNs: SLC22A4, SLC22A5 and Slc22a21). Biopharm Drug Dispos. 2013;34(1):29–44. doi:10.1002/bdd.1816
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
The Role of Teledermatology During the COVID-19 Pandemic: A Narrative Review
Ruggiero A, Martora F, Fabbrocini G, Villani A, Marasca C, Megna M, Fornaro L, Comune R, Potestio L
Clinical, Cosmetic and Investigational Dermatology 2022, 15:2785-2793
Published Date: 19 December 2022
Advances in Relationship Between Alcohol Consumption and Skin Diseases
Liu L, Chen J
Clinical, Cosmetic and Investigational Dermatology 2023, 16:3785-3791
Published Date: 29 December 2023
Advancements in Dermatological Applications of Curcumin: Clinical Efficacy and Mechanistic Insights in the Management of Skin Disorders
Mo Z, Yuan J, Guan X, Peng J
Clinical, Cosmetic and Investigational Dermatology 2024, 17:1083-1092
Published Date: 13 May 2024
Defining the Care Pathway in Patients with Psoriasis and Atopic Dermatitis
Masip M, Pagès-Puigdemont N, López-Ferrer A, de Paz HD, Serra-Baldrich E, Puig L, Riera P
Patient Preference and Adherence 2024, 18:2325-2335
Published Date: 16 November 2024
Navigating Mendelian Randomization in Sleep Medicine: Challenges, Opportunities, and Best Practices
BaHammam AS, Jahrami H
Nature and Science of Sleep 2024, 16:1811-1825
Published Date: 22 November 2024