Back to Journals » OncoTargets and Therapy » Volume 12
NEDD4 Negatively Regulates GITR via Ubiquitination in Immune Microenvironment of Melanoma
Authors Guo Y, Yang L, Lei S, Tan W, Long J
Received 15 April 2019
Accepted for publication 6 November 2019
Published 4 December 2019 Volume 2019:12 Pages 10629—10637
DOI https://doi.org/10.2147/OTT.S212317
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Sanjay Singh
Yu Guo, Lichang Yang, Shaorong Lei, Wuyuan Tan, Jianhong Long
Department of Plastic Surgery, Xiangya Hospital, Central South University, Changsha, Hunan 410008, People’s Republic of China
Correspondence: Jianhong Long
Email [email protected]
Introduction: Melanoma is a common skin cancer that is usually associated with poor clinical outcomes. Recently, the immune checkpoint GITR has been identified as a promising target for immunotherapy of melanoma. In this study, we aimed to investigate the post-translational regulation mechanism of GITR in melanoma.
Methods: Western blotting was used to evaluate the protein expression of NEDD4, GITR and Foxp3. Real-time PCR (RT-PCR) was performed to determine expression levels of NEDD4, GITR, Foxp3 and IL-2. Cell viability was detected by MTT assay. The ubiquitination of GITR was evaluated by immunoprecipitation. NEDD4 expression data and melanoma survival data were obtained from The Cancer Genome Atlas (TCGA) and cBioPortal databases.
Results: We demonstrate that E3 ligase NEDD4 binds to GITR and mediates ubiquitination and degradation of GITR. Overexpression of NEDD4 inhibits anti-tumor immunity mediated by T cells against melanoma cells. We also found that the expression of NEDD4 is increased in metastatic melanoma. High NEDD4 expression level is correlated with the poor prognosis of melanoma patients.
Discussion: In summary, our findings demonstrated that E3 ligase NEDD4 mediates ubiquitination and degradation of GITR and suppresses T-cell-mediated-killings on melanoma cells. Our work highlighted the E3 ligase NEDD4 as a novel prognosis biomarker and therapeutic target for melanoma.
Keywords: melanoma, glucocorticoid-induced TNF receptor, GITR, neural precursor cell expressed, developmentally down-regulated 4, NEDD4, ubiquitination
Introduction
Melanoma is one of the most aggressive cancer with high mortality rates and increasing incidences, which poses a severe threat to human health worldwide.1 The etiology and pathogenesis of melanoma, which is still unclear, may involve in many factors such as environment, genetic and immunity dysregulation.2–4 More recently, the vital role of inflammatory microenvironment in development of melanoma has been elucidated.4 Immunotherapeutic approaches are considered to have a good curative effect for melanoma patients.4 The development of targeted immunotherapy has led to great advances in the therapy of melanoma.4,5
One among the immunotherapeutic strategy was immune checkpoint blockade, which aimed at recovering the restrained function of T cells.6 The immune checkpoint GITR is a member of the TNF-receptor superfamily that highly expressed on activated T cells and plays a crucial role in the differentiation and function of Treg cells.7 It is reported that modulation of GITR showed compelling anti-tumor activity in vivo.8–11 In humanized mouse melanoma model, anti-GITR agonist showed potent antitumor efficacy via Treg depletion in tumors and reversal of CD8+ T cell exhaustion.8 In-human Phase I trial demonstrates that, TRX518, a humanized agonist anti-human GITR monoclonal antibody, enables immune attack on tumors and overcomes the resistance of advanced tumors, particularly when combined with PD-1 blockade.9 Compared with anti-GITR monotherapy, combined anti-GITR and anti-PD-1 therapy can have better effects without resistance to immunotherapy in melanoma.12
Abnormal expression of GITR is detected in cancer.13–15 GITR expression level on the surface of CD8+ T cells is elevated in melanoma mouse model.16 The proportion of GITR+ T cells in tumor-infiltrating leukocytes of primary melanoma is significantly lower compared with those of metastases melanoma.17 Epigenetic modification, such as DNA methylation, is involved in mechanisms regulating GITR expression, which may cause altered GITR expression in cancer patients.15 The post-translational control of GITR protein level is unclear.
Ubiquitination is one of the most common pathway that modulates protein proteolysis. Ubiquitination is implicated in the pathogenesis of cancer via regulating oncogenes, tumor suppressor genes and immune-related genes.18–20 Studies on ubiquitination in melanoma provide novel therapeutic targets for cancer. Inhibition of ubiquitin-protease system appears to have efficacy in advanced melanoma.21 Using ubiquitination site predictor UbiProber,22 we identified potential ubiquitinated lysine residues in the protein sequence of GITR, which suggests that protein expression of GITR may be regulated by ubiquitin-proteasome pathway. Using an integrated bioinformatics platform UbiBrowser,23 we identified neural precursor cell expressed, developmentally down-regulated 4 (NEDD4) to be potential E3 ligase related with GITR.
