Back to Journals » OncoTargets and Therapy » Volume 11
NDRG3 facilitates colorectal cancer metastasis through activating Src phosphorylation
Authors Li T, Sun R, Lu MD ![]() , Chang J, Meng X, Wu Huo
, Chang J, Meng X, Wu Huo
Received 12 November 2017
Accepted for publication 5 March 2018
Published 15 May 2018 Volume 2018:11 Pages 2843—2852
DOI https://doi.org/10.2147/OTT.S156814
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Ingrid Espinoza
Ting Li, Ruochuan Sun, Mingdian Lu, Jiacong Chang, Xiangling Meng,* Huo Wu*
Department of General Surgery, The First Affiliated Hospital of Anhui Medical University, He Fei, 230222, China
*These authors contributed equally to this work
Background: NDRG3 is an N-myc downregulated gene (NDRG). The aim of this article was to identify the role of NDRG3 in colorectal cancer (CRC) and to determine the mechanism underlying its function.
Methods: Using immunohistochemical staining, expression and clinicopathological variables of NDRG3 were analyzed in 170 CRC samples. Overexpression of NDRG3 was employed in SW1116 cells, downregulation of NDRG3 was achieved in RKO cells, then migration and invasion assays were performed in vitro, and a mouse model was constructed in vivo.
Results: Increased expression of NDRG3 was observed in primary CRC tissues, and this expression was correlated with distant metastasis. Consistently, ectopic expression of NDRG3 in SW1116 cells enhanced cell migration and invasion, while knockdown of NDRG3 in RKO cells significantly suppressed CRC cell metastasis. The portal vein injection models suggested that NDRG3 overexpression facilitates liver metastasis. These events were associated with the phosphorylation of Src (c-Src) at Tyr 419 site.
Conclusion: Our results showed that NDRG3 facilitates CRC migration and invasion by activating Src phosphorylation, suggesting the role of NDRG3 as a candidate oncogene.
Keywords: NDRG3, colorectal cancer, metastasis, Src
Introduction
Colorectal cancer (CRC), being the third most common cancer and the fourth most common cause of oncological death worldwide, is a disease of high morbidity and mortality.1 In recent years, with the prevalence of westernized lifestyle, high CRC rates have been reported in the developing countries in which the risk was once low.2 Despite noticeable improvements in early diagnosis and development of novel therapy, CRC causes a large number of deaths ascribed to metastatic events.3 Therefore, it is of urgent need to elucidate the molecular mechanism that is involved in the progression of CRC for early detection and target treatment.
Recently, the N-myc downregulated gene (NDRG) family has been discovered, which is involved in cell proliferation and differentiation.4 The NDRG family is composed of four members: NDRG1, NDRG2, NDRG3, and NDRG4, and they share a 57%–65% amino acid identity to one another.4,5 Phylogenetic analysis showed that NDRG1 and NDRG3 belong to one subfamily, whereas NDRG2 and NDRG4 belong to another.5 NDRG1 was first discovered and can be activated by various conditions including androgens and hypoxia.5–9 NDRG1 has been widely found to enhance differentiation, reduce proliferation rate, and suppress cancer metastasis of various types of cancer cells, indicating the important role of NDRG1 in tumor suppressing.10–12 Interestingly, NDRG1 can upregulate transcription factor p53 and is necessary for p53-mediated apoptosis.13 NDRG2 is upregulated in Alzheimer’s disease (AD) brains and is associated with AD pathogenesis.14 NDRG2 can markedly reduce cell proliferation of glioblastoma.15 So far, little is known about the function of NDRG4 in tumors.
NDRG3 is a critical gene that leads to homologous lethality in the early embryo development and regulates the male meiosis in mouse.16 NDRG3 can be detected in multiple human tissues, with high expression in testis, prostate, and ovary and low expression in placenta, lung, liver, skeletal muscle, and so on. This expression pattern is very similar to that of NDRG1.16,17 To date, however, little is known about the exact role of NDRG3 involved in tumorigenesis. The aim of this study was to explore the expression of NDRG3 in CRC and investigate its role in cell proliferation, migration, and invasion both in vitro and in vivo.
Materials and methods
Tissue specimens and cell lines
Colorectal tumor and adjacent non-tumor tissues of 170 CRC patients were obtained from Ruijin Hospital, School of Medicine, Shanghai Jiaotong University. All patients had undergone radical surgery at Ruijin Hospital from January 2009 to January 2012. The mean age of the patients was 65.6 years (range: 30–79 years). None of them had received neoadjuvant therapy such as radiotherapy or chemotherapy prior to surgery. Clinicopathological data were collected and staged according to the UICC TNM classification. This study was approved by the ethics committee of Shanghai Ruijin Hospital, and written informed consent was obtained from all the patients.
The human CRC cell lines SW1116, CaCo2, HT-29, LoVo, HCT116, SW480, SW620, and RKO were obtained from American Type Culture Collection (Manassas, VA, USA) and preserved in the Shanghai Digestive Surgery Institute. SW1116, SW480, and SW620 were cultured in Leibovitz’s L-15 medium. CaCo2 and RKO were cultured in Eagle’s minimum essential medium. HT-29 and HCT116 were cultured in McCoy’s 5a Medium. LoVo was cultured in F-12K medium. All media were supplemented with 10% fetal calf serum, 100 μg/mL streptomycin, 100 U/mL penicillin, and cultured at 37°C in 5% CO2.
RNA extraction and real-time polymerase chain reaction (PCR)
The total RNA of CRC tissues and cell lines was extracted with TRIzol reagent (Thermo Fisher Scientific, Waltham, MA, USA) according to the manufacturer’s instructions. The isolated RNA was reverse transcribed using reverse transcription kit (Promega Corporation, Fitchburg, WI, USA). Primers of NDRG3 were 5′-TCTTTCCCAACAGGGTATCAGT-3′ (forward) and 5′-GCTTAGGTGGGTAAGAACAGGA-3′ (reverse). Primers of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) GAPDH were 5′-GGACCTGACCTGCCGTCTAG-3′ (forward) and 5′-GTAGCCCAGGATGCCCTTGA-3′ (reverse). Quantitative reverse transcription PCR (RT-PCR) analysis was carried out, and GAPDH served as a constitutive control.
