Back to Journals » Infection and Drug Resistance » Volume 13
Molecular Identification, and Characterization of Mycobacterium kansasii Strains Isolated from Four Tuberculosis Regional Reference Laboratories in Iran During 2016–2018
Authors Khosravi AD ![]() , Asban B, Hashemzadeh M
, Asban B, Hashemzadeh M ![]() , Nashibi R
, Nashibi R
Received 8 January 2020
Accepted for publication 4 June 2020
Published 7 July 2020 Volume 2020:13 Pages 2171—2180
DOI https://doi.org/10.2147/IDR.S245295
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Suresh Antony
Azar Dokht Khosravi,1,2 Bahareh Asban,1 Mohammad Hashemzadeh,1,2 Roohangiz Nashibi2,3
1Department of Microbiology, Faculty of Medicine, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran; 2Infectious and Tropical Diseases Research Center, Health Research Institute, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran; 3Infectious Diseases & Tropical Medicine Ward, Razi Teaching Hospital, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran
Correspondence: Mohammad Hashemzadeh
Department of Microbiology, School of Medicine, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran
Tel +98 613330074
Fax +98 61 33332036
Email [email protected]
Background: Non-tuberculous mycobacterial (NTM) infections are growing concern in many countries around the globe including Iran. Among them, Mycobacterium kansasii (M. kansasii) causes both pulmonary and extra-pulmonary infections. Despite the high prevalence of M. kansasii isolates in Iran, unfortunately little is known about the epidemiological aspects of M. kansasii infection. Hence, the aim of the present study was to investigate the molecular identification, determination of subtypes variation and geographic distribution of clinical isolates of M. kansasii isolates.
Methods: In the present study, 108 clinical pulmonary isolates suspected to NTM were collected from four Tuberculosis Regional Reference Laboratories in Iran during 2016– 2018. The isolates were confirmed as NTM using conventional and molecular methods. Among them, M. kansasii isolates were subjected to rpoB gene sequencing. For determination of subtyping of M. kansasii isolates, polymerase chain reaction-restriction enzyme analysis (PCR-REA) based on the hsp65 gene was performed.
Results: Based on the rpoB gene sequence analysis, 33 (30.5%) isolates were identified as M. kansasii species, compared to 31 (28.7%) isolates using phenotypic methods. The subtype I was the most frequent subtype (n=24; 72.7%), followed by subtype II (n=8; 24.2%).
Conclusion: We indicated that the rate of M. kansasii isolation with clinical significance appears to be increasing in Iran, especially in highly industrialized cities. The high rate of M. kansasii subtype I may suggest that this genotype has a particular potency for colonization, and a higher epidemiological potential for causing infection in humans. More studies are needed to provide a better understanding of the biology and pathogenicity of M. kansasii subtype I.
Keywords: Mycobacterium kansasii, subtype, PCR-restriction enzyme analysis, rpoB
Introduction
Non-tuberculous mycobacterial (NTM) infections are growing concern in many countries around the globe. They are considered opportunistic pathogens and are ubiquitous in the environmental sources.1 Today, more than 200 Mycobacterium species were known and NTM infections have been on the rise, probably because of improved culturing techniques, the development of new molecular methods, increasing the number of susceptible hosts, and greater awareness of NTM diseases.2,3 In recent years the attention of the Iranian health care system to NTM has been increased due to their clinical significance and involvement in various human diseases especially among immunocompromised individuals. Based on a meta-analysis report from Iran, 10.2% of culture positive sputum samples were belonged to NTM,4 which this prevalence is raised to 11.1% recently,5 and emphasizes on the important role of these bacteria in human infections.
Mycobacterium kansasii (M. kansasii) is a slow-growing NTM and causes both pulmonary and extra-pulmonary infections in immunocompetent and immunosuppressed individuals.6 According to a previous global report, M. kansasii was the sixth most frequently isolated NTM from clinical specimens.7 Moreover in a recent report, M. kansasii with a prevalence of 69.2% was the most common cause of pulmonary NTM disease in sub-Saharan Africa.8 The frequency of this pathogen in Iran is high, exceeding 13–17% of total NTM isolations, compared to respective rates globally.9 Treatment of M. kansasii infections is usually challenging, because of multi-drug therapy and long treatment periods up to 12 months.4,10,9,11 In an economically challenged country like Iran, due to inadequate laboratory facilities, the diagnosis of tuberculosis (TB) is still only by direct smear microscopy. In many cases identification of Mycobacterium to the species level is not done and NTM diseases are frequently misdiagnosed as TB, particularly in the case of M. kansasii infections that the clinical and radiologic manifestations often indistinguishable from pulmonary TB.12
Species identification in Mycobacterium genus is challenging due to a complex and changing taxonomy, the unreliability of traditional methods and the inability of 16S rRNA to discriminate some closely related species such as M. kansasii and M. gastri.1,3,However, other targets such as rpoB gene, dnaK gene, tuf gene, hsp65 gene, sodA gene and ITS (16S–23S internal transcribed spacer) region are usually suitable for species identification.14
Seven subtypes (genotypes, I–VII) for M. kansasii have been described. Of these, I and II are the most prevalent types and have been associated with human disease, whereas the other five (III–VII) are predominantly of environmental origin.15,16
Therefore, subtyping of M. kansasii isolates from human samples may serve as a prediction for clinical diagnosis. There are three major methodologies used for the identification of M. kansasii subtypes I–VII. One is sequence analysis of either hsp65 or rpoB genes, the other is 16S-23S rDNA ITS, and the third is tuf typing.17–19 In a most recent work conducted by Jagielski et al, the former M. kansasii subtypes have been denominated as M. kansasii (former type I), M. persicum (II), M. pseudokansasii (III), M. innocens (V), and M. attenuatum (VI), based on the alignment fraction-average nucleotide identity, genome-to-genome distance, and core-genome phylogeny.20
Despite the high prevalence of M. kansasii isolates among NTM positive cultures in Iran, unfortunately little is known about the epidemiological aspects of M. kansasii infection. Hence, the aim of the present study was to investigate the molecular identification, determination of subtypes variation and geographic distribution of clinical isolates of M. kansasii from patients in four Tuberculosis Regional Reference Laboratories in Iran during years 2016–2018.
