Back to Journals » Infection and Drug Resistance » Volume 13
Molecular Investigations of Linezolid Resistance in Enterococci OptrA Variants from a Hospital in Shanghai
Authors Li P, Yang Y, Ding L, Xu X, Lin D
Received 29 February 2020
Accepted for publication 5 July 2020
Published 4 August 2020 Volume 2020:13 Pages 2711—2716
DOI https://doi.org/10.2147/IDR.S251490
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Suresh Antony
Pei Li, Yang Yang, Li Ding, Xiaogang Xu, Dongfang Lin
Institute of Antibiotics, Huashan Hospital, Fudan University, Shanghai, People’s Republic of China
Correspondence: Dongfang Lin Tel/ Fax +86-21-52888193
Email [email protected]
Purpose: The OptrA protein is a member of ATP-binding cassette (ABC) transporters, a transporter family which can confer resistance to oxazolidinone antibiotics by transferring plasmid. We aim to describe the distribution of optrA-harbored Enterococcus in Huashan hospital in 2017 and to address the effects of optrA mutations on the susceptibility of linezolid antibiotic drug.
Methods: Linezolid-resistance-related genes were tested for Enterococcus by polymerase chain reaction (PCR) and then sequenced for amino acid substitution site analysis. Broth microdilution and agar dilution test were applied to determine the minimal inhibitory concentration (MIC) of linezolid for Enterococcus containing optrA. Pulsed field gel electrophoresis (PFGE) and multilocus sequence typing (MLST) were used to evaluate the genotypes of optrA-positive isolates. To verify the functions of two main optrA variants, optrA over-expressing strains were constructed.
Results: Among 20 optrA-positive strains, only two were resistant to linezolid. No amino acid substitution existed in 23S rRNA V domain among Enterococcus faecalis. None had cfr, cfr(B) or cfr(C) genes. F101L and G4D/K/R or T150A were the main substitutions of ribosomal protein L4, L3, respectively. We found one Enterococcus faecium isolate co-contained optrA and poxtA and another E. faecalis isolate co-contained optrA and cfr(D), but they were not resistant to linezolid. Among 20 optrA-positive strains, ST-16 was the main type. Two main optrA variants KD (T112K, Y176D) and RDK (I104R, Y176D, E256K) slightly raised enterococci’s MIC of linezolid.
Conclusion: OptrA exists in linezolid non-resistant enterococci with diverse amino acid substitutions. The variants play different roles in changing the MIC of linezolid.
Keywords: linezolid, resistant, enterococci, optrA
Introduction
Enterococci are medically important opportunistic pathogen which are more common in hospital-acquired infections. Linezolid1 is the first clinically used oxazolidinone antibiotic for the treatment of multidrug-resistant (MDR) gram-positive infections, especially in vancomycin-resistant enterococci (VRE) and methicillin-resistant Staphylococcus aureus (MRSA).2 However, linezolid-resistant enterococci have been increasingly detected both in human and animals3 in recent years, which urgently demands to deepen our knowledge on its resistance mechanisms.
The resistance of linezolid in enterococci can be caused by point substitutions in 23S rRNA V domain, the main reason for linezolid resistance.4 The 23S rRNA methyltransferase coding genes cfr,4 cfr(B), cfr(C)5 and cfr(D)6 and also amino acid substitutions in ribosomal proteins L3, L4,4 L22,7 and ABC-F family members including OptrA8 and PoxtA.9 How ABC-F proteins mediate antibiotic resistance has been in a long-standing dispute with both efflux and ribosome protection hypothesis competing mutually. Current studies seem inclined to the latter.10,11 The optrA, found in 2015, was reported for its ATP hydrolysis activity to be associated with linezolid resistance.12 Also, previous study reported that there were different amino acid substitutions in optrA.13 In this study, we investigated the molecular characterization of optrA-positive enterococci and explored the function of varied types of optrA on the linezolid susceptibility.
