Back to Journals » Infection and Drug Resistance » Volume 12
Multilocus sequence typing and blaESBL characterization of extended-spectrum beta-lactamase-producing Escherichia coli isolated from healthy humans and swine in Northern Thailand
Authors Seenama C, Thamlikitkul V, Ratthawongjirakul P
Received 22 March 2019
Accepted for publication 3 July 2019
Published 19 July 2019 Volume 2019:12 Pages 2201—2214
DOI https://doi.org/10.2147/IDR.S209545
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Suresh Antony
Chakkraphong Seenama,1,2 Visanu Thamlikitkul,2 Panan Ratthawongjirakul3
1Program of Molecular Sciences in Medical Microbiology and Immunology, Department of Transfusion Medicine and Clinical Microbiology, Faculty of Allied Health Sciences, Chulalongkorn University, Bangkok, Thailand; 2Division of Infectious Diseases and Tropical Medicine, Department of Medicine, Faculty of Medicine Siriraj Hospital, Mahidol University, Bangkok, Thailand; 3Research Group of Innovative Diagnosis of Antimicrobial Resistance, Department of Transfusion Medicine and Clinical Microbiology, Faculty of Allied Health Sciences, Chulalongkorn University, Bangkok, Thailand
Purpose: Here, we investigated the genetic relationships and characteristics of extended- spectrum beta-lactamase-producing Escherichia coli (ESBL-E. coli) isolates from healthy hosts, humans in the community and swine among the livestock of Amphor Mueang, Lamphun Province, Thailand.
Patients and methods: Four hundred and nine rectal swabs were collected from healthy people and swine. A total of 212 ESBL-E. coli was isolated and phenotypically confirmed by a combination disk method. Putative ESBL-encoding genes, including blaCTX-M, blaTEM, and blaSHV, were examined by multiplex-PCR. Randomly selected 42 ESBL-E. coli isolates were whole genome sequenced to characterize the ESBL-encoding genes and identify additional antimicrobial resistance genes. The genetic relatedness of 212 ESBL-E. coli was investigated by multilocus sequence typing.
Results: Overall, blaCTX-M was the dominant ESBL-encoding gene found in 95.75% of the isolates, followed by blaTEM (60.85%) and blaSHV (2.40%). While blaCTX-M-55 was the most common blaESBL subgroup found in this study. Whole genome sequencing showed a total of 15 different antimicrobial resistance genes other than blaESBL, including sul, qnr, aph(3ʹ)-Ia, among the selected 42 ESBL-E. coli isolates. Over half of the ESBL-E. coli (56.60%) carried blaCTX-M co-existing with blaTEM. The most common sequence types (STs) identified from human isolates were ST131, ST101, and ST70 while those isolated from swine were ST10, ST48, and ST131. ST131 strains carrying blaCTX-M were the major isolated ESBL-E. coli strains, supporting a previous study that considered this strain truly pathogenic. Noticeably, 66.51% of ESBL-E. coli strains shared 19 identical STs, including a host-restricted ST131 between humans and swine, suggesting that transmission between these two hosts might be possible.
Conclusion: Proof of a direct transfer of ESBL-E. coli from animals to humans, or vice versa, is required for further elucidation. The ESBL-E. coli isolated from both types of healthy hosts may serve as a reservoir for community-acquired antimicrobial resistance.
Keywords: ESBL-E. coli, blaCTX-M, blaTEM, and blaSHV, MLST, genetic relatedness
Introduction
Antimicrobial resistant bacteria are a global problem, and there is a need for urgent surveillance and appropriate preventive strategies. According to the World Health Organization (WHO), the spread of antimicrobial resistant bacteria has increased in all regions of the world and become one of the major causes of death worldwide. Resistant bacteria lead to difficulty in treatment and infection control and may affect the treatment of other diseases.1 The significant causes of the spread of antimicrobial resistance (AMR) in the community have been attributed to the overuse of antibiotics in households and in livestock, food-chain transmission, contamination of water environments or healthy faecal carriers.2–5 All these different sources are defined as reservoirs that facilitate the silent spread of and colonization by antimicrobial resistant bacteria within different communities, even in individuals with no history of hospitalization.6 These colonized bacteria probably present as normal flora; these bacteria may serve as a source of endogenous infections that occur after medical treatment or prolonged immune suppression.
Many recent works have reported the emergence of Escherichia coli resistant to beta-lactam antibiotics expressing extended-spectrum beta-lactamase (ESBL) enzymes. The extended-spectrum beta-lactamase-producing E. coli (ESBL-E. coli) are one of the leading pathogens that appear in both community and healthcare settings worldwide and are often resistant to other antimicrobial agents, including aminoglycosides, fluoroquinolones, and sulfonamides.7 Infection caused by ESBL-E. coli can be associated with high mortality due to multi-resistance, weakening the efficacy of therapies. ESBL-E. coli prevalence has dramatically risen during the last decade. However, the majority of ESBL-E. coli are still reported from human clinical isolates. The occurrence of ESBL-E. coli in animals is also increasing in some countries.7–9 A significant transmission of ESBL-E. coli along the pig production chain has occurred through pig compartments and abattoir waiting areas.9
To date, more than 350 different ESBL variants have been characterized and classified into nine distinct families based on their amino acid sequence comparisons.10 ESBL is found in a wide range of gram-negative bacteria, but the great majority hosts belong to the family Enterobacteriaceae, including E. coli, Klebsiella spp., Enterobacter spp., and Salmonella spp. The four, most common ESBLs, namely, CTX-M, TEM, SHV, and OXA, are found among Enterobacteriaceae species.8 Previous studies performed throughout diverse regions in the world showed variable epidemiology of these ESBLs. While TEM predominated in China, SHV was the leading group of ESBLs in Canada. However, reports from South America, Spain, New York, the United Kingdom, and several parts of India revealed CTX-M as the predominant ESBL.10 The CTX-M types of β-lactamases are the most prevalent family of ESBLs in E. coli, in which the CTX-M-15 ESBL is widely distributed across the world and often disseminated with the ST131 E. coli uropathogenic clone.11
This study aimed to connect data of ESBL-E. coli transmission among healthy humans in the community and swine among the livestock of Amphor Mueang, Lamphun Province, Thailand. Multiplex-PCR and whole genome sequencing (WGS) were performed to characterize the common ESBL-encoding genes of the isolates, while the genetic relatedness was investigated by multilocus sequence typing (MLST).
Materials and methods
Bacterial samples used in this study
Nonduplicated rectal swabs of 223 healthy people (living in the local community) and 186 healthy swine (living among the livestock) in Amphor Mueang, Lamphun Province, Thailand were collected between October and November of 2013. Each rectal swab was inoculated onto MacConkey agar (Oxoid, Baskingstoke, England) supplemented with ceftriaxone (Oxoid, Baskingstoke, England) (4 mg/L) in order to screen for ESBL-E. coli strains. Samples were incubated at 35°C for 24 hrs. All isolated colonies were identified by conventional biochemical tests. The E. coli isolates were stored at −80°C until use. Written informed consent was obtained from all participating people, whose rectal swabs were collected. The study was ethically approved by the Human Research Protection Unit, Faculty of Medicine Siriraj Hospital, Mahidol University (exemption number 0517.071/EC).