Materials and Methods
Data Source and Ethic Statement
In the present study, 108 clinical isolates of pulmonary origin (sputum and broncho-alveolar lavage [BAL]) suspected to NTM were collected over a 3-year period from January 2016 to May 2018. The isolates were obtained from patients referred to four regional tuberculosis reference laboratories of Iran including Khuzestan (38 isolates/35.2%), Tehran (34 isolates/31.5%), Kermanshah (19 isolates/17.6%) and Fars (17 isolates/15.7%). Demographic data including patients’ gender, age, and the region of residence, type of specimen, date of collection, and source laboratory were recorded. None of the individual patients were commenced on therapy for NTM disease. The isolates not matching with the American Thoracic Society/Infectious Diseases Society of America (ATS/IDSA) criteria for the definition of NTM disease21 were excluded from the study. The study was approved by Institutional Ethics and Review Board (Code: (IR.AJUMS.REC.1397.234) after submission of the preliminary proposal, and necessary permission for sample collection was granted.
Phenotypic Identification
All 108 clinical NTM isolates were subjected to phenotypic identification. Each isolate was subcultured on Lowenstein-Jensen (LJ) medium and grown colonies were stained by the Ziehl-Neelsen method for the presence of acid-fast bacilli (AFB). For initial identification, conventional phenotypic and biochemical tests including colony morphology, growth at 25, 37, and 42°C, pigment production, semi-quantitative catalase test, Tween 80 hydrolysis, arylsulfatase test, heat-stable catalase (pH 7, 68°C), urease, and nitrate reduction test were performed.22 M. kansasii ATCC 12478 was used as reference strain.
Molecular Identification
DNA Extraction
Chromosomal DNA was extracted from mycobacterial colonies grown on LJ medium using an extraction and purification QIAamp DNA Mini Kit (Qiagen NV, Venlo, the Netherlands), according to the manufacturer’s instructions. DNA concentrations and purity were determined using a Nano Drop One instrument (Thermo Fisher Scientific, Waltham, MA, USA) at 260 nm and used as template in PCR amplification.
Molecular Assignment to Species Level
A 750-bp fragment of the rpoB gene was amplified using primers MycoF (5′ GGCAAGGTCACCCCGAAGGG-3′) and MycoR (5′-AGCGGCTGCTGGGTGATCATC-3′) as described earlier.23 The amplified PCR products for each isolate were purified with the Gene JETTM Gel Extraction Kit (Fermentas, Lithuania), as per manufacturer’s instructions. The sequences of the products were determined using an ABI PRISM 7500 Sequence Detection System (Applied Biosystems, Foster City, CA, United States). The sequences of rpoB gene for each isolate were confirmed by BLAST separately, and multiple sequence alignment (MSA) was done for our sequences and the existing relevant sequences belong to M. kansasii recovered from GenBank database, using MEGA7 program.24
Methods of Subtyping
PCR-Restriction Enzyme Analysis (REA)
Subtyping of M. kansasii isolates was performed using PCR-REA method for the hsp65 gene, as previously described.17 An approximately 441 bp fragment of the hsp65 gene was amplified by PCR using two specific primers Tb11 (5′- ACCAACGATGGTGTGTCCAT-3′) and Tb12 (5′-CTTGTC GAACCGCA- TACCCT-3′). Genomic DNA of M. fortuitum ATCC 49404T and double distilled water were used as positive and negative controls, respectively. PCR products of hsp65 gene were digested by the BstEII and HaeIII restriction enzymes. The fragments were compared with those of patterns deposited in a free available database (http://app.chuv.ch/prasite, n.d.), for subtyping of M. kansasii isolates.
rpoB Gene Sequencing
Primer Design for Sequence-Based Subtyping
The sequences of rpoB gene for each subtype were recovered from GenBank database and MSA were done using MEGA7 program.24 According to the MSA results, we found a region with a suitable variation for M. kansasii subtyping. Based on this region, a forward primer (5′-AAT CAA CCT GTC GCG CAA CGA-3′) and a reverse primer (5′-GTT CAT CGA AGA AGT TGA CGT-3′) were designed by using Gene runner 3.05 software.
