Back to Journals » International Journal of Nanomedicine » Volume 12
Nanoparticle–allergen complexes for allergen immunotherapy
Authors Di Felice G ![]() , Colombo P
, Colombo P
Received 14 February 2017
Accepted for publication 11 April 2017
Published 19 June 2017 Volume 2017:12 Pages 4493—4504
DOI https://doi.org/10.2147/IJN.S134630
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Thomas Webster
Gabriella Di Felice,1 Paolo Colombo2
1National Center for Drug Research and Evaluation, Istituto Superiore di Sanità, Rome, 2Institute of Biomedicine and Molecular Immunology, National Research Council, Palermo, Italy
Abstract: Allergen-specific immunotherapy was introduced in clinical settings more than 100 years ago. It remains the only curative approach to treating allergic disorders that ameliorates symptoms, reduces medication costs, and blocks the onset of new sensitizations. Despite this clinical evidence and knowledge of some immunological mechanisms, there remain some open questions regarding the safety and efficacy of this treatment. This suggests the need for novel therapeutic approaches that attempt to reduce the dose and frequency of treatment administration, improving patient compliance, and reducing costs. In this context, the use of novel adjuvants has been proposed and, in recent years, biomedical applications using nanoparticles have been exploited in the attempt to find formulations with improved stability, bioavailability, favorable biodistribution profiles, and the capability of targeting specific cell populations. In this article, we review some of the most relevant regulatory aspects and challenges concerning nanoparticle-based formulations with immunomodulatory potential, their related immunosafety issues, and the nature of the nanoparticles most widely employed in the allergy field. Furthermore, we report in vitro and in vivo data published using allergen/nanoparticle systems, discuss their impact on the immune system in terms of immunomodulatory activity and the reduction of side effects, and show that this strategy is a novel and promising tool for the development of allergy vaccines.
Keywords: allergy, nanocarriers, immunotoxicity, immune modulation, immunotherapy, allergens
Introduction
Allergic disorders are a growing global health problem affecting more than 25% of populations in industrialized countries,1,2 and there has been a significant worldwide increase in the prevalence of asthma and allergic rhinitis since 1960.3 Immunologically, allergic subjects manifest a higher frequency of allergen-specific CD4+ Th2 cells secreting IL-4, IL-5, and IL-13 than healthy subjects, leading to the production of allergen-specific immunoglobulin E (IgE) antibodies.4
Allergen-specific immunotherapy
To date, allergen-specific immunotherapy (AIT) has been the only treatment with the demonstrated capacity to ameliorate allergic symptoms, preventing the onset of new sensitizations to different allergens, as well as reducing the development of asthma in patients with allergic rhinitis.5
In particular, clinical trials using different pharmacological combinations and diverse routes of administration have demonstrated the crucial role of regulatory T- and B-cells (Treg and Breg, respectively)6,7 and the upregulation of IgG1, IgG4, IgA antibodies.8 The production of allergen-specific Treg is triggered by IL-109 and TGF-β10 which are induced during AIT due to the increase of CD25+ CD3+ FOXP3+ cells at the local and systemic level after immunotherapy.11,12 Furthermore, successful immunotherapy brings about a substantial reduction in the recruitment and activation of proinflammatory cells, including basophils, mast cells, and eosinophils in the skin, as well as in nasal or bronchial mucosae.13
Despite these advances in our understanding of the dynamics and functioning of AIT, there remain several problematic issues to resolve.
Market data show that only a limited number of patients suffering from allergic rhinoconjunctivitis decide to initiate immunotherapy, probably due to unfavorable conditions such as: 1) the high percentage of patients with local side effects,14,15 and 2) the duration of the treatment (up to 3 years) which causes low patient compliance and elevated costs.16 All of these points underline the importance of developing novel therapeutic approaches, improving the production of pharmaceutical products, and establishing optimal dosages and the more effective routes of administration in order to reduce the costs and duration of treatment and increase its safety.
Nanotechnology applied to immunotherapy
In the last decade, nanotechnologies have been applied to immunomodulatory therapies to enhance their efficacy and reduce potential side effects, mainly in addressing cancer and infectious diseases. In fact, in the last 30 years, a number of polymers have been used to design nanoparticles (NPs) that are capable of loading both proteins and nucleic acids for therapeutic applications.17,18 Although there is no universally accepted definition of NPs, they can be regarded as the primary component of a nanomaterial (NM), having defined physical limits and at least one dimension (or one diameter) in the nanoscale range.19 The definition of NMs recommended by the European Commission in 2011 is essentially based on size as the primary parameter to describe natural, incidental, or manufactured materials containing particles with any external dimension in the “nano” range (1–100 nm).20 A more comprehensive review, including recent scientific and technological developments, is underway.
NPs can take many different forms and have variable chemical compositions and physical properties, including their size/hydrodynamic radius, morphology, surface chemistry, solubility, and charge.21 These properties can be engineered to make them suitable for specific biomedical applications. The influence of physicochemical properties on the biocompatibility of NPs is critical for their use in biological applications and has been studied extensively. On the other hand, the biodistribution of NPs after entering the body through different routes, NP cellular uptake mechanisms, and their potential toxicity are primarily influenced by the physicochemical properties of the particles – especially size and surface characteristics.22
Regulatory issues
New nanotechnology-based medicinal products, developed for both therapeutic and diagnostic uses, can potentially yield innovative approaches to the field of drug delivery, improving the targeting, bioavailability, and transport of existing medicines across biological barriers or enabling new action mechanisms. The challenge for regulatory agencies is to ensure the proper evaluation of the quality, safety, and effectiveness of nanomedicines undergoing clinical development and market authorization. Since its 2006 Reflection Paper, the European Medicine Agency (EMA) has emphasized the complexity of nanomedicine properties and the consequent need to involve specialized experts in their evaluation (http://www.ema.europa.eu/docs/en_GB/document_library/Regulatory_and_procedural_guideline/2010/01/WC500069728.pdf). Although the current regulatory framework appears sufficiently robust, several critical issues persist with regard to NP characterization, detection, fate, and persistence in human organs and tissues. Moreover, the suitability of current toxicological methods to facilitate satisfactory and exhaustive risk assessment remains under discussion.23,24
Nonetheless, a number of medicinal products containing NPs in the form of liposomes, polymer protein conjugates, polymeric substances or suspensions, and nanocrystals have already been granted Marketing Authorizations within the European Community and/or at the world level under the existing regulatory framework.18,23 Several nanodrugs are undergoing basic research and preclinical and clinical development, the entire process of which can take up to 20 years. Several specific safety issues can affect this process, primarily pertaining to: 1) the set-up and validation of standardized in vitro assays, with suitable reference materials; 2) the development of ex vivo/in vivo models relevant for the route of administration; 3) the generation of in silico approaches which are predictive for biological and toxicological responses; and 4) in vivo pharmacokinetic studies.25 Finally, the interactions of NM-based medicines with the major biological systems deserve particular attention from coordinated contributions of multidisciplinary competences.