Establishment of NDRG3 knockdown and NDRG3 overexpression
Cells
The lentivirus particles for CTTN overexpression or knockdown were designed and constructed by Shanghai GenePharma Co., Ltd. (Shanghai, China) SW1116 cells were transduced with Lenti-vector or Lenti-NDRG3 at 20 multiplicity of infection in the presence of 8 μg/mL polybrene (Sigma-Aldrich Co., St Louis, MO, USA). RKO cells were transduced with Lenti-control or Lenti-sh NDRG3 at 20 multiplicity of infection in the presence of 8 μg/mL polybrene. Stable cell lines were selected in 2 μg/mL puromycin.
Immunohistochemistry (IHC)
IHC was performed as previously described.18 NDRG3 was purchased from Abcam (Cambridge, UK) and diluted as 1:1,000.
Western blotting
Total proteins were extracted using radio immunoprecipitation assay buffer containing 1% phenylmethanesulfonyl fluoride and quantified by a BCA Protein Assay Kit (Pierce, Rockford, IL, USA). Equivalent amounts of protein were loaded onto 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis and then transferred into polyvinylidene difluoride membranes. After blocked by skim milk, membranes were incubated in the primary antibodies overnight at 4°C. Primary antibodies were as follows: NDRG3 (1:1,000; Abcam), Src (1:500; Cell Signaling Technology, Boston, MA, USA), p-Src (Try419, 1:1,000; Santa Cruz Biotechnology Inc., Dallas, TX, USA), and GAPDH (1:10,000; Abcam). Membranes were then incubated with horseradish peroxidase-conjugated secondary antibody for 2 hours at room temperature and were visualized by a Tanon detection system with electrochemiluminescence reagent.
Cell counting kit-8 (CCK-8) and colony formation assay
CCK-8 and colony formation assay were performed as previously described.19
Wound healing assay
Cells were cultured in serum-free medium and grown to 100% confluence in 6-well plates. After scratching the cell monolayer with a sterile pipette tip, the cells were washed twice with 1% PBS. The wound closing procedure was observed and photographed at 0, 24, and 48 hours under an inverted phase-contrast microscope (Olympus Corporation, Tokyo, Japan).
Migration and invasion assay
For migration assay, 200 μL of serum-free medium containing 1×105 cells was added into the upper transwell chamber (8 mm, Corning Costar, Corning, NY, USA) and 600 μL of the medium with 10% serum was added into the lower transwell chamber. The chamber was cultured in 37°C with 5% CO2 for 24 hours and fixed with methanol. After staining with 0.4% crystal violet for 30 minutes, cells were photographed under an inverted phase-contrast microscope (Olympus Corporation). For invasion assay, the insert membranes were precoated with Matrigel (BD Biosciences, San Jose, CA, USA) and the assay was performed in the same way as migration assay.
Xenograft model
Equal numbers of RKO/NC, RKO/sh-NDRG3 cells (1×106) were injected into the portal vein of ten 4-week-old BALB/c nude mice. Six weeks after injection, the mice were sacrificed, and the numbers of tumor colonies in the livers were measured. Subcutaneous tumor formation experiment was performed as previously described.19 The experiment was performed according to the Guide for the Care and Use Laboratory Animals of Ruijin Hospital, Shanghai Jiaotong University School of Medicine. Ethics committee of Ruijin Hospital also approved this research for using animals.
Statistical analyses
All tests were performed by SPSS 13.0 software. Quantitative variables were analyzed by Student’s t-test. Chi-square or Fisher’s exact test was used to analyze correlation between NDRG3 expression and clinical characteristics. p<0.05 was considered to be significant and p<0.01 was considered to be highly significant.
Results
NDRG3 expression in CRC and its correlation with clinicopathological parameters
To investigate the NDRG3 expression in CRC tissues, RT-PCR and IHC staining were performed on 170 tumor and matched non-tumor tissues. As shown in Figure 1A, NDRG3 transcript levels were significantly increased in CRC samples compared to normal tissues (p<0.01). Ninety-two (54.1%) of the CRC tissues showed NDRG3-positive staining, whereas 78 of the cancer tissues showed a negative NDRG3 signal (Figure 1B). Furthermore, a correlation analysis between NDRG3 expression and the clinicopathological features revealed that the expression of NDRG3 in the CRC tissues was significantly related to invasion depth (p=0.013) and TNM stage (p=0.015) but not to other factors (Table 1).