Antimicrobial susceptibility testing and ESBL detection
Antimicrobial susceptibility was performed using the Kirby-Bauer disk diffusion method according to the recommendations of the Clinical Laboratory Standards Institute (CLSI) 2016 guidelines.12 Antimicrobial agents (Oxoid, Baskingstoke, England) used in this study included ampicillin (10 μg), amoxicillin/clavulanic acid (20/10 μg), ceftriaxone (30 μg), cefotaxime (30 μg), ceftazidime (30 μg), cefoxitin (30 μg), imipenem (10 μg), ertapenem (10 μg), meropenem (10 μg), ciprofloxacin (5 μg), gentamicin (10 μg), amikacin (30 μg), and trimethoprim/sulfamethoxazole (1.25/23.75 μg). Phenotypic confirmation tests for ESBL production were conducted with a combination disk method, using ceftazidime (30 μg) versus ceftazidime/clavulanic acid (30/10 μg) and cefotaxime (30 μg) versus cefotaxime/clavulanic acid (30/10 μg) placed on Mueller-Hinton agar (Oxoid, Baskingstoke, England) and incubated at 35°C for 24 hrs. ESBL production was indicated if there was a difference in the inhibition zone of 5 mm between cephalosporin/beta-lactamase inhibitor disks compared with cephalosporin alone (12). E. coli ATCC25922 was used as a control strain.
Genomic DNA extraction
ESBL-E. coli isolates were grown on Tryptic soy agar at 35°C for 24 hrs. A single colony of each ESBL-E. coli was regrown in Tryptic soy broth at 35°C for 24 hrs. Two hundred microliters of each cell suspension was diluted in 800 µL of sterile distilled water and boiled at 95°C for 10 min followed by a 5-min centrifugation at 12,000 rpm. The supernatant was used as DNA template for PCR amplification.
Detection of ESBL genotypes by multiplex-polymerase chain reaction (multiplex-pcr)
All the 212 ESBL-E. coli isolates (111 isolates from humans and 101 isolates from swine) were examined for the presence of ESBL encoding genes, including blaCTX-M, blaTEM, and blaSHV genes, by multiplex-PCR. A uspA gene, which is specific to E. coli species, served as an internal control. A list of primers used for the multiplex-PCR is shown in Table 1. The multiplex-PCR amplification was performed in a total volume of 25 µL, containing 60 ng of template DNA, 10 pM each forward and reverse primer, 20 mM deoxynucleotide triphosphates (dNTPs), 1.5 mM magnesium chloride and 0.5 U Taq polymerase (Qiagen, Valencia, CA, USA) in 1x PCR standard buffer. The multiplex-PCR conditions were as follows: initial denaturation at 95°C for 15 min; 30 cycles of 94°C for 30 sec, 60°C for 40 sec and 72°C for 2 min; followed by a final extension step at 72°C for 10 min. In this study, the E. coli EC137 strain harbouring blaCTX-M, blaTEM, and blaSHV (as confirmed by WGS from a previous study and kindly provided by Associate Professor Dr. Methee Chayakulkeeree, Faculty of Medicine Siriraj Hospital, Mahidol University, Bangkok, Thailand) was used as a positive control.13 E. coli ATCC25922 was used as a negative control. Multiplex-PCR products were analyzed by 1.0% gel electrophoresis containing RedSafe™ (Scientifix, NSW, Australia) under UV light. Randomly selected multiplex-PCR products were confirmed by Sanger sequencing. The sequence reads were compared to gene sequences as described in the NCBI database (Blast search http://blast.ncbi.nlm.nih.gov/Blast.cgi).
 |
Table 1 Primers used for multiplex-PCR amplification |
Whole genome sequencing analysis
Randomly selected genomic DNA of 42 confirmed ESBL-E coli isolates (27 isolates from humans and 15 isolates from swine) were whole genome sequenced and analyzed by the Pathosystems Resource Integration Center (PATRIC), Chicago, IL, USA. In brief, sequencing was carried out using a NextSeq 500 platform. The raw data was assembled using SPAdes (version 3.9). Whole genome sequencing data were used for genotypic characterization of blaESBL subgroups and other antimicrobial resistance genes.
Multilocus sequence typing and phylogenetic analysis
To investigate the phylogeny of clonal spread versus ESBL resistance genes and bacterial hosts, a sequence type (ST) analysis of 212 ESBL-E. coli isolates was performed according to the protocols described by Tartof et al.16 Seven housekeeping genes, including adk, fumC, gyrB, icd, mdh, purA, and recA, of each isolate were amplified and sequenced. For amplification, PCR reactions of individual genes were separately performed in a total volume of 100 µL, containing 50 ng of template DNA, 20 pM each primer, 200 µM dNTPs, and 5 U Taq polymerase in 10 µl of 10x PCR standard buffer. The PCR conditions were as follows: initial denaturation at 95°C for 2 min; 30 cycles of 95°C for 1 min, 54°C (adk, fumC, icd, and purA) or 58°C (recA) or 60°C (gyrB and mdh) for 1 min and 72°C for 2 min; followed by a final extension step at 72°C for 5 min. The PCR products were sent to Bioneer Sequencing Service Co., Ltd., South Korea, to perform a sequence analysis by conventional Sanger method, using an ABI 3730XL DNA Analyzer (Applied Biosystems, Foster City, CA, USA). The sequence data were imported to the E. coli MLST database, http://mlst.warwick.ac.uk/mlst/dbs/Ecoli, for MLST type determination.17 Phylogenetic analysis was performed by BioNumerics version 7.6 software (Applied Maths, Sint-Martens-Latem, Belgium).
Statistical analysis
To evaluate the correlation between antimicrobial resistant phenotypes or genotypes and bacterial host types, a Fisher’s exact test (IBM SPSS 17.0 statistical package, Chicago, IL) was used. p-values ≤0.05 were considered statistically significant.
Results
Antimicrobial susceptibility testing
There were 115 and 103 E. coli strains isolated from 223 people and 186 swine, respectively, in this study. Antibiotic susceptibility testing revealed that all the isolates (100%) from both hosts were resistant to ampicillin, ceftriaxone, and cefotaxime. On the other hand, none of these isolates showed resistance against carbapenems, including imipenem, ertapenem, and meropenem. The E. coli isolated from the swine, compared to those isolated from humans, were significantly more resistant to amoxicillin/clavulanic acid (41.74% vs 14.78%), ceftazidime (93.20% vs 51.30%), and gentamicin (82.52% vs 35.65%) (p-value <0.05). However, ciprofloxacin resistance was significantly more common in human isolates (54.78%) rather than in swine isolates (26.21%) (p-value <0.05). There was no significant difference in resistance to cefoxitin, amikacin, and trimethoprim/sulfamethoxazole between E. coli strains isolated from both hosts (p-value >0.05). Interestingly, multidrug-resistant E. coli were predominately found in both humans (84.35%) and swine (85.44%). Resistance percentages against antimicrobial agents of E. coli in this study are presented in Table 2. Among 212 of the 218 isolated E. coli, ESBL-producing strains were detected by a combination disk method. These included 111 out of the 115 (96.52%) human isolates and 101 out of the 103 (98.06%) swine isolates.