Nanotoxicity assessment
Current approaches to assessing NM toxicity are based primarily on knowledge derived from chemical safety assessments. These methods may not adequately account for the unique properties of NMs compared to those of bulk materials/chemicals such as their small size, aggregation/agglomeration capacity, and reactivity in biological matrices. For instance, these properties may influence their absorption and transportability across membranes, persistence and accumulation in organs, entrance into blood circulation, or even crossing of biological barriers.
Although nanotoxicology has become an important topic of research with significant funding, as documented by the number of projects supported by FP6, FP7, and Horizon2020 European programs, the appropriateness of existing test methods for addressing NM safety remains under discussion among regulatory authorities and within the scientific community. The ad hoc Working Party on Manufactured Nanomaterials (WPMN) established by the Organization for Economic Cooperation and Development (OECD) has the major task of reviewing the suitability of standard chemical testing protocols for NM evaluation. Three main issues can be identified: the insufficient physicochemical characterization of NMs, the inadequate standardization and validation of toxicity assays, and the lack of reference materials. Other weaknesses that should be addressed are summarized in the following points: testing methods are often time-consuming, expensive, and/or require highly experienced specialists, thus limiting their applicability to the rapidly increasing number of novel entities that require assessment; testing tends to focus on single agents, but in many cases these agents are present as parts of mixtures, resulting in interactions that may affect biological outcomes; many existing methods, in particular in vitro assays based on cell culture models, are geared toward assessing acute responses, whereas effects of chronic and repeated exposure are much less accessible to testing; animal studies allow both longer term testing and access to intact tissues and organisms, but knowledge transfer to humans is problematic due to differences between species; moreover, the increasing need to test more different substances and mixtures rules out an upscaling of animal use for ethical, scientific, practical, and financial reasons.
An approach to overcoming these obstacles has been proposed, based on the development of an intelligent testing strategy (ITS) that would allow adequate safety evaluation of NMs.26 An ITS should integrate the characterization of physicochemical properties with in vivo and in vitro studies as well as in silico models, thus allowing the association of the intrinsic properties of an NM with its potential toxicity. A generally applied scheme encompasses some cellular assays and in vivo exposure studies in rodents, taking into account the different exposure scenarios. In vivo studies in appropriate animal models allow the evaluation of complex responses resulting from the involvement of the whole organism or its major systems, as well as the investigation of biokinetic and toxicokinetic mechanisms. Due to the complexity and the costly and time-consuming nature of in vivo studies, in vitro testing can be regarded as the first-line methodology for screening purposes in ITS and for refining the consequent evaluation steps. To this aim, the in vitro endpoints more frequently investigated include cytotoxicity, genotoxicity, inflammation, and immunotoxicity.
The assessment of the impact of NM-based medicines on the immune system in terms of immunotoxicity, and of their potential to modulate (intentionally or not) its function (suppression, adjuvance, and hypersensitivity),27 has particular relevance for the object of this review.
Until now, no specific regulatory documents have been developed to assess the immunotoxicity of nanomedicines, which are currently evaluated on the basis of guidelines, such as the International Council for Harmonisation of Technical Requirements for Pharmaceuticals for Human Use Safety Guideline 8 (ICHS8) on immunotoxicity studies for human pharmaceuticals, which are in force for conventional medicinal products (http://www.ema.europa.eu/docs/en_GB/document_library/Scientific_guideline/2009/09/WC500002851.pdf). The open question is whether the set of first- and second-level tests prescribed by these guidelines is capable of adequately evaluating the immunotoxic profile of complex and broadly heterogeneous NM-based medicines that result from the combined effects of the active pharmaceutical component, the nanocarrier, any potential coating, and the final formulation.28 A prudent response to this issue is that not all the immunotoxic properties of nanomedicines can be easily identified following the recommendations of the current guidelines. As a consequence, a specific testing battery, tailored to the different categories of NMs and the wide range of immunotoxic effects that they can cause, needs to be developed.
Therapeutic potential of nanoparticles for allergies
In recent years, various studies have demonstrated that the encapsulation of allergenic extracts or single allergens within NPs can beneficially affect the interaction of the encapsulated antigen with the immune system, as well as further improve the development of safer and more effective vaccines and immunosuppressive agents.
In the first case, immunostimulatory activity is required in order to enhance the quantity and quality of the cellular and humoral immune responses to the antigens in the vaccine. NPs can engage in such “adjuvant” activities through various mechanisms associated with the physicochemical characteristics of the NP and/or the possibility of incorporating specific ligands/agonists for the specific targeting of immune cells.29 Particle size, which affects tissue penetration and access to blood vessels and the lymphatic system, can be altered to modulate antigen delivery. The ability to protect antigens at the site of administration, maintaining their stability and conformation and allowing a gradual local release (“depot” effect), as well as the possibility of displaying their spatial organization in a highly repetitive way, are connected to characteristics of NPs, such as their chemical nature, solubility, and shape. The protection of the encapsulated payload from metabolic or immunologic degradation, together with the possibility to specifically target tissues or cells, allows the reduction of antigen/drug dose while maintaining high efficacy levels and containing unwanted side effects. This feature can be particularly favorable in the case of oral/mucosal administration where the antigen is highly exposed to enzymatic or pH-dependent destruction. The conjugation of NPs with ligands of Toll-like receptors (TLRs) or other receptors selectively represented on different populations of antigen-presenting cells (APCs) not only leads to the targeting of such cells but may also drive their functional response by activating differential signal transduction pathways and releasing soluble mediators with distinct immunomodulatory activity.30
In the second case, in addition to inducing immune tolerance, NPs can be therapeutically deployed to reduce or block the unwanted and detrimental immune responses that occur in autoimmunity and allergy by directly exerting immunosuppressive effects on immune cells (APCs and T and B lymphocytes) or improving the delivery of immunosuppressive compounds/drugs by increasing their solubility and bioavailability.29,31 The upregulation of cytokines such as TGF-β and IL-10, associated with Tregs which control the activity of effector cells and induce apoptosis, mediates the direct effects on immune cells.29
An example is shown by the fullerene spherical NPs. These have been shown to have immunosuppressive effects on anaphylaxis by decreasing in vitro IgE receptor-mediated signaling and the degranulation of human mast cells and basophils, as well as preventing histamine release and the lowering of body temperature after allergen challenge in an in vivo mouse model of anaphylaxis.32 A similar experimental approach has been applied to the evaluation of the antiallergenic potential of biodegradable poly(DL-lactide-co-glycolide) (PLGA) NPs.33 The ability of PLGA NPs to inhibit mast cell-mediated allergic responses was demonstrated in vitro on the antigen-induced activation and release of beta-hexosaminidase and histamine from the RBL-2H3 rat basophilic leukemia cell line (which possesses the phenotypic properties of mucosal mast cells) and on systemic anaphylaxis induced in mice. More recently, NP approaches have also been introduced in the field of allergy diagnosis, demonstrating that the encapsulation of hydrophobic allergens within poly-ε-caprolactone NPs can improve the diagnosis of allergic contact dermatitis.34 Moreover, NPs can be specifically designed to interact with the immune system, exploiting their immunomodulatory potential for therapeutic purposes, suggesting that new NP/allergen pharmacological preparations can open the way to alternative therapeutic options or administration routes.35
In accordance with the main focus of this review on NPs which have been engineered to act as adjuvants/carriers in allergy immunotherapy, in the following section, we briefly introduce the most common categories of NMs used in this field. NP–allergen complexes will then be described according to the origin of their allergenic component, together with an analysis of their immunological profile and immunomodulatory activities.