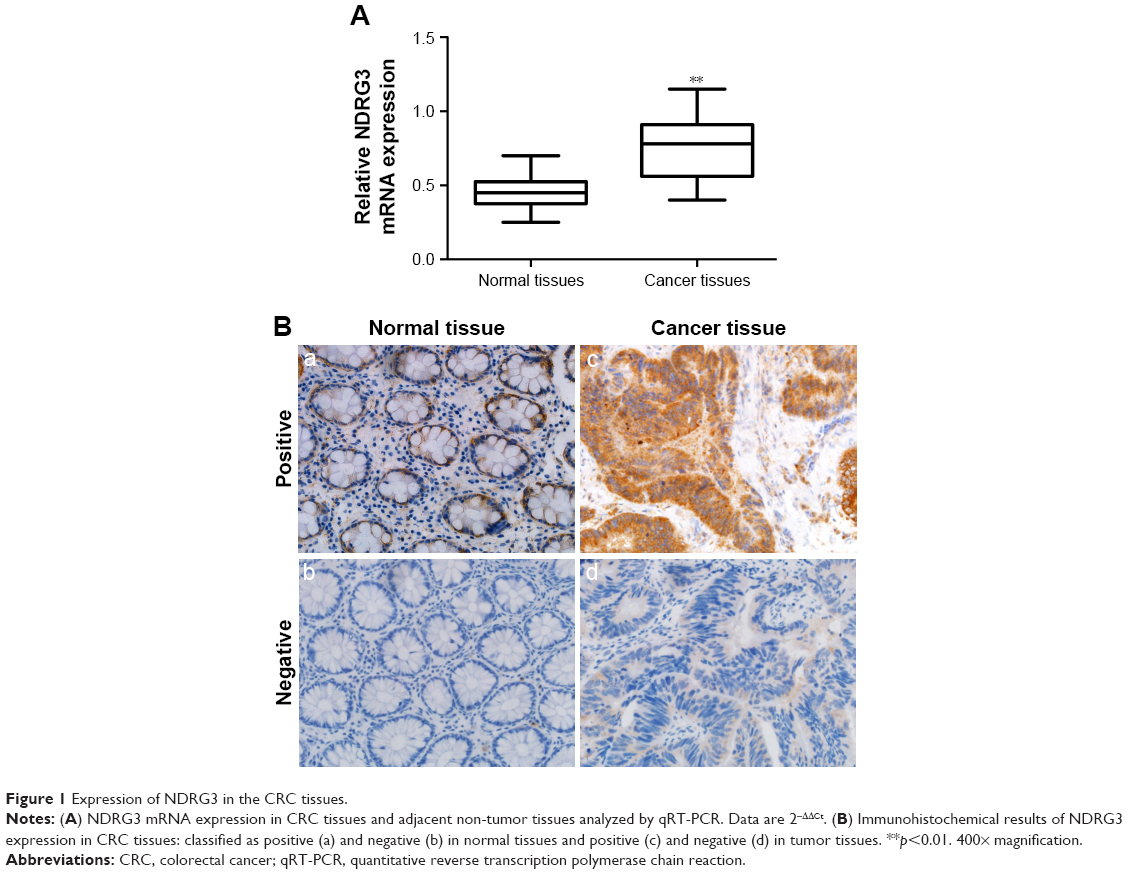 | Figure 1 Expression of NDRG3 in the CRC tissues. |
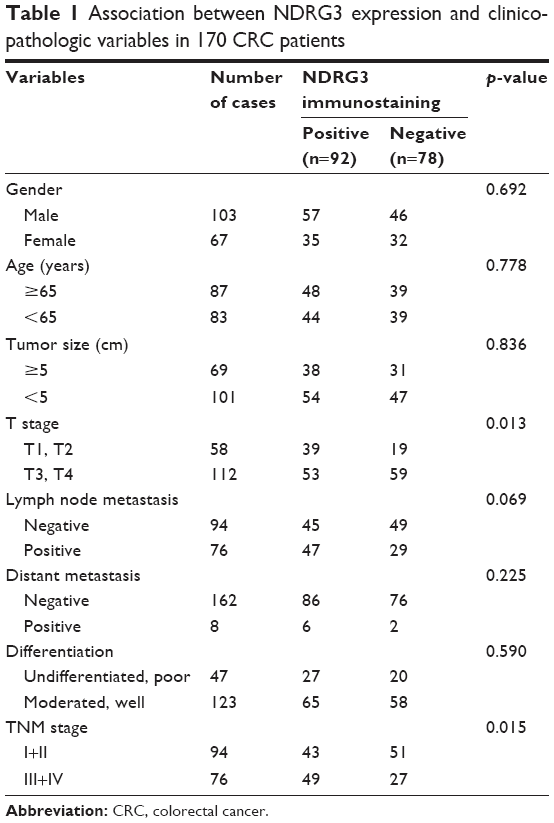 | Table 1 Association between NDRG3 expression and clinicopathologic variables in 170 CRC patients |
Construction of stable CRC cell lines
We explored the NDRG3 expression in a series of CRC cell lines and found that RKO exhibited relatively higher levels of NDRG3 expression and SW1116 exhibited low levels of NDRG3 expression (Figure 2A). In order to explore the functions of NDRG3 in CRC cell lines, we generated a SW1116 cell line ectopically overexpressing NDRG3 and applied lentivirus-mediated shRNA to knockdown NDRG3 in RKO. The effect of overexpression and knockdown was determined by Western blotting (Figure 2B).
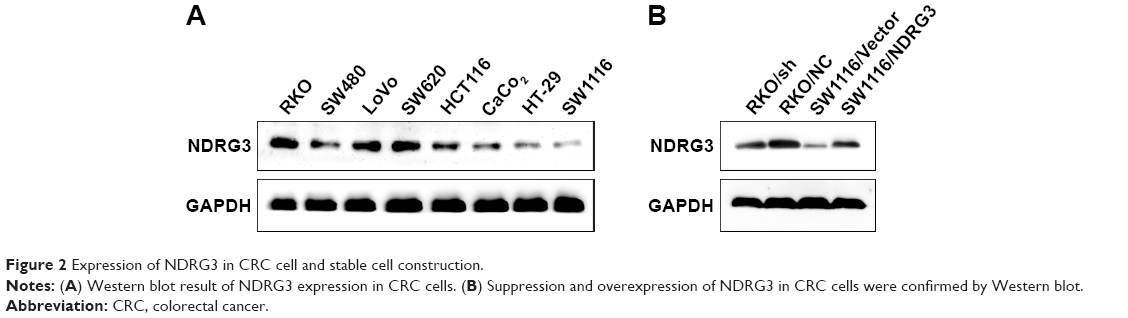 | Figure 2 Expression of NDRG3 in CRC cell and stable cell construction. |
NDRG3 promotes CRC tumor proliferation
To investigate the effect of NDRG3 expression on CRC growth, we examined the in vitro cell growth rates by CCK-8 and colony formation assays. Overexpression of NDRG3 increased the proliferation rate of the SW1116 cells, while depletion of NDRG3 reduced the proliferation rate of the RKO cells (Figure 3A–C). Consistently, there was a greater increase in both tumor volume and tumor weight in animals injected with SW1116/NDRG3 cells than in those injected with SW1116/Vector cells (Figure 4A–C). IHC analysis revealed that SW1116/NDRG3 tumors displayed a higher Ki67 index when compared with SW1116/Vector tumors (Figure 4D).
