Back to Journals » Infection and Drug Resistance » Volume 12
Molecular characterization and prevalence of antibiotic resistance in Helicobacter pylori isolates in Kuala Lumpur, Malaysia
Authors Hanafiah A ![]() , Binmaeil H
, Binmaeil H ![]() , Raja Ali RA
, Raja Ali RA ![]() , Mohamed Rose I
, Mohamed Rose I ![]() , Lopes BS
, Lopes BS
Received 11 June 2019
Accepted for publication 25 July 2019
Published 27 September 2019 Volume 2019:12 Pages 3051—3061
DOI https://doi.org/10.2147/IDR.S219069
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Joachim Wink
Alfizah Hanafiah,1 Hasyanee Binmaeil,1 Raja Affendi Raja Ali,2 Isa Mohamed Rose,3 Bruno S Lopes4
1Department of Medical Microbiology and Immunology; 2Department of Medicine; 3Department of Pathology, Faculty of Medicine, Universiti Kebangsaan Malaysia, Cheras, Kuala Lumpur 56000, Malaysia; 4Department of Medical Microbiology, School of Medicine, Medical Sciences and Nutrition, University of Aberdeen, Aberdeen AB25 2ZD, UK
Correspondence: Alfizah Hanafiah
Department of Medical Microbiology and Immunology, Faculty of Medicine, Universiti Kebangsaan Malaysia Medical Centre, 16th Floor, Preclinical Building, Jalan Yaacob Latif, Bandar Tun Razak, Cheras, Kuala Lumpur 56000, Malaysia
Tel +603 9 145 9539
Fax +603 9 145 9524
Email [email protected]
Bruno S Lopes
Department of Medical Microbiology, School of Medicine, Medical Sciences and Nutrition, University of Aberdeen, 0:025 Polwarth Building, Aberdeen AB25 2ZD, UK
Email [email protected]
Aims and objectives: Helicobacter pylori has been classified as high priority pathogen by the WHO in 2017. The emergence of antibiotic-resistant strains is one of the main causes of treatment failure in H. pylori infection. This study determined and characterized primary and secondary resistances in H. pylori in Malaysia.
Materials and methods: Gastric biopsies from antrum (n=288) and corpus (n=283) were obtained from 288 patients who underwent endoscopy at Universiti Kebangsaan Malaysia Medical Center (UKMMC), Kuala Lumpur, Malaysia. Antibiotic susceptibility to six classes of antibiotics was determined by the E-test. Mutations conferring in resistance in functional genes were identified by PCR and sequencing.
Results: Overall resistance rates to metronidazole, clarithromycin and levofloxacin were 59.3% (35/59), 35.6% (21/59) and 25.4% (15/59), respectively. Secondary isolates showed significantly higher resistance rates to clarithromycin compared to the primary isolates. Mixed infection with susceptible and resistant isolates was observed in 16.2% (6/37) of cases, of which 83.3% (n=5) had infection with the same strain. 41% (18/44) of isolates were resistant to more than one class of antibiotics of which 50% (9/18) were multidrug-resistant, two being primary and seven being secondary isolates. Mutations in rdxA, 23S rRNA and gyrA genes were associated with resistance to metronidazole, clarithromycin and levofloxacin, respectively.
Conclusion: The high level of resistance to metronidazole, clarithromycin and levofloxacin seen in H. pylori isolates in our setting warrants the need for continuous surveillance and highlights caution in use of antibiotics generally used as first-line therapy in H. pylori eradication regimen.
Keywords: Helicobacter pylori, antibiotic resistance, clarithromycin, metronidazole, levofloxacin, point mutation
Introduction
Helicobacter pylori is one of the most prevalent global pathogens that colonizes an estimated 50% of the world’s population.1 The precise epidemiology of H. pylori infection still remains unclear; although various risk factors including lower socioeconomic status, infected parents, contaminated water sources, smoking, ethnicities and geographic location may increase the risk of H. pylori infection.2 Person-to-person transmission by oral-oral, fecal-oral or gastro-oral exposure is suggested to be the most likely route of transmission.3 The evidence that H. pylori infection causes peptic ulcer disease is compelling hence all consensus guidelines recommend eradication of H. pylori for patients with symptoms.4 Standard triple therapy uses a combination of a proton pump inhibitor and two antibiotics, either clarithromycin and amoxicillin or metronidazole for a period of 7–14 days.5 However, this therapy has been questioned because of the increased eradication failure rates.6 Many factors have been implicated as causes of treatment failure, including ineffective penetration of antibiotics into the gastric mucosa, antibiotic inactivation by the low stomach pH, a lack of patient compliance, and the emergence of acquired resistance to antibiotics by H. pylori. The effectiveness of “legacy triple therapy” recommended by Maastricht III Consensus Report provides disappointingly low treatment success (i.e., below 80%).7
Antibiotic resistance in H. pylori strains has been shown to be the main factor causing eradication failure or low eradication rate in treated patients.8 The mechanisms of antibiotic resistance in H. pylori is mainly due to genetic mutations. For example, resistance to clarithromycin in H. pylori is due to point mutations in the peptidyltransferase region of domain V of the 23S rRNA. The most common mutation is an A-to-G transition at position 2143 (A2143G)9 but several other point mutations, at positions A2142G, A2144G, and T2182C, have also been described.10,11 Reports have indicated that other mutations, such as A2115G, G2141A, C2147G, T2190C, C2195T, A2223G, and C2694A, might also be associated with clarithromycin resistance.12,13 Metronidazole resistance in H. pylori is complex and is primarily associated with mutations in rdxA and frxA14 which is often a result of frameshift mutation, insertions and deletions. Resistance to fluoroquinolones arises from mutations in the quinolones resistance-determining region (QRDR) of gyrA.15 Amoxicillin resistance has been associated with amino acid substitutions (D535N, S543R and T556S) found in penicillin-binding protein 1.16 The resistance to tetracycline is due to mutations in the 16S rRNA, where single or double base-pair substitutions (A928C, AG926-927→GT and A926G/A928C) as well as triple substitution (AGA926–928→TTC) confer low to high-level tetracycline resistance in H. pylori.17 Resistance to rifampicin is due to the presence of mutations in RNA polymerase subunit B gene, rpoB.18
There is an increasing trend in primary resistance among H. pylori worldwide with guidelines recommending H. pylori culture and susceptibility testing following two unsuccessful courses of treatment.5 Antibiotic susceptibility testing is unavailable in most of the diagnostic laboratories and so epidemiological data on secondary resistance in H. pylori is essential in allowing the rational use of antibiotics following several failures in treatment.19 In this manner, the treatment success of a particular antibiotic regimen can be reliably predicted based on epidemiological data, even in the absence of individualized H. pylori culture and susceptibility testing. The aims of this study were to determine antimicrobial resistance mechanisms in primary and secondary isolates of H. pylori by identifying the mutations associated with antibiotic-resistance phenotypes in H. pylori isolated from patients with dyspepsia in Kuala Lumpur, Malaysia by presenting both phenotypic and genotypic data.