In the present study, we aim to explore whether ubiquitination is involved in post-translational regulation of GITR and the effect of NEDD4 on GITR protein level in melanoma. We found that E3 ligase NEDD4 down-regulates GITR via ubiquitination and is capable to promote the proliferation of melanoma cells by inhibiting anti-tumor immune response mediated by T cells. Overexpression of NEDD4 is detected in metastasis melanoma and is associated with the prognosis of patients. Our results identified a novel mechanism regulating GITR expression and indicated that NEDD4 played an important role in melanoma pathogenesis.
Methods
Cell Culture and Transfection
The human melanoma cell line A375 and Jurkat cells were purchased from ATCC. Jurkat cells were cultured in RPMI-1640 medium (Gibco) containing 10% fetal bovine serum (FBS, Gibco) at 37°C supplemented with 5% CO2. Cells were treated with 10 mM proteasome inhibitor MG132 (Sigma-Aldrich) or 10 mg/mL cycloheximide (CHX, Sigma-Aldrich) as indicated. Transfection of Jurkat cells was performed with 6 μg indicated plasmid by ECM 2001 Electro Cell Manipulator (BTX) using BTXPress High Performance Electroporation Solution (BTX) according to the manufacturer’s protocol. The transfected cells were transferred into RPMI-1640 medium with 10% FBS and collected for future study 48 hrs later. The mammalian expression constructs used for overexpression of indicated genes include Flag-NEDD4, Flag-GITR and HA-Ubiquitin. NEDD4 siRNAs and negative control siRNAs were purchased from Thermo Fisher Scientific. A375 cells were grown in DMEM medium (Gibco) containing 10% FBS at 37°C supplemented with 5% CO2.
Immunoprecipitation and Immunoblotting
Cells were washed by cold PBS and lysed by RIPA buffer containing 1% PMSF on ice. Centrifugation was performed and insoluble material was precipitated and discarded. Protein concentration was detected by BCA Protein Assay (Thermo Scientific). For immunoprecipitation, lysates containing equal amounts of protein were pre-cleared with Dynabeads Protein A/G (Life Technologies) and then incubated with indicated antibodies at 4°C overnight with rotation. The immunoprecipitates were then incubated with Dynabeads Protein A/G at room temperature for 2 hrs with rotation. Magnet was used to separate the beads. After washing several times with TBS, the beads were resuspended with SDS loading buffer and heated at 100°C for 5 mins. The beads were then subjected to immunoblotting with indicated antibodies. For immunoblotting, proteins were separated by 10% Bis-Tris SDS-PAGE gels and transferred to polyvinylidene difluoride (PVDF) membranes. Membranes were blocked with 5% non-fat milk in TBST buffer for 1 hr at room temperature and then probed with indicated primary antibodies at 4°C overnight. The membranes were washed three times with TBST buffer and incubated with HRP-conjugated secondary antibodies at room temperature for 1 hr. After washing for three times with TBST buffer, the membranes were developed and analyzed using chemiluminescence detection reagents (Advansta). Quantification was performed using Image J software. The load of target genes was normalized to β-actin. Antibodies used in this study were as follows: NEDD4 (Santa Cruz), GITR (Santa Cruz), Ubiquitin (Cell Signaling), Foxp3 (SAB), β-actin (Bioworld), anti-mouse-HRP (Santa Cruz), anti-rabbit -HRP (Santa Cruz).
RNA Extraction and Real-Time PCR
Total RNA was isolated using the TRIZOL reagent (Life Technologies). RNA was reverse-transcribed into cDNA using the TRUE Script 1st stand cDNA synthesis kit (Aidlab). Real-time PCR amplification of reversed cDNA was carried out with SYBR Green qPCR mix (Aidlab). All PCR reactions were performed in triplicate. Threshold cycle (Ct) values were used to calculate relative mRNA expression by the Ct relative quantification method. Primer sequences were: NEDD4: forward 5ʹ-TTGCAGCAACAACAAGAACC-3ʹ and reverse 5ʹ-TAGGTTGTCCTGAGGGGTTG-3ʹ; GITR: forward 5ʹ-GGTACAGTCCCAGGGGAAAT-3ʹ and reverse 5ʹ-GGTGCAGTCTGTCCAAGGTT-3ʹ; Foxp3: forward 5ʹ-CTTTCACCTACGCCACGCTCAT-3ʹ and reverse 5ʹ-TCCAGGTGGCAGGATGGTTTCT-3ʹ; IL-2: forward 5ʹ-CCAAACTCACCAGGATGCTCAC-3ʹ and reverse 5ʹ-CGTTGATATTGCTGATTAAGTCCCT-3ʹ; β-actin: forward 5ʹ-TCATCACCATTGGCAATGAGCG3ʹ and reverse 5ʹ-AGGTCTTTGCGGATGTCCACGT-3ʹ.
Co-Culture of Jurkat and Melanoma Cells
To perform the co-culture of Jurkat and melanoma cells, A375 cells were seeded in 96-well plate at a density of 1×104/well. After grown for 24 hrs in DMEM medium containing 10% FBS, the culture medium was discarded and cells were washed. Jurkat cell were isolated, pre-activated and transfected as indicated and resuspended in RPMI-1640 medium containing 1% FBS at a concentration of 1×106/mL and 100μL suspension was added into each A375 cells-precoated plate. A375 and Jurkat cells were co-cultured at 37°C supplemented with 5% CO2 for indicated time. After co-culture, cells in suspension were discarded and adherent cells were washed and collected. A375 cells that single cultured are kept as controls. Cell viability of A375 cells was detected by MTT assay.
