Back to Journals » Diabetes, Metabolic Syndrome and Obesity » Volume 19
Modulating the Gut-Nerve Axis and Inflammation: A Narrative Review of Dietary Patterns in Diabetic Peripheral Neuropathy
Authors Sun R, Li X, Liu X, Guo D, Chen Y, Zhao Y, Yin Y, Zhai X
Received 14 February 2026
Accepted for publication 24 April 2026
Published 21 May 2026 Volume 2026:19 602342
DOI https://doi.org/10.2147/DMSO.S602342
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Rebecca Baqiyyah Conway
Ruoxiang Sun,1,2,* Xu Li,1,* Xianping Liu,3,* Dongjie Guo,1,2 Yuanqin Chen,1,4 Yu Zhao,1,2 Yanxiang Yin,1,4 Xingyue Zhai1
1Department of Clinical Nutrition, The Second Hospital of Dalian Medical University, Dalian, Liaoning, People’s Republic of China; 2School of Public Health, Dalian Medical University, Dalian, Liaoning, People’s Republic of China; 3Department of Public Health, Dalian Health Development Center, Dalian, Liaoning, People’s Republic of China; 4College of Health and Wellness, Dalian Medical University, Dalian, Liaoning, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Xingyue Zhai, Email [email protected]
Abstract: Diabetic peripheral neuropathy (DPN) is a prevalent and debilitating complication of diabetes, arising from a complex interplay of metabolic dysregulation, oxidative stress, and chronic inflammatory pathways. This narrative review synthesizes the core pathophysiology of DPN and critically evaluates the potential role of various dietary patterns in its management. Healthy patterns, such as the Mediterranean, DASH, and plant-based diets, may offer neuroprotective potential by improving systemic metabolic and inflammatory status. Conversely, pro-inflammatory diets remain associated with exacerbated nerve damage. Furthermore, emerging strategies like low-carbohydrate diets and intermittent fasting show promise in preclinical models via mechanisms such as metabolic reprogramming and autophagy, though direct clinical evidence in DPN populations remains emerging. A critical gap persists regarding the lack of large-scale, direct intervention studies in patients with established DPN. Future research must prioritize this population to build a robust evidence base for dietary strategies in secondary prevention and personalized therapy.
Keywords: diabetic peripheral neuropathy, dietary patterns, biomarkers, inflammation, oxidative stress
Introduction
Diabetic peripheral neuropathy (DPN) represents a major global health challenge, affecting approximately 50% of individuals with diabetes during their lifetime.1,2 As the primary driver of chronic pain and non-traumatic lower-limb amputations, DPN accounts for a staggering economic burden, with annual global healthcare expenditures for diabetes-related complications approaching $1 trillion.1,3 Clinically, it is essential to distinguish between primary prevention and the management of established DPN.2 Primary prevention, centered on intensive glycemic control, has demonstrated robust efficacy in type 1 diabetes (T1DM) but offers only modest protection in type 2 diabetes (T2DM).1,2,4 Conversely, once DPN is clinically established, management pivots toward secondary and tertiary strategies—specifically alleviating neuropathic symptoms and slowing neurovascular degradation to prevent debilitating ulcers—as structural nerve damage is often irreversible.2,5 Given the limitations of traditional monotherapy, multi-targeted dietary patterns have emerged as a pivotal strategy for both preventing onset and improving clinical outcomes in established cases.5,6
Dietary patterns are a key modifiable factor and offer a powerful, multi-target strategy for DPN management. This review summarizes the core pathophysiology of DPN and evaluates the role of various dietary patterns in its development and man-agement. We assess established healthy patterns like the Mediterranean, plant-based, and DASH diets, as well as the detrimental pro-inflammatory diet. We also explore novel strategies such as low-carbohydrate diets and intermittent fasting.
Search Strategy and Selection Criteria
A comprehensive literature search was performed across PubMed, Web of Science, and Google Scholar databases for articles published from inception to February 2024. The search used combinations of keywords including “diabetic peripheral neuropathy”, “dietary patterns,” “Mediterranean diet,” “DASH diet,” “plant-based diet,” “ketogenic diet,” “intermittent fasting,” “inflammation,” “oxidative stress,” and “gut microbiota.” Selection criteria prioritized randomized controlled trials (RCTs), systematic reviews, and meta-analyses focused on DPN or related metabolic markers. Additionally, high-quality mechanistic studies using animal models were included to elucidate potential biochemical pathways connecting nutrition to nerve function. Only peer-reviewed articles published in English were considered.
The Pathogenesis of Diabetic Peripheral Neuropathy
Chronic hyperglycemia, dyslipidemia, and insulin resistance drive DPN pathogenesis by directly inducing neuronal injury and microvascular impairment.4–6 These primary drivers trigger an interconnected cascade of metabolic disruptions. Specifically, excess glucose overactivates the polyol pathway, causing osmotic stress and depleting cellular antioxidants like glutathione.7 Simultaneously, the accumulation of advanced glycation end products (AGEs) and the overactivation of the protein kinase C (PKC) pathway collaboratively generate reactive oxygen species (ROS) and activate NF-κB.8–10 This cascade upregulates pro-inflammatory cytokines and vascular factors, driving Schwann cell demyelination, vascular permeability, and ischemia.8,11 Subsequently, ROS-induced DNA breaks overactivate the PARP pathway, severely depleting NAD+ and ATP, which culminates in neuronal energy failure and apoptosis.12,13 Ultimately, these mutually reinforcing pathways converge on mitochondrial dysfunction, sustained oxidative stress, and chronic neuroinflammation.14,15 Because these core pathogenic mechanisms are intrinsically linked to nutritional status, multi-targeted dietary patterns offer a compelling and logical strategy for DPN management (Figure 1).