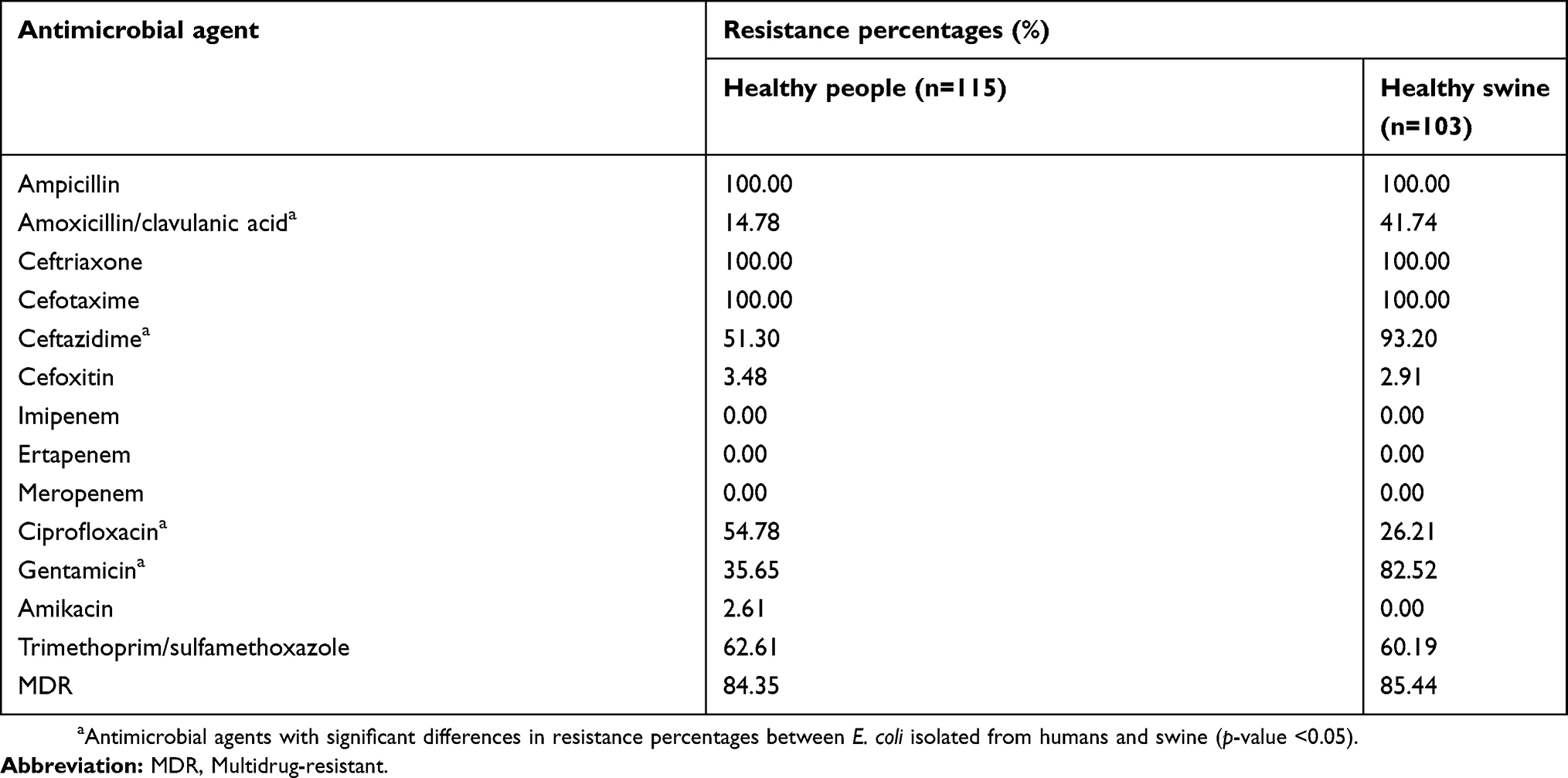 |
Table 2 An overview of the resistance percentages of E. coli isolated from healthy people (n=115) and swine (n=103) against antimicrobial agents |
ESBL gene characterization
Multiplex-PCR amplification assay was designed for blaCTX-M, blaTEM and blaSHV detection in ESBL-E. coli. The amplified products were separated by 1% agarose gel electrophoresis. Multiplex-PCR generated bands for the uspA gene used as an internal control (884 bp) and blaCTX-M (593 bp), blaTEM (455 bp), and blaSHV (747 bp) (Figure 1). All the 212 ESBL-E. coli isolates harboured uspA gene and at least one bla gene. The blaCTX-M gene was the most predominantly observed in ESBL-E. coli isolated from both hosts. There were 95.75% (203 isolates) harbouring blaCTX-M, followed by 62.74% (133 isolates) and 2.36% (5 isolates) harbouring blaTEM and blaSHV, respectively. The majority of the ESBL-E. coli (56.60%; 120 isolates) carried blaCTX-M co-existing with blaTEM, while 36.79% (78 isolates), 4.25% (9 isolates), 1.89% (4 isolates), and 0.47% (1 isolates) carried blaCTX-M alone, blaTEM alone, blaCTX-M+blaTEM+blaSHV and blaCTX-M+blaSHV, respectively. Interestingly, the presence of blaSHV alone was not observed among the strains. Statistical analysis revealed that there was no significant differences (p-value >0.05) in the distribution of ESBL genes in E. coli isolated from humans and swine. Table 3 shows a summary of the ESBL-E. coli gene distribution in humans and swine in this study.
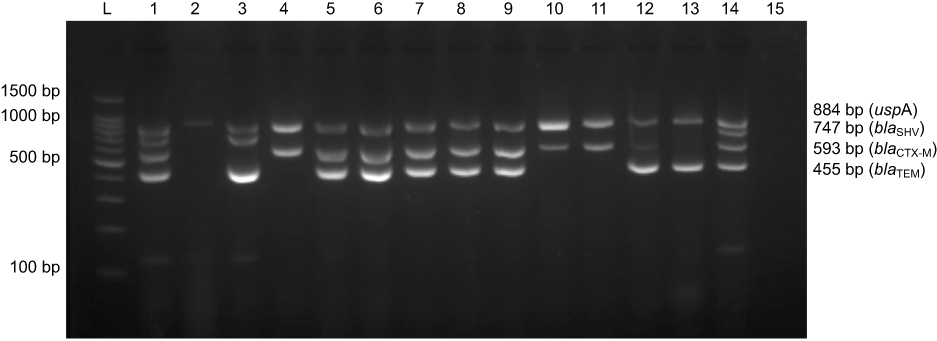 |
Figure 1 Multiplex-PCR banding patterns of blaCTX-M (593 bp), blaTEM (455 bp), blaSHV (747 bp) and uspA (884 bp) genes of E. coli generated by gel electrophoresis. L: standard DNA ladder 100 bp. Lane 1: a positive control (E. coli EC137 strain) generated 4 bands of blaCTX-M, blaTEM, blaSHV and uspA genes. Lane 2: E. coli ATCC25922 generated 1 band for the uspA gene. Lanes 3–14: E. coli isolated from the samples. Lane 15: a negative control. |
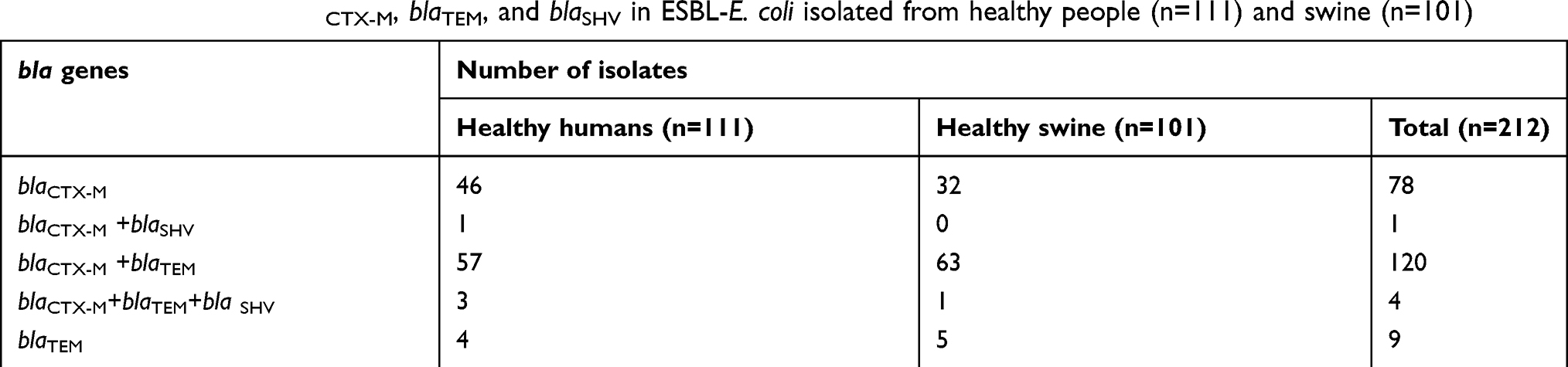 |
Table 3 Distribution of blaCTX-M, blaTEM, and blaSHV in ESBL-E. coli isolated from healthy people (n=111) and swine (n=101) |
WGS revealed that blaCTX-M-55 (56.10%; 23 isolates) was the most prevalent blaCTX-M in those selected 42 ESBL-E. coli isolates, followed by blaCTX-M-14 (43.90%; 18 isolates). However, blaCTX-M-14 was significantly predominate in isolates from humans (13 isolates) rather than those from swine (4 isolates), while there was no significant difference in blaCTX-M-55 found in ESBL-E. coli isolated from both hosts. All the blaTEM found among the 42 ESBL-E. coli isolates were only blaTEM-1B (100%; 5 isolates) (Table 4).
