Back to Journals » International Journal of Nanomedicine » Volume 20
Nanoparticles Based Therapeutic Approaches in Oncogenic Viral Infections: A Review
Authors Ahmed N ![]() , Abusalah MAHA, Abuarqoub AH
, Abusalah MAHA, Abuarqoub AH
Received 15 June 2025
Accepted for publication 14 October 2025
Published 30 October 2025 Volume 2025:20 Pages 13133—13163
DOI https://doi.org/10.2147/IJN.S547270
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Krishna Nune
Naveed Ahmed,1,* Mai Abdel Haleem A Abusalah,2,* Alqassem Hamdallah Abuarqoub2,3
1Department of Assistance Medical Sciences, Applied College, University of Tabuk, Tabuk, 71491, Saudi Arabia; 2Department of Medical Laboratory Sciences, Faculty of Allied Medical Sciences, Al-Ahliyya Amman University, Amman, Jordan; 3Al-Wafi Group for Marketing and Int’l Trade Co. Ltd, Amman, Jordan
*These authors contributed equally to this work
Correspondence: Naveed Ahmed, Department of Assistance Medical Sciences, Applied College, University of Tabuk, Tabuk, 71491, Saudi Arabia, Email [email protected]
Abstract: Nanotechnology has revolutionized biomedical research by providing novel strategies for the prevention, diagnosis, and treatment of oncogenic viral infections. Nanoparticles, possessing tunable physicochemical properties, enable targeted drug delivery, enhance vaccine efficacy, and facilitate advanced viral diagnostics, addressing key limitations of conventional therapies. Oncogenic viruses contribute substantially to the global burden of cancer through persistent infections and are influenced by environmental and lifestyle factors. The objective of this review is to synthesize and critically evaluate current advances in nanoparticle-based solutions, focusing on their mechanisms of action, therapeutic efficacy, and translation for the management of cancer-associated viral infections. Key findings include the emergence of multifunctional nanoparticles such as magnetic mesoporous silica, gold, silver, and diverse organic formulations (liposomes, polymeric, dendrimer) that function as direct antivirals and as delivery platforms. Magnetic silica nanoparticles allow magnetically guided, site-specific therapy, while organic nanoparticles improve drug stability, mucosal delivery, and biocompatibility. Metallic nanoparticles, particularly gold and silver, exhibit broad-spectrum antiviral activity through mechanisms involving viral disruption, entry inhibition, and enhanced bioavailability. Current evidence supports their efficacy in preclinical and translational models, although optimization for clinical use remains ongoing. In conclusion, advanced nanomedicine approaches offer substantial potential for tackling the challenges of oncogenic viruses and virus-associated malignancies, with ongoing innovations poised to expand their impact in cancer therapy and antiviral prevention.
Keywords: nanotechnology, nanoparticle-based therapies, gold nanoparticles, silver nanoparticles, viral infections, oncogenic viruses, antiviral agents
Introduction
In the early 1900s, cell-free filtrates were used to spread some types of animal cancers, establishing the possibility of an infectious etiology of carcinogenesis. Multiple studies have shown that retroviruses are responsible for the spread of these tumors and that retroviruses cause cancer by insertional mutagenesis or the production of viral versions of cellular genes, subsequently named oncogenes.1 The genus human papillomavirus (HPV) has been identified in cervical cancer and cutaneous squamous cell carcinoma (SCC). A plethora of viruses have been identified as the cause of human cancer. These include human T-cell leukemia virus type 1 (HTLV-1), human T-cell lymphotropic virus type 1 (TCLV-1), Merkel cell polyomavirus (MCPyV), human herpesvirus 8 (HHV-8), Kaposi’s sarcoma-associated herpesvirus (KSHV), Epstein-Barr virus (EBV), hepatitis B virus (HBV), hepatitis C virus (HCV), and human papillomaviruses (HPVs).2
Furthermore, organ transplant recipients with long-term immunosuppression and individuals with uncommon genetic skin conditions have been reported to develop cutaneous SCC caused by HPVs.3 Oncogenic viruses, such as human papillomavirus (HPV), Epstein-Barr virus (EBV), and hepatitis B/C viruses, collectively account for about 12–15% of human cancers worldwide.2 People with weakened immune systems, especially those who live in unsanitary surroundings and have limited access to healthcare, are more likely to develop malignancies caused by viruses. The KSHV and MCPyV were discovered to be virus-induced cancers, owing to the high prevalence of malignancy in immunosuppressed patients.4 Viruses may cause cancer via at least three pathways: oncogenic proteins encoded by viruses, chronic inflammation caused by viruses, and increased risk of carcinogenesis by causing genomic instability or other host cell changes.5 Given these complex interactions, the scientific objective of this review is to provide a comprehensive and critical analysis of recent progress in nanoparticle-enabled approaches to the prevention, diagnosis, and treatment of oncogenic viral infections, with particular focus on their molecular mechanisms, translational efficacy, and future challenges in clinical application. This review also aims to serve as an authoritative reference for researchers, clinicians, and nanomedicine developers working at the intersection of virology, oncology, and nanotechnology.
Molecular Mechanisms of Viral Oncogenesis in Humans
Lymphoid malignancies and solid tumors may be caused by the large DNA viruses, KSHV and EBV. The DNA genomes of KSHV and EBV are larger than those of MCPyV and HPV. MCPyV infects and probably remains overtly in dermal fibroblasts,6 while oncogenic HPVs generate chronic infections in the mucosal epithelia.7 Tumorigenesis is stimulated by a limited number of multifunctional oncoproteins produced by these small oncogenic DNA viruses.4 Hepatocytes are attacked by both HBV (tiny DNA virus) and HCV (single-stranded positive-sense RNA virus), leading to chronic liver inflammation, liver cirrhosis, and HCC. Human T cells are infected with HTLV-1 (an oncogenic retrovirus), which has been linked to T-cell lymphoma in adults.5
Therefore, these human oncogenic viruses exhibit variable disease prevalence, tumor pathology, cellular tropism, and genomes. Importantly, these viruses share many characteristics with human cancers. Asymptomatic chronic infections are caused by viruses that persist for longer periods and can be transmitted from person to person. During these protracted periods, oncogenic viruses impair immune recognition and co-opt cellular replication.5 They disrupt fundamental signaling pathways that regulate cell death and cell cycle progression to ensure their proliferation.8 As depicted in Figure 1, the stages in the development of cancer are represented sequentially, illustrating the progression from initial cellular transformation to malignant tumor formation.
 |
Figure 1 Stages in the development of cancer. |
While tumorigenesis is the unifying pathogenic consequence of all oncogenic viruses, this outcome does not benefit viral survival or transmission and is not essential for viral life cycles. The seven known human oncogenic viruses including EBV, KSHV, HBV, HCV, HTLV-1, HPVs, and MCPyV, utilize both overlapping and virus-specific mechanisms to drive cancer.8 Most encode viral oncoproteins that inactivate cellular tumor suppressors, alter cell signaling, and modulate immune responses to promote transformation; however, some (notably HBV and HCV) primarily induce tumorigenesis through chronic inflammation and repeated cycles of cell damage and regeneration, which increase mutational burden and genomic instability.8 Recent researches emphasize that in-depth study of these viruses’ proteins and molecular pathways will help identify what differentiates each virus’s oncogenic process and provide new targets for cancer prevention and therapy.8
Genomic Classification of Human Oncogenic Viruses
Oncogenic viruses are broadly categorized by the type of genetic material they carry—either DNA or RNA. DNA oncogenic viruses include EBV, KSHV, HPV, HBV, MCPyV, and herpes simplex virus types 1 and 2 (HSV-1 and HSV-2).9 HSV-1 and HSV-2, as DNA viruses, have been detected in association with several human cancers, including cervical, thyroid, and prostate cancers, and can promote tumorigenesis through inhibition of apoptosis, induction of unscheduled DNA synthesis, and dysregulation of oncogenic signaling pathways (such as the Ras/Raf/MEK/ERK cascade).10 Although the carcinogenic potential of HSV is not as well established as for EBV or HPV, increasing evidence suggests HSV infection may contribute to tumorigenesis, especially in the context of immunosuppression or chronic inflammation.10
In contrast, RNA oncogenic viruses include HTLV-1, HCV, and hepatitis D virus (HDV). HCV is a positive-sense single-stranded RNA virus that drives hepatocellular carcinoma mainly via persistent inflammation, oxidative stress, and induction of liver fibrosis, in addition to direct oncogenic effects of its proteins such as deregulation of cell-cycle control, apoptotic pathways, and promoting genomic instability.8,10 HDV, a defective RNA virus requiring HBV co-infection, exacerbates chronic liver damage and inflammation, further increasing the risk of liver cirrhosis and HCC; the oncogenic synergy in HBV/HDV co-infections elevates carcinogenesis risk compared to HBV infection alone.11
Herpesviruses
γ-Herpesviruses
KSHV and EBV are herpesviruses that belong to the Herpesviridae family. These enveloped viruses have ds DNA of size 130–150 kbps enclosed in capsids of icosahedral shape to recognize and infect their host cells, including several glycoproteins (GPsEBV Gp350), which bind and attach to CR2-CD21 B-cell receptors and cause infection of these cells.12 The major envelope of -herpesviruses is obtained at the nuclear membrane during the maturation phase, and the initial envelopment-de-envelopment is from the nuclear inner and outer leaves. All herpesviruses, including HSV-1, share these same proteins. Herpesviruses obtain their final envelopes, and glycoproteins require new viral infections, although the NEC (nuclear membrane complex) can “acquire” nucleocapsids and transport them to the cytoplasmatic compartment. Clinicians have also investigated the processes underlying the lytic cycle to enhance conventional treatment of patients with KSHV- and EBV-associated illnesses.13
Epstein–Barr Virus (EBV)
EBV, also referred to as human herpesvirus 4 (HHV-4), is a double-stranded DNA virus and one of the most pervasive herpesviruses worldwide.14 EBV infection is nearly ubiquitous, with approximately 95% of adults exposed during their lifetime and most people do not know about their presence.15 However, various illnesses develop due to the development of certain environmental factors. Infectious mononucleosis (IM) is caused by EBV, which is also linked to lymphoproliferative diseases, such as stomach cancer, nasopharyngeal carcinoma, and Burkitt lymphoma, and frequently develops in patients undergoing solid organ transplantation (PTDLs), as well as autoimmune diseases such as rheumatoid arthritis and multiple sclerosis.14,16. The last decade has also shown a link between EBV infection and the development of immunological illnesses including rheumatoid arthritis (RA) and multiple sclerosis (MS).17 Multiple studies have highlighted the significance of plasma cells in the production of demyelinating antibodies in MS.18
Molecular Mechanisms of EBV-Induced Oncogenesis
EBV contributes to oncogenesis by expressing a defined set of latent and lytic genes within infected cells, which disrupt host cell cycle regulation, inhibit apoptosis, and activate cellular oncogenes such as BCL-2 and MYC.19 The virus encodes proteins like EBNA1 (Epstein-Barr nuclear antigens 1), LMP1 (latent membrane protein 1), and LMP2 (latent membrane protein 2) that mimic or alter cellular signaling pathways (NF-κB, JAK/STAT, PI3K/Akt), repress tumor suppressors (p53, p27, PTEN (Phosphatase and tensin homolog deleted on chromosome 10)), and promote cell proliferation and survival.19,20 Oncogenic transformation typically results from sustained latency, genomic instability, and chronic immune evasion. Tumor cells frequently harbor multiple extrachromosomal copies of circular EBV DNA, and the expression of EBV latent genes in tumor tissue is a molecular hallmark of EBV-driven cancers.19 EBV has been shown to integrate in fragile, transcriptionally active regions of the host genome, particularly in epithelial cancers like nasopharyngeal carcinoma (NPC). Such integration can alter the expression of both viral MicroRNAs (miRNAs) (such as miR-BARTs) and host genes, promoting tumor progression by driving or repressing specific gene networks.8 EBV-encoded small RNAs (BART miRNAs and EBERs) act to shape a tumor-promoting microenvironment by modulating inflammation and blunting immune recognition.8
The mechanisms of EBV-driven oncogenesis are multifaceted and involve both latent and lytic viral gene expression.8. EBV replication occurs in B cells, and the lytic cycle remains incomplete because no new virions are produced. Primary (lytic) infection occurs in the oropharyngeal epithelium during the productive phase of vivo.21 Viruses can rapidly disseminate because they can enter a dormant (latent) stage in which they do not generate infectious particles. This herpesvirus has a life cycle typical of herpesviruses. This virus may cause latent infections in the epithelial cells and B lymphocytes. In this phase, only a small subset of viral proteins is produced, whereas the rest of the genome is bound to a circular episome.22 Multiple studies have identified these proteins as pivotal in suppressing host immune responses and allowing persistent infections to take hold in target cells. The viral genome must be replicated via the host DNA polymerase throughout the cell cycle progression, and the EBNA-1 and EBNA-2 (Epstein-Barr nuclear antigens 2) are necessary for this process and for the virus to enter a “persisting” state.23
Over the past 60 years, numerous studies have demonstrated that marmoset-infected EBV cells can multiply and develop in vitro, while producing all essential proteins. The six nuclear antigens, EBNA-1 to EBNA-6, are among the vital proteins, latent membrane proteins (LMPs), and two non-polyadenylated RNAs (EBERs). LMP1 is a CD40 receptor that mimics integral membrane proteins. Infected cells proliferate indefinitely because their receptors are permanently activated. Hidden proteins control cell division and apoptosis, two forms of programmed cell death, such that their respective pathways are never activated.21 EBV latent proteins, especially LMP1 and LMP2A, induce a profound epigenetic reprogramming of the host cell.8 They upregulate DNA methyltransferases (DNMT1, DNMT3A/B), resulting in silencing of tumor suppressor genes such as CDH1 (E-cadherin), PTEN, and p16. EBNA family proteins and BZLF1 (BamHI Z fragment leftward open reading frame 1) further recruit histone-modifying complexes, suppressing genes responsible for apoptosis and cell cycle arrest.8
However, EBV may develop a productive phase in which over hundred proteins are produced for use in the lytic phase, manufacturing and producing new virions, and spreading the viral particles in various organs and tissues. The lytic state has been associated with disease severity (AR) in MS and RA and two autoimmune disorders associated with EBV. These three groups of lytic proteins are based on their role in viral replication. For replication, the early and late expression of viral IE proteins is required. The former must be present for the viral genome to replicate; they are referred to as concatemers, and after being released from their nucleocapsids, they acquire access to the plasma membrane through a well-defined process.18 Two proteins encoded by the BFRF1 and BFLF2 genes play an important role in the cytoplasmic migration and translocation of newly arrived viruses by shifting the nuclear membrane.24 Virions in the cytoplasm acquire proteins (late) required for tegument and envelope formation. The genome of EBV, isolated from B95.8 cells, has been cloned using DNA recombination procedures,25 and many in vitro systems have been generated for BAC construction. The role of each EBV protein has been determined using this method. To demonstrate the importance of this early EBV protein, a BFLF2 knockout mutant was developed.26 This study established that this gene expresses a protein that acts as a barrier to the nuclear egress of viruses and helps in the packing of DNA. Vaccine development has primarily focused on eliminating this lytic protein, because its absence slows capsid maturation and promotes the release of DNA-free virions (VLPs or virus-like particles).27
In vitro, drugs such as phorbol esters (TPA-phorbol 12-myristate 13-acetate) and histone deacetylase inhibitors (sodium butyrate-NaB) may stimulate the transition of latently infected cells into the lytic phase of viral replication. After an instant, early genes activate the lytic gene-triggering cascade, and all lytic proteins essential for virion assembly are produced. The protease inhibitor BZT enhances viral replication via autophagy machinery.28
Emerging Nanomedicine Approaches Against EBV-Driven Malignancies
Emerging nanomedicine approaches are rapidly transforming the diagnosis and treatment of EBV-driven malignancies.29,30 Advances in multifunctional nanoparticles including gold, magnetic, liposome-based, and protein-derived platforms, have enabled both precise EBV biomarker detection and targeted therapeutic delivery.29,31 For instance, gold nanoparticles have been employed to deliver miRNA or siRNA agents aimed at silencing EBV oncoproteins such as LMP1, demonstrating significant tumor suppression in nasopharyngeal carcinoma models.32 Liposome-based and ferritin-derived protein nanoparticle vaccines presenting EBV glycoproteins (gp350, gH/gL, gp42) have been shown to induce robust neutralizing responses in both B cells and epithelial cells in preclinical models.29,31 This has led to recent clinical trials such as NIH’s Phase I study of EBV gp350-ferritin nanoparticle vaccine evaluating safety and immunogenicity in humans.33
A striking innovation in EBV nanomedicine is the combination of nanoparticle platforms with cell-based therapies. Prussian blue nanoparticles (PBNPs) conjugated onto cytotoxic T lymphocytes (CTLs) targeting EBV antigens have produced biohybrid “nanoimmunotherapy” constructs, retaining both the antiviral/antitumor properties of CTLs and the drug delivery, imaging, and photothermal capabilities of PBNPs.34 Further, mRNA-loaded lipid nanoparticles targeting EBV latent proteins like LMP2 have been shown to sensitize relapsed EBV-related tumors to immune checkpoint blockade, illustrating synergies between RNA therapeutics and nanocarrier-mediated delivery.35 In this study, ionizable lipid nanoparticles carrying LMP2-mRNA have shown synergy with immune checkpoint inhibitors (anti-PD-1), producing pronounced antitumor effects and reversal of CD8+ T cell exhaustion in preclinical NPC studies. These lipid platforms facilitate targeted delivery to tumor-draining lymph nodes, maximize antigen presentation, and minimize systemic toxicity.35