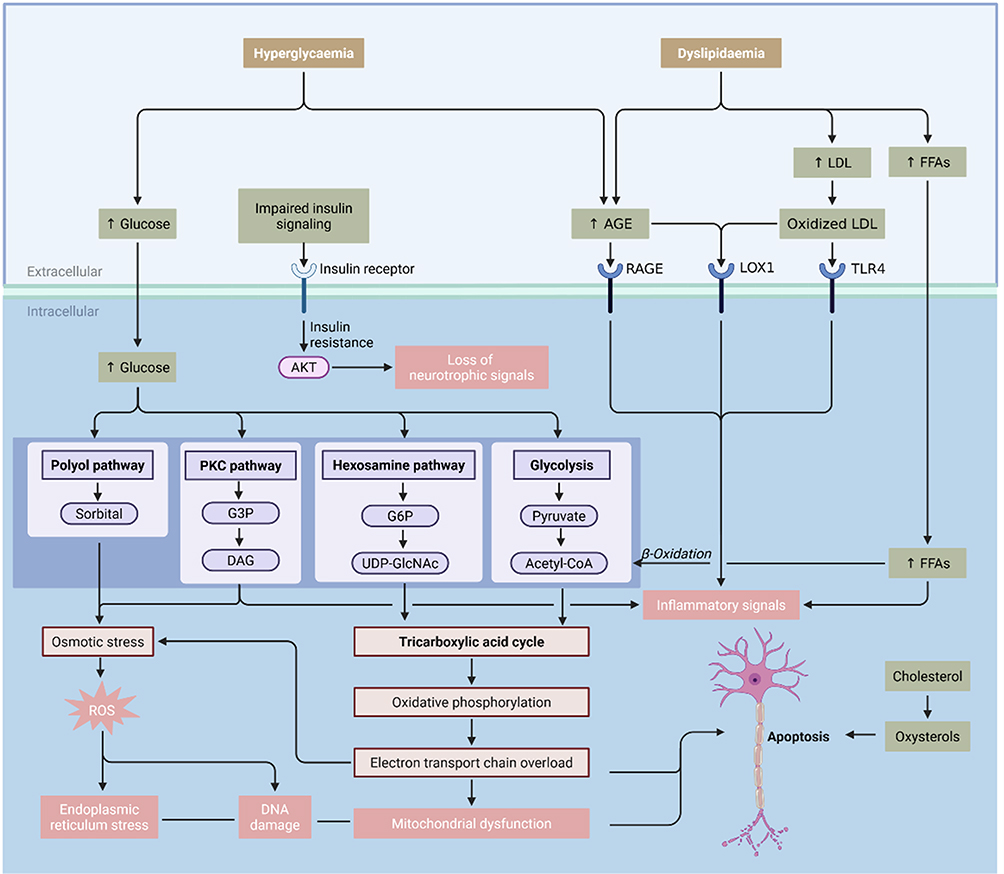 |
Figure 1 Interplay of Hyperglycaemia and Dyslipidaemia in the Pathogenesis of Diabetic Pe-ripheral Neuropathy. Abbreviations: LDL, low-density lipoprotein; FFAs, free fatty acids; AGE, advanced glycation end products; RAGE, receptor for advanced glycation end products; LOX1, lectin-like oxidized low-density lipoprotein receptor-1; TLR4, Toll-like receptor 4; G3P, glycer-aldehyde-3-phosphate; DAG, diacylglycerol; G6P, glucose-6-phosphate; UDP-GlcNAc, uridine diphosphate N-acetylglucosamine; Acetyl-CoA, acetyl coenzyme A; ROS, reactive oxygen spe-cies; AKT, protein kinase B; PKC, protein kinase C. |
Biomarkers Associated with DPN and Their Clinical Significance
The pathogenesis of diabetic peripheral neuropathy (DPN) is complex, with its onset and development resulting from the combined effects of multiple pathophysio-logical changes. Therefore, identifying sensitive and specific biomarkers is not only crucial for the early diagnosis, disease assessment, and prognosis of DPN, but it also provides a window for understanding its intrinsic pathological network. More im-portantly, these biomarkers—which reflect glycemic homeostasis, lipotoxicity, in-flammatory status, and microvascular damage—can largely be modulated by dietary and nutritional interventions. This section aims to systematically review the key bi-omarkers associated with DPN and to reveal their potential to serve as both a bridge connecting dietary interventions with neuropathic improvement and as targets for monitoring. The major biomarkers discussed in this section are summarized in Table 1.
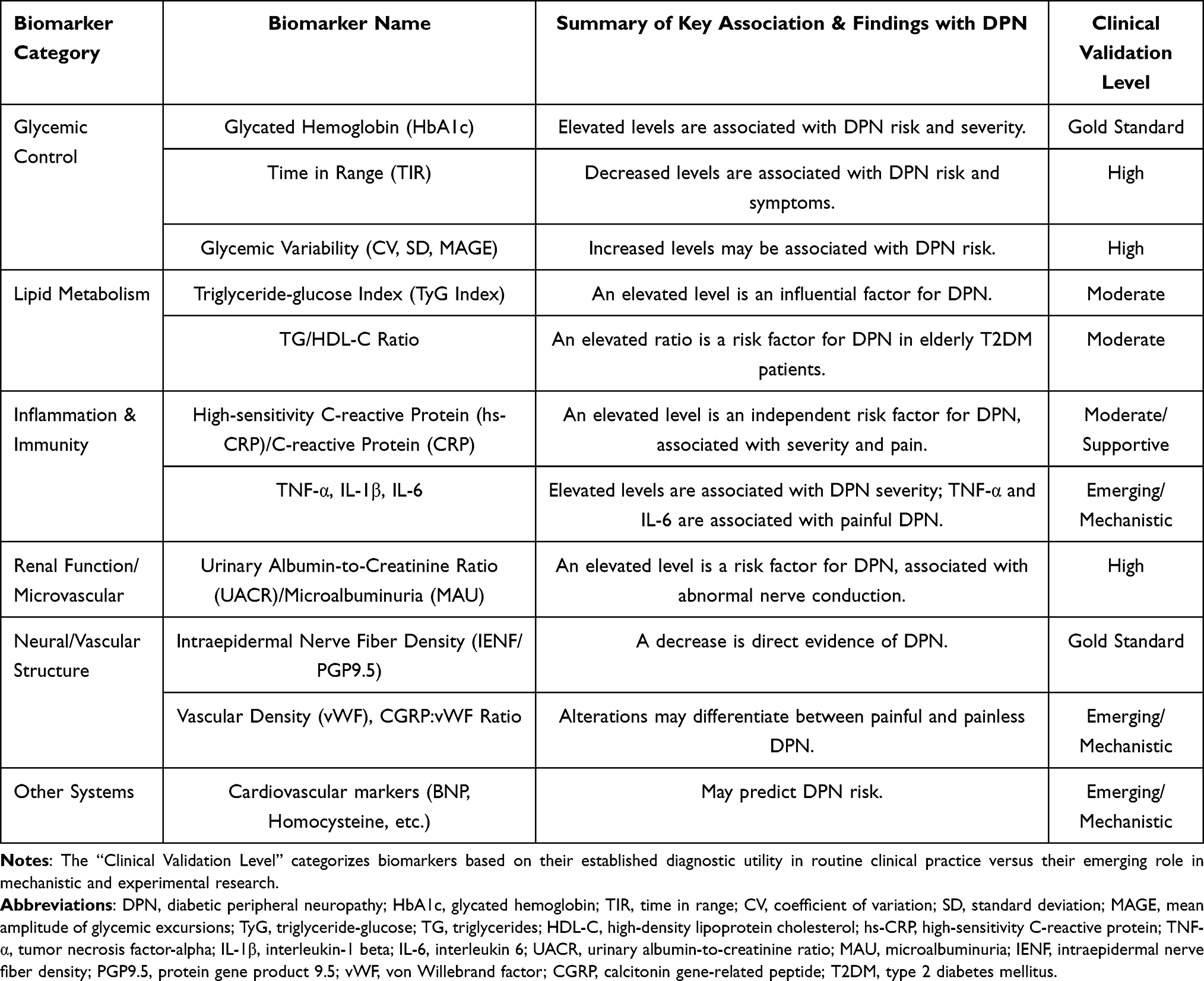 |
Table 1 Overview of Major Biomarkers Associated with DPN and Their Clinical Significance |
Biomarkers Related to Glycemic Control and Metabolic Dysregulation
Chronic hyperglycemia and its associated metabolic dysregulation are the initi-ating and central factors in the onset and development of DPN. As the “gold standard” for reflecting average blood glucose levels over the past 2–3 months, glycated hemo-globin (HbA1c) is closely correlated with the incidence and severity of DPN. Multiple studies have confirmed that an elevated HbA1c level is an independent risk factor for DPN.16–19 A systematic review by Casadei also emphasized the potential of HbA1c as a biomarker for peripheral neuropathy in the diabetic foot.20 Strict glycemic con-trol that maintains HbA1c within the standard range is fundamental to preventing and delaying the progression of DPN. However, research suggests that for established neuropathy, its progression appears difficult to completely reverse despite subsequent improvements in glycemic control,21 highlighting the importance of early interven-tion.
Time in Range (TIR) is a new indicator for assessing glycemic variability and quality of control that has emerged with the popularization of continuous glucose monitoring technology. A study by Wang Danyu demonstrated that TIR levels in pa-tients with T2DM are negatively correlated with the prevalence of DPN, where lower TIR levels are associated with a higher prevalence of DPN.22
The Triglyceride-glucose (TyG) index is a simple and reliable indicator for as-sessing insulin resistance (IR).
