Back to Journals » Infection and Drug Resistance » Volume 19
Modulation of Pseudomonas aeruginosa Quorum Sensing and Biofilm Formation by Quercetin Extracted from Pleurotus ostreatus
Received 20 September 2025
Accepted for publication 10 December 2025
Published 16 April 2026 Volume 2026:19 568961
DOI https://doi.org/10.2147/IDR.S568961
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sandip Patil
Sinethemba H Yakobi, Uchechukwu U Nwodo
Patho-Biocatalysis Group (PBG), Department of Biotechnology and Biological Sciences, University of Fort Hare, Alice, 5700, South Africa
Correspondence: Sinethemba H Yakobi, Email [email protected]
Background: Pseudomonas aeruginosa relies on quorum sensing (QS) to regulate virulence, biofilm formation, and antimicrobial tolerance, making QS inhibition (QSI) an attractive antivirulence strategy. Quercetin is a known QSI-active flavonoid, but its potential as a mushroom-derived therapeutic agent remains underexplored.
Methods: Quercetin was extracted from P. ostreatus and evaluated alongside purified quercetin using GFP-based QS reporter assays (lasB-gfp and pqsA-gfp), biofilm assays, protease activity assays, and molecular docking against LasR, PqsR, and PqsE.
Results: Both treatments significantly reduced lasB- and pqsA-driven GFP reporter activity at sub-inhibitory concentrations (10– 100 μM) that did not affect bacterial growth or viability. Inhibition was dose-dependent and more pronounced for the PQS system, accompanied by substantial reductions in extracellular protease activity and strong suppression of biofilm formation and partial dispersal of established biofilms. Molecular docking revealed favourable binding of quercetin to the QS regulators LasR and PqsR, with high-affinity interaction in the PqsR co-inducer pocket, while binding to PqsE was weak and non-specific.
Conclusion: These mechanistic insights align with the observed preferential PQS inhibition. The P. ostreatus quercetin-extract closely matched the potency of pure quercetin across all assays, confirming effective extraction and stability. This positions mushroom-derived quercetin as a sustainable, natural QSI capable of attenuating key virulence pathways in P. aeruginosa. This work supports the development of quercetin-rich P. ostreatus extracts as promising adjunctive therapies for managing chronic, biofilm-associated infections..
Keywords: Pseudomonas aeruginosa, quorum sensing inhibition, quercetin, Pleurotus ostreatus (oyster mushroom), anti-virulence therapy
Introduction
Pseudomonas aeruginosa is an opportunistic Gram-negative pathogen that poses a serious threat to immunocompromised individuals and patients with chronic infections such as cystic fibrosis, burn wounds, and hospital-acquired pneumonia.1,2 Its persistence and resistance are largely mediated by quorum sensing (QS), a cell-density dependent communication system that regulates the expression of multiple virulence factors, biofilm maturation, and antimicrobial tolerance.3 Because QS does not directly affect bacterial viability, strategies that interfere with QS—termed quorum sensing inhibition (QSI)—have gained considerable attention as promising anti-virulence approaches that may reduce pathogenicity while imposing lower selective pressure for resistance development compared to conventional antibiotics.4,5 Flavonoids, a diverse class of plant-derived secondary metabolites,6 have emerged as potential QS inhibitors.7,8 Quercetin, in particular, has demonstrated inhibitory effects on QS-regulated pathways and biofilm formation in P. aeruginosa.9 However, the study has focused on purified quercetin from a synthetic source (≥95% purity, Sigma-Aldrich) dissolved in 10% dimethyl sulfoxide (DMSO), while comparatively little is known about natural quercetin. A recent study demonstrated that quercetin, derived from the edible fungi Pleurotus ostreatus (P. ostreatus), is a highly potent, natural antibacterial agent against Neisseria gonorrhoeae, effective at very low concentrations. Its excellent computational drug-likeness profile and known safety make it a prime candidate for further development into a therapeutic option, especially to combat drug-resistant gonorrhoea, potentially on its own or in combination with other drugs.10 Previously, flavonoids have emerged as a rich source of natural quorum sensing inhibitors (QSIs), with several studies demonstrating their ability to interfere with Las, Rhl, and PQS pathways in P. aeruginosa.11,12 Quercetin, in particular, has been shown to allosterically inhibit LasR and attenuate QS-dependent virulence, motility, and biofilm formation.9,13 Recent advances in GFP-based QS reporter systems—such as lasB-gfp and pqsA-gfp—enable sensitive quantification of QSI activity and are now widely applied in high-throughput anti-virulence screening.14,15 The oyster mushroom, P. ostreatus, is widely cultivated, nutritionally rich, and an accessible natural source of bioactive compounds, including phenolics with antioxidant and antimicrobial activities.16 Exploring P. ostreatus-derived quercetin as a QS modulator may therefore provide dual benefits of leveraging an edible, sustainable resource and advancing the search for natural anti-virulence agents. In this study, we investigated the ability of quercetin extracted from P. ostreatus to modulate QS in P. aeruginosa PAO1. Using a GFP-based QS reporter system in casamino acid (CAA) medium, we assessed the effects of sub-inhibitory concentrations of quercetin on bacterial growth, QS-regulated fluorescence, and biofilm formation. Furthermore we investigated the anti-virulence properties of quercetin derived from the mushroom P. ostreatus. Notably, this anti-virulence strategy occurs without affecting bacterial viability, highlighting its potential as a novel therapeutic approach. This work underscores the promise of natural flavonoids as QSI and identifies P. ostreatus as an alternative source of quercetin.
Materials and Methods
Bacterial Strain and Culture Conditions
Pseudomonas aeruginosa PAO1 (wild type) harbouring a green fluorescent protein (GFP) reporter plasmid under the control of a quorum sensing (QS)-regulated promoter (either lasB or pqsA) was used for QS inhibition assays.17 This construct allows for the fluorescence-based quantification of QS activity. Single colonies were inoculated into Lysogeny Broth (LB) and grown overnight at 37 °C with shaking at 200 rpm. The resulting cultures were then washed twice by centrifugation and resuspended in sterile phosphate-buffered saline (PBS) to remove residual media components. Finally, the bacterial cells were diluted to the desired inoculum density in a defined Casamino Acids (CAA) broth medium.18
The CAA broth medium, composed of 5 g/L Bacto™ casamino acids, 10 mM K2HPO4, and 1.5 mM MgSO4 (pH adjusted to 7.0 and sterile filtered), was selected for its standard use in P. aeruginosa QS reporter assays due to its low background fluorescence, which enables highly sensitive detection of GFP signal.