Categories of nanomaterials/nanoparticles applied in allergen immunotherapy
Different categories of NPs have been developed and studied as adjuvants/carriers to improve the efficacy and safety of allergy immunotherapy. Their classification is not univocal because overlapping criteria can be applied (size and morphology, physicochemical features, function, etc). An attempt to group NPs used for the preparation of allergen complexes according to their chemical nature could include these major classes of products: polyesters, polysaccharide polymers and carbohydrate-based particles, liposomes, protamine-based NPs, and polyanhydrides.
Polyesters
Polymeric nanocarriers have been investigated as vaccine delivery systems due to their ability to provide the controlled release of antigens or adjuvants.36,37 Polyesters are a group of molecules whose biocompatibility, safety, and biodegradability in human applications are well-documented and which have already been approved by the US Food and Drug Administration (FDA).38 Several applications have been described using PLGA particles in sutures and implants, and their use in humans has been proposed for decades.39,40 Poly(glutamic acid) (PGA) and PLGA have been primarily investigated as biodegradable synthetic polymer nanocarriers. It is possible to modify their size, surface properties, and release profile (in particular, the biodegradation rate) to affect the immunogenicity of the encapsulated antigens as well as to optimize delivery through different administration routes (oral, mucosal, and systemic).41
Polysaccharide polymers and carbohydrate-based particles
Another molecular group that has been studied in this context is polysaccharides, which are easily produced polymers derived from natural sources. In this respect, poly(D-glucosamine) (known as chitosan) is a very abundant natural polysaccharide which is derived from shrimp and other crustacean shells. Chitosan is composed of randomly distributed β-(1-4)-linked D-glucosamine (deacetylated unit) and N-acetyl-D-glucosamine (acetylated unit) and has several advantageous properties, such as low production costs, biocompatibility, biodegradability, and absence of toxicity.42 As a natural, nontoxic polysaccharide, it has already been approved for human use in the US and Europe for bandages and other hemostatic agents. Notably, for applications in the field of immunomodulatory agents, chitosan can provide a cationic charge that may facilitate endocytosis as well as adjuvance that acts by promoting the maturation of dendritic cells (DCs). Another important property of chitosan is its mucoadhesiveness that makes it particularly attractive for targeting mucosal tissues and promoting mucosal immunity.36,43 Other carbohydrate-based particles (Sepharose beads) have also been suggested as a novel particulate adjuvant to improve allergy vaccinations through the enhancement of antigen uptake by DCs.44
Liposomes
Liposomes are spherical vesicles composed of one or more bilayer phases of phospholipids encompassing aqueous compartments; this double chemical nature allows the entrapment and delivery of both hydrophobic and hydrophilic molecules. Due to their high biocompatibility and biodegradability, nanosized liposomes have found widespread application as drug and gene carriers.45 The major advantages of their application in drug delivery are the possibility of enhancing the solubility and bioavailability of the protein cargo, as well as its in vivo stability, to protect it from undesired interactions with other cells/molecules and optimize specific targeting of the action site of a drug.46
Protamine-based NPs
Protamines are biodegradable arginine-rich peptides of approximately 4 kDa that have been used for decades in human medications.47,48 Protamine NPs complexed with DNA or RNA oligonucleotides can be generated via electrostatic interaction, showing optimal biocompatibility and high stability. Moreover, since protamine contains a nuclear localization signal,49 protamine-based NPs can easily transport their content into the nucleus, thus improving the efficiency of specific gene therapy. In the context of this review, they are attractive elements for the formation of complexes with oligonucleotides, strong activators of Toll like receptors.
Polyanhydrides
Among biodegradable polymers, amphiphilic polyanhydride nanocarriers present unique properties in their capacity to modulate immune responses, together with a favorable safety profile. In fact, they have been demonstrated to be taken up by DCs and to induce antigen-specific proliferation of both CD4+ and CD8+ T cells.50 Their properties, such as release kinetics, antigen retention, and adjuvant effect, can be tailored based on polymer composition, with particular attention to the degree of hydrophobicity. Surface modifications can be introduced to enhance or reduce their mucus permeability and bioadhesion to specific target cells. For specific application in the field of allergy immunotherapy, Gantrez® AN 119 (poly[methyl vinyl ether-co-maleic anhydride]) has been investigated. In fact, Gantrez AN 119 is a copolymer of methyl vinyl ether and maleic anhydride that can easily react with amino groups, making it easy to load or link different types of proteins, including allergens.51
Nanoparticle-allergen complexes
Different classes of allergens have been studied in association with nanostructured adjuvants and/or carriers engineered for allergy immunotherapy. In vitro studies on human immune cells as well as mouse models of sensitization have been applied to the functional evaluation of such complexes. In the next section, these complexes have been categorized according to the origin of the allergenic molecules, putting special emphasis on the novel beneficial immunoregulatory properties introduced by their formation into complexes and investigating different routes of administration in vivo.