The aforementioned indicators, which reflect long-term glycemic levels, glycemic variability, and insulin resistance, constitute the core monitoring targets for evaluating the efficacy of dietary interventions, such as adjusting the quality and quantity of car-bohydrates and increasing dietary fiber.
Biomarkers Related to Glycemic Control and Metabolic Dysregulation
Dyslipidemia is a recognized risk factor for DPN, and its neurotoxic effects (ie., lipotoxicity) can be evaluated using various biomarkers. Conventional lipid markers have demonstrated an association with DPN. Multiple studies have confirmed that hypertriglyceridemia (high TG) and low high-density lipoprotein cholesterol (HDL-C) are risk factors for DPN.17–19
To more comprehensively reflect the integrated effects of lipotoxicity, researchers have further explored the value of composite indices. The TG/HDL-C ratio is the ratio of triglycerides to high-density lipoprotein.
In addition, some molecules directly related to the pathogenic mechanisms of li-pids have also come under investigation. Oxidized low-density lipoprotein (oxLDL) is theoretically considered to be an important pro-atherosclerotic and neurotoxic factor. However, a case-control study by Rosales-Hernandez et al found no significant dif-ference in plasma oxLDL levels between DPN patients and control subjects, suggesting that its pathogenic role in human DPN requires further investigation.23
Inflammatory and Immune-Related Biomarkers
Chronic low-grade inflammation is a key pathological link connecting metabolic disorders with nerve damage; therefore, biomarkers that assess systemic and local in-flammatory status hold a significant position in DPN research.
As a systemic inflammatory marker, elevated levels of high-sensitivity C-reactive protein (hs-CRP/CRP) are associated with the incidence and severity of DPN.
As an important pro-inflammatory cytokine, Tumor Necrosis Factor-alpha (TNF-α) levels are elevated in DPN patients and are associated with reduced nerve conduction velocity and pain.24,25 Pro-inflammatory cytokines from the interleukin (IL) family, such as IL-1β and IL-6, are elevated in DPN patients and correlate with the severity of DPN and pain.26,27 Conversely, anti-inflammatory cytokines like IL-10 may be de-creased in DPN patients.25
Biomarkers Related to Renal Function and Microvascular Damage
DPN and diabetic nephropathy (DN) are both microvascular complications that often coexist and share similar pathological foundations. Therefore, indicators reflect-ing renal microvascular damage, such as the urinary albumin-to-creatinine ratio (UACR), also provide indirect clues for DPN risk assessment.
The urinary albumin-to-creatinine ratio (UACR) is a sensitive indicator for early DN and has also been found to be associated with DPN.
Other Relevant Hematological Indicators
Serum uric acid (SUA) is receiving increasing attention due to the relationship between hyperuricemia and DPN. Hyperuricemia is not only related to purine metab-olism but is also closely linked to modern dietary factors such as high-fructose diets, which adds a new dimension to its potential as a target for nutritional intervention.