Nucleotide Sequencing
For definitive subtyping of M. kansasii isolates, nearly a 250-bp fragment of the rpoB gene was amplified using a set of above-mentioned designed primers. The cycling condition was 95◦C for 1 min, followed by 30 cycles of 95◦C for 30s, 64◦C for 30s, and 72◦C for 30s and finalized with 72◦C for 5 min. The amplified PCR products of rpoB gene for each M. kansasii isolate were purified with the Gene JETTM Gel Extraction Kit (Fermentas, Lithuania), as described in the manufacturer’s instructions. The sequences of the products were determined by application of an ABI PRISM 7500 Sequence Detection System (Applied Biosystems, Foster City, CA, United States) according to the standard protocol of the supplier. Phylogenetic trees were obtained from DNA sequences based on 250 bp fragments using the Neighbor-Joining (NJ) method and Kimura’s two parameter (K2P) distance correction model with 1000 bootstrap replications supported by the MEGA7 software.
Results
All 108 NTM-suspected clinical isolates were identified as NTM using conventional culture and phenotypic identification tests (Table 1). These were obtained from patients with positive cultures met (ATS/IDSA) diagnostic criteria for NTM disease.21 Tables 2 and 3 describe the proportions of NTM isolates within our population and those meeting ATS/IDSA