Nanoscale biosensors and rapid diagnostic platforms are also at the forefront, with systems that incorporate gold or carbon nanomaterials, graphene, or biomimetic elements (host receptors such as CD21) enabling highly sensitive and specific detection of EBV DNA, RNA, or glycoproteins in clinical samples and saliva.36 These developments facilitate early detection, monitoring, and risk stratification for EBV-associated malignancies and autoimmune diseases such as multiple sclerosis.36,37
Overall, nanotechnology has enhanced drug and vaccine delivery efficiency, enabled multiplexed viral detection, and enabled innovative combined immunotherapeutic approaches against EBV-driven cancers.29,31 While preclinical evidence is promising, ongoing clinical trials and translational studies continue to define the optimal strategies and safety profiles for these platforms in humans.29,31
Challenges and Limitations in EBV Nanomedicine
Despite these breakthroughs, significant challenges remain. Achieving efficient in vivo delivery of nucleic acid therapeutics (such as RNAi or CRISPR (Clustered Regularly Interspaced Short Palindromic Repeats)/Cas9) into all EBV–infected tumor cells is difficult, in part due to biological barriers and tumor heterogeneity.8 Nanoparticle toxicity and immune reactivity require careful safety profiling and rigorous optimization of nanoparticle formulation and dosing.38 Long-term risks—such as bioaccumulation or chronic inflammation—must be addressed through further preclinical and clinical studies. Manufacturing scalability, regulatory approval, and interpatient heterogeneity add further obstacles to the clinical translation of nanomedicine approaches.38 Moreover, the high copy number of EBV episomes and viral evolution complicate curative strategies, making combination therapies and adaptive nanotechnologies a research priority.19
KSHV (Kaposi’s Sarcoma-Associated Herpesvirus)
The KSHV, also known as human herpesvirus-8 (HHV-8), is a gamma-herpesvirus.39,40 The virus contains a large double-stranded DNA genome and cycles between lytic and latent infection, with lifelong persistence primarily in endothelial and B cells.39 The earliest known male resident of the Mediterranean region had this trait. HIV-infected patients with KS were first identified in the early 1980s. Disorder multicentric Castleman disease (MCD) is associated with primary effusion lymphoma (PEL) and MCD. T lymphocytes, dendritic cells, monocytes, and endothelial cells are susceptible to infection.13 KSHV infection is mostly asymptomatic in immunocompetent individuals but is highly oncogenic in immunocompromised populations, such as people living with HIV/AIDS.39,40
Molecular Mechanisms Underlying KSHV-Induced Tumorigenesis
Oncogenesis by KSHV involves a complex interplay of latent and lytic viral gene products. Latency is the predominant state in Kaposi’s sarcoma lesions.39 However, little is known regarding the pathogenic pathways involved in the progression of this illness. According to various models used to describe viral expression during the latent phase, the virus may express proteins that mimic those typically expressed by the host cell to control propagation and cell apoptosis through checkpoints of the cell cycle or signal transduction activated by various kinases.41 The latent nuclear antigen (LANA), a multifunctional protein critical for the maintenance of viral latency and persistence in host cells, is encoded by open reading frame 73 (ORF73) of KSHV. LANA serves as an essential oncogenic driver through several mechanisms: it tethers the viral genome to host chromatin, disrupts key tumor suppressor pathways involving p53 and retinoblastoma protein (pRb), and regulates both viral and cellular gene expression to promote cell survival and proliferation.41 High expression of ORF73/LANA is a hallmark of KSHV latent infection and is indispensable for the growth and survival of Kaposi’s sarcoma tumor cells.41 Other latent proteins, like v-cyclin and vFLIP (viral FLICE-inhibitory protein), disrupt the cell cycle and activate NF-κB, promoting cell proliferation and survival. KSHV also encodes homologs of cytokines and chemokines (eg, v-IL-6 (viral Interleukin-6), v-GPCR (viral G protein-coupled receptor)), which promote inflammation, angiogenesis, and a pro-tumorigenic microenvironment.39 Lytic cycle gene products—expressed in a minority of tumor cells—further support paracrine signaling, sustaining tumor growth and angiogenesis.39
In addition, according to various models used to describe viral expression during the latent phase, the virus may express proteins that mimic those typically expressed by the host cell to control propagation and cell apoptosis through checkpoints of the cell cycle or signal transduction activated by various kinases. The LANA protein of KSHV, which controls the ongoing survival of the virus inside host cells, is encoded by ORF73.42 Disruption of p53 function contributes to dysregulation of cellular growth and survival LANA may interact with GSK3 (eg, c-Myc and cyclin D) to further regulate the cellular localization of β-catenin and subsequent activation of proliferative genes. The lytic state of KSHV is produced by K-bZIP and RTA proteins, which are homologs of the virus, that is, the EBV BZLF1 and BRLF1 early and direct genes. According to literature on EBV, both late and early proteins are required for virion maturation and viral replication.43,44
KSHV increases and amplifies IL-10 and IL-6 cytokine production, constitutively activating nuclear factor kappa B (NF-kB) and transcription factor STAT3.45,46 Recent studies by KSHV researchers have led to a model that explains how this virus contributes to the onset of Kaposi sarcoma by tracking the disease course using a battery of different diagnostic markers. The synergistic effects of the latent and lytic stages on disease severity are briefly discussed.47 Unfortunately, traditional therapies are often ineffective in patients with Primary Effusion Lymphoma (PEL).48 Therefore, a thorough understanding of the cellular processes (such as the fact that it maintains the course of the illness) is required to develop novel treatment techniques. By illuminating the cellular and viral processes associated with these cancers, researchers hope to enhance the therapeutic results of the existing medications.13
Using in vitro models, several pharmaceutical medicines have been shown to enhance (and prevent) the lytic state to suppress specific processes, such as apoptosis or autophagy. FDA-approved in 2003 for multiple myeloma treatment, bortezomib (BZT) has been shown to promote viral replication in KSHV-infected and EBV-infected B95.8 cells by modulating the ubiquitin-proteasome system and autophagy.49 Therefore, health care experts should consider the unusual side effects of this medicine. The effectiveness of therapy increases when it is combined with other medications.50 One of the routes investigated in this study is the autophagy system. By controlling the length of proteins and how quickly damaged organelles (such as the mitochondria) are degraded. How much space does the endoplasmic reticulum take up, and does this biological system maintain the cell in a steady state? In addition, chemoresistance is often associated with the dysregulation of autophagy, which occurs in several malignancies.51 Usually, this process is responsible for the degradation of pathogens and maintenance of immune system functionality. However, β-herpesviruses employ an autophagy mechanism to accelerate viral particle maturation and reproduction.51
Emerging Nanomedicine Strategies Against KSHV-Related Cancers
Recent pharmacological investigations have demonstrated that certain drugs that constrain autophagy at either the early or later stages, such as chloroquine, work in concert with platinum-based cancer therapies to slow disease progression in patients. Therefore, finding a method to administer these pharmaceutical medications without using harmful solvents is essential. Many innovative approaches have been developed in nanomedicine and nanodevices to mitigate the unwanted pharmacological consequences of these therapies.52 In addition, the earliest medical applications of nanosystems were to improve the effectiveness of existing treatments by increasing their dosage and bioavailability. Several antiviral mechanisms of NPs have been identified. Some are obligatory, whereas others are intended to improve vaccination performance of vaccinations.53
Nanotechnology offers innovative strategies against KSHV-driven malignancies.54,55 Nanoparticles, such as polymeric nanocarriers, facilitate targeted delivery of antiviral drugs or RNA-based therapeutics to KSHV-infected cells, enhancing stability, uptake, and tissue-specific targeting.56 Recent studies have demonstrated the efficacy of nano-formulated miRNAs in suppressing KSHV gene expression and tumor cell proliferation.55 Other platforms such as microneedle arrays loaded with pH-responsive nanoparticles, have achieved localized delivery of antiviral agents (eg, ganciclovir) with improved bioavailability and reduced off-target effects.54 Furthermore, nanoparticle-based biosensors enable sensitive detection of KSHV DNA, supporting diagnosis and monitoring.56
Challenges and Limitations in KSHV Nanotherapeutic Translation
Despite these advances, several challenges remain. Efficient and uniform nanoparticle delivery to all tumor sites is hindered by tissue barriers and tumor heterogeneity. Safety concerns include potential cytotoxicity, long-term nanoparticle accumulation, and possible immunogenic reactions.54 Current manufacturing standards for advanced delivery systems like microneedle arrays require further standardization and quality control.54 Additionally, clinical translation is complicated by inconsistent biodistribution, variability in patient immune status, and a lack of large-scale, long-term studies to assess efficacy and safety in diverse populations.54
Human Papillomavirus (HPV)
The HPV is a non-enveloped, double-stranded, circular DNA virus of the Papillomaviridae family that infects cutaneous and mucosal epithelia.57 HPV is one of the most common sexually transmitted infections (STIs) worldwide and is causally responsible for approximately 5% of all human cancers, particularly those of the cervix, anogenital tract, and head and neck regions.57 More than 200 varieties of HPVs have been characterized based on their DNA sequences, although only a subset is known to cause serious health issues. By infecting epithelial cells, HPVs are classified as cutaneous or mucosal HPVs. The anogenital, pulmonary, and oropharyngeal epithelia are infected by the final group of HPVs, making them important in cancer biology. At least 20 different HPV types have been associated with anogenital lesions, which are further divided into high-risk (HR-HPV) types that are associated with the risk of premalignant squamous and low-risk (LR-HPV) types. The HR-HPV types such as HPV-16 and HPV-18 are strongly associated with oncogenesis.57
Cervical cancer is the most common illness caused by HPV infection.58 In 2020, 342,000 women died of cervical cancer out of a total of more than 6,00,000. Cervical cancer is the 4th most frequent cancer in women globally, after breast, lung, and colorectal cancers.58 Cervical cancer is one of the leading causes of death in low-income countries.5 In addition, approximately 99% of malignant cervical lesions are caused by HR-HPVs, of which at least 70% are caused by HPV18 and HPV16.59 As most HPV infections are effectively cleared by the host immune system without causing clinical symptoms, persistent infection with high-risk HPV (HR-HPV) types is a necessary—but not sufficient—condition for the development of cervical cancer. Additional cofactors such as viral persistence, host genetic susceptibility, and immunosuppression are required for the progression from infection to malignancy.60
One strand of the circular DNA genome, comprising small, non-enveloped viruses (HPVs), is translated. The HPV genome is approximately 8 kb long. It is divided into three sections: the early area, which encodes all non-structural proteins from E1 to E7 for the regulation of expression of viral genes, as well as a few ones such as E1 and E2 involved in the replication of DNA, and the latter involves encoding structural proteins, that is, L1 and L2, which are necessary for the development of the viral capsid and the long control region (LCR), which is required for transcription and replication.61 Although host proteins are not strictly required for HPV replication, they are still required. In humans, HPVs exclusively infect the epithelial cells (mostly keratinocytes). Only cells in the basal layer of the mucosa, where they first enter the body, have the potential to divide and multiply. Asymmetric cell division in the basal cells replenishes this cell pool. Two cells are produced because of this asymmetric division, one of which is used to replenish the basal cell population and the other leaves the basal layer. Terminally differentiated basal cells leave the cell cycle and DNA replication stops and transitions to the suprabasal layer. Cells specialize as they ascend to the skin’s surface but eventually die.62
This renewal mechanism in the stratified squamous epithelium of the cervix is intricately associated with the HPV life cycle. Sexual contact spreads HPV infection from a man’s anogenital area (mostly the scrotum, foreskin, glans, and corona sulcus) to the genital tract of a woman. Hair follicles or microabrasions allow HPVs to enter deeper layers, and from there, they go to the squamocolumnar junction, where there is only one layer of epithelial cells. HPV binds to the L1 capsid protein and infects basal cells by interacting with proteoglycans that contain heparan sulfate. Both laminin-5 and laminin-6, which promote cell adhesion and act as cellular co-receptors for HPVs, are released by the keratinocytes. Virus particles enter keratinocytes after specific binding at the cell surface via the actin cytoskeleton-dependent endocytic pathway.63 According to Popa et al (2015), the HPV capsid is destroyed inside basal cells, the viral DNA is covered in the L2 protein, the blisters are packed, and the virus is then transferred to the nucleus, where the nuclear envelope is broken down during early mitosis. Promyelocytic leukemia nuclear bodies, also known as promyelocytic oncogenic domains, interact with the HPV L2 protein complex genome during mitosis and nuclear membrane reformation.64 Transcription and replication of HPV and other DNA viruses occur in these nuclear bodies.5
The copies of the viral genome in episomes remain low in infected basal cells during the initial phases of HPV replication.65 HPV DNA replication requires an interaction between the E1 and E2 proteins of the virus and host DNA replication machinery. The E2 Transcription factor recruits DNA helicase E1 to the replication fork of the virus when it binds to specific locations on the LCR DNA. This allows the assembly of the DNA replication complex. The viral genome replicates modestly early in HPV infection, with approximately 50–100 copies of viral DNA per cell. Persistent infection caused by HPV requires the continuous production of viral DNA in host cells. A large amount of the HPV genome is produced when basal cells move towards the epithelial region. This causes the development of progeny viruses, which are shed in the form of virion-laden squames from the infected epithelia. However, the HPV genome is frequently incorporated into the DNA of host cells in premalignant and malignant cervical lesions. The viral life cycle terminates when HPV DNA is integrated because it can no longer produce contagious particles.5 To what extent can HPV maintain DNA replication in cells that have stopped dividing? Infected, growth-arrested host cells can actively multiply HPV due to the virus’s sophisticated tactics to avoid apoptotic signals. Early E5, E6, and E7 proteins.5
HPV-Driven Carcinogenesis Mechanism
In contrast to HPV E6 and E7 oncoproteins, the role of HPV E5 in carcinogenic alterations of the cervical epithelium is not well understood. E5, a short hydrophobic polypeptide produced by both low- and high-risk HPV strains, influences vital processes in cancer development. As previously mentioned, the E5 oncoprotein is involved in HPV-driven cellular transformation during the nascent phase of cervical carcinogenesis’s nascent phases.66 When HPV DNA integrates into the host genome, the viral oncoproteins E6 and E7 are upregulated while the E2 and E5 genes are lost.66 The persistent expression of E6 and E7 leads to immortalization of HPV-infected cells, induction of genomic instability, inhibition of apoptosis, and continued replication of HPV DNA.61.
Furthermore, P53 activity is associated with transformation characteristics of HR-HPV E6. p53 is a TSG that is altered in almost half of all human cancers. By inhibiting the division of cells with impaired DNA and inducing programmed cell death in cancer cells, P53 slows the growth of tumors. When DNA breaks in dividing cells, a critical mechanism called DNA repair prevents cancer development. The modulation of cellular transformation by E6 results in the accumulation of damaged DNA and genome instability.67 This is primarily caused by the absence of effective P53-initiated DNA repair and downregulation of apoptosis in HPV-infected cells. Murine Double Minute 2 (MDM2), an E3 ligase, keeps P53 protein at low levels in healthy cells.5
In addition, P53 accumulates in the nucleus in response to genotoxic stress due to the confluence of cues that work together to inhibit MDM2. P53 is homo-tetramerized and functions as a transcription factor that regulates gene expression during several cellular activities, including apoptosis, cell cycle arrest, and DNA repair. In the G1 phase, cell cycle arrest is induced by stimulating of the P53 pathway, which provides cells with a chance to restore their impaired DNA. However, P53 initiates apoptosis when sufficient DNA impairment is found. Insufficiency of the P53 protein ensures that HPV-infected cells do not undergo activation of these anti-stress biological activities. When the E6-associated protein (E6AP) associated with ubiquitin ligase binds to HPV E6, the complex attracts P53 and promotes P53’s ubiquitinylation and subsequent destruction by the proteasome. The HPV E6 oncoprotein inhibits p53 activity through multiple mechanisms in addition to promoting its proteasomal degradation. E6 interferes with the transcriptional activity of p53 by preventing its phosphorylation and by inhibiting the binding of the coactivator CBP/p300. This disruption shifts the CBP/p300 complex from an activating to a repressive state, thereby preventing p53-dependent gene transactivation and weakening p53’s ability to induce cell-cycle arrest and apoptosis.68
Consequently, P53 and p300 suppress the expression of Ataxia Telangiectasia and Rad3-related (ATR), a gene that encodes phosphatidylinositol 3-kinase, which is crucial for the restoration of UV-impaired DNA. HPVs types 5 and 8 may be associated with non-melanomic skin cancer because E6 upregulates P300 and ATR while simultaneously inhibiting the development of thymine dimers, which increases the carcinogenic impending of UV rays. Infected keratinocytes require HPV E6 for survival. Diverse proteins in the cell are responsible for E6’s immortalizing effects.65 Figure 2 demonstrates the oncogenic behavior of HPV oncoproteins E6 and E7 and their effects on host cell cycle regulation.