DPN and Dietary Patterns
Diabetic peripheral neuropathy (DPN) pathogenesis is complex, involving multi-ple links such as hyperglycemia, dyslipidemia, insulin resistance, oxidative stress, chronic inflammation, and microvascular dysfunction. Lifestyle factors, particularly dietary habits, play a crucial role in the management of diabetes and its complications. Compared to studies on single nutrients, analysis of dietary patterns can more com-prehensively reflect an individual’s overall dietary structure and its complex relation-ship with health. Therefore, exploring the association between different dietary pat-terns and DPN, along with their potential mechanisms, is of great importance for the prevention and management of DPN (summarized in Table 2).
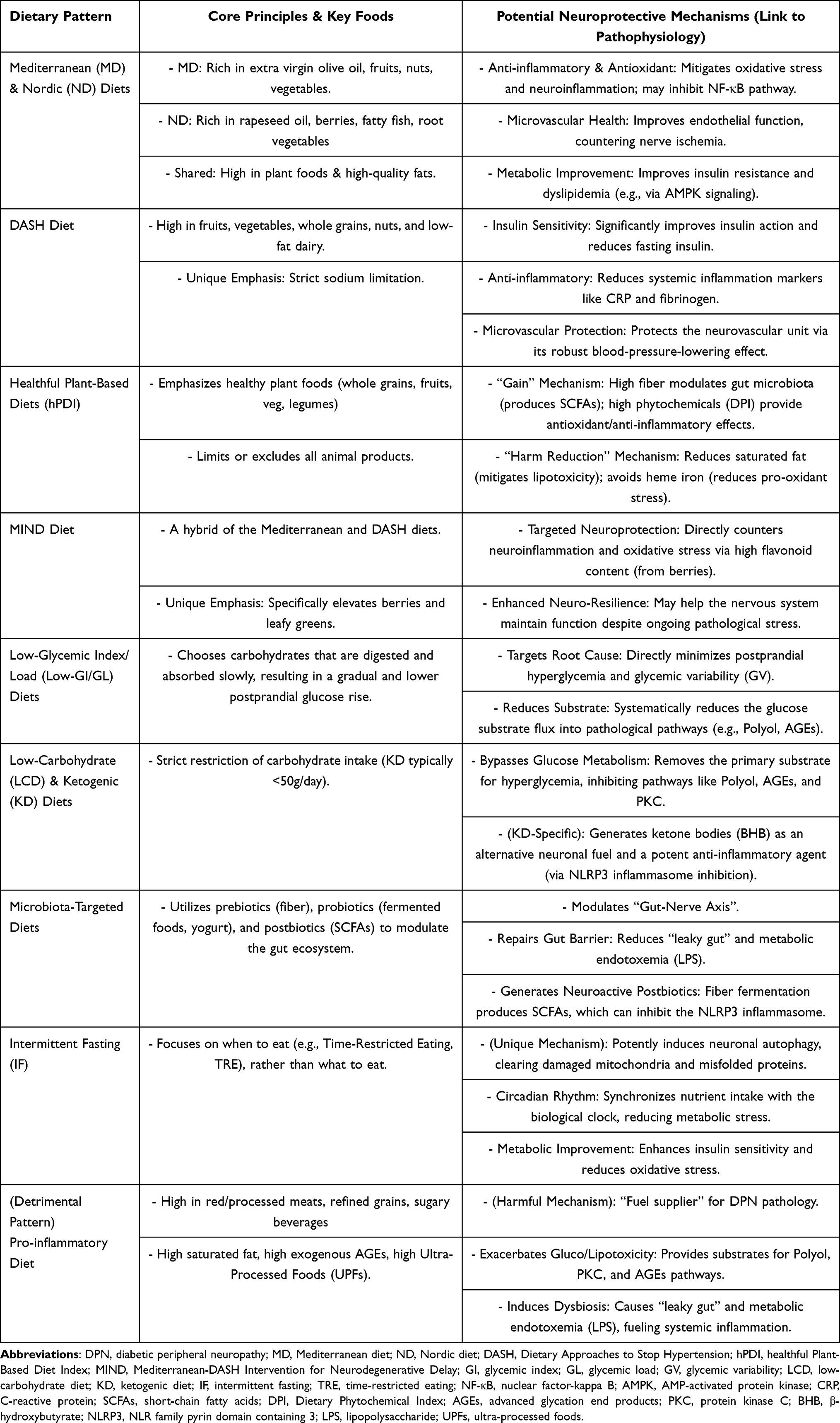 |
Table 2 Summary of Dietary Patterns and Their Potential Neuroprotective Mechanisms in DPN |
The Mediterranean Dietary and Nordic Dietary Pattern
Healthy dietary patterns adapted to regional foods and cultures, such as the Mediterranean and Nordic diets, offer important insights for DPN management. Alt-hough their food components differ—with the former emphasizing olive oil, fruits, and nuts, and the latter featuring rapeseed oil, berries, and fatty fish28–30—their under-lying nutritional principles are highly consistent. Both patterns appear to exert neuro-protective potential by targeting similar core pathological mechanisms.
To begin with, a key advantage of these dietary patterns is their potent an-ti-inflammatory and antioxidant capacity, which directly counteracts critical drivers of DPN. The Mediterranean diet (MD) is rich in polyphenols and antioxidants from extra virgin olive oil, fruits, and vegetables, which can mitigate oxidative stress and neu-roinflammation.28 Similarly, the Nordic diet (ND) provides strong antioxidant support through its high content of berries and root vegetables. Clinical evidence supports this: a randomized controlled trial showed that the ND significantly im-proved markers of inflammation in individuals with impaired glucose metabolism.30 Another study found that adherence to core ND components was associated with lower concentrations of high-sensitivity C-reactive protein (hs-CRP),31 a key bi-omarker for DPN severity. By mitigating systemic inflammation, both diets may inhibit the activation of pro-inflammatory cascades, such as the NF-κB pathway, thereby protecting neurons from damage.
Furthermore, both patterns can effectively protect microvascular health, which is crucial for alleviating the nerve damage caused by ischemia and hypoxia. Endothelial dysfunction is a central event in the course of DPN. A randomized controlled trial by de Mello et al explicitly reported that the ND improved markers of endothelial func-tion.30 This finding is supported by large cohort data; the EPIC-Potsdam study, for instance, found that ND adherence was associated with a possible reduced risk of car-diovascular events like myocardial infarction and stroke.32 While direct evidence for the MD on DPN is still evolving,33–35 its robust, well-documented benefits for re-ducing systemic cardiovascular risk are undisputed, which indirectly supports its po-tential benefits for the neurovascular unit.
