Back to Journals » Infection and Drug Resistance » Volume 13
NDM-1 Carbapenemase-Producing Enterobacteriaceae are Highly Susceptible to Ceragenins CSA-13, CSA-44, and CSA-131
Authors Chmielewska SJ, Skłodowski K, Piktel E, Suprewicz Ł ![]() , Fiedoruk K
, Fiedoruk K ![]() , Daniluk T, Wolak P
, Daniluk T, Wolak P ![]() , Savage PB
, Savage PB ![]() , Bucki R
, Bucki R ![]()
Received 12 May 2020
Accepted for publication 28 July 2020
Published 28 September 2020 Volume 2020:13 Pages 3277—3294
DOI https://doi.org/10.2147/IDR.S261579
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Suresh Antony
Sylwia Joanna Chmielewska,1 Karol Skłodowski,1 Ewelina Piktel,1 Łukasz Suprewicz,1 Krzysztof Fiedoruk,1 Tamara Daniluk,1 Przemysław Wolak,2 Paul B Savage,3 Robert Bucki1
1Department of Medical Microbiology and Nanobiomedical Engineering, Medical University of Białystok, Białystok, Poland; 2The Faculty of Medicine and Health Sciences of the Jan Kochanowski University in Kielce, Kielce, Poland; 3Department of Chemistry and Biochemistry, Brigham Young University, Provo, UT, USA
Correspondence: Robert Bucki
Department of Medical Microbiology and Nanobiomedical Engineering, Medical University of Białystok, Mickiewicza 2C, Białystok 15-222, Poland
Tel +48 85 748 5493
Fax +48 85 748 5416
Email [email protected]
Background and Purpose: Treatment of infections caused by NDM-1 carbapenemase-producing Enterobacteriaceae (CPE) represents one of the major challenges of modern medicine. In order to address this issue, we tested ceragenins (CSAs – cationic steroid antimicrobials) as promising agents to eradicate various NDM-1-producing Gram-negative enteric rods.
Materials and Methods: Susceptibility to CSA-13, CSA-44, and CSA-131 of four reference NDM-1 carbapenemase-producing strains, ie, Escherichia coli BAA-2471, Enterobacter cloacae BAA-2468, Klebsiella pneumoniae subsp. pneumoniae BAA-2472, and K. pneumoniae BAA-2473 was assessed by MIC/MBC testing of planktonic cells as well as biofilm formation/disruption assays. To define the mechanism of CSAs bactericidal activity, their ability to induce generation of reactive oxygen species (ROS), permeabilization of the inner and outer membranes, and their mechanical and adhesive properties upon CSA addition were examined. Additionally, hemolytic assays were performed to assess CSAs hemocompatibility.
Results: All tested CSAs exert substantial bactericidal activity against NDM-1-producing bacteria. Moreover, CSAs significantly prevent biofilm formation as well as reduce the mass of developed biofilms. The mechanism of CSA action comprises both increased permeability of the outer and inner membrane, which is associated with an extensive ROS generation. Additionally, atomic force microscopy (AFM) analysis has shown morphological alterations in bacterial cells and the reduction of stiffness and adhesion properties. Importantly, CSAs are characterized by low hemolytic activity at concentrations that are bactericidal.
Conclusion: Development of ceragenins should be viewed as one of the valid strategies to provide new treatment options against infections associated with CPE. The studies presented herein demonstrate that NDM-1-positive bacteria are more susceptible to ceragenins than to conventional antibiotics. In effect, CSA-13, CSA-44, and CSA-131 may be favorable for prevention and decrease of global burden of CPE.
Keywords: antibiotic resistance, NDM-1, CSA-13, CSA-44, CSA-131, ceragenins, CPE
Introduction
The discovery of antibiotics was a remarkable moment in the history of mankind, which revolutionized medicine by saving the lives of previously incurable patients suffering from infectious disease.1 Nevertheless, the extraordinary advantages of these drugs in reducing morbidity and mortality have been limited by the appearance of resistant bacteria.2 The rapid rise and dissemination of multiple types of antimicrobial-resistant (MDR) bacteria have posed a crucial threat to global public health.3 Importantly, the number of new antibiotics registries is considerably limited.4 Nowadays, the advent of resistance among pathogenic bacteria is imperiling the utility of the first-line antibiotics. Currently, carbapenems, such as imipenem, meropenem, and ertapenem, are recommended as the most effective in treatment of serious infections caused by MDR bacteria.3,5 Unfortunately, the indiscriminate, injudicious use of carbapenems has led to the appearance of bacteria resistant to these last-resort antibiotics.1 The consequences of this phenomenon increase the risk of prolonged illness, a greater mortality, and economic losses.1
Numerous organizations, such as the Centers for Disease Control and Prevention (CDC) and the European Centre for Disease Prevention (ECDC), are warning against the return to the pre-antibiotic era.1 It is estimated that annually, in the United States, more than 2.8 million antibiotic-resistant infections occur, resulting in more than 35,000 deaths. In 2019, the CDC published a list describing the microorganisms that were considered urgent threats. This list was headlined by carbapenem- producing or -resistant Enterobacteriaceae (CPE, CRE), that are classified by WHO as “critical” pathogens for research and development of new antibiotics.6 Moreover, it should be highlighted that in 2017, more than 13,000 cases of healthcare-associated CPE infections were reported in the USA and at least 1,100 people died.7 Additionally, analysis of ECDC reports concludes that the highest percentage of carbapenem-producing (CP) Klebsiella pneumoniae strains was detected in Greece (63.9%); followed by Romania (29.5%) and Italy (26.8%).8 Despite ongoing control efforts to combat the CPE, so far no significant results have been achieved.1,9 Regrettably, recent predictions point out that bacterial infections will result in 10 million annual deaths by 2050, more than those presently induced by cancer.10
Nowadays, one of the most clinically and epidemiologically important mechanisms of resistance to carbapenems is NDM-1 (New Delhi Metallo-β-lactamase-1). The first bacterium harboring blaNDM-1 was K. pneumoniae isolated in 2008 from a 59-year-old Swedish man who was previously hospitalized in India. It should be emphasized that, since then, strains producing NDM-1 carbapenemases have been revealed in almost all countries and in most species among Enterobacterales bacteria threatening epidemic outbreaks.3,11-13 In effect, NDM-1 mediated resistance has become one of the most meaningful current problems in the field of drug resistance.13 It should be noted that bacteria harboring NDM-1 are not only resistant to β-lactams, but also to another groups of antibiotics, hence frequently are classified as pandrug-resistant (PDR) pathogens.13 To date, the treatment options for NDM-1 infections have remained very limited, with colistin, tigecycline, fosfomycin, aminoglycosides, as well as ciprofloxacin as the basic therapy.11 Unfortunately, the effective use of these agents is undermined by their significant toxicity and the increasing number of resistant strains.1 Consequently, there is an imperative and urgent need to introduce new antimicrobial therapeutic strategies.14
Ceragenins have been viewed as one of the valid solutions to the impending crisis of antimicrobial resistance.15 The molecular architecture of ceragenins is founded on a cholic acid, which is appended by amine groups, therefore ceragenins are also known as cationic steroid antimicrobials (CSAs).16 Describing in more detail, CSAs demonstrate positive charges arranged on one face and hydrophobic residues on the other.17 The mechanism of the bactericidal action proposed for these compounds comprises an electrostatic connection between ceragenins and the negatively charged molecules of the bacterial surface. This association enables the further direct and rapid antimicrobial activity of CSAs resulting in the microbial membrane insertion, ultimately leading to changes in the membrane organization of phospholipids and sudden membrane depolarization.16 This affinity of CSAs for bacterial membrane components is due to the cell selectivity. The recent data showed that the study compounds selectively associate with prokaryotes over eukaryotic cells, which possibly arises from the higher net of the negative charge present on prokaryotic membranes.18
The therapeutic appropriateness of ceragenins has been emphasized by multiple publications.16 CSAs are assumed to be a promising alternative to conventional antibiotics due to a broad spectrum of their activity against potent microbial pathogens, such as Gram-positive and Gram-negative bacteria as well as fungi and, importantly, MDR strains.15,16,19 What is more, the development of resistance to CSAs is considerably lower compared to conventional antibiotics.16 So far, no bacteria have accomplished high levels of resistance to ceragenins.20 Apart from antibacterial effects, these molecules also display antibiofilm, sporicidal, antiviral, antiparasitic, and anticancer activities.21–24 The therapeutic potential of CSAs is also underlined by their low toxicity, good stability under physiological conditions, and relatively inexpensive production.20,25 Because of promising therapeutic properties of CSAs, we investigated antibacterial features of CSA-13, CSA-44, and CSA-131 against rods belonging to Enterobacteriaceae family producing NDM-1.
Materials and Methods
Bacterial Strains and Tested Compounds
Four NDM-1 carbapenemase-producing strains: Escherichia coli BAA-2471, Enterobacter cloacae BAA-2468, Klebsiella pneumoniae subsp. pneumoniae BAA-2472, and K. pneumoniae BAA-2473 were purchased from American Type Culture Collection (ATCC, USA). E. coli BAA-2471 and K. pneumoniae subsp. pneumoniae BAA-2472 were isolated from the respiratory sample, whereas E. cloacae BAA-2468 as well as K. pneumoniae BAA-2473 from the human urine.