 |
Figure 2 Oncogenic behavior of E6 and E7. |
Disruption of the pRB pathway and overexpression of telomerase reverse transcriptase (TERT) contribute to the immortalization of normal keratinocytes. E6 from oncogenic HPVs activates the TERT promoter in cervical cells, increasing TERT activity and is necessary for cervical carcinogenesis.51 It is generally known that E7, another important oncogene, is responsible for the cervical epithelium’s malignant transformation.51 It is a mandatory oncoprotein for transcription in the host, affecting immune evasion and irregular cell cycle progression. E7’s influence on the pattern of expression of host genes is mediated by interactions with many transcription factors and chromatin remodeling complexes.51 These interactions allow E7 to exert a far-reaching effect on the expression patterns of the host genes. The cooperation between E7 and the pRB family was significant.65 The pRB tumor suppressor is well recognized as an essential for the regulation of cellular processes such as cell cycle progression and cell death.65 To release transcription factor E2F and activate the genes necessary for entering the S-phase of the cell cycle, E7 controls the proteasomal degradation of pRB.65
The initial proteins, E5–E7, produced by HPVs are crucial for the malignant transformation of the cervical epithelium because they play a crucial role in the life cycle of HPV. Critical cell cycle checkpoints are inhibited and these proteins increase host genomic instability.69
In addition, other HPV proteins are also partially responsible for the cervical transformation. In human keratinocytes, the intermediate filament network collapses when HPV16 E4 interacts with the cytokeratins. CIN-I (Cervical intraepithelial neoplasia grade I) or LSIL (low-grade squamous intraepithelial lesions) are pre-cancerous squamous lesions in the tissue of cervical that can develop to HSIL (high-grade squamous intraepithelial lesions). This stage includes the earlier entities CIN2, CIN3, moderate and severe dysplasia, and ultimately, cervical. pRB, a tumor suppressor, is well recognized as a crucial regulator of cellular processes such as cell cycle propagation and cell death. To release transcription factor E2F and activate the genes necessary for entering the S-phase of the cell cycle, E7 controls the proteasomal degradation of pRB. The early proteins E5 to E7 produced by HPVs are crucial for the malignant transformation of the cervical epithelium because they play an essential role in the life cycle of HPV.70 Cigarette smoking64 and alcohol use71 are two additional cancer risk factors that support maintenance of chronic HPV infection. HPV infection also causes persistent inflammation, which plays an important role in cancer progression of cancer.72 Pro-inflammatory factors have been implicated in the progression of humans.51
Nanoparticle-Based Strategies for HPV Management
Nanotechnology is revolutionizing the management of HPV-associated cancers by introducing advanced diagnostic, therapeutic, and vaccine modalities. Nanoparticle platforms, including inorganic (gold, silver, silica, zinc oxide), polymeric, and lipid-based carriers, have enabled targeted drug delivery, gene therapy, and immune modulation with higher specificity and reduced systemic toxicity compared to conventional treatments.73 Recent innovations include nanoparticle-assisted gene editing (such as CRISPR/Cas9 and siRNA (small interfering RNA) payloads) targeting HPV E6/E7 oncogenes.73 For example, PEGylated liposomes and pH-responsive cationic nanoliposomes have been shown to deliver CRISPR/Cas9 complexes efficiently to cervical cancer cells, achieving precise genome editing with minimal off-target effects and significant tumor inhibition in vivo.73,74
Nanovaccines are a major focus, leveraging nanoparticles (eg, PLGA (poly(d, l-lactic-co-glycolic acid)), lipid-based, virus-like particles) as carriers for peptide, mRNA, or protein antigens.75 Clinical trials have assessed several nanovaccine candidates such as BNT113 (RNA-lipoplex) and PDS0101 (lipid nanoparticle with E6/E7 peptides), in patients with HPV-positive cervical and head-and-neck cancers, often in combination with immune checkpoint inhibitors.76 These formulations enhance antigen uptake, promote dendritic cell maturation, and trigger robust cytotoxic T-lymphocyte responses, leading to improved tumor regression and survival rates compared to traditional vaccine approaches.75
Additionally, nano-enabled biosensors, such as graphene oxide and carbon nanotube composites functionalized with DNA probes, have markedly increased the sensitivity and specificity of molecular HPV detection, enabling rapid and reliable diagnosis of HPV infections and associated malignancies at early stages.77 Advances in multiplexed colorimetric and electrochemical biosensing further support point-of-care testing and population-based screening for high-risk HPV subtypes.78
Challenges and Limitations in HPV Nanotherapeutics
Despite this progress, multiple biological and technological hurdles limit the clinical translation of HPV nanotherapeutics. Biological barriers include inefficient nanoparticle penetration through tumor tissue, complex interactions with stromal and immune cells, and rapid clearance by the reticuloendothelial system.73 Tumor heterogeneity and the immunologically “cold” microenvironment of advanced cancers can lead to variable therapeutic efficacy and resistance, complicating uniform outcomes in patient cohorts.73
Safety concerns persist around nanotoxicity, encompassing oxidative stress, inflammation, and DNA damage caused by certain nanoparticle formulations.79 Long-term effects such as bioaccumulation, off-target toxicity, and unexpected immune activation must be exhaustively characterized in diverse patient populations using advanced models (eg, organoids, immunocompetent animals).73 Regulatory challenges include the lack of standardized guidelines for nanoparticle safety assessment, as well as batch-to-batch variability due to complex synthesis conditions, which impedes reproducibility and large-scale manufacturing.79 As noted in recent reviews, the absence of unified international protocols for evaluation, approval, and monitoring remains a significant bottleneck for bringing nano-enabled HPV therapies into routine clinical practice.73
Future research should prioritize optimizing delivery routes, minimizing immunogenicity and toxicity, enabling scalable production, and establishing stringent regulatory frameworks for reliable integration of nanomedicines targeting HPV-driven cancers.79 Such advances hold promise for improving outcomes and broadening patient access to precision HPV diagnostics and therapeutics.
Hepatitis Viruses
HCV infection also ranks highly among the causes of chronic liver disease. Approximately 71 million people worldwide were chronically infected with human immunodeficiency virus (HIV) before the advent of direct-acting antivirals (DAAs) treatment.80 Achieving a persistent virologic response, often known as SVR or virological cure, is associated with a significantly reduced risk of developing hepatocellular carcinoma (HCC). A meta-analysis of results from cohorts receiving interferon-based therapy revealed a decline in the likelihood of HCC recurrence of more than 70% after SVR, regardless of the degree of underlying hepatic fibrosis. Despite the continued high risk, individuals with cirrhosis also show a remarkable decrease.81 The landscape of HCV-induced HCC caused by HCV has changed due to new DAAs, and numerous studies have shown that patients who achieve SVR after taking DAAs have a considerably lower chance of developing HCC.82 Elderly, Hispanic ethnicity, male sex, obesity and diabetes, genotype 3 HCV, smoking, alcohol abuse, and co-infection with HBV or HIV are additional factors that enhance the developmental risk of residual HCC in patients with HCV. A developing substantiation advocates that a combination of these indicators may enhance risk stratification and aid in the identification of patients whose risk of HCC is still present despite SVR.83
However, viruses, immune cells, and genetic susceptibility play unique, but complementary, roles in the complicated process of hepatic carcinogenesis. HCC progression has been associated with chronic inflammation of the liver, deregulation of cell signaling, and oxidative stress. Oncogenic viruses often interact with host factors (such as immunological dysregulation) to generate preneoplastic illnesses rather than cancer.84 In the hepatic niche, prolonged inflammation caused by viral infection directly results in enhanced production of reactive oxygen species (ROS) and proinflammatory cytokines.85 According to some researchers, platelet dysfunction and other irregularities in hemostasis can be factors in the development of hepatic cancer.86
Hepatitis B Virus
HBV is a partially double-stranded DNA virus of the Hepadnaviridae family, responsible for acute and chronic liver infection and a major global health burden.87 With over 296 million chronic cases worldwide, HBV is a leading cause of cirrhosis and HCC.87 HBV can be incorporated into the DNA of hepatic cells, resulting in chromosomal rearrangement, genome instability, and mutations in tumor suppressor and proto-oncogene genes.88 In the absence of cirrhosis, this appears to be the primary mechanism by which HBV leads to HCC.89
Molecular Mechanisms Underlying HBV -Induced Tumorigenesis
Oncogenic transformation linked to HBV is multifactorial and involves both direct and indirect pathways. Chronic infection triggers persistent hepatic inflammation and regeneration, which drive genetic and epigenetic changes in hepatocytes. Directly, the HBV genome can integrate into host DNA, causing disruption or activation of cellular genes, particularly those controlling cell cycle and apoptosis. The viral HBx protein is a key effector, known to transactivate cellular oncogenes, inhibit tumor suppressor genes (eg, p53), and alter signaling pathways, facilitating malignant transformation and promoting HCC. Additionally, sustained immune-mediated hepatocyte turnover and microenvironmental changes further increase cancer risk.
In addition, in patients with cirrhosis, various variables are involved in the progression of HBV-associated HCC. Beginning with the NF-kappaB, Raf, c-Jun, MAPK, Jak-Stat, FAK, and protein kinase C pathways, phosphatidylinositol-3 kinase signaling cascades, the Src-dependent, and the PI3K/Akt pathway, the HBV-expressed X protein interacts with a variety of signal transduction and transcription factors.89
Furthermore, tumor-suppressing genes are silenced, and chromosome instability is induced by HBx, which also causes hyper-or hypomethylation of genomic DNA.89 It has both pro- and anti-apoptotic effects, and has been shown to increase TERT expression and telomerase activity.90 Dysregulation of IGF-II11 is associated with the HBx protein. These mechanisms culminate in unconfirmed cell proliferation and malignancy.91
The utmost essential uncontrolled pathways are the PI3K/Akt/mTOR, Wnt/FZD/-catenin, IRS1/IGF1, and MAPK (Ras/Raf/mitogen-activated protein kinases) routes. As a regulator of stem cells, WNT connects with LRP and FZD to prevent β-catenin degradation. Catenin then enters the nucleus and combines with T-cell-specific transcription factor/lymphoid enhancer-binding factor (TCF/LEF) to activate transcription. Unregulated liver cell proliferation, survival, and HCC lead to the synthesis of WNT target genes.91 The same outcome as WNT activation is obtained by upregulating PI3K/AKT and Ras/ERK1/2, which lead to the overexpression and activation of cyclin D1, c-Myc, and NF-B, respectively. In addition, HBV-induced FasL exposure caused by HBV causes hepatocytes to undergo increased apoptosis and compensatory regeneration.92 In Asian countries, aflatoxin exposure and HBV infection are frequently coupled. In these circumstances, mutations in the p53 tumor suppressor gene, which is a gatekeeper characterize HBV-related carcinogenesis.93
Emerging Nanomedicine Strategies Against HBV-Related Cancers
Nanotechnology offers promising solutions for HBV diagnosis, drug delivery, gene therapy, and immunization. Nanoparticles including lipid-based, polymeric, and biomimetic systems, can deliver antiviral agents (such as nucleos(t)ide analogues or siRNA), increasing drug bioavailability, targeting hepatocytes, and overcoming resistance.94 Polymer and lipid nanoparticles can also encapsulate gene-editing tools, such as CRISPR/Cas9, or siRNAs for targeted degradation of viral covalently closed circular DNA (cccDNA) or silencing of key viral transcripts, achieving potent HBV inhibition in preclinical studies.94 Biomimetic nanoparticles guided by peptide ligands (eg, PreS/2-21) further enhance hepatocyte-specific delivery, gene editing precision, and immune evasion.94 Metal nanoparticles and microfluidic platforms have also improved biosensing and early detection of HBV antigens at the point of care.
Challenges and Limitations in HBV Nanotherapeutic Translation
Despite these advances, several translational obstacles remain. Biological barriers, including complex liver architecture and rapid clearance by Kupffer cells, can limit nanoparticle distribution and therapeutic efficacy.94 Long-term safety concerns involve nanoparticle accumulation, hepatotoxicity, immune activation, and non-specific uptake.94 Gene-editing approaches targeting HBV cccDNA or integrated genomes must be assessed for off-target effects, delivery efficiency, and sustained antiviral action.94 Regulatory, manufacturing, and cost-related hurdles complicate clinical adoption; batch-to-batch variability and scalability of multifunctional nanoparticles need to be resolved for widespread implementation. Robust preclinical models that recapitulate the complexity of chronic HBV infection and associated metabolic comorbidities are urgently needed to validate nanomedicine strategies.94 Finally, multi-omics-guided personalization and durable suppression of chronic infection without toxicity remain ambitious but achievable goals for the field.95
Hepatitis D Virus
HDV is a small, defective, circular single-stranded RNA virus and the causative agent of hepatitis D, a severe form of viral hepatitis that only occurs in individuals co-infected with HBV.96,97 HDV requires the hepatitis B surface antigen (HBsAg) provided by HBV for its assembly, replication, and infectivity.98 HDV infection affects approximately 12–20 million people globally and is linked to rapid progression of liver disease, including cirrhosis and HCC.99
Molecular Mechanisms Underlying HDV -Induced Tumorigenesis
HDV drives oncogenesis primarily through chronic hepatic inflammation, immunological dysregulation, and persistent liver injury, leading to fibrosis and ultimately HCC.100 Molecular studies show that HDV-infected livers exhibit upregulation of pathways for DNA damage response, cell cycle progression, and genomic instability, including distinct signatures for sonic hedgehog, GADD45, and mitotic checkpoint control.100 HDV exerts oncogenic synergy with HBV, with transcriptomic analyses demonstrating more pronounced genetic instability and more aggressive malignancy than HBV or HCV alone.100 Additionally, HDV proteins can alter immune surveillance, cell proliferation and apoptosis, facilitating transformation of hepatocytes into a chronic inflammatory microenvironment.100,101
Other processes, such as changes in the immune system response, epigenetic alterations, or ER oxidative stress, may be associated with HDV infection. HDV disrupts the above-mentioned pathways (Smad3, TGF, STAT3, NF-102 to increase cell survival, cell proliferation, and malignant formation. Additionally, HDV prevents an immunological reaction to IFN, which prolongs cell life.103
Emerging Nanomedicine Strategies Against HDV-Related Cancers
Nanotechnology offers promising avenues for HDV management, primarily via advanced drug delivery systems and diagnostics. Polymeric, lipid, and inorganic nanoparticles are being investigated for targeted hepatic delivery of experimental anti-HDV agents such as entry inhibitors, nucleic acid analogues, and siRNA/CRISPR-based therapies.104 Nanocarrier platforms may enhance antiviral compound bioavailability, improve selective uptake by liver cells, and minimize systemic toxicity.105 Additionally, nanoparticle-based biosensors facilitate rapid, sensitive detection of HDV RNA and antigens in patient samples, supporting earlier diagnosis and treatment monitoring.106 Surface-modified nanoparticles and ligand-targeted approaches are under development to refine specificity and reduce off-target accumulation.106
Challenges and Limitations in HDV Nanotherapeutic Translation
Implementation of nanotechnology for HDV remains limited by biological barriers, uncertain safety, and regulatory complexities. Nanoparticle biodistribution in the liver, rapid clearance, and potential immune activation pose translational risks.107 Toxicity from off-target effects (eg, oxidative stress, inflammation) and long-term bioaccumulation remain inadequately evaluated, especially for chronic therapies.73 Manufacturing and scale-up challenges such as reproducibility, stability, and regulatory approval, further constrain the clinical use of nanocarriers.105 The rarity of HDV, diverse patient liver microenvironments, and co-infection with HBV complicate trial design, personalization, and broad adoption, highlighting the need for harmonized safety testing, multi-center collaboration, and long-term biosurveillance.104 Hepatitis C Virus
HCV is a single-stranded, positive-sense RNA virus within the Flaviviridae family that primarily infects liver cells (hepatocytes).108 HCV is transmitted mainly through blood-to-blood contact and represents a major global health challenge, with approximately 58 million people chronically infected and 1.5 million new cases annually.108 Acute infection progresses to chronic hepatitis in the majority of cases, leading to liver fibrosis, cirrhosis, and HCC.108
Molecular Mechanisms Underlying HCV -Induced Tumorigenesis
In contrast to the Hepatitis B virus, human cytomegalovirus does not bind to human DNA. Three main routes are implicated in HCV-induced hepatocarcinogenesis brought on by HCV: immune response deregulation, alterations in the function of antigen-presenting cells, and chronic inflammation.109
HCV’s oncogenicity is multi-factorial. Chronic infection induces persistent inflammation and immune-mediated tissue injury, driving fibrosis and ultimately cirrhosis.110 HCV proteins including core, NS3, NS5A, and NS5B, modulate host signaling pathways involved in cell proliferation, apoptosis, oxidative stress, and lipid metabolism.110 Notably, the HCV core protein and non-structural proteins interfere with key tumor suppressors (such as p53 and Rb), disrupt cell cycle regulation, and activate Wnt/β-catenin and NF-κB pathways, fostering an environment conducive to hepatic malignant transformation.110 The high mutation rate and formation of quasispecies allow HCV to evade immune clearance and sustain chronic infection.110
Furthermore, when the viral protein NS5A interacts with cellular elements,111 two critical cellular processes are altered: protein synthesis (because ER activity is pushed toward the creation of viral proteins instead of cellular proteins) and cell division.112 Oxidative stress is induced by accumulation of fatty acids (long chains) and cholesterol in virus-infected cells. This damage to endoplasmic reticulum function, in turn, triggers the activation of the NFkB pathway.113
In addition, the NRF2-kelck-like ECH-associated protein (KEAP), PI3K-mTOR pathway, angiogenesis, cancer stem cells, and receptor tyrosine kinases are among the downstream pathways that TERT, p53, catenin, Rb, remodeling of chromatin/modifications of epigenetics, hepatocyte differentiation, and HBV-related carcinogenesis follow.109 The NS3 proteins and HCV core may also increase pro-cancerous cytokines such as IFN, IL-1, IL-6, and TNF.