Additionally, these diets address the upstream metabolic disturbances that initiate neuropathy, and their effects are now traceable through specific biomarkers. Insulin resistance and dyslipidemia are primary drivers of DPN. The MD, with its high fiber and MUFA content, is effective in improving these metabolic markers.27,28 Research on the ND provides even more detailed evidence at the molecular level. An interven-tion study by Fritzen et al found that the ND improves overall metabolism by acti-vating AMPK signaling in adipose tissue,26 a master regulator that enhances insulin sensitivity. Furthermore, metabolomics studies have identified biomarkers like pipe-colic acid betaine, which is linked to whole grain intake. The level of this compound is inversely associated with fasting insulin and unfavorable lipid profiles,36 providing strong evidence for how the ND precisely regulates metabolic pathways.
Finally, these fiber-rich dietary patterns are closely linked to the gut microbiota, a frontier that opens new possibilities for personalized nutrition. The gut microbiome is a key hub connecting diet and host health. Interestingly, a 6-month trial by Roager et al found that while the ND did not cause major shifts in the overall microbiota, the par-ticipants’ baseline microbial signature (specifically the Prevotella-to-Bacteroides ratio) predicted their metabolic response to the diet, such as changes in plasma cholesterol.37 This suggests that the benefits of healthy diets may not be uniform and could be influenced by an individual’s enterotype. This finding points to a new direction for DPN management, where analyzing a patient’s gut microbiota could help tailor the most effective dietary interventions for them.
In conclusion, although direct clinical evidence for using MD or ND to treat DPN is still accumulating and can be inconsistent,35,38 the mechanistic evidence is strong and convergent. The diets’ abilities to act simultaneously on inflammation, vascular health, metabolism, and the microbiota demonstrate their significant neuroprotective potential. The fact that these distinct regional diets achieve similar benefits underscores a crucial point: for managing diabetic complications, adhering to the principles of a diet rich in plant foods, high-quality fats, and antioxidants is likely more important than following a specific food list.
The DASH Dietary Pattern
The Dietary Approaches to Stop Hypertension (DASH) diet is another well-researched, plant-forward dietary pattern.39 Initially developed to lower blood pressure, the DASH diet emphasizes a high intake of fruits, vegetables, whole grains, and nuts. Unlike the Mediterranean and Nordic diets, it specifically promotes low-fat dairy products and places a stronger emphasis on strict sodium limitation.39 Its nu-trient-rich composition provides a strong foundation for improving overall metabolic health.
The relevance of the DASH diet to DPN lies in its proven ability to improve insulin sensitivity and glycemic control. A key randomized controlled trial by Ard et al showed that adding the DASH diet to a lifestyle intervention significantly enhanced insulin action, an effect not seen with lifestyle changes alone.40 A meta-analysis further supports this, finding that the DASH diet can significantly reduce fasting insu-lin levels.41 More recent research has begun to identify the specific glucose homeo-stasis pathways improved by DASH, suggesting the benefits likely arise from its plant-based components.42 Furthermore, long-term adherence to the DASH diet is associated with a lower risk of developing a metabolically unhealthy phenotype, which is a major upstream risk factor for the development of DPN.43 For patients with type 2 diabetes, following the DASH diet has been shown to yield comprehensive benefits. These include significant reductions in glycated hemoglobin (A1C), fasting blood glu-cose, LDL cholesterol, and blood pressure.44 These metabolic improvements directly counteract the hyperglycemic and dyslipidemic states that fuel the core pathological pathways of DPN.
Furthermore, the DASH diet directly targets the chronic inflammation and vas-cular dysfunction central to DPN’s pathogenesis. A clinical trial by Azadbakht et al demonstrated that the DASH diet significantly reduced levels of C-reactive protein (CRP) and fibrinogen in diabetic individuals.45 This anti-inflammatory effect is consistent with other findings linking the DASH diet to lower hs-CRP levels.46 No-tably, the key biomarkers improved by the DASH diet—such as A1C and triglycer-ides—are the same factors identified in recent studies as strong predictors for the de-velopment and severity of neuropathy.47 The diet’s strong, well-established effect on lowering blood pressure also helps protect the microvasculature, potentially improving nerve blood supply and reducing ischemic damage.44,48
In conclusion, the DASH diet offers a multi-faceted intervention that addresses the complex pathology of DPN. It simultaneously improves metabolic control, enhances insulin sensitivity, and reduces inflammation.40,41,44–46 Its well-documented effects on blood pressure also suggest a distinct benefit for protecting the microvasculature that supports nerve health.44,48 By acting on several key pathological pathways at once, the DASH diet represents a robust strategy for the prevention and management of diabetic peripheral neuropathy.