Pollen allergens
Birch pollen is a major allergenic source in Central Europe as well as in North America, and almost all birch allergic patients are sensitized to the Bet v 1 allergen; thus, the development of a product containing this molecule would be a useful drug for all birch allergic patients. In addition, the Bet v 1 allergenic molecule displays cross reactivity with several others allergens from different pollens and plant derived food. In particular, it has been reported that more than 70% of patients with birch pollen allergy react to at least 1 Bet v 1-associated allergenic food source, including vegetables, nuts, and legumes demonstrating the relevant role of this family of allergens.52 In a prophylactic experimental set-up, Schöll et al demonstrated that PLGA particles loaded with rBet v 1 (the major allergen of birch pollen) support the immunogenicity of the allergen in an animal system. This research group treated Bet v 1 allergic mice subcutaneously (SC) with different allergen formulations and evaluated their relative immunological properties. Bet v 1 allergen was encapsulated in PLGA in the presence or absence of Al(OH)3, or used absorbed in Al(OH)3 only, mimicking commercially available products. PLGA-entrapped Bet v 1 proved to be successful in modifying the ongoing Th2 response, leading to a decrease in IgG1 antibody production and an increase in IgG2a antibody titer after a few shots. These serological data were reinforced by the fact that allergen-loaded PLGA nanoparticles modulate the Th2 response (reduction of IgG1 antibodies) meanwhile IFN-γ and IL-10 were produced after the allergen-specific stimulation of splenocytes.53
Profilins are a widespread family of highly cross-reactive allergens which have been isolated in a large number of sources, so much so that this allergen is considered a true panallergenic molecule.54 Xiao et al synthesized a complex containing PLGA with a recombinant profilin from Caryota mitis (rCmP). The Authors showed that the rCmP-loaded PLGA NPs effectively inhibited the generation of allergen-specific IgE and the secretion of the pro-inflammatory Th2 cytokine IL-4, facilitating the generation of allergen-specific IgG2a and the secretion of the Th1 cytokines in vivo.55 The same NPs were employed in a different immunotherapy regimens by Salari et al, targeting another relevant cross-reactive allergen, Che a 3 protein belonging to the polcalcin family, an allergen showing high levels of identity with polcalcin from olive, birch, alder, rapeseed, and timothy grass pollens.54 BALB/c mice were sensitized to rChe a 3 and then treated sublingually, either with soluble rChe a 3 or PLGA-encapsulated rChe a 3. In vitro and ex vivo assays demonstrated significantly increased antigen-specific IgG2a. In addition, IL-4 levels in restimulated splenocytes were significantly reduced, while IFN-γ, IL-10, and TGF-β levels, as well as Foxp3 expression, were significantly higher than in control groups. This suggests that this formulation can induce a stronger Th1/Treg pathway than the purified allergen.56
A different strategy was proposed by Marazuela et al, describing the use of PLGA as a vehicle for the intranasal (IN) administration of the OLE109–130 peptide (a major Ole e 1 T cell epitope) in a mouse sensitization model. The pre-treatment of BALB/c mice with OLE109–130–PLGA complex before sensitization to the whole Ole e 1 (a major allergen of olive pollen, with a high degree of homology to allergens from other allergenic sources57) led to the significant inhibition of allergen-specific IgE and IgG1 levels, with a marked increase of specific IgG2a antibodies. Moreover, IL-5 and IL-10 levels in spleen cell cultures were suppressed in peptide-PLGA pre-treated mice, suggesting that pre-treatment with the OLE109–130–PLGA complex is effective at preventing subsequent allergic sensitization to Ole e 1.58,59
A different class of polymers belonging to this family of particles has also been tested by Broos et al, who demonstrated that 200 nm-sized biodegradable poly(g-glutamic acid) (γ-PGA) NPs activate human monocyte-derived dendritic cells (MoDCs). In this study, it was established that MoDCs from grass allergic patients stimulated with a mixture of γ-PGA and Phleum pratense extract augment the production of allergen-specific IL-10, a relevant cytokine for the induction of peripheral tolerance.60
Another polylactide particle used to generate new allergen complexes is poly(D,L-lactide-co-glycolide) (PLG). PLG microparticles were introduced in clinical contexts several years ago, exhibiting a capacity to slow the release of the entrapped antigens.61 In particular, Batanero et al used this polymer to entrap nOle e 1 allergen. The intraperitoneal (IP) immunization of such complexes in mice elicited high levels of specific IgG2a antibodies and low levels of both total IgE and specific IgG1 antibodies, demonstrating that this product induces a Th1-like immune response. On the other hand, immunization with nOle e 1 absorbed in alum only induced high levels of both specific IgG1 and total IgE, showing the adjuvancy capability of the particle in the context of the allergen specific immunological response.62
Carbohydrate-based particles (CBPs) covalently coupled to the timothy grass pollen allergen Phl p 5 (a major grass allergen) also exhibited desirable adjuvant properties. When compared with alum-absorbed Phl p 5, Grönlund et al reported that both adjuvants induced significantly increased allergen-specific antibody responses compared with soluble Phl p 5, but, unlike alum, CBP-Phl p 5 did not induce granulomatous tissue reactions. This demonstrated that this kind of formulation can reduce one of the most common local side effects observed following intradermal or subcutaneous injections.63
Recently, it has been demonstrated that liposomes coated with a neoglycolipid constructed from mannotriose and dipalmitoylphosphatidylethanolamine (Man3-DPPE) (oligomannose-coated liposomes, OMLs) can induce a strong Th1 immune response against encapsulated antigens in mice, with significant IFN-γ production and suppressed IL-4 production.64,65 These findings open the possibility that the administration of OMLs with entrapped allergens may control allergic reactions to Japanese cedars, using the major allergen Cry j 1, an allergen belonging to the Pectase lysase family. The pre-treatment of unsensitized mice with Cry j 1/OMLs blocked total and allergen-specific IgE levels in sera from Cry j 1 sensitized animals, with decreased IgG1 as well and significantly increased levels of specific IgG2a.66–68
Liposome-protamine-DNA (LPD) NPs are safe, effective, and non-toxic adjuvants that induce Th1-like immune responses. Nouri et al demonstrated that the encapsulation of rChe a 1, rChe a 2, and rChe a 3 allergens (produced in a recombinant hybrid form [rHM]) from Chenopodium album resulted in a reduction in specific IgE and a marked increase in IgG2a in a murine model. Furthermore, LPD-rHM induced allergen-specific responses with high IFN-γ production, as well as the expression of T-bet (a transcription factor that controls the expression of Th1 cytokines) in stimulated splenocytes.69
Gómez et al investigated the possibility of using a particle composed of Gantrez NPs with Lolium protein extract plus lipopolysaccharides (LPSs). The immunotherapeutic capacity of this formulation was evaluated in a model of mice which had been pre-sensitized to Lolium perenne. Once the animals were sensitized, allergens/NPs were administered and then mice were challenged with Lolium extract IP and the intensity of anaphylaxis was studied. After immunization, the animals treated with LPS–Lolium–NP showed IgG2a antibody levels 100-times higher than controls or Lolium–alum challenged mice. This observation demonstrated that the combination of the copolymer with LPS can induce a Th1 polarization that can counteract the existing Th2 polarization.70
Parietaria pollen is characterized by the presence of two major allergens which are capable of inducing IgE production in the majority of allergic patients. For this reason, recombinant Parietaria major allergens were entrapped within two independent classes of NPs whose bioavailability was evaluated by means of a basophil activation assay, showing that complex formation does not inhibit their availability to the IgE antibodies of allergic patients.71,72
Refer to Table 1 for a summary of this section.