Emerging Nanomedicine Strategies Against HCV-Related Cancers
Nanotechnology is transforming HCV management through innovations in targeted drug delivery, molecular diagnostics, and vaccine development. Nanoparticle formulations including polymeric, lipid-based, and inorganic carriers, enhance the stability, hepatic targeting, and effectiveness of DAAs, siRNA, and anti-HCV combinations, allowing for reduced dosage and systemic toxicity.114 For example, nanoparticles functionalized with galactose or hyaluronic acid ligands facilitate hepatocyte-specific delivery of anti-HCV therapeutics.114 Nanoparticle-based RNA interference strategies have shown potent inhibition of HCV replication and protein expression in preclinical models.115 Furthermore, nanoparticle-supported biosensors including gold nanoparticles and quantum dot platforms, enable rapid, sensitive, and multiplexed detection of HCV RNA and antigens in clinical samples, improving early diagnosis and monitoring.114 Nanoparticle-based HCV vaccines (especially those presenting the E1E2 glycoprotein complex) are being investigated for their ability to induce strong humoral and cellular immunity, although clinical translation remains pending.116
Challenges and Limitations in HCV Nanotherapeutic Translation
Key challenges for HCV nanomedicine include biological barriers to liver-specific delivery, potential nanotoxicity, immune responses, and long-term biosafety concerns.114 The rapid hepatic clearance, non-specific uptake by the reticuloendothelial system, and variability in patient microenvironment limit therapeutic efficacy and consistency.114 Adverse events linked to certain nanoparticle types include oxidative stress, inflammation, and unintended cellular injury.114 Manufacturing standardization, regulatory approval, and batch reproducibility remain obstacles to clinical translation and large-scale production.114 Another critical roadblock is the absence of an effective, licensed HCV vaccine despite promising nanoparticle-based approaches.114 Overcoming these issues requires improved nanoparticle design, robust preclinical models, and harmonized international guidelines for nanomedicine evaluation.117
Nanoparticles and Human Viral Infection
Viruses are among the most impactful disease-causing agents with high pandemic potential, as demonstrated by the global burden and economic costs of repeated epidemics and outbreaks.118 The most common viral disease outbreaks in humans are associated with animal reservoirs.119 Around 11 of the 14 notable epidemics that humanity has encountered globally over the past 120 years have been caused by viruses. In addition, several viral diseases and infections, such as the common cold, can result in significant economic losses worldwide, apart from significant morbidity and mortality rates.120 Influenza A and B viruses, metapneumovirus, parainfluenza viruses, rhinovirus, coronavirus, enterovirus, and syncytial respiratory viruses are the primary viruses that cause lower respiratory tract infections.121
Conventional antiviral treatments often suffer from poor target specificity, drug resistance, and undesirable side effects, necessitating the development of more effective alternatives.114 Nanotechnology presents a promising frontier in viral disease management by enabling the design of NPs with tunable physicochemical properties that facilitate advanced antiviral strategies. The versatility of NPs arises from their ability to directly inactivate viruses, inhibit viral attachment and entry through receptor blocking, deliver antiviral drugs and genetic material precisely to infected cells, and even modulate host immune responses.114 Recent studies report that gold, silver, silica-based, polymeric, and lipid-based NPs can be engineered for multifunctional antiviral and vaccine applications, significantly improving bioavailability, maximizing tissue targeting, and reducing systemic toxicity.122 NP-enabled point-of-care diagnostics, including biosensors and rapid antigen tests, have become essential for timely virus detection, outbreak control, and pandemic preparedness.122 Few viruses, such as MERS, respiratory syncytial virus, and influenza A virus, have been treated with NPs. NPs used to treat viral infections are frequently inorganic and functionalized, and are used first as antimicrobials alone and then as antimicrobials plus nanocarriers.123 The principal types of nanoparticles used in cancer therapeutics are visually summarized in Figure 3, which categorizes nanoparticles based on composition and biomedical application.
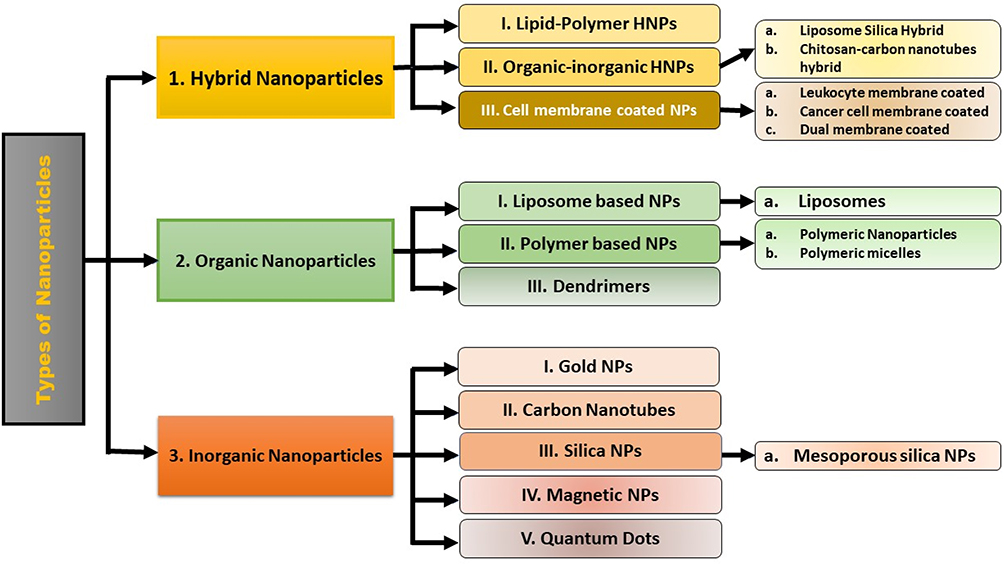 |
Figure 3 Types of nanoparticles used in cancer therapeutics. |
Antiviral techniques mediated by nanoparticles can be subdivided into four major groups: inactivation of viruses, blockage of viral entry, suppression of viral replication, and inhibition of apoptosis. While most existing data come from in vitro studies, there is increasing emphasis on in vivo research including inhaled and intranasal nanomedicines for respiratory viruses, to assess bio-distribution, targeted delivery, and safety in physiological contexts.114
One significant recent advancement is the success of lipid nanoparticles in vaccine technology, notably in the Messenger RNA (mRNA) vaccines developed for SARS-CoV-2, which have demonstrated high efficacy, rapid scalability, and strong immune activation.124 Nanocarriers such as dendrimers and polymeric NPs are also being investigated for nucleic acid delivery (eg, siRNA, DNA, mRNA) against emerging and persistent viral threats, enabling genome-level intervention and long-lasting immunity.124. The contribution of nanomedicine to SARS-CoV-2 infection is significant. SARS-CoV-2 has resulted in millions of illnesses and fatalities125 and is developing as a major risk to healthcare and a global budget of.126
Recent advances in nanotechnology have revealed multiple, distinct mechanisms by which nanoparticles exert antiviral effects. Inorganic nanoparticles such as silver nanoparticles (AgNPs), gold nanoparticles (AuNPs), zinc oxide, and cerium oxide inactivate viruses through direct binding to viral envelopes or capsids, generation of ROS that damage viral proteins and nucleic acids, and disruption of viral entry into host cells.127 These effects have been demonstrated against both enveloped viruses (eg, influenza, herpes simplex, coronaviruses) and non-enveloped viruses, though the latter often require longer exposure or higher concentrations for inactivation. Specifically, nanoceria (CeO2) disrupts viral lipid bilayers and adsorbs viral nucleic acids, effectively inactivating a broad range of pathogens.127
Nanoparticles also serve as efficient drug and nucleic acid delivery vehicles, improving stability, cellular uptake, and targeted release of therapeutics.122,128. Polymeric, lipid-based, and dendrimer nanoparticles have been successfully used to deliver antiviral drugs, mRNA vaccines, and gene-silencing agents (eg, siRNA) with precise tissue targeting and controlled release For instance, the lipid nanoparticle platforms used in mRNA COVID-19 vaccines have set a new standard for rapid vaccine deployment and robust immune activation.122
Clinical studies and early-phase clinical trials have begun to translate these nanotechnologies into real-world antiviral therapies.128 Nanoparticle-based formulations of established antiviral drugs have demonstrated improved viral suppression and immune responses in patient populations with HIV and viral hepatitis, and nanoparticle-based peptide and subunit vaccines have been shown safe and immunogenic in recent trials against dengue and COVID-19.128 Immunomodulatory nanoparticles, such as VLPs or nanocarriers loaded with Toll-like receptors (TLR) agonists, are actively being developed to boost host antiviral defense mechanisms and provide cross-protection against viral variants.128
Nanotechnological strategies for combating viral infections can be broadly classified into three core categories: diagnostic technologies, vaccine platforms, and therapeutic interventions.129 Recent advances include the engineering of ACE2-coated quantum dots and surface-functionalized nanoparticles. These innovations have been utilized not only for developing rapid point-of-care viral diagnostics and biosensors but also for producing personal protective equipment (such as antiviral gloves, masks, and nasal filters) and even antiviral chewing gum capable of binding and neutralizing respiratory pathogens like SARS-CoV-2, thereby preventing viral transmission.129 Viral gastroenteritis, especially in Western countries,130 is most frequently caused by rotavirus, norovirus, and adenovirus, and is associated with major health and economic burdens. Importantly, the mortality rate for viral diarrhea is inversely correlated with a country’s level of socioeconomic development, reflecting disparities in healthcare access and sanitation.131 While nanoparticle-based therapies remain under exploration, certain nanoplatforms such as gold/copper sulfide (Au/CuS) core/shell nanoparticles, have shown efficacy in inactivating norovirus GI.1 in vitro,124 and novel nanoparticle-based vaccines (eg, rotavirus VP6-ferritin NPs and norovirus-rotavirus recombinant PEG NPs) have exhibited promise in preclinical models.131
Although these approaches are promising, translational challenges remain. Two critical concerns for oral and gastrointestinal nano-therapies are: (1) the interactions between administered nanoparticles and the intestinal mucosa, which can affect absorption and host-microbiome dynamics; and (2) the need to fully evaluate the impact of NPs on natural ecosystems, given their persistence, potential toxicity, and wide dispersibility. The ideal delivery route for antiviral nanoparticles targeting enteric viruses is oral administration, pending rigorous in vivo validation.124 The therapeutic approaches utilizing nanoparticles for airways viral infections are systematically outlined in Table 1, including key sites of action, viral targets, and types of nanoparticles employed.
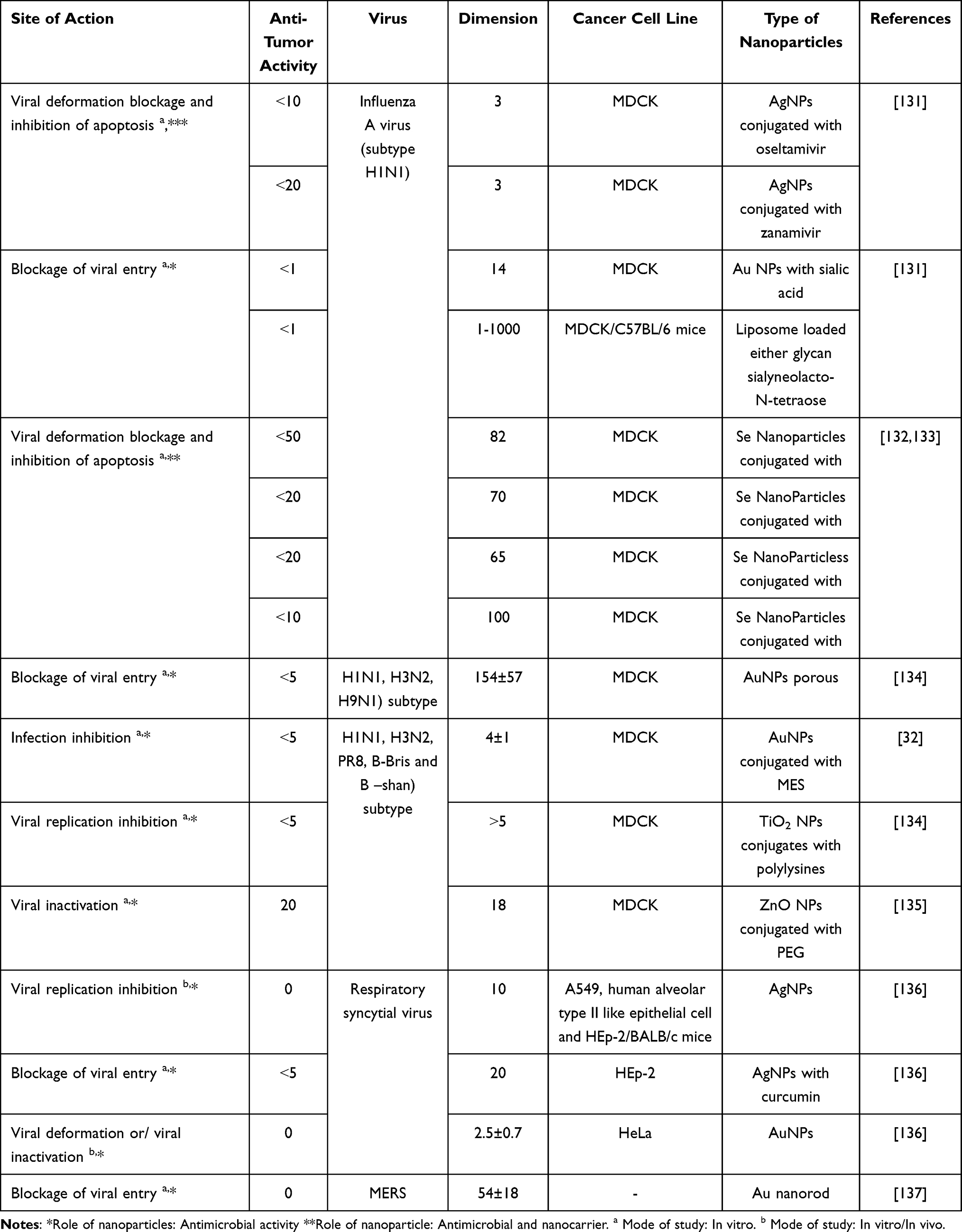 |
Table 1 Nanoparticles for Airways Viral Infections Treatment Methods |
STIs caused by viruses including HSV, HIV, HBV, and HPV represent a significant global health challenge. Additionally, HCV has become increasingly important due to changes in risk behaviors.138 Many of these infections remain incurable, emphasizing the urgent need for advanced antiviral therapies. While current HIV treatments rely on combination antiretrovirals—targeting integrase, CCR5, fusion/entry, protease, and reverse transcriptase—resistance and treatment failure remain major obstacles.139
In this context, NP-based strategies have been developed for HIV and other STIs, utilizing a range of mechanisms: viral inactivation, entry inhibition, cryoprotection, suppression of viral replication, and even breaking viral latency.134 Table 2 provides an overview of nanoparticle-based strategies for treating STIs with viral etiology, listing sites of action, target pathogens, and nanoparticle formulations. Among the most practical and near-term applications is the incorporation of nanoparticles into topical products (eg, lubricants), providing local protection against sexual transmission.134 Since the use of NPs related to non-CD4+T cells has not proven to be effective in eliminating the infection and its use to prevent viral invasion, damage the virus, and worsen the situation, the use of NPs to treat HIV is still under consideration.139 Resistance to antiretroviral medication may be the leading cause of HIV treatment failure may turn out to be antiretroviral medication resistance. However, for systemic antiviral use, in vivo studies, clinical trials, and mechanistic investigations are essential to verify efficacy, safety, and targeted delivery for each virus type.134
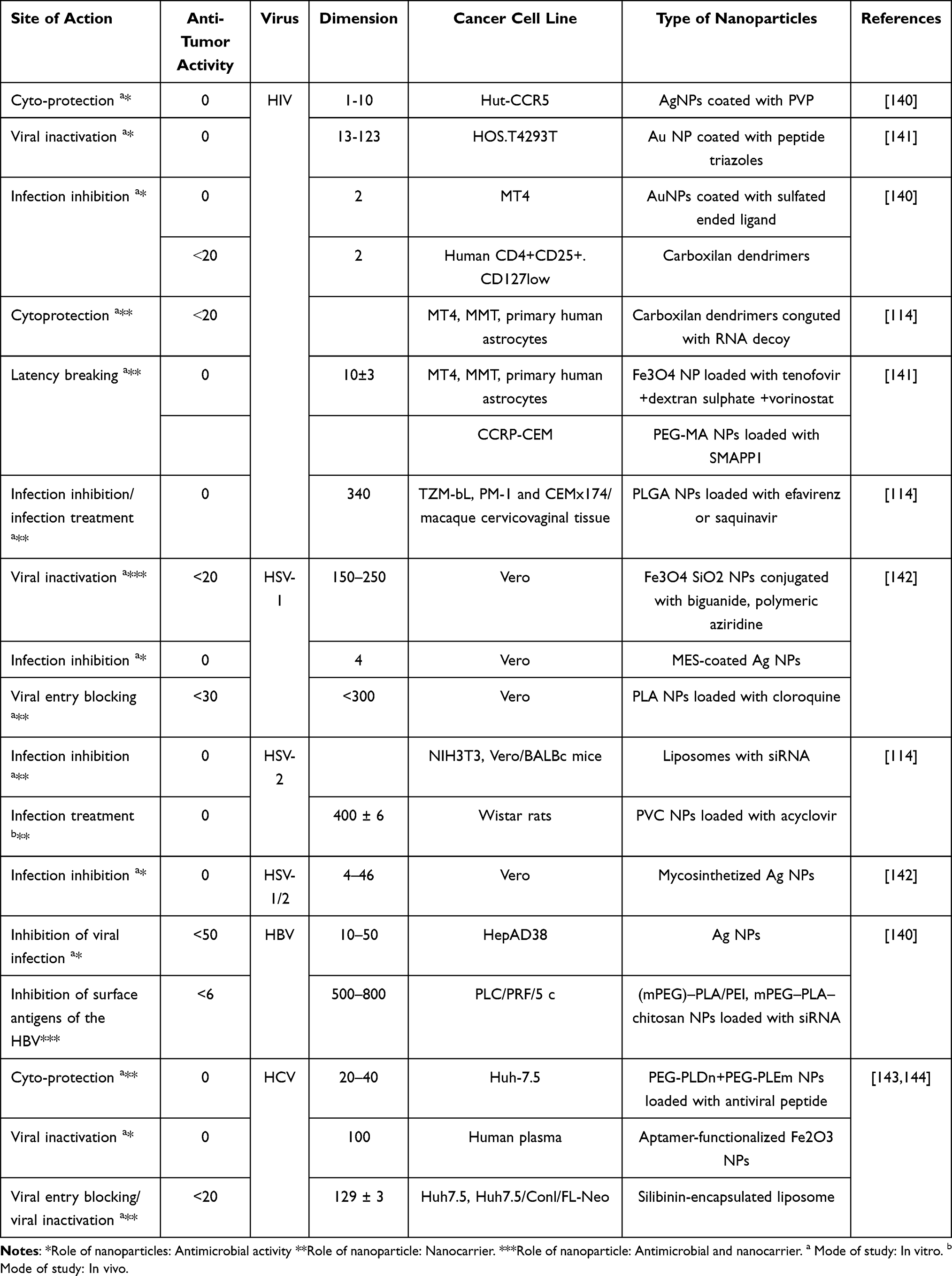 |
Table 2 Nanoparticles for Therapeutic Methods Against STIs with Viral Infections |
Herpes genitalis, resulting from HSV-1 or HSV-2 infection, remains difficult to eradicate due to the viruses’ ability to establish latency in sensory nerves and to reactivate under various conditions.142 Nanoparticle-based approaches, particularly silver, gold, and dendrimer nanoparticles, have shown promising results in inhibiting HSV infection by directly inactivating virions, blocking viral entry, and interfering with replication largely in in vitro studies.142 Translation to in vivo and clinical studies is necessary for real-world application, and differentiation between HSV-1 and HSV-2 infection remains clinically relevant for prevention strategies.