guidelines. According to phenotypic tests, 31 isolates (28.7%) were assigned to M. kansasii. For definite identification, all 108 isolates were subjected to rpoB gene sequencing and PRA, which revealed that 33 isolates (30.5%) showed more than 99% homology with M. kansasii species and were included in our study (Table 1). The M. kansasii isolates were obtained from samples belonged to 25 male (75.7%) and 8 female (24.3%) patients with mean age of 56.7% years, and the age group >60 years (n= 15, 45.4%), was the highly infected group (Table 2).
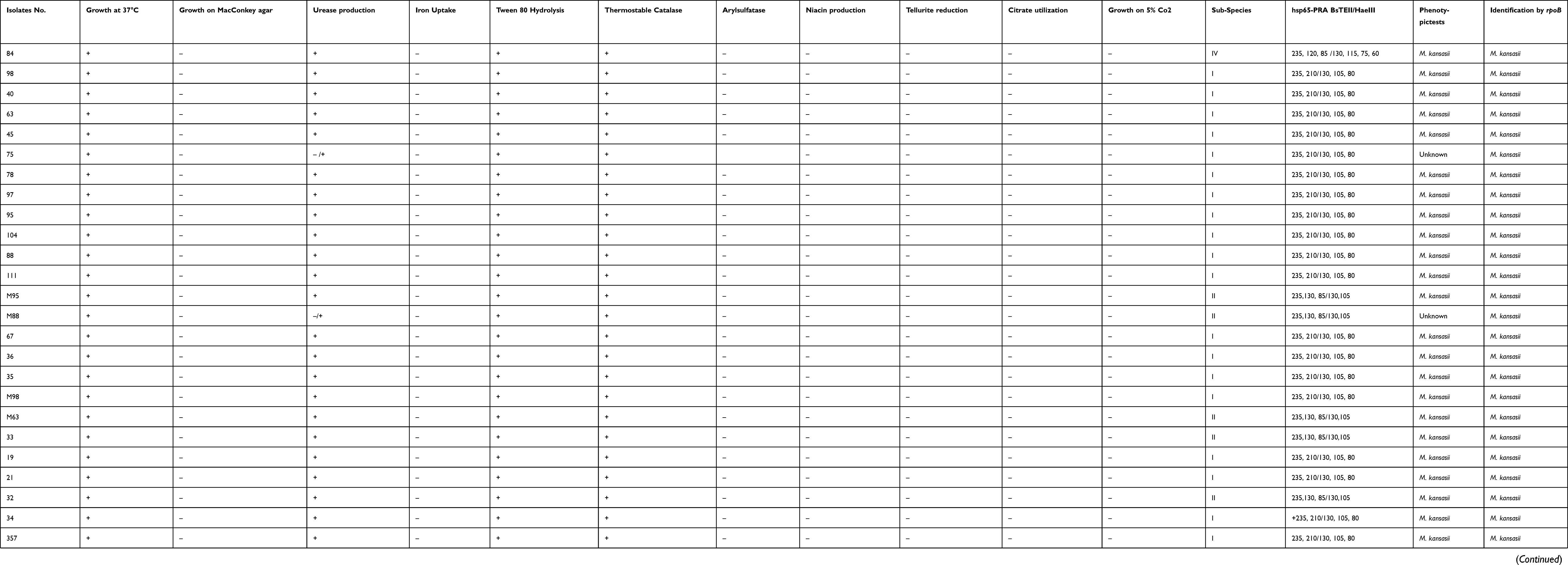 |
Table 1 Results of M. kansasii Identification by Phenotypic and Genotypic Tests |
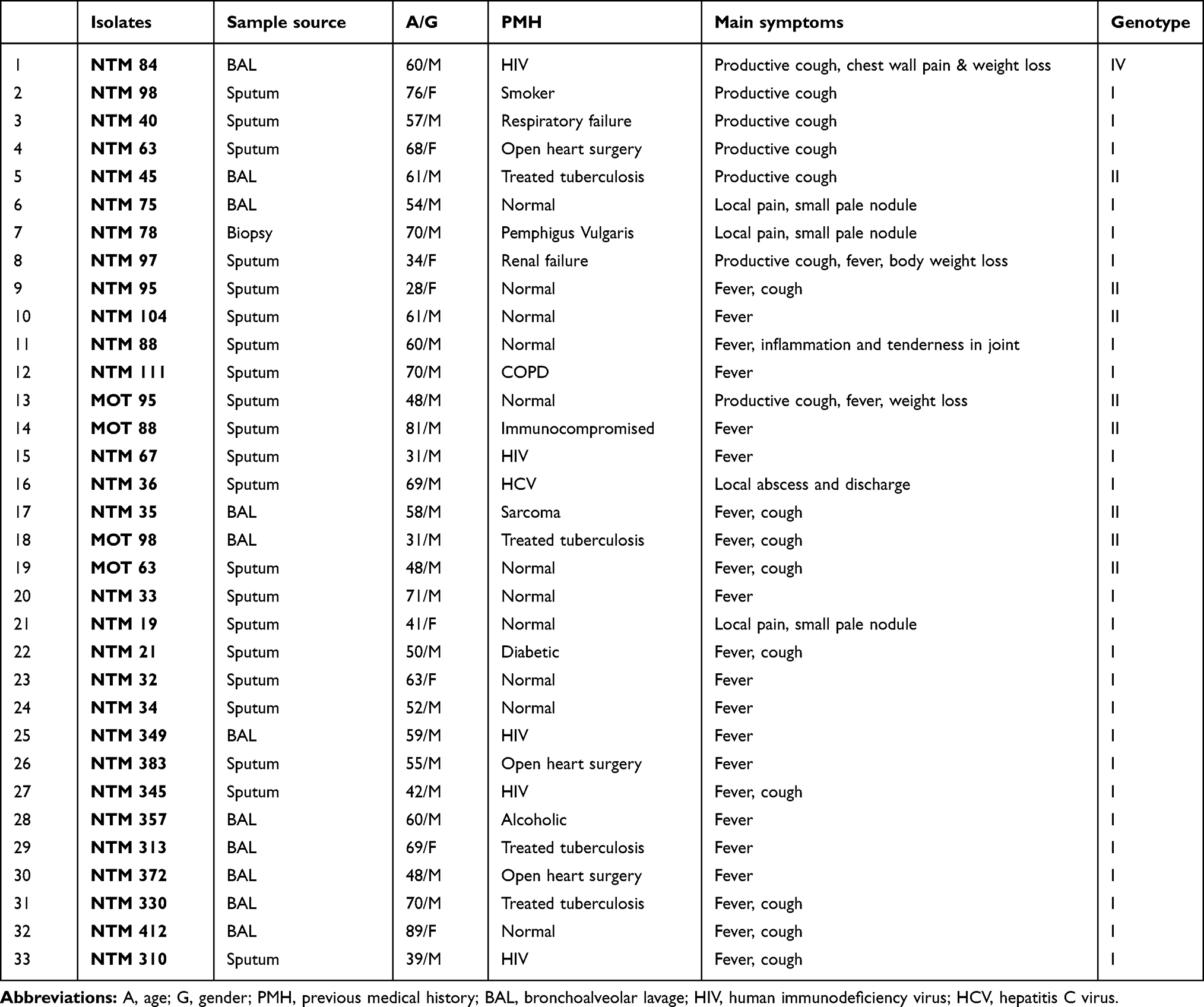 |
Table 2 Baseline Characteristics of Patients with M. kansasii-Positive Culture from Respiratory Samples |
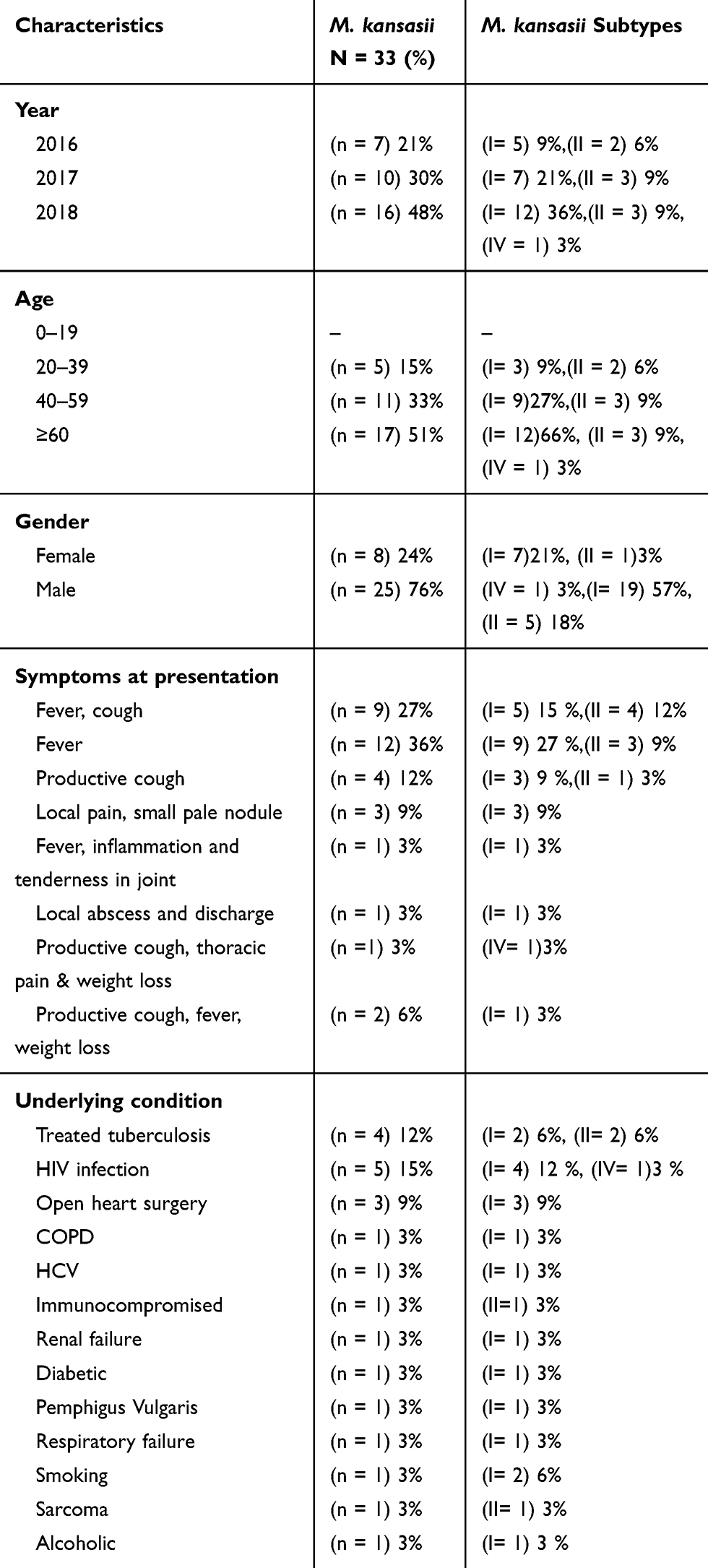 |
Table 3 Frequency of M. kansasii Isolates and Subtypes by Characteristic, Symptoms at Presentation and Previous Medical History |
The isolates were mostly obtained from sputum samples (n=21, 63%), followed by BAL (n=11, 33%). All of 33 patients had positive AFB on three or more available specimens. The analyzed M. kansasii isolates were originated from TB centers in Khuzestan (n= 12, 36%), Tehran (n= 11, 33%), Kermanshah (n= 6, 18%), and Fars (n= 4, 12%). As shown in Figure 