Ceragenins: CSA-13, CSA-44, and CSA-131 were synthesized as described previously.18 The chemical structures of CSAs are shown in Figure 1. In the next step the prepared CSAs were dissolved in PBS (phosphate-buffered saline, Thermo Fisher Scientific, USA). Then, the suspension was sonicated using a washer IS-4 (InterSonic) for 30 minutes at room temperature, before the investigation. Finally, appropriate CSA concentrations were used for the tests. In turn, antibiotics (ertapenem, imipenem, colistin, gentamicin, tobramycin, ciprofloxacin, tigecycline, and fosfomycin) were acquired from Sigma-Aldrich (USA), except meropenem (AstraZeneca, Poland).
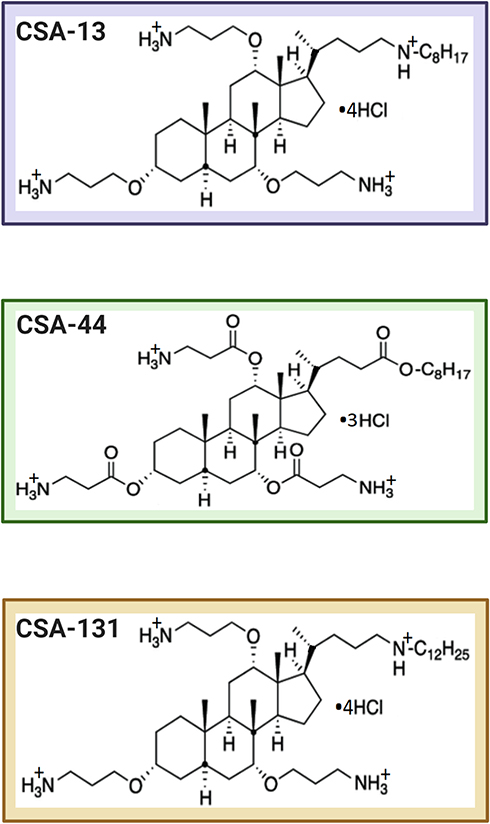 |
Figure 1 The chemical structures of tested CSAs. |
MIC/MBC Assay
Minimal inhibitory concentrations (MICs) and minimal bactericidal concentrations (MBCs) of the tested compounds against E. coli BAA-2471, E. cloacae BAA-2468, K. pneumoniae subsp. pneumoniae BAA-2472, and K. pneumoniae BAA-2473 were determined using the serial microdilution method in Mueller-Hinton broth (Sigma-Aldrich, USA). The MIC values for the antibiotics were interpreted in accordance with the European Committee on Antimicrobial Susceptibility Testing (EUCAST). MBC values were determined by plating each sample (10 μL) on MacConkey agar (Biomaxima, Poland) and defined as the lowest concentration of the compounds studied resulting in at least a 99.9% killing of the initial bacterial inoculum.
Killing Assay
To determine the bactericidal activities of CSA-13, CSA-44, and CSA-131 against the strains producing NDM-1, a killing assay was performed. Individual colonies of bacteria were suspended to ~108 CFU/mL and serially diluted to 105 CFU/mL with sterile PBS. The assays were performed using ceragenins within the concentration range 0.1–0.9 μg/mL. After 60 minutes of incubation at 37°C, samples were diluted from 10–1000-fold. Thereafter, 10 μL aliquots of each dilution were plated on the MacConkey agar and incubated overnight at 37°C to establish the number of viable colonies. Colony-forming units (CFU/mL) of the individual samples were determined from the dilution factor.
Prevention of Biofilm Formation
Formation of biofilms by NDM-positive strains in the presence of different concentrations of CSAs (1–100 μg/mL) was assessed using the resazurin-based fluorimetric method. After incubation of bacterial samples with ceragenins (37°C for 24, 48, and 72 hours), the plates were washed 3-times with PBS to remove unattached bacteria and stained using resazurin (Sigma-Aldrich, USA) in a final concentration of 0.2 mg/mL per well. After that, fluorescence intensity was measured (λex=520 nm/λem=590 nm) using Labsystem Varioscan Lux (Thermo Fisher Scientific, USA).
Disruption of Established Biofilms
Biofilms of the studied bacterial strains were grown in 96-well plates at 37°C for 24, 48, and 72 hours in LB broth (LB, Bio Corp, USA). After incubation, the plates were washed with PBS to remove planktonic cells. Next CSA-13, CSA-44, and CSA-131 were added in varied concentrations (1–100 μg/mL). After 1 hour incubation, plates were washed with PBS and biofilms were removed from plates by sonication using a washer IS-4 (InterSonic) for 15 minutes. After that, samples were diluted 10–1000-fold and 10 μL aliquots of each dilution were plated on MacConkey agar and incubated overnight at 37°C to determine the number of viable colonies. The CFU/mL of the individual samples was defined from the dilution factor.
Outer Membrane Permeabilization Assay
N-phenyl-1-napthylamine (NPN, Sigma-Aldrich, USA) uptake assay was used to assess the outer membrane permeability of NDM-1 producing strains. Bacterial cells were resuspended in PBS (OD600=0.1) prior to incubation with CSA-13, CSA-44, and CSA-131 in concentrations ranging from 1–10 µg/mL. NPN was added to a final concentration of 0.5 mM, and the mixture was incubated for 5 minutes. Fluorescence (λex=348 nm/λem=408 nm) was measured using Labsystem Varioscan Lux.
Inner Membrane Permeabilization Assay
To evaluate β-galactosidase activity, o-nitrophenyl-β-
ROS Generation Assessment
Ceragenin-induced generation of ROS was measured using 2′,7′-dichlorofluorescein diacetate (DFCH-DA, Sigma-Aldrich, USA) as a fluorescent probe. Bacterial cells (OD600=0.1) were pipetted into 96-well black plates. Then CSA-13, CSA-44, and CSA-131 in the concentrations ranging from 1–10 µg/mL were added to each well. After, DFCH-DA in PBS at 20 µM was prepared and mixed. Fluorescence was measured for 60 minutes immediately after addition of the dye at excitation/emission wavelengths of 488/535 nm.
The Atomic Force Microscope Imaging
Characterization of mechanical properties of Klebsiella pneumoniae BAA-2473 cells untreated and treated with 1 μg/mL, 5 μg/mL, and 10 μg/mL of CSA-13, CSA-44, and CSA-131 were performed using an atomic force microscope NanoWizard 4 BioScience AFM (JPK Instruments, Bruker) equipped with a liquid cell setup. Silicon Nitride cantilevers (Bruker MSCT) described by a spring constant of 0.37 N/m were used. Due to the lateral forces during contact mode scanning, the force curves-based imaging mode with the resolution of 128 pixels per line was used, to image bacterial surfaces (JPK QI™ mode - Quantitative Imaging). The topography maps sizes of 5 μm×5 μm and 3 μm×3 μm were recorded. To assess the wider spectrum of cells changes after treatment, QI maps were used to determine bacteria surface stiffness (a slope mode) and adhesion forces between the cells and the AFM probe.
Hemolytic Activity
Hemolytic activity of CSA-13, CSA-44, and CSA-131 was determined using human red blood cells (RBCs) suspended in PBS hematocrit ~5%. RBCs were incubated for 1, 6, and 12 hours at 37°C after addition of the compounds in the concentration of 1–100 μg/mL. To evaluate the relative hemoglobin concentration in supernatants after centrifugation at 2000 g, the optical absorbance was measured at 540 nm (Labsystem Varioscan Lux). Simultaneously, the positive control (100% of hemolysis) was taken from the wells in which 1% Triton X-100 (Sigma-Aldrich, USA) was used to disrupt the cell membrane, whereas RBCs in PBS were used as a negative control. The relative absorbance compared to that treated with 1% Triton X-100 was defined as the percentage of hemolysis.
Statistical Analysis
All statistical analyses were conducted using Graph Pad Prism, version 8 (San Diego, CA). Collected data are reported as the mean±standard deviation (SD) of three-to-six experiments. One-tailed Student’s t-test was used to compare differences between all referential strains, and a P-value<0.05 was considered to be statistically significant.
Results
Ceragenins CSA-13, CSA-44, and CSA-131 Exert Antibacterial Activity Against NDM-1 Producing Strains
According to EUCAST criteria, E. coli BAA-2471 belongs to the MDR bacteria category, whereas E. cloacae BAA 2468, K. pneumoniae subsp. pneumoniae BAA 2472, and K. pneumoniae BAA 2473 are classified as XDR (extensively drug resistant) strains due to the resistance to almost all antibiotics used. As shown in Table 1, CSA-13, CSA-44, and CSA-131 possess a significantly higher activity with MIC values ranging from 1–2 μg/mL than gentamicin (MIC >256 μg/mL), tobramycin (MIC 32 >256 μg/mL), ciprofloxacin (MIC 64 >256 μg/mL) or fosfomycin (MIC ≥256 μg/mL, except E. coli BAA 2471).
