Back to Journals » Infection and Drug Resistance » Volume 15
Molecular Characterization of Carbapenem-Resistant Acinetobacter baumannii with Special Reference to Carbapenemases: A Systematic Review
Authors Gupta N, Angadi K, Jadhav S ![]()
Received 17 August 2022
Accepted for publication 29 November 2022
Published 22 December 2022 Volume 2022:15 Pages 7631—7650
DOI https://doi.org/10.2147/IDR.S386641
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Prof. Dr. Héctor Mora-Montes
Neetu Gupta, Kalpana Angadi, Savita Jadhav
Department of Microbiology, Symbiosis Medical College for Women (SMCW) & Symbiosis University Hospital and Research Centre (SUHRC), Symbiosis International (Deemed University), Lavale, Pune, India
Correspondence: Savita Jadhav, Department of Microbiology, Symbiosis Medical College for Women (SMCW) & Symbiosis University Hospital and Research Centre (SUHRC), Symbiosis International (Deemed University), Lavale, Pune, India, Tel +919284434364, Email [email protected]
Abstract: Carbapenemases are β-lactamase enzymes that hydrolyze a variety of β-lactams including carbapenem and belong to different Ambler classes (A, B, D). These enzymes can be encoded by plasmid or chromosomal-mediated genes. The major issues associated with carbapenemases-producing organisms are compromising the activity and increasing the resistance to carbapenems which are the last resort antibiotics used in treating serious infections. The global increase of pathogen, carbapenem-resistant A. baumannii has significantly threatened public health. Thus, there is a pressing need for a better understanding of this pathogen, to know the various carbapenem resistance encoding genes and dissemination of resistance genes from A. baumannii which help in developing strategies to overcome this problem. The horizontal transfer of resistant determinants through mobile genetic elements increases the incidence of multidrug, extensive drug, and Pan-drug resistant A. baumannii. Therefore, the current review aims to know the various mechanisms of carbapenem resistance, categorize and discuss carbapenemases encoding genes and various mobile genetic elements, and the prevalence of carbapenemase genes in recent years in A. baumannii from various geographical regions.
Keywords: carbapenemases, multidrug-resistance, metallo-β-lactamases, oxacillinases
Introduction
Genus Acinetobacter is comprised of a group of species that are gram-negative coccobacilli, aerobe, non-fermenting, encapsulated, and present in the environment. Members from the genus Acinetobacter are regarded as non-motile, but there are reports which found that this pathogen showed surface swarming and produced channels (“ditches”) in the semisolid agar used usually as a motility medium.1–3 Acinetobacter can invade the body in various ways and lead to fatal infections.1 Among Acinetobacter spp., A. baumannii is the bacterial pathogen from the ESKAPE group (Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii (A. baumannii), Pseudomonas aeruginosa, and Enterobacter species) and has gained major attention in recent years as a nosocomial pathogen, especially in intensive care and burn units.4 However, community-acquired infections usually occur in people who have preexisting comorbid conditions.5 A. baumannii is part of the Acinetobacter calcoaceticus-baumannii complex, which also includes other species like Acinetobacter pittii, Acinetobacter nosocomialis, A. dijkshoorniae, A. seifertii which are pathogenic and Acinetobacter calcoaceticus which is nonpathogenic.6 A. baumannii shows increased resistance to most antibiotics and the slower pace in the development of newer antibiotics and limited treatment options have resulted in increased mortality following various infections due to this organism.1,7
The carbapenem class of drug is commonly used to treat the life-threatening infections caused by MDR Acinetobacter species.1,8 Carbapenems are bactericidal, broad-spectrum, beta-lactam antibiotics and like all beta-lactam antibiotics inhibit bacterial cell wall synthesis by binding to penicillin-binding proteins (PBPs 1a, 1b, 2, and 3) and inactivating these enzymes. Certain examples of worldwide used carbapenem are imipenem, meropenem, doripenem, ertapenem, panipenem, and biapenem.9,10 Taking into consideration carbapenem activity against A. baumannii, it has been shown that imipenem and doripenem are more potent than meropenem. Multidrug-resistant (MDR) A. baumannii including resistance to carbapenem drugs has been reported globally and is an alarming sign for clinicians due to the burden of infections caused by carbapenem-resistant A. baumannii. Both the Center for Disease Control (CDC) and the World Health Organization (WHO) have ranked carbapenem-resistant A. baumannii (CRAB) as a high-priority antibiotic-resistant pathogen to tackle.8–12
The main mechanisms in carbapenem resistant Acinetobacter species are, (i) the production of enzymes (eg, carbapenemase) which hydrolyze these drugs and non-enzymatic mechanisms which include modification in the porins expression or alterations in the gene encoding porins, OMP (Caro, OprD) that result in loss or defects in porins and limit the drug entry into periplasmic space where PBPs are located, (ii) new or altered structure of penicillin-binding proteins (PBPs) that decrease the affinity of β-lactam to PBP and, (iii) overexpression of drug efflux pumps.13–15 Degradation of β-lactam by β-lactamase enzymes is the most prevalent drug resistance mechanism in A. baumannii.16 But the combination of different drug resistance mechanisms can be observed in the same microorganism. It is important to note that this organism undergoes rapid genetic mutation and acquires foreign antibiotic-resistant determinants by mobile genetic elements and thus expands the spectrum of resistance.17–21 The global spread of resistant organisms because of the exchange of different resistance determinants was possible with the increase of tourism and human migration in the recent era. Therefore, knowing the epidemiology and change in the genetic pattern of the organism helps to understand this organism better.17–23
In this review, we performed a systematic search of the literature to discuss the various mechanisms of carbapenem resistance, especially gene encoding carbapenemases in A. baumannii, to highlight the various mobile genetic elements in A. baumannii and their role in the spread of carbapenemase genes and to review the prevalence of various carbapenemase encoding genes from 2019–2021 in various parts of the world to understand the present situation.
Materials and Methods
Search Strategy with Inclusion and Exclusion Criteria
Articles for the incidence and dissemination of various carbapenemase genes in A. baumannii were searched for in Pubmed for the years 2019–2021 using the Boolean query “carbapenem-resistant genes and Acinetobacter baumannii”. The search of articles was restricted by selecting full and free full text articles for these years.
Review and original articles were also searched for using various word combinations for Acinetobacter baumannii carbapenemase genes, their historical perspectives, mobile genetic elements and mechanism of carbapenem resistance from the Google and Google Scholar search engines.
All those articles found relevant to our review were also included in the study. Articles showing the isolation of only one or two carbapenemase genes (OXA-51 and OXA-23), tknown to be common to Acinetobacter baumannii, were excluded from the study.
The presence of various carbapenemase genes was also searched for from website: https://www.ncbi.nlm.nih.gov/pathogens/isolates#/refgene/BETA-LACTAM.
Data Extraction
Information from various searched studies extracted and included in the present study are the prevalence of various carbapenemase genes, presence of mobile genetic elements, geographical location and year of isolation. For historical perspective the type/site of infection in patients and samples from which they were isolated was also added wherever a search was found. Data identification was done by first author of studies and year of publication. Various types and subtypes of carbapenemase genes present in A. baumannii were also included from above mention links.
Data Analysis
Findings from various studies is structured in two tables provided and also summarized under various headings and subheadings below.
Mechanisms of Carbapenem Resistance
Non-Enzymatic Mechanisms of Resistance
Penicillin-Binding Proteins (PBPs)
Mutations that change the production level or decrease the binding affinity of PBPs lead to resistance but the mechanism is associated with only low-level carbapenem resistance.24 In a study from Spain, A. baumannii isolated from the blood showed the production of oxacillinases and the lack of PBP2 were the most frequently observed mechanisms of resistance to carbapenems.25 In a study from Germany, the Im R clone displayed a significantly diminished expression of all PBPs except the 24kD protein. There was no saturation of this PBP, even with a higher concentration of imipenem, explaining the low affinity of imipenem to this target protein.26
Porin Channels and Outer Membrane Proteins (OMPs)
Various studies identify the differential expression of OMPs in antibiotic-resistant A. baumannii strains. OmpA imparts drug resistance to A. baumannii by allowing the slow diffusion of negatively charged beta-lactam antibiotics. OprD downregulation was observed in multidrug-resistant and pan-drug-resistant A. baumannii.27 Decreased membrane porin density (Omp22–23, Omp43, Omp44, Omp47, Omp33–36, Omp37 and CarO) was associated with pan-drug-resistance in A. baumannii.28 Several other OMPs in A. baumannii associated with carbapenem non-susceptibility are CarO or carbapenem associated protein, HMP-AB (heat-modifiable protein in A. baumannii) and OmpW.29 Various examples of the reduced number of porin channels and poor expression of genes resulting in porin loss or efficacy have been described in the review by Bonomo and Szabo.30
Efflux Pumps