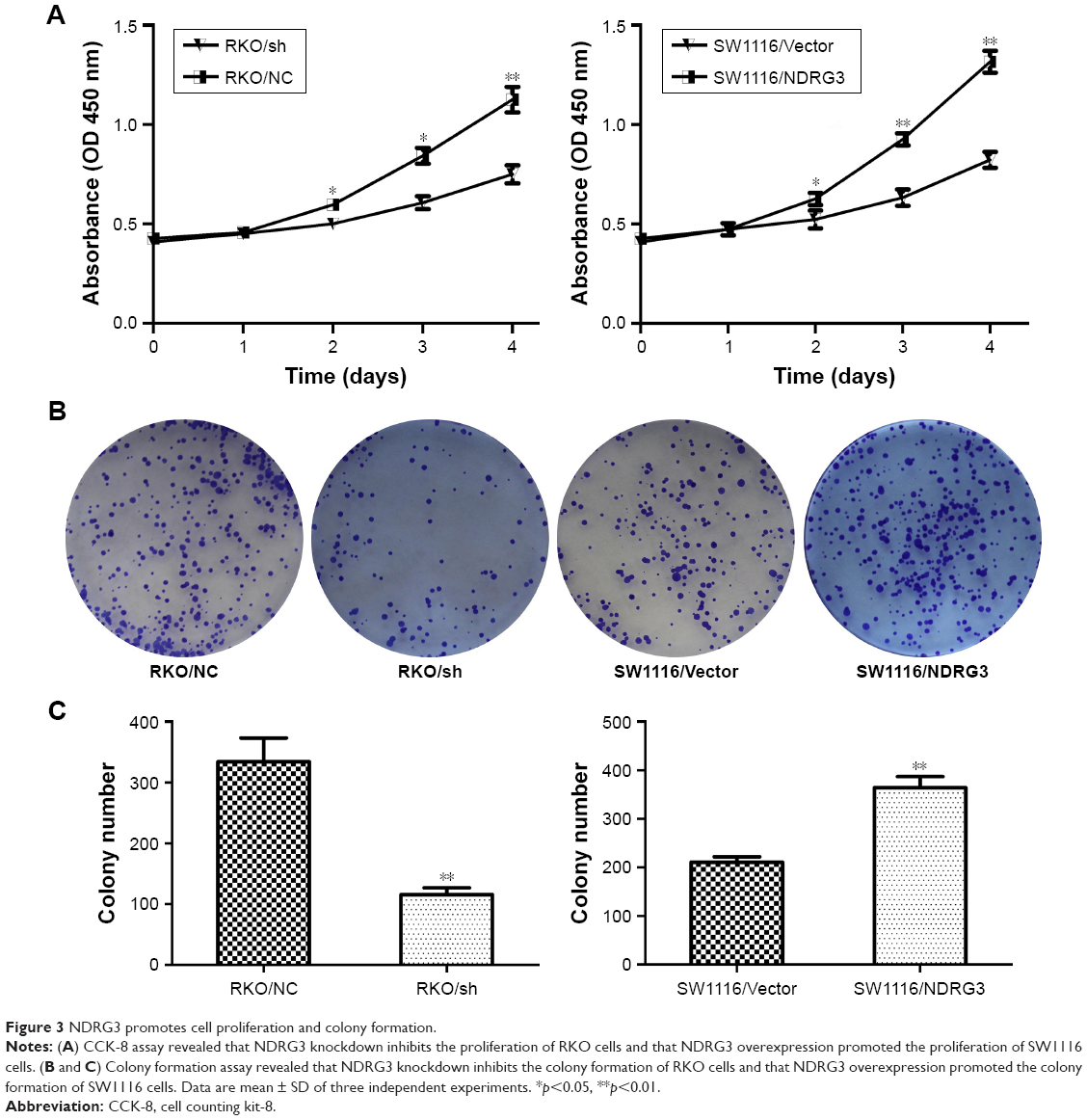 | Figure 3 NDRG3 promotes cell proliferation and colony formation. |
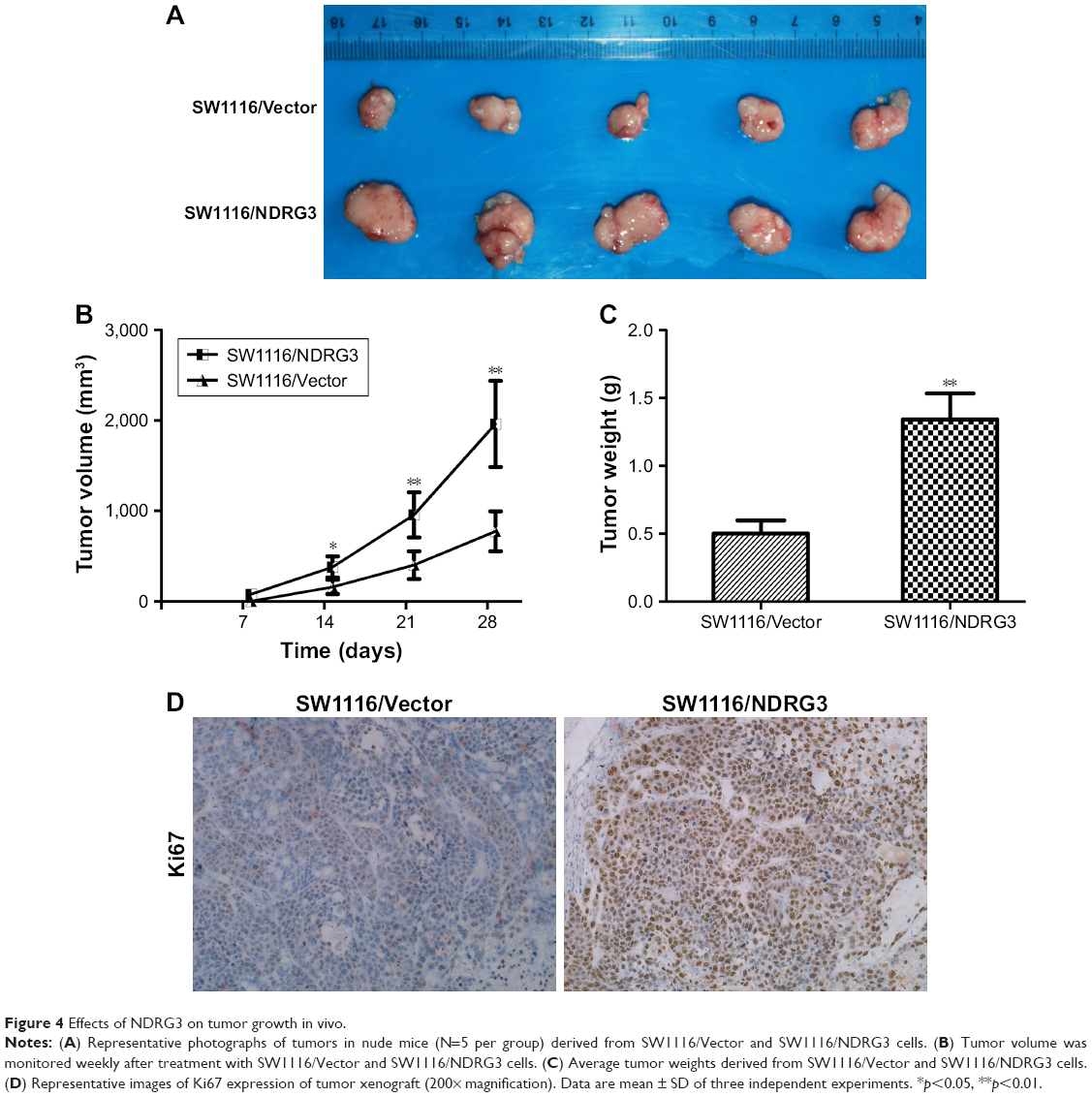 | Figure 4 Effects of NDRG3 on tumor growth in vivo. |
NDRG3 enhances wound healing, migration, and invasion of CRC cells
Wound healing assay was used to evaluate the influence of NDRG3 on cellular migration. After 48 hours of incubation, the distance between wound edges of SW1116/NDRG3 cells was markedly shorter than those of SW1116/Vector cells. In contrast, a longer distance was noticed in NDRG3 knockdown cells RKO compared with the negative control cells. The wound areas of the experimental group and controls were significantly different (Figure 5A–C).