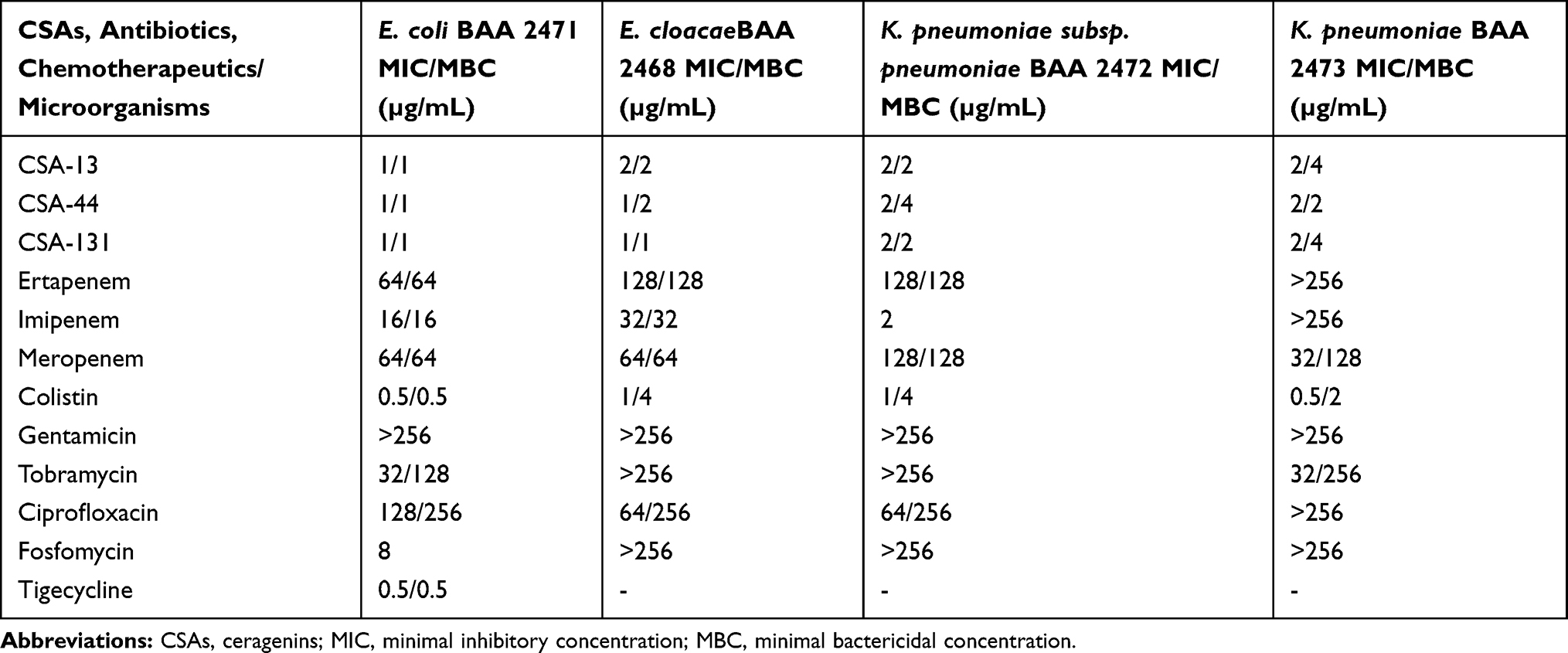 |
Table 1 MIC and MBC Values of CSA-13, CSA-44, CSA-131, and Antibiotics/Chemotherapeutics Tested Against Escherichia coli BAA 2471, Enterobacter cloacae BAA 2468, Klebsiella pneumoniae subsp. pneumoniae BAA 2472, and Klebsiella pneumoniae BAA 2473 |
Furthermore, CSA-13 displays the most prominent bactericidal effect against a planktonic form of all strains (Figure 2A–D). Bacterial growth inhibition was reported at 0.1 μg/mL; 0.3 μg/mL; 0.2 μg/mL, and 0.4 μg/mL in the case of E. coli BAA 2471 (A); E. cloacae BAA 2468 (B); K. pneumoniae subsp. pneumoniae BAA 2472 (C); and K. pneumoniae BAA 2473 (D), respectively. Equally high antibacterial activity was also noted for CSA-44 and CSA-131, since bactericidal doses of ceragenins mentioned were not higher than 0.8 μg/mL. Interestingly, the bactericidal activity of the ceragenins tested was not dependent on the NDM-1 producing strain.
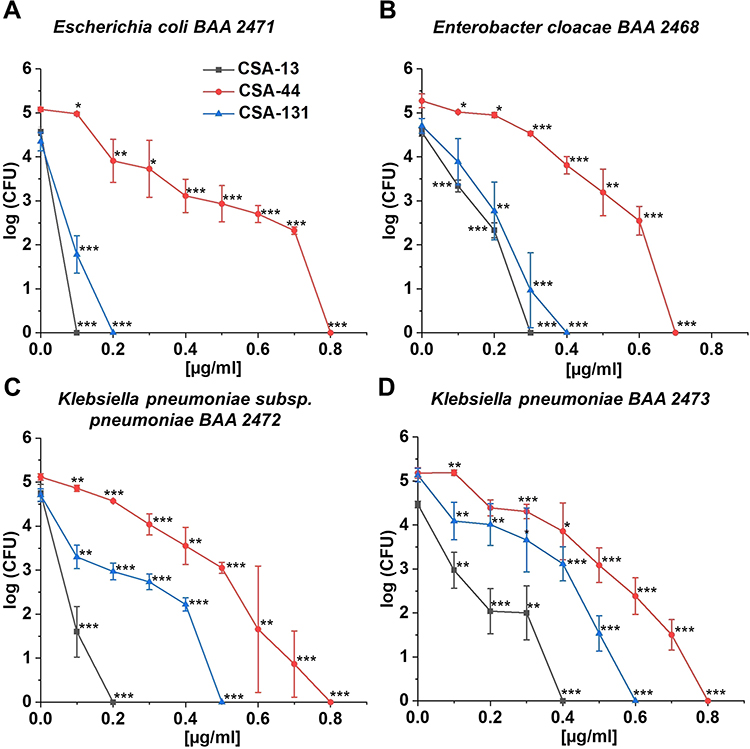 |
Figure 2 Bactericidal activity of CSA-13 (black squares), CSA-44 (red circles), and CSA-131 (blue triangles) against Escherichia coli BAA 2471 (A), Enterobacter cloacae BAA 2468 (B), Klebsiella pneumoniae subsp. pneumoniae BAA 2472 (C), and Klebsiella pneumoniae BAA 2473 (D). Killing activities of CSAs studied at 1–100 µg/mL was determined using a standard colony counting assay. Results show mean±SD from six measurements. * indicates statistical significance at ≤0.05, ** ≤0.01, and *** ≤0.001. |
Ceragenins CSA-13, CSA-44, and CSA-131 Prevent Biofilm Formation by NDM-1-Producing Strains and Disrupt Biofilms Formed
All of the ceragenins tested inhibited the formation of both young and mature biofilms, at 24–72 hours (Figure 3A–D), in a concentration-dependent manner. CSA-13 significantly inhibited biofilm formation by E. coli BAA 2471 (Figure 3A), whereas CSA-44 effectively prevented biofilm formation by E. cloacae BAA 2468 (Figure 3B). Additionally, relevant antibiofilm activities of CSA-131 were determined against K. pneumoniae subsp. pneumoniae BAA 2472 (Figure 3C) and K. pneumoniae BAA 2473 (Figure 3D).
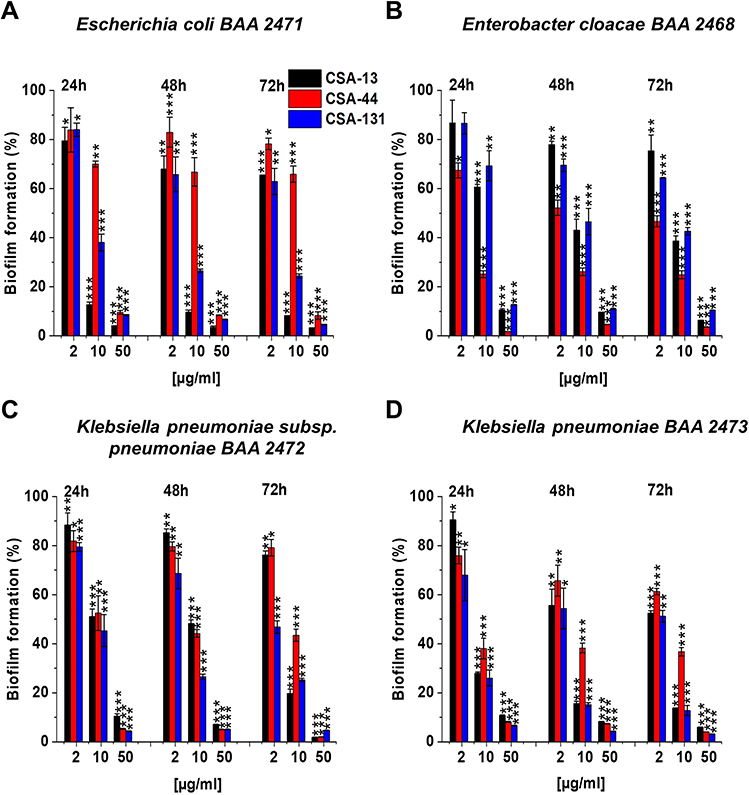 |
Figure 3 Prevention of biofilm formation by Escherichia coli BAA 2471 (A), Enterobacter cloacae BAA 2468 (B), Klebsiella pneumoniae subsp. pneumoniae BAA 2472 (C), and Klebsiella pneumoniae BAA 2473 (D) during treatment with CSA-13, CSA-44, and CSA-131 compounds. Formation of biofilm in the presence of CSAs ranging from 1–100 μg/mL was assessed using the resazurin-based fluorimetric method after 24, 48, and 72 hours incubation. Results show mean±SD from 3–6 measurements. * indicates statistical significance ≤0.05, ** ≤0.01, and *** ≤0.001. |
To assess biofilm survival of NDM-1 producing strains in the presence of CSAs, the killing assay method was performed. As shown in Figure 4A–L CSA-44 exhibited the highest potential to efficiently eliminate bacteria embedded in the biofilm. Nevertheless, CSA-13 and CSA-131 have also been found to possess the ability to disrupt mature biofilms (Figure 4A–L). These results suggest that CSAs should be considered as a good candidate for inhibition of biofilm formation as well as reduction of the mature biofilm.