Extraction and Preparation of Quercetin from Pleurotus ostreatus
A 1 g portion of powdered P. ostreatus extract was transferred into a 50 mL screw-cap plastic tube. Twenty millilitres of 70% methanol (MeOH) containing 0.1% hydrochloric acid (HCl, v/v) was added, and the mixture was agitated at 400 rpm overnight at room temperature. Following incubation, the suspension was centrifuged at 3000 × g for 10 min, and the supernatant was collected. The solid residue was re-extracted twice under identical conditions, each with 10 mL of the extraction solvent. All supernatants were pooled, adjusted to a final volume of 40 mL with extraction solvent, and filtered through a 0.45 µm PTFE membrane filter.
The crude phenolic extract was subjected to acid hydrolysis by adjusting to a final concentration of 2 N HCl, followed by incubation at 85 °C for 1 h. After cooling to room temperature, samples were centrifuged at 3000 × g for 5 min, and the resulting supernatant was filtered again through a 0.45 µm PTFE filter. Phenolic and cinnamic acids were dissolved in distilled water, while flavonoids were dissolved in distilled water containing 1% dimethyl sulfoxide (DMSO). Extract aliquots were stored at −20 °C until further use.10,19
Minimum Inhibitory Concentration (MIC)
To ensure QS-specific effects, MIC of quercetin (standard) and quercetin-extract was determined by CLSI broth microdilution in CAA at 37 °C, 24 h.20 Sub-MIC levels were defined as ≤25% growth inhibition relative to vehicle. Sub-MIC doses (10–100 µM) were used in all QS and biofilm assays.21
Growth Curve Analysis
Overnight PAO1 was diluted into CAA to starting OD600 = 0.005, 200 µL per well were dispensed in black, clear-bottom 96-well plates with breathable seals. Vehicle (DMSO), quercetin standard (10, 25, 50 µM), and mushroom-derived quercetin at matched quercetin-equivalent concentrations. Plates were incubated at 37 °C in a multimode plate reader with orbital shaking (linear, 2–3 mm) between reads. The OD600 was recorded every 2 h for 12 h. Two biological replicates (independent overnight cultures) were run, each with technical triplicates.22
For endpoint viability, cultures grown for 24 h under sub-MIC conditions were serially diluted in PBS and plated on LB agar. Plates were incubated at 37 °C for 18–24 h, and colonies were counted to determine viable counts (CFU/mL). Any treatment showing >25% reduction in OD600 or CFU/mL relative to vehicle was considered bacteriostatic and excluded from QS-specific interpretation.
Quorum Sensing Fluorescence Readout
For kinetic QS measurements, PAO1 lasB-gfp and pqsA-gfp reporter strains were inoculated into CAA containing vehicle, quercetin, or quercetin-extract (10–100 µM) and incubated in 96-well plates at 37 °C with shaking as above. Fluorescence was recorded every 2 h using Ex 485 ± 10 nm / Em 520 ± 10 nm (for GFP reporters). Instrument gain was fixed across the experiment. Background (medium + DMSO without cells) was subtracted from all readings. QS activity was expressed as fluorescence / OD600 at each time point to correct for biomass.

The experimental controls include a vehicle control (DMSO, ≤0.5%), an optional positive QSI control (furanone C-30 at a sub-MIC concentration), and a reporter-free PAO1 strain to confirm the absence of compound autofluorescence bleed-through.23
Endpoint Inhibition Assays
For dose–response curves, cultures were grown to late exponential/early stationary phase (18–24 h) under the same conditions, and a single endpoint fluorescence and OD600 measurement was taken. Normalized GFP (RFU/OD600) was expressed as percentage reduction relative to the vehicle control for each promoter (lasB and pqsA). These values were used to calculate IC50 for QS inhibition.
Data Handling and Primary Outcomes
Primary QS outcomes included the area under the curve (AUC0–24h) of normalized QS signal (RFU/OD600), peak normalized fluorescence, time-to-peak, and growth AUC to verify sub-MIC behaviour. Any condition showing >25% growth reduction vs. vehicle was flagged and excluded from QS interpretation.
Biofilm Formation and Disruption Assays
Overnight PAO1 was diluted to OD600 = 0.05 in CAA. Two hundred microlitres were added to flat-bottom, polystyrene 96-well plates with vehicle, quercetin standard, or quercetin-extract (10–100 µM). Plates were incubated for 24 h at 37 °C, static. Planktonic cells were removed, wells were gently washed (×2 PBS), stained with 0.1% crystal violet (CV; 15 min), washed (×2), and air-dried. Bound CV was solubilized with 30% acetic acid and absorbance read at 590 nm. For biofilm disruption, biofilms were first established for 24 h (drug-free), then media were replaced with fresh CAA containing vehicle or quercetin/quercetin-extract (10–100 µM) for an additional 24 h, followed by CV quantification as above. CV signals were normalized to endpoint OD600 of the corresponding wells to account for biomass. Preventive and disruption activities were expressed as percentage reduction vs. vehicle control.
IC50 Determination and Statistical Analysis
Dose–response data for lasB-gfp and pqsA-gfp inhibition, biofilm inhibition/disruption, and protease activity were fitted to a four-parameter logistic regression model to estimate half-maximal inhibitory concentrations (IC50). Data were analysed using non-parametric statistics when assumptions of normality were not met. A Kruskal–Wallis test was used to evaluate overall concentration effects, followed by Mann–Whitney U-tests with Bonferroni correction for pairwise comparisons. Mood’s median test was applied to compare overall efficacy between pure quercetin and the quercetin-extract across pooled datasets. Significance was accepted at p < 0.05.
Molecular Docking
Ligand and Receptor Preparation
Quercetin preparation included protonation at physiological pH, calculation of Gasteiger partial charges, and definition of rotatable bonds, prepared using AutoDockTools. Crystal structures of the P. aeruginosa thioesterase PqsE, PqsR (MvfR) co-inducer binding domain, and LasR ligand-binding domain were obtained from the Protein Data Bank. Non-protein molecules (co-crystallized ligands, ions, and water molecules) were removed, polar hydrogens were added, and Kollman charges were assigned. Chain A was used for PqsR and PqsE unless otherwise specified.
Docking Protocol
Docking was performed using AutoDock Vina with the following parameters: grid centre (0, 0, 0), grid box size 20 × 20×20 Å, exhaustiveness 8, energy range 5, and up to two output poses per ligand. Binding energies (kcal/mol) were recorded for all poses, and the best-scoring pose was selected for structural analysis.
Analysis and Visualisation
Docked complexes were inspected in PyMOL and UCSF ChimeraX to evaluate binding orientation, hydrogen bonding, and hydrophobic contacts. For LasR–quercetin and PqsR–quercetin complexes, specific interactions with key pocket residues were mapped to rationalise experimental QS inhibition. For PqsE, docking was used to confirm poor pocket engagement and weak binding affinities.