There are various families of efflux pumps; ABC (ATP-binding cassette) transporter family, RND (resistance nodulation-cell division), MATE (multidrug and toxic compound extrusion), MFS (major facilitator superfamily), SMR (small multidrug resistance). The ABC family mainly exists in Gram-positive bacteria. The CraA, AmvA, CmlA, MdfA, and Tet pumps (part of the MFS family) have been identified in A. baumannii. In A. baumannii for an increase in the MIC of antibiotics, AbeM is the only efflux pump in the MATE family. For some antibiotics, AbeS (part of the SMR family) is a low-level resistance pump in the A. baumannii. AdeABC, AdeDE, AdeIJK, AdeXYZ, and AdeFGH efflux pump, which is associated with resistance to carbapenem, belongs to the RND family prevalent in the Acinetobacter species. The adeABC is the most studied pump gene that has a strong association with carbapenem resistance. It is composed of AdeA — membrane fusion protein; AdeB — inner membrane protein channel; and AdeC — outer membrane protein channel. Overexpression of adeABC is managed by genes -sensor kinase, adeS, and response regulator adeR. Overexpression of adeABC occurs due to mutation of adeRS or the insertion sequence of ISAba1 element upstream of the adeABC operon in A. baumannii. adeABC include β-lactams, fluoroquinolones, tetracycline (tigecycline), macrolide (linamides) and chloramphenicol, aminoglycosides.29–32 Another RND-type efflux pump in A. baumannii is AdeIJK, which is regulated by AdeN, a TetR-type regulator. Located upstream from the AdeFGH operon, AdeL, a LysR-type transcriptional regulator transcribed in the opposite direction and found to be responsible for the overproduction of AdeFGH.33 A study from China showed the overexpression of AdeABC efflux pump genes was closely associated with carbapenem (meropenem) resistance in A. baumannii.34 A study from South Korea also showed that the expression of the AdeABC efflux pump is an important resistance determinant in obtaining antibiotic resistance to the carbapenem group in A. baumannii.32
Enzymatic Mechanism of Resistance
Beta-lactamase enzymes are categorized into four classes based on sequence motifs and different hydrolytic mechanisms. Carbapenems are hydrolysed by Class A, B and D enzymes. Carbapenemases are classified according to their active sites into two groups, (i) serine carbapenemases and (ii) metallo-b-lactamases (MBLs).35–37
Class A Carbapenemases
In these active sites of enzymes contain serine. These serine-β-lactamases can be inactivated by β-lactamase inhibitors like clavulanic acid and tazobactam.37
Class B Carbapenemases
These β-lactamases are MBLs having Zn or another heavy metal in their catalytic site. Metallo beta-lactamase enzymes require a water molecule and a zinc ion to trigger and disrupt the Beta-lactam ring of the drug. These enzymes, like serine-β-lactamases, are not inhibited by clavulanate, sulbactam, or tazobactam; instead they are inhibited by metal ion chelators like ethylenediamine tetraacetic acid (EDTA), 1,10-o-phenanthroline or dipicolinic acid. Although MBL-type carbapenemases are less common than OXAs, their hydrolytic activity is 100–1000 times more potent.35–40
Class D Carbapenemases
Genes encoding oxacillinase enzymes (OXAs) are the main cause of carbapenem resistance in Acinetobacter baumannii. These enzymes are serine-dependent; usually not inhibited by clavulanic acid, sulbactam, and tazobactam whereas in vitro their activities may be inhibited at a concentration of 100 mM by sodium chloride (NaCl).35,40
Genes Encoding Carbapenemases in A. baumannii
Increased carbapenem resistance in Acinetobacter baumannii has been primarily driven by acquisition of resistance determinants and activation of intrinsic resistance mechanisms such as the chromosomal β-lactamases, blaOXA-51-like.41 A. baumannii carbapenem resistance is mainly due to the Ambler class D (OXA-type) carbapenemases and Ambler class B (Metallo-beta-lactamases) carbapenemase and less frequently with Ambler class A.39,42 As of 5 June 2022, 370 blaOXA-51 variants, 46 blaOXA-23 variants, 24 blaOXA-134 variants,12 variants of blaOXA-24, 9 blaOXA-143, and 7blaOXA-58 variants of β-lactamases have been identified (https://www.ncbi.nlm.nih.gov/pathogens/isolates#/refgene/BETA-LACTAM, https://www.ncbi.nlm.nih.gov/nuccore/MK682761.1) In A. baumannii class D carbapenemases (oxacillinases), class B (metallo-β-lactamases), and class A (β-lactamases) collected from various published studies and in the above link are shown in Table 1. OXA-48 variants are widespread in the Enterobacteriaceae family but reported from A. baumannii as well.43
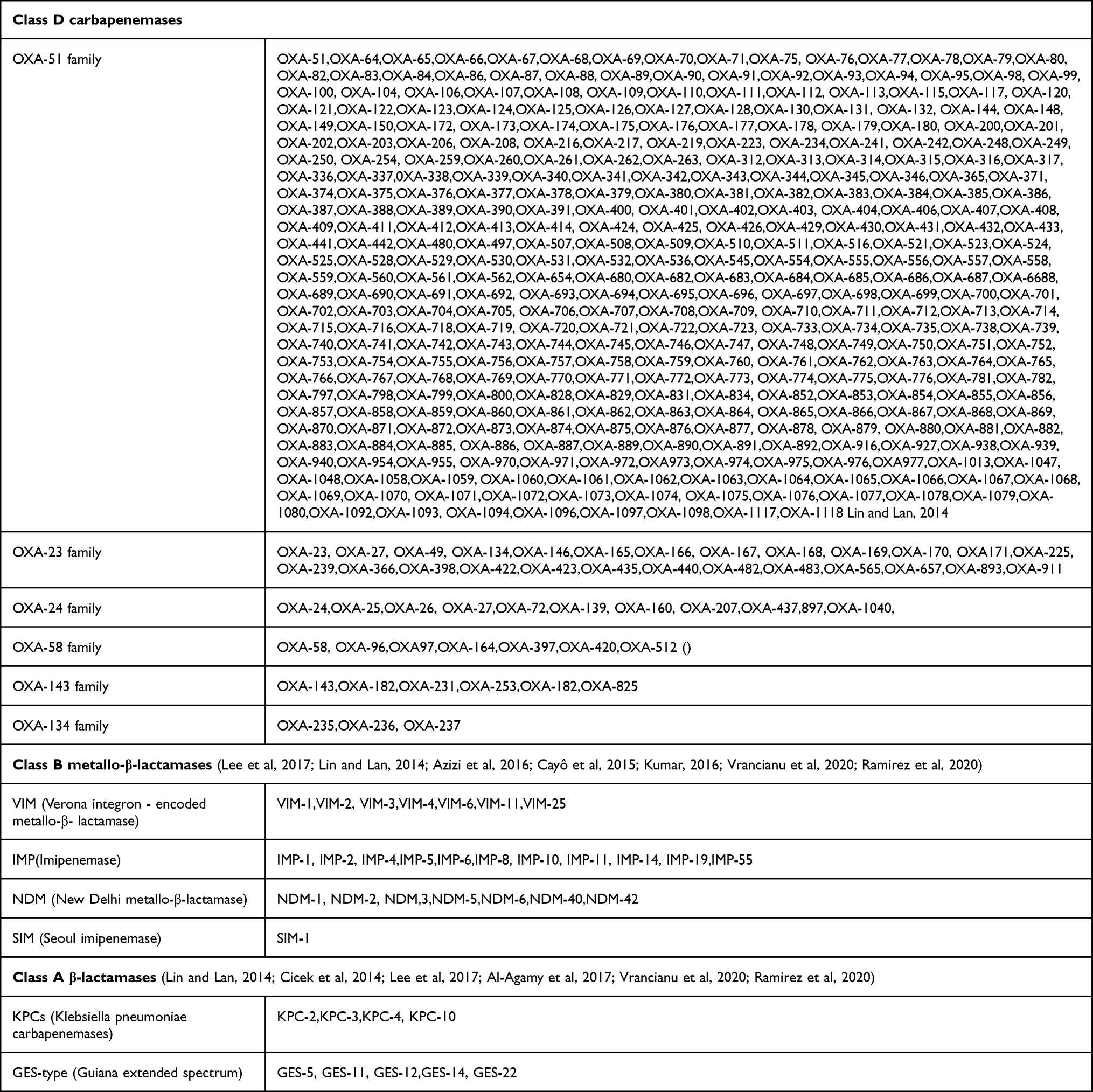 |
Table 1 Carbapenemases in A.baumannii (Higgins et al, 2010; Lin and Lan, 2014; Cicek et al, 2014; Evans and Amyes, 2014; Cayô et al, 2015; Azizi et al, 2016; Kumar, 2016; Lee et al, 2017; Al-Agamy et al, 2017; Vrancianu et al, 2020; Ramirez et al, 2020) |
Role of Mobile Genetic Elements (MGEs) in the Spread of Drug Resistance
Genome plasticity mechanisms include mutations and mobile genetic elements such as R Plasmids, insertion sequence, composite transposons, integrons, and resistance islands which result in the dissemination of antimicrobial resistance determinants encoding carbapenemase enzymes.44,45 The simplest form of mobile genetic elements (MGEs) are insertion sequences (IS) which are present in all domains of life.46 Same insertion sequences present on both sides of an antibiotic resistance gene form a compound transposon.39 Intercellular mobility of these MGEs occurs through self-replicative plasmids encoding genes for conjugative transfer and with a conjugative transposon.31
Insertion Sequences
In A. baumannii, the genes with resistant determinant encoding carbapenemases enzymes are regulated by the upstream presence of insertion sequences (IS): ISAba1 (IS4 family), ISAba2 (IS3 family), ISAba3 (IS1 family), ISAba4 (IS982 family), IS1008, ISAba125 (IS30 family), ISAba9, ISAba10 and IS18 (IS30 family), and ISAba825 (IS982 family). Due to the presence of promoter regions in the insertion sequence or after the insertion event the formation of a new promoter region, insertion sequences perform a major role to activate or increase the expression of downstream resistant genes and result in increased resistance to carbapenems.38,39,42,47
ISAba1 plays a major role in the carbapenem resistance gene transfer and expression in A. baumannii. This insertion sequence association with blaOXA-51-like, blaOXA-23-like, and blaOXA-58-like is reported in many studies.38,39,42 In one study the author reported the presence of ISAba1 in A. lwoffii which demonstrates the mobility of this IS in this genus.48 In A. baumannii isolates, the insertion ISAba2, ISAba3, and ISAba4, IS18 elements have also been identified upstream to blaOXA-58-like and blaOXA-23-like genes in many studies. The study described the novel ISAba4 element upstream to blaOXA-23 gene in A. baumannii.49 In a study from Egypt, insertion sequence ISAba1 was detected in all tested isolates, while ISAba2 was detected in 2.7% and ISAba3 was detected in 4% of isolates.15 A study done in Tehran, Iran revealed ISAba1 (45.1%) and ISAba4 (12.9%) were detected upstream of blaOXA-23-like genes of isolates.50 The presence of ISAba1 upstream to blaOXA-51-like genes was seen in 32.2% of all isolates. In a study from Vietnam, PCR amplification of upstream and downstream sequences of blaOXA-58 gene revealed the ISAba3 presence at both locations in one multidrug-resistant isolate that showed high resistance to imipenem due to overexpression of the blaOXA-58 gene and very high periplasmic β-lactamase activity.24 In a study of Weiyuan Wu, S1008-ΔISAba3 was the most common IS upstream of the blaOXA-58-like gene in A. baumannii clinical isolates, followed by ISAba3 and ISAba1. All blaOXA-23-like genes but one had an upstream insertion of ISAba1. The insertion of IS1008 (IS6 family) provided a hybrid promoter and increased the transcription level of the blaOXA-58 gene.47 The study detected, the ISAba825-ISAba3-like hybrid promoter upstream of blaOXA-58 leading to carbapenem resistance.51 A study from France identified an ISAba10, 1203 bp novel insertion sequence. This element was found to be inserted into the ISAba1 element upstream blaOXA-23 gene in an A. baumannii to provide an additional promoter sequence showing high minimum inhibitory concentrations (MICs) to carbapenems.52 ISs such as ISAba1, ISAba10, ISAba825, and ISAba125 elements with the spread of resistance are also involved in the disruption of the carO gene that codes for an important outer membrane channel and participates in the influx of carbapenem drug in A. baumannii. The absence of this outer membrane protein in isolates has been correlated with reduced carbapenem susceptibility.52,53
Transposon
Composite transposons have resistance genes in their center and are flanked by insertion sequences on both ends.39 Several studies reported that the blaNDM gene is located between two copies of the ISAba125 element, forming a composite transposon named Tn125.54,55 In a study from India, two clinical isolates were collected (ACN21 from Sir Ganga Ram Hospital, Delhi, and CIAT758 from Tata Medical Center, Kolkata) and of blaOXA-23, blaNDM-1 and blaOXA-58 genetic arrangement identified in complete genomes. In the ACN21 genome, the variant of the blaOXA-58 gene, blaOXA-420, and blaNDM-1 gene was present. The blaOXA-420 gene is bracketed by two elements of ISAba3 and forms a composite transposon. blaNDM-1 gene with downstream ISAba125 and upstream ble-MBL, bracketed by two copies of ISAba125 but additional insertion sequence, ISAba14 was identified upstream ATP binding protein and forming a Tn125-like composite transposon. The CIAT758 genome harbors blaOXA-23 on the chromosome and are bracketed by two copies of ISAba1 and bla OXA-58 gene with two copies of ISAba3. Downstream of ISAba3 possesses another insertion element, IS1008.56,57 A study described how Tn125 composite transposon in NDM-1 producing isolates disrupted by IS26 might potentially mobilize gene blaNDM-1.58