Materials and methods
This study was approved by the Medical Research Ethic Committee of the University (UKM 1.5.3.5/244/ETP-2013-042) and written informed consent was obtained from each patient. The study was conducted in accordance with the Declaration of Helsinki.
Patient sampling
A total of 288 consecutive patients consisting of 129 Malays, 110 Chinese, 29 Indians and 20 from other ethnic background with chronic dyspepsia undergoing oesophagogastroduodenoscopy (OGDS) at the Universiti Kebangsaan Malaysia Medical Center (UKMMC), Kuala Lumpur, Malaysia from April 2014 to August 2015 were enrolled in the study. Patients who had history of eradication failure and those that did not take antibiotics for at least 4 weeks prior to the time of enrollment were included in the study. Patients’ age was between 18 and 89 years with the mean age of 52.41±16.44 years and median age of 55. For each patient, three biopsies were obtained from the antrum and corpus of the stomach, each placed in different tubes for culture, rapid urease test and histopathological examination.
H. pylori culture
Biopsies for culture were placed in Brucella broth (BBL; Becton Dickson, USA) containing 15% glycerol and transported to the Microbiology laboratory in UKMMC. Gastric biopsies were plated onto Columbia agar (Oxoid, Basingstoke, UK) supplemented with 7% defibrinated sheep blood (Oxoid) and Dent’s supplement (Oxoid). The inoculated agar plates were incubated at 37°C up to 5–10 days in microaerophilic conditions generated using microaerophilic CampyGen gas pack (Oxoid). Culture was recorded as positive when H. pylori growth identified by Gram staining showed gram-negative curved/spiral rods and positive for urease, catalase and oxidase tests.20 Colonies were subcultured on several agar plates and used for antibiotic susceptibility testing and DNA extraction.
Rapid urease test
Biopsies were immersed in urease medium and a color change was recorded between 10 mins and 24 hrs because of hydrolysis of urea into carbon dioxide and ammonia by urease-producing Helicobacter strains. The ammonium ion increases the pH of the medium, which was detected by the indicator phenol red, resulting in color change from yellow to reddish pink.
Histopathological examination
Biopsies for histopathological examination were fixed in 10% formalin and transported to the Pathology Laboratory in UKMMC. Paraffin-embedded section of the biopsies was prepared, cut and stained with Warthin-Starry staining for detection of H. pylori.
Antibiotic susceptibility testing
Due to strain fastidiousness, Clinical Laboratory Standard Institute (CLSI) recommended guidelines for antibiotic resistance in H. pylori by determination of the minimum inhibitory concentration (MIC) to different classes of antibiotics, as there are no standardized disc diffusion methods defined for testing in H. pylori susceptibility. The MIC of H. pylori isolates was determined to six classes of antibiotics (metronidazole, clarithromycin, amoxicillin, levofloxacin, tetracycline and rifampicin) by E-test. Bacterial suspension equivalent to McFarland 3.0 standard (approximately 107 cfu/mL) was prepared from a pure culture of H. pylori. Two hundred microliter of bacterial suspension were dispensed using sterile tip onto Muller-Hinton agar with 5% sheep blood and streaked in 3 different directions using sterile swab such that it produced confluent growth. The E-test strips (bioMérieux, Marcy-l’Etoile, France) were placed on each plate and the plates were incubated in a microaerophilic environment at 37°C for 72 hrs. Resistance breakpoint for metronidazole was >8 mg/L,21 amoxicillin,22 levofloxacin,23 tetracycline24 were >1 mg/L and for rifampicin >4 mg/L.25 The epidemiological cut off for clarithromycin was defined as ≤0.25 mg/L (susceptible), 0.5 mg/L (intermediate), ≥1 mg/mL for resistant according to CLSI guidelines.26 H. pylori strain ATCC 43526 was included as a control. The MIC50 and MIC90 of antibiotics for primary and secondary H. pylori isolates were calculated for each antibiotic.
DNA extraction, PCR and sequencing
H. pylori pure cultures on Columbia blood agar were scraped off and suspended in phosphate-buffered saline. The DNA was extracted using FavorPrep™ Tissue Genomic DNA Extraction Mini kit according to the manufacturer’s instructions (Favorgen Biotech Corporation, Ping-Tung, Taiwan). The extracted DNA was stored at −20°C until further use.
Mutations in rdxA, frxA, 23S rRNA and gyrA genes were determined by targeted gene sequencing approach as described earlier.27–29 The amplification conditions consisted of an initial denaturation at 95 °C for 3 mins, followed by 30 cycles of denaturation at 95°C for 30 s; annealing at 60°C (for rdxA) or 55°C (for frxA) or 54°C (for 23S rRNA) or 50°C (for gyrA) for 30 s and extension at 72°C for 1 min with final extension at 72°C for 10 mins. The PCR amplification was performed in a PCR Thermal Cycler machine (iCycler; Bio-Rad Laboratories, Hercules, CA, USA). The amplified products were purified using Geneaid PCR cleanup kit (Geneaid, New Taipei City, Taiwan) and sequenced using a BigDye terminator v3.1 sequencing kit on ABI 3730×L sequencer (MyTACG Bioscience Enterprise, Selangor, Malaysia). Comparative sequence analyses between resistant and susceptible strains were carried out using BioEdit software (http://www.mbio.ncsu.edu/bioedit/bioedit.html). The DNA sequence results were aligned and compared to the wild-type isolates of H. pylori 23S rRNA (Accession no. KC556778), H. pylori 128A rdxA (Accession no. AF180396) and H. pylori gyrase A subunit (gyrA) (Accession no. L29481).