1, the higher rate of M. kansasii infection was observed in the Khuzestan followed by Tehran provinces. According to patients’ archive information, the most frequent previous medical history (PMH) was human immunodeficiency virus (HIV) infection (n=5, 15%), and treated TB (n=4, 0.12%) (Table 3).
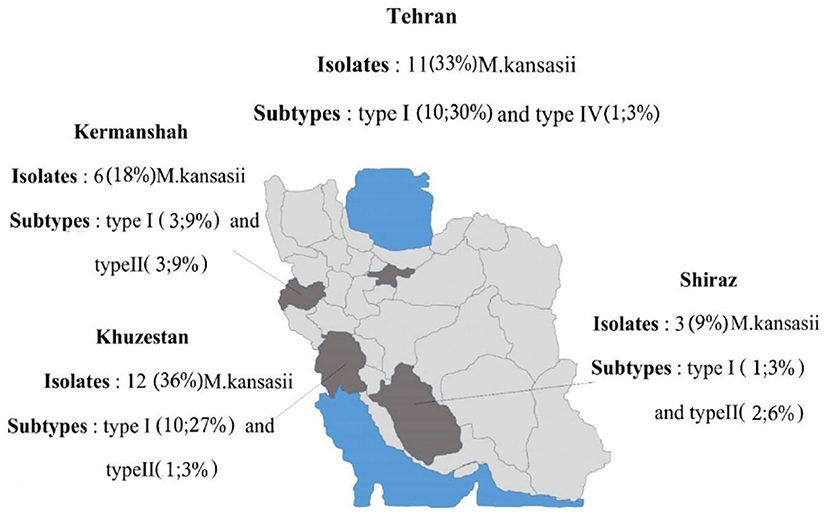 |
Figure 1 Location and geographic distribution of M. kansasii strains during 2016–2018. |
The M. kansasii isolates were genotyped by hsp65-based PRA method and partial rpoB gene sequencing. Three different subtypes of I, II, and IV were identified by both methods as shown in Table 4. The subtype I was the most frequent subtype (n=24; 73%), followed by subtype II (n=8; 24%). As presented in Table 4, the results of typing using rpoB gene sequencing (designed in this study), were identical with PRA method. Distribution of M. kansasii subtypes in parts of Iran under investigation is shown in Figure 1. Based on the sequencing results of the partial ropB gene, a dendrogram (the maximum likelihood tree generated using the Tamura 3-parameter model in MEGA7 software) illustrating the genetic distances between different M. kansasii types (I–VI), was generated (Figure 2). For all strains under the study, the typing results obtained with PCR-REA of hsp65 gene and sequencing were in 100% agreement.