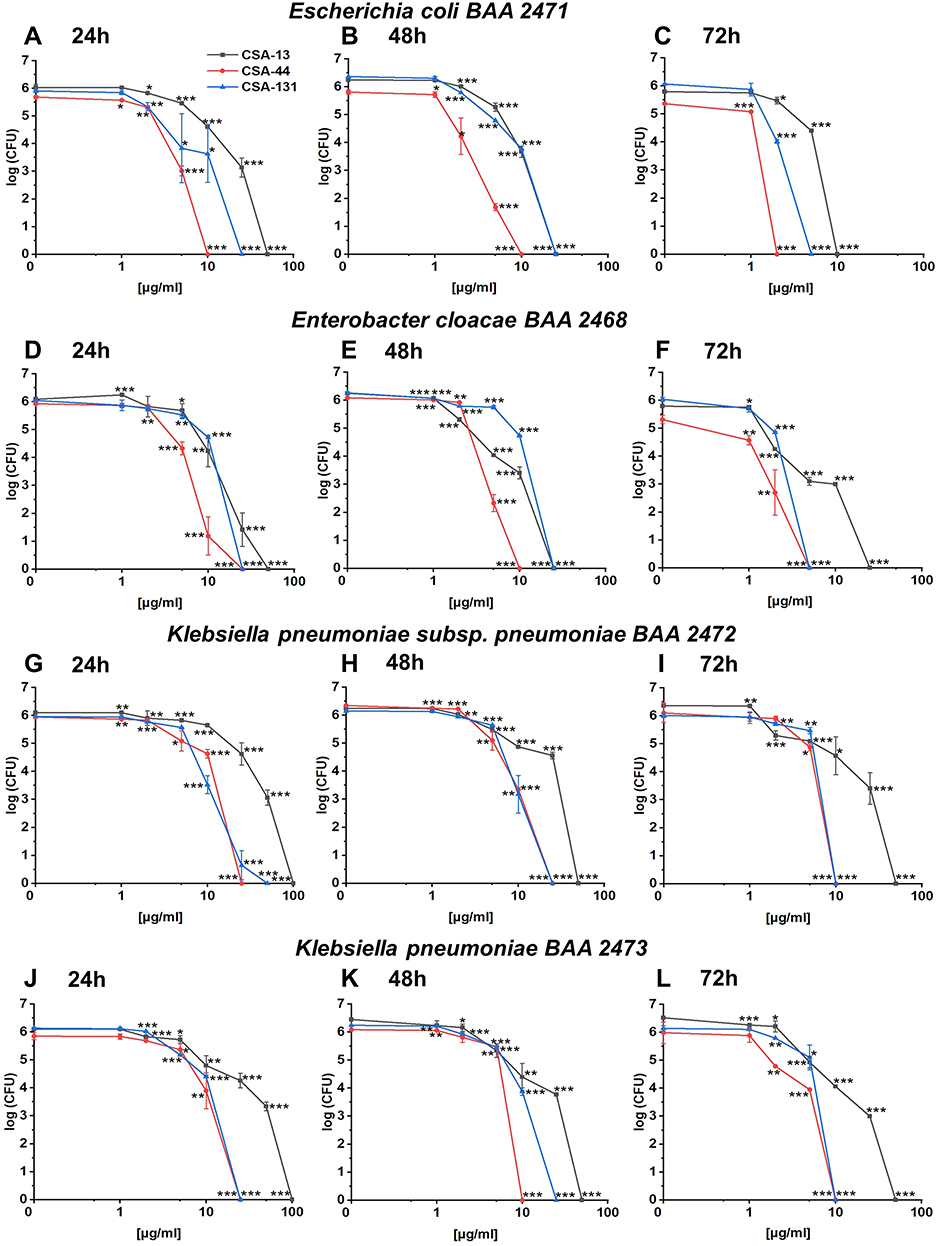 |
Figure 4 Disruption of the biofilms formed by Escherichia coli BAA 2471 (A–C), Enterobacter cloacae BAA 2468 (D–F), Klebsiella pneumoniae subsp. pneumoniae BAA 2472 (G–I), and Klebsiella pneumoniae BAA 2473 (J–L) using CSA-13, CSA-44, and CSA-131 compounds. Reduction of the biofilm formed at 24, 48, and 72 hours incubation with CSAs in concentration of 1–100 µg/mL was performed using killing assay method. Results show mean±SD from 3–6 measurements. * indicates statistical significance ≤0.05, ** ≤0.01, and *** ≤0.001. |
Ceragenins CSA-13, CSA-44, and CSA-131 Cause Disruption of the Inner and Outer Bacterial Membrane Followed by Increased ROS Generation
The CSAs mechanism of action involved interaction with targeted cell membranes, and permeabilization of the outer and inner membrane. Those initial step enabling the cell death.26 The ability of CSAs to disrupt the outer and inner membrane of Gram-negative rods producing NDM-1 was determined using the NPN uptake assay or ONPG hydrolysis assays. The outer membrane (OM) of Gram-negative bacteria is considered to be relatively impervious to different hydrophobic chemicals, including NPN. Nevertheless, some permeabilizing agents have been recognized to alter the uptake of substances like NPN, thus making them an effective indicator for studying the cell permeability. Because NPN is hydrophobic it cannot penetrate intact membranes, and therefore exhibits weak fluorescence emission. However, in the case of damage to the outer membrane, enhanced uptake of NPN as well as increased fluorescence can be observed.27
Data presented in Figure 5A–D indicate that all CSAs studied were able to permeabilize the outer bacterial membrane in a dose-dependent manner. It is notable that CSA-131 at doses of 1–10 μg/mL caused a 1.5–2-fold increase in the outer membrane disruption compared to the control (Figure 5A–D). Among the compounds tested, CSA-131 displayed the strongest interaction with K. pneumoniae subsp. pneumoniae BAA 2472 outer membrane (Figure 5C), even 3-fold compared to the control (at concentration of 5–10 μg/mL).
 |
Figure 5 Disruption of the outer membrane of Escherichia coli BAA 2471 (A), Enterobacter cloacae BAA 2468 (B), Klebsiella pneumoniae subsp. pneumoniae BAA 2472 (C), and Klebsiella pneumoniae BAA 2473 (D) upon treatment with CSA-13, CSA-44, and CSA-131 at doses of 1–10 μg/mL was investigated using NPN uptake assay. Results show mean±SD, n=3; * indicates statistical significance ≤0.05, ** ≤0.01, and *** ≤0.001. |
On the other hand, to measure the permeabilization of the inner bacterial membrane, ONPG hydrolysis assays were performed (Figure 6A–D). The principle of this assay is based on detection of β-galactosidase catalytic activity. In this assay, traversal of the cytoplasmic membrane by ONPG (colorless) is measured by detection of the hydrolysis product: yellow ortho-nitrophenol (ONP). It should be noted that ONPG has to first pass the outer membrane through the non-specific porins. The appearance of a yellow color as a result of the enzymatic hydrolysis of ONPG is used to monitor the permeabilization of the bacterial inner membrane.27,28
 |
Figure 6 Permeability of the inner bacterial membrane of Escherichia coli BAA 2471 (A), Enterobacter cloacae BAA 2468 (B), Klebsiella pneumoniae subsp. pneumoniae BAA 2472 (C), and Klebsiella pneumoniae BAA 2473 (D) was assessed using ONPG hydrolysis assay. The insertion of CSA-13, CSA-44, and CSA-131 ranging 1–10 μg/mL into the bacterial membrane was monitored by the enhancement of β-galactosidase activity. Results show: mean±SD, n=3; * indicates statistical significance ≤0.05, ** ≤0.01, and *** ≤0.001. |
According to our study, CSA-131 exerted the greatest disruption of the inner membrane integrity in E. coli BAA 2471 (Figure 6A), K. pneumoniae subsp. pneumoniae BAA 2472 (Figure 6C), and K. pneumoniae BAA 2473 (Figure 6D). However, insignificant differences between individual ceragenins were observed with K. pneumoniae subsp. pneumoniae BAA 2472 (Figure 6C) as well as K. pneumoniae BAA 2473 strains (Figure 6D). With regard to CSA-131 (Figures 6A–D), over 1- to approximately 4-fold enhancement of permeabilization of the inner membrane was observed at doses ranging from 1–10 μg/mL compared to the control. The data obtained for E. cloacae BAA 2468 (Figure 6B) indicated that, in turn, the strongest β-galactosidase activity was detected in the case of CSA-44 (from over twice to approximately 3-times compared to the control at a concentration of 2–10 μg/mL).
Reactive oxygen species (ROS) have always played a critical role in the bacterial response to the lethal stress. To investigate the induction of ROS by NDM-1 producing strains, DFCH-DA-utilizing fluorimetric assay was used. Results presented in Figure 7A–D show that CSAs contribute to generation of ROS in a dose-dependent manner. Interestingly, the highest production of ROS at a concentration of 1–5 μg/mL was observed upon CSA-44 treatment; ie, more than a 1- to a roughly 3-fold increase in oxidative damage was observed, as compared to the control. Surprisingly, addition of CSA-131 at doses 10 μg/mL resulted in even >10 to >12-fold potentiation of ROS generation among all strains producing carbapenemases (Figure 7A–D). Based on the results collected it is important to point out that all CSAs can cause disruption of the inner and outer bacterial membrane and affect the redox status by increasing ROS formation.