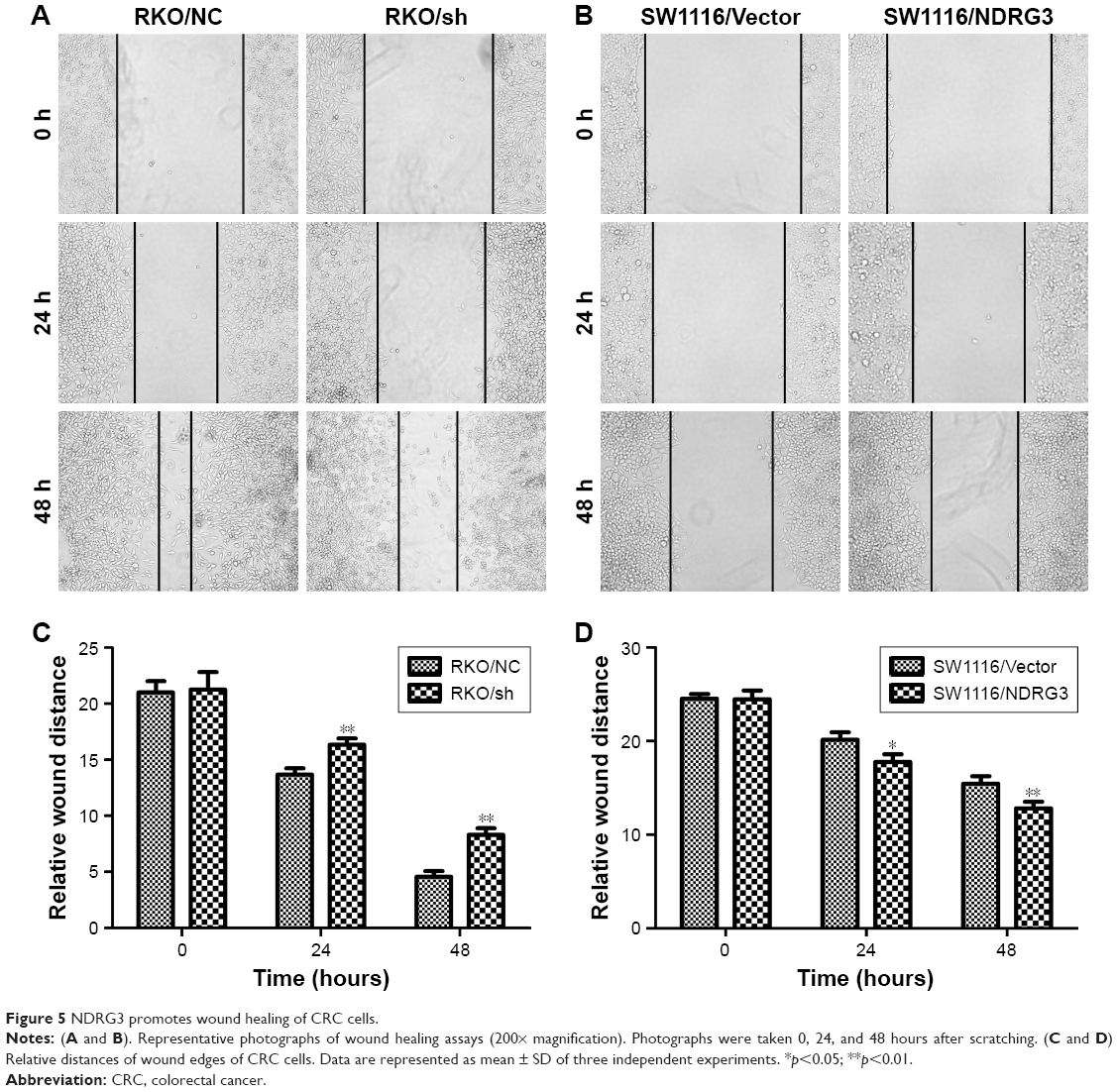 | Figure 5 NDRG3 promotes wound healing of CRC cells. |
Transwell assays were performed to further detect the influence of NDRG3 on cellular migration and invasion. The amount of cells that migrated into the lower chamber was significantly increased in SW1116/NDRG3 group compared with the vector control (Figure 6A and C). In contrast, the RKO/KD group was moderately decreased compared with mock group (Figure 6B and D). Similar data were obtained in invasion assays with SW1116 and RKO cells (Figure 6A–D). Thus, together, these results implicate NDRG3 in the regulation of characteristic cellular behaviors found in CRC.