 |
Table 4 Restriction Patterns and M. kansasii Sub-Typing by PRA-Hsp65 and rpoB Gene Sequencing |
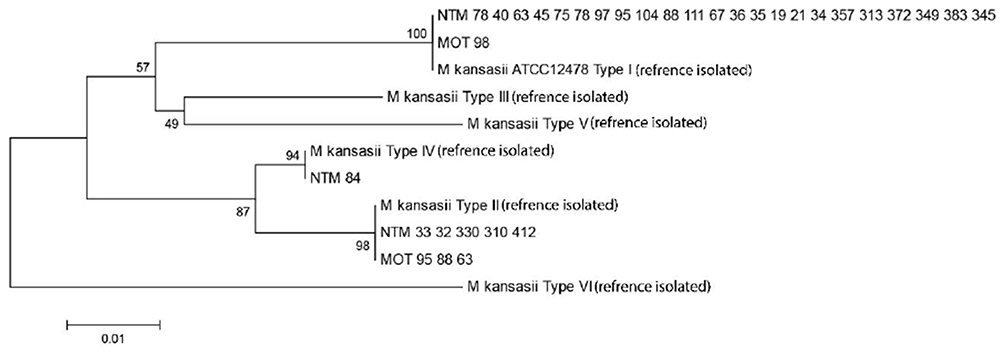 |
Figure 2 A dendrogram (the maximum likelihood tree generated using the Tamura 3-parameter model in MEGA6 software) illustrating the genetic distances between different M. kansasii types (I–VI), based on the sequencing results of the partial ropB gene. |
Discussion
Currently, the incidence and prevalence of NTM diseases are increasing, and their impact on human health has been a topic of interest.25,26 Iran is an endemic country where TB remains a major public health problem. Although the epidemiology of TB is well described, the prevalence of NTM disease in Iran remains largely undetermined. M. kansasii is one of the most frequently encountered NTM species among culture positive sputum samples in Iran.4 Unfortunately, there are no reliable data on the epidemiology of M. kansasii infections in Iranian patients. Hence, to our best knowledge, this is one of the limited studies on the frequency, epidemiology and distribution of M. kansasii species from different regions in Iran, as the first report belonged to Velayati et al from Iran.27 In the present study, we applied a combination of phenotypic and molecular methods for the species identification of NTM isolates. Based on the rpoB gene sequencing and PRA method, 33 (30.5%) isolates were identified correctly as M. kansasii species, compared to 31 (28.7%) isolates using phenotypic methods, so it was confirmed that sequence-based identification offers much more resolution than the phenotypic methods, as shown in several studies.28–30
There are several reports on the frequency of M. kansasii from Iran and around the world. In studies from Iran, the clinical M. kansasii respiratory isolates has been shown to vary from 11% to 26%.4,5,30 Additionally, in South American and some European countries, M. kansasii was reported as the second most common cause of NTM diseases.31 Also, high rates of M. kansasii incidence have been consistently reported in Japan and South Africa.32,33 This wide range of differences might be explained by different sample sizes, difference in diagnostic criteria and changing in NTM epidemiology and distribution. The results of this study showed that the majority of Iranian patients with M. kansasii lung infection were from highly industrialized provinces. Among the 33 patients positive for M. kansasii, 23 (69.7%) lived in highly industrialized provinces of Tehran and Khuzestan, which this finding is in line with other studies.28,34
The hsp65-PRA assay is one of the most common techniques currently used for M. kansasii subtypes identification.29 Here, we applied both PRA and partial rpoB gene sequencing, designed in this study. Of the seven currently recognized M. kansasii subtypes (genotypes, I–VII), genotype I has predominated among clinical samples, making up 42–100% of M. kansasii clinical isolates.16 This was confirmed by this study too, and we found that the subtype I is the predominant genotype of M. kansasii (72%), as reported worldwide.7,35,36 In comparison, the frequency of this genotype was found to be lower in Switzerland (68%),37 and much higher in Brazil (98%).38 However, only 24% and 3% of M. kansasii isolates in this study were classified as subtype II and subtype IV, respectively, which is lower from other similar previous studies.7 Another study conducted in Spain revealed the absence of genotype II among M. kansasii clinical isolates, with 91 (97.8%) of the 93 isolates tested representing genotype I.39 In contrast to our study, Tortoli et al, reported that M. kansasii type II has been found more likely to be recovered from patients with HIV-positive status, whereas type I has also been associated with pulmonary infections in HIV-negative patients with pre-existing pulmonary diseases.40
The high prevalence of type I among the isolates of M. kansasii of HIV-positiveindividuals indicates that this type had a greater capacity for colonization and/or pathogenic activity for human beings, while the data available from type II suggest that this type less able to overcome natural resistance mechanisms. Many epidemiological and pathogenic aspects of M. kansasii are still far from being understood, and further investigation is necessary to elucidate the virulence factors of the M. kansasii subtypes I and II. However, the present study emphasizes on the identification of M. kansasii to the subtype level, may be not only an interesting epidemiological tool, but also a process relevant to clinical management, as it allows the differentiation of pathogenic from the nonpathogenic subtypes. Performance of drug susceptibility testing and comparing the results of drug resistance between M. kansasii subtypes, though was desired but has not been done in this study, and this was the limitation of our study due to the extreme lack of budget, however, will be our goal for future studies.
In conclusion, we showed that the rate of M. kansasii isolation with clinical significance appears to be increasing in Iran, especially in highly industrialized cities. Management of M. kansasii infections requires a multidisciplinary effort from primary care physicians, pulmonologists and infectious diseases specialists. The high detection rate of M. kansasii subtype I in clinical samples may suggest that this genotype has a particular propensity for colonization, and thus a higher epidemiological potential for causing infection in humans. More comprehensive studies, on large collections of M. kansasii isolates, are needed to provide a better understanding of the biology and pathogenicity of M. kansasii subtype I.