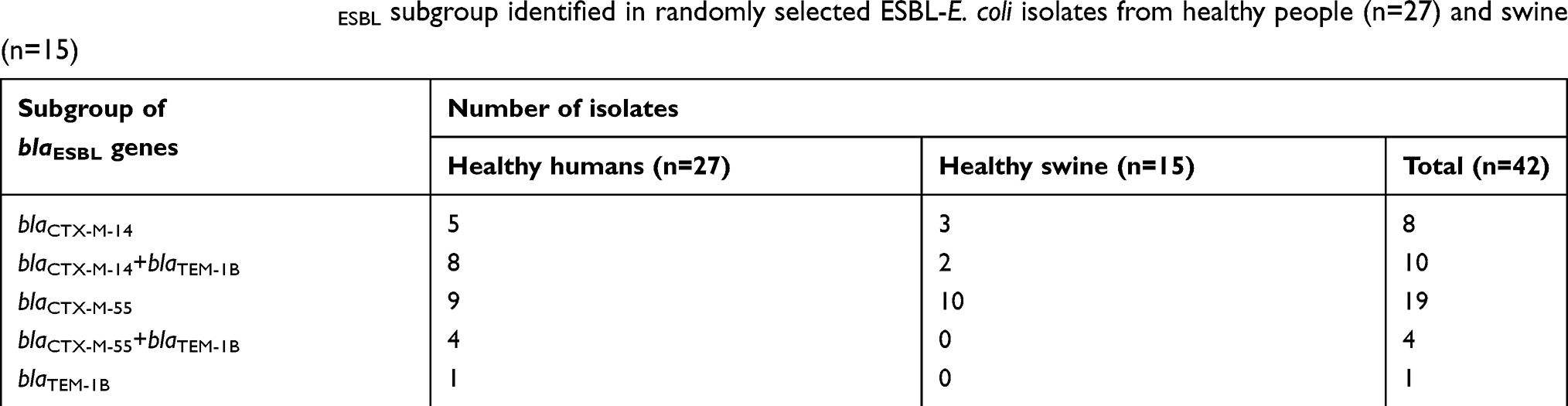 |
Table 4 Summary of the blaESBL subgroup identified in randomly selected ESBL-E. coli isolates from healthy people (n=27) and swine (n=15) |
Detection of additional antimicrobial resistance genes
Apart from the blaESBL subgroup, WGS revealed 15 additional different antimicrobial resistance genes among the 42 ESBL-E. coli isolates. These included sul1, sul2 and sul3 (encoding resistance to sulfonamides), aadA22 and aph(3ʹ)-Ia (encoding resistance to aminoglycosides), strA, strB, aadA2 and aadA5 (encoding resistance to streptomycin), qnrS1 (encoding resistance to fluoroquinolones), tetA, tetB and tetD (encoding resistance to tetracyclines), arr2 (encoding resistance to rifampicin), and mefB (encoding resistance to macrolides) (Table 5). qnrS1 was the dominant antimicrobial resistance gene other than blaESBL found in 61.90% (26 isolates) of 42 ESBL-E. coli isolates, followed by sul subgroup (45.24%, 19 isolates). Although, there was no correlation between the significant pattern of those resistance genes and the ST of 42 ESBL-E. coli isolates. Several whole genome sequenced isolates carried multiple antimicrobial resistance genes, as summarized in Table 5.
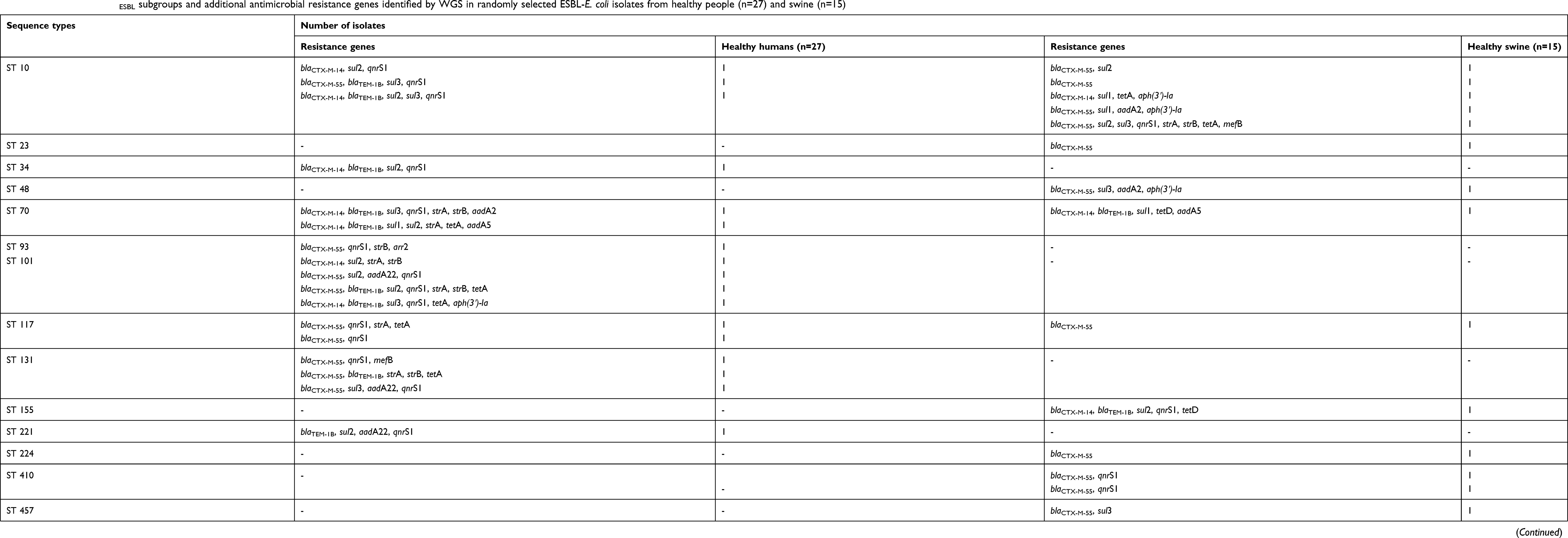 |
Table 5 Distribution of blaESBL subgroups and additional antimicrobial resistance genes identified by WGS in randomly selected ESBL-E. coli isolates from healthy people (n=27) and swine (n=15) |
Multilocus sequence typing analysis
A total of 7 housekeeping genes, including adk, fumC, gyrB, icd, mdh, purA, and recA were amplified and sequenced for each ESBL-E. coli isolate. Sixty-six unique sequence types (STs) were identified among the 212 ESBL-E. coli isolates. Among these, only 25 STs (n=132) clustered into 16 clonal complexes (CCs) (Table 6). The five, most common CCs were CC10 (n=48), CC131 (n=24), CC23 (n=14), CC155 (n=14), and CC101 (n=9). In total, ST131 (n=24, 11.32%) was the most predominant ST, followed by ST10 (n=18, 8.49%), ST48 (n=17, 8.02%), ST34 (n=9, 4.25%), and ST101 (n=9, 4.25%). An MLST dendrogram of these isolates was constructed with BioNumerics software. Figure 2 demonstrates the ESBL-E. coli STs isolated from human and swine sample sources. In the MLST dendrogram, each circle represents an ST, and the size of a circle is proportional to the number of ESBL-E. coli isolates belonging to this ST.