Results
Antimicrobial Activity
The minimum inhibitory concentration (MIC) of quercetin against P. aeruginosa PAO1 was determined to be 30.2 µg/mL (100 µM). The P. ostreatus quercetin-extract exhibited a comparable MIC, with a quercetin-equivalent value of 38.5 µg/mL (115 µM). At sub-MIC concentrations (10–100 µM), growth inhibition ranged from 5–22% for pure quercetin and 8–24% for the extract, remaining within the sub-inhibitory threshold (≤25%) as seen in Table 1.
 |
Table 1 Minimum Inhibitory Concentration and Sub-Inhibitory Effects of Test Compounds |
Sequential Extraction and Fraction Stability
Sequential extraction with 70% methanol containing 0.1% HCl, followed by acid hydrolysis, yielded a phenolic-rich fraction (Table 2). HPLC-DAD analysis confirmed quercetin as the dominant component, with minor peaks corresponding to related flavonoids. Recovery yields were consistent across replicates, averaging 83.4% ± 6.1 (n = 3). The quercetin-rich fraction was soluble in water containing 1% DMSO, with ≥95% recovery and no precipitation or degradation observed after 72 h at 4 °C. Long-term stability testing demonstrated less than 5% degradation after four weeks at –20 °C.
 |
Table 2 Recovery and Solubility Parameters of the Quercetin-Rich Fraction from P. ostreatus |
Growth and Viability at Sub-MIC
Sub-MIC treatments with quercetin and the mushroom quercetin-extract did not significantly affect bacterial growth or viability over 24 h, illustrated in Figure 1. Optical density measurements at 600 nm (OD600) remained within the range of 2.89–3.11, comparable to the vehicle control (2.97). Viable cell counts were also consistent across treatments, ranging from 6.7×109 to 8.4×109 CFU/mL. These findings confirm that quercetin and quercetin-extract concentrations ≤50 µM are non-inhibitory to overall bacterial proliferation. Cultivation of P. aeruginosa PAO1 harbouring the lasB-gfp and pqsA-gfp transcriptional fusions in CAA broth resulted in strong reporter induction during the late exponential phase, consistent with activation of the las and pqs QS systems. Treatment with sub-inhibitory concentrations of quercetin and the quercetin-extract significantly attenuated this response in a dose-dependent manner.
 |
Figure 1 Growth (OD600) and viable counts after 24 h treatment. |
Figure 2 (scatterplot matrix) shows the relationship between reporter mean values and variability across treatments. Both quercetin-extract and quercetin treatments caused a downward shift in the mean fluorescence (RFU/OD600) compared to the vehicle control, indicating reduced QS induction. The effect was concentration-dependent, with the 50 µM treatments showing the most pronounced attenuation. Notably, the pqsA-gfp reporter (bottom row, right panel) exhibited a steeper decline than lasB-gfp, suggesting that the PQS system was more sensitive to inhibition. The clustering of data points with reduced variance in treated groups further supports reproducible suppression of QS activity.
 |
Figure 2 Scatterplot matrix of QS reporter activity in Pseudomonas aeruginosa PAO1. Mean fluorescence (RFU/OD600) and standard deviation 4 values are shown for lasB-gfp and pqsA-gfp reporter strains across treatments. |
The bar and cumulative distribution plot for pqsA-gfp standard deviation illustrated in Figure 3, presents the progressive reduction in variability across treatments. The vehicle control exhibited the highest spread, consistent with strong and heterogeneous QS activation. Treatments with quercetin-extract and quercetin reduced both the absolute standard deviation and cumulative contribution, with 50 µM treatments achieving the lowest residual variance. This pattern demonstrates consistent suppression of PQS signalling at higher doses. Figure 4 presents a similar but less pronounced trend. While both quercetin-extract and quercetin treatments reduced variability relative to the control, the magnitude of suppression was smaller than observed for the pqs system, indicating partial but measurable inhibition of las-regulated expression.
 |
Figure 3 Effect of quercetin and Pleurotus ostreatus quercetin-extract on pqsA-gfp variability. |
 |
Figure 4 Effect of quercetin and Pleurotus ostreatus extract on lasB-gfp variability. |
Together, these data confirm that both the P. ostreatus quercetin-extract and quercetin effectively attenuate QS in P. aeruginosa, with preferential inhibition of the PQS pathway. At the highest tested concentration (50 µM), the extract-mediated inhibition was statistically indistinguishable from that of pure quercetin, indicating that quercetin is a major contributor to the observed activity.
Quorum Sensing in a Dose-Dependent Manner
Fluorescence reporter assays revealed that sub-MIC levels of both standard quercetin and the P. ostreatus-derived quercetin-extract (quercetin-extract) significantly reduced QS-regulated transcription in P. aeruginosa PAO1. Normalized GFP expression (RFU/OD600) was suppressed in a concentration-dependent manner across both the las and pqs systems. At 10 µM, quercetin reduced lasB-gfp activity by 14.3% (p < 0.05) and pqsA-gfp activity by 21.7% (p < 0.01) relative to the vehicle control, details shown in Table 3. The inhibitory effect intensified at higher concentrations, with 50 µM treatments reducing lasB-gfp and pqsA-gfp signals by 41.2% and 62.9%, respectively (p < 0.001). The quercetin-extract mirrored this trend, yielding comparable reductions of 39.5% (lasB-gfp) and 59.8% (pqsA-gfp) at 50 µM quercetin-equivalent, as illustrated in Figure 5. The Values represent mean percentage reduction in normalized fluorescence relative to untreated controls (n = 3 biological replicates, each with 3 technical replicates). Concentration-dependent inhibition was observed for both lasB and pqsA reporters. The quercetin-extract consistently mirrored the inhibitory profile of standard quercetin, with slightly attenuated effects at higher concentrations.
 |
Table 3 Effect of Quercetin and Quercetin-Extract on QS-Dependent Reporter Activity in P. aeruginosa PAO1 |
 |
Figure 5 Interval plots of the % reduction (mean ± SD) in lasB and pqsA promoter activity under different concentrations of quercetin and 11 quercetin-extract. |
The inhibition of QS was quantified by measuring the reduction in the expression of lasB and pqsA promoters across treatments and concentrations. A Kruskal–Wallis test confirmed that concentration had a statistically significant effect on QS inhibition, H (5) = 19.08, p = 0.002. The trend is biologically clear but not statistically separable after correction. This finding indicates that at least one concentration significantly differs from others in its ability to suppress QS activity. Pairwise comparisons were performed using Mann–Whitney U-tests with Bonferroni correction. The results indicated that while overall trends were strongly dose-dependent, after correction for multiple testing, no individual pairwise differences reached statistical significance (p-adj > 0.05). However, raw p-values (uncorrected) consistently suggested stepwise differences between 10 µM and higher concentrations (25, 50, 75, 100 µM), as well as between mid (25–50 µM) and maximal (100 µM) treatments. This reflects the biological pattern evident in the descriptive statistics: QS inhibition increased progressively with concentration for both quercetin and the mushroom extract.