Acknowledgments
This work is part of MSc. thesis of Bahareh Asban, which was approved in Infectious and Tropical Diseases Research Center, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran. We are grateful to research affairs of the university for financial support of the study (Grant No. OG-9718).
Disclosure
The authors declare no conflicts of interest in this research work.
References
1. Jeon D. Infection source and epidemiology of nontuberculous mycobacterial lung disease. Tuberc Respir Dis (Seoul). 2019;82(2):94–101. doi:10.4046/trd.2018.0026
2. Forbes BA. Mycobacterial taxonomy. J Clin Microbiol. 2017;55(2):380–383. doi:10.1128/JCM.01287-16
3. McShane PJ, Glassroth J. Pulmonary disease due to nontuberculous mycobacteria: current state and new insights. Chest. 2015;148(6):1517–1527. doi:10.1378/chest.15-0458
4. Nasiri MJ, Dabiri H, Darban-Sarokhalil D, Shahraki AH. Prevalence of non-tuberculosis mycobacterial infections among tuberculosis suspects in Iran: systematic review and meta-analysis. PLoS One. 2015;10(6):e0129073. doi:10.1371/journal.pone.0129073
5. Mortazavi Z, Bahrmand A, Sakhaee F, et al. Evaluating the clinical significance of nontuberculous mycobacteria isolated from respiratory samples in Iran: an often overlooked disease. Infect Drug Resist. 2019;12:1917–1927. doi:10.2147/IDR.S214181
6. Johnston JC, Chiang L, Elwood K. Mycobacterium kansasii. Microbiol Spectr. 2017;5:1. doi:10.1128/microbiolspec.TNMI7-0011-2016
7. Bakuła Z, Safianowska A, Nowacka-Mazurek M, Bielecki J, Jagielski T. Subtyping of mycobacterium kansasii by PCR-restriction enzyme analysis of the hsp65 gene. Biomed Res Int. 2013;2013.
8. Okoi C, Anderson STB, Antonio M, Mulwa SN, Gehre F, Adetifa IMO. Non-tuberculous Mycobacteria isolated from pulmonary samples in sub-Saharan Africa - a systematic review and meta analyses. Sci Rep. 2017;7(1):12002. doi:10.1038/s41598-017-12175-z
9. Falkinham JO
10. Nasiri MJ, Dabiri H, Fooladi AA, Amini S, Hamzehloo G, Feizabadi MM. High rates of nontuberculous mycobacteria isolation from patients with presumptive tuberculosis in Iran. New Microbes New Infect. 2018;21:12–17. doi:10.1016/j.nmni.2017.08.008
11. Philley JV, DeGroote MA, Honda JR, et al. Treatment of non-tuberculous mycobacterial lung disease. Curr Treat Options Infect Dis. 2016;8(4):275–296. doi:10.1007/s40506-016-0086-4
12. Matveychuk A, Fuks L, Priess R, Hahim I, Shitrit D. Clinical and radiological features of Mycobacterium kansasii and other NTM infections. Respir Med. 2012;106(10):1472–1477. doi:10.1016/j.rmed.2012.06.023
13. Liang B, Du C, Tian X, et al. Rapid identification of clinically relevant Mycobacterium species by multicolor melting curve analysis. J Clin Microbiol. 2019;57(1).
14. Dai J, Chen Y, Dean S, Morris JG, Salfinger M, Johnson JA. Multiple-genome comparison reveals new loci for Mycobacterium species identification. J Clin Microbiol. 2011;49(1):144–153. doi:10.1128/JCM.00957-10
15. Bakuła Z, Brzostek A, Borówka P, et al. Molecular typing of Mycobacterium kansasii using pulsed-field gel electrophoresis and a newly designed variable-number tandem repeat analysis. Sci Rep. 2018;8(1):4462. doi:10.1038/s41598-018-21562-z
16. Bakuła Z, Kościuch J, Safianowska A, et al. Clinical, radiological and molecular features of Mycobacterium kansasii pulmonary disease. Respir Med. 2018;139:91–100. doi:10.1016/j.rmed.2018.05.007
17. Bakuła Z, Modrzejewska M, Safianowska A, et al. Proposal of a new method for subtyping of Mycobacterium kansasii based upon PCR restriction enzyme analysis of the tuf gene. Diagn Microbiol Infect Dis. 2016;84(4):318–321. doi:10.1016/j.diagmicrobio.2015.12.009
18. Huang CC, Chen JH, Hu ST, et al. Combined rpoB duplex PCR and hsp65 PCR restriction fragment length polymorphism with capillary electrophoresis as an effective algorithm for identification of mycobacterial species from clinical isolates. BMC Microbiol. 2012;12:137. doi:10.1186/1471-2180-12-137
19. Iwamoto T, Saito H. Comparative study of two typing methods hsp65 PRA and ITS sequencing revealed a possible evolutionary link between Mycobacterium kansasii type I and II isolates. FEMS Microbiol Lett. 2005;254:129–133. doi:10.1111/j.1574-6968.2005.00013.x
20. Jagielski T, Borówka P, Bakuła Z, et al. Genomic insights into the Mycobacterium kansasii complex: an update. Front Microbiol. 2020;10:2918. doi:10.3389/fmicb.2019.02918
21. Griffith DE, Aksamit T, Brown-Elliott BA, et al. An official ATS/IDSA statement: diagnosis, treatment, and prevention of nontuberculous mycobacterial diseases. Am J Respir Crit Care Med. 2007;175(4):367–416. doi:10.1164/rccm.200604-571ST
22. Mahon CR, Lehman DC, Manuselis G
23. Adékambi T, Colson P, Drancourt M. rpoB-based identification of non-pigmented and late-pigmenting rapidly growing mycobacteria. J Clin Microbiol. 2003;41(12):5699–5708. doi:10.1128/JCM.41.12.5699-5708.2003
24. Kumar S, Stecher G, Tamura K. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol Biol Evol. 2016;33(7):1870–1874. doi:10.1093/molbev/msw054
25. Falkinham JO. Current epidemiologic trends of the nontuberculous mycobacteria (NTM). Curr Environ Health Rep. 2016;3(2):161–167.