Plant-Based Dietary Patterns
Plant-based dietary patterns are defined by an emphasis on foods derived from plants, such as fruits, vegetables, legumes, whole grains, nuts, and seeds, while dis-couraging or completely excluding most or all animal products.49 Recent global health initiatives, such as the EAT-Lancet Commission report, also advocate for a predominantly plant-based “planetary health diet” to simultaneously promote human health and environmental sustainability.50 For more precise research, indices like the healthful Plant-Based Diet Index (hPDI) have been developed. The hPDI distinguishes between healthy plant foods (eg., whole grains, fruits, vegetables) and less healthy ones (eg., refined grains, sugary drinks).51 Broadly, greater adherence to healthy plant-based diets is associated with a lower risk of major chronic diseases, including type 2 diabetes.52
Strong evidence supports the benefits of plant-based diets for improving key metabolic markers relevant to DPN. Recent meta-analyses of randomized controlled trials (RCTs) show that vegetarian and vegan diets significantly reduce both glycated hemoglobin (A1C) and body mass index (BMI) in patients with type 2 diabetes.53,54 Data from large prospective cohorts, such as the UK Biobank, further confirm that adherence to plant-based diets is associated with a lower risk of incident type 2 dia-betes, thereby reducing the foundational risk for DPN.55 Most critically, a direct link to neuropathy has been established. A case-control study by Asadi et al found that a higher Dietary Phytochemical Index (DPI), reflecting a diet rich in plant-derived bio-active compounds, was associated with a 75% reduced odds of having diabetic sen-sorimotor polyneuropathy (DSPN) after adjusting for confounders.56
The neuroprotective potential of plant-based diets stems from a dual “gain and harm reduction” mechanism.49 The “gain” mechanism arises from a high intake of beneficial components. Abundant dietary fiber promotes a healthy gut microbiota, which in turn produces beneficial metabolites like short-chain fatty acids (SCFAs) that modulate systemic inflammation and improve metabolic health.57 The high intake of phytochemicals provides direct antioxidant and anti-inflammatory effects. This is supported by the DPI study56 and other research linking the hPDI to lower levels of hs-CRP.46
Simultaneously, a “harm reduction” mechanism works by limiting deleterious components. The exclusion or reduction of animal products lowers the intake of satu-rated fats, which mitigates the lipotoxicity that drives mitochondrial dysfunction in neurons. The avoidance of red and processed meat is particularly important, as their consumption is linked to increased inflammation and a higher risk of T2D.58 It also significantly reduces the intake of heme iron. Excess heme iron is a potent pro-oxidant that can generate reactive oxygen species, directly contributing to the oxidative stress central to DPN’s pathogenesis.59 Furthermore, plant-based diets often have a lower energy density, which aids in weight management. Evidence from a two-year RCT by Turner-McGrievy et al confirmed that a vegan diet was more effective for weight loss than a moderate low-fat diet.60 This indirect benefit of weight control can compre-hensively improve insulin resistance and the overall metabolic environment.
Looking forward, the distinction between different qualities of plant-based diets is crucial. The foundational work by Satija et al demonstrated that while an hPDI was associated with a substantially lower risk of T2D, an unhealthful plant-based diet (uPDI) rich in refined grains and sugary drinks was associated with a higher risk.51 This highlights that simply avoiding animal products is not enough. Future research on DPN should therefore use these indices to differentiate diet quality and provide more precise and effective dietary recommendations.
The Mediterranean, Nordic, DASH, and healthy Plant-Based diets, despite their different geographical origins and specific food lists, converge on a core set of princi-ples that collectively target the complex pathophysiology of DPN. Their shared foun-dation is a high intake of whole, minimally processed plant foods—including fruits, vegetables, whole grains, and legumes—and a limitation of red and processed meats, refined grains, and sugar-sweetened beverages.
This common core results in a multi-pronged mechanistic attack on DPN. First, by emphasizing high-fiber, unrefined carbohydrates, all four patterns are typically lower in glycemic index and load, directly counteracting the primary driver of DPN: hyper-glycemia and its associated glucotoxicity. Second, their richness in phytochemicals, vitamins, and minerals provides a powerful antioxidant and anti-inflammatory de-fense. A systematic review of RCTs confirmed that adherence to such healthy, plant-rich patterns is indeed associated with a reduction in biomarkers of inflamma-tion.61 Third, by favoring unsaturated fats (from olive oil, rapeseed oil, or nuts) over saturated fats, these diets improve lipid profiles and endothelial function, thereby mitigating the lipotoxicity and microvascular damage that contribute to nerve ische-mia.
However, their subtle differences offer unique, complementary benefits. The DASH diet’s explicit focus on sodium restriction and high potassium and magnesium intake provides a distinct advantage for blood pressure control and vascular health. The complete exclusion of heme iron in vegan diets offers the most potent strategy for reducing this specific source of oxidative stress. The Mediterranean and Nordic diets, with their emphasis on specific high-quality oils, robustly target lipotoxicity.
The MIND Dietary Pattern
The MIND (Mediterranean-DASH Intervention for Neurodegenerative Delay) diet is a dietary pattern designed specifically to promote brain health. It is a hybrid of the Mediterranean (MD) and DASH diets. The MIND diet adopts the core principles of these two patterns but has a unique focus. It specifically highlights 10 brain-healthy food groups, such as leafy greens and berries, and 5 food groups to limit, including red meat, sweets, and fried foods.62
Although the MIND diet was created to prevent Alzheimer’s disease, its benefits also apply to the key risk factors for DPN. A key prospective study was the first to show that adherence to the MIND diet is associated with a lower risk of developing type 2 diabetes.63 Another study found that following the MIND diet helps reduce the risk of developing a metabolically unhealthy phenotype, which is closely linked to insulin resistance and systemic inflammation.43 These findings show that the MIND diet can address the upstream drivers of DPN.
The diet’s neuroprotective mechanisms are linked to its emphasis on specific nu-trients and bioactive compounds. The MIND diet places special importance on the in-take of berries and leafy greens.62 Berries are an excellent source of flavonoids. A large prospective study by Devore et al confirmed that a high intake of flavonoids, particularly from berries, is associated with slower cognitive decline.64 Similarly, a study by Morris et al found that daily consumption of leafy green vegetables was linked to a significant slowing of cognitive decline.65 These foods are rich in anti-oxidants and anti-inflammatory compounds. They can directly counter oxidative stress and neuroinflammation, which are core pathological mechanisms in DPN. This is supported by research showing that a MIND-like diet can protect the brain from the damage caused by environmental pollutants.66
The MIND diet may also enhance neuro-resilience, or the nervous system’s ability to withstand damage. A unique study by Dhana et al found that adherence to the MIND diet was associated with better cognitive function, even in the presence of Alz-heimer’s disease pathology.67 This suggests the diet’s benefits go beyond just pre-venting damage and may help the nervous system maintain function despite ongoing stress. This mechanism is highly relevant to DPN, where peripheral nerves are under constant metabolic pressure.
When compared to the Mediterranean and DASH diets, all three patterns share a common foundation. They are all plant-forward and recommend limiting red meat, saturated fats, and added sugars. All three can positively influence the biomarkers and pathophysiological processes of DPN by improving insulin sensitivity, lipid profiles, and inflammation. The key innovation of the MIND diet, however, is its specificity. Unlike the broad recommendations for fruits and vegetables in the MD and DASH diets, the MIND diet precisely elevates berries and leafy greens to a central role because of their strong, specific evidence for neuroprotection.64,65 Therefore, the MIND diet can be seen as a refined version of the MD and DASH diets, optimized to maximize benefits for the nervous system. In summary, the MIND diet is a highly promising strategy for the prevention and management of diabetic peripheral neuropathy due to its proven effects on metabolic health and its targeted neuroprotective mechanisms.