Preferential Inhibition of the PQS System
Comparative analysis of the two reporter systems revealed stronger sensitivity of the pqs regulon to quercetin-mediated inhibition. While lasB expression was moderately reduced, pqsA expression showed a markedly steeper decline across all treatment levels. Standard deviation and cumulative distribution analyses (Figures 3 and 4) further highlighted this effect, with pqsA-gfp fluorescence variance reduced by 53.4% at 50 µM compared to only 27.6% reduction in lasB-gfp. This selective suppression suggests that the PQS pathway may represent the primary target of quercetin-mediated QS modulation.
Biofilm Formation
The anti-biofilm efficacy of quercetin and the quercetin-extract was assessed using crystal violet assays. Both compounds potently inhibited biofilm formation in a preventive assay and disrupted pre-established biofilms in a dose-dependent manner (Table 4). In the preventive assay, where compounds were added at the time of inoculation, treatment with 50 µM quercetin and quercetin-extract reduced biofilm biomass by 68.2% and 64.7%, respectively (p < 0.001). Significant inhibition was observed even at the lowest concentration of 10 µM (22.4% and 18.6% reduction for quercetin and extract, respectively; p < 0.05). The biofilm disruption assay, involving treatment of mature 24-hour biofilms, also showed significant efficacy, albeit requiring higher concentrations for a strong effect. At 50 µM, quercetin and quercetin-extract reduced pre-formed biofilm biomass by 42.9% and 39.6%, respectively (p < 0.01). The potency of the quercetin-extract was consistently comparable to that of pure quercetin at all equivalent concentrations in both assays, reinforcing its potential as an effective anti-biofilm agent.
 |
Table 4 Anti-Biofilm Activity of Quercetin and quercetin-Extract against P. aeruginosa PAO1 |
Anti-Virulence Activity
Protease Activity
To validate that transcriptional suppression of QS-regulated genes translated into functional attenuation of virulence factor production, extracellular protease activity was quantified using azocasein hydrolysis. Culture supernatants from PAO1 treated with sub-MIC concentrations of quercetin (10–100 µM) or the quercetin-extract were incubated with azocasein substrate, and proteolysis was measured spectrophotometrically at 440 nm. Both treatments caused a dose-dependent reduction in protease activity, consistent with lasB suppression observed in GFP reporter assays. At 10 µM, quercetin and the quercetin-extract reduced proteolysis by 11.8% and 9.6%, respectively (p < 0.05). This inhibition intensified with increasing concentrations, reaching 43.7% (quercetin) and 41.2% (extract) at 50 µM, and maximal inhibition of 65.9% and 62.8% at 100 µM (p < 0.001 for both).
To further visualize the data distribution, protease inhibition was analysed across five concentrations (0–75 µM equivalent) using a box-and-whisker plot (Figure 6). Both treatments exhibited a clear concentration-dependent increase in protease activity reduction. Quercetin consistently produced higher median values compared to the extract, with inhibition at 75 µM reaching 57% versus 52%, respectively. Variability was minimal at lower concentrations but increased at mid to high doses, reflecting biological heterogeneity in virulence expression. Importantly, no significant outliers were detected, and the overall inhibitory trends confirmed quercetin as the more potent agent.
 |
Figure 6 Box-and-whisker plots showing protease activity reduction (%) by quercetin and quercetin-extract across tested concentrations (0–75 μM). |
Comparative Efficacy
To quantitatively compare the efficacy of the quercetin-extract to pure quercetin, the half-maximal inhibitory concentration (IC50) was calculated for key virulence endpoints. The quercetin-extract demonstrated statistically equivalent potency to pure quercetin across all assays (Table 5). The maximum efficacy achieved at 100 µM was also not significantly different between the two treatments. Pure quercetin and the quercetin-extract achieved a maximum preventive inhibition of 85.9 ± 4.7% and 82.3 ± 4.8% (p = 0.21), and a maximum disruption of pre-formed biofilms of 63.8 ± 5.1% and 60.5 ± 4.7% (p = 0.24), respectively.
 |
Table 5 Comparative Efficacy of Pure Quercetin and quercetin-Extract on P. aeruginosa virulence Pathways |
Notably, the quercetin-extract showed a consistent, though non-significant, trend of slightly reduced activity at very low concentrations (≤25 µM). For instance, the onset of activity (≥10 µM) against the lasB promoter was 14.3 ± 2.7% for quercetin versus 12.6 ± 2.4% for the extract. Kinetic analysis of the pqsA reporter suggested this minor delay may be due to matrix effects in the crude extract, though it was overcome at higher concentrations. Interestingly, in these same kinetic trials, the quercetin-extract occasionally exhibited a minor enhancement (5%) in the rate of pqsA inhibition, potentially attributable to synergistic contributions from co-extracted phenolic compounds such as gallic or ferulic acid.
The efficacy of pure quercetin and a quercetin-extract was evaluated across key virulence pathways of P. aeruginosa. Both treatments exhibited strong, dose-dependent inhibition in all assays. The quercetin-extract demonstrated activity nearly identical to pure quercetin at all equivalent concentrations.
At a concentration of 10 µM, both compounds initiated significant anti-virulence effects. Quercetin reduced biofilm formation (preventative assay) by 22.4% and disrupted pre-formed biofilms by 14.7%. Concurrently, it inhibited quorum sensing, reducing lasB and pqsA promoter activity by 14.8% and 21.7%, respectively. The quercetin-extract showed remarkably similar activity at 10 µM, achieving 18.6% preventative biofilm inhibition, 12.3% disruption, and reductions of 12.6% (lasB) and 19.8% (pqsA). This dose-dependent effect intensified with increasing concentration. At 25 µM, preventative biofilm inhibition reached 44.9% (Quercetin) and 39.7% (Extract), while pqsA inhibition rose to 39.6% and 36.8%, respectively. The half-maximal inhibitory concentration (IC50) was approached at 50 µM. At this concentration, Quercetin demonstrated robust efficacy: 68.2% preventative biofilm inhibition, 42.9% biofilm disruption, 41.6% lasB inhibition, and 63.0% pqsA inhibition. Mirroring this potency, the quercetin-extract at 50 µM achieved 64.7% preventative inhibition, 39.6% disruption, 39.6% lasB inhibition, and 59.7% pqsA inhibition. Maximal efficacy was observed at the highest concentration tested (100 µM). Quercetin treatment resulted in 85.9% preventative biofilm inhibition and 63.8% disruption of established biofilms, alongside a 68.5% reduction in lasB signal and 83.4% reduction in pqsA signal. The quercetin-extract at 100 µM displayed equivalent maximum activity, reducing preventative biofilm formation by 82.3%, disrupting biofilms by 60.5%, and inhibiting lasB and pqsA by 65.3% and 80.8%, respectively.