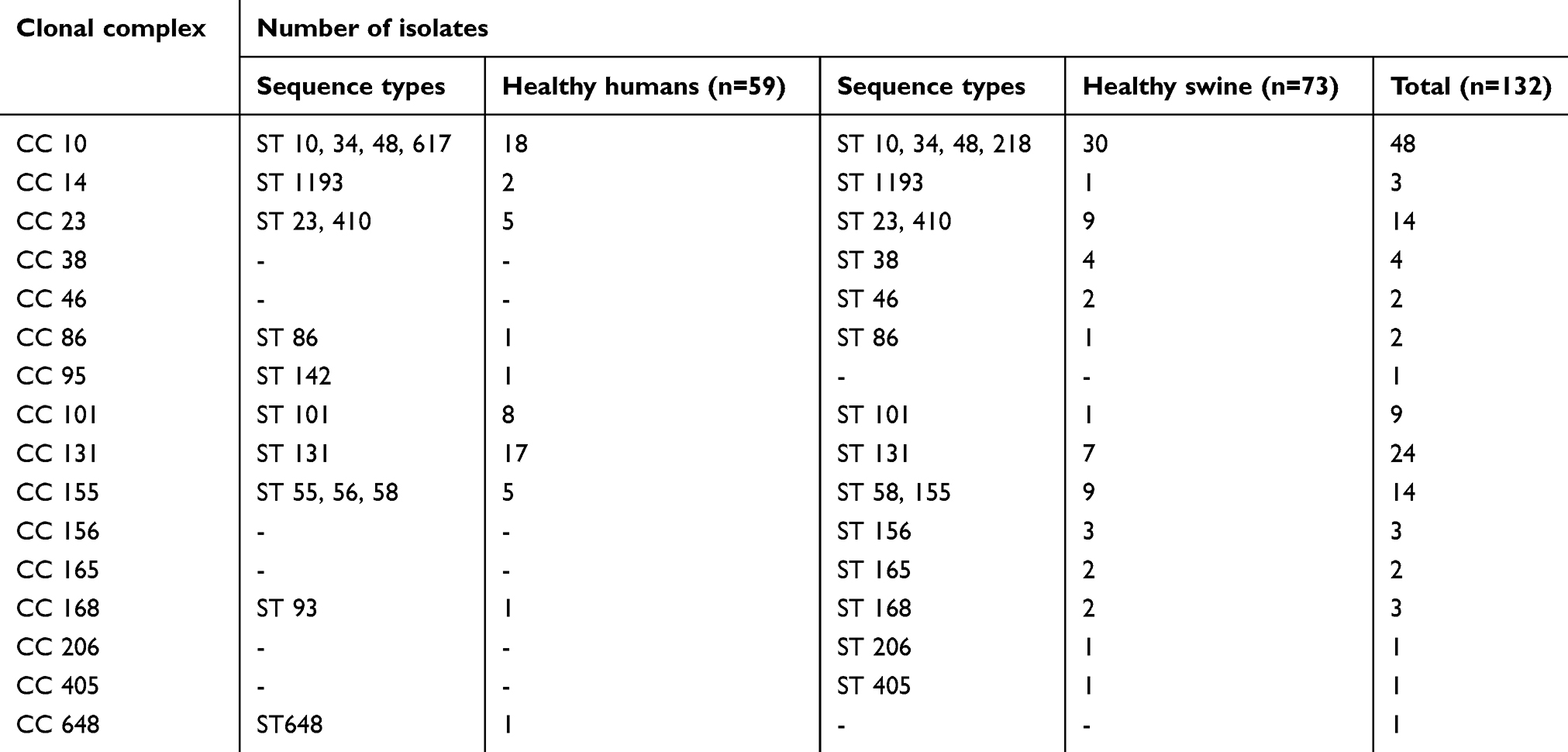 |
Table 6 Distribution of the clonal complexes and their sequence type members detected in ESBL-E.coli isolates from humans and swine |
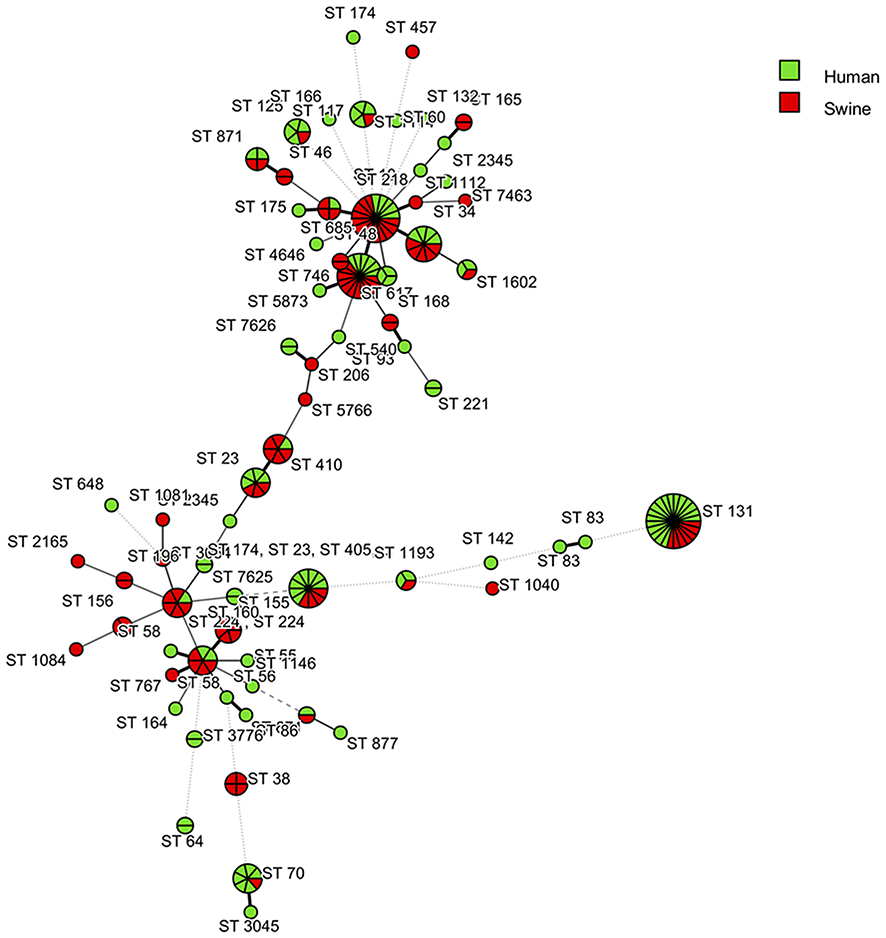 |
Figure 2 MLST dendrogram created based on STs of the 212 ESBL-E.coli isolates from human and swine coloured according to the strain’s origin. Each circle represents a given allelic profile and is named according to the MLST sequence type. Each circle corresponds to an ST, and the size of the circle is related to the number of isolates in this study. (BioNumerics version 7.6 software, Applied Maths, Sint-Martens-Latem, Belgium).Abbreviations: MLST, Multilocus sequence typing; ST, Sequence type; ESBL, Extended-spectrum beta-lactamase. |
Interestingly, 141 out of 212 (66.51%) ESBL-E. coli shared identical STs between human and swine isolates. All these isolates belonged to 19 shared STs, for example, ST10 (CC10), ST23 (CC23), ST34 (CC10), ST48 (CC10), ST58 (CC155), ST101 (CC101), and ST131 (CC131). In the human source, a total of 47 STs were identified, and ST131 (n=17) was the most prevalent ST, followed by ST101 (n=8), ST70 (n=6), ST48 (n=6), and ST10 (n=5). There were 28 out of 47 STs that were identified in only human isolates, including ST55 and ST56 (CC155), ST93 (CC168), ST142 (CC95), and ST648 (CC648). In the swine source, a total of 38 STs were identified, and ST10 (n=13) was the most prevalent, followed by ST48 (n=11) and ST131 (n=7). There were 19 of 38 STs that were identified in only swine isolates, including ST218 (CC10), ST155 (CC155), ST168 (CC168), and ST405 (CC405).