Emerging research demonstrates that nanoparticle-based platforms for HPV130 and HBV.139 predominantly focus on vaccine delivery and immune modulation.145 For hepatitis B, primary nanoparticle-based interventions include the use of nanocarriers for viral inactivation, inhibition of entry, and cytoprotective drug delivery145,146 Acute hepatitis is the primary cause of HBV infection, which rarely progresses to fulminant hepatitis. Instead, HCB often results in chronic infections, rather than acute icteric hepatitis.139 Within the next 10–50 years, infection may cause 20% or more of the population to develop cirrhosis or liver cancer.139
Nanoparticles can be engineered with various surface modifications and functionalizations including size, shape, and charge, to enhance their uptake by the liver. For example, nanoparticles that are positively charged and larger than 200 nm in diameter are more efficiently absorbed by hepatic cells,147 as the liver’s unique vascular structure and resident macrophages preferentially sequester such particles. However, excessive accumulation or uptake of these nanoparticles can induce inflammatory responses in the liver, which may contribute to the development of hepatitis or liver injury,148.
Ongoing advancements in nanomaterials, such as PEGylated and targeted nanoparticles, hold significant promises for local and systemic antiviral interventions, but require careful assessment of biocompatibility, long-term safety, and environmental impact.122 Despite these advances, translational challenges remain, including the need for long-term toxicity assessment, biodistribution modeling, immune compatibility, and environment impact studies for large-scale NP deployment.114 Careful nano-bio interface engineering, regulatory harmonization, and ecotoxicological assessment will be critical to realize the full therapeutic potential of antiviral nanomedicine.
Magnetic Silica Nanoparticles
Magnetic mesoporous silica nanoparticles (MSNs) represent a significant advance in nanomedicine, offering a large surface area and adjustable pore size that facilitate high loading and efficient delivery of antiviral agents.149 Their inherent biocompatibility ensures minimal toxicity and robust interaction with biological environments, making them suitable for clinical applications in viral infections. Magnetic properties introduced by iron oxide components allow MSNs to be guided by external magnetic fields, resulting in precise, site-specific delivery to infected tissues and organs, a feature that greatly enhances treatment efficacy while reducing off-target effects.150
These nanoparticles are extensively investigated as smart carriers for both antiviral drugs and vaccine antigens, with their mesoporous structure enabling controlled, sustained release profiles tailored to maximize therapeutic effects and minimize dosing frequency.150 The external manipulation enabled by magnetic responsiveness permits non-invasive concentration of therapeutics directly at sites of infection, thereby improving local drug concentrations and decreasing systemic toxicities compared to conventional approaches.150 Recent experimental evidence demonstrates that virus-like magnetic MSNs loaded with antiviral compounds or protein antigens robustly stimulate immune responses, leading to significant reductions in viral titers, enhanced pathogen clearance, and improved survival rates in animal infection models.151
Moreover, the possibility of multi-functionalization allows magnetic MSNs to carry imaging agents or multiple therapeutic moieties, enabling simultaneous diagnosis, monitoring, and treatment of viral diseases, a strategy that reflects the growing field of theranostic nanomedicine.150
Organic Nanoparticles: Liposomes, Polymeric Nanoparticles & Dendrimers
Organic nanoparticles, including liposomes, polymeric nanoparticles, and dendrimers, are being widely adopted for antiviral therapy due to their customizable architecture, ability to encapsulate diverse drug molecules, and superior safety profiles compared to many inorganic carriers.114
Liposomes are spherical vesicles composed of phospholipid bilayers that can load both hydrophilic drugs in their aqueous core and hydrophobic drugs within their membrane, providing protection from enzymatic degradation and controlled release at infection sites.124 This versatility permits liposomal formulation of numerous antiviral agents, which has demonstrated improved stability and prolonged circulation time in vivo. Targeted and sustained delivery using liposomes has enhanced therapeutic efficacy against viruses such as influenza, HIV, and herpes simplex virus.114
Polymeric nanoparticles, made from biodegradable materials such as PLGA (polylactic-co-glycolic acid) and chitosan, can be engineered in size, surface charge, and composition to optimize mucosal penetration and targeted drug delivery.114 These carriers have demonstrated notable benefits in delivering antivirals across mucosal barriers such as those found in the respiratory and gastrointestinal tracts, resulting in improved protection and treatment outcomes in preclinical models. Moreover, polymeric nanoparticles enable the development of next-generation vaccines through antigen presentation and immune modulation.114
Dendrimers are highly branched, tree-like polymers with precise control over molecular size, shape, and surface functionality, allowing multivalent interactions with viral envelopes and efficient packaging of nucleic acid therapeutics.114 Dendrimer-based antivirals have shown effectiveness by directly inhibiting viral entry and replication, including against HIV and respiratory viruses, and by facilitating delivery of RNA-based or DNA-based genetic materials for therapeutic intervention.114 These nanocarriers demonstrate enhanced pharmacokinetics, cellular uptake, and specificity, significantly advancing the prospects for effective antiviral treatments and novel vaccine designs.114
Carbon-Based and Other Nanoparticles
Carbon-based nanoparticles including carbon nanotubes and graphene oxide, are gaining recognition in antiviral research due to their distinctive surface chemistry, high aspect ratio, and ability to interact directly with viral structures.114 These materials can physically disrupt viral envelopes and capsids or bind to viral surface proteins, thereby preventing viral attachment, entry, and replication in host cells.114. Additionally, graphene oxide exhibits excellent adsorption capacity and can be functionalized for targeted delivery of antiviral drugs, enhancing their efficacy and minimizing off-target effects.114 Research has shown that carbon nanomaterials are effective against a range of viruses, including influenza, dengue, and herpes simplex virus.152
Cerium oxide nanoparticles, due to their unique redox properties, act as efficient ROS scavengers, protecting host tissues from viral-induced oxidative stress and inflammation.153 Their potent antioxidant activity stabilizes cellular membranes and mitigates the damage associated with viral replication and immune responses, making them valuable adjuncts in both antiviral drug development and supportive care.153 Recent studies have also indicated that cerium oxide nanoparticles can modulate innate immunity and inhibit viral replication, further broadening their potential in nanomedicine-based antiviral applications.154
Inorganic Nanoparticles
Inorganic NPs are novel candidates for new therapies because of their excellent biocompatibility and regulated biodistribution patterns. Inorganic NPs are best used for detecting substances for biosensing and drug delivery, whereas drug delivery systems control drug release in the intended organ to control undesirable supply to undesirable areas.155 The most popular NPs that can be used in cancer therapeutics are addressed by how they target specific tumor sites, their toxicity, and their effectiveness when used in humans.
Silver Nanoparticles (AgNPs)
Because of their unique optical, electrical, and thermal characteristics, AgNPs are superior to many other metal NPs in many ways. AgNPs have been found to exhibit potent antibacterial and specific inhibitory effects, which are supported by their rich physicochemical characteristics, simplicity of synthesis, adaptability, and intriguing therapeutic potential. Owing to advancements in radiotherapy, thermal ablation, medication, gene delivery, and their sensitive diagnostic capabilities, AgNPs are versatile agents with a wide range of uses.156 Their physicochemical characteristics indicate that they have therapeutic effects on several types of cancerous cells. AgNPs are being used as a paradigm shift in the treatment of oncogenic viral diseases by acting as outstanding bioimaging and sensing agents. AgNPs are synthesized using various methods, such as biological synthesis using bacterial strains, chemical synthesis with medicines or chemotherapeutics, and green synthesis of algae and plants. Various cancer cell lines, such as MCF7 (breast cancer cells), A549 (lung cancer cells), HeLa (cervical cancer cells), HepG2 (liver cancer cells), and HCT-116 (colon cancer cells), are susceptible to these methods.123
Gold Nanoparticles (AuNPs)
Nanoscale particles (1–100 nm) with distinct characteristics influence various industries including chemistry and biology. Owing to their particular modes, Nanoparticles have the potential to be an ideal tool for nanomedicine because of their interactions with ligand molecules, regulated distribution patterns, cytotoxicity reduction, and monodispersity. Several diagnostic uses, such as cancer therapy, require AuNPs.157 There are several methods for producing NP, including chemical, green, and biological methods. Plant-based green synthesis is the most popular option because it is safer, less expensive, and more environment-friendly. This type of heating exhibits anti-tumor activity and can be combined with botanicals, stabilizers, chemotherapeutics, and several reducing agents to enhance their properties.158 A plasmonic reverberation shift from 520 to 800–1200 nm (complex forms) was brought about by combining NPs of a particular size and suitable form with gold NPs. In infrared light, the tissues of the body become transparent, giving a range between 800 and 1200 nm the potential to have therapeutically positive effects in deep tissues using photothermal and photoimaging techniques, but this will rely on the effectiveness of these techniques.158
Another important characteristic was the accessible surface area. Notably, nanoparticles have a very high surface-area-to-volume ratio because their surface area is inversely proportional to their size. Its significance continues to grow because of its compatibility and response.158 Various gold NPs such as nanorods, nanocages, nanostars, nanocubes, and nanospheres have become effective tools for studying human cancer and cell biology. AuNPs have been fabricated with various morphologies and sizes, including nanorods, nanospheres, nanocages, nano-shells, and nanostars. These AuNPs depend on the planning technique used. The enormous potential for PTT/PDT, drug distribution, and malignant growth bioimaging provided by these AuNPs has electrical and optical properties for simple surface functionalization.159
Mechanism of Action of AgNP and AuNP
Acute myeloid leukemia, epidermoid larynx carcinoma, colon cancer, oral squamous cell carcinoma, lung cancer, breast cancer, cervical cancer, and other cancers have been the subject of extensive in vivo and in vitro studies of the anticancer effects of AgNPs. The surface charge, size, and form are characteristics that naturally occur in AgNPs. The particles should be very small to produce optimal high biological reactivity, preferably at the nanoscale. Smaller particle sizes exhibit better biological responses. In addition, longer exposure times and higher doses produce more potent anticancer actions. AgNPs have been combined with CPT and carboplatin (anticancer drugs used in breast, colon, lung, and cervical cancers).160 Medications combined with AgNPs dramatically lower or restrict the capacity of malignant cells to proliferate, diminish cell viability, and alter the cell shape.159
AgNP-containing drugs increase cytotoxicity by destroying the mitochondrial membrane, inducing cell death, and increasing ROS oxygen species levels. GSH maintains ROS levels throughout the early stages of apoptosis; however, when GSH spikes dangerously, it throws the cells’ redox equilibrium out of balance. The combination of the drug and AgNPs increased the efficacy of p35 and caspases while decreasing the influence of Bcl-2 proteins. By altering Akt expression, they control the signaling molecules involved in cytotoxicity, cell survival, and cell death.159 By inducing mTO phosphorylation, Akt signaling increases tumor development, cell size, and proliferation. Inhibition of Akt expression and phosphorylation in cancer cells was achieved by conjugation with AgNPs. Owing to faulty tumor vasculature, nanoparticles can aggregate in tumor cells and have an enhanced permeability and retention (EPR)) effect without the addition of a targeted ligand molecule, which is significant for antitumor efficacy. This results in less lymphatic drainage, less interstitial fluid absorption, and a preference for the retention of NPs in the tumor, which damages cancerous cells. Scientists are increasingly interested in the use of noble metals.161
They focused on AuNPs, which are helpful for bioimaging, cancer therapy, medication administration, and other applications.162 NPs bind to appropriate surface ligands that lead them to tumor cells to confirm the killing of cancer cells at a specific place. There are two alternative methods of targeting cancer cells. The first method involves conjugation of AuNPs with polyethylene glycol (PEG), which inhibits accretion and improves their retention in the circulation. The 2nd method involves conjugating antibodies specific to AuNPs that bind to different biomarkers to identify cancer cells. These strategies can enhance cellular uptake and target more efficient cancer treatment.
Nanoparticle Toxicity and Safety Considerations in Cancer and Viral Therapy Nanoparticle-based therapeutics have greatly advanced antiviral and cancer therapy, but their clinical application requires a robust and evidence-driven appraisal of toxicity and potential adverse effects.163 Toxicity profiles of NPs are highly variable and depend on parameters such as particle size, shape, composition, surface chemistry, and dose. Various morphologies, such as needle-shaped or spherical particles, may elicit different cellular responses and degrees of apoptosis. Dose and duration of exposure remain critical, with longer treatment times and higher doses often increasing cytotoxic potential.163 In addition, emerging evidence reveals that multiple physicochemical parameters including size, shape, and especially surface charge, modulate NP toxicity by influencing their cellular uptake and interactions with biological membranes.1
Among the five most common metallic NPs (Ag, MoO3, Al, Fe2O4, and TiO2), it has been determined that AgNPs are the most dangerous because of their high concentration and alteration of NP size. As the concentration dependence increases, lactate hydrogenase leaks into the membrane, disrupting mitochondria’s functions.164 Some potential considerations regarding AgNPs could affect their toxicity in the body and restrict their use. These considerations could include the non-specific distribution of NPs that cause cytotoxicity in many organs and varying NP sizes, such as how easily small NPs can overcome barriers such as the testis barrier and blood-brain barrier and potentially cause cytotoxic effect.157 Owing to their larger size and slower release of Ag ions, AgNPs showed toxicity at higher concentrations, starting at an average length of 40 nm. AgNP cytotoxicity has been reported in vitro to have both short- and long-term harmful properties that manifest in vivo. AgNPs have been used in several in vitro experiments and are thought to be the most toxic and biologically reactive.164 In addition, it was discovered that the MCF7 cells were cytotoxic to AgNPs of 5 to 50 nm size at a concentration of 60 g/mL after incubating the cells for 20 minutes with average inhibitory concentrations and that the AgNPs also increased the oxidative stress response, which is essential for the NP-induced apoptosis.165
Furthermore, positively charged NPs more strongly disrupt membrane integrity and induce oxidative stress, leading to increased cytotoxicity compared to neutral or negatively charged NPs.1 Concerns have been raised regarding the possible cytotoxic effects of AuNPs on biological structures.1 Recent research highlights that gold NPs exhibit toxicity based on surface charge, which significantly influences cellular uptake and biological responses. Strongly charged particles (especially cationic) display greater toxicity, destabilizing membrane integrity and triggering excessive cellular stress.1 Given the unpredictable and heterogeneous nature of human tissues and cells, it is essential to examine the biological effects of each AuNP formulation rigorously before clinical use.1 Therefore, advancement and regulatory approval of AuNPs for cancer theranostic applications urgently require standard, validated testing protocols that incorporate comprehensive toxicity screening, surface modification strategies (eg, PEGylation), and an understanding of protein corona formation.1
A major mechanism of NP-induced toxicity is the generation of ROS. Metallic nanoparticles including silver, gold, copper oxide, and cerium oxide, are known to generate ROS, which can disrupt the cellular redox equilibrium, induce lipid peroxidation, damage mitochondria, and prompt cell death.166,167 This oxidative stress can activate signaling cascades, for example, the NF-κB pathway—that fuel local and systemic inflammatory responses and further disrupt tissue homeostasis. Notably, dose-dependent toxicities are common: while low NP concentrations may have therapeutic or negligible effects, higher doses and chronic exposures are much more likely to trigger harmful biological outcomes such as apoptosis or necrosis.168 Notably, silver nanoparticles have demonstrated selective cytotoxic effects preferentially on malignant, virus-associated cells through ROS-mediated cell death, but careful dose optimization is necessary, as normal cells may also be affected.123 Cerium oxide nanoparticles, although widely used for their antioxidant properties, can paradoxically induce oxidative stress and membrane damage at higher doses in cancer models.169 These effects underscore the importance of judicious nanoparticle design and dosing in clinical applications.