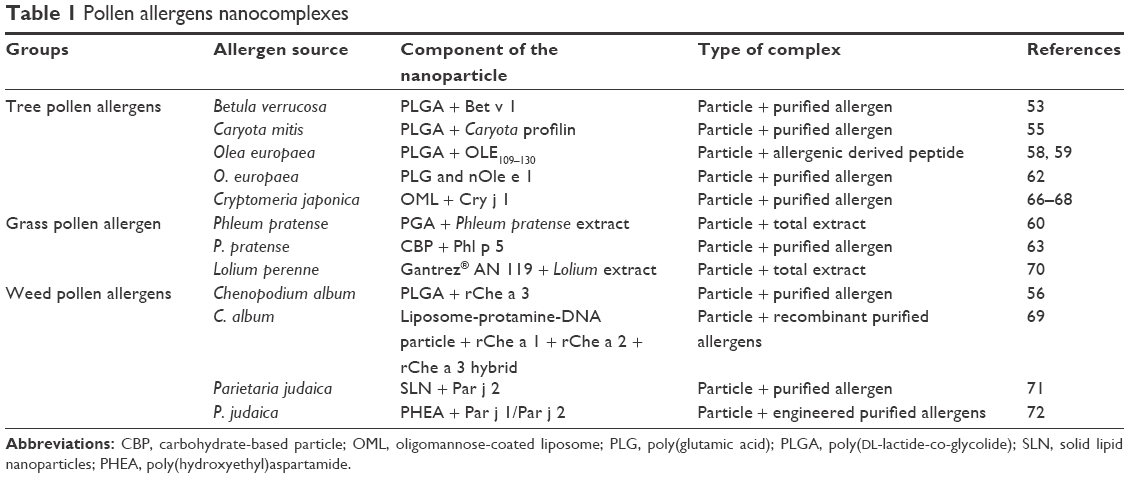 | Table 1 Pollen allergens nanocomplexes |
Arthropoda allergens
Joshi et al developed a strategy based on the co-encapsulation of PLGA with one of the major house dust mite (HDM) allergens (Der p 2) and unmethylated cytosine-phosphate-guanine (CpG) nucleotides.73 CpG oligonucleotides are known to be potent adjuvants, shifting immune responses to the Th1 type since they are TLR-9 agonists.74,75 The authors showed that the co-delivery of PLGA particles loaded with an antigen and CpG can stimulate a potent antigen-specific immune response, as well as that the magnitude of the immune response is correlated to the size of the PLGA particles used for immunization demonstrating that even the dimension of the NP can influence the allergen specific immune response.73
Comparable results using CpG oligonucleotides were further reported by Martìnez Gómez et al using the bee venom phospholipase A2 (PLA2) allergen as a model. Both naïve and bee venom allergic mice were immunized with microparticles containing only PLA2, showing that this treatment induced a weak antibody response. On the other hand, when PLA2-PLGA NPs were combined with CpG, a strong PLA2-specific antibody response was observed, and the presence of CpG was required for the induction of the Th1-associated isotype IgG2a. The effect of the CpG nucleotide was further strengthened when protamine was co-encapsulated for the complexation of CpG.76
Chitosan has been used for its mucoadhesiveness that makes this particle attractive for mucosal immunity.43 Following this line of evidence, Liu et al tested the immunotherapeutic efficiency of the IN administration of Dermatophagoides farinae extract (Der f) entrapped in chitosan microparticles. BALB/c mice were IP sensitized with Der f extract absorbed in alum, followed by IN treatment with PBS, chitosan, Der f or Der f-chitosan nano-vaccine for 6 weeks. The mice were subsequently challenged intranasally with Der f extract for 1 week, showing that this complex can induce immunological protection in a murine model of allergic asthma by inducing regulatory T cells and Th1-type reaction.77 In a similar way, Li et al entrapped a major epitope peptide of the mite group 2 allergen Der f 2 (Der f 2 47–67) in chitosan microparticles which were IP injected into BALB/c mice which had been previously intranasally challenged with a Der f extract allergen. Mice immunized with this protocol displayed decreased airway hyperreactivity, lung inflammation, and mucus production. Antibody production analysis showed a reduced serum level of Der f-specific IgE and increased specific IgG2a,78 Furthermore, another strategy utilizing chitosan was to mix it with hydroxybenzotriazole (HOBt), hydrophobic biomolecules, ie, phenylalanine (Phe), and hydrophilic polymers, ie, poly(ethylene glycol) methyl ether (mPEG), to produce positively charged NPs (20–50 nm in size) capable of entrapping negatively charged HDM extract (Dermatophagoides pteronyssinus). The HDM-entrapped CS-Phe-mPEG showed biocompatibility favoring T cell immune response and a synergistic effect of chitosan, Phe and mPEG with a clear increase in IFN-γ and IL-10 production in both allergic and healthy volunteers was reported.79
Liposomes have been complexed with cockroach allergens, a frequently cited source of severe asthma. Meechan et al studied the therapeutic efficacy of an IN liposome-adjuvant vaccine made of a refined Periplaneta americana arginine kinase (AK, Per a 9), compared to a liposome-entrapped P. americana crude extract (CRE) vaccine in adult BALB/c mice. The liposome-entrapped native AK attenuated airway inflammation after CRE provocation and caused a shift from Th2 to Th1 and Treg responses. Furthermore, this study demonstrated that IN liposome adjuvanted cockroach allergens containing native Per a 9 are more effective than complexes with liposome and crude extracts at attenuating allergy airway inflammation and allowing a more precise standardization of the product.80
Refer to Table 2 for a summary of this section.
 | Table 2 Arthropoda allergens nanocomplexes |
Other allergens (animal, food)
Thunberg et al showed that CBP-rFel d 1 (the major cat allergen) induces rapid allergen-specific antibody production and prevents the induction of allergic immune responses in mice sensitized to rFel d 1. Interestingly, in contrast to alum absorbed rFel d 1, CBPrFel d 1 remains at the injection site longer, enabling prolonged antigen exposure, working as an adjuvant for a safer and more efficient allergy vaccination.81
Another relevant feature of these types of complexes was introduced by Weinberger et al who showed that the coupling of a carbohydrate to a protein can mask B-cell epitopes, making the protein “hypoallergenic”. In particular, they showed that the hypoallergenicity of neoglycoconjugates depends on the carbohydrate, the nature of the allergen, and the chemistry used for the coupling reaction, suggesting this strategy as a promising one that may lead to safer formulations.82
Food allergies affect approximately 5% of the population, severely impacting patient quality of life. Several strategies have been reported in the formation of NP complexes with single allergens or allergenic extracts.
Protamines have been used for a long time in clinical settings as molecules which are easy to handle in the making of complex nucleic acids via electrostatic interaction. For this reason, protamine-nAra h 2-CpG particles were prepared and injected SC into naïve BALB/c mice, showing an increase in Ara h 2-specific IgG2a with no detectable IgE specific antibodies, and, of particular interest, Pali-Schöll et al reported that granuloma formation was completely absent at injection sites.83
Furthermore, De S Rebouças et al suggested that poly(anhydride) NPs could be loaded with raw or roasted peanut proteins. These amphiphilic nanocarriers can modulate immune response per se, and their adjuvant capacity was evaluated in an animal model of food allergy. NP formulations were studied after oral immunization of C57BL/6 mice, showing low specific IgE induction. Furthermore, oral immunization with spray-dried NPs loaded with peanut proteins showed a significant decrease in splenic Th2 cytokines and the enhancement of both Th1 and regulatory cytokines.84
In a different way, Srivastava et al investigated the efficacy and safety of peanut oral immunotherapy using CpG-coated poly(lactic-coglycolic acid) NPs containing a peanut extract (CpG/PNNPs). In particular, C3H/HeJ mice were rendered peanut allergic by means of oral sensitization with peanut and cholera toxin. Mice were subjected to four weekly gavages with CpG/PN-NPs, vehicle (PBS), NPs alone, peanut alone, CpG NPs, or peanut NPs. Mice with peanut allergy treated with CpG/PN-NPs but not with a vehicle or other components were significantly protected from anaphylaxis. Treatment was associated with decreased levels of Th2 cytokines (IL-4, IL-5, and IL-13) but increased IFN-γ levels, together with decreased peanut-specific IgE/IgG1 levels and an increase in IgG2a levels.85
Refer to Table 3 for a summary of this section.