Transposons Tn2006, Tn2007, Tn2008, Tn2008B, and Tn2009 harboring blaOXA-23 have been reported in A. baumannii. In Tn2007, ISAba4 promoter upstream gene blaOXA–23. In comparison other transposons carry ISAba1, in Tn2006 and Tn2009, blaOXA–23 genes have been reportedly flanked on both sides by ISAba1 and named as compound or class 1 transposons. But in Tn2006, two ISAba1 copies inversely oriented compared to the Tn2009 were present in the same direction. Tn2008 and Tn2008B have only one copy of ISAba1 upstream to OXA-23 and lack the second copy of ISAba1. These transposons share a common region “blaOXA−23- ΔATPase”. Tn2006, Tn2008, and Tn2009 all found in the conjugative plasmids. Transposon Tn2007 is immovable and not considered a transposon. Tn2006 and Tn2008 are reported to be globally disseminated.59,60 One study identified, the blaOXA-23 gene in carbapenem-resistant A. baumannii was carried by Tn2009 (54%), Tn2006 (44%), and in Tn2008 (1.6%).61 A study showed the blaNDM genes in a novel Tn7 family transposon designated Tn6924 or its variants which indicates the significance of this transposon in spreading resistance genes.62
Integrons
Integrons are immobile but can be transferred through mobile genetic elements e.g. plasmids or transposons. Integrons are composed of an intI gene encoding an integrase, proximal primary recombination site attI, and a strong promoter gene, where mobile gene cassettes, carriage efflux pump genes, and drug resistance determinants, can be acquired or excised by a site-specific recombination mechanism catalyzed by the integrase. Various gene cassettes in integrons rearrange under antibiotic selective pressure.44 Classification of integrons based on the amino acid sequence of the IntI integrases. Common types of integrons in A. baumannii are the transportable class I (Tn402 derivatives) integron, followed by class II (Tn7 family) and class III integrons. Among these, the class 1 integron has the high prevalence in A. baumannii followed by class II.44,63,64 Resistance to carbapenem by metallo beta-lactamases has been reported by integrons in A. baumannii in many studies from parts of the world.64–66 In one study, Class 1 and 2 integrons were detected in 74.1% and 12.5% of isolates respectively, and Class 3 integrons were not detected among these isolates. Various gene cassettes in Class 1 integron-carrying strains (ampC, aacA4-catB8, ISAba1-blaOXA-23-GES-14, aadA2-cm1A6-GES-14-qacF, VIM-25-GES-24-qacF, dfrA5-ISAba1-blaOXA-51-blaOXA-40 and aadA2-GES-11-IMP-1) and class 2 integron-bearing A. baumannii strains (IMP-4, VIM-2-VEB-aacA4 and dfrA2-sat-2-aadA4) were observed.44 In a study from Korea, MBL gene, blaSIM-1 was detected in class 1 integrons borne gene cassette among seven clinical isolates of A. baumannii.52 In a study from Iran class I integrons and II integrons were detected in 63.9% and 78.2% of A. baumannii isolates respectively and 49.6% had both classes of integron genes. In any of the isolates, class III integron was not detected. All of the isolates positive for integron were resistant to the drugs imipenem, meropenem, and ceftriaxone.67 A study from Iran detected the most prevalent gene cassette arrays class 1 integron carrying isolates were blaIMP-19, aacA31, blaOXA-21, aadA-1 and blaVIM-1, qacED-1.65 In a study from South India, Class-1 integrase genes were found in all strains of A. baumannii whereas class 2 integrase genes were found in 6% of isolates.68 A study from Uganda found, Class 1 integrons in 62% of CRAB isolates.69 The importance of integrons in a clinical setting is that frequent use of a single antibiotic lead to overexpression of various antibiotic resistance gene determinants due to the presence of a common promoter.29
AbaR-Type Genomic Resistance Islands
Carbapenem resistance can also be acquired in A. baumannii by AbaR-type genomic islands that can harbor multiple antimicrobial resistance determinants. AbaRs are transposons that incorporate themselves into the chromosomal comM genes, encoding an ATPase domain, and play as important vehicles for acquiring antimicrobial resistance genes.70 In 2006, the first A. baumannii resistance island (AbaR1) was described in the multidrug-resistant AYE A. baumannii strain from France with a large genomic island of 86kbp in the chromosome in which there was a cluster of 45 resistance genes.71 In A. baumannii strains there have been many resistant islands described so far including AbaR1, AbaR3, AbaR4, AbaR5-Aba19, AbaR25, and others.72 In A. baumannii, different backbones are usually found in different epidemic clones. The AbaR3-type resistant island found in Global Clone 1 (GC1) has been mapped to the Tn6019 backbone and invariably associated with Tn6018 or its elements with multiple antimicrobial resistance regions (MARRs). AbaR4-type islands are mostly found in Global Clone 2 (GC2), Tn6022 is the predominant backbone element and sometimes bears the blaOXA-23 gene (Bi et al, 2020). Plasmids are also reported to carry AbaRs which can be exchanged both intra- and interspecies.73 AbGRI1-type islands have been identified in GC2, originate from a plasmid-borne ancestral form -AbGRI1-0, and consist of a Tn6022, a Tn6172, and a plasmid-borne fragment between the two linker transposons as a transposable unit.74 Acinetobacter baumannii strain AbaR25 was linked to an international clone (a variant of AbaR4) that carried blaOXA-23-like carbapenemase gene within the resistance island.72
Historical Perspectives
Increasing antibiotic resistance in Acinetobacter has been noticed in various surveys since 1975. Before that Acinetobacter infections could be easily treated with a single or combination of antibiotics.75 By the year 1998, the carbapenem-hydrolyzing beta-lactamases were identified throughout the world in clinical isolates of Acinetobacter species.76
Class D β-Lactamases (OXAs)
The first reported case of the OXA-Beta-lactamase enzyme with potent carbapenemase activity was plasmid-encoded and described by Panton et al in 1993.77 The gene was detected from a multidrug-resistant A. baumannii isolated from the blood culture of a patient from Edinburgh, Scotland in 1985, and the same year imipenem was approved for its first use.77 It was named first as ARI-1 (Acinetobacter resistant to imipenem) and renamed after sequencing as OXA-23. blaOXA23-like gene was discovered on the chromosome of Acinetobacter resistans which was the natural source of this enzyme and was mobilized to A. baumannii through ISAba1, insertion sequence provided by A. baumannii.39,43,76,78 The blaOXA-23 gene can be located either on the chromosome or on plasmids.76,78 These genes have been found in many species of Acinetobacter as well as members of the Enterobacteriaceae family.76 OXA-27 and OXA-49 variants of OXA-23 were identified in CRAB from Singapore and China respectively.79,80 OXA-423, a new variant of OXA-23 in A. baumannii was reported from China in 2020 from a sputum sample isolate collected in 2013.81 An extremely drug-resistant (XDR) A. baumannii AB030 isolate from the IC5 lineage, isolated in Canada showed resistance to fluoroquinolones, carbapenem, aminoglycosides, and tigecycline. Carbapenem resistance was due to the presence of blaOXA-65 and two copies of blaOXA-23. In AB030 both copies of blaOXA-23 are flanked by ISAba1 insertion sequences.82 A study identified the strains with co-occurrence of resistance genes on the chromosome and plasmids occurring with blaOXA-23 and showed that the presence of multiple copies and locations had no significant effect on the susceptibility to carbapenems and most antibiotics.83
The largest group of OXA beta-lactamases mentioned to date are the OXA-51-like. OXA-51/69-like beta-lactamase is mentioned as intrinsic, a naturally occurring chromosomal enzyme in A. baumannii.43 The study described the emergence of a novel carbapenemase (OXA-51) in genetically distinct carbapenem-resistant clinical isolates of A. baumannii from Argentina. OXA-51 presence, in different clones of A. baumannii, obtained from three hospitals within Buenos Aires indicates that the horizontal transfer of the encoding gene had occurred.84 However, plasmids harboring ISAba1 – blaOXA-51-like have been detected in A. baumannii, Acinetobacter nosocomialis and Acinetobacter pittii.85,86 blaOXA-51-like genes, which were thought to reside exclusively in Acinetobacter species but also found in members of the Enterobacteriaceae family.87 This affects the accuracy of using blaOXA-51-like detection as a tool for differentiating A. baumannii from other Acinetobacter species and non-Acinetobacter Gram-negative genera.
OXA-24/40 B-lactamase was first identified from the chromosome of A. baumannii isolated from a bronchial aspirate of a patient admitted to the Medical Intensive Care Unit in the Ramóny Cajal Hospital, Madrid, Spain in 1997.88 The first representative to be isolated from this group was OXA-24, subsequently renamed OXA-40 after sequencing which showed that these were identical enzymes.89 However, now the gene OXA-24 carrying plasmids in carbapenem-resistant Acinetobacter baumannii has been identified worldwide. OXA-25 and OXA-26 variants were identified in Spain and in Belgium respectively, from isolates collected between 1995 and 1997.79 The blaOXA-72 gene was first reported in Thailand from an Acinetobacter strain (GenBank accession no. AY739646) in 2004.90 The OXA-58 gene was identified from the plasmid of a strain isolated from a 24-year-old female with a skin burn infection at the Rangueil University Hospital (Toulouse, France) in August 2003. They cloned and characterized a novel carbapenem-hydrolyzing oxacillinase, OXA-58.91 The OXA-58 gene is associated with hospital outbreaks in Europe, the USA, South America, Australia, and Africa.89,92–96 A novel plasmid-borne carbapenemase gene, OXA-143 identified from an A. baumannii strain isolated from blood culture in the Brazilian Intensive care unit in 2004.97 A carbapenem-resistant A. baumannii (strain Ac-141) was isolated from the urine culture of a patient admitted to a tertiary teaching hospital located in southern Brazil, 2007 and sequencing of the blaOXA-143-like gene identified a single mutation. This new OXA-143 variant was named OXA-231.98 Three novel class D β-lactamase, OXA-235, OXA-236, OXA-237 were reported in 2013, from the plasmids of A. baumannii strains isolated from the United States and Mexico between 2005 and 2009.99 The first report of OXA-48-like producing A. baumannii from a nursing home resident with faecal colonization was in northern Portugal.100
Class B β-Lactamases (MBLs)