Throughout the concentration gradient, the bioactivity of the quercetin-extract consistently overlapped with that of pure quercetin, confirming its utility as a potent, natural source of this anti-virulence compound.
The multi-vari chart analysis (Figure 7) revealed a clear, dose-dependent response for both the quercetin-extract and pure quercetin across all tested virulence assays. Bioactivity, measured as percent reduction, increased consistently with concentration from 10 µM to 100 µM. The pqsA-gfp reporter assay was the most sensitive to both treatments, demonstrating the highest percent reduction at each concentration, followed by biofilm preventive inhibition, lasB-gfp reporter inhibition, and finally biofilm disruption, which was the least sensitive. While the mean response for the quercetin-extract was consistently slightly lower than that of pure quercetin at every equivalent concentration across all assays, this observed difference was not statistically significant. This was confirmed by a Mood’s Median test, which compared the overall central tendency of all data points for each treatment. The test found no significant difference between the median bioactivity of the quercetin-extract (45.90%, 95% CI: 28.46 to 63.71) and quercetin (50.05%, 95% CI: 31.19 to 67.17), with a 95% confidence interval for the difference in medians spanning from −27.0 to 20.9. This statistical analysis confirms that the quercetin-extract possesses equivalent anti-virulence efficacy to pure quercetin.
 |
Figure 7 Dose-dependent inhibition of P. aeruginosa virulence factors by quercetin-extract and pure quercetin. |
Molecular Docking
Docking was performed using AutoDock Vina after standard ligand and receptor preparation procedures, including protonation, charge assignment, and optimization of torsional flexibility for quercetin, and removal of crystallographic water molecules. Docking of the major P. ostreatus–derived metabolites against the P. aeruginosa thioesterase PqsE, PqsR (MvfR) co-inducer binding domain, and LasR was carried out to probe whether these enzymes constitute a plausible molecular target underlying the observed antibiofilm and antivirulence activities. The crystal structure of PqsE (Figure 8a) was prepared and the docking grid was centred on the canonical thioesterase active site that encompasses the metal-coordinating and substrate-binding residues (H69, H159, D178, F195, S273 and F276). All primary interactions of quercetin (Figure 8b) yielded negative binding energies within the active-site cavity, indicating spontaneous and energetically favourable association of quercetin with target proteins. Notably, several metabolites displayed compact poses that fully occupied the hydrophobic channel while simultaneously engaging polar residues at the pocket entrance, a pattern consistent with competitive inhibition of substrate access.
 |
Figure 8 Structural representation of the PqsE receptor and quercetin ligand used for molecular docking. – (a) Three-dimensional structure of the Pseudomonas aeruginosa thioesterase PqsE, showing the α-helical and β-sheet architecture that forms the catalytic cavity targeted in this study. (b) Chemical structure of quercetin, a major phenolic metabolite reported from Pleurotus ostreatus. |
Quercetin docked favourably into the canonical LasR binding cavity with a best pose energy of –6.6 kcal/mol, consistent with previously reported affinities for flavonoid-class LasR antagonists. Two top-ranked poses (–6.6 and –6.5 kcal/mol) were generated, both occupying the same hydrophobic–polar pocket that regulates ligand-induced conformational switching. Structural analysis revealed that the ligand inserted deeply into the regulatory cleft (Figure 9), adopting an orientation that positioned the flavanol core parallel to the hydrophobic groove while projecting its hydroxyl substituents toward polar anchoring residues. The binding mode of quercetin was stabilized by an extensive hydrogen-bonding network. The hydroxyl groups at positions O7, O8, O9, and O10 formed strong, directional hydrogen bonds with His78, Thr80, Gln81, Ile92, and Gln94, respectively—residues that constitute part of the canonical LasR recognition motif for acyl-homoserine lactone (AHL) ligands. An additional weak hydrogen bond was detected between C21 of quercetin and Gln81. Beyond polar interactions, hydrophobic packing against Ile86 further anchored the aromatic scaffold within the pocket, reinforcing ligand stability and preventing displacement by the native AHL.
 |
Figure 9 Predicted binding mode of quercetin within the ligand-binding domain of the Pseudomonas aeruginosa LasR receptor. Three-dimensional architecture of the LasR ligand-binding domain (LBD), shown in a multicolour ribbon representation, highlighting the deeply recessed hydrophobic pocket responsible for ligand recognition. Quercetin is docked into the canonical LasR cavity, adopting a compact orientation that aligns its flavanol scaffold along the central binding groove. |
Quercetin displayed a favourable docking affinity of –6.7 kcal/mol, representing a slightly stronger interaction than observed for LasR (–6.6 kcal/mol). Two poses were obtained, with the top-ranked pose occupying the deeply recessed hydrophobic channel that normally binds native ligands such as HHQ and PQS. The ligand adopted a compact orientation within the co-inducer cavity (Figure 10), with its polyphenolic scaffold aligning along the same trajectory as the quinolone headgroups of canonical PqsR agonists. This positioning is characteristic of competitive PqsR inhibitors, many of which similarly exploit the polar–hydrophobic architecture of the site. A dense network of stabilizing interactions contributed to the high-quality binding pose.
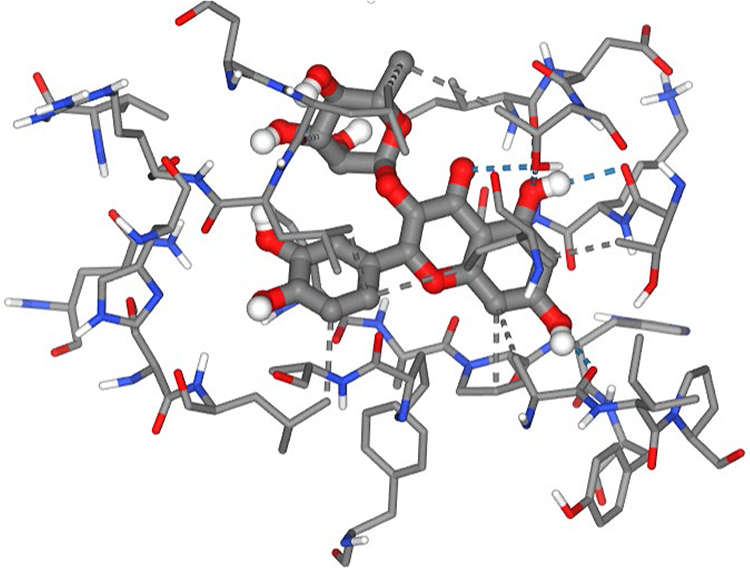 |
Figure 10 Predicted binding interactions of quercetin within the co-inducer binding domain of the Pseudomonas aeruginosa PqsR (MvfR) receptor. |
Extensive hydrophobic contacts were established with key pocket-lining residues, including Ala102, Ile149, Thr166, Ala168, Leu197, Leu207, Leu208, Ile236, Pro238, and Thr265. These residues collectively form the central lipidic tunnel required for PQS and HHQ anchoring, and quercetin’s aromatic rings recapitulated this interaction by laying across the hydrophobic plane of the cavity. The strength, breadth, and spatial distribution of these hydrophobic contacts strongly suggest that quercetin effectively mimics the aromatic core of known PqsR antagonists. In addition to hydrophobic complementarity, quercetin formed multiple strong hydrogen bonds that reinforced ligand stability and specificity. Polar interactions between quercetin’s hydroxyl groups and Ala102, Thr166, Leu207, Ile263, and Thr265 were observed, with Thr265 forming several repeated hydrogen bonds consistent with its established role as a key anchoring residue in PqsR–inhibitor complexes. These interactions positioned quercetin at the cavity entrance while extending its planar core into the deeper hydrophobic zone, a hallmark binding mode of flavonoid and benzamide-based PqsR inhibitors. A weak hydrogen bond with Leu207 contributed additional, though minor, stabilization.