RAPD-PCR fingerprinting
In patients with a discordant antibiotic susceptibility pattern between the isolates from the antrum and corpus of the stomach, DNA fingerprinting via random amplified polymorphic DNA analyses (RAPD-PCR) was performed using primers 1254, 1281 and 1284 as described by earlier30 in order to find out whether these patients were infected with different or same strains. PCR was carried out in 25 μL volumes containing 20 ng of genomic DNA, 3 mM MgCl2, 400 μM of each dNTP, 0.8 µM of primer and 0.1 U/µL of EconoTaq® DNA polymerase (Lucigen®, Middleton, WI, USA) in the buffer provided. T100™ Thermal Cycler (Bio-Rad Laboratories) was used for amplification and the cycling program was denaturation at 94°C for 1 mins, followed by 45 cycles of 36°C for 1 min, 72°C for 1 min; and a final extension at 72°C for 10 mins. The PCR products were analyzed by electrophoresis in a 2% agarose gel while using 0.5× TBE buffer under a lower voltage (75 V, 2.5 hrs). Gels were stained with GelStain (Transbionovo, Beijing, China) and photographed under UV light under a transilluminator (Alphaimager® Hp; Cell Bioscience).
Statistical analysis
Statistical analysis was performed using SPSS software version 23 (SPSS Inc, Chicago, IL, USA). Differences between groups were statistically evaluated using Chi-square (χ2) test and Fisher’s exact test with a 95% CI was used to define differences in the proportion. Differences were considered significant when P-value was <0.05.
Results
H. pylori infection status and overall susceptibility profile
A patient was regarded as H. pylori positive when culture or histology or rapid urease test and histology showed a positive result. Two hundred and forty-one (83.7%) patients had not undergone any therapy for H. pylori infection, whereas 47 (16.3%) patients had previous history of eradication failure. The frequency of H. pylori infection observed either by culture or histology or rapid urease test was 16.7% (48/288). H. pylori infection was significantly higher in patients with previous treatment failure (34%, 16/47) compared to untreated patients (13.3%, 32/241) (χ2=12.21, P=0.0001, OR=3.371, 95% CI=1.659–6.848). H. pylori infection was detected in 8.5% Malays (11/129), 20.9% Chinese (23/110), 27.6% Indians (8/29) and 30% other ethnicities (6/20). Among H. pylori-infected patients, 70.8% (34/48) had infection in both antrum and corpus, 18.8% (9/48) had infection only in antrum and 10.4% (5/48) had infection only in corpus.
A total of 59 H. pylori were isolated from 37 H. pylori-infected patients. Twenty-two patients had H. pylori isolated from both antrum and corpus, 8 from antrum and 7 from corpus. H. pylori were not isolated from 11 infected-patients positive for rapid urease test and/or histology.
Overall susceptibility profile of H. pylori to the tested antibiotics, regardless type of isolate (primary or secondary) and site of biopsy (antrum or corpus) showed that all isolates were susceptible to amoxicillin, tetracycline and rifampicin. Resistance to metronidazole, clarithromycin and levofloxacin were observed in 59.3% (35/59), 35.6% (21/59) and 25.4% (15/59) of the isolates, respectively. Fifteen isolates (25.4%) were susceptible to all tested antibiotics. Among the 37 patients, 73% (n=27) were infected with resistant H. pylori strains. Single, dual and triple H. pylori resistant infection were observed in 43.2% (16/37), 16.2% (6/37) and 13.5% (5/37) of the patients, respectively.
Mixed infection
Mixed infection by susceptible and resistant H. pylori strains was observed in 16.2% (6/37) of the patients where infection by metronidazole-susceptible and -resistant identified in four patients, clarithromycin-susceptible and-resistant in one patient and levofloxacin-susceptible and -resistant in one patient. RAPD DNA fingerprinting analysis showed that H. pylori strains from antrum and corpus in one patient had different DNA patterns suggesting a mixed infection in this patient. In the other five patients, no substantial differences of DNA pattern between antrum and corpus isolates suggesting an infection with a single clone of H. pylori strain.
Antibiotic susceptibility profiles of primary and secondary H. pylori isolates
Primary isolate is defined as H. pylori strain isolated from patient untreated for Helicobacter infection, whereas secondary isolate originates from patients with eradication failure. Among the 59 isolates, resistance to metronidazole, clarithromycin and levofloxacin were observed in 56.1% (23/41), 12.2% (5/41) and 17.1% (7/41) of the primary isolates and 66.7% (12/18), 88.9% (16/18) and 44.4% (8/18) of the secondary isolates. Majority of the secondary isolates (88.9%) were significantly resistant to clarithromycin (P=0.0001) compared to the primary isolates (12.2%) (Table 1) and were also resistant to metronidazole and levofloxacin compared to the primary isolates but the difference was not statistically significant.
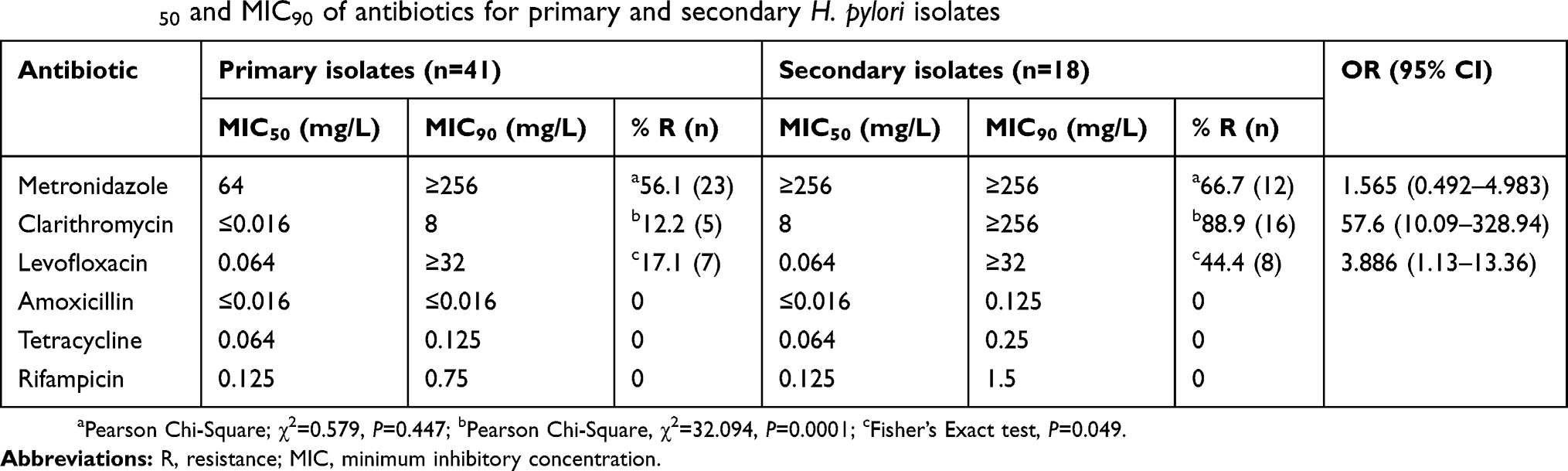 |
Table 1 MIC50 and MIC90 of antibiotics for primary and secondary H. pylori isolates |
Distribution of MICs value of antibiotics among primary and secondary isolates
Secondary isolates had higher MIC50 values for metronidazole and clarithromycin compared to the primary isolates (Table 1). The MIC50 for levofloxacin showed similar value for primary and secondary isolates. Secondary H. pylori isolates show a higher MIC90 for clarithromycin compared to the primary isolates, whereas for metronidazole and levofloxacin, the MIC90 were similar between primary and secondary isolates. For amoxicillin, tetracycline and rifampicin, a higher MIC90 values were observed compared to the primary isolates but still in the susceptible range. Various distributions of MIC values of the antibiotics were observed among primary and secondary H. pylori isolates (Figure S1). For metronidazole, high MIC value (MIC ≥256 mg/L) was found in 46.3% of the primary isolates (19/41) and 55.6% (10/18) of the secondary isolates. Clarithromycin MIC ≥1 mg/L was detected in 12.2% (5/41) of the primary isolates and 88.9% (16/18) were detected among the secondary isolates. 17.1% (7/41) and 44.4% (8/18) of the primary and secondary isolates, respectively, showed levofloxacin MIC of ≥1 mg/L. However, a predominance of levofloxacin MIC ≥32 mg/L was found among the secondary isolates (33.3%; 6/18) compared to the primary isolates (12.2%; 5/41).