IMP-1 was described in Acinetobacter strains collected between 1998–2001 in the clinical microbiology laboratory of the São Paulo Hospital, Brazil. This study showed the first occurrence of IMP-1 from collected isolates in 1998, when 29% of tested isolates carried the blaIMP gene but from January 1999 to December 2001 all CRAB strains isolated in the hospital carried IMP-1 showing the emergence of this mechanism of resistance in hospital.101 The IMP-2 variant after sequencing was first reported in a clinical isolate Acinetobacter baumannii AC-54/97 from Italy which was isolated in 1997 from an intensive care unit of a hospital.102 IMP-4, a novel metallo-beta-lactamase was identified from the isolates collected in Hong Kong between 1994 and 1998.103 A new variant of blaIMP (IMP5) inserted in a class 1 integron (named In76) was reported from Portugal from a clinical strain A. baumannii 65FFC isolated in 1998 from the urine of the patient.104 IMP-6's first appearance in A. baumannii A3227 isolated from the tracheal secretion of a patient, reported from Brazil.105 IMP-8 was reported in A. phenon 6/ct 13TU, from China.106 IMP-10 was reported from two A. baumannii strains isolated from blood and bronchoalveolar lavage (BAL) samples of two different patients collected in São Paulo, Brazil, in 2000.107 IMP-11 (accession no. AB074436) in Japan IMP-19 reported in A. baumannii from Japan.108 A. baumannii strain 56 recovered from tracheal aspirate sample from an adult ICU, Iran showed MBL activity, and analysis of IMP sequence identified a novel allelic IMP-55 variant.109 The first reported VIM-2 enzyme found on a Class 1 integron of strains collected in 1998 and 1999 was from respiratory samples in Seoul, Korea.110 The first report of the VIM-1 determinant in A. baumannii in the world was from Greece, from the isolates collected from March 2004 to March 2005.111 The first VIM-4 MBL determinant was a part of a class 1 integron reported in Acinetobacter species isolated from a sputum sample collected between 2001 and 2006 in Greece.112 First description of the blaVIM-11 gene spreading among A. baumannii strains in southern Taiwan was recovered between 2002 and 2006. blaVIM-11 was associated with a class 1 integron, the study also reported the presence of VIM-3 in A. baumannii.113 SIM-1 is the only variant of the SIM enzyme reported in A. baumannii and this new MBL determinant, was reported from isolates collected from 2003–2004 sputum and urine samples in Seoul, Korea. The SIM-1 variant in this study was found encoded by gene cassette associated with a class 1 integron.114 The blaNDM-1 report with the coexistence of the blaOXA-23 gene and armA gene in A. baumannii was reported from patients admitted to intensive care units in Chennai, India.17 However, the detection of NDM-1 in A. baumannii 161/07 in Germany in the year 2007 in transposons structure which was composed of two copies of insertion sequence ISAba125 suggests that NDM-1 in A. baumannii might be disseminated earlier than it was thought.54 NDMs variant NDM-2 was detected from the isolate from a central venous line catheter reported from Egypt.115 A new variant blaNDM-3 from multidrug-resistant A. baumannii isolated from a patient with burn injuries from India was reported.116 NDM-6 in an Acinetobacter baumannii isolated from urine from northern Spain.117 First report of the existence of blaSPM-1 among the clinical isolates of A. baumannii was from Iran.118 Genes, including SPM (São Paulo metallo-β-lactamase) and GIM (German imipenemase), were reported in A. baumannii in many studies.118–120
Class A β-Lactamases
KPC-2, KPC-3, KPC-4, and novel variant KPC-10 were reported from the A. baumannii isolates collected from January to May 2009 from 17 hospitals in Puerto Rico.121 GES-11, a new extended-spectrum-lactamase conferring reduced susceptibility to carbapenem was detected as a part of a class 1 integron in Acinetobacter baumannii BM4674 isolated from the tibia fracture of a patient, in France in 2008.122 Plasmid-located blaGES-11 and blaGES-12 variants and a likely chromosomally located blaGES-14 novel variant reported from the isolates of A. baumannii collected between January 2008 to December 2009 from the hospitals of Belgium.123 GES-5 conferring carbapenem resistance in A. baumannii was reported from the isolates collected from January to December 2010 from patients admitted to a tertiary hospital in Riyadh, Saudi Arabia.124 The first report of the existence of blaSPM with GES-1 beta-lactamase was from Iran in A. baumannii.118
Presence of Various Carbapenemase Genes in Different Countries
From a nationwide surveillance study from Thailand MDR A. baumannii harboured blaOxa23 (85.8%), blaOXA58(7.9%), blaOXA-40(1%), blaVIM (2.9%), and blaIMP (2%), and in XDR A. baumannii detected blaOXA23(93%), blaOXA 58 (3.9%), blaVIM (1.3%), and blaIMP(0.8%).125
Analysis of three years (2019–2021) of articles from Table 2 showed that after OXA-51-like genes, which are intrinsic to A. baumannii, OXA-23 was predominantly seen in most of the studies from various countries which is already known and also seen previously to these studies. Multilocus sequence typing (MLST) data reported that the OXA−23 gene, particularly associated with clone CC92, especially ST75, ST92, ST138 was spreading rapidly in China.126
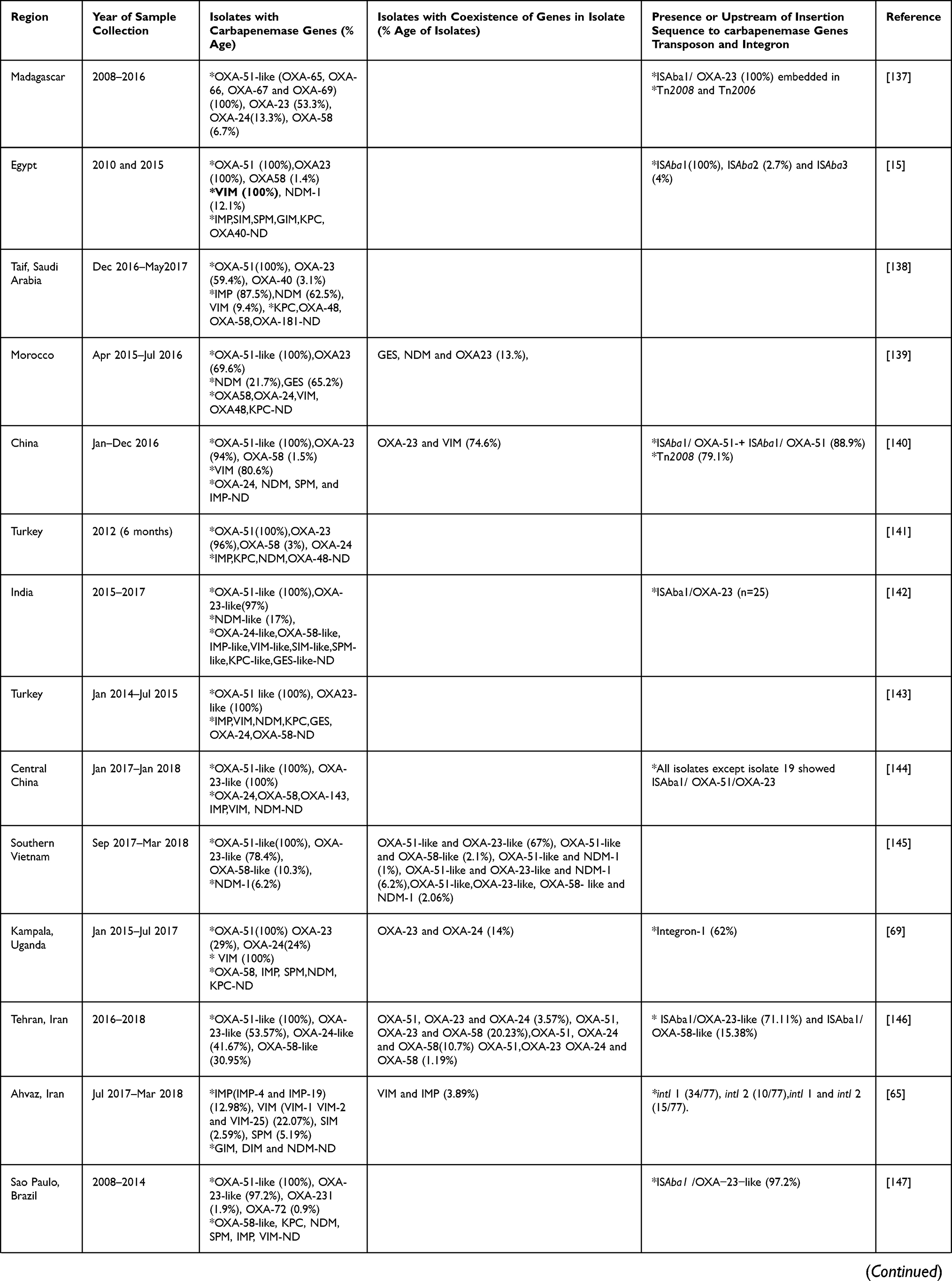 | 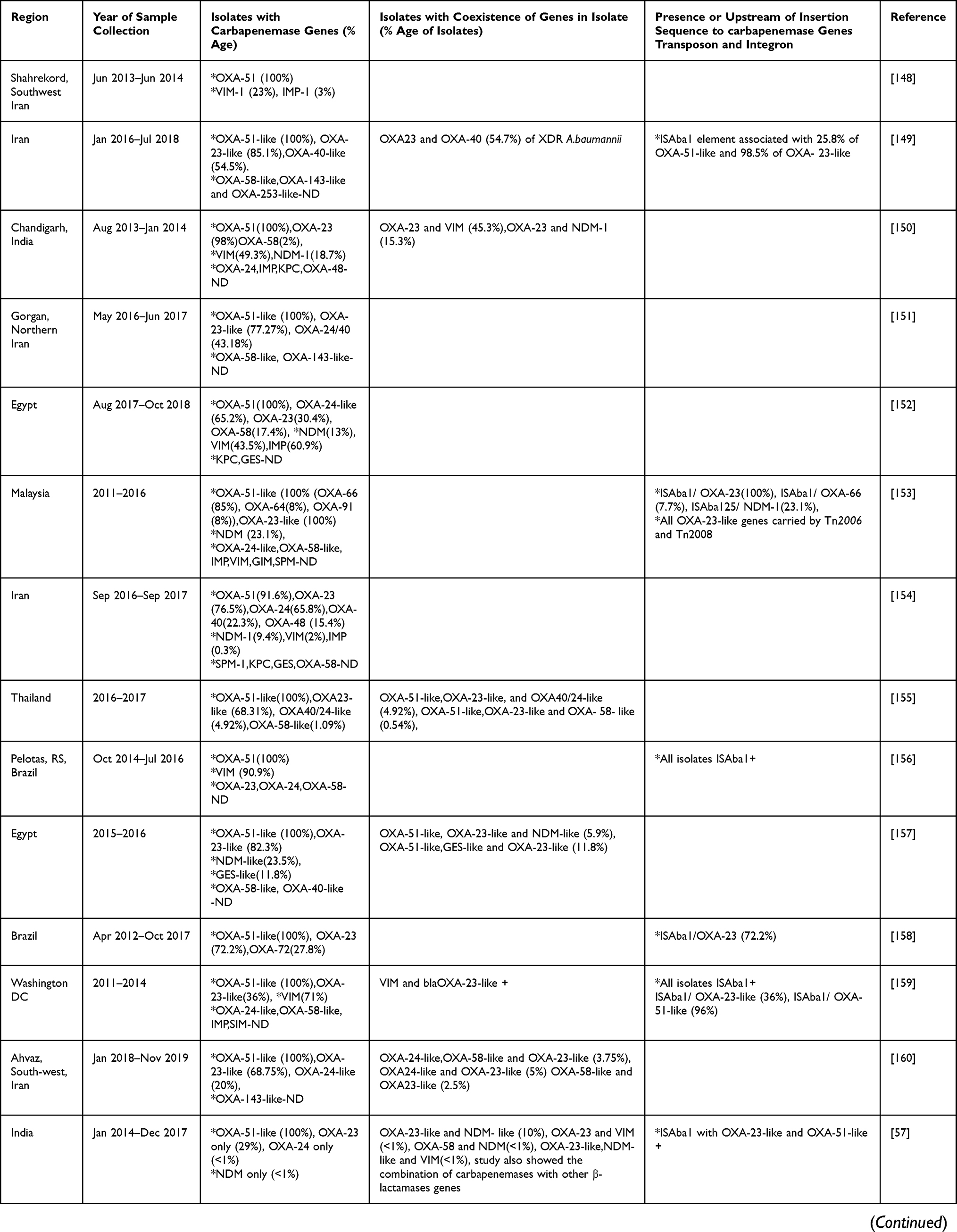 | 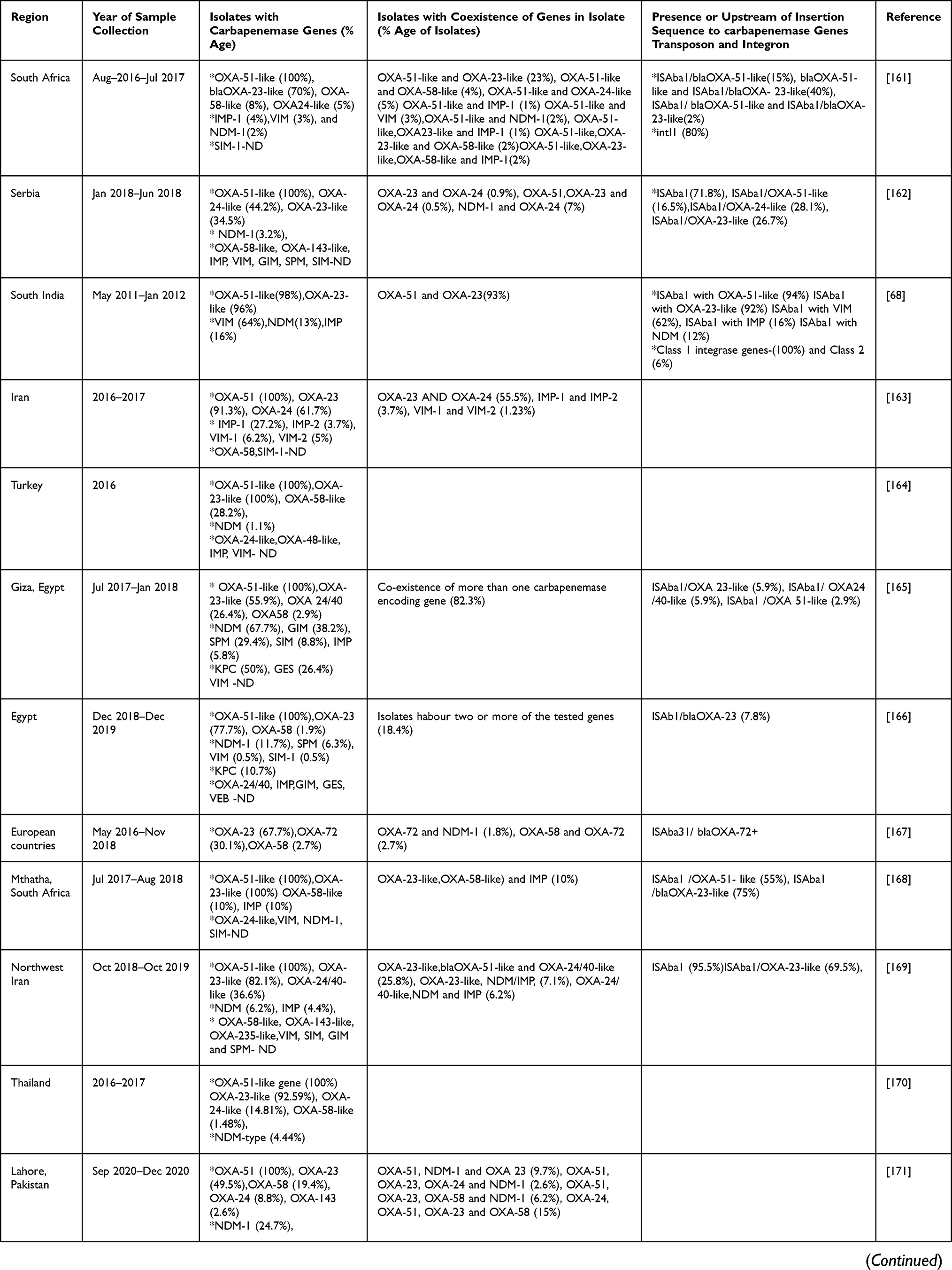 |  |
Table 2 Carbapenemase Genes Dissemination in A. baumannii from Various Countries in the Last Three Years (2019–2021) |
In studies from Egypt and Serbia the percentage of OXA-24-like exceeded the OXA-23-like percentage whereas in other studies OXA-24 percentage was either less than OXA-23 or not detected. In other studies, done in Egypt, China, Turkey, Vietnam, India (Chandigarh), South Africa, and Lahore, OXA-58-like was found in more isolates than OXA-24-like, while in most studies OXA-24-like genes were detected followed by OXA-23 or not detected. The studies where MBLs have been seen predominantly rather than OXA-23-like after OXA-51-like were from Saudi Arabia (IMP and NDM), Uganda (VIM), Brazil (VIM), Washington, DC (VIM), and Egypt (NDM). In a study from Egypt and Uganda, 100% of isolates showed the presence of VIM. In MBLs IMP, VIM, NDM were commonly identified in A. baumannii. Other carbapenemases identified were GES-type from the study of Morocco, SIM and SPM from Iran, and GIM, SPM, and SIM from Egypt. This highlights the predominance of OXAs other than OXA-23, MBLs and Class A carbapenemases in different regions also.
Limitation
The search was restricted to only the PubMed engine and Google for the prevalence of carbapenemase genes. In the present review, details of MLST clones were not included. The remaining search engines would have given more data to analyse and discuss which is not included in PubMed.
Strategies in Controlling the Dissemination of Resistant Determinants
Active antimicrobial stewardship program in hospitals needs the time to reduce the selective antibiotic pressure and the development of resistance to broad spectrum antibiotics. The use of various care bundles in Infection Control Practices (ICP) is needed to control the spread of these organisms among patients, staff, and patient attendants to become infected or colonized. A colonized individual with a drug resistant organism can further spread the organism to the community and on migration to various geographical regions which can complicate the present situation. Taking into consideration treatment options, in a retrospective (2015–2017) study from Chiang Mai University Hospital, Thailand conducted on critically ill CRAB infected patients, the author concluded that there is a reduction in 30-day mortality on addition of drug meropenem to colistin, high clinical and microbiological responses, and no difference in nephrotoxicity when compared to drug colistin monotherapy.127 The author also found in one of his studies that a loading dose of colistin methanesulfonate resulted in favourable patient outcomes with a better clinical, microbiological response and survival rate than a non-loading dose. But a loading dose of colistin methanesulfonate found with an increase in nephrotoxicity, therefore rigorous renal function monitoring is required when a loading dose of colistin is administered to the patient.128 The author also found increased nephrotoxicity when a loading dose of colistin is given in XDR-AB infected cancer patients but in comparison to the above study not resulted in significant better outcome comparing to nonloading dose.129 One more study conducted by the same author compared the combination of loading dose colistin with imipenem and loading dose colistin with meropenem for treatment of CRAB infected patients. He found that the loading dose of colistin and meropenem treatment gave a better survival rate, clinical and microbiological response, but no significant difference in nephrotoxicity while comparing the two combination regimens.130 In one scientific review, the author discussed Infection control precautions including hand hygiene, cohorting, and environmental decontamination, to control MDR A. baumannii outbreak and various treatment options.29 A study from Korea showed the efficacy of various combination regimens including drug colistin, sulbactam, minocycline, and tigecycline in the treatment of CRAB.131 Before treatment, the important thing to differentiate colonization from infection to avoid misuse of antibiotics and if carbapenem resistance genes are detected, confirming the in-vitro susceptibility need to be performed as many times genes do not express and it helps in treating the patient in case of pan-drug resistant and extensively drug resistant organism. At the same time, knowing the presence of genes in phenotypic carbapenem sensitive organisms, the doctor understands that the failure of a drug can occur during treatment due to some inducer and is able to choose the drug on a case by case basis.132 As some carbapenemases retain activity against beta-lactamase inhibitors, they can be used in combination with other agents for treatment. So knowing the type, presence and expression of the carbapenemase gene helps in ways to avoid therapy failure.