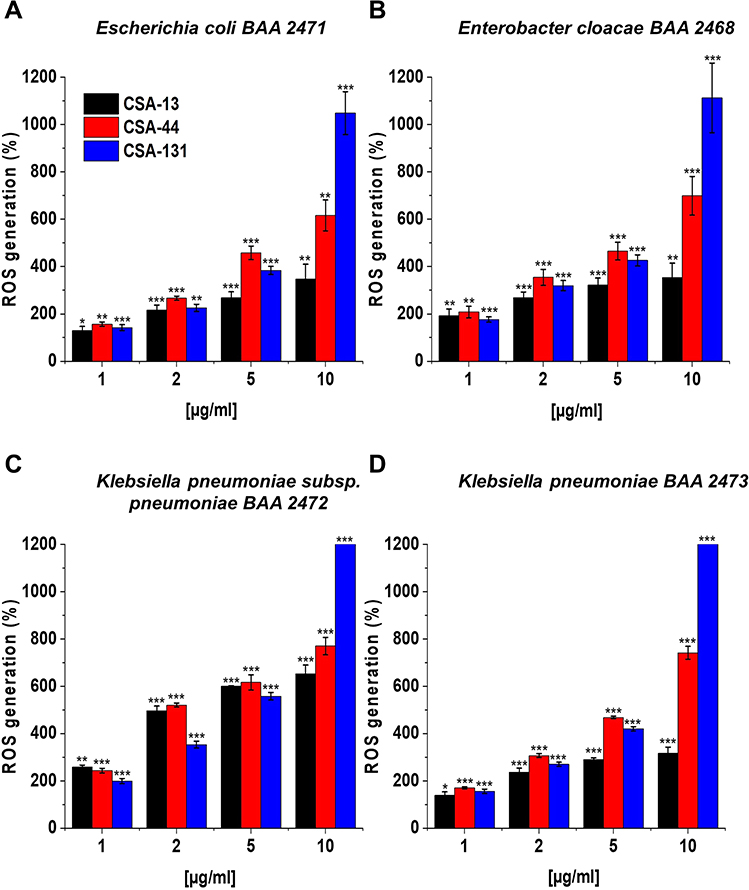 |
Figure 7 Induction of reactive oxygen species (ROS) generation by Escherichia coli BAA 2471 (A), Enterobacter cloacae BAA 2468 (B), Klebsiella pneumoniae subsp. pneumoniae BAA 2472 (C), and Klebsiella pneumoniae BAA 2473 (D) was evaluated by DFCH-DA fluorimetric assay. Formation of ROS upon treatment with CSA-13, CSA-44, and CSA-131 at a concentration of 1–10 μg/mL was presented. Results show: mean±SD, n=3; * indicates statistical significance ≤0.05, ** ≤0.01, and *** ≤0.001. |
Ceragenins CSA-13, CSA-44, and CSA-131 Affect the Morphology, Mechanical, and Adhesive Properties of Bacterial Cells
Visualization of the morphological alteration of K. pneumoniae BAA-2473 cells subjected to incubation with ceragenins (CSA-13, CSA-44, and CSA-131), using AFM (Figure 8), was performed to elaborate on the mechanisms leading to ceragenins-mediated membrane destructions. Figure 9 presents the representative morphology of bacterial cell changes. The analysis of the results indicates that the mechanism of ceragenins action might involve ceragenins interaction with the negatively charged bacterial membrane molecules such as lipopolysaccharide and phosphatidylglycerol. Such interaction results in membrane molecules rearrangement leading to membrane disruption.29 Figure 9A1–I1 and A3–I3 present the bacterial changes in response to CSA-13 and CSA-131 treatment, respectively. Extensive morphological alterations in bacterial cells, ie, microcracks and surface wrinkling, were observed compared to the untreated cells. The shape of the K. pneumoniae BAA-2473 became more oval and the surface of wrinkling was observed, especially in higher CSAs concentrations. The stiffness of the bacterial surface and the adhesion force also decreased markedly upon CSAs addition. In the case of CSA-13, stiffness decreased by 25% and adhesion by 57% compared to the untreated cells. A similar effect was observed upon CSA-131 treatment (reduction of stiffness by 35% and adhesion by 57%). These results are consistent with the observed changes in the shape of cells and leakage of their contents, indicating that changes in the organization of membrane lipids may also be associated with changes in the mechanical properties of bacterial surfaces. Subsequently, morphological changes of K. pneumoniae BAA-2473 after CSA-44 addition have been shown in Figure 9A2–I2. It is worth highlighting that the changes observed are less pronounced compared to CSA-13 and CSA-131. Significant differences in the shape (microcracks and bacteria surface wrinkling) were observed only in the highest concentration of CSA-44. Furthermore, the stiffness of the bacterial surface and the adhesion forces decreased to a lesser extent, ie, stiffness only by 4.6% and adhesion by 40%, respectively.
 |
Figure 8 Schematic model of mechanical properties of AFM. |
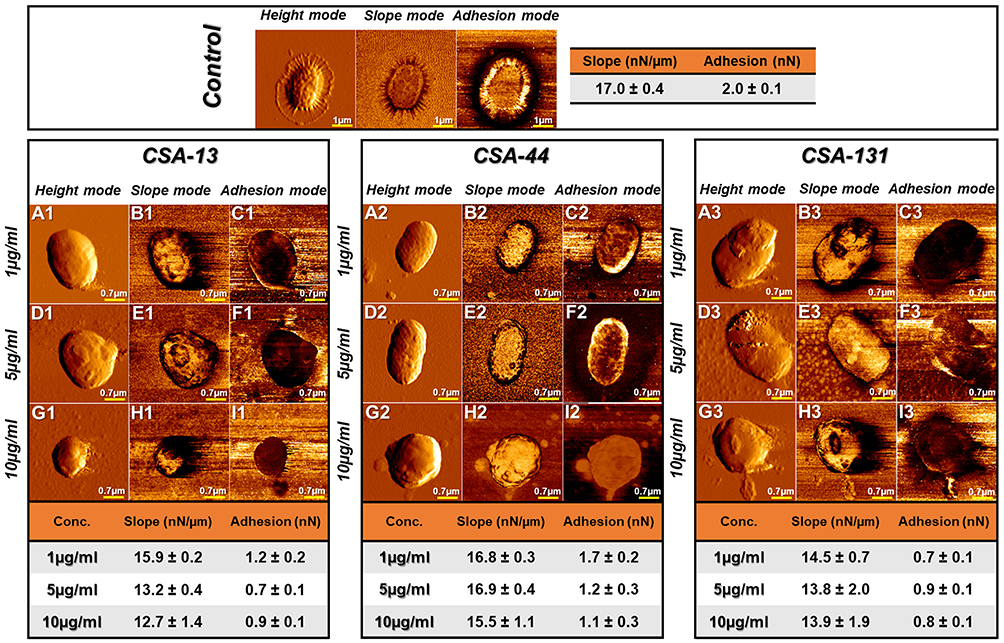 |
Figure 9 Atomic force microscopy measurements of Klebsiella pneumoniae BAA-2473 cells subjected to ceragenin CSA-13, CSA-44, and CSA-131 treatment. Examination of AFM topography and visualization of stiffness and adhesion indicate that all CSAs affect the morphology, mechanical, and adhesive properties of Klebsiella pneumoniae BAA-2473. The left column of panels (in both control and CSA-13, CSA-44, and CSA-131 sectors) shows the height mode (topography), the middle column shows the slope mode (stiffness), and the right column is represented by the adhesion mode (forces between the cells and the AFM probe). The mean values of slope and adhesion (plus standard deviation) are presented in the tables. |
Ceragenins CSA-13, CSA-44, and CSA-131 Exert Low Hemolytic Activity
Ceragenins may cause red blood cell (RBC) hemolysis at high concentrations as a consequence of interference with the integrity of plasma membrane and loss of the membrane’s ability to control homoeostasis.30 It should be highlighted that assessment of hemoglobin release from RBC can provide significant information of potential toxicity of those compounds.30,31 Data in Figure 10A–C indicated that all studied ceragenins did not affect RBC membrane permeability at the concentration range of 1–25 μg/mL after 1 hour of incubation (Figure 10A–C). Furthermore, ceragenins in concentrations of 1–10 μg/mL after 6 and 12 hours of incubation, were non-hemolytic (Figure 10A–C). Importantly, concentrations of CSA-13, CSA-44, or CSA-131 mentioned above were sufficient to inhibit the growth or even efficiently kill NDM-1 producing bacteria (Figure 2A–D). What is more, at these doses CSAs prevent biofilm formation and cause reduction of the biofilm formed (Figure 3A–D and Figure 4A–L).