Beyond oxidative stress, nanoparticles may induce autophagic flux disturbances and endoplasmic reticulum stress, disrupting protein folding, cellular trafficking, and potentially leading to apoptosis or unpredictable immune reactions.166 Genotoxic effects, DNA strand breaks, chromosomal aberrations, and altered cell cycle progression, are now well documented both in vitro and in vivo.166 Spherical NPs generally pose less risk than rod-shaped or needle-like constructs due to differences in cellular uptake and mechanical disruption mechanisms.166
Biodistribution and bioaccumulation are additional factors influencing chronic toxicity, with nanoparticles sometimes persisting or accumulating in key organs.170 This can result in organ-specific toxicity manifesting as nephropathy, hepatotoxicity, neurotoxicity, or pulmonary damage.170 The EPR effect can facilitate selective accumulation in tumor tissues, offering therapeutic advantages, but also carries a risk for adverse tissue reactions and off-target toxicity.163 For polymeric and organic nanoparticles, toxicity may also arise from degradation by-products and altered immunogenicity, making thorough characterization essential prior to clinical deployment.163
Furthermore, nanoparticles can cross physiological barriers and accumulate in susceptible organs, as demonstrated for AgNPs in the brain, liver, and spleen, increasing the risk of chronic toxicity, neurotoxicity, and immunological consequences.1,171 Therefore, analyzing NP biodistribution is essential for predicting uptake and toxicity profiles during in vivo applications. When administered by different routes, the potential for bioaccumulation can escalate cellular injury, a risk eliminated neither by novel nor traditional delivery methods.1,171 Surface charge and protein corona formation also play crucial roles, as the adsorption of proteins and biomolecules onto NP surfaces (“protein corona”) can alter biological identity, immune recognition, and toxicity 206,207. A schematic comparison of conventional versus nanoparticle-based therapeutics is presented in Figure 4 to highlight the advantages of nanomedicine over traditional approaches.
 |
Figure 4 Schematic representation of the conventional vs nanoparticle-based therapeutics in various cancers. |
Stability, agglomeration, and shelf-life of AgNPs critically impact their toxic potential. Aging may alter the release of ionic silver, thereby raising the toxicity. Currently, there remain knowledge gaps about the exact mechanisms and long-term health effects of AgNPs and their aggregation in organs, necessitating more comprehensive in vivo studies and long-term follow-up to clarify their safety profiles.1 The cytotoxic effects of AgNPs also depend on attributes such as shape, concentration, and size. The potential cytotoxicity of NPs should be thoroughly studied to better understand their biocompatibility. AgNP toxicity can be decreased in two ways: by lowering the concentration of silver ions and by removing silver NPs. These strategies have been used to reduce the toxic environmental effects of NPs.172
Furthermore, organ-specific effects have been highlighted in recent human and animal studies.173 In the liver, inorganic and carbon-based NPs can trigger hepatocyte inflammation, apoptosis, and fibrotic changes through mitochondrial damage and immune activation.174 Renal toxicities can include glomerular damage and tubular degeneration linked to ROS and direct uptake.173 NPs crossing the blood-brain barrier have been implicated in neurotoxicity, influencing neurotransmitter release, synaptic plasticity, and may cause behavioral alterations.175 Cardiovascular effects include endothelial dysfunction, pro-thrombotic activity, and arrhythmogenic risks, notably for metal oxide and carbon NPs.176 In the reproductive system, both male and female, disruption of hormonal regulation and antioxidant defense have been observed, leading to impaired fertility following NP exposure.175
The evaluation of nanoparticle safety relies on standardized in vitro and in vivo assays such as cell viability, oxidative stress biomarker analysis, and immunological profiling. Preclinical animal models are vital for assessing biodistribution, chronic toxicity, and long-term effects, preceding any human trials.177 Modern approaches emphasize “safe-by-design” strategies, utilizing surface functionalization, optimal particle sizing, and tailored dosing regimens to maximize efficacy while minimizing off-target toxicity.178 Ongoing regulatory surveillance and harmonization remain essential, given the complexity and novelty of nanomedicines.177
Mechanistic studies have shown that NPs can trigger cellular damage by promoting oxidative stress, disrupting mitochondrial function, altering autophagy flux, and inducing inflammation, which may ultimately result in apoptosis, genotoxicity, or even dysregulation of homeostatic pathways such as protein folding or antigen presentation.179 The balance between therapeutic efficacy and potential adverse effects is central to safe clinical translation, requiring multidisciplinary risk assessment and continuous post-market monitoring.1
While clinical studies in humans remain comparatively limited, available evidence from occupational, therapeutic, and environmental exposure supports caution.180 Reported effects range from transient inflammation and respiratory impairment to longer-term cardiovascular, hepatic, or neurological changes associated with chronic or high-dose NP contact.180 This underscores the need for robust toxicity screening, continued post-market monitoring, and standardized regulatory assessment for NP-based medical products.
Multifunctional Nanoparticles: Clinical Applications and Future Directions
Emerging nanomedicine platforms are being engineered for multi-functionality, combining antiviral, anticancer, and immunomodulatory effects within a single nanosystem. Multifunctional nanoparticles, such as antibody-conjugated, magnetic, and stimuli-responsive carriers, can enable the precise and simultaneous delivery of diverse therapeutic payloads,181 including chemotherapy, RNA-based drugs (siRNA or mRNA), and even CRISPR/Cas9 gene editors, directly to virus-infected or transformed cells.182 These nanocarriers are also being developed for stimuli-triggered release, enhancing selectivity and minimizing off-target toxicity. Such features hold promises for improving the efficacy and safety of therapies against virus-induced malignancies.114
For instance, multifunctional nanoparticles can achieve active or passive targeting, imaging, hyperthermia, and drug delivery, thereby overcoming biological barriers and heterogeneity in tumor tissues and viral reservoirs. Disease-targeting integrated nanosystems employing both magnetic and signaling components such as gold nanorods for photothermal activation and iron oxide nanoworms for drug targeting, allow for amplified and site-selective accumulation in tumor regions, as demonstrated in preclinical mouse models.183,184 Furthermore, magnetic nanoparticles (MNPs) have demonstrated precise site-specific drug and gene delivery when guided by external magnetic fields, reducing systemic toxicity and improving local efficacy.185 The incorporation of targeting ligands such as antibodies further enhances specificity; antibody-conjugated MNPs have been shown to accumulate preferentially in tumor cells, leading to enhanced detection and simultaneous therapy.185,186
As detailed in Gautam et al (2024), targeted nanoplatforms not only enable tissue-specific accumulation through active and passive mechanisms, but also facilitate real-time imaging and drug delivery by integrating diagnostic and therapeutic (theranostic) capabilities.182 Engineered nanostructures with targeting ligands and imaging components (eg, magnetic resonance agents, quantum dots) allow for early detection of virus-driven tumors, real-time monitoring of therapy, and guided delivery of therapeutics.182,185 Advanced approaches highlighted include the use of bioengineered nanoparticles for sentinel lymph node mapping, image-guided surgical excision, and adaptive therapy strategies responsive to tumor microenvironmental cues. Additionally, the authors emphasize the critical importance of surface engineering such as PEGylation and ligand attachment, to prolong circulation time, improve biocompatibility, and enable multi-modal interventions including photodynamic and photothermal therapies.182 Such platforms will improve the ability to stratify patients, detect recurrence, and adapt treatment regimens, moving clinical nanomedicine toward smarter, more proactive care.182
Gautam et al also discuss novel clinical translation pathways, such as the use of multiplexed nanocarrier systems tailored to genetic and proteomic profiles for patient-specific therapy, and highlight ongoing clinical trials using viral-oncogenesis-targeted nanomedicines for improved outcomes with minimized adverse effects.182 These innovations are positioned as important steps beyond conventional monotherapies and earlier reviews, addressing the complexity of tumor microenvironments and resistance mechanisms.182
In addition, the development of next-generation nanovaccines, virion-mimicking VLPs, dendrimer-based systems, or hybrid organic-inorganic platforms, offers innovative routes for both prophylactic and therapeutic immunization against oncoviruses and their related cancers.128 These nanoparticles are tailored to present viral antigens or adjuvants in highly immunogenic contexts, eliciting robust adaptive and innate immune responses with greater potency and longevity than conventional vaccines. Importantly, such platforms are also being explored for use in therapeutic cancer vaccines and as carriers for checkpoint inhibitors or cytokines.128
Clinically, these strategic advances translate into improved pharmacokinetics, reduced adverse effects, and enhanced therapeutic index when compared to conventional single-modality approaches. Current literature also highlights that personalized nanocarrier designs, adaptive cargo loading, and surface functionalization enable the tailoring of therapies to patient-specific tumor genetics, viral mutations, and immune microenvironment characteristics.187,188
Future strategies emphasize patient-specific nanotherapies powered by artificial intelligence and big data analytics. By integrating genomic, proteomic, and imaging data, machine learning algorithms will allow clinicians to predict optimal nanoparticle designs, dosing regimens, and target patient subgroups in real time.189 This individualized approach can increase response rates, reduce toxicity, and adapt therapies as the tumor or viral status evolves hallmarks of precision oncology.182,189
By merging these foundational principles and recent developments including those from Gautam et al (2024), the review advances the literature: (1) by presenting emerging nano-enabled targeting strategies for virus-induced cancers, and (2) by discussing direct clinical progress in lymph node targeting, image-guided therapy, and real-time response assessment. This approach bridges the gap between established knowledge and novel clinical translations, helping to chart future directions in multifunctional nanoparticle-based therapy.
Translation of these advanced nanomedicine concepts into clinical practice must overcome significant barriers, including long-term safety, regulatory approval, cost-effectiveness, and scalable manufacturing.114 Nevertheless, as regulatory agencies develop nanotechnology-specific guidance and as interdisciplinary research bridges biological, engineering, and computational sciences, nanomedicine is poised to play a transformative role in both the prevention and treatment of oncovirus-associated malignancies.114,128
Conclusion
The revolutionary nature of nanotechnology can be seen in how it is already being used to improve the healthcare system in different ways. Owing to recent advancements in their design and engineering, nanomedicines have many benefits over traditional approaches for the diagnosis, prevention, and treatment of viral infections. Nanomedicine approaches are preferred for various reasons, including small particle size, capacity to change the surface to acquire the desired selectivity, and biocompatibility. These innovative techniques hold great promise for antiviral medicines, in addition to addressing the issues of antiviral resistance, poor drug solubility and bioavailability, rapid drug release, and a limited therapeutic window. The area of oncogenic viral infections is rapidly expanding, leading to the development of theranostics, which can lead to accurate diagnosis in addition to effective therapy and real-time monitoring. Nanomaterials, used as adjuvants in antiviral vaccinations and research to enhance the immune system, have shown promise in the prevention and treatment of oncogenic viral infections.
Acknowledgments
Naveed Ahmed would like to acknowledge Applied College, University of Tabuk, for providing the facilities to conduct this informative review.
Disclosure
Alqassem Hamdallah Abuarqoub is affiliated with Al-Wafi Group for Marketing and Int’l Trade Co. Ltd. The company had no role in the design, conduct, manuscript preparation, or decision to publish this review article. The authors report no other conflicts of interest in this work.
References
1. Muhaimin M, Chaerunisaa AY, Dewi MK, Khatib A, Hazrina A. The toxicological profile of active pharmaceutical ingredients-containing nanoparticles: classification, mechanistic pathways, and health implications. Pharmaceuticals. 2025;18(5):703.
2. Moore PS, Chang Y. Why do viruses cause cancer? Highlights of the first century of human tumour virology. Nat Rev Cancer. 2010;10(12):878–889. doi:10.1038/nrc2961
3. Howley PM, Pfister HJ. Beta genus papillomaviruses and skin cancer. Virology. 2015;479:290–296. doi:10.1016/j.virol.2015.02.004
4. Feng H, Shuda M, Chang Y, Moore PS. Clonal integration of a polyomavirus in human Merkel cell carcinoma. Science. 2008;319(5866):1096–1100. doi:10.1126/science.1152586
5. Schiffman M, Castle PE, Jeronimo J, Rodriguez AC, Wacholder S. Human papillomavirus and cervical cancer. Lancet. 2007;370(9590):890–907. doi:10.1016/S0140-6736(07)61416-0
6. Liu W, Yang R, Payne AS, et al. Identifying the target cells and mechanisms of Merkel cell polyomavirus infection. Cell Host Microbe. 2016;19(6):775–787. doi:10.1016/j.chom.2016.04.024
7. Doorbar J. Molecular biology of human papillomavirus infection and cervical cancer. Clin Sci. 2006;110(5):525–541. doi:10.1042/CS20050369
8. Xiao Q, Liu Y, Li T, et al. Viral oncogenesis in cancer: from mechanisms to therapeutics. Signal Transduction Targeted Ther. 2025;10(1):151. doi:10.1038/s41392-025-02197-9
9. Alibek K, Baiken Y, Kakpenova A, et al. Implication of human herpesviruses in oncogenesis through immune evasion and suppression. Infect Agent Cancer. 2014;9(1):3. doi:10.1186/1750-9378-9-3
10. Galati L, Chiantore MV, Marinaro M, Di Bonito P. Human oncogenic viruses: characteristics and prevention strategies-lessons learned from human papillomaviruses. Viruses. 2024;16(3):416.
11. Roulot D, Layese R, Brichler S, et al. Hepatitis D virus infection markedly increases the risk of hepatocellular carcinoma in patients with viral B cirrhosis. Clin Gastroenterol Hepatol. 2024;2024:1.
12. Turk SM, Jiang R, Chesnokova LS, Hutt-Fletcher LM. Antibodies to gp350/220 enhance the ability of Epstein-Barr virus to infect epithelial cells. J Virol. 2006;80(19):9628–9633. doi:10.1128/JVI.00622-06
13. Liang Q, Chang B, Brulois KF, et al. Kaposi’s sarcoma-associated herpesvirus K7 modulates Rubicon-mediated inhibition of autophagosome maturation. J Virol. 2013;87(22):12499–12503. doi:10.1128/JVI.01898-13
14. Li P, Jiang Z, Shi J, et al. A self-assembled nanoparticle vaccine elicits effective neutralizing antibody response against EBV infection. Front Immunol. 2025;15. doi:10.3389/fimmu.2024.1530364
15. Young LS, Yap LF, Murray PG. Epstein–Barr virus: more than 50 years old and still providing surprises. Nat Rev Cancer. 2016;16(12):789–802. doi:10.1038/nrc.2016.92
16. Prevention CfDCa. Epstein-Barr Virus and Infectious Mononucleosis. 2024. Available from: https://www.cdc.gov/epstein-barr/about/index.html.
17. Petersson F. Nasopharyngeal carcinoma: a review. Paper presented at: Seminars in diagnostic pathology. 2015.
18. Dawson CW, Port RJ, Young LS. The role of the EBV-encoded latent membrane proteins LMP1 and LMP2 in the pathogenesis of nasopharyngeal carcinoma (NPC). Paper presented at: Seminars in cancer biology. 2012.
19. Yu H, Robertson ES. Epstein-barr virus history and pathogenesis. Viruses. 2023;15(3):714. doi:10.3390/v15030714
20. Patel PD, Alghareeb R, Hussain A, Maheshwari MV, Khalid N. The association of epstein-barr virus with cancer. Cureus. 2022;14(6):e26314. doi:10.7759/cureus.26314
21. Thorley-Lawson DA. EBV persistence—introducing the virus. Epstein Barr Virus. 2015;2015:151–209.
22. Murata T, Tsurumi T. Switching of EBV cycles between latent and lytic states. Rev Med Virol. 2014;24(3):142–153. doi:10.1002/rmv.1780
23. Frappier L. Epstein–Barr Virus Is an Agent of Genomic Instability. Nature Publishing Group UK London; 2023.
24. Wang J-T, Doong S-L, Teng S-C, Lee C-P, Tsai C-H, Chen M-R. Epstein-Barr virus BGLF4 kinase suppresses the interferon regulatory factor 3 signaling pathway. J Virol. 2009;83(4):1856–1869. doi:10.1128/JVI.01099-08
25. Dai Y-C, Liao Y-T, Juan Y-T, et al. The novel nuclear targeting and BFRF1-interacting domains of BFLF2 are essential for efficient Epstein-Barr virus virion release. J Virol. 2020;94(3):e01498–01419. doi:10.1128/JVI.01498-19
26. Granato M, Feederle R, Farina A, et al. Deletion of Epstein-Barr virus BFLF2 leads to impaired viral DNA packaging and primary egress as well as to the production of defective viral particles. J Virol. 2008;82(8):4042–4051. doi:10.1128/JVI.02436-07
27. Feederle R, Neuhierl B, Baldwin G, et al. Epstein-Barr virus BNRF1 protein allows efficient transfer from the endosomal compartment to the nucleus of primary B lymphocytes. J Virol. 2006;80(19):9435–9443. doi:10.1128/JVI.00473-06
28. Xia J, He Y, Meng B, et al. NEK2 induces autophagy‐mediated bortezomib resistance by stabilizing Beclin‐1 in multiple myeloma. Mol oncol. 2020;14(4):763–778. doi:10.1002/1878-0261.12641
29. Li P, Yu Z, Jiang Z, et al. A liposome-based nanoparticle vaccine induces effective immunity against EBV infection. Vaccines. 2025;13(4):360.
30. Yang X, Zeng X, Huang J, et al. Loop-mediated isothermal amplification linked a nanoparticles-based biosensor for detecting Epstein-Barr virus. Appl Microbiol Biotechnol. 2024;108(1):91. doi:10.1007/s00253-023-12948-9
31. Dai Y, Zhang B, Yang L, Tao S, Yu Y, Li C. Recent progress in the vaccine development against Epstein-Barr virus. Viruses. 2025;17(7):936.
32. Yousaf A, Pervaiz J, Farzand A, et al. Scientific prominence of Epstein-Barr Virus (EBV) Concomitant with Nasopharyngeal carcinoma: Scientific prominence of EBV associated NPC. Electronic Journal of Medical Research. 2025;1(1):28–44.
33. Health NIo. NIH launches clinical trial of Epstein-Barr virus vaccine. 2022. Available from: https://www.nih.gov/news-events/news-releases/nih-launches-clinical-trial-epstein-barr-virus-vaccine.