Multi-drug resistance
Among metronidazole, clarithromycin and levofloxacin-resistant isolates from both primary and secondary cases (n=44), 59.1% (26/44) were resistant to one antibiotic, 20.5% (9/44) were resistant to two antibiotics, and 20.5% (9/44) were resistant to all three antibiotics. 7.4% (2/27) of the primary isolates were multi-drug resistant compared to 41.1% (7/17) of the secondary isolates.
Mutations in rdxA, frxA, 23S rRNA and gyrA genes
Amino acid substitutions/deletion were identified in 41 sites of the RdxA protein among 59 H. pylori isolates which result in 50 different mutations. Among the metronidazole-resistant strains (Table 2), 40% (14/35) of the strains had missense mutations, 5.7% (2/35) had a codon deletion and 54.3% (19/35) had frameshift/nonsense mutations. In addition, frameshift and nonsense mutations were identified only in metronidazole-resistant strains. As the majority of the isolates had high-level of metronidazole resistance (MIC of 32 to >256 mg/L), no association between type of mutation and MIC value was observed. The rdxA alleles of metronidazole-susceptible strains showed that six isolates (25%) did not harbor any specific mutations, 8.3% (2/24) had a codon deletion and 66.7% (16/24) had missense mutations. Screening for the mutation in frxA in 14 metronidazole-resistant isolates demonstrated that 50% (n=7) of the isolates had frameshift or nonsense mutations, whereas the rest except one isolate (ETP-26C) harbored at least 3 missense mutations (Table 2).
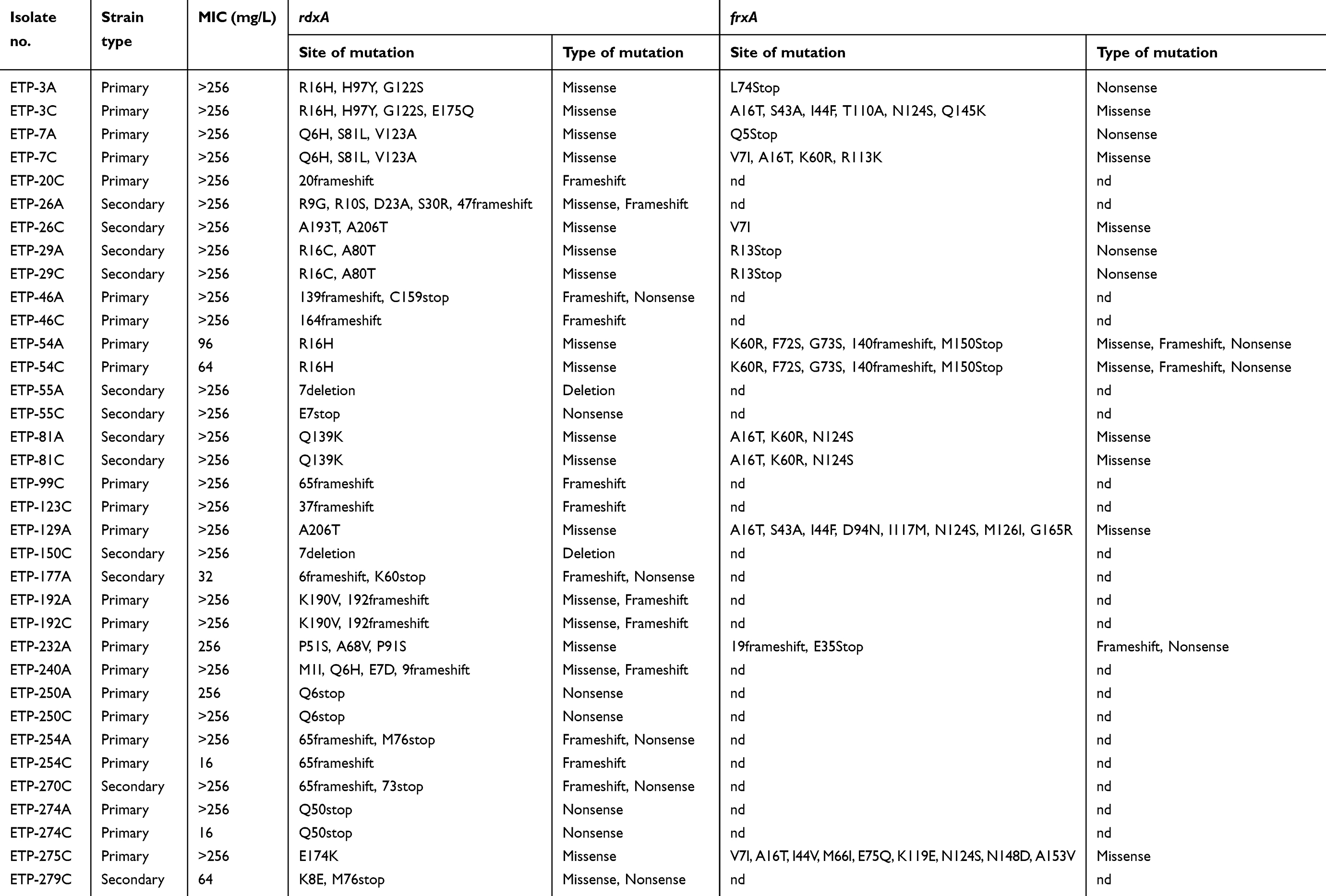 |
Table 2 Metronidazole resistance, rdxA and frxA mutations |
23S rRNA V domain (nucleotide position at 1995 to 2274) was sequenced in all 59 H. pylori strains. There were three-point mutations, which included single nucleotide transition at A2142G, A2143G and T2182G (Table S1). Mutations at nucleotide position 2142 and 2143 that confer A to G change were identified in resistant strains only with majority of the strains had A2143G (90.5%) and only two strains had A2142G mutation (Table 3). None of these strains were found to have double mutations of A2142G and A2143G but either of these mutations occurred in all resistant strains. A point mutation at position 2182 from T to C were identified in both clarithromycin-susceptible (89.5%; 34/38) and -resistant strains (100%; 21/21). Four clarithromycin-susceptible isolates had no mutation in 23S rRNA gene. Among the clarithromycin-resistant strains harboring A2143G mutation, 8 of 19 (42.1%) strains had high MIC value (64 to >256 mg/L) (Table 3). The other 11 strains have various MIC values ranging from 2 to 16 mg/L. Two strains harbored A2142G mutation in 23S rRNA had MIC value for clarithromycin of 32 and >256 mg/L each.