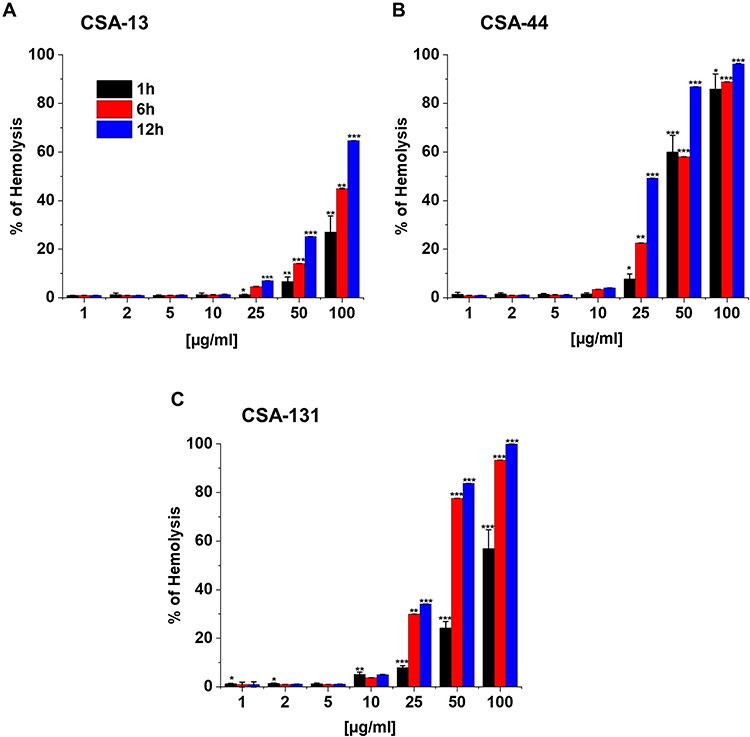 |
Figure 10 Hemoglobin release from human red blood cells (RBCs) after 1, 6, and 12 hours incubation in the presence of CSA-13 (A), CSA-44 (B), and CSA-131 (C) ranging from 1–100 μg/mL. Results show: mean±SD, n=3; * indicates statistical significance ≤0.05, ** ≤0.01, and *** ≤0.001. |
Discussion
Despite significant scientific progress, antibiotic resistance is still a global public health concern.32,33 What is more, due to the dearth of development of new antibiotics in the treatment of CPE infections, significant increasing in morbidity and mortality are observed all over the world.33
Bacteria resistance to carbapenems is generally associated with three fundamental mechanisms: production of carbapenemases, expression of efflux pumps, and porin mutations, which may result in reduction of the outer membrane permeability.5,34 The production of carbapenemases is a crucial mechanism underlying carbapenem resistance among bacteria throughout the world (Figure 11A).5 According to the Ambler classification method, carbapenemases can be classified as 1) Class A Klebsiella pneumoniae carbapenemase (KPC); 2) Class B Metallo-β-Lactamases (MBLs) such as New Delhi MBL (NDM-1), and 3) Class D oxacillinases (OXA), eg, OXA-48-type carbapenemases.5,35 Studies performed in Poland between 2016–2018 have demonstrated that 47.1–86.9% of all CPE strains harbor the NDM enzyme.36 Therefore, NDM-1 carbapenemases have been recognized as one of the most dangerous concerns.13
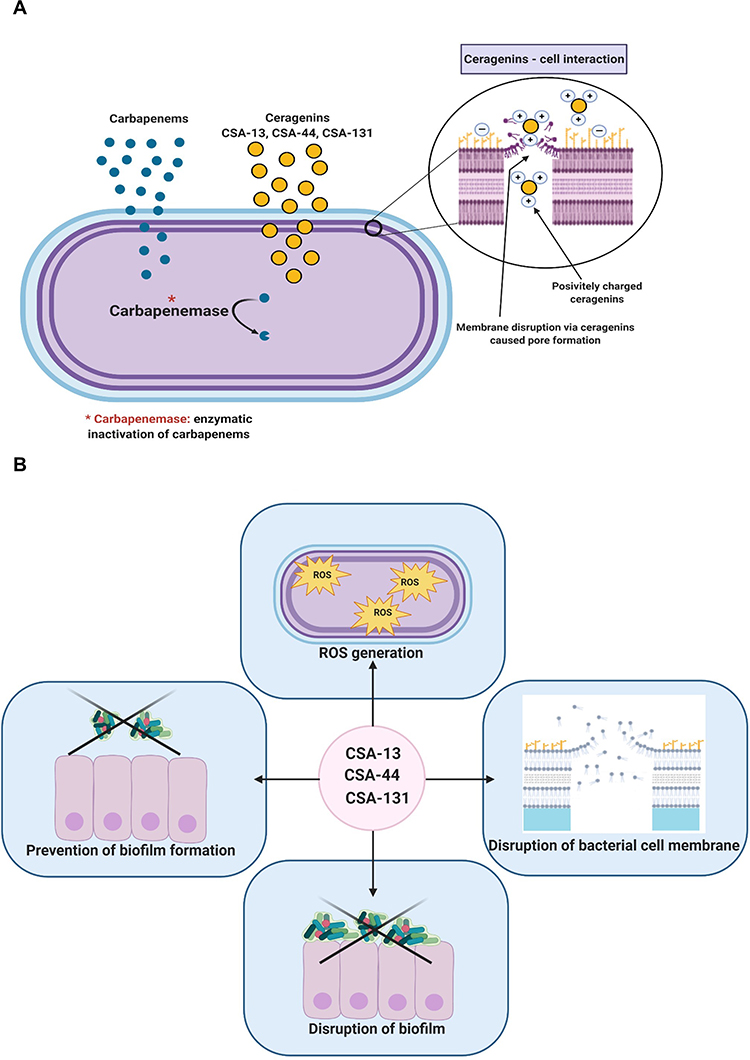 |
Figure 11 Mechanism of carbapenem resistance among bacteria belonging to Enterobacteriaceae family. Proposed mechanism of action which involves selective association with microbial membranes, mediated by ion pairing of positively charged CSAs with negatively charged microbial membranes (A). The pleiotropic activity of CSA-13, CSA-44, and CSA-131 (B). The figure was prepared using BioRender. |
Ceragenins could be developed as a therapeutic strategy against bacteria producing NDM-1 carbapenemases. CSAs possess potent antimicrobial activities against a broad spectrum of microorganisms including Gram-positive bacteria, Gram-negative bacteria, fungi, virus, or parasites.21,37 According to the literature, CSAs may be effective, ie, against hypervirulent Pseudomonas aeruginosa strain LESB58, colistin-resistant K. pneumoniae, carbapenem resistant Acinetobacter baumannii, methicillin, or even vancomycin resistant Staphylococcus aureus. In turn, among fungi, CSAs can be useful against fluconazole-resistant Candida albicans or C. auris. Therefore, CSAs can be considered as a new approach to eradicate MDR pathogens.16,20,29,38,39
Moreover, bacteria are unable to achieve high levels of resistance to CSAs. It should be pointed out that ceragenins appear to be well tolerated and exhibits no degradation in the presence of bacterial and host proteases, making these properties a significant area of the antimicrobial research.20
An important criterion in the assessment of effectiveness of CSAs is maintenance of the sufficient antibacterial activity with highly drug-resistant organisms. MIC values were determined for this purpose. Based on the current study, we point out that all ceragenins (CSA-13, CSA-44, CSA-131) are active against NDM-1 producing strains, ie, E. coli BAA 2471; E. cloacae BAA 2468; K. pneumoniae subsp. pneumoniae BAA 2472, and K. pneumoniae BAA 2473. The MIC range of all ceragenins tested was from 1–2 µg/mL, indicating at least 16-fold to more than 256-fold lower values than those for gentamicin, tobramycin, ciprofloxacin, and fosfomycin (except E. coli BAA 2471). According to carbapenems, ie, ertapenem, imipenem (except K. pneumoniae subsp. pneumoniae BAA 2472), and meropenem, ceragenins gave at least 8-fold to even more than 256-fold lower values of MICs against the strains expressing an NDM-1 resistance mechanism. Although the antibiotics/chemotherapeutics mentioned above have been recommended in the treatment of infections caused by NDM-1 producing strains, our results demonstrated that these drugs were ineffective in eradicating strains tested due to the high levels of resistance. It should be underlined that our data are in agreement with those of Ozbek-Celik et al,38 who reported that MICs values for meropenem-resistant clinical isolates of K. pneumoniae were 0.5–32 µg/mL for CSA-13 and CSA-44 (MIC90, 32 µg/mL) and 0.5–16 µg/mL in the case of CSA-131 (MIC90, 32 µg/mL). In addition, Bozkurt Guzel et al40 also determined the antimicrobial activity of CSA-13 against E. coli ATCC 25922 with the MIC value 4 µg/mL. Furthermore, it must be emphasized that E. coli ATCC 25922 does not produce any carbapenemases. Our observations may lead to the conclusion that the ceragenins analyzed are more effective than carbapenems, aminoglycosides, colistin (except E. coli BAA 2471), ciprofloxacin, and fosfomycin against the strains tested in our study.
The high activity of CSA-13, CSA-44, and CSA-131 was confirmed by the killing assay. The investigation showed that CSA-13, at 5-fold lower concentrations than MIC values, still had a potent bactericidal activity against K. pneumoniae BAA 2473. Moreover, at concentrations higher than 0.8 µg/mL, no bacteria growth was observed.
With regard to novel fields of research that focus on preventing adherence and biofilm formation of bacteria, the purpose of this study was to investigate activities of CSAs against formation of biofilm as well as disruption of the biofilms preformed. A hallmark of strains harboring blaNDM-1 gene is the formation of biofilm, which facilitates the persistence of these pathogenic isolates in the human body. To a great extent, biofilm formation substantially increases bacterial resistance to antibiotics, which is important in the context of infections.41
Few articles describing the antibiofilm activity of CSAs have been published so far. Interestingly, these compounds can be useful in preventing biofilm formation by P. aeruginosa, E. coli (associated with UTI), or Streptococcus pneumoniae.16,25,42 Additionally, the ability of CSAs to modulate mixed species biofilms formed by organisms like P. aeruginosa and S. aureus or C. albicans and S. aureus was observed.37,43 Additionally, Durnaś et al19, and Bozkurt-Guzel et al21 demonstrated the significant inhibition of biofilm generation by fungi as C. albicans. In addition, data reported by Hashemi et al39 indicate antibiofilm activities of CSAs against C. auris.
Accordingly, all ceragenins tested (CSA-13, CSA-44, CSA-131) have been shown to penetrate the extracellular matrix and eradicate a biofilm in a dose-dependent manner.20,25 Importantly, even non-toxic doses of ceragenins (at least 5–10 μg/mL) inhibit the biofilm formation in the 40–50% range.