Materials and Methods
Bacterial Strains
We examined 573 strains collected in 2017 at Huashan hospital, a tertiary hospital from Shanghai in China (Table 1). All were enterococci identified by bioMerieux Vitek-2 Compact and were stored in 30% glycerol broth under −80°C. The culture medium used in this study were BHIA (Brain-Heart Infusion Agar), BHIB (BrainHeart Infusion Broth), MHA (Mullex-Hinton Agar) and MHB (Mullex-Hinton Broth) (Thermo Fisher Oxoid, England).
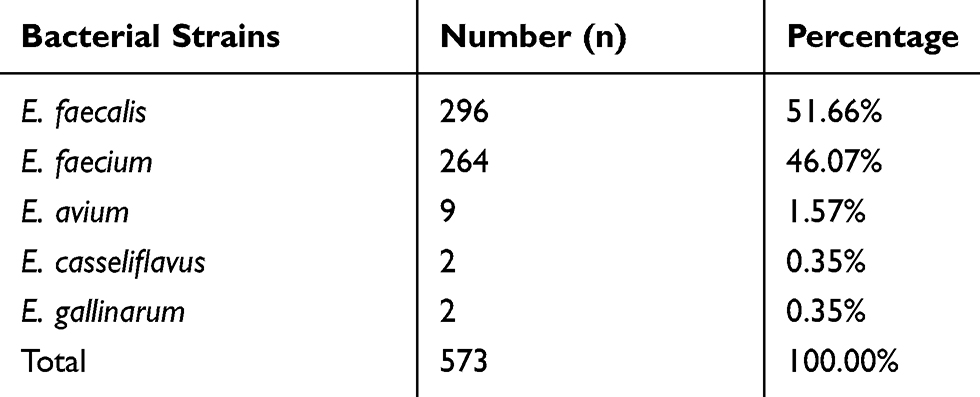 |
Table 1 The Distribution of Different Species of Enterococcus Isolates from Huashan Hospital in 2017 |
Screening of optrA and Other Linezolid Resistance Related Genes in Enterococci
The length of optrA is 1968 bp.8 We screened 573 enterococci mentioned above by using PCR. All DNA samples were amplified using primers optrA-F1 (5ʹ-CAGGTGGTCAGCGAACTAAGA-3ʹ) and optrA-R1 (5ʹ-AGCCAAGAGCAGTTCTGACC-3ʹ), of which the product size was 792 bp. The thermal conditions were 1 cycle of 94°C for 5 min followed by 30 cycles of 94°C for 30 s, 56°C for 30 s, and 72°C for 30 s and 72°C for 5 min to stop the extension step. Then, amplifications were carried out in the optrA-positive strains for getting the whole sequence of optrA and detecting the existence of cfr,14 cfr(B), cfr(C), poxtA and mutations in rplC, rplD,15 rplV, and 23S rRNA V domain16 which were associated with linezolid resistance. Primers of these amplifications were shown in Supplementary 1.
Antimicrobial Susceptibility Testing
The MIC for all optrA-positive isolates were determined by broth microdilution and agar dilution tests following the recommendations given in the Clinical and Laboratory Standards Institute (CLSI) documents M100-ED28.17 Enterococcus faecalis ATCC29212 served as the quality control strain. These antimicrobial susceptibility tests involved 18 antibiotics (Shanghai Food and Drug Administration, Shanghai, China) which were listed in Supplementary 4. All tests were performed by using broth microdilution except for fosfomycin which was practiced with agar dilution. In CLSI, the approved MIC testing of fosfomycin is agar dilution, and the agar media should be supplemented with 25 μg/mL of glucose-6-phosphate.
Molecular Typing of optrA-Positive Enterococci
The phylogenetic distance among isolates harboring optrA was analyzed by PFGE. The sequence typing (ST) of all optrA-positive enterococci were identified through MLST. The MLST analysis was on the basis of amplifying the seven housekeeping genes gdh, gyd, psts, gki, aroE, xpt, yqil in E. faecalis and the other seven genes atpa, ddl, gdh, purk, gyd, psts, adk in Enterococcus faecium. According to the sequences provided in PubMLST (https://pubmlst.org/data/), primers for MLST were designed. All STs were determined by using the Enterococcus MLST database (https://pubmlst.org/efaecalis/and https://pubmlst.org/efaecium/).