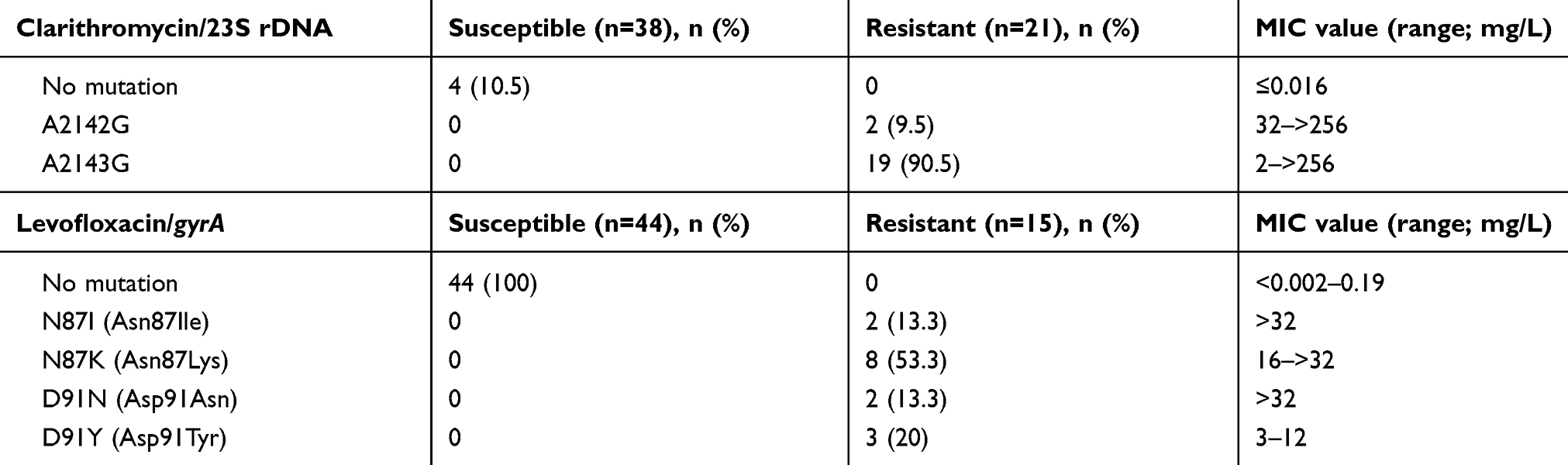 |
Table 3 23S rRNA, gyrA mutations and the range of minimum inhibitory concentration (MIC) values in H. pylori-susceptible and -resistant isolates |
QRDR of gyrA (nucleotide at position 32–600) of H. pylori isolates were sequenced. Upon putative translation, four different amino acid variants, in gyrase A subunit at amino acid position 87 [N87I, N87K] and position 91 [D91N, D91Y] were detected to be exclusively present in resistant strains only (Table S1). Ten of 15 levofloxacin-resistant strains exhibited amino acid substitution at position Asn-87 (66.7%), whereas substitution at position Asp-91 was present in 5 other strains (33.3%). For levofloxacin-resistant isolates, strains harboring mutations N87K, N87I and D91N had higher MIC values of levofloxacin compared to the strains harboring mutation D91Y (Table 3).
Discussion
Over the last decades, studies showed decreasing prevalence of H. pylori infection worldwide.31 This is also noted in the present study, where we observed a lower prevalence rate of H. pylori infection (16.7%) compared to our previous study (30.8%).32 Since Malaysia is a multiracial country with Indian population consistently showing the highest prevalence rate of H. pylori infection and Malay the least, the proportion of ethnic composition plays an important role in determining the overall H. pylori infection in Malaysia.33,34
Infection with antibiotic-resistant H. pylori, poor patient compliance,35 drug regimens and treatment duration are important factors which contribute to the treatment failure and our study shows that 34% of our patients with treatment failure were infected with resistant H. pylori strains. Although a decline in H. pylori infection is seen, an increase in rates of antimicrobial resistance occurs in many parts of the world.36 Results of the present study show that majority of our clinical isolates were resistant to at least one antibiotic. An overall high resistance rates observed in this study were due to the inclusion of both primary and secondary resistance.
Metronidazole is an antimicrobial agent widely used to treat patients with parasitic infection, periodontal and gynecological diseases in many developing countries.5 This may explain the high rates of primary resistance to metronidazole among H. pylori strains.36 In this study, a total of 59.3% of strains were resistant to metronidazole. Primary and secondary resistance was reasonably high and accounted for 56.1% and 66.7% of the isolates, respectively. Previous studies in Malaysia also reported high rates of primary resistance in H. pylori toward metronidazole (32.3–37.4%).37,38 We observed an increasing trend of primary metronidazole resistance in our previous study37 from 37.4% to 56.1%. High prevalence of metronidazole resistance rate has also been reported in other Southeast Asian countries such as Indonesia (46.8%),5 Vietnam (69.9%),39 Thailand (36%),40 and Singapore (38.4%).41 Moreover, secondary resistance to metronidazole in the present study was higher than the primary resistance.