 | Table 3 Animal and food allergens nanocomplexes |
Conclusion
Several studies on allergen immunotherapy performed with commercially available products in comparison with pharmaceutical treatment have shown that immunotherapy may be very beneficial for patients and health care systems, reducing costs and bringing relief to patients.86 However, some questions still remain regarding the safety of these treatments and the need to reduce the dose and time of administration to improve patient compliance. In recent years, nanotechnologies have strongly emerged in vaccinology and, more currently, in the allergen immunotherapy field, with several studies in animals showing very promising future perspectives.35,87 Of particular interest in this field is how nanocarrier/allergen complexes can be formulated to obtain improved stability, slow release kinetics, selective targeting of specific immune cells, and lower immunotoxicity vs optimized immunomodulatory potential. Although such applications are still at an investigational stage, in vitro and in vivo studies have started to open the way to deeper knowledge about the interaction between the immune system and NPs/allergen complexes. The results suggest that several NP-based formulations can be applied in the near future to allergen immunotherapy to solve its unmet needs.
Disclosure
The authors report no conflicts of interest in this work.
References
Bousquet J, Anto J, Auffray C, et al. MeDALL (Mechanisms of the Development of ALLergy): an integrated approach from phenotypes to systems medicine. Allergy. 2011;66(5):596–604. | ||
Newson RB, van Ree R, Forsberg B, et al. Geographical variation in the prevalence of sensitization to common aeroallergens in adults: the GA(2) LEN survey. Allergy. 2014;69(5):643–651. | ||
Devereux G. The increase in the prevalence of asthma and allergy: food for thought. Nat Rev Immunol. 2006;6(11):869–874. | ||
Akdis CA, Blaser K, Akdis M. Genes of tolerance. Allergy. 2004;59(9):897–913. | ||
Jutel M, Agache I, Bonini S, et al. International consensus on allergy immunotherapy. J Allergy Clin Immunol. 2015;136(3):556–568. | ||
Akdis M, Verhagen J, Taylor A, et al. Immune responses in healthy and allergic individuals are characterized by a fine balance between allergen-specific T regulatory 1 and T helper 2 cells. J Exp Med. 2004;199(11):1567–1575. | ||
Durham SR, Ying S, Varney VA, et al. Grass pollen immunotherapy inhibits allergen-induced infiltration of CD4+ T lymphocytes and eosinophils in the nasal mucosa and increases the number of cells expressing messenger RNA for interferon-gamma. J Allergy Clin Immunol. 1996;97(6):1356–1365. | ||
Sandrini A, Rolland JM, O’Hehir RE. Current developments for improving efficacy of allergy vaccines. Expert Rev Vaccines. 2015;14(8):1073–1087. | ||
Nouri-Aria KT, Wachholz PA, Francis JN, et al. Grass pollen immunotherapy induces mucosal and peripheral IL-10 responses and blocking IgG activity. J Immunol. 2004;172(5):3252–3259. | ||
Pilette C, Nouri-Aria KT, Jacobson MR, et al. Grass pollen immunotherapy induces an allergen-specific IgA2 antibody response associated with mucosal TGF-beta expression. J Immunol. 2007;178(7):4658–4666. | ||
Radulovic S, Jacobson MR, Durham SR, Nouri-Aria KT. Grass pollen immunotherapy induces Foxp3-expressing CD4+ CD25+ cells in the nasal mucosa. J Allergy Clin Immunol. 2008;121(6):1467–1472, 1472e1. | ||
Shamji MH, Durham SR. Mechanisms of immunotherapy to aeroallergens. Clin Exp Allergy. 2011;41(9):1235–1246. | ||
Akdis M, Akdis CA. Mechanisms of allergen-specific immunotherapy: multiple suppressor factors at work in immune tolerance to allergens. J Allergy Clin Immunol. 2014;133(3):621–631. | ||
Canonica GW, Cox L, Pawankar R, et al. Sublingual immunotherapy: World Allergy Organization position paper 2013 update. World Allergy Organ J. 2014;7(1):6. | ||
Radulovic S, Wilson D, Calderon M, Durham S. Systematic reviews of sublingual immunotherapy (SLIT). Allergy. 2011;66(6):740–752. | ||
Kiel MA, Röder E, Gerth van Wijk R, Al MJ, Hop WC, Rutten-van Mölken MP. Real-life compliance and persistence among users of subcutaneous and sublingual allergen immunotherapy. J Allergy Clin Immunol. 2013;132(2):353–360.e2. | ||
Etheridge ML, Campbell SA, Erdman AG, Haynes CL, Wolf SM, McCullough J. The big picture on nanomedicine: the state of investigational and approved nanomedicine products. Nanomedicine. 2013;9(1):1–14. | ||
Weissig V, Pettinger TK, Murdock N. Nanopharmaceuticals (part 1): products on the market. Int J Nanomedicine. 2014;9:4357–4373. | ||
Duncan R, Gaspar R. Nanomedicine(s) under the microscope. Mol Pharm. 2011;8(6):2101–2141. | ||
Official Journal of the European Union. Commission Recommendation of 18 October 2011 on the definition of nanomaterial. European Commission; 2011. Available from: http://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2011:275:0038:0040:EN:PDF. Accessed May 26, 2017. | ||
Wu L, Zhang J, Watanabe W. Physical and chemical stability of drug nanoparticles. Adv Drug Deliv Rev. 2011;63(6):456–469. | ||
Nyström AM, Fadeel B. Safety assessment of nanomaterials: implications for nanomedicine. J Control Release. 2012;161(2):403–408. | ||
Ehmann F, Sakai-Kato K, Duncan R, et al. Next-generation nanomedicines and nanosimilars: EU regulators’ initiatives relating to the development and evaluation of nanomedicines. Nanomedicine (Lond). 2013;8(5):849–856. | ||
Havel H, Finch G, Strode P, et al. Nanomedicines: from bench to bedside and beyond. AAPS J. 2016;18(6):1373–1378. | ||
Kaur IP, Kakkar V, Deol PK, Yadav M, Singh M, Sharma I. Issues and concerns in nanotech product development and its commercialization. J Control Release. 2014;193:51–62. | ||
Farcal L, Torres Andón F, Di Cristo L, et al. Comprehensive in vitro toxicity testing of a panel of representative oxide nanomaterials: first steps towards an intelligent testing strategy. PLoS One. 2015;10(5):e0127174. | ||