Cell Viability Assay
To assess the cell viability of A375 cells after co-culture with Jurkat, cell survival rate was detected by MTT Assay Kit (Beyotime Biotechnology) according to the manufacturer’s instructions. The absorbance was determined at 570 nm with microplate reader (Tecan). Three independent experiments were conducted and all MTT assays were performed in triplicate.
Analysis of TCGA Data
The expression profiles and clinical data for melanoma were downloaded from TCGA (https://cancergenome.nih.gov/). We obtained melanoma mRNA RNA-Seq-HTSeq-fragments per kilobase of exon model per million mapped reads (FPKM) data from TCGA database to analyze the expression of NEDD4. The FPKM data were standardized using trans per million (TPM) for a better comparison. An unpaired t test was performed to determine the difference between NEDD4 in primary and metastasis melanoma. The melanoma survival data in TCGA, including overall survival (OS) and disease-free survival (DFS), were obtained from cBioPortal for Cancer Genomics database.24,25 We conducted Kaplan–Meier survival analysis according to the expression of NEDD4.
Statistical Analyses
Statistical analysis of comparisons between two groups was performed with unpaired T test. Multiple comparisons were analyzed by one-way analysis of variance. Data was presented as an average of at least three independent experiments. Continuous variables were expressed as mean ± SD. A value of p<0.05 was considered statistically significant. All of the data were analyzed by SPSS version 22.0 software (IBM).
Results
Ubiquitin-Proteasome Pathway Regulates Protein Expression of GITR
To explore the post-translational modifications of GITR, we first determined the protein level of GITR after the treatment of proteasome inhibitor MG132 or protein synthesis inhibitor cycloheximide (CHX) in Jurkat cells. As shown in Figure 1A and B, the GITR protein level was increased after MG132 treatment and reduced after CHX treatment, suggesting that GITR is degraded by the proteasome. Jurkat cells were transfected with HA-ubiquitin plasmid. The ubiquitination level of GITR was increased and protein level was decreased 48 hrs after transfection of ubiquitin (Figure 1C). These results indicate that the degradation of GITR is via ubiquitin-proteasome pathway.
 |
Figure 1 GITR is degraded via the ubiquitin–proteasome pathway. GITR protein level was detected by immunoblotting after treatment with 20 μM MG132 (A) or 100 ng/mL cycloheximide (CHX) (B) for 0, 3 or 6 hrs. (C) Jurkat cells were transfected with HA-ubiquitin or control plasmid. Forty-eight hours after transfection, endogenous GITR was immunoprecipitated (IP) from cell lysates with anti-GITR antibody and the immunoprecipitates were analyzed by immunoblotting (IB) with anti-HA antibody (*p<0.05; n = 3). |
NEDD4 Binds to GITR
To explore the relationship between NEDD4 and GITR, Jurkat cells were collected, lysed and subjected to immunoprecipitation (IP) and Western blot analysis. As shown in Figure 2A and B, the IP was performed with anti-GITR or anti-NEDD4 antibody, and interaction of NEDD4 and GITR was detected. A clear association between NEDD4 and GITR can be observed.
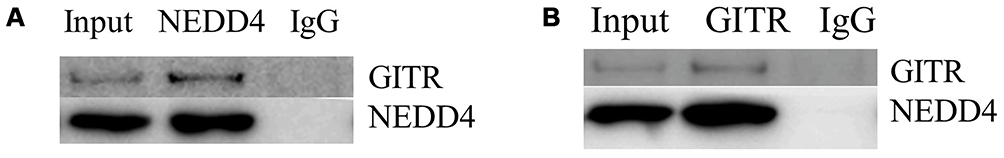 |
Figure 2 NEDD4 binds to GITR. Jurkat cells were collected and lysates were immunoprecipitated with anti-NEDD4 (A) or anti-GITR (B) antibody followed by immunoblotting with anti-GITR or anti-NEDD4 antibody (n=3). Cell lysates were then immunoblotted with anti-NEDD4 and anti-GITR antibodies. |
NEDD4 Mediates Degradation and Ubiquitination of GITR
We explored whether NEDD4 inhibits GITR expression via ubiquitination. Jurkat cells were transfected with Flag-NEDD4 or vector plasmid. As shown in Figure 3A, GITR protein level was decreased and amount of poly-ubiquitinated GITR was increased significantly after the addition of NEDD4. Consistently, knocking down of NEDD4 by siRNA led to markedly increased GITR and impaired ubiquitination of GITR (Figure 3B). The above results suggest that NEDD4 may promote the degradation of GITR through the ubiquitination mechanism. We then tested whether NEDD4 would affect the expression of Treg marker Foxp3 and IL-2. As shown in Figure 3A, protein expression of Foxp3 was increased in NEDD4-Flag transfected cells. Loss of NEDD4 by siRNA transfection resulted in decreased protein expression of Foxp3 (Figure 3B). We observed significantly higher levels of IL-2 and Foxp3 mRNA in NEDD4-Flag transfected cells compared with control group (Figure 3C and D). The mRNA levels of IL-2 and Foxp3 were clearly decreased after NEDD4 knockdown (Figure 3C and D).