An overall high resistant to clarithromycin was detected in 35.6% of strains, where the rates of strains with secondary resistance (88.9%) were significantly higher than strains with primary resistance (12.2%). Previous studies in Malaysia reported low primary resistance rates to clarithromycin (2.1–6.8%).37 A dramatic increase in primary clarithromycin resistance was seen in our setting from 2.1% (2011 report) to 12.2% (the present study) and is of concern suggesting the evolution of antibiotic-resistant H. pylori. The rise of clarithromycin resistance rate has also been observed in other local reports.38,42 Clarithromycin is the key antibiotic in the first-line H. pylori therapy regimen. A significant difference between primary and secondary resistance to clarithromycin indicates that infection with clarithromycin-resistant H. pylori isolates results in the failure of eradication therapy. Analysis of the clarithromycin MIC distribution shows that clarithromycin may still be a highly effective drug, but only in patients infected with susceptible strains.
A higher primary resistance rate of levofloxacin was found in our study compared to other local reports.37,38 However, the activity of levofloxacin is still high as more than 80% and 50% of primary and secondary isolates, respectively, were inhibited at low MIC values. Increasing trend in levofloxacin resistance worldwide43 is a major concern as it is recommended for salvage treatment regimens in clarithromycin failed treatments.5
H. pylori isolates in our region is consistently susceptible to amoxicillin, tetracycline and rifampicin as shown in the present study and others.37,38 A stable primary resistance to amoxicillin and tetracycline has been reported in Singapore from 2002 to 2014.41 Even though amoxicillin is one of the key antibiotics in first-line triple therapy for H. pylori eradication,5 the finding suggests that exposure to amoxicillin did not induce the development of resistance. A wide range of tetracycline and rifampicin MIC values and the shift in MIC for these drugs toward higher values could be of concern as both drugs are proposed in third-line rescue regimen for H. pylori eradication therapy.
Some of our patients (16.2%) were infected with strains of different antibiotic susceptibilities from the antrum and corpus. Majority of our patients with discordant antibiotic susceptibility between antrum and corpus were infected with a single H. pylori strains as shown by identical RAPD fingerprinting analysis. This suggested that antibiotic-resistant H. pylori strains develop from a pre-existing susceptible strain rather than co-infection with a different strain.44 The presence of heteroresistant strains in the present study is common45,46 indicating that no single biopsy site can be considered representative of antimicrobial susceptibility testing and can lead to underestimation of antimicrobial resistance.47 Multiple biopsies from different sites of the stomach are needed especially in patients with pangastritis in order to verify the presence of heteroresistance to antibiotics.47
Multi-drug resistance H. pylori strains have been reported from many countries.24,39,48,49 Multidrug-resistance in secondary isolates was higher than those in primary indicating the role of antibiotics in the generation of resistance in H. pylori.
Many point mutations in rdxA have been reported and these positions were not uniform and highly distributed among different H. pylori strains.50 Frameshift and nonsense mutations contributed to metronidazole resistance as these mutations were identified only in metronidazole-resistant strains, which is in agreement with previous reports.50 About 45% of our metronidazole-resistant strains harboring missense mutation and a codon deletion had intact rdxA reading frames. However, half of these isolates harbored frameshift or nonsense mutations in frxA. Presence of many missense mutations in both rdxA and frxA might contribute to metronidazole resistance in our H. pylori isolates.
More than 40% of our strains harboring A2143G mutation in 23S rRNA gene had high clarithromycin MIC value suggesting that this mutation is associated with highly resistant H. pylori strains. A previous study reported the A2143G mutation has a much stronger impact on clarithromycin resistance than the A2142G.51
Asn87Lys mutation in the gyrA was predominant among our H. pylori isolates. In other regions such as Europe countries and Japan, the most common mutation was located at amino acid 9125,52 suggesting that there may be a geographical variation in gyrA genotype distribution. Furthermore, we found that more than half of our levofloxacin-resistant strains harbored Asn87Lys mutation associated with high MIC values. Results show that screening of gyrA mutations at amino acids 87 and 91 could be adequate for detecting levofloxacin-resistant among our strains.
Conclusion
The difference between high prevalence of H. pylori in Indian population and low prevalence in Malays in Malaysia is likely to be environmental in origin strains remains confined to predefined cohorts which in the case of Malaysia, is ethnic based supported by distinctness of strains between different races. The increasing incidence of H. pylori resistance to metronidazole, clarithromycin and levofloxacin in Malaysia warrants systematic surveillance to optimize the selection of first-line and second-line treatment regimens which can be suitable for people of specific ethnic groups due to their likelihood of being infected with a lineage-specific strain belonging to ethnic cohort. This may reduce secondary resistance due to treatment failure and prevent the development of resistance in H. pylori isolates. It is recommended that antibiotic susceptibility testing be performed before administration of the first-line antibiotics to reduce the development of secondary resistance especially to clarithromycin which is an effective drug for the treatment of metronidazole-resistant H. pylori. Rapid point of care assays and/or implementation of molecular assays for detection of mutations in rdxA, frxA, 23S rRNA and gyrA can aid in linking genotypic results with the phenotype for treatment of disease.
Ethics approval and informed consent
The study was approved by the Medical Research Ethic Committee of the University (UKM 1.5.3.5/244/ETP-2013-042) and written informed consent was obtained from each patient and the study was conducted in accordance with the Declaration of Helsinki.
Data sharing statement
Data will be shared and available from the corresponding author on reasonable request.
Acknowledgment
We would like to thank the Universiti Kebangsaan Malaysia for providing both the permission and the facilities to conduct and publish this research. The research was funded by a grant from Universiti Kebangsaan Malaysia under Economic Transformation Programme Research Fund Scheme (grant no. ETP-2013-042).
Author contributions
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; gave final approval of the version to be published; and agree to be accountable for all aspects of the work.
Disclosure
HB received a scholarship from the Universiti Kebangsaan Malaysia under the postgraduate “Zamalah” program. The authors declare no other conflicts of interest in this work.
References
1. Hooi JKY, Lai WY, Ng WK. Global prevalence of Helicobacter pylori infection: systematic review and meta-analysis. Gastroenterology 2017;153:420–429.
2. Ozbey G, Demirel U, Aygun C, Ertas HB. Investigation of the association between clinical outcome and the cag pathogenicity-island and other virulence genes of Helicobacter pylori isolates from patients with dyspepsia in Eastern Turkey. Brazilian J Microbiol. 2013;44(4):1267–1274. doi:10.1590/S1517-83822013000400034
3. Brown LM. Helicobacter pylori: epidemiology and routes of transmission. Epidemiol Rev. 2000;22(2):283–297. doi:10.1093/oxfordjournals.epirev.a018040
4. Jones NL, Koletzko S, Goodman K, et al. Joint ESPGHAN/NASPGHAN guidelines for the management of Helicobacter pylori in children and adolescents (update 2016). J Pediatr Gastroenterol Nutr. 2017;64(6):991–1003.