Dobrovolskaia MA, Shurin M, Shvedova AA. Current understanding of interactions between nanoparticles and the immune system. Toxicol Appl Pharm. 2016;299:78–89. | ||
Giannakou C, Park MV, de Jong WH, van Loveren H, Vandebriel RJ, Geertsma RE. A comparison of immunotoxic effects of nanomedicinal products with regulatory immunotoxicity testing requirements. Int J Nanomedicine. 2016;11:2935–2952. | ||
Smith DM, Simon JK, Baker JR Jr. Applications of nanotechnology for immunology. Nat Rev Immunol. 2013;13(8):592–605. | ||
Klinman DM, Sato T, Shimosato T. Use of nanoparticles to deliver immunomodulatory oligonucleotides. Wiley Interdiscip Rev Nanomed Nanobiotechnol. 2016;8(4):631–637. | ||
Zolnik BS, González-Fernández A, Sadrieh N, Dobrovolskaia MA. Nanoparticles and the immune system. Endocrinology. 2010;151(2):458–465. | ||
Ryan JJ, Bateman HR, Stover A, et al. Fullerene nanomaterials inhibit the allergic response. J Immunol. 2007;179(1):665–672. | ||
Tahara K, Tadokoro S, Yamamoto H, Kawashima Y, Hirashima N. The suppression of IgE-mediated histamine release from mast cells following exocytic exclusion of biodegradable polymeric nanoparticles. Biomaterials. 2012;33(1):343–351. | ||
Cortial A, Nosbaum A, Rozières A, et al. Encapsulation of hydrophobic allergens into nanoparticles improves the in vitro immunological diagnosis of allergic contact dermatitis. Nanomedicine. 2015;11(4):1029–1033. | ||
Di Felice G, Barletta B, Bonura A, Butteroni C, Corinti S, Colombo P. Nanoparticles adjuvants in allergology: new challenges and pitfalls. Curr Pharm Des. 2015;21(29):4229–4239. | ||
Kim MG, Park JY, Shon Y, Kim G, Shim G, Oh YK. Nanotechnology and vaccine development. Asian J Pharm Sci. 2014;9(5):227–235. | ||
Zhao L, Seth A, Wibowo N, et al. Nanoparticle vaccines. Vaccine. 2014;32(3):327–337. | ||
Soppimath KS, Aminabhavi TM, Kulkarni AR, Rudzinski WE. Biodegradable polymeric nanoparticles as drug delivery devices. J Control Release. 2001;70(1–2):1–20. | ||
Coombes AG, Meikle MC. Resorbable synthetic polymers as replacements for bone graft. Clin Mater. 1994;17(1):35–67. | ||
Postlethwait RW, Smith BM. A new synthetic absorbable suture. Surg Gynecol Obstet. 1975;140(3):377–380. | ||
Han FY, Thurecht KJ, Whittaker AK, Smith MT. Bioerodable PLGA-based microparticles for producing sustained-release drug formulations and strategies for improving drug loading. Front Pharmacol. 2016;7:185. | ||
Ahmed TA, Aljaeid BM. Preparation, characterization, and potential application of chitosan, chitosan derivatives, and chitosan metal nanoparticles in pharmaceutical drug delivery. Drug Des Devel Ther. 2016;10:483–507. | ||
Landriscina A, Rosen J, Friedman AJ. Biodegradable chitosan nanoparticles in drug delivery for infectious disease. Nanomedicine (Lond). 2015;10(10):1609–1619. | ||
Kovacsovics-Bankowski M, Clark K, Benacerraf B, Rock KL. Efficient major histocompatibility complex class I presentation of exogenous antigen upon phagocytosis by macrophages. Proc Natl Acad Sci U S A. 1993;90(11):4942–4946. | ||
Bernasconi V, Norling K, Bally M, Höök F, Lycke NY. Mucosal vaccine development based on liposome technology. J Immunol Res. 2016;2016:5482087. | ||
Akbarzadeh A, Rezaei-Sadabady R, Davaran S, et al. Liposome: classification, preparation, and applications. Nanoscale Res Lett. 2013;8(1):102. | ||
Owens DR. Insulin preparations with prolonged effect. Diabetes Technol Ther. 2011;13 (Suppl 1):S5–S14. | ||
Schulman S, Bijsterveld NR. Anticoagulants and their reversal. Transfus Med Rev. 2007;21(1):37–48. | ||
Martins RP, Ostermeier GC, Krawetz SA. Nuclear matrix interactions at the human protamine domain: a working model of potentiation. J Biol Chem. 2004;279(50):51862–51868. | ||
Torres MP, Wilson-Welder JH, Lopac SK, et al. Polyanhydride microparticles enhance dendritic cell antigen presentation and activation. Acta Biomater. 2011;7(7):2857–2864. | ||
Gómez S, Gamazo C, San Roman B, Vauthier C, Ferrer M, Irachel JM. Development of a novel vaccine delivery system based on Gantrez nanoparticles. J Nanosci Nanotechnol. 2006;6(9–10):3283–3289. | ||
Geroldinger-Simic M, Zelniker T, Aberer W, et al. Birch pollen-related food allergy: clinical aspects and the role of allergen-specific IgE and IgG4 antibodies. J Allergy Clin Immunol. 2011;127(3):616–622.e1. | ||
Schöll I, Weissenböck A, Förster-Waldl E, et al. Allergen-loaded biodegradable poly(D,L-lactic-co-glycolic) acid nanoparticles down-regulate an ongoing Th2 response in the BALB/c mouse model. Clin Exp Allergy. 2004;34(2):315–321. | ||
McKenna OE, Asam C, Araujo GR, Roulias A, Goulart LR, Ferreira F. How relevant is panallergen sensitization in the development of allergies? Pediatr Allergy Immunol. 2016;27(6):560–568. | ||
Xiao X, Zeng X, Zhang X, et al. Effects of Caryota mitis profilin-loaded PLGA nanoparticles in a murine model of allergic asthma. Int J Nanomedicine. 2013;8:4553–4562. | ||
Salari F, Varasteh AR, Vahedi F, Hashemi M, Sankian M. Down-regulation of Th2 immune responses by sublingual administration of poly (lactic-co-glycolic) acid (PLGA)-encapsulated allergen in BALB/c mice. Int Immunopharmacol. 2015;29(2):672–678. | ||
Valenta R, Twaroch T, Swoboda I. Component-resolved diagnosis to optimize allergen-specific immunotherapy in the Mediterranean area. J Investig Allergol Clin Immunol. 2007;17 (Suppl 1):36–40. | ||
Marazuela EG, Prado N, Moro E, et al. Intranasal vaccination with poly(lactide-co-glycolide) microparticles containing a peptide T of Ole e 1 prevents mice against sensitization. Clin Exp Allergy. 2008;38(3):520–528. | ||
Marazuela EG, Rodríguez R, Fernández-García H, García MS, Villalba M, Batanero E. Intranasal immunization with a dominant T-cell epitope peptide of a major allergen of olive pollen prevents mice from sensitization to the whole allergen. Mol Immunol. 2008;45(2):438–445. | ||