Pro-Inflammatory Dietary Pattern
In contrast to healthy dietary patterns, a diet characterized by a high intake of red and processed meats, refined grains, and sugar-sweetened beverages is widely recog-nized as pro-inflammatory. Its nutritional profile—high in saturated fats, exogenous advanced glycation end products (AGEs), and low in fiber—contributes to this effect.68 To quantify this inflammatory potential, researchers have developed tools such as the Dietary Inflammatory Index (DII).69 A core feature of this pattern is the high consumption of ultra-processed foods (UPFs), which are industrial formulations con-taining numerous additives.70
Substantial evidence links this dietary pattern to negative neurological outcomes. In human populations, adherence to a pro-inflammatory diet is significantly associated with an increased risk of developing type 2 diabetes,71 while high consumption of UPFs also elevates T2D risk.72 Animal models provide more direct evidence of nerve damage. High-fat diets have been shown to induce neuronal inflammation73 and can produce peripheral nerve damage similar to that seen in diabetes, even in the ab-sence of overt hyperglycemia.74 This suggests that dyslipidemia itself is a critical factor in nerve injury. Furthermore, a “cafeteria diet,” which models this eating pattern, has been shown to cause nerve dysfunction even in a prediabetic state, highlighting the direct detrimental effect of the diet itself.75
Mechanistically, this pro-inflammatory dietary pattern acts as a “fuel supplier” for the pathological network of DPN, simultaneously igniting multiple damaging path-ways. First, the high load of sugar and saturated fat provides the substrate for core metabolic disturbances. This directly exacerbates hyperglycemia and lipotoxicity, thereby overloading the polyol, protein kinase C (PKC), and AGEs pathways.76 The role of saturated fatty acids is particularly detrimental. The animal study by Rumora et al clearly demonstrated that a diet rich in saturated fats induces neuropathy through mechanisms involving mitochondrial dysfunction and neuronal lipotoxicity.27 Ad-ditionally, foods like processed meats, which are often cooked at high temperatures, are a major source of exogenous AGEs. These compounds activate their receptor (RAGE), directly triggering oxidative stress and inflammatory responses.8
Second, this dietary pattern acts as a “disruptor” of the gut microbiota, exacer-bating nerve damage via the gut-nerve axis. The high-fat, low-fiber structure disrupts the balance of intestinal microorganisms, leading to dysbiosis.77 The study by Guo et al directly linked these diet-induced changes in the gut microbiota to peripheral neu-ropathy phenotypes and to the expression of genes in the nerve related to inflammation, lipid metabolism, and antioxidant defense.78 A deeper mechanism involves the impairment of intestinal barrier integrity, leading to a “leaky gut.” This allows bacterial endotoxins like lipopolysaccharide (LPS) to translocate into the bloodstream, a condi-tion known as metabolic endotoxemia. This process fuels a chronic, low-grade systemic inflammatory state that persistently activates the immune system and intensifies neu-roinflammation.79,80
In summary, the pro-inflammatory dietary pattern provides a continuous patho-logical drive for the development and progression of DPN. It achieves this by directly activating multiple metabolic damage pathways and indirectly by disrupting gut ho-meostasis. Therefore, a foundational strategy in the nutritional intervention for pa-tients with DPN is the strict avoidance of this dietary pattern.
Low-Glycemic Index/Load Diets
The glycemic index (GI) and glycemic load (GL) are key metrics for assessing the quality, rather than the quantity, of carbohydrates.81 The GI ranks carbohy-drate-containing foods based on their effect on postprandial blood glucose levels, while the GL accounts for both the GI and the amount of carbohydrate consumed. The core principle of a low-GI/GL diet is to choose foods that are digested and absorbed slowly, resulting in a gradual and lower rise in blood glucose and insulin.76 This principle is a shared, underlying component of many healthy dietary patterns, including the Mediterranean, DASH, and healthy plant-based diets.
A large body of high-level evidence confirms that low-GI/GL diets effectively improve key metabolic markers in individuals with diabetes.82,83 Multiple system-atic reviews and meta-analyses of randomized controlled trials (RCTs) consistently show that low-GI/GL diets significantly reduce glycated hemoglobin (A1C) compared to high-GI/GL diets. Importantly, these diets also address glycemic variability.84 A dedicated meta-analysis found that a low-GI diet significantly improves measures of glucose fluctuations in patients with type 2 diabetes.85 This is a critical point, as re-search by Pai et al has directly identified high glycemic variability as an independent risk factor for the development of painful diabetic peripheral neuropathy (PDPN).86
Mechanistically, the neuroprotective potential of a low-GI/GL diet lies in its ability to target the root cause of DPN: hyperglycemia and its fluctuations.87 Postprandial glucose spikes are potent triggers of oxidative stress, a primary driver of diabetic complications.88 Research has even shown that acute glucose fluctuations can gener-ate a more intense oxidative stress response than sustained chronic hyperglyce-mia.89 By creating a smoother postprandial glucose profile, a low-GI/GL diet sys-tematically reduces the substrate flux into multiple, well-established pathological pathways.
This concept is further supported by pharmacological evidence. Al-pha-glucosidase inhibitors, such as acarbose, function by slowing intestinal carbohy-drate absorption, effectively mimicking the physiological effect of a low-GI meal.90 In the UKPDS 44 trial, acarbose was shown to improve glycemic control over three years, providing a proof of principle for this mechanism.
In summary, a low-GI/GL diet is a foundational intervention that directly targets the initial steps of DPN pathogenesis. By minimizing postprandial hyperglycemia and glycemic variability, it systematically inhibits multiple interconnected pathological pathways. This reduces glucotoxicity, oxidative stress, and neuroinflammation. Therefore, incorporating low-GI/GL principles into daily eating is a core and effective strategy for the prevention and management of DPN.