Determination on the Function of Various Types of optrA
From the study published in 2015,8 the optrA found in plasmid pE394 (GenBank accession no. KP399637.1) was defined as the wildtype. A plasmid called E. faecalis JH2-2_pAM401ΩoptrA was constructed to confirm that the optrA could result in the resistance of linezolid in enterococci. Therefore, in our research, in order to investigate the effects of the amino acid substitutions of optrA on linezolid resistance, we followed the steps in the aforementioned article for the construction of the plasmid OG1RF_pABG5.0ΩoptrA (wildtype and mutations). pABG5.018 served as the shuttle vector which could exist both in Escherichia coli and E. faecalis.
In-Fusion HD Cloning Kits (Takara) were used in the experiment. After the successful construction, the plasmid was transferred into OG1RF,19 a linezolid susceptible E. faecalis (MIC=2μg/mL), and then the optrA-positive clones were selected for further antimicrobial susceptibility test.
Results
The Distribution of optrA and Its Molecular Types
In all 573 strains isolated from Huashan hospital in 2017, we detected 20 optrA-positive isolates, numbered as 8, 37, 91, 154, 155, 185, 216, 223, 280, 297, 366, 397, 402, 404, 413, 421, 422, 446, 550, and 551, which accounted for 3.48% of the total strains. In Table 2, we could see that there were 14 optrA variants with 1 to 6 alterations at different positions. I104R, T112K, Y176D, and E256K were the most common amino acid substitutions; 18 of 20 strains were separated into 14 various STs, and the remaining two strains could not be identified in known STs.
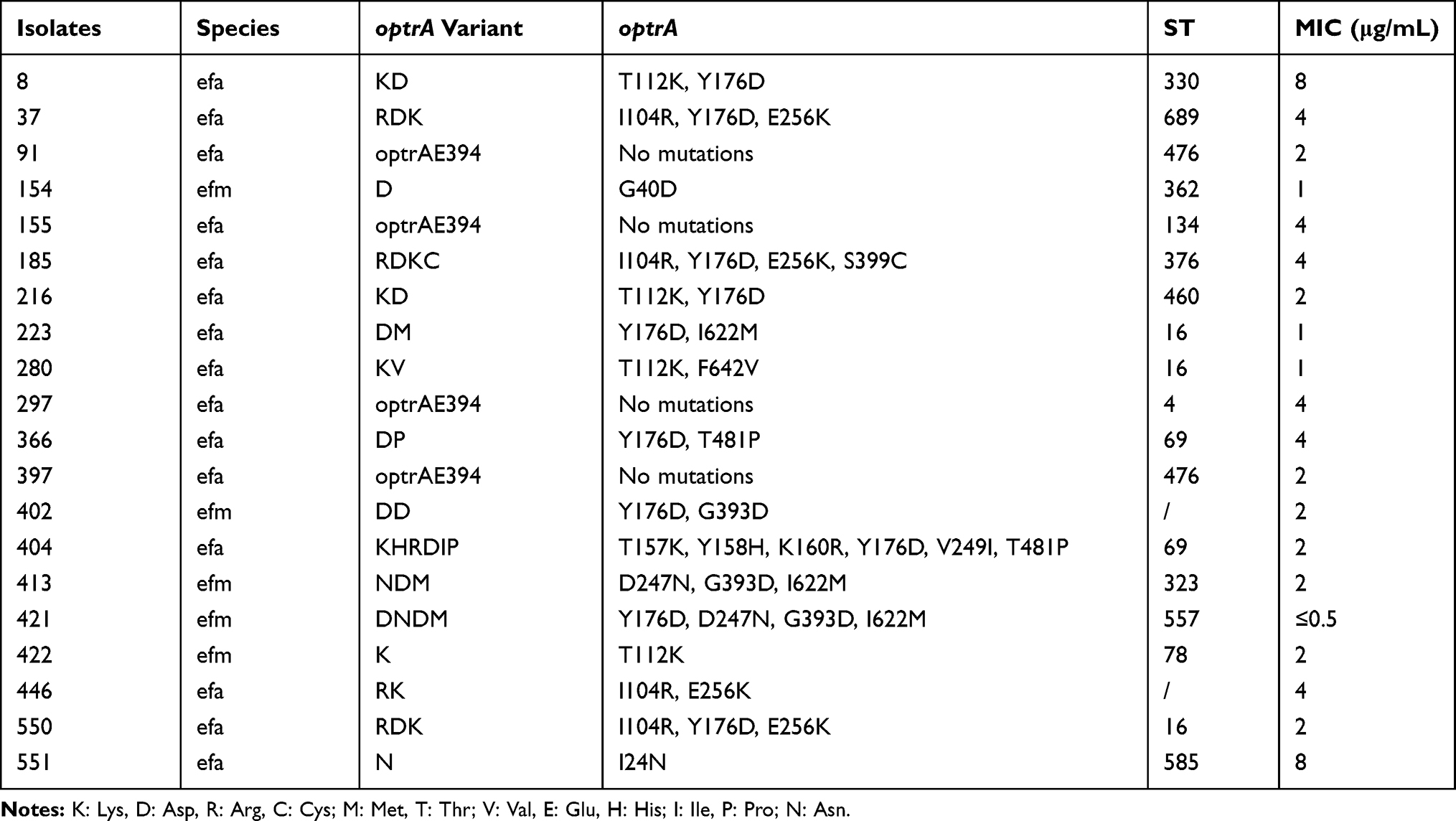 |
Table 2 Twenty optrA-Positive Strains Were Numbered and Their MLST Types Were Listed as Shown Below. We Found the Existence of Various optrA Mutations Among Those 20 Which Presented Diverse MICs of Linezolid. According to CLSI, The Criteria of Judging Susceptibility Included Susceptible (MIC≤2μg/mL), Intermediate (MIC=4μg/mL), Resistant (MIC≥8μg/mL) |