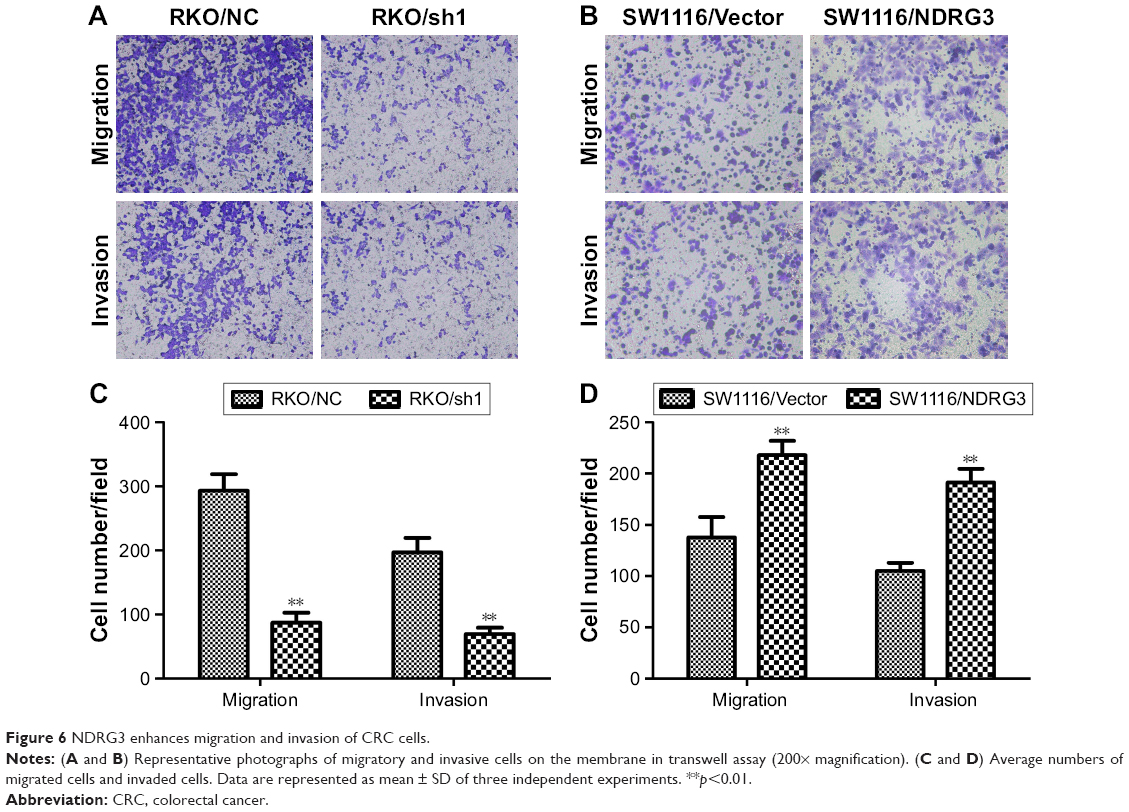 | Figure 6 NDRG3 enhances migration and invasion of CRC cells. |
NDRG3 facilitates tumor metastasis in vivo
To determine the function of NDRG3 in liver metastasis, we applied the portal vein injection model. As shown in Figure 7, there were significantly more visible liver metastasis nodules in mice injected with the SW1116/NDRG3 cells compared with SW1116/Vector control. These data suggested that NDRG3 expression facilitates tumor metastases in vivo. In accordance with these observations, we found that phosphorylation (Tyr 419) of Src was dramatically activated in NDRG3-overexpressing CRC cells compared with the control groups, implicating the crucial role of Src in NDRG3-mediated CRC cell metastasis (Figure 7C).
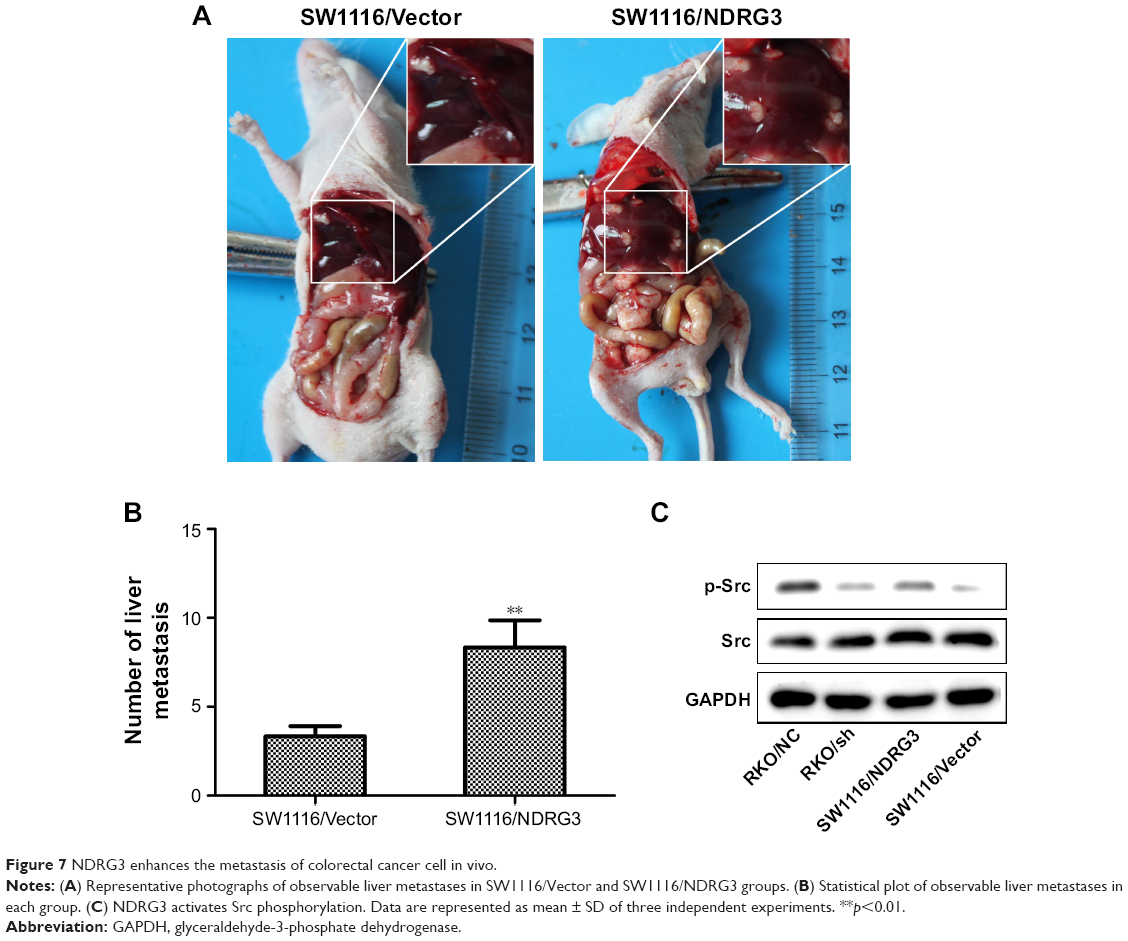 | Figure 7 NDRG3 enhances the metastasis of colorectal cancer cell in vivo. |
Discussion
As an essential oncoprotein family member, myc participates in the transcriptional program driving, including cell proliferation20,21 and inhibiting cell differentiation.22 Myc regulates cell proliferation and differentiation mainly through transcriptional regulation of its target genes.23 Human N-myc down-regulated gene (NDRG) family members have been found intimately involved in cell proliferation and differentiation. The role of NDRG1 in suppressing metastatic progression has been reported in a series of studies.24–26 Liu et al found that NDRG1 attenuates the downstream signaling of c-Src which involves in the p130Cas-CrkIIRac1 pathway, and this leads to suppression of cellular migration.27 Mi et al28 showed that NDRG reduces cav1 expression by promoting its ubiquitylation and degradation via the proteasome in CRC cells. In addition, cav1 mediates the suppressive function of NDRG1 in epithelial–mesenchymal transition, migration, and invasion.28 NDRG3 has been classified in the same subfamily as NDRG1 and found to be expressed in multiple human tissues. Exogenous NDRG3 expression in PC-3 cells enhances tumor growth and increases tumor angiogenesis through upregulating angiogenic chemokines including CXCL1 (chemokine ligand 1), CXCL3 (chemokine ligand 3), and CXCL5 (chemokine ligand 5).17,29 Pan et al found that NDRG3 was tightly associated with meiosis progression in testis and NDRG3 was required for the lactate-induced double-strand break repair via modulating the ERK1/2 pathway.29 A recent study demonstrated that NDRG3 is a hypoxia-inducible lactate sensor and plays key roles