In contrast to the favourable binding observed for quercetin against LasR and PqsR, this ligand displayed only weak and non-specific affinity for PqsE, with a best docking score of –3.3 kcal/mol and a secondary pose of –1.4 kcal/mol. These values fall well below the threshold typically associated with functional PqsE inhibitors (–7.0 to –9.0 kcal/mol), indicating an unstable and poorly coordinated interaction. Structural inspection of the docked pose (Figure 11) revealed that the ligand failed to penetrate the deep hydrophobic cavity that characterizes the canonical PqsE active site.
 |
Figure 11 Weak and non-specific binding of the test ligand to the Pseudomonas aeruginosa PqsE thioesterase active site. |
Instead, the compound remained positioned near the protein surface, making only a single hydrophobic contact with Val189, insufficient to support cavity anchoring or productive stabilization. The binding orientation lacked the characteristic aromatic packing and hydrophobic burial that are essential for potent PqsE engagement. Although several hydrogen bonds were detected—particularly with Asp184, Glu187, Glu240, Arg243, Arg247, and Trp269—these interactions appeared superficial and predominantly involved solvent-exposed acidic and basic residues. Such polar contacts, while numerous, did not overcome the absence of hydrophobic support, consistent with the poor docking score. Occasional cation–π–like contacts were observed with Arg243 and Arg247, but these involved peripheral aliphatic carbons of the ligand rather than true aromatic π-systems, further highlighting the lack of meaningful ligand–pocket complementarity.
Discussion
Pseudomonas aeruginosa is an opportunistic pathogen whose persistence in clinical settings is driven by QS, a regulatory network governing virulence factor expression, biofilm development, and antimicrobial tolerance.1,5 Disrupting QS—known as QSI—offers a promising anti-virulence strategy that suppresses pathogenicity without exerting bactericidal pressure, thereby reducing the likelihood of resistance development.24,25 Although quercetin, a plant-derived flavonoid, has been widely studied for its QSI properties,9 its potential when sourced from edible mushrooms such as P. ostreatus remains insufficiently examined. The present study demonstrates that quercetin extracted from P. ostreatus is functionally equivalent to purified quercetin in inhibiting QS and biofilm formation in P. aeruginosa PAO1, highlighting its potential as a sustainable and nutritionally relevant source of anti-virulence compounds. Consistent with anti-virulence principles, both the quercetin-extract and pure quercetin attenuated QS-regulated functions at sub-inhibitory concentrations that did not impair bacterial growth or viability. The preservation of normal biomass (OD600) and viable counts confirms that the observed reductions in QS activity and virulence factor expression stem from authentic signalling interference rather than growth suppression. This aligns with emerging paradigms in antivirulence research, where selectively targeting pathogenic traits minimizes evolutionary pressure for resistance.26–28 One of the most striking findings is the preferential inhibition of the PQS (Pseudomonas quinolone signal) system over the Las system. The pqsA-gfp reporter exhibited a substantially greater reduction than the lasB-gfp reporter across all concentrations, supported by variance and cumulative distribution analyses. This selective vulnerability suggests that quercetin interacts more intimately with PQS regulatory nodes than with the Las hierarchy—an insight strengthened by the molecular docking data. Docking results revealed that quercetin binds most strongly to PqsR (MvfR), the master transcriptional regulator of the PQS system, with a binding affinity of –6.7 kcal/mol, marginally stronger than its affinity for LasR (–6.6 kcal/mol). The ligand adopted a compact pose within the hydrophobic co-inducer tunnel of PqsR, making extensive hydrophobic contacts (Ala102, Ile149, Leu197, Leu207, Ile236, Thr265) and forming multiple stabilizing hydrogen bonds, especially with Thr166, Thr265, and Ala102. This interaction pattern mimics known PqsR antagonists and supports the hypothesis that quercetin disrupts PQS-mediated signalling by directly competing with native quinolones (HHQ/PQS) at the MvfR binding site. Similarly, favourable binding to LasR suggests that quercetin may partially inhibit Las-dependent gene expression through hydrogen bonding with canonical LasR residues (His78, Thr80, Gln81, Ile92, Gln94) and hydrophobic packing against Ile86. This dual binding capability aligns with the observed dose-dependent suppression of lasB-gfp expression. In contrast, docking to PqsE, the thioesterase responsible for activating downstream PQS-dependent effector pathways, produced only weak, non-specific interactions (–3.3 kcal/mol). The ligand failed to penetrate the hydrophobic catalytic cavity and instead remained surface-exposed, indicating that PqsE is unlikely to be a direct molecular target of quercetin. This functional selectivity—strong binding to PqsR, moderate binding to LasR, and negligible affinity for PqsE—provides a mechanistic explanation for the experimental phenotype: dominant PQS inhibition, partial Las inhibition, and no effects attributable to PqsE disruption.
These docking results strengthen the mechanistic conclusion that quercetin’s QSI activity is mediated predominantly through interference with the PQS transcriptional regulator PqsR, with additional contributions from LasR antagonism. Furthermore, the decline in pqsA-gfp activity relative to lasB-gfp also suggests that quercetin may interact more directly with PQS-regulatory components or interfere with PQS autoinducer synthesis or perception.14,29,30 This aligns with prior evidence that flavonoids can destabilize PQS-regulated signalling cascades by binding to transcriptional regulators such as PqsR.31 Given the centrality of PQS in orchestrating biofilm robustness and persistent infections, selective targeting of this system represents a therapeutically advantageous mechanism.