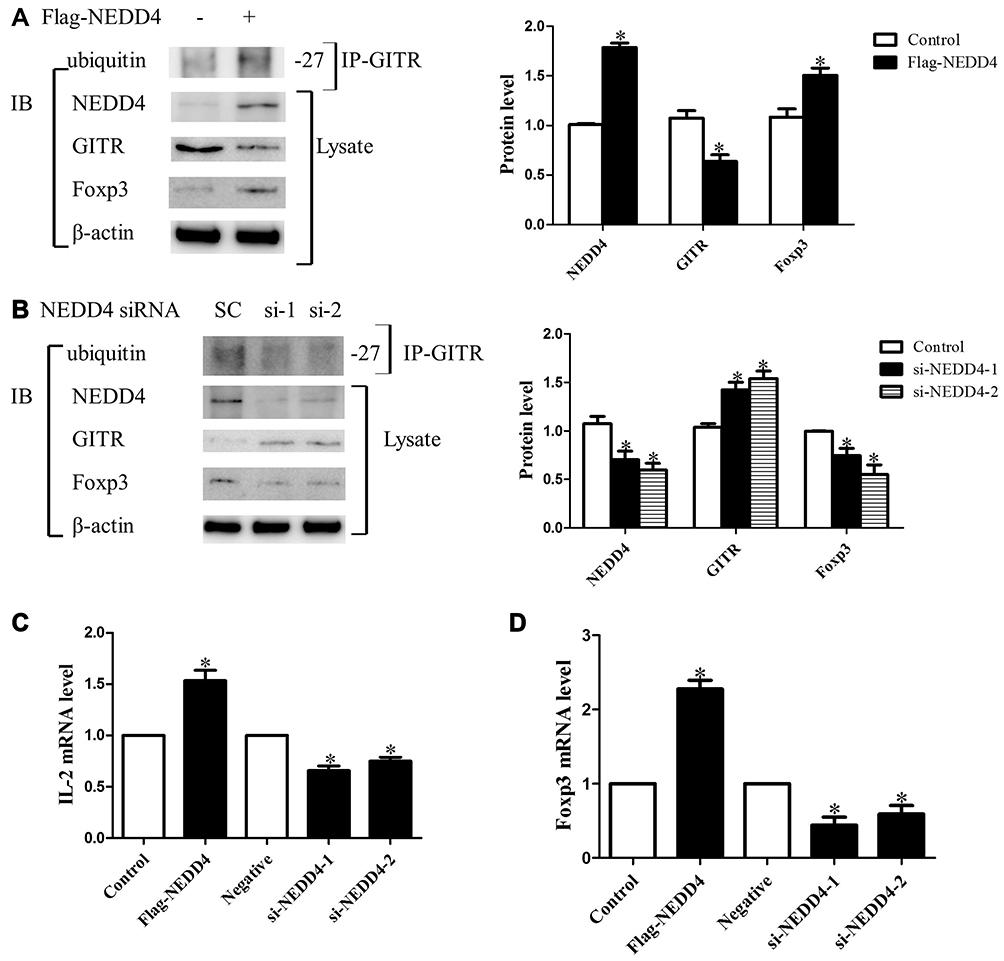 |
Figure 3 NEDD4 mediates degradation and ubiquitination of GITR. Jurkat cells were transfected with Flag-NEDD4 (A) or transfected with NEDD4-RNAi (B). Forty-eight hours after transfection, cells were collected and cell lysates were immunoblotted with NEDD4, GITR, Foxp3 and β-actin. Ubiquitination level of GITR was detected with immunoprecipitation using anti-GITR antibody followed by immunoblotting using anti-ubiquitin antibody. IL-2 (C) and Foxp3 (D) mRNA expression was measured by real time-PCR and normalized to β-actin (*p<0.05; n = 3). |
NEDD4 Inhibit T Cell-Mediated Immune Attack Upon Melanoma Cells
To confirm the inhibitory effect of T cells on the proliferation of melanoma cells, we explored the viability of A375 cells when cultured alone or together with anti-CD3 plus anti-CD28 antibodies activated Jurkat cells by MTT assay. As shown in Figure 4A, cell viability of melanoma cells was significantly decreased after 72 hrs of co-culture with activated Jurkat cells when compared with control. We then explored the potential role of NEDD4 in Jurkat cell-mediated inhibition of melanoma cell proliferation. Jurkat cells were activated with anti-CD3 plus anti-CD28 antibodies for 24 hrs and then transfected with Flag-NEDD4 or Flag-GITR or co-transfected with Flag-NEDD4 and Flag-GITR. The transfection efficiency was confirmed by RT-PCR (Figure 4C) and Western-blot (Figure 4D). Cells were then co-cultured with melanoma cell line A375 for 72 hrs. As expected, a significantly lower survival rate of A375 was observed after co-culture with Flag-GITR Jurkat cells compared with control (Figure 4B). The survival rate of A375 was significantly higher after co-culture with Flag-NEDD4 Jurkat cells compared with control, which was restored by co-transfection with Flag-NEDD4 and Flag-GITR (Figure 4B). These results indicate that NEDD4 mediated GITR degradation may lead to reduced anti-tumor effect of immune cells and increased proliferation of melanoma cells.