in the promotion of hypoxia responses in hypoxia inducible factor-1-independent manner.30 NDRG3 plays a key role in spermatogenesis. NDRG3 is upregulated when Raji cells are treated with Silurus asotus lectin (SAL), suggesting that NDRG3 was regulated by signal transduction through the binding of SAL to globotriaosylceramide (Gb3).31 These previous studies have led to the current investigation. In the present study, we showed that NDRG3 expression is substantially downregulated in CRC tissues compared with adjacent normal tissues. This expression is significantly associated with invasion depth, indicating poor prognosis. These data suggest that NDRG3 may be associated with CRC tumorigenicity and metastasis. We performed CCK-8 and colony formation assays to further determine the importance of NDRG3 in cell proliferation. The results presented in the present study showed that ectopic expression of NDRG3 in the SW1116 cells inhibited cell proliferation and impaired the capability of wound healing, migration, and invasion, while knockdown of NDRG3 in RKO cells produced the opposite results. Xenograft model results supported the experimental outcomes in vitro. Src is a non-receptor tyrosine kinase whose expression and activity are involved in the regulation of oncogenic processes including proliferation, angiogenesis, differentiation, and survival motility.32,33 Upon dephosphorylation of Src at Tyr530 site, the molecular interactions are destabilized, thus resulting in the autophosphorylation of Tyr419. This event allows the opening of SH2 and SH3 domains to interact with G-protein-coupled receptors, receptor tyrosine kinases, and focal adhesion kinase.34–36 Evidence showed that Src orchestrates tumor progression through phosphorylating tyrosine residues mainly acting as the downstream of RTKs and integrins.37 The results presented in the present study show that ectopic NDRG3 in SW1116 cells increased the phosphorylation of Src, while silencing of NDRG3 in RKO cells produced the opposite result. NDRG3 enhances CRC metastasis through the activation of c-Src at Tyr419. However, how NDRG3 regulates Src remains unknown and is worth a next approach.
Conclusion
Our data show that NDRG3 promotes CRC cell proliferation, migration, and invasion, suggesting that NDRG3 may be an oncogene in CRC.
Acknowledgment
This study was supported by Anhui Science and Technology research project (number 1501ID04040).
Disclosure
The authors report no conflicts of interest in this work.