5. Malfertheiner P, Megraud F, O’Morain CA, et al. Management of Helicobacter pylori infection – the Maastricht IV/Florence Consensus Report. Gut. 2012;61(5):646–664. doi:10.1136/gutjnl-2012-302084
6. Megraud F. H. pylori antibiotic resistance: prevalence, importance, and advances in testing. Gut. 2004;53(9):1374–1384. doi:10.1136/gut.2003.022111
7. Wu W, Yang Y, Sun G. Recent insights into antibiotic resistance in Helicobacter pylori eradication. Gastroenterol Res Pract. 2012;2012. doi:10.1155/2012/585674
8. Alfizah H, Norazah A, Hamizah R, Ramelah M. Resistotype of Helicobacter pylori isolates: the impact on eradication outcome. J Med Microbiol. 2014;63(5):703–709. doi:10.1099/jmm.0.069781-0
9. De Francesco V, Zullo A, Ierardi E, Vaira D. Minimal inhibitory concentration (MIC) values and different point mutations in the 23S rRNA gene for clarithromycin resistance in Helicobacter pylori. Dig Liver Dis. 2009;41(8):610–611. doi:10.1016/j.dld.2009.01.001
10. Boyanova L, Markovska R, Yordanov D, Gergova G, Mitov I. Clarithromycin resistance mutations in Helicobacter pylori in association with virulence factors and antibiotic susceptibility of the strains. Microb Drug Resist. 2016;22(3):227–232. doi:10.1089/mdr.2015.0199
11. Matta AJ, Zambrano DC, Pazos AJ. Punctual mutations in 23S rRNA gene of clarithromycin-resistant Helicobacter pylori in Colombian populations. World J Gastroenterol. 2018;24(14):1531. doi:10.3748/wjg.v24.i14.1531
12. Kim JM, Kim JS, Kim N, et al. Gene mutations of 23S rRNA associated with clarithromycin resistance in Helicobacter pylori strains isolated from Korean patients. J Microbiol Biotechnol. 2008;18(9):1584–1589.
13. Rimbara E, Noguchi N, Kawai T, Sasatsu M. Novel mutation in 23S rRNA that confers low-level resistance to clarithromycin in Helicobacter pylori. Antimicrob Agents Chemother. 2008;52(9):3465–3466. doi:10.1128/AAC.00445-08
14. Tsugawa H, Suzuki H, Muraoka H, et al. Enhanced bacterial efflux system is the first step to the development of metronidazole resistance in Helicobacter pylori. Biochem Biophys Res Commun. 2011;404(2):656–660. doi:10.1016/j.bbrc.2010.12.034
15. Rimbara E, Noguchi N, Kawai T, Sasatsu M. Fluoroquinolone resistance in Helicobacter pylori: role of mutations at position 87 and 91 of GyrA on the level of resistance and identification of a resistance conferring mutation in GyrB. Helicobacter. 2012;17(1):36–42. doi:10.1111/j.1523-5378.2011.00912.x
16. Tseng Y, Wu D, Chang C, et al. Amoxicillin resistance with β‐lactamase production in Helicobacter pylori. Eur J Clin Invest. 2009;39(9):807–812. doi:10.1111/j.1365-2362.2009.02166.x
17. Toledo H, López-Solís R. Tetracycline resistance in Chilean clinical isolates of Helicobacter pylori. J Antimicrob Chemother. 2009;65(3):470–473. doi:10.1093/jac/dkp457
18. Heep M, Beck D, Bayerdorffer E, Lehn N. Rifampin and rifabutin resistance mechanism in Helicobacter pylori. Antimicrob Agents Chemother. 1999;43(6):1497–1499.
19. Graham DY, Lee Y, Wu M. Rational Helicobacter pylori therapy: evidence-based medicine rather than medicine-based evidence. Clin Gastroenterol Hepatol. 2014;12(2):177–186.e3. doi:10.1016/j.cgh.2013.05.028
20. Fadilah N, Hanafiah A, Razlan H, Wong ZQ, Mohamed Rose I, Rahman MM. Multiplex PCR for detection of Helicobacter pylori infection in gastric biopsies with lower inflammatory score. Br J Biomed Sci. 2016;73(4):180–187. doi:10.1080/09674845.2016.1220705
21. Kim SY, Woo CW, Lee YM, et al. Genotyping CagA, VacA subtype, IceA1, and BabA of Helicobacter pylori isolates from Korean patients, and their association with gastroduodenal diseases. J Korean Med Sci. 2001;16(5):579–584. doi:10.3346/jkms.2001.16.5.579
22. Francesco VD, Zullo A, Hassan C, Giorgio F, Rosania R, Ierardi E. Mechanisms of Helicobacter pylori antibiotic resistance: an updated appraisal. World J Gastrointest Pathophysiol. 2011;2(3):35–41. doi:10.4291/wjgp.v2.i3.35
23. Kim JM, Kim JS, Kim N, Jung HC, Song IS. Distribution of fluoroquinolone MICs in Helicobacter pylori strains from Korean patients. J Antimicrob Chemother. 2005;56(5):965–967. doi:10.1093/jac/dki334
24. Monno R, Fumarola L, Capolongo C, et al. Susceptibility of Helicobacter pylori to antibiotics including tigecycline. J Med Microbiol Diagn. 2015;S5:1.
25. Chisholm SA, Owen RJ. Frequency and molecular characteristics of ciprofloxacin-and rifampicin-resistant Helicobacter pylori from gastric infections in the UK. J Med Microbiol. 2009;58(10):1322–1328. doi:10.1099/jmm.0.011270-0
26. CLSI. Performance Standards for Antimicrobial Susceptibility Testing; Twentieth Informational Supplement. MIC Interpretive Standards for Helicobacter Pylori. Wayne (PA): Clinical and Laboratory Standards Institute (CLSI); 2010.