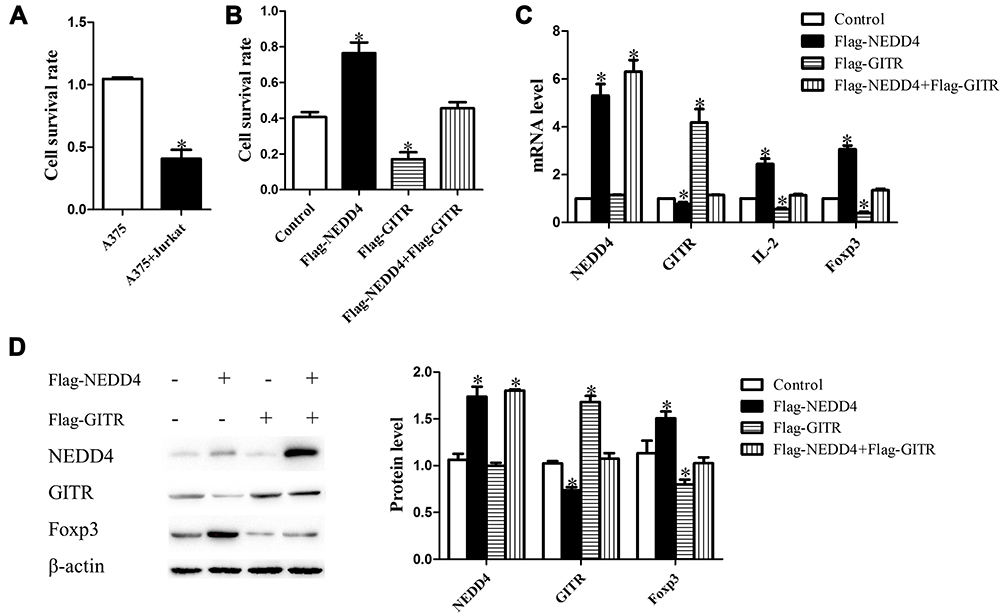 |
Figure 4 3.4 NEDD4 inhibit T cell-mediated immune attack upon melanoma cells. (A) Jurkat cells activated with anti-CD3 plus anti-CD28 antibodies were co-incubated or not with A375, respectively. Survival fraction of A375 cells was detected with MTT assays (n = 3). Jurkat cells were activated with anti-CD3 plus anti-CD28 antibodies and then transfected with Flag-NEDD4 or Flag-GITR or co-transfected with Flag-NEDD4 and Flag-GITR. Forty-eight hours after transfection, cells were co-incubated with A375. (B) Survival fraction of A375 cells was detected with MTT assays (*p<0.05; n = 3). Cells were then collected and NEDD4, GITR, IL-2 and Foxp3 mRNA expression was measured by real time-PCR and normalized to β-actin (C). Cell lysates were immunoblotted with NEDD4, GITR, Foxp3 and β-actin (D). |
NEDD4 Is Highly Expressed in Metastasis Melanoma and Associated with Poor Prognosis
We analyzed the TCGA data for melanoma to explore the clinical significance of NEDD4 expression in melanoma patients. We first determined the expression of NEDD4 in melanoma patients. As shown in Figure 5A, NEDD4 mRNA was significantly higher in metastasis melanoma patients compared with that in primary melanoma patients. We analyzed the TCGA data for melanoma to evaluate the relationship between NEDD4 and prognosis of melanoma patients by Kaplan–Meier analysis. Patients with genetic alterations in NEDD4 expressed significantly higher gene level compared with those without alterations (Figure 5B). The Kaplan-Meier analysis results showed that alteration carriers with high NEDD4 expression tended to associate with poor overall survival (Figure 5C). Alteration carriers with higher NEDD4 expression level were significantly correlated with poor disease-free survival (Figure 5D). These results suggest that NEDD4 may function as an oncogene in melanoma.