34. Burga RA, Patel S, Bollard CM, Y Cruz CR, Fernandes R. Conjugating Prussian blue nanoparticles onto antigen-specific T cells as a combined nanoimmunotherapy. Nanomedicine. 2016;11(14):1759–1767. doi:10.2217/nnm-2016-0160
35. Xiang Y, Tian M, Huang J, et al. LMP2-mRNA lipid nanoparticle sensitizes EBV-related tumors to anti-PD-1 therapy by reversing T cell exhaustion. J Nanobiotechnol. 2023;21(1):324. doi:10.1186/s12951-023-02069-w
36. Anik Ü, Çaylayik NS. Development of Epstein Barr virus biosensor by mimicking its infection mechanism for early detection of multiple sclerosis. Sci Rep. 2025;15(1):28089. doi:10.1038/s41598-025-13030-2
37. Anık Ü, Çaylayık NS. Early detection of multiple sclerosis using spectroscopic epstein-barr virus biosensors based on infection mechanism mimicry. Anal Chem. 2025;97(37):20466–20474. doi:10.1021/acs.analchem.5c03964
38. Li Y, Yang X, Tao L, et al. Challenges in the diagnosis of Epstein-Barr virus-positive inflammatory follicular dendritic cell sarcoma: extremely wide morphologic spectrum and immunophenotype. Am J Surg Pathol. 2023;47(4):476–489. doi:10.1097/PAS.0000000000002011
39. Jary A, Veyri M, Gothland A, Leducq V, Calvez V, Marcelin AG. Kaposi’s sarcoma-associated herpesvirus, the etiological agent of all epidemiological forms of Kaposi’s sarcoma. Cancers. 2021;13(24):6208. doi:10.3390/cancers13246208
40. Kumwenda T, Hodson DZ, Rambiki K, et al. Diagnosis and management of kaposi sarcoma-associated herpesvirus inflammatory cytokine syndrome in resource-constrained settings: a case report and an adapted case definition. Trop Med Infect Dis. 2024;9(12). doi:10.3390/tropicalmed9120307
41. Zhang P, Wang J, Zhang X, Wang X, Jiang L, Gu X. Identification of AIDS-associated kaposi sarcoma: a functional genomics approach. Front Genet. 2019;10:1376. doi:10.3389/fgene.2019.01376
42. Purushothaman P, Dabral P, Gupta N, Sarkar R, Verma SC. KSHV genome replication and maintenance. Front Microbiol. 2016;7:54. doi:10.3389/fmicb.2016.00054
43. Ueda K. KSHV genome replication and maintenance in latency. Human Herpesviruses. 2018;2018:299–320.
44. Wei F, Gan J, Wang C, Zhu C, Cai Q. Cell cycle regulatory functions of the KSHV oncoprotein LANA. Front Microbiol. 2016;7:334. doi:10.3389/fmicb.2016.00334
45. Rose-John S. Interleukin-6 family cytokines. Cold Spring Harbor Perspect Biol. 2018;10(2):a028415. doi:10.1101/cshperspect.a028415
46. Cengiz Seval G, Beksac M. The safety of bortezomib for the treatment of multiple myeloma. Expert Opin Drug Saf. 2018;17(9):953–962. doi:10.1080/14740338.2018.1513487
47. Wan Q, Tucker AJ, Zhao J. Oncogenic mechanisms of kaposi’s sarcoma-associated herpesvirus on cell metabolism and cell transformation. J Med Virol. 2025;97(8):e70565. doi:10.1002/jmv.70565
48. Okada S, Goto H, Yotsumoto M. Current status of treatment for primary effusion lymphoma. Intractable Rare Dis Res. 2014;3(3):65–74. doi:10.5582/irdr.2014.01010
49. Arcuri LJ, Americo AD. Treatment of relapsed/refractory multiple myeloma in the bortezomib and lenalidomide era: a systematic review and network meta-analysis. Ann. Hematol. 2021;100(3):725–734. doi:10.1007/s00277-021-04404-3
50. C-y S, J-y L, Z-b C, Zhang L, Chen L, Hu Y. Efficacy and safety of bortezomib maintenance in patients with newly diagnosed multiple myeloma: a meta-analysis. Biosci Rep. 2017;37(4):BSR20170304. doi:10.1042/BSR20170304
51. Cao Y, Xie L, Shi F, et al. Targeting the signaling in Epstein–Barr virus-associated diseases: mechanism, regulation, and clinical study. Signal Transduction Targeted Ther. 2021;6(1):15. doi:10.1038/s41392-020-00376-4
52. Kanda T. EBV-encoded latent genes. Human Herpesviruses. 2018;2018:377–394.
53. Meyers JM, Munger K. The viral etiology of skin cancer. J Invest Dermatol. 2014;134(e1):E29–32. doi:10.1038/skinbio.2014.6
54. Liu C, Yin X, Xu H, et al. Microneedle-array-mediated transdermal delivery of GCV-functionalized zeolitic imidazolate framework-8 nanoparticles for KSHV treatment. Int J Mol Sci. 2024;25(23):12946. doi:10.3390/ijms252312946
55. Li F, Cao D, Yao L, et al. Targeted delivery of miR-34a-5p by phenylborate-coupled polyethylenimide nanocarriers for anti-KSHV treatment. Front Bioeng Biotechnol. 2023;11:1343956. doi:10.3389/fbioe.2023.1343956
56. Mancuso M, Jiang L, Cesarman E, Erickson D. Multiplexed colorimetric detection of Kaposi’s sarcoma associated herpesvirus and Bartonella DNA using gold and silver nanoparticles. Nanoscale. 2013;5(4):1678–1686. doi:10.1039/c3nr33492a
57. Baba SK, Alblooshi SSE, Yaqoob R, et al. Human papilloma virus (HPV) mediated cancers: an insightful update. J Transl Med. 2025;23(1):483.
58. Hussain FS, Abro NQ, Ahmed N, Memon SQ, Memon N. Nano-antivirals: a comprehensive review. Front Nanotechnol. 2022;4. doi:10.3389/fnano.2022.1064615
59. Arbyn M, Weiderpass E, Bruni L, et al. Estimates of incidence and mortality of cervical cancer in 2018: a worldwide analysis. Lancet Glob Health. 2020;8(2):e191–e203. doi:10.1016/S2214-109X(19)30482-6
60. Mohammed FA, Tune KK, Jett M, Muhie S. Cervical cancer stages, human papillomavirus integration, and malignant genetic mutations: integrative analysis of datasets from four different cohorts. Cancers. 2023;15(23):5595. doi:10.3390/cancers15235595
61. Graham SV. Human papillomavirus: gene expression, regulation and prospects for novel diagnostic methods and antiviral therapies. Fut Microbiol. 2010;5(10):1493–1506. doi:10.2217/fmb.10.107
62. Graham SV. Keratinocyte differentiation-dependent human papillomavirus gene regulation. Viruses. 2017;9(9):245. doi:10.3390/v9090245
63. Schelhaas M, Shah B, Holzer M, et al. Entry of human papillomavirus type 16 by actin-dependent, clathrin-and lipid raft-independent endocytosis. PLoS Pathogens. 2012;8(4):e1002657. doi:10.1371/journal.ppat.1002657
64. Guion LG, Sapp M. The role of promyelocytic leukemia nuclear bodies during HPV infection. Front Cell Infect Microbiol. 2020;10:35. doi:10.3389/fcimb.2020.00035
65. Doorbar J, Quint W, Banks L, et al. The biology and life-cycle of human papillomaviruses. Vaccine. 2012;30:F55–F70.
66. Venuti A, Paolini F, Nasir L, et al. Papillomavirus E5: the smallest oncoprotein with many functions. Mol Cancer. 2011;10(1):1–18. doi:10.1186/1476-4598-10-140
67. Aubrey BJ, Kelly GL, Janic A, Herold MJ, Strasser A. How does p53 induce apoptosis and how does this relate to p53-mediated tumour suppression? Cell Death Differ. 2018;25(1):104–113. doi:10.1038/cdd.2017.169
68. Li S, Hong X, Wei Z, et al. Ubiquitination of the HPV oncoprotein E6 is critical for E6/E6AP-mediated p53 degradation. Front Microbiol. 2019;10:2483.
69. Venuti A, Paolini F, Nasir L, et al. Papillomavirus E5: the smallest oncoprotein with many functions. Mol Cancer. 2011;10(1):140.
70. Nayar R, Wilbur DC. The Bethesda system for reporting cervical cytology: a historical perspective. Acta Cytologica. 2017;61(4–5):359–372. doi:10.1159/000477556
71. Oh HY, Seo -S-S, Kim MK, et al. Synergistic effect of viral load and alcohol consumption on the risk of persistent high-risk human papillomavirus infection. PLoS One. 2014;9(8):e104374. doi:10.1371/journal.pone.0104374
72. Boccardo E, Lepique AP, Villa LL. The role of inflammation in HPV carcinogenesis. Carcinogenesis. 2010;31(11):1905–1912. doi:10.1093/carcin/bgq176
73. Sharma U, Shekhar H, Sharma B, et al. Emerging nanotherapeutics for HPV-associated cancers: current advances, challenges, and future directions. Discov Oncol. 2025;16(1):1653. doi:10.1007/s12672-025-02439-w
74. Lopes-Nunes J, Oliveira PA, Cruz C. Nanotherapy for human papillomavirus-associated cancers: breakthroughs and challenges. Trends Pharmacol Sci. 2024;45(9):781–797. doi:10.1016/j.tips.2024.07.004
75. Zhao X, Zhang Y, Trejo-Cerro O, et al. A safe and potentiated multi-type HPV L2-E7 nanoparticle vaccine with combined prophylactic and therapeutic activity. NPJ Vaccines. 2024;9(1):119. doi:10.1038/s41541-024-00914-z
76. Zhang J, Fan J, Skwarczynski M, Stephenson RJ, Toth I, Hussein WM. Peptide-based nanovaccines in the treatment of cervical cancer: a review of recent advances. Int J Nanomed. 2022;17:869–900. doi:10.2147/IJN.S269986
77. Pan L, Li B, Chen J, et al. Nanotechnology-based weapons to combat human papillomavirus infection associated diseases. Front Chem. 2021;9:798727. doi:10.3389/fchem.2021.798727
78. Mahmoodi P, Rezayi M, Rasouli E, et al. Early-stage cervical cancer diagnosis based on an ultra-sensitive electrochemical DNA nanobiosensor for HPV-18 detection in real samples. J Nanobiotechnol. 2020;18(1):11. doi:10.1186/s12951-020-0577-9
79. Fallatah MM, Alradwan I, Alfayez N, et al. Nanoparticles for cancer immunotherapy: innovations and challenges. Pharmaceuticals. 2025;18(8):1086. doi:10.3390/ph18081086
80. Ferrarese A, Zanetto A, Gambato M, et al. Liver transplantation for viral hepatitis in 2015. World J Gastroenterol. 2016;22(4):1570. doi:10.3748/wjg.v22.i4.1570
81. D’Ambrosio R, Della Corte C, Colombo M. Hepatocellular carcinoma in patients with a sustained response to anti-hepatitis C therapy. Int J Mol Sci. 2015;16(8):19698–19712. doi:10.3390/ijms160819698
82. Ioannou GN, Green PK, Berry K. HCV eradication induced by direct-acting antiviral agents reduces the risk of hepatocellular carcinoma. J Hepatol. 2018;68(1):25–32. doi:10.1016/j.jhep.2017.08.030
83. Klöhn M, Schrader JA, Brüggemann Y, Todt D, Steinmann E. Beyond the usual suspects: Hepatitis E virus and its implications in hepatocellular carcinoma. Cancers. 2021;13(22):5867. doi:10.3390/cancers13225867
84. Kanda T, Goto T, Hirotsu Y, Moriyama M, Omata M. Molecular mechanisms driving progression of liver cirrhosis towards hepatocellular carcinoma in chronic hepatitis B and C infections: a review. Int J Mol Sci. 2019;20(6):1358. doi:10.3390/ijms20061358
85. Yang P, Markowitz GJ, Wang X-F. The hepatitis B virus-associated tumor microenvironment in hepatocellular carcinoma. Natl Sci Rev. 2014;1(3):396–412. doi:10.1093/nsr/nwu038
86. Zanetto A, Senzolo M, Campello E, et al. Influence of hepatocellular carcinoma on platelet aggregation in cirrhosis. Cancers. 2021;13(5):1150. doi:10.3390/cancers13051150
87. Jeng WJ, Papatheodoridis GV, Lok ASF. Hepatitis B. Lancet. 2023;401(10381):1039–1052. doi:10.1016/S0140-6736(22)01468-4
88. Tu T, Budzinska MA, Shackel NA, Urban S. HBV DNA integration: molecular mechanisms and clinical implications. Viruses. 2017;9(4):75. doi:10.3390/v9040075
89. Herman JG, Baylin SB. Gene silencing in cancer in association with promoter hypermethylation. N Engl J Med. 2003;349(21):2042–2054. doi:10.1056/NEJMra023075
90. Zhang X, Dong N, Zhang H, You J, Wang H, Ye L. Effects of hepatitis B virus X protein on human telomerase reverse transcriptase expression and activity in hepatoma cells. J Lab Clin Med. 2005;145(2):98–104. doi:10.1016/j.lab.2004.11.018
91. Phesse T, Flanagan D, Vincan E. Frizzled7: a promising Achilles’ heel for targeting the Wnt receptor complex to treat cancer. Cancers. 2016;8(5):50. doi:10.3390/cancers8050050
92. Ebert G, Allison C, Preston S, et al. Eliminating hepatitis B by antagonizing cellular inhibitors of apoptosis. Proc Natl Acad Sci. 2015;112(18):5803–5808. doi:10.1073/pnas.1502400112
93. Hussain S, Schwank J, Staib F, Wang X, Harris C. TP53 mutations and hepatocellular carcinoma: insights into the etiology and pathogenesis of liver cancer. Oncogene. 2007;26(15):2166–2176. doi:10.1038/sj.onc.1210279
94. Li G, Dai Z, Guo J. Therapeutic nanomaterials in NAFLD: current advances and potential applications in patients with concurrent HBV infection. Int J Nanomed. 2025;20:3803–3823. doi:10.2147/IJN.S510271
95. Zhuang AQ, Chen Y, Chen SM, et al. Current status and challenges in anti-hepatitis B virus agents based on inactivation/inhibition or elimination of hepatitis b virus covalently closed circular DNA. Viruses. 2023;15(12). doi:10.3390/v15122315
96. Negro F, Lok AS. Hepatitis D: a review. JAMA. 2023;330(24):2376–2387. doi:10.1001/jama.2023.23242
97. Degasperi E, Sandmann L, Wedemeyer H, Lampertico P. the delta cure working G. Hepatitis D Virus infection: pathophysiology, epidemiology and treatment. Report from the third delta cure meeting 2024. Liver Int. 2025;45(7):e70189. doi:10.1111/liv.70189
98. Pisaturo M, Russo A, Grimaldi P, Martini S, Coppola N. current and future therapeutic options for chronic hepatitis D virus infection. Front Cellular and Infect Microbiol. 2025;14:1382017. doi:10.3389/fcimb.2024.1382017
99. Lampertico P, Degasperi E, Sandmann L, Wedemeyer H. Hepatitis D virus infection: pathophysiology, epidemiology and treatment. Report from the first international delta cure meeting 2022. JHEP Rep. 2023;5(9):100818. doi:10.1016/j.jhepr.2023.100818
100. Farci P, Niro GA, Zamboni F, Diaz G. Hepatitis D virus and hepatocellular carcinoma. Viruses. 2021;13(5):830. doi:10.3390/v13050830
101. Puigvehí M, Moctezuma-Velázquez C, Villanueva A, Llovet JM. The oncogenic role of hepatitis delta virus in hepatocellular carcinoma. JHEP Rep. 2019;1(2):120–130. doi:10.1016/j.jhepr.2019.05.001
102. Shih HH, Sheen I-J, Su C-W, Peng W-L, Lin L-H, Wu J-C. Hepatitis D virus isolates with low replication and epithelial-mesenchymal transition-inducing activity are associated with disease remission. J Virol. 2012;86(17):9044–9054. doi:10.1128/JVI.00130-12
103. Pugnale P, Pazienza V, Guilloux K, Negro F. Hepatitis delta virus inhibits alpha interferon signaling. Hepatology. 2009;49(2):398–406. doi:10.1002/hep.22654
104. Gopalakrishna H, Mironova M, Dahari H, Koh C, Heller T. Advances and challenges in managing Hepatitis D virus: evolving strategies. Curr Hepatol Rep. 2024;23(1):32–44. doi:10.1007/s11901-024-00643-w
105. Singh L, Kruger HG, Maguire GEM, Govender T, Parboosing R. The role of nanotechnology in the treatment of viral infections. Ther Adv Infect Dis. 2017;4(4):105–131. doi:10.1177/2049936117713593
106. Liu W-H, Cui J-Y, Yu M, et al. A comprehensive review of diagnostic approaches for hepatitis D. Front Mol Biosci. 2025;12:1598784. doi:10.3389/fmolb.2025.1598784
107. Shevtsov M, Zhao L, Protzer U, van de Klundert MAA. Applicability of metal nanoparticles in the detection and monitoring of hepatitis B virus infection. Viruses. 2017;9(7):193. doi:10.3390/v9070193
108. Martinello M, Solomon SS, Terrault NA, Dore GJ. Hepatitis C. Lancet. 2023;402(10407):1085–1096. doi:10.1016/S0140-6736(23)01320-X
109. Dash S, Aydin Y, Widmer KE, Nayak L. Hepatocellular carcinoma mechanisms associated with chronic HCV infection and the impact of direct-acting antiviral treatment. J Hepatocell Carcinoma. 2020;Volume 7:45–76. doi:10.2147/JHC.S221187
110. Ali AM, Hassan MR, Aburub F, et al. explainable machine learning approach for hepatitis C diagnosis using SFS feature Selection. Machines. 2023;11:391. doi:10.3390/machines11030391
111. Li Y, Boehning DF, Qian T, Popov VL, Weinman SA. Hepatitis C virus core protein increases mitochondrial ROS production by stimulation of Ca2+ uniporter activity. FASEB J. 2007;21(10):2474–2485. doi:10.1096/fj.06-7345com
112. Chang M-L. Metabolic alterations and hepatitis C: from bench to bedside. World J Gastroenterol. 2016;22(4):1461. doi:10.3748/wjg.v22.i4.1461
113. Yousaf A. DNA replication: causes of replication stress and DNA damage response pathways: A review on DNA replication. Electron J Med Res. 2025;1(1):8–27.
114. Ghorai S, Shand H, Patra S, et al. Nanomedicine for the treatment of viral diseases: smaller solution to bigger problems. Pharmaceutics. 2024;16(3):407. doi:10.3390/pharmaceutics16030407
115. Elberry MH, Darwish NHE, Mousa SA. Hepatitis C virus management: potential impact of nanotechnology. Virol J. 2017;14(1):88. doi:10.1186/s12985-017-0753-1
116. Toth EA, Andrianov AK, Fuerst TR. Prospects for developing an Hepatitis C virus E1E2-based nanoparticle vaccine. Rev Med Virol. 2023;33(5):e2474. doi:10.1002/rmv.2474
117. Duncan JD, Urbanowicz RA, Tarr AW, Ball JK. Hepatitis C virus vaccine: challenges and prospects. Vaccines. 2020;8(1):90.