A relationship between the MLST-based dendrogram and the distribution of ESBL genes in E. coli is illustrated in Figure 3. Almost all human and swine isolates carried blaCTX-M, either alone or in combination with blaTEM and/or blaSHV. Only 4 human isolates, including ST221 (n=2), ST1193 (n=1), and ST3376 (n=1), and 5 swine isolates, including ST10 (n=2), ST48 (n=1), ST58 (n=1), and ST224 (n=1), had only blaTEM. Among the three most common shared STs found in both human and swine isolates, ST131 (n=24), ST10 (n=18), and ST48 (n=17), there were noteworthy patterns of ESBL encoding genes. Among the ST131 isolates, 15 isolates harboured blaCTX-M+blaTEM (11 human and 4 swine isolates), whereas the remaining 9 isolates harboured blaCTX-M (6 human and 3 swine isolates).
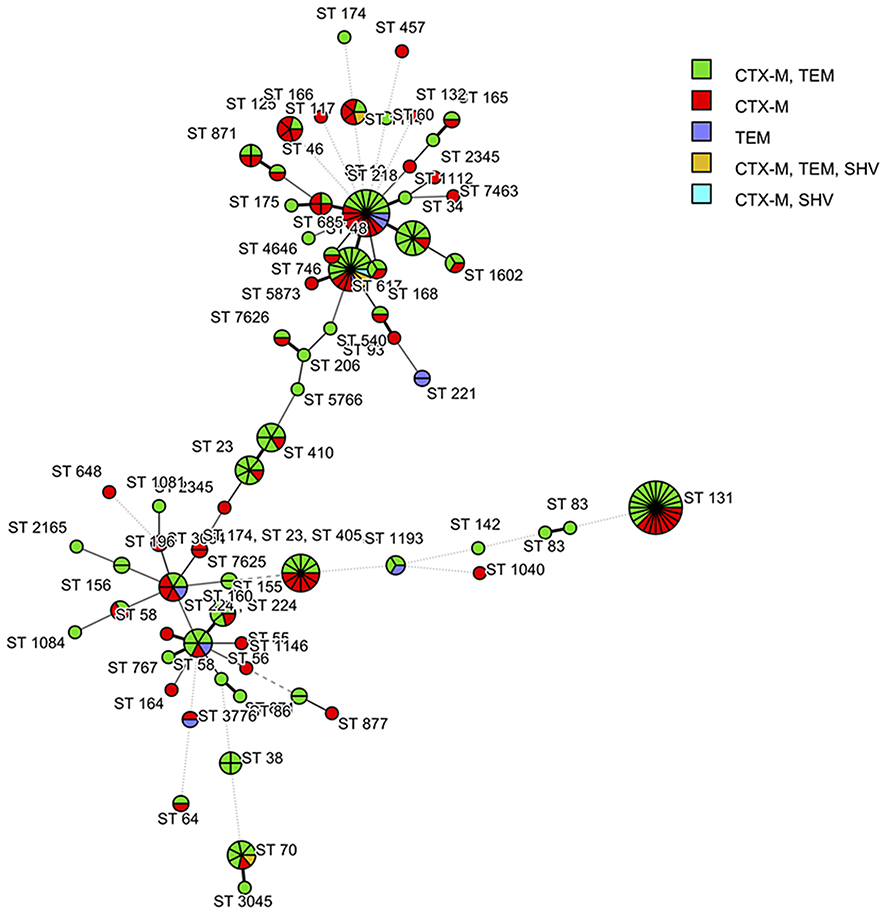 |
Figure 3 The relationship between the MLST-based dendrogram and the distribution of the ESBL encoding genes of the 212 E. coli strains isolated from human and swine coloured according to ESBL genes. (BioNumerics version 7.6 software, Applied Maths, Sint-Martens-Latem, Belgium).Abbreviations: MLST, Multilocus sequence typing; ST, Sequence type; ESBL, Extended-spectrum beta-lactamase. |
Among ST10 isolates, there were 8 isolates harbouring blaCTX-M+blaTEM (3 human and 5 swine isolates), whereas the other 8 isolates harboured blaCTX-M (2 human and 6 swine isolates). The other two swine isolates harboured only blaTEM. A variety of ESBL encoding genes was found among the ST48 isolates. There were 10 isolates harbouring blaCTX-M+blaTEM (2 human and 8 swine isolates), 3 isolates harbouring blaCTX-M (2 human and 1 swine isolates), 2 isolates harbouring blaCTX-M+blaTEM+blaSHV (1 human and 1 swine isolate), 1 human isolate harbouring blaCTX-M+blaSHV, and one swine isolate harbouring only blaTEM.
Discussion
ESBL is one of the beta-lactamase enzymes and is considered the essential mechanism of resistance to b-lactam antibiotics among gram-negative bacilli. The ESBL enzymes are usually acquired by horizontal gene transfer, which rapidly supports the spreading of resistant bacterial strains.18 Several publications have revealed data of spreading of ESBL-E. coli in communities worldwide.7–9 In Thailand, ESBL-E. coli was previously isolated from the food of healthy adults and animal farm workers, as well as farm animals and environmental samples.19 In our study, we assessed the spreading of ESBL-E. coli among humans and swine from a community by MLST. ESBL-E. coli was isolated from healthy humans living in Amphor Mueang, Lamphun Province and from healthy swine living in livestock in the same community as the sampled humans.