Beyond transcriptional QS interference, quercetin and the quercetin-extract significantly impaired biofilm formation and partially disrupted mature biofilms. Preventive effects were more pronounced than disruption of established biofilms, consistent with the role of QS in early biofilm architecture stabilization. Nevertheless, the capacity to reduce biomass in pre-formed biofilms is clinically relevant, especially in chronic infections where biofilm persistence contributes to antibiotic recalcitrance.32 Quercetin also significantly reduced extracellular protease production, a key Las-regulated virulence trait. This functional suppression not only corroborates the lasB-gfp reporter results but also demonstrates that transcriptional inhibition translates into phenotypic attenuation. The dose-dependent reduction in protease activity underscores the broad-spectrum anti-virulence potential of quercetin. Importantly, the quercetin-extract consistently matched the efficacy of pure quercetin, with no significant differences in IC50 values for lasB inhibition, pqsA inhibition, biofilm inhibition, biofilm disruption, or protease reduction. This parity suggests that extraction does not compromise compound potency and may even benefit from subtle synergistic effects with co-extracted phenolics (gallic acid, ferulic acid). The equivalence of mushroom-derived quercetin to the purified standard highlights P. ostreatus as a scalable, edible, and economical source of QS inhibitors. Given the extensive cultivation and proven safety of P. ostreatus, mushroom-derived quercetin is an attractive candidate for nutraceutical, dietary, or adjunctive therapeutic applications. Such approaches may have particular relevance in cystic fibrosis airway infections, burn wound colonization, and ventilator-associated pneumonia, where biofilm-mediated persistence undermines antibiotic efficacy.33,34 Unlike conventional antibiotics, quercetin-based QSIs attenuate virulence without inhibiting growth, impose low selective pressure for resistance, may synergize with existing antimicrobials by destabilizing biofilms, and are derived from natural, renewable sources.8,35,36 This study demonstrates that quercetin derived from P. ostreatus is a potent, selective QSI in P. aeruginosa, with mechanistic evidence pointing toward strong PqsR antagonism, moderate LasR interference, and negligible PqsE targeting. By integrating biochemical, phenotypic, and in silico interaction data, these findings substantiate the role of mushroom-derived flavonoids as promising anti-virulence agents.
Limitations and Future Directions
While this study provides robust in vitro evidence of quercetin-mediated QS inhibition, several limitations must be acknowledged. First, the experiments were performed under controlled laboratory conditions using GFP-based reporter strains, which may not fully capture the complexity of in vivo infection environments. Host factors, nutrient conditions, and polymicrobial interactions can influence QS dynamics and compound activity. Second, the molecular mechanism underlying preferential PQS inhibition remains to be elucidated. Future work should employ molecular docking, transcriptomics, and mutational analyses to pinpoint quercetin’s targets within the QS regulatory hierarchy. Third, although P. ostreatus extracts were standardized to quercetin-equivalents, batch variability and the contribution of co-metabolites warrant further investigation.
Conclusion
Quercetin, whether purified or extracted from P. ostreatus, effectively inhibits quorum sensing and biofilm-associated virulence in P. aeruginosa PAO1 at non-inhibitory concentrations. The stronger suppression of the PQS system is supported mechanistically by molecular docking, which shows high-affinity binding of quercetin to PqsR and favourable interaction with LasR, but weak, non-specific binding to PqsE. The quercetin-extract matched the potency of pure quercetin across all assays, confirming P. ostreatus as a viable natural source of QSI-active flavonoids. These findings highlight mushroom-derived quercetin as a sustainable anti-virulence candidate with potential to complement existing treatments for chronic, biofilm-mediated infections.
Compliance with Ethical Standards
Research Involving Human Participants and/or Animals: This study did not involve human participants or animals.
Acknowledgments
The authors acknowledge the Patho-Biocatalysis Group (PBG).
Funding
The Department of Science and Innovation and the Technology Innovation Agency.
Disclosure
The study funder had no role in the study design, data collection, data analysis, data interpretation, or report writing. The authors declare no conflicts of interest related to this study.
References
1. Abdelaziz MA, El-Aziz AMA, El-Sokkary MMA, Barwa R. Characterization and genetic analysis of extensively drug-resistant hospital acquired Pseudomonas aeruginosa isolates. BMC Microbiol. 2024;24(1):225. doi:10.1186/s12866-024-03321-5
2. Yakobi SH, Ramaloko W, Maningi NE. Molecular characterization and inhibition of cyclic-di-GMP receptor proteins in Pseudomonas aeruginosa: INSIGHTS from docking and dynamics studies. Biochem Biophys Rep. 2025;43:102124. doi:10.1016/j.bbrep.2025.102124
3. Al Saqr A, Aldawsari M, Khafagy E-S, Shaldam M, Hegazy W, Abbas H. A novel use of allopurinol as a quorum-sensing inhibitor in Pseudomonas aeruginosa. Antibiotics. 2021;10(11):1385. doi:10.3390/antibiotics10111385
4. Abbas HA, Shaldam MA. Glyceryl trinitrate is a novel inhibitor of quorum sensing in Pseudomonas aeruginosa. African Health Sci. 2017;16(4):1109. doi:10.4314/ahs.v16i4.29
5. Boakye A, Seidu MP, Adomako A, Laryea MK, Borquaye LS. Marine-derived furanones targeting quorum-sensing receptors in Pseudomonas aeruginosa: molecular insights and potential mechanisms of inhibition. Bioinformat Biol Insights. 2024;18:11779322241275843. doi:10.1177/11779322241275843
6. Yakobi SH, Gasa N, Pooe OJ. A molecular and phenotypic analysis of the prevalence and patterns of antimicrobial resistance in Neisseria gonorrhoeae isolates from Kwazulu Natal, South Africa. Sci Afr. 2024;25e02334. doi:10.1016/j.sciaf.2024.e02334
7. Di Giacomo C, Malfa GA, Tomasello B, Bianchi S, Acquaviva R. Natural compounds and glutathione: beyond mere antioxidants. Antioxidants. 2023;12(7):1445. doi:10.3390/antiox12071445
8. Rasamiravaka T, Labtani Q, Duez P, El Jaziri M. The formation of biofilms by Pseudomonas aeruginosa: a review of the natural and synthetic compounds interfering with control mechanisms. Biomed Res Int. 2015;2015:759348. doi:10.1155/2015/759348