The detection and amino acid substitutions of other linezolid-resistant-related genes were presented in Supplementary 2. All genes amplified above were compared with the sequences from E. faecalis and E. faecium attainable in NCBI. (https://www.ncbi.nlm.nih.gov/nuccore/NC_01-7316.1 and https://www.ncbi.nlm.nih.gov/genome/?term=Enterococcus+faecium+DO). None amino acid substitution of 23S rRNA V domain was detected in E. faecalis. None of these 20 strains contained cfr, cfr(B), cfr(C) genes while 397 harbored cfr(D) gene. Nineteen strains had F101L mutations of L4, while no mutation of L22 was found, and G4K/D/R were the main mutation sites of L3. Only one strain possessed both optrA and poxtA.
For further study on whether there was the dissemination of optrA in Huashan hospital, we performed PFGE and MLST. Similarity among 20 strain patterns was figured out by computerized band analysis with Bionumerics software version 3.3. This part of results could be seen in Supplementary 3. No obvious homology was found in these 20 strains harboring optrA.
Antimicrobial Susceptibility Testing
The results of the antimicrobial susceptibility testing were shown in Supplementary 4. All strains were resistant to tetracycline, kanamycin, erythromycin, and ciprofloxacin while they were susceptible to vancomycin and teicoplanin. The distribution of resistance in the remaining 8 antibiotics were followed by ampicillin (5/20, 25%), penicillin (5/20, 25%), linezolid (2/20, 10%), rifampicin (5/20, 20%), chloramphenicol (15/20, 75%), and fosfomycin (3/20, 15%). Only one strain was found with tigecycline MIC as 4 μg/mL. The imipenem MIC of 4 strains were equal to or over 8 μg/mL. More detailed tests on linezolid were further put into practice. Only two strains were found as linezolid resistant, obtained by broth microdilution separately.
Function of Two Types of optrA Variants in Linezolid Resistance
Isolates 8 (optrA-KD) and 37 (optrA-RDK) were found containing the most common alterations of optrA among these 20 strains. Based on this result, they were chosen for the transformation. The successful transformants (designated OG1RF_pABG5.0ΩoptrA-KD and OG1RF_pABG5.0ΩoptrA-RDK) exhibited 2 to 4 folds increase in the MIC of linezolid compared with the recipient strain OG1RF (Table 3).