In this study, the majority of the isolated ESBL-E. coli comprised multidrug-resistant strains; these bacteria may serve as an unintentional source of difficult-to-treat endogenous infections due to their multidrug-resistant phenotypes that limit the options for antibiotic therapy. The risk factors involved with the faecal carriage of AMR include the use of antibiotics without prescriptions such as beta-lactams and fluoroquinolones use for lower urinary tract infections.20 The use of antibiotics in food animals contributes to selective pressures that promote AMR in livestock.11 Our data show that, apart from beta-lactam antibiotics, ESBL-E. coli was further resistant to fluoroquinolones, aminoglycosides, and trimethoprim/sulfamethoxazole. WGS revealed several transferable antimicrobial resistance genes other than blaESBL co-existing in ESBL-E. coli. Many isolates carried three or more resistance gene categories, reflecting the multidrug-resistant phenotype of the isolates. Although, not all isolates that were antimicrobial susceptibility tested were whole genome sequenced in this study. A mechanism of plasmid-mediated quinolone resistance (PMQR) includes fluoroquinolone target protection by the Qnr protein (encoded by qnr) and aminoglycoside-modifying enzyme production (encoded by aac-6ʹ-Ib-cr) that induce resistance against fluoroquinolones and aminoglycosides simultaneously. Interestingly, the PMQRs have been previously shown to co-exist on plasmids carrying blaCTX-M.21 This may explain the association of fluoroquinolone and/or aminoglycoside resistance that was found among the ESBL-E. coli strains in our study. sul1, sul2, and sul3 are known as plasmid-encoded sulfonamide resistance genes that induce resistance against sulfonamides. A coexistence of ESBL genes with sul genes in ESBL-E. coli has previously been reported.22
In the last two decades, the majority of the ESBL strains identified in human clinical isolates were SHV or TEM types, until 10 years ago when CTX-M became the most widespread type of ESBL.11 Our study showed that blaCTX-M was the most frequently detected (95.75%) ESBL encoding gene either alone or in combination with other bla genes, followed by blaTEM (60.85%) and blaSHV (2.40%). Both groups of human and swine isolates had no significant differences in the distribution of these bla genes. Similar to our findings, previous studies showed that the blaCTX-M family had the highest prevalence among those typical E. coli ESBL encoding genes, followed by blaTEM and blaSHV either from clinical, animal, or environmental isolates.23–25 The prevalence of blaCTX-M-15 has risen over time worldwide and is dominant in most countries. Exceptions are China, Japan, Spain, and South-East Asia, where blaCTX-M-14 genes are dominant.11 Here, blaCTX-M-55 and blaCTX-M-14 were the most prevalent identified blaCTX-M genes. However, only 42 randomly selected ESBL-E. coli samples out of 212 samples were investigated for a subgroup of blaCTX-M and may not be a representative of the blaCTX-M distribution in the country. In Thailand, blaCTX-M-55 has been previously demonstrated to be the most common CTX-M in clinical isolates, farm waste and canals, followed by blaCTX-M-14.25
MLST presents data on the genetic relationship between the STs of ESBL-E. coli strains and host origins. The dominant ST identified in our study was ST131, which was the first and third most common ST identified in healthy humans and swine, respectively. In general, ST131 E. coli is the most common ST obtained from human clinical isolates globally. Previous studies revealed that ST131 was predominant among ESBL-E. coli, ranging from 28 to 38%, isolated from human clinical samples in Sweden, Germany and Thailand.25–27 However, ST131 seems to be host restricted, and there have been very few reports on ST131 from either animals (healthy or sick), foods, or environments.25,28 Our study provides another line of evidence showing a possibility that ST131 could disseminate to various animal species and ecosystems.
ST131 E. coli has a broad spectrum of infections both in community and hospital settings. ST131 E. coli also has a large number of virulence-associated genes, which make these bacteria truly pathogenic.28 ST131 isolates are commonly reported to produce ESBL, mostly by CTX-M-15, and almost all isolates are resistant to fluoroquinolones.28 Our findings showed that all ST131 harboured blaCTX-M either alone or in combination with other bla genes. However, only two strains of ST131 were randomly identified for a subgroup by WGS, and CTX-M-55 was found (data not shown). Thus, in our study, it is too early to conclude which CTX-M subgroup is mostly produced by the ST131 ESBL-E. coli. In addition, most of the ST131 isolates were resistant to ciprofloxacin and gentamicin. A global expansion of the clone ST131 carrying blaCTX-M is considered one major reason for the rise in ESBL-E. coli. Increased virulence of ST131 strains has been previously discussed, but the proof of this virulence is unclear.26 Lavigne et al proposed a key explanation for the success in the spread of ST131 E. coli using a zebrafish model; CTX-M-producing ST131 and non-CTX-M-producing ST131 had decreased virulence but improved persistence during infection compared with non-ST131 E. coli.29
The next most common ST of ESBL-E. coli identified in this study was ST10, which was obtained from both human and swine isolates. In contrast to ST131, ST10 is commonly isolated from a variety of sources.20 Our finding is supported by a previous study that showed that ST10 ESBL-E. coli is globally disseminated in humans and animals.20,30 ST48 ESBL-E. coli was the third most common ST obtained in our study. This clone was previously isolated from both hospitalized and non-hospitalized patients as well as from poultry.31,32 Nearly all ST10 and ST48 harboured blaCTX-M either alone or in combination with other bla genes; the two exceptions to the former scenario and the one exception to the latter scenario all carried only blaTEM. Unlike ST131, in which blaCTX-M was strictly associated, both ST10 and ST48 seemed to be less restricted in their association with blaCTX-M.
In a study from Denmark, highly similar clones of ESBL-E. coli were observed in swine livestock and farm workers, suggesting that swine may play a significant role as vectors in the transfer ESBL-E. coli via close contact.33 Nonetheless, evidence showing a direct transfer of ESBL-E. coli from animals to humans is still inadequate. The presence of shared STs, particularly the human-restricted ST131 and the non-host-restricted ST10 and ST 48 isolates, between people in the community and the swine on farms in our study seemed to be transferred by means other than close contact. Commensal ESBL-E. coli in the gut of animals and humans may be an important source of bacteria causing opportunistic infections or act as a resistant gene reservoir, serving as a source of bacterial spread.18 However, it would be interesting to further investigate the ESBL-E. coli isolates that shared identical STs by more discriminatory typing methods, such as WGS, to prove the relatedness among isolates and characterize the mechanisms of transfer between humans and animals.
Conclusion
Proof of a direct transfer of ESBL-E. coli from animals to human is required for further elucidation. However, this study showed several shared identical STs of ESBL-E. coli from the healthy humans in a community and the swine in the community livestock, suggesting epidemiological links or that transmission between these two hosts is possible. ESBL-E. coli isolates from both hosts indicate that healthy humans and livestock animals can serve as a reservoir for AMR. Minimizing the transmission between hosts is essential to control the spread of ESBL- E. coli in community settings. Additional monitoring of the epidemiology of ESBL-E. coli in humans and livestock, as well as the proof of possible transmission routes, are needed.
Acknowledgments
This work was supported by the 90th Anniversary Chulalongkorn University Fund and the Chulalongkorn University Ratchadaphiseksomphot Endowment Fund (CU-GR_62_07_37_01), and the Allied Health Sciences Fund (AHS-CU 60003).
Disclosure
The authors declare that they have no conflicts of interest related to the publication of this article.
References
1. World Health Organization [homepage on the Internet]. Geneva: Antimicrobial Resistance: Global Report on Surveillance; 2014. Available from: http://www.who.int/drugresistance/documents/surveillancereport/en/.
2. Angulo FJ, Nargund VN, Chiller TC. Evidence of an association between use of anti-microbial agents in food animals and antimicrobial resistance among bacteria isolated from humans and the human health consequences of such resistance. J Vet Med B Infect Dis Vet Public Health. 2004;51:374–379. doi:10.1111/j.1439-0450.2004.00789.x
3. Börjesson S, Ny S, Egervärn M, et al. Limited dissemination of extended-spectrum β-lactamase- and plasmid-encoded AmpC-producing Escherichia coli from food and farm animals, Sweden. Emerg Infect Dis. 2016;22(4):634–640. doi:10.3201/eid2204.151142
4. Korzeniewska E, Harnisz M. Extended-spectrum β-lactamase (ESBL)-positive Enterobacteriaceae in municipal sewage and their emission to the environment. J Environ Manage. 2013;128:904–911. doi:10.1016/j.jenvman.2013.06.051
5. Fernández-Reyes M, Vicente D, Gomariz M, et al. High rate of fecal carriage of extended-spectrum β-lactamase-producing Escherichia coli in healthy children in Gipuzkoa, Northern Spain. Antimicrob Agents Chemother. 2014;58(3):1822–1824. doi:10.1128/AAC.02045-12
6. Jakobsen L, Kurbasic A, Skjøt-Rasmussen L, et al. Escherichia coli isolates from broiler chicken meat, broiler chickens, pork, and pigs share phylogroups and antimicrobial resistance with community-dwelling humans and patients with urinary tract infection. Foodborne Pathog Dis. 2010;7(5):537–547. doi:10.1089/fpd.2009.0409
7. Wu G, Day MJ, Mafura MT, et al. Comparative analysis of ESBL-positive Escherichia coli isolates from animals and humans from the UK, the Netherlands and Germany. PLoS One. 2013;8(9):e75392. doi:10.1371/journal.pone.0075392
8. Guenther S, Ewers C, Wieler LH. Extended-spectrum β-Lactamases producing E. coli in wildlife, yet another form of environmental pollution? Front Microbiol. 2011;2:246. doi:10.3389/fmicb.2011.00246
9. Schmithausen RM, Schulze-Geisthoevel SV, Stemmer F, El-Jade M, Reif M, Hack S. Analysis of transmission of MRSA and ESBL-E among pigs and farm personnel. PLoS One. 2015;10(9):e0138173. doi:10.1371/journal.pone.0138173
10. Yazdi M, Nazemi A, Mirinargasi M, Jafarpour M, Sharifi SH. Genotypic versus phenotypic methods to detect extended-spectrum β-lactamases (ESBLs) in uropathogenic Escherichia coli. Ann Biol Res. 2012;3:2454–2458.