Conclusion
A. baumannii with various carbapenem resistance encoding genes is widespread worldwide and the rapid spread of resistance due to mobile genetic elements has declared this organism as a high- priority antibiotic-resistant pathogen and global threat in medical care. But as the situation is still not well documented certain parts of the world need to be studied from various regions to know more about the genetics of this drug resistant superbug.
It is observed from various studies that OXA-23 is the most frequently detected carbapenemase in the CRAB worldwide but recent studies have been reported with an increased prevalence of other carbapenemase genes in various regions over OXA-23 and the importance of various mobile genetic elements. There is the need for active surveillance to understand the exact situation in various countries, to track the spread, to start the appropriate controlling measures, to break the chain of this bug and, most importantly, to start timely proper treatment of patients based on the presence of various genes to avoid therapeutic failure and mortality.
Therefore, compiled genotypic systematic data have been prepared for the thorough knowledge of its molecular characterization with its global genetic diversity.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Li S, Duan X, Peng Y, Rui Y. Molecular characteristics of carbapenem-resistant Acinetobacter spp. from clinical infection samples and fecal survey samples in Southern China. BMC Infect Dis. 2019;19(1):1–12. doi:10.1186/s12879-019-4423-3
2. Barker J, Maxted H. Observations on the growth and movement of Acinetobacter on semi-solid media. J Med Microbiol. 1975;8(3):443–446. doi:10.1099/00222615-8-3-443
3. Gupta N, Gandham N, Jadhav S, Mishra RN. Isolation and identification of Acinetobacter species with special reference to antibiotic resistance. J Nat Sci Biol Med. 2015;6(1):159–162. doi:10.4103/0976-9668.149116
4. Hamidian M, Nigro SJ. Emergence, molecular mechanisms and global spread of carbapenem-resistant Acinetobacter baumannii. Microb Genom. 2019;5(10). doi:10.1099/mgen.0.000306
5. Ramirez MS, Bonomo RA, Tolmasky ME. Carbapenemases: transforming Acinetobacter baumannii into a yet more dangerous menace. Biomolecules. 2020;10(5):720. doi:10.3390/biom10050720
6. Harding CM, Hennon SW, Feldman MF. Uncovering the mechanisms of Acinetobacter baumannii virulence. Nat Rev Microbiol. 2018;16(2):91–102. doi:10.1038/nrmicro.2017.148
7. Perez F, Hujer AM, Hujer KM, Decker BK, Rather PN, Bonomo RA. Global challenge of multidrug-resistant Acinetobacter baumannii. Antimicrob Agents Chemother. 2007;51(10):3471–3484. doi:10.1128/AAC.01464-06
8. Mezzatesta ML, D’Andrea MM, Migliavacca R, et al. Epidemiological characterization and distribution of carbapenem-resistant Acinetobacter baumannii clinical isolates in Italy. Clin Microbiol Infect. 2012;18(2):160–166. doi:10.1111/j.1469-0691.2011.03527.x
9. Zhanel GG, Wiebe R, Dilay L, et al. Comparative review of the carbapenems. Drugs. 2007;67(7):1027–1052. doi:10.2165/00003495-200767070-00006
10. Codjoe FS, Donkor ES. Carbapenem resistance: a review. Med Sci. 2017;6(1):1. doi:10.3390/medsci6010001
11. Manchanda V, Sanchaita S, Singh NP. Multidrug resistant Acinetobacter. J Glob Infect Dis. 2010;2(3):291–304. doi:10.4103/0974-777X.68538
12. Said D, Willrich N, Ayobami O, Noll I, Eckmanns T, Markwart R. The epidemiology of carbapenem resistance in Acinetobacter baumannii complex in Germany (2014–2018): an analysis of data from the national Antimicrobial Resistance Surveillance system. Antimicrob Resist Infect Control. 2021;10(1):1–13. doi:10.1186/s13756-021-00909-8
13. Vashist J, Tiwari V, Das R, Kapil A, Rajeswari MR. Analysis of penicillin-binding proteins (PBPs) in carbapenem resistant Acinetobacter baumannii. Indian J Med Res. 2011;133(3):332.
14. Elshamy AA, Aboshanab KM. A review on bacterial resistance to carbapenems: epidemiology, detection and treatment options. Future Sci. 2020;6(3):FSO438. doi:10.2144/fsoa-2019-0098
15. Abouelfetouh A, Torky AS, Aboulmagd E. Phenotypic and genotypic characterization of carbapenem-resistant Acinetobacter baumannii isolates from Egypt. Antimicrob Resist Infect Control. 2019;8:185. doi:10.1186/s13756-019-0611-6
16. Handal R, Qunibi L, Sahouri I, et al. Characterization of carbapenem-resistant Acinetobacter baumannii strains isolated from hospitalized patients in Palestine. Int J Microbiol. 2017;2017. doi:10.1155/2017/8012104
17. Karthikeyan K, Thirunarayan MA, Krishnan P. Coexistence of bla OXA-23 with bla NDM-1 and armA in clinical isolates of Acinetobacter baumannii from India. J Antimicrob Chemother. 2010;65(10):2253–2254. doi:10.1093/jac/dkq273
18. Suresh A, Ranjan A, Jadhav S, et al. Molecular genetic and functional analysis of pks-harboring, extra-intestinal pathogenic Escherichia coli from India. Front Microbiol. 2018;9:2631. doi:10.3389/fmicb.2018.02631
19. Ranjan A, Shaik S, Nandanwar N, et al. Comparative genomics of Escherichia coli isolated from skin and soft tissue and other extraintestinal infections. mBio. 2017;8:e01070–17. doi:10.1128/mBio.01070-17
20. Shaik S, Ranjan A, Tiwari SK, et al. Comparative genomic analysis of globally dominant ST131 clone with other epidemiologically successful extraintestinal pathogenic Escherichia coli (ExPEC) lineages. mBio. 2017;8:e01596–17. doi:10.1128/mBio.01596-17
21. Mirza S, Savita jadhav RN, Das NK. Coexistence of β-lactamases in community-acquired infections in a tertiary care hospital in India. Int J Microbiol. 2019;2019:5. doi:10.1155/2019/7019578
22. Avasthi TS, Kumar N, Baddam R, et al. Genome of multidrug-resistant uropathogenic Escherichia coli strain NA114 from India. J Bacteriol. 2011;193(16):4272–4273. doi:10.1128/JB.05413-11
23. Jadhav S, Hussain A, Devi S, et al. Virulence characteristics and genetic affinities of multiple drug resistant uropathogenic Escherichia coli from a semi Urban locality in India. PLoS One. 2011;6(3):e18063. doi:10.1371/journal.pone.0018063
24. Nguyen AT, Pham SC, Ly AK, Nguyen CV, Vu TT, Ha TM. Overexpression of bla OXA-58 gene driven by IS Aba3 Is associated with imipenem resistance in a clinical Acinetobacter baumannii isolate from Vietnam. Biomed Res Int. 2020;2020:1–9. doi:10.1155/2020/7213429
25. Fernández-Cuenca F, Martínez-Martínez L, Conejo MC, Ayala JA, Perea EJ, Pascual A. Relationship between β-lactamase production, outer membrane protein and penicillin-binding protein profiles on the activity of carbapenems against clinical isolates of Acinetobacter baumannii. J Antimicrob Chemother. 2003;51(3):565–574. doi:10.1093/jac/dkg097
26. Gehrlein M, Leying H, Cullmann W, Wendt S, Opferkuch W. Imipenem resistance in Acinetobacter baumannii is due to altered penicillin-binding proteins. Chemotherapy. 1991;37(6):405–412. doi:10.1159/000238887
27. Uppalapati SR, Sett A, Pathania R. The outer membrane proteins OmpA, CarO, and OprD of Acinetobacter baumannii confer a two-pronged defense in facilitating its success as a potent human pathogen. Front Microbiol. 2020;11:589234. doi:10.3389/fmicb.2020.589234
28. Vrancianu CO, Gheorghe I, Czobor IB, Chifiriuc MC. Antibiotic resistance profiles, molecular mechanisms and innovative treatment strategies of Acinetobacter baumannii. Microorganisms. 2020;8(6):935. doi:10.3390/microorganisms8060935
29. Nguyen M, Joshi SG. Carbapenem resistance in Acinetobacter baumannii, and their importance in hospital‐acquired infections: a scientific review. J Appl Microbiol. 2021;131(6):2715–2738. doi:10.1111/jam.15130
30. Bonomo RA, Szabo D. Mechanisms of multidrug resistance in Acinetobacter species and Pseudomonas aeruginosa. Clin Infect Dis. 2006;43(Supplement_2):S49–S56. doi:10.1086/504477
31. Vrancianu CO, Pelcaru CF, Alistar A, et al. Escaping from ESKAPE. Clinical significance and antibiotic resistance mechanisms in Acinetobacter baumannii: a review. Biointerface Res Appl Chem. 2021;11(1):8190–8203.