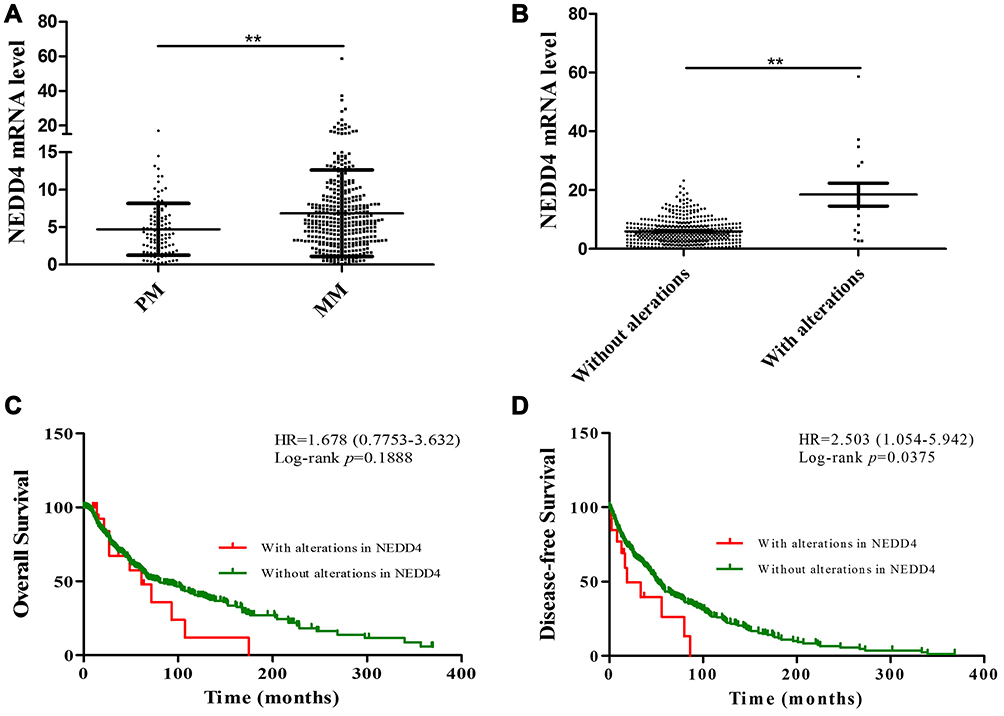 |
Figure 5 NEDD4 expression is highly expressed in metastasis melanoma and correlated with poor prognosis of patients. Using TCGA data, NEDD4 expression in primary (PM) and metastasis (MM) melanoma was compared (A). Expression of NEDD4 in patients with alterations was compared to those without alterations (B). Kaplan–Meier curves for overall survival (C) or disease-free survival (D) of melanoma patients carrying NEDD4 mutation or not (**p<0.01). |
Discussion
Immune checkpoint GITR is a promising drug target for immunotherapy of melanoma and plays an essential role in the pathogenesis of the disease. In this study, we explored the post-translational regulation mechanism of GITR. We found that E3 ligase NEDD4 binds to and inhibits GITR via ubiquitination to inhibit the anti-tumor effect of T cells upon melanoma cells. NEDD4 expression is increased in metastasis melanoma and is associated with shorter disease-free survival in patients.
GITR is altered in several diseases in addition of cancer and is associated with prognosis of patients.13–15,26 Abnormal GITR has been reported to be a result of both genetic and epigenetic changes.15,26–28 GITR polymorphisms contribute to the individual differences in the proportion of peripheral GITR+ cells in patients and are correlated with susceptibility and prognosis of autoimmune diseases.26 Previous research also revealed that altered GITR expression in tumors is partly due to epigenetic modification mechanisms, such as DNA methylation.15,27 Transcription factor methyl-CpG-binding domain Protein 4 (MBD4), which is overexpressed in Treg cells, binds to and recruits DNA methyltransferases 1 (DNMT1) to GITR promoter and inhibits its expression and functions.27 Interestingly, there is a relationship between genetic variants in GITR promoter and autoimmune and inflammatory diseases.26,28 It is possible that GITR promoter polymorphism may affect the binding of DNMT1, thus result in dysregulation of GITR expression in patients. In this study, we found that E3 ligase NEDD4 binds to GITR and mediates its ubiquitination and degradation.
In our study, we found that NEDD4 induces degradation of GITR via ubiquitin-proteasome pathway. NEDD4 is an E3 ubiquitin-protein ligase that participants in the founding of the homologous to E6-AP carboxyl terminus (HECT)-type E3 ligases family. It mainly mediates ubiquitination and degradation of membrane proteins. NEDD4 promotes growth and inhibits apoptosis of tumor cells.29 Repression of NEDD4 results in reversion of epithelial–mesenchymal transition (EMT) features, which suggest that NEDD4 is an oncogene and could serve as an attracting drug target in cancer.30 Several studies found that the treatment of NEDD4 inhibitors leads to the upregulation of tumor suppressor PTEN and impaired melanoma cell proliferation.31,32 However, the precise role of NEDD4 in the development of melanoma has not been fully investigated. Clinical benefit after GITR agonist treatment is documented to be correlated with tumor microenvironment containing high ratio of Treg cells.8 GITR upregulates the NF-κB family member p50 to inhibit Foxp3 via regulation of its histone acetylation status, which subverts the induction of Foxp3+ Tregs.33 According to our results, Jurkat cells that overexpressed NEDD4 displayed reduced anti-tumor effect on A375 cells, with increased Foxp3 and IL-2. Therefore, it is reasonable to speculate that NEDD4 may activate tumor-suppressive Treg cells via mediating GITR degradation, thus contributing to melanoma progression.