References
Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J, Jemal A. Global cancer statistics, 2012. CA Cancer J Clin. 2015;65(2):87–108. | ||
Favoriti P, Carbone G, Greco M, Pirozzi F, Pirozzi RE, Corcione F. Worldwide burden of colorectal cancer: a review. Updates Surg. 2016;68(1):7–11. | ||
Brenner H, Kloor M, Pox CP. Colorectal cancer. Lancet. 2014;383(9927):1490–1502. | ||
Zhang J, Li F, Liu X, et al. The repression of human differentiation-related gene NDRG2 expression by Myc via Miz-1-dependent interaction with the NDRG2 core promoter. J Biol Chem. 2006;281(51):39159–39168. | ||
Zhou RH, Kokame K, Tsukamoto Y, Yutani C, Kato H, Miyata T. Characterization of the human NDRG gene family: a newly identified member, NDRG4, is specifically expressed in brain and heart. Genomics. 2001;73(1):86–97. | ||
Kokame K, Kato H, Miyata T. Homocysteine-respondent genes in vascular endothelial cells identified by differential display analysis. GRP78/BiP and novel genes. J Biol Chem. 1996;271(47):29659–29665. | ||
Ulrix W, Swinnen JV, Heyns W, Verhoeven G. The differentiation-related gene 1, Drg1, is markedly upregulated by androgens in LNCaP prostatic adenocarcinoma cells. FEBS Lett. 1999;455(1–2):23–26. | ||
Salnikow K, Blagosklonny MV, Ryan H, Johnson R, Costa M. Carcinogenic nickel induces genes involved with hypoxic stress. Cancer Res. 2000;60(1):38–41. | ||
Salnikow K, Su W, Blagosklonny MV, Costa M. Carcinogenic metals induce hypoxia-inducible factor-stimulated transcription by reactive oxygen species-independent mechanism. Cancer Res. 2000;60(13):3375–3378. | ||
Kurdistani SK, Arizti P, Reimer CL, Sugrue MM, Aaronson SA, Lee SW. Inhibition of tumor cell growth by RTP/rit42 and its responsiveness to p53 and DNA damage. Cancer Res. 1998;58(19):4439–4444. | ||
Guan RJ, Ford HL, Fu Y, Li Y, Shaw LM, Pardee AB. Drg-1 as a differentiation-related, putative metastatic suppressor gene in human colon cancer. Cancer Res. 2000;60(3):749–755. | ||
Bandyopadhyay S, Pai SK, Gross SC, et al. The Drg-1 gene suppresses tumor metastasis in prostate cancer. Cancer Res. 2003;63(8):1731–1736. | ||
Stein S, Thomas EK, Herzog B, et al. NDRG1 is necessary for p53-dependent apoptosis. J Biol Chem. 2004;279(47):48930–48940. | ||
Mitchelmore C, Buchmann-Moller S, Rask L, West MJ, Tronscoso JC, Jensen NA. NDRG2: a novel Alzheimer’s disease associated protein. Neurobiol Dis. 2004;16(1):48–58. | ||
Deng Y, Yao L, Chau L, et al. N-Myc downstream-regulated gene 2 (NDRG2) inhibits glioblastoma cell proliferation. Int J Cancer. 2003;106(3):342–347. | ||
Zhao W, Tang R, Huang Y, et al. Cloning and expression pattern of the human NDRG3 gene. Biochim Biophys Acta. 2001;1519(1–2):134–138. | ||
Wang W, Li Y, Li Y, et al. NDRG3 is an androgen regulated and prostate enriched gene that promotes in vitro and in vivo prostate cancer cell growth. Int J Cancer. 2009;124(3):521–530. | ||
Wu H, Jing X, Cheng X, et al. Asporin enhances colorectal cancer metastasis through activating the EGFR/src/cortactin signaling pathway. Oncotarget. 2016;7(45):73402–73413. | ||
Wu H, Cheng X, Ji X, et al. Cortactin contributes to the tumorigenicity of colorectal cancer by promoting cell proliferation. Oncol Rep. 2016;36(6):3497–3503. | ||
Malagolini N, Dall’Olio F, Turrini I, Cessi C, Serafini-Cessi F. Effect of ethanol on human colon carcinoma CaCo-2 and HT-29 cell lines during the maturation process. Alcohol Clin Exp Res. 1994;18(6):1386–1391. | ||
Dimberg A, Bahram F, Karlberg I, Larsson LG, Nilsson K, Oberg F. Retinoic acid-induced cell cycle arrest of human myeloid cell lines is associated with sequential down-regulation of c-Myc and cyclin E and posttranscriptional up-regulation of p27(Kip1). Blood. 2002;99(6):2199–2206. | ||
Ayer DE, Eisenman RN. A switch from Myc:Max to Mad:Max heterocomplexes accompanies monocyte/macrophage differentiation. Genes Dev. 1993;7(11):2110–2119. | ||
Eisenman RN. Deconstructing myc. Genes Dev. 2001;15(16):2023–2030. | ||
Liu W, Iiizumi-Gairani M, Okuda H, et al. KAI1 gene is engaged in NDRG1 gene-mediated metastasis suppression through the ATF3-NFkappaB complex in human prostate cancer. J Biol Chem. 2011;286(21):18949–18959. | ||
Mao Z, Sun J, Feng B, et al. The metastasis suppressor, N-myc downregulated gene 1 (NDRG1), is a prognostic biomarker for human colorectal cancer. PLoS One. 2013;8(7):e68206. | ||
Liu W, Xing F, Iiizumi-Gairani M, et al. N-myc downstream regulated gene 1 modulates Wnt-beta-catenin signalling and pleiotropically suppresses metastasis. EMBO Mol Med. 2012;4(2):93–108. | ||
Liu W, Yue F, Zheng M, et al. The proto-oncogene c-Src and its downstream signaling pathways are inhibited by the metastasis suppressor, NDRG1. Oncotarget. 2015;6(11):8851–8874. | ||
Mi L, Zhu F, Yang X, et al. The metastatic suppressor NDRG1 inhibits EMT, migration and invasion through interaction and promotion of caveolin-1 ubiquitylation in human colorectal cancer cells. Oncogene. 2017;36(30):4323–4335. | ||
Pan H, Zhang X, Jiang H, et al. Ndrg3 gene regulates DSB repair during meiosis through modulation the ERK signal pathway in the male germ cells. Sci Rep. 2017;7:44440. | ||
Park KC, Lee DC, Yeom YI. NDRG3-mediated lactate signaling in hypoxia. BMB Rep. 2015;48(6):301–302. | ||
Kawano T, Sugawara S, Hosono M, Tatsuta T, Nitta K. Alteration of gene expression induced by Silurus asotus lectin in Burkitt’s lymphoma cells. Biol Pharm Bull. 2008;31(5):998–1002. | ||
Guarino M. Src signaling in cancer invasion. J Cell Physiol. 2010;223(1):14–26. | ||
Wheeler DL, Iida M, Dunn EF. The role of Src in solid tumors. Oncologist. 2009;14(7):667–678. | ||
Cooper JA, Gould KL, Cartwright CA, Hunter T. Tyr527 is phosphorylated in pp60c-src: implications for regulation. Science. 1986;231(4744):1431–1434. | ||
Okada M, Nakagawa H. A protein tyrosine kinase involved in regulation of pp60c-src function. J Biol Chem. 1989;264(35):20886–20893. | ||
Nada S, Okada M, MacAuley A, Cooper JA, Nakagawa H. Cloning of a complementary DNA for a protein-tyrosine kinase that specifically phosphorylates a negative regulatory site of p60c-src. Nature. 1991;351(6321):69–72. | ||
Feigelson SW, Grabovsky V, Winter E, et al. The Src kinase p56(lck) up-regulates VLA-4 integrin affinity. Implications for rapid spontaneous and chemokine-triggered T cell adhesion to VCAM-1 and fibronectin. J Biol Chem. 2001;276(17):13891–13901. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.