Broos S, Lundberg K, Akagi T, et al. Immunomodulatory nanoparticles as adjuvants and allergen-delivery system to human dendritic cells: implications for specific immunotherapy. Vaccine. 2010;28(31):5075–5085. | ||
Gupta RK, Singh M, O’Hagan DT. Poly(lactide-co-glycolide) microparticles for the development of single-dose controlled-release vaccines. Adv Drug Deliv Rev. 1998;32(3):225–246. | ||
Batanero E, Barral P, Villalba M, Rodríguez R. Encapsulation of Ole e 1 in biodegradable microparticles induces Th1 response in mice: a potential vaccine for allergy. J Control Release. 2003;92(3):395–398. | ||
Grönlund H, Vrtala S, Wiedermann U, et al. Carbohydrate-based particles: a new adjuvant for allergen-specific immunotherapy. Immunology. 2002;107(4):523–529. | ||
Kojima N, Biao L, Nakayama T, Ishii M, Ikehara Y, Tsujimura K. Oligomannose-coated liposomes as a therapeutic antigen-delivery and an adjuvant vehicle for induction of in vivo tumor immunity. J Control Release. 2008;129(1):26–32. | ||
Shimizu Y, Takagi H, Nakayama T, et al. Intraperitoneal immunization with oligomannose-coated liposome-entrapped soluble leishmanial antigen induces antigen-specific T-helper type immune response in BALB/c mice through uptake by peritoneal macrophages. Parasite Immunol. 2007;29(5):229–239. | ||
Ishii M, Kato C, Hakamata A, Kojima N. Targeting with oligomannose-coated liposomes promotes maturation and splenic trafficking of dendritic cells in the peritoneal cavity. Int Immunopharmacol. 2011;11(2):164–171. | ||
Ishii M, Kojima N. Mucosal adjuvant activity of oligomannose-coated liposomes for nasal immunization. Glycoconj J. 2010;27(1):115–123. | ||
Ishii M, Koyama A, Iseki H, Narumi H, Yokoyama N, Kojima N. Anti-allergic potential of oligomannose-coated liposome-entrapped Cry j 1 as immunotherapy for Japanese cedar pollinosis in mice. Int Immunopharmacol. 2010;10(9):1041–1046. | ||
Nouri HR, Varasteh A, Jaafari MR, Davies JM, Sankian M. Induction of a Th1 immune response and suppression of IgE via immunotherapy with a recombinant hybrid molecule encapsulated in liposome-protamine-DNA nanoparticles in a model of experimental allergy. Immunol Res. 2015;62(3):280–291. | ||
Gómez S, Gamazo C, San Roman B, et al. A novel nanoparticulate adjuvant for immunotherapy with Lolium perenne. J Immunol Methods. 2009;348(1–2):1–8. | ||
Bondì ML, Montana G, Craparo EF, et al. Lipid nanoparticles as delivery vehicles for the Parietaria judaica major allergen Par j 2. Int J Nanomedicine. 2011;6:2953–2962. | ||
Licciardi M, Montana G, Bondì ML, et al. An allergen-polymeric nanoaggregate as a new tool for allergy vaccination. Int J Pharm. 2014;465(1–2):275–283. | ||
Joshi VB, Adamcakova-Dodd A, Jing X, et al. Development of a poly (lactic-co-glycolic acid) particle vaccine to protect against house dust mite induced allergy. AAPS J. 2014;16(5):975–985. | ||
Mohri K, Nishikawa M, Takahashi Y, Takakura Y. DNA nanotechnology-based development of delivery systems for bioactive compounds. Eur J Pharm Sci. 2014;58:26–33. | ||
Creticos PS, Schroeder JT, Hamilton RG, et al; Immune Tolerance Network Group. Immunotherapy with a ragweed-toll-like receptor 9 agonist vaccine for allergic rhinitis. N Engl J Med. 2006;355(14):1445–1455. | ||
Martìnez Gómez JM, Fischer S, Csaba N, et al. A protective allergy vaccine based on CpG- and protamine-containing PLGA microparticles. Pharm Res. 2007;24(10):1927–1935. | ||
Liu Z, Guo H, Wu Y, Yu H, Yang H, Li J. Local nasal immunotherapy: efficacy of Dermatophagoides farinae-chitosan vaccine in murine asthma. Int Arch Allergy Immunol. 2009;150(3):221–228. | ||
Li J, Liu Z, Wu Y, Wu H, Ran P. Chitosan microparticles loaded with mite group 2 allergen Der f 2 alleviate asthma in mice. J Investig Allergol Clin Immunol. 2008;18(6):454–460. | ||
Jirawutthiwongchai J, Klaharn IY, Hobang N, et al. Chitosan-phenylalanine-mPEG nanoparticles: from a single step water-based conjugation to the potential allergen delivery system. Carbohydr Polym. 2016;141:41–53. | ||
Meechan P, Tungtrongchitr A, Chaisri U, et al. Intranasal, liposome-adjuvanted cockroach allergy vaccines made of refined major allergen and whole-body extract of Periplaneta americana. Int Arch Allergy Immunol. 2013;161(4):351–362. | ||
Thunberg S, Neimert-Andersson T, Cheng Q, et al. Prolonged antigen-exposure with carbohydrate particle based vaccination prevents allergic immune responses in sensitized mice. Allergy. 2009;64(6):919–926. | ||
Weinberger EE, Himly M, Myschik J, et al. Generation of hypoallergenic neoglycoconjugates for dendritic cell targeted vaccination: a novel tool for specific immunotherapy. J Control Release. 2013;165(2):101–109. | ||
Pali-Schöll I, Szöllösi H, Starkl P, et al. Protamine nanoparticles with CpG-oligodeoxynucleotide prevent an allergen-induced Th2-response in BALB/c mice. Eur J Pharm Biopharm. 2013;85(3 Pt A):656–664. | ||
De S Rebouças J, Irache JM, Camacho AI, et al. Immunogenicity of peanut proteins containing poly(anhydride) nanoparticles. Clin Vaccine Immunol. 2014;21(8):1106–1112. | ||
Srivastava KD, Siefert A, Fahmy TM, Caplan MJ, Li XM, Sampson HA. Investigation of peanut oral immunotherapy with CpG/peanut nanoparticles in a murine model of peanut allergy. J Allergy Clin Immunol. 2016;138(2):536–543.e4. | ||
Jutel M, Agache I, Bonini S, et al. International Consensus on Allergen Immunotherapy II: mechanisms, standardization, and pharmacoeconomics. J Allergy Clin Immunol. 2016;137(2):358–368. | ||
Gamazo C, Gastaminza G, Ferrer M, Sanz ML, Irache JM. Nanoparticle based-immunotherapy against allergy. Immunotherapy. 2014;6(7):885–897. |
 © 2017 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2017 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.