32. Ju Y, Kim YJ, Chang CL, Choi GE, Hyun KY. Relationship between AdeABC Efflux pump genes and carbapenem in multidrug-resistant Acinetobacter baumannii. Biomed Sci Lett. 2021;27(2):59–68. doi:10.15616/BSL.2021.27.2.59
33. Zhang Y, Li Z, He X, et al. Overproduction of efflux pumps caused reduced susceptibility to carbapenem under consecutive imipenem-selected stress in Acinetobacter baumannii. Infect Drug Resist. 2017;11:457–467. doi:10.2147/IDR.S151423
34. Xu CF, Bilya SR, Xu W. adeABC efflux gene in Acinetobacter baumannii. New Microbes New Infect. 2019;30:100549. doi:10.1016/j.nmni.2019.100549
35. Kyriakidis I, Vasileiou E, Pana ZD, Tragiannidis A. Acinetobacter baumannii antibiotic resistance mechanisms. Pathogens. 2021;10(3):373. doi:10.3390/pathogens10030373
36. Ranjan A, Shaik S, Mondal A, et al. Molecular epidemiology and genome dynamics of New Delhi metallo-β-lactamase-producing extraintestinal pathogenic Escherichia coli strains from India. Antimicrob Agents Chemother. 2016;60:6795–6805. doi:10.1128/AAC.01345-16
37. Diene SM, Rolain JM. Carbapenemase genes and genetic platforms in Gram-negative bacilli: Enterobacteriaceae, Pseudomonas and Acinetobacter species. Clin Microbiol Infect. 2014;20(9):831–838. doi:10.1111/1469-0691.12655
38. Poirel L, Nordmann P. Carbapenem resistance in Acinetobacter baumannii: mechanisms and epidemiology. Clin Microbiol Infect. 2006;12(9):826–836. doi:10.1111/j.1469-0691.2006.01456
39. Pagano M, Martins AF, Barth AL. Mobile genetic elements related to carbapenem resistance in Acinetobacter baumannii. Braz J Microbiol. 2016;47:785–792. doi:10.1016/j.bjm.2016.06.005
40. Bush K, Jacoby GA. Updated functional classification of β-lactamases. Antimicrob Agents Chemother. 2010;54(3):969–976. doi:10.1128/AAC.01009-09
41. Djahmi N, Dunyach-Remy C, Pantel A, Dekhil M, Sotto A, Lavigne JP. Epidemiology of carbapenemase-producing Enterobacteriaceae and Acinetobacter baumannii in Mediterranean countries. Biomed Res Int. 2014;2014:1–11. doi:10.1155/2014/305784
42. Ibrahim ME. Prevalence of Acinetobacter baumannii in Saudi Arabia: risk factors, antimicrobial resistance patterns and mechanisms of carbapenem resistance. Ann Clin Microbiol Antimicrob. 2019;18(1):1–12. doi:10.1186/s12941-018-0301-x
43. Evans BA, Amyes SG. OXA β-lactamases. Clin Microbiol Rev. 2014;27(2):241–263. doi:10.1128/CMR.00117-13
44. Goudarzi H, Azad M, Seyedjavadi SS, et al. Characterization of integrons and associated gene cassettes in Acinetobacter baumannii strains isolated from intensive care unit in Tehran, Iran. J Acute Dis. 2016;5(5):386–392. doi:10.1016/j.joad.2016.08.004
45. Suresh A, Shaik S, Baddam R, et al. Evolutionary dynamics based on comparative genomics of pathogenic Escherichia coli lineages harbouring polyketide synthase (pks) island. mBio. 2021;12:e03634–20. doi:10.1128/mBio.03634-20
46. Vandecraen J, Chandler M, Aertsen A, Van Houdt R. The impact of insertion sequences on bacterial genome plasticity and adaptability. Crit Rev Microbiol. 2017;43(6):709–730. doi:10.1080/1040841X.2017.1303661
47. Wu W, He Y, Lu J, Lu Y, Wu J, Liu Y. Transition of bla OXA-58-like to bla OXA-23-like in Acinetobacter baumannii clinical isolates in southern China: an 8-year study. PLoS One. 2015;10(9):e0137174. doi:10.1371/journal.pone.0137174
48. Segal H, Garny S, Elisha BG. Is IS ABA-1 customized for Acinetobacter? FEMS Microbiol Lett. 2005;243(2):425–429. doi:10.1016/j.femsle.2005.01.005
49. Corvec S, Poirel L, Naas T, Drugeon H, Nordmann P. Genetics and expression of the carbapenem-hydrolyzing oxacillinase gene bla OXA-23 in Acinetobacter baumannii. Antimicrob Agents Chemother. 2007;51(4):1530–1533. doi:10.1128/AAC.01132-06
50. Bahador A, Raoofian R, Farshadzadeh Z, et al. The prevalence of ISAba1 and ISAba4 in Acinetobacter baumannii species of different international clone lineages among patients with burning in Tehran, Iran. Jundishapur J Microbiol. 2015;8(7). doi:10.5812/jjm.17167v2
51. Ravasi P, Limansky AS, Rodriguez RE, Viale AM, Mussi MA. ISAba825, a functional insertion sequence modulating genomic plasticity and bla OXA-58 expression in Acinetobacter baumannii. Antimicrob Agents Chemother. 2011;55(2):917–920. doi:10.1128/AAC.00491-10
52. Lee Y, Kim CK, Lee H, Jeong SH, Yong D, Lee K. A novel insertion sequence, IS Aba10, inserted into IS Aba1 adjacent to the bla OXA-23 gene and disrupting the outer membrane protein gene carO in Acinetobacter baumannii. Antimicrob Agents Chemother. 2011;55(1):361–363. doi:10.1128/AAC.01672-09
53. Mussi MA, Limansky AS, Viale AM. Acquisition of resistance to carbapenems in multidrug-resistant clinical strains of Acinetobacter baumannii: natural insertional inactivation of a gene encoding a member of a novel family of β-barrel outer membrane proteins. Antimicrob Agents Chemother. 2005;49(4):1432–1440. doi:10.1128/AAC.49.4.1432-1440.2005
54. Pfeifer Y, Wilharm G, Zander E, et al. Molecular characterization of bla NDM-1 in an Acinetobacter baumannii strain isolated in Germany in 2007. J Antimicrob Chemother. 2011;66(9):1998–2001. doi:10.1093/jac/dkr256
55. Bontron S, Nordmann P, Poirel L. Transposition of Tn 125 encoding the NDM-1 carbapenemase in Acinetobacter baumannii. Antimicrob Agents Chemother. 2016;60(12):7245–7251. doi:10.1128/AAC.01755-16
56. Vijayakumar S, Wattal C, Oberoi JK, et al. Insights into the complete genomes of carbapenem-resistant Acinetobacter baumannii harbouring bla OXA-23, bla OXA-420 and bla NDM-1 genes using a hybrid-assembly approach. Access Microbiol. 2020;2:8. doi:10.1099/acmi.0.000140
57. Vijayakumar S, Anandan S, Prabaa D, et al. Insertion sequences and sequence types profile of clinical isolates of carbapenem-resistant A. baumannii collected across India over four year period. J Infect Public Health. 2020;13(7):1022–1028. doi:10.1016/j.jiph.2019.11.018
58. Jones LS, Toleman MA, Weeks JL, Howe RA, Walsh TR, Kumarasamy KK. Plasmid carriage of bla NDM-1 in clinical Acinetobacter baumannii isolates from India. Antimicrob Agents Chemother. 2014;58(7):4211–4213. doi:10.1128/AAC.02500-14
59. Chen Y, Gao J, Zhang H, Ying C. Spread of the blaOXA–23-containing Tn 2008 in carbapenem-resistant Acinetobacter baumannii isolates grouped in CC92 from China. Front Microbiol. 2017;8:163. doi:10.3389/fmicb.2017.00163
60. Nigro SJ, Hall RM. Structure and context of Acinetobacter transposons carrying the oxa23 carbapenemase gene. J Antimicrob Chemother. 2016;71(5):1135–1147. doi:10.1093/jac/dkv440
61. Yoon EJ, Kim JO, Yang JW, et al. The bla OXA-23-associated transposons in the genome of Acinetobacter spp. represent an epidemiological situation of the species encountering carbapenems. J Antimicrob Chemother. 2017;72(10):2708–2714. doi:10.1093/jac/dkx205
62. Mann R, Rafei R, Gunawan C, Harmer CJ, Hamidian M. Variants of Tn 6924, a novel Tn 7 family transposon carrying the bla NDM Metallo-β-Lactamase and 14 Copies of the aphA6 amikacin resistance genes found in Acinetobacter baumannii. Microbiol Spectr. 2022;10(1):e01745–21. doi:10.1128/spectrum.01745-21
63. Deylam Salehi M, Ferdosi-Shahandashti E, Yahyapour Y, Khafri S, Pournajaf A, Rajabnia R. Integron-mediated antibiotic resistance in Acinetobacter baumannii isolated from intensive care unit patients, Babol, North of Iran. Biomed Res Int. 2017;2017:1–8. doi:10.1155/2017/7157923
64. Nikibakhsh M, Firoozeh F, Badmasti F, Kabir K, Zibaei M. Molecular study of metallo-β-lactamases and integrons in Acinetobacter baumannii isolates from burn patients. BMC Infect Dis. 2021;21(1):1–6. doi:10.1186/s12879-021-06513
65. Amin M, Navidifar T, Shooshtari FS, Goodarzi H. Association of the genes encoding metallo-β-lactamase with the presence of integrons among multidrug-resistant clinical isolates of Acinetobacter baumannii. Infect Drug Resist. 2019;12:1171–1180. doi:10.2147/IDR.S196575
66. Yum JH. High prevalence and genotypic characterization of metallo-β-lactamase (MBL)-producing Acinetobacter spp. isolates disseminated in a Korean Hospital. Korean J Clin Lab Sci. 2019;51(4):444–452. doi:10.15324/kjcls.2019.51.4.444
67. Halaji M, Rezaei A, Zalipoor M, Faghri J. Investigation of class I, II, and III integrons among Acinetobacter Baumannii isolates from hospitalized patients in Isfahan, Iran. Oman Med J. 2018;33(1):37–42. doi:10.5001/omj.2018.07
68. Kalal BS, Chandran SP, Yoganand R, Nagaraj S. Molecular characterization of carbapenem-resistant Acinetobacter baumannii strains from a tertiary care center in South India. Infectio. 2020;24(1):27–34. doi:10.22354/in.v24i1.824
69. Aruhomukama D, Najjuka CF, Kajumbula H, et al. blaVIM-and blaOXA-mediated carbapenem resistance among Acinetobacter baumannii and Pseudomonas aeruginosa isolates from the Mulago Hospital intensive care unit in Kampala, Uganda. BMC Infect Dis. 2019;19(1):1–8. doi:10.1186/s12879-019-4510-5
70. Sung JY, Koo SH, Cho HH, Kwon KC. AbaR7, a genomic resistance island found in multidrug-resistant Acinetobacter baumannii isolates in Daejeon, Korea. Ann Lab Med. 2012;32(5):324–330. doi:10.3343/alm.2012.32.5.324
71. Fournier PE, Vallenet D, Barbe V, et al. Comparative genomics of multidrug resistance in Acinetobacter baumannii. PLoS Genet. 2006;2(1):e7. doi:10.1371/journal.pgen.0020007
72. Nowak P, Paluchowska P. Acinetobacter baumannii: biology and drug resistance—role of carbapenemases. Folia Histochemica Et Cytobiologica. 2016;54(2):61–74. doi:10.5603/FHC.a2016.0009
73. Bi D, Xie R, Zheng J, et al. Large-scale identification of AbaR-type genomic islands in Acinetobacter baumannii reveals diverse insertion sites and clonal lineage-specific antimicrobial resistance gene profiles. Antimicrob Agents Chemother. 2019;63(4):e02526–18. doi:10.1128/AAC.02526-18
74. Bi D, Zheng J, Xie R, et al. Comparative analysis of AbaR-type genomic islands reveals distinct patterns of genetic features in elements with different backbones. Msphere. 2020;5(3):e00349–20. doi:10.1128/mSphere.00349-20
75. Bergogne-Berezin E, Towner KJ. Acinetobacter spp. as nosocomial pathogens: microbiological, clinical, and epidemiological features. Clin Microbiol Rev. 1996;9(2):148–165. doi:10.1128/CMR.9.2.148
76. Queenan AM, Bush K. Carbapenemases: the versatile β-lactamases. Clin Microbiol Rev. 2007;20(3):440–458. doi:10.1128/CMR.00001-07
77. Paton R, Miles RS, Hood J, Amyes SGB. ARI 1: β-lactamase-mediated imipenem resistance in Acinetobacter baumannii. Int J Antimicrob Agents. 1993;2(2):81–87. doi:10.1016/0924-8579(93
78. Zarrilli R, Pournaras S, Giannouli M, Tsakris A. Global evolution of multidrug-resistant Acinetobacter baumannii clonal lineages. Int J Antimicrob Agents. 2013;41(1):11–19. doi:10.1016/j.ijantimicag.2012.09.008
79. Afzal-Shah M, Woodford N, Livermore DM. Characterization of OXA-25, OXA-26, and OXA-27, molecular class D β-lactamases associated with carbapenem resistance in clinical isolates of Acinetobacter baumannii. Antimicrob Agents Chemother. 2001;45(2):583–588. doi:10.1128/AAC.45.2.583-588.2001
80. Brown S, Amyes S. OXA β-lactamases in Acinetobacter: the story so far. J Antimicrob Chemother. 2006;57(1):1–3. doi:10.1093/jac/dki425
81. Yang Z, Wang P, Song P, Li X. Carbapenemase OXA-423: a Novel OXA-23 Variant in Acinetobacter baumannii. Infect Drug Resist. 2020;13:4069–4075. doi:10.2147/IDR.S277364
82. Singh M, De Silva PM, Al-Saadi Y, et al. Characterization of extremely drug-resistant and hypervirulent Acinetobacter baumannii AB030. Antibiotics. 2020;9(6):328. doi:10.3390/antibiotics9060328
83. Wang Z, Li H, Zhang J, Wang H. Co-Occurrence of bla OXA-23 in the chromosome and plasmid: increased fitness in carbapenem-resistant Acinetobacter baumannii. Antibiotics. 2021;10(10):1196. doi:10.3390/antibiotics10101196
84. Brown S, Young HK, Amyes SGB. Characterisation of OXA-51, a novel class D carbapenemase found in genetically unrelated clinical strains of Acinetobacter baumannii from Argentina. Clin Microbiol Infect. 2005;11(1):15–23. doi:10.1111/j.1469-0691.2004.01016.x
85. Rodríguez CH, Nastro M, Famiglietti A. Carbapenemases in Acinetobacter baumannii. Review of their dissemination in Latin America. Revista Argentina de Microbiología. 2018;50(3):327–333. doi:10.1016/j.ram.2017.10.006
86. Lee YT, Kuo SC, Chiang MC, et al. Emergence of carbapenem-resistant non-baumannii species of Acinetobacter harboring a bla OXA-51-like gene that is intrinsic to A baumannii. Antimicrob Agents Chemother. 2012;56(2):1124–1127. doi:10.1128/AAC.00622-11
87. Leski TA, Bangura U, Jimmy DH, et al. Identification of bla OXA-51-like, bla OXA-58, bla DIM-1, and bla VIM carbapenemase genes in hospital Enterobacteriaceae isolates from Sierra Leone. J Clin Microbiol. 2013;51(7):2435–2438. doi:10.1128/JCM.00832-13
88. Bou G, Oliver A, Martínez-Beltrán J. OXA-24, a novel class D β-lactamase with carbapenemase activity in an Acinetobacter baumannii clinical strain. Antimicrob Agents Chemother. 2000;44(6):1556–1561. doi:10.1128/AAC.44.6.1556-1561.2000
89. Poirel L, Naas T, Nordmann P. Diversity, epidemiology, and genetics of class D β-lactamases. Antimicrob Agents Chemother. 2010;54(1):24–38. doi:10.1128/AAC.01512-08
90. Lu PL, Doumith M, Livermore DM, Chen TP, Woodford N. Diversity of carbapenem resistance mechanisms in Acinetobacter baumannii from a Taiwan Hospital: spread of plasmid-borne OXA-72 carbapenemase. J Antimicrob Chemother. 2009;63(4):641–647. doi:10.1093/jac/dkn553
91. Poirel L, Marqué S, Héritier C, Segonds C, Chabanon G, Nordmann P. OXA-58, a novel class D β-lactamase involved in resistance to carbapenems in Acinetobacter baumannii. Antimicrob Agents Chemother. 2005;49(1):202–208. doi:10.1128/AAC.49.1.202-208.2005
92. Merkier AK, Catalano M, Ramírez MS, et al. Polyclonal spread of blaOXA-23 and blaOXA-58 in Acinetobacter baumannii isolates from Argentina. J Infect Dev Ctries. 2008;2(03):235–240. doi:10.3855/jidc.269
93. Poirel L, Mansour W, Bouallegue O, Nordmann P. Carbapenem-resistant Acinetobacter baumannii isolates from Tunisia producing the OXA-58-like carbapenem-hydrolyzing oxacillinase OXA-97. Antimicrob Agents Chemother. 2008;52(5):1613–1617. doi:10.1128/AAC.00978-07
94. Sevillano E, Fernandez E, Bustamante Z, et al. Emergence and clonal dissemination of carbapenem-hydrolysing OXA-58-producing Acinetobacter baumannii isolates in Bolivia. J Med Microbiol. 2012;61(1):80–84. doi:10.1099/jmm.0.032722-0
95. Bertini A, Poirel L, Bernabeu S, et al. Multicopy bla OXA-58 gene as a source of high-level resistance to carbapenems in Acinetobacter baumannii. Antimicrob Agents Chemother. 2007;51(7):2324–2328. doi:10.1128/AAC.01502-06
96. Héritier C, Dubouix A, Poirel L, Marty N, Nordmann P. A nosocomial outbreak of Acinetobacter baumannii isolates expressing the carbapenem-hydrolysing oxacillinase OXA-58. J Antimicrob Chemother. 2005;55(1):115–118. doi:10.1093/jac/dkh500
97. Higgins PG, Poirel L, Lehmann M, Nordmann P, Seifert H. OXA-143, a novel carbapenem-hydrolyzing class D β-lactamase in Acinetobacter baumannii. Antimicrob Agents Chemother. 2009;53(12):5035–5038. doi:10.1128/AAC.00856-09
98. Gionco B, Pelayo JS, Venancio EJ, Cayô R, Gales AC, Carrara-Marroni FE. Detection of OXA-231, a new variant of bla OXA-143, in Acinetobacter baumannii from Brazil: a case report. J Antimicrob Chemother. 2012;67(10):2531–2532. doi:10.1093/jac/dks223
99. Higgins PG, Pérez-Llarena FJ, Zander E, Fernández A, Bou G, Seifert H. OXA-235, a novel class D β-lactamase involved in resistance to carbapenems in Acinetobacter baumannii. Antimicrob Agents Chemother. 2013;57(5):2121–2126. doi:10.1128/AAC.02413-12
100. Goncalves D, Cecilio P, Ferreira H. First detection of OXA-48-like producing Acinetobacter baumannii in the faecal flora of nursing home residents in Northern Portugal. In: Twenty-Third European Congress of Clinical Microbiology and Infectious Diseases. Berlin, Germany: European Congress; 2013.