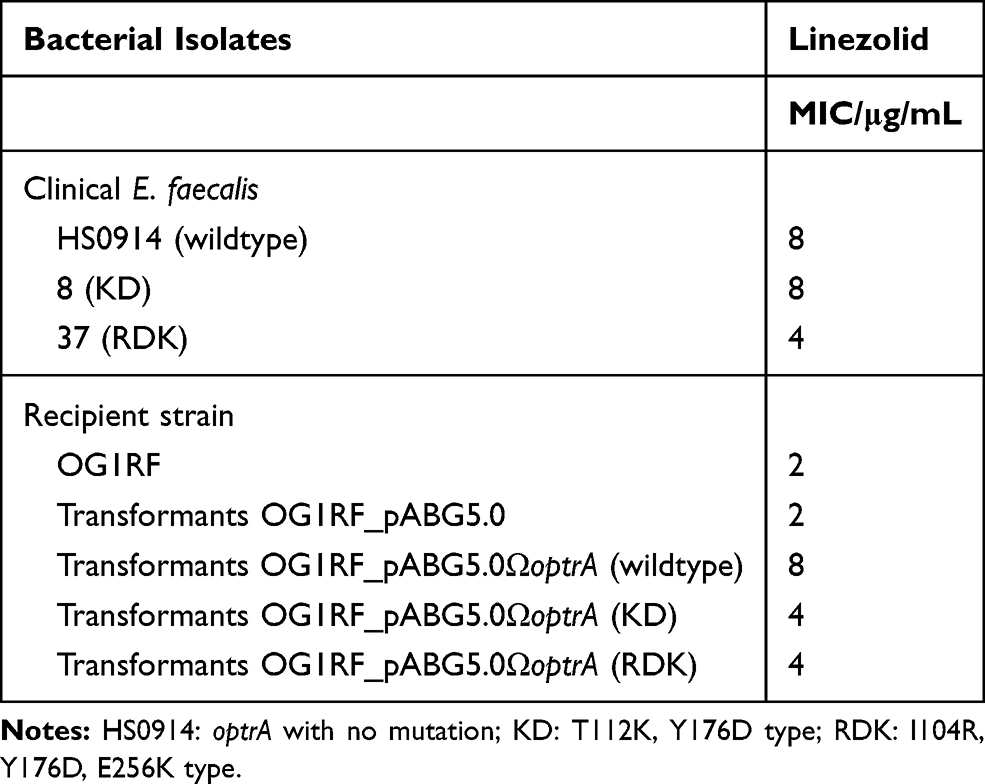 |
Table 3 MICs for Clinical E. faecalis with optrA Variants, Their Transformants and Recipient Strains |
Discussion
In 2017, E. faecalis and E. faecium were two main types of bacterial pathogens isolated from Huashan hospital. The detection rate of optrA in 573 strains was not high. ST-16 was the type that commonly existed among 14 STs (3/20) as mentioned in Table 2 which coincided with the common type of enterococci harboring optrA in other districts.20,21 These comparisons indicated that optrA was carried by low-homology enterococci isolates.
The results of the antimicrobial susceptibility tests have revealed that multidrug-resistant enterococci were not the minority. Resistance levels of linezolid in enterococci containing optrA were unevenly distributed.
Through the method of broth microdilution, only two strains were detected with low linezolid resistant level (MIC=8 μg/mL). This conceded that optrA was not only present in linezolid-resistant enterococci. At the same time, 20 strains harboring optrA had different numbers of amino acid substitutions. In the preliminary experiments, the successful construction of optrA over-expressing strains had shown that KD (T112K, Y176D) (BankIt2317497 KD MT122998) and RDK (I104R, Y176D, E256K) (BankIt2317550 RDK MT122999) optrA variants were able to raise linezolid MIC but could not reach the MIC as shown in the wildtype optrA in pE349. Potential mechanisms of mutations that affected the level of resistance of the bacteria to linezolid deserve further study. However, not all types of variants were used to construct the plasmid for over-expressing the optrA. Interestingly, 91 and 397 with no mutation of optrA were susceptible to linezolid (MIC=2 μg/mL). Whether optrA gene expressed and translated or there were some other mechanisms antagonizing the ribosome protection mechanism which led to the loss of drug resistance phenotype will be another avenue of research.