118. Sankatsing VDV, van Summeren J, Abreha FM, et al. Economic impact of respiratory syncytial virus infections in children under 5 years of age attending primary care in Italy: a prospective cohort study in two regions. Influenza Other Respir Viruses. 2025;19(2):e70074.
119. Bloom DE, Cadarette D. Infectious disease threats in the twenty-first century: strengthening the global response. Front Immunol. 2019;10:549. doi:10.3389/fimmu.2019.00549
120. Hua S, Wu SY. Advances and challenges in nanomedicine. Front Pharmacol. 2018;9:1397. doi:10.3389/fphar.2018.01397
121. Singh L, Kruger HG, Maguire GE, Govender T, Parboosing R. The role of nanotechnology in the treatment of viral infections. Therap Adv Infectious Dis. 2017;4(4):105–131.
122. Islam S, Ahmed MMS, Islam MA, Hossain N, Chowdhury MA. Advances in nanoparticles in targeted drug delivery–A review. Results Surfaces Interfaces. 2025;19:100529. doi:10.1016/j.rsurfi.2025.100529
123. Wan C, Tai J, Zhang J, et al. Silver nanoparticles selectively induce human oncogenic γ-herpesvirus-related cancer cell death through reactivating viral lytic replication. Cell Death Dis. 2019;10(6):392.
124. Huang Y, Guo X, Wu Y, et al. Nanotechnology’s frontier in combatting infectious and inflammatory diseases: prevention and treatment. Signal Transduction Targeted Ther. 2024;9(1):34. doi:10.1038/s41392-024-01745-z
125. Lysenko V, Lozovski V, Lokshyn M, et al. Nanoparticles as antiviral agents against adenoviruses. Adv Nat Sci. 2018;9(2):025021.
126. Aguilera-Correa JJ, Esteban J, Vallet-Regí M. Inorganic and polymeric nanoparticles for human viral and bacterial infections prevention and treatment. Nanomaterials. 2021;11(1):137. doi:10.3390/nano11010137
127. Umezawa E, Fujino K, Yamanaka HI, et al. Nonwoven fabric coated with cerium oxide nanoparticles for viral inactivation and transmission Inhibition. Sci Rep. 2025;15(1):10340. doi:10.1038/s41598-025-94199-4
128. Kumarasamy RV, Natarajan PM, Umapathy VR, Roy JR, Mironescu M, Palanisamy CP. Clinical applications and therapeutic potentials of advanced nanoparticles: a comprehensive review on completed human clinical trials. Front Nanotechnol. 2024;6. doi:10.3389/fnano.2024.1479993
129. Liang Z, Shan G, Shan S, Ding Z, Zhang J, Chu B. Comprehensive review on emerging nanotechnologies for combating COVID-19 and future pandemic preparedness. Int J Pharm. 2025;682:125970. doi:10.1016/j.ijpharm.2025.125970
130. Aguilera Correa JJ, Esteban J, Vallet Regí M. Nanoparticles for human viral and bacterial infections prevention and treatment. Ene. 2021;9:56.
131. Raji V, Pal K, Zaheer T, et al. Gold nanoparticles against respiratory diseases: oncogenic and viral pathogens review. Therapeut Delivery. 2020;11(8):521. doi:10.4155/tde-2020-0071
132. Coleman CM, Venkataraman T, Liu YV, et al. MERS-CoV spike nanoparticles protect mice from MERS-CoV infection. Vaccine. 2017;35(12):1586–1589. doi:10.1016/j.vaccine.2017.02.012
133. Zhang W, Yang H, Kong X, et al. Inhibition of respiratory syncytial virus infection with intranasal siRNA nanoparticles targeting the viral NS1 gene. Nature Med. 2005;11(1):56–62. doi:10.1038/nm1174
134. Ma Y, Xiao X, Wang Y, et al. Insight into antiviral activity of Ag/TiO2 nanocomposites against influenza H1N1 virus and its antiviral mechanism. Int J Nanomed. 2024;Volume 19:11305–11320. doi:10.2147/IJN.S469684
135. Bhatti A, DeLong RK. Nanoscale interaction mechanisms of antiviral activity. ACS Pharmacol Transl Sci. 2023;6(2):220–228. doi:10.1021/acsptsci.2c00195
136. Jain N, Jain P, Rajput D, Patil UK. Green synthesized plant-based silver nanoparticles: therapeutic prospective for anticancer and antiviral activity. Micro Nano Systems Letters. 2021;9(1):1–24.
137. Huang X, Li M, Xu Y, et al. Novel gold nanorod-based HR1 peptide inhibitor for middle east respiratory syndrome coronavirus. ACS Appl Mater Interfaces. 2019;11(22):19799–19807. doi:10.1021/acsami.9b04240
138. Aguilera-Correa J, Esteban J, Vallet-Regí M. Inorganic and polymeric nanoparticles for human viral and bacterial infections prevention and treatment. Nanomaterials. 2021;11(137):137.
139. Oprea M, Panaitescu DM. Nanocellulose hybrids with metal oxides nanoparticles for biomedical applications. Molecules. 2020;25(18):4045. doi:10.3390/molecules25184045
140. Negahdari B, Darvishi M, Saeedi AA. Gold nanoparticles and hepatitis B virus. Artific Cells Nanomed Biotechnol. 2019;47(1):455–461. doi:10.1080/21691401.2018.1553786
141. Maksimovic‐Ivanic D, Fagone P, McCubrey J, Bendtzen K, Mijatovic S, Nicoletti F. HIV‐protease inhibitors for the treatment of cancer: repositioning HIV protease inhibitors while developing more potent NO‐hybridized derivatives? Int J Cancer. 2017;140(8):1713–1726. doi:10.1002/ijc.30529
142. Jwaziri AK, Salavatiha Z, Kiani SJ, Khales P, Vazirzadeh M, Tavakoli A. Magnesium oxide nanoparticles: a new frontier in antiviral therapy against herpes simplex virus type 1. Adv Virol. 2025;2025(1):3088529. doi:10.1155/av/3088529
143. Liu C-H, Lin -C-C, Hsu W-C, et al. Highly bioavailable silibinin nanoparticles inhibit HCV infection. Gut. 2017;66(10):1853–1861. doi:10.1136/gutjnl-2016-312019
144. Lee M-Y, Yang J-A, Jung HS, et al. Hyaluronic acid–gold nanoparticle/interferon α complex for targeted treatment of hepatitis C virus infection. ACS Nano. 2012;6(11):9522–9531. doi:10.1021/nn302538y
145. Gaglia MM, Munger K. More than just oncogenes: mechanisms of tumorigenesis by human viruses. Curr Opin Virol. 2018;32:48–59. doi:10.1016/j.coviro.2018.09.003
146. Yerramsetti S, Cohen T, Atun R, Menzies NA. Global estimates of paediatric tuberculosis incidence in 2013–19: a mathematical modelling analysis. Lancet Glob Health. 2022;10(2):e207–e215. doi:10.1016/S2214-109X(21)00462-9
147. He Y, Wang Y, Wang L, Jiang W, Wilhelm S. Understanding nanoparticle-liver interactions in nanomedicine. Expert Opin Drug Deliv. 2024;21(6):829–843. doi:10.1080/17425247.2024.2375400
148. Meng X, Zhu G, Yang Y-G, Sun T. Targeted delivery strategies: the interactions and applications of nanoparticles in liver diseases. Biomed Pharmacother. 2024;175:116702.
149. Nicolae CL, Pîrvulescu DC, Antohi AM, Niculescu AG, Grumezescu AM, Croitoru GA. Silica nanoparticles in medicine: overcoming pathologies through advanced drug delivery, diagnostics, and therapeutic strategies. Rom J Morphol Embryol. 2024;65(2):173–184.
150. Payamifar S, Khalili Y, Foroozandeh A, Abdouss M, Hasanzadeh M. Magnetic mesoporous silica nanoparticles as advanced polymeric scaffolds for efficient cancer chemotherapy: recent progress and challenges. RSC Adv. 2025;15(20):16050–16074. doi:10.1039/D5RA00948K
151. Liu M, Zhao Y, Shi Z, Zink JI, Yu Q. Virus-like magnetic mesoporous silica particles as a universal vaccination platform against pathogenic infections. ACS Nano. 2023;17(7):6899–6911. doi:10.1021/acsnano.3c00644
152. Serrano-Aroca Á, Takayama K, Mishra YK, de la Fuente-Nunez C. Carbon-based nanomaterials for antiviral applications. Adv Funct Mater. 2024;34(38):2402023.
153. Zandi M, Hosseini F, Adli AH, et al. State-of-the-art cerium nanoparticles as promising agents against human viral infections. Biomed Pharmacother. 2022;156:113868.
154. Shcherbakov AB. CeO2 nanoparticles and cerium species as antiviral agents: critical review. Eur J Med Chem Rep. 2024;10:100141.
155. Bukhari B, Naveed M, Makhdoom SI, et al. A comparison between organic and inorganic nanoparticles: prime nanoparticles for tumor curation. Nano. 2021;16(13):2130011.
156. Auffan M, Rose J, Bottero J-Y, Lowry GV, Jolivet J-P, Wiesner MR. Towards a definition of inorganic nanoparticles from an environmental, health and safety perspective. Nat Nanotechnol. 2009;4(10):634–641. doi:10.1038/nnano.2009.242
157. Huang Y-J, S-hJN H. TRAIL-functionalized gold nanoparticles selectively trigger apoptosis in polarized macrophages. Nanotheranostics. 2017;1(3):326. doi:10.7150/ntno.20233
158. Priya MRK, Iyer PR. Antiproliferative effects on tumor cells of the synthesized gold nanoparticles against Hep2 liver cancer cell line. Egyp Liver J. 2020;10(1):15. doi:10.1186/s43066-020-0017-4
159. Liu R, Pei Q, Shou T, Zhang W, Hu J, Li W. Apoptotic effect of green synthesized gold nanoparticles from Curcuma wenyujin extract against human renal cell carcinoma A498 cells. Int J Nanomed. 2019;14:4091–4103. doi:10.2147/IJN.S203222
160. Lu Z, Qi L, Y-r L, et al. Novel albumin nanoparticle enhanced the anti-insulin-resistant-hepatoma activity of metformin. Int J Nanomed. 2020;15:5203. doi:10.2147/IJN.S253094
161. Wang K, Kievit FM, Jeon M, Silber JR, Ellenbogen RG, Zhang M. Nanoparticle‐mediated target delivery of TRAIL as gene therapy for glioblastoma. Adv Healthcare Mater. 2015;4(17):2719–2726. doi:10.1002/adhm.201500563
162. Abusalah MAH, Abd Rahman ENSE, Choudhary OP, Choudhary OP. Evolving trends in stem cell therapy: an emerging and promising approach against various diseases. Int J Surg. 2024;110(11):6862–6868. doi:10.1097/JS9.0000000000001948
163. Pavelić K, Kraljević Pavelić S, Bulog A, et al. Nanoparticles in medicine: current status in cancer treatment. Int J Mol Sci. 2023;24(16):12827. doi:10.3390/ijms241612827
164. Nunez C, Estevez SV, Del Pilar Chantada M. Inorganic nanoparticles in diagnosis and treatment of breast cancer. J Biol Inorganic Chem. 2018;23(3):331–345. doi:10.1007/s00775-018-1542-z
165. Zhang N, Yu J, Liu P, Chang J, Ali D, Tian X. Gold nanoparticles synthesized from Curcuma wenyujin inhibits HER-2/neu transcription in breast cancer cells (MDA-MB-231/HER2). Arab J Chem. 2020;13(10):7264–7273.
166. Havelikar U, Ghorpade KB, Kumar A, et al. Comprehensive insights into mechanism of nanotoxicity, assessment methods and regulatory challenges of nanomedicines. Discov Nano. 2024;19(1):165. doi:10.1186/s11671-024-04118-1
167. Murugesan S, Balasubramanian S, Perumal E. Copper oxide nanoparticles induced reactive oxygen species generation: a systematic review and meta-analysis. Chem Biol Interact. 2025;405:111311. doi:10.1016/j.cbi.2024.111311
168. Das SK, Sen K, Ghosh B, Ghosh N, Sinha K, Sil PC. Molecular mechanism of nanomaterials induced liver injury: a review. World J Hepatol. 2024;16(4):566–600. doi:10.4254/wjh.v16.i4.566
169. Lin W, Huang YW, Zhou XD, Ma Y. Toxicity of cerium oxide nanoparticles in human lung cancer cells. Int J Toxicol. 2006;25(6):451–457. doi:10.1080/10915810600959543
170. Xuan L, Ju Z, Skonieczna M, Zhou PK, Huang R. Nanoparticles-induced potential toxicity on human health: applications, toxicity mechanisms, and evaluation models. MedComm. 2023;4(4):e327. doi:10.1002/mco2.327
171. Sati A, Mali SN, Ranade TN, Yadav S, Pratap A. Silver nanoparticles (AgNPs) as a double-edged sword: synthesis, factors affecting, mechanisms of toxicity and anticancer potentials—An updated review till march 2025. Biol Trace Elem Res. 2025. doi:10.1007/s12011-025-04688-w
172. Wicki A, Witzigmann D, Balasubramanian V, Huwyler J. Nanomedicine in cancer therapy: challenges, opportunities, and clinical applications. J Control Release. 2015;200(200):138–157. doi:10.1016/j.jconrel.2014.12.030
173. Ma Y, Yang H, Niu S, Guo M, Xue Y. Mechanisms of micro- and nanoplastics on blood-brain barrier crossing and neurotoxicity: current evidence and future perspectives. NeuroToxicol. 2025;109:92–107. doi:10.1016/j.neuro.2025.06.003
174. Sharma S, Parveen R, Chatterji BP. Toxicology of nanoparticles in drug delivery. Curr Pathobiol Rep. 2021;9(4):133–144. doi:10.1007/s40139-021-00227-z
175. Qamar W, Gulia S, Athar M, et al. An insight into impact of nanomaterials toxicity on human health. Peer J. 2024;12:e17807.
176. Huang C, Liu X, Wu Q, et al. Cardiovascular toxic effects of nanoparticles and corresponding molecular mechanisms. Environ Pollut. 2024;356:124360. doi:10.1016/j.envpol.2024.124360
177. Neetika SM, Thakur P, Thakur P, et al. Cancer treatment and toxicity outlook of nanoparticles. Environ Res. 2023;237(Pt 1):116870. doi:10.1016/j.envres.2023.116870
178. Damasco JA, Ravi S, Perez JD, Hagaman DE, Melancon MP. Understanding nanoparticle toxicity to direct a safe-by-design approach in cancer nanomedicine. Nanomaterials. 2020;10(11). doi:10.3390/nano10112186
179. Khalili Fard J, Jafari S, Eghbal MA. A review of molecular mechanisms involved in toxicity of nanoparticles. Adv Pharm Bull. 2015;5(4):447–454. doi:10.15171/apb.2015.061
180. Kumah EA, Fopa RD, Harati S, Boadu P, Zohoori FV, Pak T. Human and environmental impacts of nanoparticles: a scoping review of the current literature. BMC Public Health. 2023;23(1):1059. doi:10.1186/s12889-023-15958-4
181. Nsairat H, Lafi Z, Al-Najjar BO, et al. How advanced are self-assembled nanomaterials for targeted drug delivery? A comprehensive review of the literature. Int J Nanomed. 2025;20:2133–2161. doi:10.2147/IJN.S490444
182. Gautam D, Joshi G, Talwan P, Bisht P. Nanotechnology in Cancer Therapy: targeted Drug Delivery and the Role in Viral Oncogenesis. In: Nanoparticles in Cancer Therapy. CRC Press; 2024:301–318.
183. Yao Y, Zhou Y, Liu L, et al. Nanoparticle-based drug delivery in cancer therapy and its role in overcoming drug resistance. Front Mol Biosci. 2020;7:193. doi:10.3389/fmolb.2020.00193
184. Ulbrich K, Holá K, Šubr V, Bakandritsos A, Tuček J, Zbořil R. Targeted drug delivery with polymers and magnetic nanoparticles: covalent and noncovalent approaches, release control, and clinical studies. Chem Rev. 2016;116(9):5338–5431. doi:10.1021/acs.chemrev.5b00589
185. Yu MK, Park J, Jon S. Targeting strategies for multifunctional nanoparticles in cancer imaging and therapy. Theranostics. 2012;2(1):3–44. doi:10.7150/thno.3463
186. Wu M, Huang S. Magnetic nanoparticles in cancer diagnosis, drug delivery and treatment. Mol Clin Oncol. 2017;7(5):738–746. doi:10.3892/mco.2017.1399
187. Lan HR, Zhang YN, Han YJ, et al. Multifunctional nanocarriers for targeted drug delivery and diagnostic applications of lymph nodes metastasis: a review of recent trends and future perspectives. J Nanobiotechnol. 2023;21(1):247. doi:10.1186/s12951-023-01990-4
188. Minh Hoang CN, Nguyen SH, Tran MT. Nanoparticles in cancer therapy: strategies to penetrate and modulate the tumor microenvironment – a review. Smart Mater Med. 2025;6(2):270–284. doi:10.1016/j.smaim.2025.07.004
189. Fountzilas E, Pearce T, Baysal MA, Chakraborty A, Tsimberidou AM. Convergence of evolving artificial intelligence and machine learning techniques in precision oncology. Npj Digital Med. 2025;8(1):75. doi:10.1038/s41746-025-01471-y
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Green Synthesis and Characterization of Silver Nanoparticles Using Zingiber officinale Extracts to Investigate Their Antibacterial Potential
Ramzan M, Abusalah MAHA, Ahmed N, Yean CY, Zeshan B
International Journal of Nanomedicine 2024, 19:13319-13338
Published Date: 11 December 2024