101. Tognim MC, Gales AC, Penteado AP, Silbert S, Sader HS. Dissemination of IMP-1 metallo-β-lactamase–producing Acinetobacter species in a Brazilian teaching hospital. Infect Control Hosp Epidemiol. 2006;27(7):742–747. doi:10.1086/504356
102. Riccio ML, Franceschini N, Boschi L, et al. Characterization of the metallo-β-lactamase determinant of Acinetobacter baumannii AC-54/97 reveals the existence of bla IMP allelic variants carried by gene cassettes of different phylogeny. Antimicrob Agents Chemother. 2000;44(5):1229–1235. doi:10.1128/AAC.44.5.1229-1235
103. Chu YW, Afzal-Shah M, Houang ET, et al. IMP-4, a novel metallo-β-lactamase from nosocomial Acinetobacter spp. collected in Hong Kong between 1994 and 1998. Antimicrob Agents Chemother. 2001;45(3):710–714. doi:10.1128/AAC.45.3.710-714.2001
104. Da Silva GJ, Correia M, Vital C, et al. Molecular characterization of bla IMP-5, a new integron-borne metallo-β-lactamase gene from an Acinetobacter baumannii nosocomial isolate in Portugal. FEMS Microbiol Lett. 2002;215(1):33–39. doi:10.1016/S0378-1097(02)00896-0
105. Gales AC, Tognim MC, Reis AO, Jones RN. Emergence of an IMP-like metallo-enzyme in an Acinetobacter baumannii clinical strain from a Brazilian teaching hospital. Diagn Microbiol Infect Dis. 2003;45(1):77–79. doi:10.1016/s0732-8893(02)00500-x
106. Wang H, Guo P, Sun H, et al. Molecular epidemiology of clinical isolates of carbapenem-resistant Acinetobacter spp. from Chinese hospitals. Antimicrob Agents Chemother. 2007;51(11):4022–4028. doi:10.1128/AAC.01259-06
107. Cayô R, Rodrigues-Costa F, Matos AP, Carvalhaes CG, Jové T, Gales AC. Identification of a new integron harboring bla IMP-10 in carbapenem-resistant Acinetobacter baumannii clinical isolates. Antimicrob Agents Chemother. 2015;59(6):3687–3689. doi:10.1128/AAC.04991-14
108. Yamamoto M, Nagao M, Matsumura Y, et al. Interspecies dissemination of a novel class 1 integron carrying bla IMP-19 among Acinetobacter species in Japan. J Antimicrob Chemother. 2011;66(11):2480–2483. doi:10.1093/jac/dkr336
109. Azizi O, Shakibaie MR, Badmasti F, et al. Class 1 integrons in non-clonal multidrug-resistant Acinetobacter baumannii from Iran, description of the new blaIMP-55 allele in In1243. J Med Microbiol. 2016;65(9):928–936. doi:10.1099/jmm.0.000315
110. Yum JH, Yi K, Lee H, et al. Molecular characterization of metallo-b-lactamase-producing Acinetobacter baumannii and Acinetobacter genomospecies 3 from Korea: identification of two new integrons carrying the bla VIM-2 gene cassettes. J Antimicrob Chemother. 2002;49(5):837–840. doi:10.1093/jac/dkf043
111. Tsakris A, Ikonomidis A, Pournaras S, et al. VIM-1 metallo-β-lactamase in Acinetobacter baumannii. Emerg Infect Dis. 2006;12(6):981–983. doi:10.3201/eid1206.051097
112. Figueiredo S, Poirel L, Papa A, Koulourida V, Nordmann P. First identification of VIM-4 metallo-β-lactamase in Acinetobacter spp. Clin Microbiol Infect. 2008;14(3):289–290. doi:10.1111/j.1469-0691.2007.01942.x
113. Lee MF, Peng CF, Hsu HJ, Chen YH. Molecular characterisation of the metallo-β-lactamase genes in imipenem-resistant Gram-negative bacteria from a university hospital in southern Taiwan. Int J Antimicrob Agents. 2008;32(6):475–480. doi:10.1016/j.ijantimicag.2008.07.009
114. Lee K, Yum JH, Yong D, et al. Novel acquired metallo-β-lactamase gene, bla SIM-1, in a class 1 integron from Acinetobacter baumannii clinical isolates from Korea. Antimicrob Agents Chemother. 2005;49(11):4485–4491. doi:10.1128/AAC.49.11.4485-4491.2005
115. Kaase M, Nordmann P, Wichelhaus TA, Gatermann SG, Bonnin RA, Poirel L. NDM-2 carbapenemase in Acinetobacter baumannii from Egypt. J Antimicrob Chemother. 2011;66(6):1260–1262. doi:10.1093/jac/dkr135
116. Kumar M. Identification of a novel NDM variant, blaNDM-3, from a multidrug-resistant Acinetobacter baumannii. Infect Control Hosp Epidemiol. 2016;37(6):747–748. doi:10.1017/ice.2016.66
117. Xanthopoulou K, Urrutikoetxea-Gutiérrez M, Vidal-Garcia M, et al. First report of New Delhi Metallo-β-Lactamase-6 (NDM-6) in a clinical Acinetobacter baumannii isolate from Northern Spain. Front Microbiol. 2020;11:589253. doi:10.3389/fmicb.2020.589253
118. Shahcheraghi F, Abbasalipour M, Feizabadi MM, Ebrahimipour GH, Akbari N. Isolation and genetic characterization of metallo-β-lactamase and carbapenemase producing strains of Acinetobacter baumannii from patients at Tehran hospitals. Iran J Microbiol. 2011;3(2):68.
119. Mohamed NM, Raafat D. Phenotypic and genotypic detection of metallo-beta-lactamases in imipenem-resistant Acinetobacter baumannii isolated from a tertiary hospital in Alexandria, Egypt. Res J Microbiol. 2011;6(10):750–760. doi:10.17311/jm.2011.750.760
120. Girija SA, Jayaseelan VP, Arumugam P. Prevalence of VIM-and GIM-producing Acinetobacter baumannii from patients with severe urinary tract infection. Acta Microbiol Immunol Hung. 2018;65(4):539–550. doi:10.1556/030.65.2018.038
121. Robledo IE, Aquino EE, Santé MI, et al. Detection of KPC in Acinetobacter spp. in Puerto Rico. Antimicrob Agents Chemother. 2010;54(3):1354–1357. doi:10.1128/AAC.00899-09
122. Moubareck C, Brémont S, Conroy MC, Courvalin P, Lambert T. GES-11, a novel integron-associated GES variant in Acinetobacter baumannii. Antimicrob Agents Chemother. 2009;53(8):3579–3581. doi:10.1128/AAC.00072-09
123. Bogaerts P, Naas T, El Garch F, et al. GES extended-spectrum β-lactamases in Acinetobacter baumannii isolates in Belgium. Antimicrob Agents Chemother. 2010;54(11):4872–4878. doi:10.1128/AAC.00871-10
124. Al-Agamy MH, Jeannot K, El-Mahdy TS, et al. First detection of GES-5 carbapenemase-producing Acinetobacter baumannii isolate. Microb Drug Resist. 2017;23(5):556–562. doi:10.1089/mdr.2016.0152
125. Yungyuen T, Chatsuwan T, Plongla R, et al. Nationwide surveillance and molecular characterization of critically drug-resistant Gram-negative bacteria: results of the Research University Network Thailand study. Antimicrob Agents Chemother. 2021;65(9):e00675–21. doi:10.1128/AAC.00675-21
126. Ruan Z, Chen Y, Jiang Y, et al. Wide distribution of CC92 carbapenem-resistant and OXA-23-producing Acinetobacter baumannii in multiple provinces of China. Int J Antimicrob Agents. 2013;42(4):322–328. doi:10.1016/j.ijantimicag.2013.06.019
127. Katip W, Uitrakul S, Oberdorfer P. A comparison of colistin versus colistin plus meropenem for the treatment of carbapenem-resistant Acinetobacter baumannii in critically ill patients: a propensity score-matched analysis. Antibiotics. 2020;9(10):647. doi:10.3390/antibiotics9100647
128. Katip W, Uitrakul S, Oberdorfer P. Clinical efficacy and nephrotoxicity of the loading dose colistin for the treatment of carbapenem-resistant Acinetobacter baumannii in critically ill patients. Pharmaceutics. 2021;14(1):31. doi:10.3390/pharmaceutics14010031
129. Katip W, Uitrakul S, Oberdorfer P. Clinical outcomes and nephrotoxicity of colistin loading dose for treatment of extensively drug-resistant Acinetobacter baumannii in cancer patients. Infect Drug Resist. 2017;10:293. doi:10.2147/IDR.S144314
130. Katip W, Oberdorfer P, Kasatpibal N. Effectiveness and nephrotoxicity of loading dose colistin–meropenem versus loading dose colistin–imipenem in the treatment of carbapenem-resistant Acinetobacter baumannii infection. Pharmaceutics. 2022;14(6):1266. doi:10.3390/pharmaceutics14061266
131. Seok H, Choi WS, Lee S, et al. What is the optimal antibiotic treatment strategy for carbapenem-resistant Acinetobacter baumannii (CRAB)? A multicentre study in Korea. J Glob Antimicrob Resist. 2021;24:429–439. doi:10.1016/j.jgar.2021.01.018
132. Gandham NR, Gupta N, Jadhav SV, Misra RN. Isolation of Acinetobacter baumannii from cerebrospinal fluid following craniotomy. Med J DY Patil Univ. 2012;5:151–153. doi:10.4103/0975-2870.103347
133. Higgins PG, Schneiders T, Hamprecht A, Seifert H. In vivo selection of a missense mutation in adeR and conversion of the novel bla OXA-164 gene into bla OXA-58 in carbapenem-resistant Acinetobacter baumannii isolates from a hospitalized patient. Antimicrob Agents Chemother. 2010;54(12):5021–5027. doi:10.1128/AAC.00598-10
134. Lin MF, Lan CY. Antimicrobial resistance in Acinetobacter baumannii: from bench to bedside. Journal of Clinical Cases: World J Clin Cases. 2014;2(12):787. doi:10.12998/wjcc.v2.i12.787
135. Cicek AC, Saral A, Iraz M, et al. OXA-and GES-type β-lactamases predominate in extensively drug-resistant Acinetobacter baumannii isolates from a Turkish University Hospital. Clin Microbiol Infect. 2014;20(5):410–415. doi:10.1111/1469-0691.12338
136. Lee CR, Lee JH, Park M, et al. Biology of Acinetobacter baumannii: pathogenesis, antibiotic resistance mechanisms, and prospective treatment options. Front Cell Infect Microbiol. 2017;7:55. doi:10.3389/fcimb.2017.00055
137. Simo Tchuinte PL, Rabenandrasana MA, Kowalewicz C, et al. Phenotypic and molecular characterisations of carbapenem-resistant Acinetobacter baumannii strains isolated in Madagascar. Antimicrob Resist Infect Control. 2019;8(1):1–9. doi:10.1186/s13756-019-0491-9
138. El-Badawy MF, Abdelwahab SF, Alghamdi SA, Shohayeb MM. Characterization of phenotypic and genotypic traits of carbapenem-resistant Acinetobacter baumannii clinical isolates recovered from a tertiary care hospital in Taif, Saudi Arabia. Infect Drug Resist. 2019;12:3113. doi:10.2147/IDR.S206691
139. El Hafa H, Nayme K, El Hamzaoui N, et al. Dissemination of carbapenem-resistant Acinetobacter baumannii strains carrying the blaGES, blaNDM and blaOXA23 in Morocco. Germs. 2019;9(3):133. doi:10.18683/germs.2019.1168
140. Huang ZY, Li J, Shui J, Wang HC, Hu YM, Zou MX. Co-existence of bla OXA-23 and bla VIM in carbapenem-resistant Acinetobacter baumannii isolates belonging to global complex 2 in a Chinese teaching hospital. Chin Med J. 2019;132(10):1166–1172. doi:10.1097/CM9.0000000000000193
141. Boral B, Unaldi Ö, Ergin A, Durmaz R, Eser ÖK. A prospective multicenter study on the evaluation of antimicrobial resistance and molecular epidemiology of multidrug-resistant Acinetobacter baumannii infections in intensive care units with clinical and environmental features. Ann Clin Microbiol Antimicrob. 2019;18(1):1–9. doi:10.1186/s12941-019-0319-8
142. Vijayakumar S, Mathur P, Kapil A, et al. Molecular characterization & epidemiology of carbapenem-resistant Acinetobacter baumannii collected across India. Indian J Med Res. 2019;149(2):240. doi:10.4103/ijmr.IJMR_2085_17
143. Coskun US, Caliskan E, Cicek AC, Turumtay H, Sandalli C. β-lactamase genes in carbapenem resistance Acinetobacter baumannii isolates from a Turkish university hospital. J Infect Dev Ctries. 2019;13(01):50–55. doi:10.3855/jidc.10556
144. Guo J, Li C. Molecular epidemiology and decreased susceptibility to disinfectants in carbapenem-resistant Acinetobacter baumannii isolated from intensive care unit patients in central China. J Infect Public Health. 2019;12(6):890–896. doi:10.1016/j.jiph.2019.06.007
145. Hoang Quoc C, Nguyen Thi Phuong T, Nguyen Duc H, et al. Carbapenemase genes and multidrug resistance of Acinetobacter baumannii: a cross sectional study of patients with pneumonia in Southern Vietnam. Antibiotics. 2019;8(3):148. doi:10.3390/antibiotics8030148
146. Tafreshi N, Babaeekhou L, Ghane M. Antibiotic resistance pattern of Acinetobacter baumannii from burns patients: increase in prevalence of blaOXA-24-like and blaOXA-58-like genes. Iran J Microbiol. 2019;11(6):502–509.