Screening of other linezolid-resistant genes showed that the cfr, cfr(B), and cfr(C) detection rates were zero, which were consistent to the results of MIC. Only one E. faecalis was detected to carry cfr(D).In this year, a study from europe characterized the novel cfr(D) gene identified in an E. faecium clinical isolate.6 According to the results mentioned above, it might probably that the cfr(D) gene tended to transfer between two kinds of Enterococcus. Whether it played a role of conferring linezolid resistance in Enterococcus still needs to be explored.
F101L mutation of L3 protein was common among 19 strains. Whether it participated in the process of linezolid resistance remains to be confirmed. For lack of the mature techniques of gene knockout in Enterococcus, it is not easy to verify the concrete functions of these substitutions.
The newly discovered PoxtA, which also belongs to ABC-F proteins, shares 32% homology identity with OptrA. Of the 20 strains, only 402 had OptrA (Y176D, G393D) and PoxtA (no alterations). Surprisingly these two displayed no synergistic effects because its MIC showed that it was a susceptible one. Whether they existed in one plasmid, like pE035 obtained from swine carrying both optrA and poxtA,22 or neither was expressed or had other negative interactions is vague at the moment. Moreover, since 421 was extremely susceptible to linezolid (MIC<0.5 μg/mL), whether its regulatory region was interfered still needs to be checked.
In conclusion, although the detection rate of optrA in Enterococcus spp. isolates was not high, and there was no obvious evidence of its dissemination in hospital, its role in linezolid resistance still needs to be noted. OptrA mutations affected the linezolid resistance at varying degrees. Detailed mechanisms such as how optrA variants interact with linezolid are of more need to be clarified.
Ethical Statement
The strains we used in this study were obtained from the biological sample and strains bank of the Institute of Antibiotics, Huashan Hospital, Shanghai, China. They came from the normal clinical testing and were stored in the strains bank. The ethics committee of Huashan Hospital authorized our study and written informed consent is not required. This study would not do harm to rights, benefits, and health of the subjects, and the privacy and personal identity information of the subjects will not be included in this study.
Acknowledgments
We thank Jinxin Zheng (Medical Molecular Virology Laboratory of Fudan University) for providing plasmids pABG5.0. We are grateful to the following colleagues at Huashan Hospital for their advice, assistance: Xuefei Zhang, Dan Li, Qingqing Xu, Li Ding, Leilei Wang, Renru Han, Mengyun Yin.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest for this work.
References
1. Leach KL, Brickner SJ, Noe MC. Linezolid, the first oxazolidinone antibacterial agent. Ann N Y Acad Sci. 2011;1222(1):49–54. doi:10.1111/j.1749-6632.2011.05962.x
2. Cattoir V, Giard J-C. Antibiotic resistance in Enterococcus faecium clinical isolates. Expert Rev Anti Infect Ther. 2014;12:239–248. doi:10.1586/14787210.2014.870886
3. Bi R, Qin T, Fan W, Ma P, Gu B. The emerging problem of linezolid-resistant enterococci. J Glob Antimicrob Resist. 2018;13:11–19. doi:10.1016/j.jgar.2017.10.018
4. Miller WR, Munita JM, Arias CA. Mechanisms of antibiotic resistance in enterococci. Expert Rev Anti Infect Ther. 2014;12:1221–1236. doi:10.1586/14787210.2014.956092
5. Tang Y, Dai L, Sahin O, Wu Z, Liu M, Zhang Q. Emergence of a plasmid-borne multidrμg resistance gene cfr(C) in foodborne pathogen Campylobacter. J Antimicrob Chemother. 2017;72:1581–1588. doi:10.1093/jac/dkx023