9. Shastri T, Binsuwaidan R, Siddiqui AJ, et al. Quercetin exhibits broad-spectrum antibiofilm and antiquorum sensing activities against gram-negative bacteria: in vitro and in silico investigation targeting antimicrobial therapy. Can J Infect Dis Med Microbiol. 2025;2025(1):2333207. doi:10.1155/cjid/2333207
10. Yakobi S, Zuma L, Gasa N, Pooe O, Hesham AE-L. Antimicrobial potential of organic phenolic compounds from wild mushroom extracts: impact on proliferation and kinetic growth of multidrug-resistant neisseria gonorrhoeae strains. J Food Biochem. 2024;2024(1):2336255. doi:10.1155/2024/2336255
11. Skogman ME, Kanerva S, Manner S, Vuorela PM, Fallarero A. Flavones as quorum sensing inhibitors identified by a newly optimized screening platform using chromobacterium violaceum as reporter bacteria. Molecules. 2016;21(9):1211. doi:10.3390/molecules21091211
12. Alum EU, Gulumbe BH, Izah SC, et al. Natural product-based inhibitors of quorum sensing: a novel approach to combat antibiotic resistance. Biochem Biophys Rep. 2025;43:102111. doi:10.1016/j.bbrep.2025.102111
13. Ouyang J, Feng W, Lai X, et al. Quercetin inhibits Pseudomonas aeruginosa biofilm formation via the vfr-mediated lasIR system. Microb Pathogenesis. 2020;149:104291. doi:10.1016/j.micpath.2020.104291
14. Lu L, Li M, Yi G, et al. Screening strategies for quorum sensing inhibitors in combating bacterial infections. J Pharm Anal. 2022;12(1):1–18. doi:10.1016/j.jpha.2021.03.009
15. Zhang L, Tan L, Liu M, et al. Quantitative measurement of cell-surface displayed proteins based on split-GFP assembly. Microb Cell Fact. 2024;23(1):108. doi:10.1186/s12934-024-02386-1
16. Yakobi SH, Mkhize S, Pooe OJ, Mohamed SA. Screening of antimicrobial properties and bioactive compounds of Pleurotus ostreatus extracts against staphylococcus aureus, escherichia coli, and neisseria gonorrhoeae. Biochem Res Int. 2023;2023:1777039. doi:10.1155/2023/1777039
17. Urban JH, Vogel J. A green fluorescent protein (GFP)-based plasmid system to study post-transcriptional control of gene expression in vivo. Methods Mol Biol. 2009;540:301–319. doi:10.1007/978-1-59745-558-9_22
18. Lichtenberg M, Kvich L, Larsen SLB, Jakobsen TH, Bjarnsholt T, Cardona ST. Inoculum concentration influences Pseudomonas aeruginosa phenotype and biofilm architecture. Microbiol Spectr. 2022;10(6):e03131–22. doi:10.1128/spectrum.03131-22
19. Gangwar M, Gautam MK, Sharma AK, Tripathi YB, Goel RK, Nath G. Antioxidant capacity and radical scavenging effect of polyphenol rich mallotus philippenensis fruit extract on human erythrocytes: an in vitro study. Sci World J. 2014;2014(1):279451. doi:10.1155/2014/279451
20. Yakobi SH, Pooe OJ. Identification of emerging multidrug-resistant neisseria gonorrhoeae isolates against five major antimicrobial agent options. Med Sci. 2023;11(2):28. doi:10.3390/medsci11020028
21. Elawady R, Aboulela AG, Gaballah A, Ghazal AA, Amer AN. Antimicrobial Sub-MIC induces Staphylococcus aureus biofilm formation without affecting the bacterial count. BMC Infect Dis. 2024;24(1):1065. doi:10.1186/s12879-024-09790-3
22. Rogers AT, Bullard KR, Dod AC, Wang Y. Bacterial growth curve measurements with a multimode microplate reader. Biol Protoc. 2022;12(9):e4410. doi:10.21769/BioProtoc.4410
23. Al Sulaiman D, Chang JYH, Ladame S. Subnanomolar detection of oligonucleotides through templated fluorogenic reaction in hydrogels: controlling diffusion to improve sensitivity. Angew Chem Int Ed. 2017;56(19):5247–5251. doi:10.1002/anie.201701356
24. Hao S, Yang D, Zhao L, et al. EGCG-mediated potential inhibition of biofilm development and quorum sensing in Pseudomonas aeruginosa. Int J Mol Sci. 2021;22(9):4946. doi:10.3390/ijms22094946
25. Patel R, Soni M, Soyantar B, et al. A clash of quorum sensing vs quorum sensing inhibitors: an overview and risk of resistance. Arch Microbiol. 2023;205(4):107. doi:10.1007/s00203-023-03442-x
26. Dehbanipour R, Ghalavand Z. Anti-virulence therapeutic strategies against bacterial infections: recent advances. Germs. 2022;12(2):262–275. doi:10.18683/germs.2022.1328
27. Dickey SW, Cheung GYC, Otto M. Different drugs for bad bugs: antivirulence strategies in the age of antibiotic resistance. Nat Rev Drug Discov. 2017;16(7):457–471. doi:10.1038/nrd.2017.23
28. Li J, Jia T, Yang L. Targeting anti-virulence factor strategies of bacterial pathogens. Biosaf Health. 2025;7(1):1–4. doi:10.1016/j.bsheal.2025.01.006
29. Sabir S, Das T, Kuppusamy R, et al. Novel quinazolinone disulfide analogues as pqs quorum sensing inhibitors against Pseudomonas aeruginosa. Bioorg Chem. 2023;130:106226. doi:10.1016/j.bioorg.2022.106226
30. Tan SY-Y, Chua S-L, Chen Y, et al. Identification of five structurally unrelated quorum-sensing inhibitors of Pseudomonas aeruginosa from a natural-derivative database. Antimicrob Agents Chemother. 2013;57(11):5629–5641. doi:10.1128/AAC.00955-13
31. Mansuri ML, Parihar P, Solanki I, Parihar MS. Flavonoids in modulation of cell survival signalling pathways. Genes Nutr. 2014;9(3):400. doi:10.1007/s12263-014-0400-z
32. Sahoo K, Meshram S. Biofilm formation in chronic infections: a comprehensive review of pathogenesis, clinical implications, and novel therapeutic approaches. Cureus. 2024;16(10):e70629. doi:10.7759/cureus.70629
33. Elfadadny A, Ragab RF, AlHarbi M, et al. Antimicrobial resistance of Pseudomonas aeruginosa: navigating clinical impacts, current resistance trends, and innovations in breaking therapies. Front Microbiol. 2024;15:1374466. doi:10.3389/fmicb.2024.1374466
34. Rossi E, La Rosa R, Bartell JA, et al. Pseudomonas aeruginosa adaptation and evolution in patients with cystic fibrosis. Nat Rev Microbiol. 2021;19(5):331–342. doi:10.1038/s41579-020-00477-5
35. Yakobi S, Zuma L, Pooe O. Molecular docking and structure-activity relationship analysis of target compounds against glyceraldehyde-3-phosphate dehydrogenase in azithromycin-resistant neisseria gonorrhoeae. ChemistrySelect. 2024;9(9):e202303341. doi:10.1002/slct.202303341
36. Yakobi S, Zuma L, Pooe O, Dar AA. Investigation into the interaction between penicillin-resistant and penicillin-susceptible gonococcal penicillin-binding protein 2 and target phenolic ligands through molecular docking studies and structure-activity relationship analysis. Adv Pharmacolog Pharmaceu Sci. 2024;2024(1):2585922. doi:10.1155/2024/2585922
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.