11. Bevan ER, Jones AM, Hawkey PM. Global epidemiology of CTX-M β-lactamases: temporal and geographical shifts in genotype. J Antimicrob Chemother. 2017;72(8):2145–2155. doi:10.1093/jac/dkx146
12. CLSI. Performance Standards for Antimicrobial Susceptibility Testing; Approved Standard 26th Edition. Document M100S. Pennsylvania: Clinical and Laboratory Standard Institute (CLSI); 2016.
13. Tangkoskul T, Tiengrim S, Onsomang S, et al. Molecular characterization of extended-spectrum β-lactamases and its correlation with clinical laboratory standards institute interpretive criteria for disk diffusion susceptibility testing in Enterobacteriaceae isolates in Thailand. Southeast Asian J Trop Med Public Health. 2012;43(6):1461–1469.
14. Chen J, Griffiths MW. PCR differentiation of Escherichia coli from other gram-negative bacteria using primers derived from the nucleotide sequences flanking the gene encoding the universal stress protein. Lett Appl Microbiol. 1998;27(6):369–371. doi:10.1046/j.1472-765X.1998.00445.x
15. Monstein HJ, Ostholm-Balkhed A, Nilsson MV, Nilsson M, Dornbusch K, Nilsson LE. Multiplex-PCR amplification assay for the detection of blaSHV, blaTEM and blaCTX-M genes in Enterobacteriaceae. APMIS. 2007;115:1400–1408.
16. Tartof SY, Solberg OD, Manges AR, Riley LW. Analysis of a uropathogenic Escherichia coli clonal group by multilocus sequence typing. J Clin Microbiol. 2005;43(12):5860–5864. doi:10.1128/JCM.43.12.5860-5864.2005
17. Escherichia coli MLST Database [homepage on the Internet]. Coventry: University of Warwick Escherichia coli MLST Database; 2018. Available from: http://mlst.warwick.ac.uk/mlst/dbs/Ecoli.
18. Hawkey PM, Jones AM. The changing epidemiology of resistance. J Antimicrob Chemother. 2009;64(Suppl1):i3–10.
19. Boonyasiri A, Tangkoskul T, Seenama C, Saiyarin J, Tiengrim S, Thamlikitkul V. Prevalence of antibiotic resistant bacteria in healthy adults, foods, food animals, and the environment in selected areas in Thailand. Pathog Glob Health. 2014;108(5):235–245. doi:10.1179/2047773214Y.0000000148
20. Ewers C, Bethe A, Semmler T, Guenther S, Wieler LH. Extended-spectrum β-lactamase producing and AmpC-producing Escherichia coli from livestock and companion animals, and their putative impact on public health: a global perspective. Clin Microbiol Infect. 2012;18(7):646–655. doi:10.1111/j.1469-0691.2012.03850.x
21. Cattoir V, Nordmann P. Plasmid-mediated quinolone resistance in gram-negative bacterial species: an update. Curr Med Chem. 2009;16(8):1028–1046. doi:10.2174/092986709787581879
22. Arabi H, Pakzad I, Nasrollahi A, et al. Sulfonamide resistance genes (sul) M in extended-spectrum β-lactamase (ESBL) and non-ESBL producing Escherichia coli isolated from Iranian hospitals. Jundishapur J Microbiol. 2015;8(7):e19961. doi:10.5812/jjm.19961v2.
23. Bubpamala J, Khuntayaporn P, Thirapanmethee K, Montakantikul P, Santanirand P, Chomnawang MT. Phenotypic and genotypic characterizations of extended-spectrum β-lactamase-producing Escherichia coli in Thailand. Infect Drug Resist. 2018;11:2151–2157. doi:10.2147/IDR.S174506
24. Dahms C, Hübner NO, Kossow A, Mellmann A, Dittmann K, Kramer A. Occurrence of ESBL-producing Escherichia coli in livestock and farm workers in Mecklenburg-Western Pomerania, Germany. PLoS One. 2015;10(11):e0143326. doi:10.1371/journal.pone.0143326
25. Runcharoen C, Raven KE, Reuter S, et al. Whole genome sequencing of ESBL-producing Escherichia coli isolated from patients, farm waste and canals in Thailand. Genome Med. 2017;9(1):81. doi:10.1186/s13073-017-0471-8
26. Brolund A, Edquist PJ, Mäkitalo B, et al. Epidemiology of extended-spectrum β-lactamase-producing Escherichia coli in Sweden 2007–2011. Clin Microbiol Infect. 2014;20(6):344–352. doi:10.1111/1469-0691.12742
27. Pietsch M, Eller C, Wendt C, et al. Molecular characterization of extended-spectrum β-lactamase (ESBL)-producing Escherichia coli isolates from hospital and ambulatory patients in Germany. Vet Microbiol. 2017;200:130–137. doi:10.1016/j.vetmic.2015.11.028
28. Nicolas-Chanoine MH, Bertrand X, Madec JY. Escherichia coli ST131, an intriguing clonal group. Clin Microbiol Rev. 2014;27(3):543–574.
29. Lavigne JP, Vergunst AC, Goret L, et al. Virulence potential and genomic mapping of the worldwide clone Escherichia coli ST131. PLoS One. 2012;7(3):e34294. doi:10.1371/journal.pone.0034294
30. Day MJ, Rodríguez I, van Essen-Zandbergen A, et al. Diversity of STs, plasmids and ESBL genes among Escherichia coli from humans, animals and food in Germany, the Netherlands and the UK. J Antimicrob Chemother. 2016;71(5):1178–1182. doi:10.1093/jac/dkv485
31. Smet A, Martel A, Persoons D, et al. Characterization of extended-spectrum beta-lactamases produced by Escherichia coli isolated from hospitalized and nonhospitalized patients: emergence of CTX-M-15-producing strains causing urinary tract infections. Microb Drug Resist. 2010;16(2):129–134. doi:10.1089/mdr.2009.0132
32. Blaak H, Hamidjaja RA, van Hoek AH, de Heer L, de Roda Husman AM, Schets FM. Detection of extended-spectrum β-lactamase (ESBL)-producing Escherichia coli on flies at poultry farms. Appl Environ Microbiol. 2014;80(1):239–246. doi:10.1128/AEM.02616-13
33. Hammerum AM, Larsen J, Andersen VD, et al. Characterization of extended-spectrum β-lactamase (ESBL)-producing Escherichia coli obtained from Danish pigs, pig farmers and their families from farms with high or no consumption of third- or fourth-generation cephalosporins. J Antimicrob Chemother. 2014;69(10):2650–2657. doi:10.1093/jac/dku180
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.