26. Winthrop KL, Henkle E, Walker A, Cassidy M, Hedberg K, Schafer S. On the reportability of nontuberculous mycobacterial disease to public health authorities. Ann Am Thorac Soc. 2017;14(3):314–317. doi:10.1513/AnnalsATS.201610-802PS
27. Velayati AA, Farnia P, Mozafari M, et al. Molecular epidemiology of nontuberculous mycobacteria isolates from clinical and environmental sources of a metropolitan city. PLoS One. 2014;9(12):e114428. doi:10.1371/journal.pone.0114428
28. Khosravi AD, Hashemi Shahraki A, Hashemzadeh M, Sheini Mehrabzadeh R, Teimoori A. Prevalence of non-tuberculous mycobacteria in hospital waters of major cities of Khuzestan Province, Iran. Front Cell Infect Microbiol. 2016;6:42. doi:10.3389/fcimb.2016.00042
29. Kim SH, Shin JH. Identification of nontuberculous mycobacteria using multilocus sequence analysis of 16S rRNA, hsp65, and rpoB. J Clin Lab Anal. 2018;32(1):e22184. doi:10.1002/jcla.22184
30. Shahraki AH, Trovato A, Mirsaeidi M, et al. Mycobacterium persicum sp. nov., a novel species closely related to Mycobacterium kansasii and Mycobacterium gastri. Int J Syst Evol Microbiol. 2017;67(6):1766–1770. doi:10.1099/ijsem.0.001862
31. Hoefsloot W, Van Ingen J, Andrejak C, et al. The geographic diversity of nontuberculous mycobacteria isolated from pulmonary samples: an NTM-NET collaborative study. Eur Respir J. 2013;42(6):1604–1613. doi:10.1183/09031936.00149212
32. Kobashi Y, Mouri K, Miyashita N, Oka M. Clinical usefulness of QuantiFERON TB-2G test for the early diagnosis of pulmonary Mycobacterium kansasii disease. Jpn J Infect Dis. 2009;62:239–241.
33. Van Halsema CL, Chihota VN, Gey van Pittius NC, et al. Clinical relevance of nontuberculous mycobacteria isolated from sputum in a gold mining workforce in South Africa: an observational, clinical study. Biomed Res Int. 2015;2015.
34. Yim JJ, Park YK, Lew WJ, Bai GH, Han SK, Shim YS. Mycobacterium kansasii pulmonary diseases in Korea. J Korean Med Sci. 2005;20(6):957–960. doi:10.3346/jkms.2005.20.6.957
35. Li Y, Pang Y, Tong X, Zheng H, Zhao Y, Wang C. Mycobacterium kansasii subtype I is associated with clarithromycin resistance in China. Front Microbiol. 2016;7:2097. doi:10.3389/fmicb.2016.02097
36. Thomson R, Tolson C, Huygens F, Hargreaves M. Strain variation amongst clinical and potable water isolates of M. kansasii using automated repetitive unit PCR. Int J Med Microbiol. 2014;304(3–4):484–489. doi:10.1016/j.ijmm.2014.02.004
37. Taillard C, Greub G, Weber R, et al. Clinical implications of Mycobacterium kansasii species heterogeneity: Swiss National Survey. J Clin Microbiol. 2003;41(3):1240–1244. doi:10.1128/JCM.41.3.1240-1244.2003
38. da Silva Telles MA, Chimara E, Ferrazoli L, Riley LW. Mycobacterium kansasii: antibiotic susceptibility and PCR-restriction analysis of clinical isolates. J Med Microbiol. 2005;54(10):975–979. doi:10.1099/jmm.0.45965-0
39. Santin M, Alcaide F, Benitez MA, et al. Incidence and molecular typing of Mycobacterium kansasii in a defined geographical area in Catalonia, Spain. Epidemiol Infect. 2004;132(3):425–432. doi:10.1017/S095026880300150X
40. Tortoli E, Simonetti MT, Lacchini C, Penati V, Urbano P. Tentative evidence of AIDS-associated biotype of Mycobacterium kansasii. J Clin Microbiol. 1994;32(7):1779–1782. doi:10.1128/JCM.32.7.1779-1782.1994
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.