6. Franois G, Mohamed S, Loren D, et al. Molecular and functional analysis of the novel cfr(D) linezolid resistance gene identified in Enterococcus faecium. J Antimicrob Chemother. 2020. doi:10.1093/jac/dkaa125
7. Mendes RE, Deshpande LM, Jones RN. Linezolid update: stable in vitro activity following more than a decade of clinical use and summary of associated resistance mechanisms. Drug Resist Updat. 2014;17:1–12. doi:10.1016/j.drup.2014.04.002
8. Wang Y, Lv Y, Cai J, et al. A novel gene, optrA, that confers transferable resistance to oxazolidinones and phenicols and its presence in enterococcus faecalis and enterococcus faecium of human and animal origin. J Antimicrob Chemother. 2015;70:2182. doi:10.1093/jac/dkv116
9. Antonelli A, D’Andrea MM, Brenciani A, et al. Characterization of poxtA, a novel phenicol-oxazolidinone-tetracycline resistance gene from an MRSA of clinical origin. J Antimicrob Chemother. 2018;73:1763–1769. doi:10.1093/jac/dky088
10. Sharkey LK, Edwards TA, O’Neill AJ. ABC-F proteins mediate antibiotic resistance throμgh ribosomal protection. Mbio. 2016;7:e01975. doi:10.1128/mBio.01975-15
11. Wang Y, Li X, Wang Y, Schwarz S, Shen J, Xia X. Intracellular accumulation of linezolid and florfenicol in optrA-producing enterococcus faecalis and staphylococcus aureus. Molecules. 2018;23:3195. doi:10.3390/molecules23123195
12. Zhong XB, Xiang H, Wang TD, et al. A novel inhibitor of the new antibiotic resistance protein OptrA. Chem Biol Drug Des. 2018;92:1458–1467. doi:10.1111/cbdd.13311
13. Cui L, Wang Y, Lv Y, et al. Nationwide surveillance of novel oxazolidinone resistance gene optrA in enterococcus isolates in China from 2004 to 2014. Antimicrob Agents Chemother. 2016;60:7490–7493. doi:10.1128/AAC.01256-16
14. Wang L, He Y, Xia Y, Wang H, Liang S. Investigation of mechanism and molecular epidemiology of linezolid-resistant enterococcus faecalis in China. Infect Genet Evol. 2014;26:14–19. doi:10.1016/j.meegid.2014.05.001
15. Diaz L, Kiratisin P, Mendes RE, Panesso D, Singh KV, Arias CA. Transferable plasmid-mediated resistance to linezolid due to cfr in a human clinical isolate of enterococcus faecalis. Antimicrob Agents Chemother. 2012;56:3917–3922. doi:10.1128/AAC.00419-12
16. Bourgeois-Nicolaos N, Massias L, Couson B, Butel MJ, Andremont A, Doucet- Populaire F. Dose dependence of emergence of resistance to linezolid in enterococcus faecalis in vivo. J Infect Dis. 2007;195:1480–1488. doi:10.1086/513876
17. CLSI. Performance Standards for Antimicrobial Susceptibility Testing. CLSI Supplement M100.
18. Guiton PS, Hung CS, Kline KA, et al. Contribution of autolysin and sortase A during enterococcus faecalis DNA-dependent biofilm development. Infect Immun. 2009;77:3626–3638. doi:10.1128/IAI.00219-09
19. Dunny GM, Brown BL, Clewell DB. Induced cell aggregation and mating in Streptococcus faecalis: evidence for a bacterial sex pheromone. Proc Natl Acad Sci USA. 1978;75:3479–3483. doi:10.1073/pnas.75.7.3479
20. Zheng JX, Bai. B, Lin ZW, et al. Characterization of biofilm formation by Enterococcus faecalis isolates derived from urinary tract infections in China. J Med Microbiol. 2017. doi:10.1099/jmm.0.000647
21. Zhou WQ, Gao S, Xu HJ, et al. Distribution of the optrA gene in enterococcus isolates at a tertiary care hospital in China. J Glob Antimicrob Resist. 2019;17:180–186. doi:10.1016/j.jgar.2019.01.001
22. Hao W, Shan X, Li D, Schwarz S, Zhang SM, Li XS. Analysis of a poxtA- and optrA-co-carrying conjμgative multiresistance plasmid from Enterococcus faecalis. J Antimicrob Chemother. 2019;70:1771–1775. doi:10.1093/jac/dkz109
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.