Our results indicate that all CSAs significantly inhibited biofilm formation and possessed the ability to disrupt the biofilm formed by strains harboring the blaNDM-1 gene. What is more, the most valid finding of our investigation is the promising application of CSA-13, CSA-44, and CSA-131 as potential therapeutics in preventing the biofilm-related infections even caused by NDM-1 producing strains.
Previous results suggested that the mechanism of ceragenins action is the damage to the plasma membrane leading to its dysfunction.16,21,37 In order to confirm this hypothesis with regard to MDR bacteria such as NDM-1 producing strains, NPN uptake and ONPG hydrolysis assays were used. We found that all analyzed CSAs were able to permeabilize the outer membrane of NDM-1 producing bacteria in a dose-dependent manner. It should be emphasized that the minimum concentration required for the disruption of the outer membrane integrity was determined as 1 µg/mL. Furthermore, the studies presented herein demonstrated that CSA-13, CSA-44, and CSA-131 exert significant disruption of the inner membrane integrity in a dose-dependent manner. It must be noted that even at 1 µg/mL concentration the insertion of CSAs into the bacterial membrane was detected.
Additionally, we found that, apart from the disruption of the bacterial membrane, the generation of reactive oxygen species (ROS) also plays a key role in the antibacterial activity of ceragenins. Interestingly, ROS may cause numerous toxic effects, including DNA and RNA damage, membrane depolarization, protein carbonylation, and lipid peroxidation. ROS-mediated damage presumably triggers the subsequent rounds of ROS production leading to an unstoppable process that could be the terminal step in the life of bacteria. In effect, accumulated lesions ultimately overwhelm the capacity of a cell to repair which ensures the cell death.44
Increased production of ROS is assumed to be associated with the mechanism of action of ceragenins, which results in the formation of pores within the cell membrane. As a consequence, the integrity of bacterial cells was lost due to an imbalance in the bacterial redox status.26 For instance, Niemirowicz et al26 demonstrated that treatment of representative clinical isolates of C. albicans with magnetic nanoparticles conjugated with CSA-13 contributes to generation of ROS. However, to date this mechanism of CSAs action have not been investigated in CPE strains. Remarkably, we noted that ROS generation depends on ceragenin, and CSA-44 caused only an ~3-fold increase in oxidative damage, whereas CSA-131 caused a from >10 to >12-fold increase compared to control.
Based on the results collected herein, the proposed mode of the antibacterial action of CSA-13, CSA-44, and CSA-131 against the bacteria producing NDM-1 producing involves the disruption of the inner and outer bacterial membrane combined with the induction of an oxidative stress via the generation of ROS.
Investigating the structure and mechanical properties of bacterial surfaces with high resolution is the paramount importance to better understand the mechanism of ceragenins action.45 In order to identify changes in K. pneumoniae BAA-2473 cell membrane damage upon CSA-13, CSA-44, and CSA-131 treatment an AFM study was performed. The results obtained indicate that all tested ceragenins affect the bacterial structure which consequently leads to changes in the bacterial shape, ie, microcracks and surface wrinkling, and finally disrupt the cell membrane with the leakage of the intracellular contents. The data presented in this study are in agreement with our previous research demonstrating the changes in the bacterial structure and an increase in the membrane permeability of S. aureus X30 after treatment with CSA-13 and CSA-13+MNP@NH2.29 Additionally, disorganization of Gram-negative bacteria membranes such as P. aeruginosa in response to MNP@CSA-13 was also confirmed.30 The similar analysis of AFM also indicates the membrane destruction of C. albicans cells after MNP@CSA-13 addition.26
Finally, a crucial aspect that must be considered in the development of novel compounds is their safety. In relations to studies describing the in vivo application of CSA, the potent antibacterial and anti-inflammatory activities of CSA-13 in a mouse model of P. aeruginosa infection has been observed.16 Similar low toxicity of CSA-13 in the mouse model demonstrated by Saha et al46 underlines the possibility of the application of this compound for human therapeutics. Additionally, pharmacokinetic features of CSA-13 are comparable to conventional antibiotics and might be placed close to fluoroquinolones.16,23,25,30 In order to check this property of CSAs, hemolysis of RBC was performed. The evaluation of hemoglobin release from RBC can be a helpful clue in determining the potential toxicity of ceragenins.30,31 According to our investigation, the toxicity of CSA-13, CSA-44, and CSA-131 has been established to be low relative to bactericidal and anti-biofilm doses. Importantly, the concentration of CSAs ranging from 1–25 μg/mL did not cause hemolysis after 1 hour of incubation. It is significant to underline that the bactericidal activity as well as the inhibition of biofilm formation or the disruption of biofilm preformed have still been observed at these doses of CSA-13, CSA-44, and CSA-131. Due to minimal hemolytic effects, the results obtained suggest promising advantages of CSAs in the treatment of infections caused by NDM-1 producing strains.
Conclusions
This is the first report evaluating the activity of CSA-13, CSA-44, and CSA-131 against CPE strains producing NDM-1. Globally, the prevalent expansion of multidrug or even extensively drug resistant pathogens demand the development of novel antibacterial compounds. Our study results suggest a promising future of CSA-13, CSA-44, and CSA-131 as potential therapeutics for treating infections caused by NDM-1 producing strains. Moreover, CSAs demonstrate many compelling advantages, amidst them higher antibacterial activity than antibiotics/chemotherapeutics tested, notable antibiofilm capacity, and even the potential to efficiently eradicate bacteria embedded within biofilm (Figure 11B). Another important finding of our study is that CSAs at concentrations enabling observation of the benefits mentioned still remain non-hemolytic. What is more, mechanisms of CSA-13, CSA-44, and CSA-131 involve the disruption of the inner and outer bacterial membrane combined with the increased ROS generation (Figure 11B). Additionally AFM imagine has shown morphological changes in bacterial cells, ie, microcracks and surface wrinkling as well as reduction of stiffness and adhesion properties. Significantly, our investigation indicates that CSA-13, CSA-44, and CSA-131 can be developed as novel and efficient weapons in the fight against NDM-1 strains.
Ethics Statement
Hemolytic activity of tested agents was evaluated in blood samples from adult healthy volunteers under IRB approval: R-I-002/382/2012. This study was approved by the institutional review board (IRB) of The Medical University of Bialystok. The study was in accordance with the Declaration of Helsinki and written informed consent was provided by the blood donors.
Acknowledgments
The authors would like to express very great appreciation to Dr Piotr Deptuła for his AFM measurements which significantly improved the quality of the manuscript. This work was supported by the National Science Center, Poland, under Grant: UMO-2018/31/B/NZ6/02476 and UMwB SUB/1/DN/19/001/1162.
Disclosure
PBS is a consultant for N8 Medical, Inc and reports personal fees from N8 Medical, Inc, during the conduct of the study. The authors report no other potential conflicts of interest for this work.
References
1. Aslam B, Wang W, Arshad MI, et al. Antibiotic resistance: a rundown of a global crisis. Infect Drug Resist. 2018;11:1645–1658. doi:10.2147/IDR.S173867
2. Gashaw M, Berhane M, Bekele S, et al. Emergence of high drug resistant bacterial isolates from patients with health care associated infections at Jimma University medical center: a cross sectional study. Antimicrob Resist Infect Control. 2018;7(1):138. doi:10.1186/s13756-018-0431-0
3. Cheng P, Li F, Liu R, et al. Prevalence and molecular epidemiology characteristics of carbapenem-resistant. Infect Drug Resist. 2019;12:2505–2518. doi:10.2147/IDR.S208122
4. Da Silva GJ, Mendonça N. Association between antimicrobial resistance and virulence in Escherichia coli. Virulence. 2012;3(1):18–28. doi:10.4161/viru.3.1.18382
5. Cui X, Zhang H, Du H. Carbapenemases in Enterobacteriaceae: detection and Antimicrobial Therapy. Front Microbiol. 2019;10:1823. doi:10.3389/fmicb.2019.01823
6. Global priority list of antibiotic-resistant bacteria to guide research, discovery and development of new antibiotics. Available from: https://www.who.int/medicines/publications/WHO-PPL-Short_Summary_25Feb-ET_NM_WHO.pdf?ua=1.
7. Biggest Threats and Data, 2019 AR Threats Report; 2019. Available from: https://www.cdc.gov/drugresistance/biggest-threats.html.
8. Data from the ECDC Surveillance Atlas - Antimicrobial resistance; 2018. Available from: https://www.ecdc.europa.eu/en/antimicrobial-resistance/surveillance-and-disease-data/data-ecdc.
9. Livorsi DJ, Chorazy ML, Schweizer ML, et al. A systematic review of the epidemiology of carbapenem-resistant Enterobacteriaceae in the United States. Antimicrob Resist Infect Control. 2018;7(1):55. doi:10.1186/s13756-018-0346-9
10. Gupta A, Mumtaz S, Li CH, Hussain I, Rotello VM. Combatting antibiotic-resistant bacteria using nanomaterials. Chem Soc Rev. 2019;48(2):415–427. doi:10.1039/C7CS00748E
11. Kong Z, Cai R, Cheng C, et al. First Reported Nosocomial Outbreak Of NDM-5-Producing. Infect Drug Resist. 2019;12:3557–3566. doi:10.2147/IDR.S218945
12. Dane Krajowego Ośrodka Referencyjnego ds. Lekowrażliwości Drobnoustrojów (KORLD), dotyczące pałeczek Enterobacterales wytwarzających karbapenemazy NDM, KPC, VIM i OXA-48 na terenie Polski w latach 2006 – 2018; 2018. Available from: https://korld.nil.gov.pl/pdf/Raport%20KORLD%202019_EL_2.pdf.