27. Marais A, Bilardi C, Cantet F, Mendz GL, Mégraud F. Characterization of the genes rdxA and frxA involved in metronidazole resistance in Helicobacter pylori. Res Microbiol. 2003;154(2):137–144. doi:10.1016/S0923-2508(03)00030-5
28. Ribeiro ML, Vitiello L, Miranda MC, et al. Mutations in the 23S rRNA gene are associated with clarithromycin resistance in Helicobacter pylori isolates in Brazil. Ann Clin Microbiol Antimicrob. 2003;2(1):11. doi:10.1186/1476-0711-2-11
29. Wueppenhorst N, Stueger H, Kist M, Glocker E. Identification and molecular characterization of triple-and quadruple-resistant Helicobacter pylori clinical isolates in Germany. J Antimicrob Chemother. 2009;63(4):648–653. doi:10.1093/jac/dkp003
30. Akopyanz N, Bukanov NO, Westblom TU, Kresovich S, Berg DE. DNA diversity among clinical isolates of Helicobacter pylori detected by PCR-based RAPD fingerprinting. Nucleic Acids Res. 1992;20(19):5137–5142. doi:10.1093/nar/20.19.5137
31. Peleteiro B, Bastos A, Ferro A, Lunet N. Prevalence of Helicobacter pylori infection worldwide: a systematic review of studies with national coverage. Dig Dis Sci. 2014;59(8):1698–1709. doi:10.1007/s10620-014-3063-0
32. Alfizah H, Rizal A, Isa M, Aminuddin A, Jasmi A, Ramelah M. Four year analysis of Helicobacter pylori infection among patients with dyspepsia in Universiti Kebangsaan Malaysia Medical Centre. Med Health. 2010;5:13–21.
33. Sasidharan S, Lachumy SJT, Ravichandran M, Latha LY, Gegu SRS. Epidemiology of Helicobacter pylori among multiracial community in Northern Peninsular, Malaysia: effect of age across race and gender. Asian Pac J Trop Med. 2011;4(1):72–75. doi:10.1016/S1995-7645(11)60037-0
34. Yu CJ, Yan P, Yee LY. Prevalence of Helicobacter pylori infection among patients attending gastroenterology endoscopy unit at Serdang Hospital. Malaysian J Med Health Sci. 2015;11(1):11–17.
35. Huang JQ, Hunt RH. Treatment after failure: the problem of “non-responders”. Gut. 1999;45 Suppl 1:I40–I44.
36. Ghotaslou R, Leylabadlo HE, Asl YM. Prevalence of antibiotic resistance in Helicobacter pylori: a recent literature review. World J Methodol. 2015;5(3):164–174. doi:10.5662/wjm.v5.i3.164
37. Ahmad N, Zakaria WR, Mohamed R. Analysis of antibiotic susceptibility patterns of Helicobacter pylori isolates from Malaysia. Helicobacter. 2011;16(1):47–51. doi:10.1111/j.1523-5378.2010.00816.x
38. Teh X, Khosravi Y, Lee WC, et al. Functional and molecular surveillance of Helicobacter pylori antibiotic resistance in Kuala Lumpur. PLoS One. 2014;9(7):e101481. doi:10.1371/journal.pone.0101481
39. Binh TT, Shiota S, Nguyen LT, et al. The incidence of primary antibiotic resistance of Helicobacter pylori in Vietnam. J Clin Gastroenterol. 2013;47(3):233–238. doi:10.1097/MCG.0b013e3182676e2b
40. Vilaichone R, Gumnarai P, Ratanachu-Ek T, Mahachai V. Nationwide survey of Helicobacter pylori antibiotic resistance in Thailand. Diagn Microbiol Infect Dis. 2013;77(4):346–349. doi:10.1016/j.diagmicrobio.2013.08.010
41. Ang TL, Fock KM, Ang D, Kwek ABE, Teo EK, Dhamodaran S. The changing profile of Helicobacter pylori antibiotic resistance in Singapore: a 15‐year study. Helicobacter. 2016;21(4):261–265. doi:10.1111/hel.12291
42. Goh K, Navaratnam P. High Helicobacter pylori resistance to metronidazole but zero or low resistance to clarithromycin, levofloxacin, and other antibiotics in Malaysia. Helicobacter. 2011;16(3):241–245. doi:10.1111/j.1523-5378.2011.00841.x
43. Thung I, Aramin H, Vavinskaya V, et al. the global emergence of Helicobacter pylori antibiotic resistance. Aliment Pharmacol Ther. 2016;43(4):514–533. doi:10.1111/apt.13497
44. Kim JJ, Kim JG, Kwon DH. Mixed‐infection of antibiotic susceptible and resistant Helicobacter pylori isolates in a single patient and underestimation of antimicrobial susceptibility testing. Helicobacter. 2003;8(3):202–206.
45. Rimbara E, Noguchi N, Tanabe M, Kawai T, Matsumoto Y, Sasatsu M. Susceptibilities to clarithromycin, amoxycillin and metronidazole of Helicobacter pylori isolates from the antrum and corpus in Tokyo, Japan, 1995–2001. Clin Microbiol Infect. 2005;11(4):307–311. doi:10.1111/j.1469-0691.2005.01099.x
46. Selgrad M, Tammer I, Langner C, et al. Different antibiotic susceptibility between antrum and corpus of the stomach, a possible reason for treatment failure of Helicobacter pylori infection. World J Gastroenterol. 2014;20(43):16245–16251. doi:10.3748/wjg.v20.i43.16245
47. Mascellino MT, Porowska B, De Angelis M, Oliva A. Antibiotic susceptibility, heteroresistance, and updated treatment strategies in Helicobacter pylori infection. Drug Des Devel Ther. 2017;11:2209. doi:10.2147/DDDT.S136240
48. Karczewska E, Wojtas-Bonior I, Sito E, Zwolińska-Wcisło M, Budak A. Primary and secondary clarithromycin, metronidazole, amoxicillin and levofloxacin resistance to Helicobacter pylori in southern Poland. Pharmacol Rep. 2011;63(3):799–807.
49. Di Giulio M, Di Campli E, Di Bartolomeo S, et al. In vitro antimicrobial susceptibility of Helicobacter pylori to nine antibiotics currently used in Central Italy. Scand J Gastroenterol. 2016;51(3):263–269. doi:10.3109/00365521.2015.1092577
50. Butlop T, Mungkote N, Chaichanawongsaroj N. Analysis of allelic variants of rdxA associated with metronidazole resistance in Helicobacter pylori: detection of common genotypes in rdxA by multiplex allele-specific polymerase chain reaction. Genet Mol Res. 2016;15(3). doi:10.4238/gmr.15038674
51. De Francesco V, Margiotta M, Zullo A, et al. Clarithromycin-resistant genotypes and eradication of Helicobacter pylori. Ann Intern Med. 2006;144(2):94–100. doi:10.7326/0003-4819-144-2-200601170-00006
52. Fujimura S, Kato S, Iinuma K, Watanabe A. In vitro activity of fluoroquinolone and the gyrA gene mutation in Helicobacter pylori strains isolated from children. J Med Microbiol. 2004;53(10):1019–1022. doi:10.1099/jmm.0.45642-0
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.