Prognosis of advanced melanoma patients is far from satisfactory, which makes it very meaningful to identify novel biomarkers that predict early recurrence or distant organ metastasis of the disease.34 GITR expression in metastasis tumor is significantly higher than in primary tumor.14 Tang C et al35 found that after Ipilimumab treatment combined with radiotherapy, melanoma patients who achieved markedly clinical benefit exhibited significant increases in the GITR expressing T cells count compared to those who did not. These results indicate that GITR may be a potential biomarker for diagnostic and immunotherapy of tumors. We found that NEDD4 is highly expressed in metastatic melanoma and is associated with poor survival of melanoma patients, which provides NEDD4 as a potential biomarker for diagnosis and prognosis of melanoma.
In conclusion, we identified E3 ligase NEDD4 as a new negative regulator that involved in post-translational modifications of GITR. It mediates ubiquitination and degradation of GITR and suppresses T cell mediated-killings on melanoma cells. NEDD4 is highly expressed in metastasis melanoma and associated with poor prognosis of patients. Hence, NEDD4 may be a potential drug target and valuable prognosis indicator for melanoma patients.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (No. 81571945), Hunan Provincial Key Research and Development Plan (No. 2016SK2092) and Xiangya Famous Medicine Project in Central South University.
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Whiteman DC, Green AC, Olsen CM. The growing burden of invasive melanoma: projections of incidence rates and numbers of new cases in six susceptible populations through 2031. J Invest Dermatol. 2016;136(6):1161–1171. doi:10.1016/j.jid.2016.01.035
2. Bald T, Quast T, Landsberg J, et al. Ultraviolet-radiation-induced inflammation promotes angiotropism and metastasis in melanoma. Nature. 2014;507(7490):109–113. doi:10.1038/nature13111
3. Fu S, Wu H, Zhang H, Lian CG, Lu Q. DNA methylation/hydroxymethylation in melanoma. Oncotarget. 2017;8(44):78163–78173. doi:10.18632/oncotarget.18293
4. Nakamura K, Okuyama R. Immunotherapy for advanced melanoma: current knowledge and future directions. J Dermatol Sci. 2016;83(2):87–94. doi:10.1016/j.jdermsci.2016.05.009
5. Ascierto PA, Flaherty K, Goff S. Emerging strategies in systemic therapy for the treatment of melanoma. Am Soc Clin Oncol Educ Book. 2018;38:751–758.
6. Wang W, Liu J, He Y, McLeod HL. Prospect for immune checkpoint blockade: dynamic and comprehensive monitorings pave the way. Pharmacogenomics. 2017;18(13):1299–1304. doi:10.2217/pgs-2017-0053
7. Knee DA, Hewes B, Brogdon JL. Rationale for anti-GITR cancer immunotherapy. Eur J Cancer. 2016;67:1–10. doi:10.1016/j.ejca.2016.06.028
8. Mahne A, Mauze S, Joyce-Shaikh B, et al. Dual roles for regulatory T cell depletion and co-stimulatory signaling in agonistic GITR targeting for tumor immunotherapy. Cancer Res. 2017;77(5):1108–1118. doi:10.1158/0008-5472.CAN-16-0797
9. Zappasodi R, Sirard C, Li Y, et al. Rational design of anti-GITR-based combination immunotherapy. Nat Med. 2019;25(5):759–766. doi:10.1038/s41591-019-0420-8
10. Melero I, Hirschhorn-Cymerman D, Morales-Kastresana A, Sanmamed MF, Wolchok JD. Agonist antibodies to TNFR molecules that costimulate T and NK cells. Clin Cancer Res. 2013;19(5):1044–1053. doi:10.1158/1078-0432.CCR-12-2065
11. Scirka B, Szurek E, Pietrzak M, et al. Anti-GITR antibody treatment increases TCR repertoire diversity of regulatory but not effector T cells engaged in the immune response against B16 melanoma. Arch Immunol Ther Exp (Warsz). 2017;65(6):553–564. doi:10.1007/s00005-017-0479-1