147. Tavares LC, Vasconcellos FM, Sousa WV, et al. Emergence and persistence of high-risk clones among MDR and XDR A. baumannii at a Brazilian teaching hospital. Front Microbiol. 2019;9:2898. doi:10.3389/fmicb.2018.02898
148. Khaledi M, Shahini Shams Abadi M, Validi M, Zamanzad B, Vafapour R, Gholipour A. Phenotypic and genotypic detection of metallo-β-lactamases in A. baumanii isolates obtained from clinical samples in Shahrekord, southwest Iran. BMC Res Notes. 2019;12(1):1–6. doi:10.1186/s13104-019-4636-y
149. Ranjbar R, Farahani A. Study of genetic diversity, biofilm formation, and detection of Carbapenemase, MBL, ESBL, and tetracycline resistance genes in multidrug-resistant Acinetobacter baumannii isolated from burn wound infections in Iran. Antimicrob Resist Infect Control. 2019;8(1):1. doi:10.1186/s13756-019-0612-5
150. Sharma M, Singhal L, Gautam V, Ray P. Distribution of carbapenemase genes in clinical isolates of Acinetobacter baumannii & a comparison of MALDI-TOF mass spectrometry-based detection of carbapenemase production with other phenotypic methods. Indian J Med Res. 2020;151(6):585. doi:10.4103/ijmr.IJMR_1383_18
151. Ezadi F, Jamali A, Heidari A, Javid N, Ardebili A. Heteroresistance to colistin in oxacillinase-producing carbapenem-resistant Acinetobacter baumannii clinical isolates from Gorgan, Northern Iran. J Glob Antimicrob Resist. 2020;21:380–385. doi:10.1016/j.jgar.2019.11.010
152. El-Kazzaz W, Metwally L, Yahia R, Al-Harbi N, El-Taher A, Hetta HF. Antibiogram, prevalence of OXA carbapenemase encoding genes, and RAPD-genotyping of multidrug-resistant Acinetobacter baumannii incriminated in hidden community-acquired infections. Antibiotics. 2020;9(9):603. doi:10.3390/antibiotics9090603
153. Rao M, Rashid FA, Shukor S, Hashim R, Ahmad N. Detection of antimicrobial resistance genes associated with carbapenem resistance from the whole-genome sequence of Acinetobacter baumannii isolates from Malaysia. Can J Infect Dis Med Microbiol. 2020. doi:10.1155/2020/5021064
154. Azimi L, Fallah F, Karimi A, et al. Survey of various carbapenem-resistant mechanisms of Acinetobacter baumannii and Pseudomonas aeruginosa isolated from clinical samples in Iran. Iran J Basic Med Sci. 2020;23(11):1396–1400. doi:10.22038/IJBMS.2020.44853.10463
155. Thirapanmethee K, Srisiri-A-Nun T, Houngsaitong J, Montakantikul P, Khuntayaporn P, Chomnawang MT. Prevalence of OXA-type β-lactamase genes among carbapenem-resistant Acinetobacter baumannii clinical isolates in Thailand. Antibiotics. 2020;9(12):864. doi:10.3390/antibiotics9120864
156. de Freitas SB, Amaral SC, Ferreira MR, et al. Molecular characterization of carbapenem-resistant Acinetobacter baumannii associated with nosocomial infection in the Pelotas, RS, Brazil. Curr Microbiol. 2020;77(10):2724–2734. doi:10.1007/s00284-020-02060-w
157. Fam NS, Gamal D, Mohamed SH, et al. Molecular characterization of Carbapenem/Colistin-resistant Acinetobacter baumannii clinical isolates from Egypt by whole-genome sequencing. Infect Drug Resist. 2020;13:4487–4493. doi:10.2147/IDR.S288865
158. Nodari CS, Cayô R, Streling AP, et al. Genomic analysis of carbapenem-resistant Acinetobacter baumannii isolates belonging to major endemic clones in South America. Front Microbiol. 2020;11:584603. doi:10.3389/fmicb.2020.584603
159. Bansal G, Allen-McFarlane R, Eribo B. Antibiotic susceptibility, clonality, and molecular characterization of carbapenem-resistant clinical isolates of Acinetobacter baumannii from Washington DC. Int J Microbiol. 2020;2120159. doi:10.1155/2020/2120159
160. Mortazavi SM, Farshadzadeh Z, Janabadi S, et al. Evaluating the frequency of carbapenem and aminoglycoside resistance genes among clinical isolates of Acinetobacter baumannii from Ahvaz, south-west Iran. New Microbes New Infect. 2020;38:100779. doi:10.1016/j.nmni.2020.100779
161. Anane YA, Apalata T, Vasaikar S, Okuthe GE, Songca S. Molecular detection of carbapenemase-encoding genes in multidrug-resistant acinetobacter baumannii clinical isolates in South Africa. Int J Microbiol. 2020;1–10. doi:10.1155/2020/7380740
162. Lukovic B, Gajic I, Dimkic I, et al. The first nationwide multicenter study of Acinetobacter baumannii recovered in Serbia: emergence of OXA-72, OXA-23 and NDM-1-producing isolates. Antimicrob Resist Infect Control. 2020;9(1):1–2. doi:10.1186/s13756-020-00769-8
163. Rezaei A, Fazeli H, Faghri J. Investigation of carbapenem resistant Acinetobacter baumannii ST2 in Iran. Acta Microbiol Immunol Hung. 2021;68(1):20–26. doi:10.1556/030.2020.01164
164. Gozalan A, Unaldı O, Guldemir D, et al. Molecular characterization of carbapenem-resistant Acinetobacter baumannii blood culture isolates from three hospitals in Turkey. Jpn J Infect Dis. 2021;74(3):200–208. doi:10.7883/yoken.JJID.2020.478
165. Wasfi R, Rasslan F, Hassan SS, Ashour HM, El-Rahman A, Ola A. Co-existence of carbapenemase-encoding genes in Acinetobacter baumannii from cancer patients. Infect Dis Ther. 2021;10(1):291–305. doi:10.1007/s40121-020-00369-4
166. Hassan RM, Salem ST, Hassan SI, Hegab AS, Elkholy YS, Chang Y-F. Molecular characterization of carbapenem-resistant Acinetobacter baumannii clinical isolates from Egyptian patients. PLoS One. 2021;16(6):e0251508. doi:10.1371/journal.pone.0251508
167. Kostyanev T, Xavier BB, García-Castillo M, et al. Phenotypic and molecular characterizations of carbapenem-resistant Acinetobacter baumannii isolates collected within the EURECA study. Int J Antimicrob Agents. 2021;57(6):106345. doi:10.1016/j.ijantimicag.2021.106345
168. Adjei AY, Vasaikar SD, Apalata T, Okuthe EG, Songca SP. Phylogenetic analysis of carbapenem-resistant Acinetobacter baumannii isolated from different sources using Multilocus Sequence Typing Scheme. Infect Genet Evol. 2021;96:105132. doi:10.1016/j.meegid.2021.105132
169. Vahhabi A, Hasani A, Rezaee MA, et al. Carbapenem resistance in Acinetobacter baumannii clinical isolates from northwest Iran: high prevalence of OXA genes in sync. Iran J Microbiol. 2021;13(3):282. doi:10.18502/ijm.v13i3.6388
170. Khuntayaporn P, Kanathum P, Houngsaitong J, Montakantikul P, Thirapanmethee K, Chomnawang MT. Predominance of international clone 2 multidrug-resistant Acinetobacter baumannii clinical isolates in Thailand: a nationwide study. Ann Clin Microbiol Antimicrob. 2021;20(1):1. doi:10.1186/s12941-021-00424-z
171. Ejaz H, Ahmad M, Younas S, et al. Molecular epidemiology of extensively-drug resistant Acinetobacter baumannii sequence type 2 co-harboring bla NDM and bla OXA from clinical origin. Infect Drug Resist. 2021;14:1931–9. doi:10.2147/IDR.S310478
172. Zhang Y, Ding F, Luo Y, et al. Distribution pattern of carbapenemases and solitary contribution to resistance in clinical strains of Acinetobacter baumannii. Ann Palliat Med. 2021;10:9184–9191. doi:10.21037/apm-21-1805
173. Namaei MH, Yousefi M, Askari P, Roshanravan B, Hashemi A, Rezaei Y. High prevalence of multidrug-resistant non-fermentative Gram-negative bacilli harboring blaIMP-1 and blaVIM-1 metallo-beta-lactamase genes in Birjand, south-east Iran. Iran J Microbiol. 2021;13(4):470. doi:10.18502/ijm.v13i4.6971
174. Khalil MA, Ahmed FA, Elkhateeb AF, et al. Virulence characteristics of biofilm-forming acinetobacter baumannii in clinical isolates using a Galleria Mellonella Model. Microorganisms. 2021;9(11):2365. doi:10.3390/microorganisms9112365
175. Xu X, Xu C, Salisu RB, Xu W. Beta-lactamase gene expression level of hospital-acquired CRAB isolated from children in Picu. Infect Drug Resist. 2021;14:3195. doi:10.2147/IDR.S322604
176. Kumari M, Verma S, Venkatesh V, et al. Emergence of blaNDM-1 and blaVIM producing Gram-negative bacilli in ventilator-associated pneumonia at AMR Surveillance Regional Reference Laboratory in India. PLoS One. 2021;16(9):e0256308. doi:10.1371/journal.pone.0256308
177. Gandham N, Gupta N, Vyawahare C, Mirza SB, Misra RN. Molecular characterization identifies upstream presence of ISAba1 to OXA carbapenemase genes in carbapenem-resistant Acinetobacter baumannii. J Lab Physicians. 2022;14(01):006–10. doi:10.1055/s-0041-1732809
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.