13. Chmielewska S, Leszczyńska K. Carbapenemase of intestinal rods- the beginning of post-antibiotic era? Post Microb. 2019;58:271–289.
14. Yu X, Zhang W, Zhao Z, et al. Molecular characterization of carbapenem-resistant Klebsiella pneumoniae isolates with focus on antimicrobial resistance. BMC Genomics. 2019;20(1):822. doi:10.1186/s12864-019-6225-9
15. Raheem N, Straus SK. Mechanisms of Action for Antimicrobial Peptides With Antibacterial and Antibiofilm Functions. Front Microbiol. 2019;10:2866. doi:10.3389/fmicb.2019.02866
16. Wnorowska U, Fiedoruk K, Piktel E, et al. Nanoantibiotics containing membrane-active human cathelicidin LL-37 or synthetic ceragenins attached to the surface of magnetic nanoparticles as novel and innovative therapeutic tools: current status and potential future applications. J Nanobiotechnology. 2020;18(1):3. doi:10.1186/s12951-019-0566-z
17. Ding B, Taotofa U, Orsak T, Chadwell M, Savage PB. Synthesis and characterization of peptide-cationic steroid antibiotic conjugates. Org Lett. 2004;6(20):3433–3436. doi:10.1021/ol048845t
18. Ding B, Guan Q, Walsh JP, et al. Correlation of the antibacterial activities of cationic peptide antibiotics and cationic steroid antibiotics. J Med Chem. 2002;45(3):663–669. doi:10.1021/jm0105070
19. Durnaś B, Wnorowska U, Pogoda K, et al. Candidacidal Activity of Selected Ceragenins and Human Cathelicidin LL-37 in Experimental Settings Mimicking Infection Sites. PLoS One. 2016;11(6):e0157242. doi:10.1371/journal.pone.0157242
20. Hashemi MM, Mmuoegbulam AO, Holden BS, et al. Susceptibility of Multidrug-Resistant Bacteria, Isolated from Water and Plants in Nigeria, to Ceragenins. Int J Environ Res Public Health. 2018;15(12):12. doi:10.3390/ijerph15122758
21. Bozkurt-Guzel C, Hacioglu M, Savage PB. Investigation of the in vitro antifungal and antibiofilm activities of ceragenins CSA-8, CSA-13, CSA-44, CSA-131, and CSA-138 against Candida species. Diagn Microbiol Infect Dis. 2018;91(4):324–330. doi:10.1016/j.diagmicrobio.2018.03.014
22. Piktel E, Prokop I, Wnorowska U, et al. Ceragenin CSA-13 as free molecules and attached to magnetic nanoparticle surfaces induce caspase-dependent apoptosis in human breast cancer cells via disruption of cell oxidative balance. Oncotarget. 2018;9(31):21904–21920. doi:10.18632/oncotarget.25105
23. Durnaś B, Piktel E, Wątek M, et al. Anaerobic bacteria growth in the presence of cathelicidin LL-37 and selected ceragenins delivered as magnetic nanoparticles cargo. BMC Microbiol. 2017;17(1):167. doi:10.1186/s12866-017-1075-6
24. Piktel E, Pogoda K, Roman M, et al. Sporicidal activity of ceragenin CSA-13 against Bacillus subtilis. Sci Rep. 2017;7(1):44452. doi:10.1038/srep44452
25. Wnorowska U, Piktel E, Durnaś B, Fiedoruk K, Savage PB, Bucki R. Use of ceragenins as a potential treatment for urinary tract infections. BMC Infect Dis. 2019;19(1):369. doi:10.1186/s12879-019-3994-3
26. Niemirowicz K, Durnaś B, Tokajuk G, et al. Formulation and candidacidal activity of magnetic nanoparticles coated with cathelicidin LL-37 and ceragenin CSA-13. Sci Rep. 2017;7(1):4610. doi:10.1038/s41598-017-04653-1
27. Labus K. Effective detection of biocatalysts with specified activity by using a hydrogel-based colourimetric assay - β-galactosidase case study. PLoS One. 2018;13(10):e0205532. doi:10.1371/journal.pone.0205532
28. Gravel J, Paradis-Bleau C, Schmitzer AR. Adaptation of a bacterial membrane permeabilization assay for quantitative evaluation of benzalkonium chloride as a membrane-disrupting agent. Medchemcomm. 2017;8(7):1408–1413. doi:10.1039/C7MD00113D
29. Niemirowicz K, Piktel E, Wilczewska AZ, et al. Core-shell magnetic nanoparticles display synergistic antibacterial effects against. Int J Nanomedicine. 2016;11:5443–5455. doi:10.2147/IJN.S113706
30. Niemirowicz K, Surel U, Wilczewska AZ, et al. Bactericidal activity and biocompatibility of ceragenin-coated magnetic nanoparticles. J Nanobiotechnology. 2015;13(1):32. doi:10.1186/s12951-015-0093-5
31. Leszczyńska K, Namiot A, Cruz K, et al. Potential of ceragenin CSA-13 and its mixture with pluronic F-127 as treatment of topical bacterial infections. J Appl Microbiol. 2011;110(1):229–238. doi:10.1111/j.1365-2672.2010.04874.x
32. Peyclit L, Baron SA, Rolain JM. Drug Repurposing to Fight Colistin and Carbapenem-Resistant Bacteria. Front Cell Infect Microbiol. 2019;9:193. doi:10.3389/fcimb.2019.00193
33. Lee CR, Lee JH, Park KS, Kim YB, Jeong BC, Lee SH. Global Dissemination of Carbapenemase-Producing Klebsiella pneumoniae: epidemiology, Genetic Context, Treatment Options, and Detection Methods. Front Microbiol. 2016;7:895.
34. Eichenberger EM, Thaden JT. Epidemiology and Mechanisms of Resistance of Extensively Drug Resistant Gram-Negative Bacteria. Antibiotics. 2019;8:2.
35. Sheu CC, Chang YT, Lin SY, Chen YH, Hsueh PR. Infections Caused by Carbapenem-Resistant. Front Microbiol. 2019;10:80. doi:10.3389/fmicb.2019.00080
36. Raport Krajowego Ośrodka Referencyjnego ds. Lekowrażliwości Drobnoustrojów Występowanie Enterobacteriaceae (głównie Klebsiella pneumoniae), wytwarzających karbapenemazę New Delhi (NDM) na terenie Polski w okresie I – III kwartał 2017 roku; 2017. Available from: https://korld.nil.gov.pl/pdf/Raport_NDM_18-12-2017_strona.pdf.
37. Hacioglu M, Haciosmanoglu E, Birteksoz-Tan AS, Bozkurt-Guzel C, Savage PB. Effects of ceragenins and conventional antimicrobials on Candida albicans and Staphylococcus aureus mono and multispecies biofilms. Diagn Microbiol Infect Dis. 2019;95(3):114863. doi:10.1016/j.diagmicrobio.2019.06.014
38. Ozbek-Celik B, Damar-Celik D, Mataraci-Kara E, Bozkurt-Guzel C, Savage PB. Comparative In Vitro Activities of First and Second-Generation Ceragenins Alone and in Combination with Antibiotics Against Multidrug-Resistant. Antibiotics. 2019;8:3.
39. Hashemi MM, Rovig J, Holden BS, et al. Ceragenins are active against drug-resistant Candida auris clinical isolates in planktonic and biofilm forms. J Antimicrob Chemother. 2018;73(6):1537–1545. doi:10.1093/jac/dky085
40. Bozkurt Guzel C, Oyardi OB, Savage P. Comparative in vitro antimicrobial activities of CSA-142 and CSA-192, second-generation ceragenins, with CSA-13 against various microorganisms. J Chemother. 2018;30(6–8):332–337. doi:10.1080/1120009X.2018.1534567
41. Wnorowska U, Watek M, Durnas B, et al. Extracellular DNA as an essential component and therapeutic target of microbial biofilm. Med Stud Studia Medyczne. 2015;31(2):132–138. doi:10.5114/ms.2015.52912
42. Moscoso M, Esteban-Torres M, Menéndez M, García E. In vitro bactericidal and bacteriolytic activity of ceragenin CSA-13 against planktonic cultures and biofilms of Streptococcus pneumoniae and other pathogenic streptococci. PLoS One. 2014;9(7):e101037. doi:10.1371/journal.pone.0101037
43. Olekson MA, You T, Savage PB, Leung KP. Antimicrobial ceragenins inhibit biofilms and affect mammalian cell viability and migration. FEBS Open Bio. 2017;7(7):953–967. doi:10.1002/2211-5463.12235
44. Hong Y, Zeng J, Wang X, Drlica K, Zhao X. Post-stress bacterial cell death mediated by reactive oxygen species. Proc Natl Acad Sci U S A. 2019;116(20):10064–10071. doi:10.1073/pnas.1901730116
45. Gammoudi I, Mathelie-Guinlet M, Morote F, et al. Morphological and nanostructural surface changes in Escherichia coli over time, monitored by atomic force microscopy. Colloids Surf B Biointerfaces. 2016;141:355–364. doi:10.1016/j.colsurfb.2016.02.006
46. Saha S, Savage PB, Bal M. Enhancement of the efficacy of erythromycin in multiple antibiotic-resistant gram-negative bacterial pathogens. J Appl Microbiol. 2008;105(3):822–828. doi:10.1111/j.1365-2672.2008.03820.x
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.