12. GITR agonism overcomes resistance to immunotherapy in solid tumors. Cancer Discov. 2019;9(7):824.
13. Benevides L, Cardoso CR, Tiezzi DG, Marana HR, Andrade JM, Silva JS. Enrichment of regulatory T cells in invasive breast tumor correlates with the upregulation of IL-17A expression and invasiveness of the tumor. Eur J Immunol. 2013;43(6):1518–1528. doi:10.1002/eji.201242951
14. Pedroza-Gonzalez A, Verhoef C, Ijzermans JN, et al. Activated tumor-infiltrating CD4+ regulatory T cells restrain antitumor immunity in patients with primary or metastatic liver cancer. Hepatology. 2013;57(1):183–194. doi:10.1002/hep.26013
15. Liu Y, Quang P, Braggio E, et al. Novel tumor suppressor function of glucocorticoid-induced TNF receptor GITR in multiple myeloma. PLoS One. 2013;8(6):e66982. doi:10.1371/journal.pone.0066982
16. Cote AL, Zhang P, O’Sullivan JA, et al. Stimulation of the glucocorticoid-induced TNF receptor family-related receptor on CD8 T cells induces protective and high-avidity T cell responses to tumor-specific antigens. J Immunol. 2011;186(1):275–283. doi:10.4049/jimmunol.1001308
17. Edwards J, Tasker A, Pires da Silva I, et al. Prevalence and cellular distribution of novel immune checkpoint targets across longitudinal specimens in treatment-naive melanoma patients: implications for clinical trials. Clin Cancer Res. 2019;25:3247–3258. doi:10.1158/1078-0432.CCR-18-4011
18. Haymaker C, Yang Y, Wang J, et al. Absence of Grail promotes CD8(+) T cell anti-tumour activity. Nat Commun. 2017;8(1):239. doi:10.1038/s41467-017-00252-w
19. Wang Y, Ou Z, Sun Y, et al. Androgen receptor promotes melanoma metastasis via altering the miRNA-539-3p/USP13/MITF/AXL signals. Oncogene. 2017;36(12):1644–1654. doi:10.1038/onc.2016.330
20. Luo Z, Zhang X, Zeng W, et al. TRAF6 regulates melanoma invasion and metastasis through ubiquitination of Basigin. Oncotarget. 2016;7(6):7179–7192. doi:10.18632/oncotarget.6886
21. Olaverria Salavaggione GN, Duggan MC, Carson WE. Analysis of MLN4924 (pevonedistat) as a potential therapeutic agent in malignant melanoma. Melanoma Res. 2018;1. doi:10.1097/CMR.0000000000000474
22. Chen X, Qiu JD, Shi SP, Suo SB, Huang SY, Liang RP. Incorporating key position and amino acid residue features to identify general and species-specific Ubiquitin conjugation sites. Bioinformatics. 2013;29(13):1614–1622. doi:10.1093/bioinformatics/btt196
23. Li Y, Xie P, Lu L, et al. An integrated bioinformatics platform for investigating the human E3 ubiquitin ligase-substrate interaction network. Nat Commun. 2017;8(1):347. doi:10.1038/s41467-017-00299-9
24. Gao J, Aksoy BA, Dogrusoz U, et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci Signal. 2013;6(269):pl1. doi:10.1126/scisignal.2004088
25. Cerami E, Gao J, Dogrusoz U, et al. The cBio cancer genomics portal: an open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012;2(5):401–404. doi:10.1158/2159-8290.CD-12-0095
26. Tomizawa R, Watanabe M, Inoue N, et al. Association of functional GITR gene polymorphisms related to expression of glucocorticoid-induced tumour necrosis factor-receptor (GITR) molecules with prognosis of autoimmune thyroid disease. Clin Exp Immunol. 2011;165(2):141–147. doi:10.1111/cei.2011.165.issue-2
27. Wang S, Li Y, Zhu F, et al. DNMT1 cooperates with MBD4 to inhibit the expression of Glucocorticoid-induced TNFR-related protein in human T cells. FEBS Lett. 2017;591(13):1929–1939. doi:10.1002/feb2.2017.591.issue-13
28. Wu B, Ji X, Han R, et al. GITR promoter polymorphism contributes to risk of coal workers’ pneumoconiosis: a case-control study from China. Immunol Lett. 2014;162(2 Pt B):210–216. doi:10.1016/j.imlet.2014.10.023
29. Wen W, Li J, Wang L, et al. Inhibition of NEDD4 inhibits cell growth and invasion and induces cell apoptosis in bladder cancer cells. Cell Cycle. 2017;16(16):1509–1514. doi:10.1080/15384101.2017.1338220
30. Feng S, Yang G, Yang H, et al. NEDD4 is involved in acquisition of epithelial-mesenchymal transition in cisplatin-resistant nasopharyngeal carcinoma cells. Cell Cycle. 2017;16(9):869–878. doi:10.1080/15384101.2017.1308617
31. Aronchik I, Kundu A, Quirit JG, et al. The antiproliferative response of indole-3-carbinol in human melanoma cells is triggered by an interaction with NEDD4-1 and disruption of wild-type PTEN degradation. Mol Cancer Res. 2014;12(11):1621–1634. doi:10.1158/1541-7786.MCR-14-0018
32. Quirit JG, Lavrenov SN, Poindexter K, et al. Indole-3-carbinol (I3C) analogues are potent small molecule inhibitors of NEDD4-1 ubiquitin ligase activity that disrupt proliferation of human melanoma cells. Biochem Pharmacol. 2017;127(undefined):13–27. doi:10.1016/j.bcp.2016.12.007
33. Xiao X, Shi X, Fan Y, et al. GITR subverts Foxp3(+) Tregs to boost Th9 immunity through regulation of histone acetylation. Nat Commun. 2015;6:8266. doi:10.1038/ncomms9266
34. Guo Y, Long J, Lei S. Promoter methylation as biomarkers for diagnosis of melanoma: a systematic review and meta-analysis. J Cell Physiol. 2018;234(11).
35. Tang C, Welsh JW, de Groot P, et al. Ipilimumab with stereotactic ablative radiation therapy: Phase I results and immunologic correlates from peripheral T cells. Clin Cancer Res. 2016.
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.