Low-Carbohydrate and Ketogenic Diets
Low-carbohydrate diets (LCDs) are a class of dietary patterns that alter macro-nutrient distribution by restricting carbohydrate intake. This restriction exists on a spectrum, from moderate LCDs (<45% of energy from carbohydrates) to very-low-carbohydrate ketogenic diets (KDs), which typically limit carbohydrates to less than 50 grams per day.91
Accumulating evidence suggests potential short-term benefits of LCDs on glycemic control. Multiple systematic reviews and meta-analyses indicate that in patients with type 2 diabetes, LCDs may be associated with a greater reduction in glycated hemoglobin (A1C) at 6 months compared to high-carbohydrate diets.92,93 More encouragingly, preclinical studies suggest an emerging neuroprotective potential. In a type 1 diabetic mouse model, Enders et al reported that a KD might help mitigate and potentially reverse mechanical allodynia and improve epidermal nerve fiber density.94 Another study in diabetic rats observed that a KD could alleviate neuropathic pain.95
Mechanistically, the primary neuroprotective potential of LCDs stems from their ability to bypass glucose metabolism. By drastically reducing carbohydrate influx, LCDs remove the primary substrate for hyperglycemia, thereby directly inhibiting the flux through damaging pathological pathways like the polyol, AGEs, and PKC path-ways.88
The ketogenic diet introduces a unique and deeper layer of mechanisms. In a state of carbohydrate restriction, the liver produces ketone bodies, primarily β-hydroxybutyrate (BHB). Ketones serve as a highly efficient alternative fuel source for neurons, which is particularly beneficial when glucose metabolism is impaired.96 More importantly, ketone bodies also function as signaling molecules.97 First, BHB exerts direct anti-inflammatory effects. Groundbreaking work by Youm et al demon-strated that BHB specifically blocks the activation of the NLRP3 inflammasome, thereby inhibiting the release of pro-inflammatory cytokines like IL-1β and directly countering neuroinflammation.98 Second, BHB is an endogenous histone deacety-lase (HDAC) inhibitor. By inhibiting HDACs, BHB can epigenetically modify gene expression, leading to the upregulation of antioxidant genes and enhancing cellular defenses against oxidative stress.99
Despite the promising mechanisms and short-term benefits, significant contro-versies regarding the long-term application of LCDs must be critically addressed. First, the pronounced initial advantage in glycemic control appears to attenuate over time. Both major meta-analyses and a two-year RCT have shown that the difference in A1C reduction between LCDs and other diets is often no longer statistically significant at 12 to 24 months.92,100,101 This is likely due to challenges with long-term dietary adherence, as highly restrictive diets can be difficult to sustain amidst social and cul-tural food environments.101
Second, cardiometabolic safety, particularly regarding dyslipidemia, remains a primary concern. While many individuals on LCDs see improvements in triglycerides and HDL cholesterol, a significant portion experiences an increase in low-density lip-oprotein cholesterol (LDL-C).102 Although some evidence suggests this may be a shift towards larger, less atherogenic LDL particles, the National Lipid Association advises caution, emphasizing that elevated LDL-C remains a risk factor for cardio-vascular disease regardless of particle size.102 The long-term cardiovascular impact, especially in diets high in saturated fats, is not yet fully understood and requires careful monitoring.
Third, the long-term effects on renal function are another area of debate. While studies in individuals with normal kidney function have not shown adverse effects, there is a theoretical concern that a sustained high-protein intake could increase glo-merular hyperfiltration. Therefore, caution is advised when applying LCDs in patients with pre-existing chronic kidney disease.103 Other potential long-term issues include the risk of micronutrient deficiencies if the diet is not well-formulated and the potential for loss of lean body mass if protein intake is inadequate.101
In conclusion, low-carbohydrate and ketogenic diets represent emerging dietary strategies with multi-faceted mechanistic potential, demonstrating initial short-term efficacy in improving overall glycemic control and suggesting a theoretical basis for neuroprotection.However, given the challenges with long-term adherence, the attenuation of effects over time, and unresolved safety questions, they should be considered as a medical nutrition therapy to be implemented under clinical supervision, rather than a universal, lifelong recommendation for all individuals.104
Microbiota-Targeted Diets
As the understanding of the gut microbiota’s central role in host health deepens, nutritional research is shifting from broad dietary patterns to more specific “microbi-ota-targeted diets”.105 This approach is not a fixed menu but rather a strategy de-signed to actively shape the structure and function of the gut microbial community through precise nutritional inputs. The core tools of this strategy include several key components designed to modulate the gut ecosystem. These include prebiotics, which are typically non-digestible dietary fibers like fructooligosaccharides and inulin that selectively promote the growth of beneficial bacteria;106 and probiotics, which are live microorganisms commonly found in yogurt and fermented foods that confer a health benefit to the host when consumed in adequate amounts.107 A combination of these is known as synbiotics. Furthermore, the approach considers postbiotics, which are the beneficial metabolites produced by microorganisms, such as short-chain fatty acids (SCFAs).105
A growing body of evidence indicates that gut dysbiosis is a key component in the pathophysiology of DPN. An animal model study by Guo et al directly demonstrated that the peripheral neuropathy induced by a high-fat diet was associated with specific gut microbiota signatures, nerve lipid profiles, and nerve gene expression pathways related to inflammation and antioxidant defense.78 Another study found that ginger, a food rich in phytochemicals, could alleviate neuropathic pain in diabetic rats by beneficially modulating the gut microbiome.108 These findings provide a strong ra-tionale for intervening in DPN by targeting the gut microbiota.
Microbiota-targeted diets exert their effects on DPN through multiple mechanisms, centrally involving the modulation of the gut-nerve axis. First, these diets work to re-store gut barrier integrity and reduce metabolic endotoxemia. An unhealthy diet can compromise the tight junctions of the intestinal epithelium, leading to a “leaky gut” that allows bacterial endotoxins like lipopolysaccharide (LPS) to enter the bloodstream and cause systemic low-grade inflammation.79 Prebiotics and probiotics can enhance gut barrier function by nourishing intestinal cells and producing metabolites like butyrate, thereby reducing this primary inflammatory trigger from the source.105
Second, microbiota-targeted diets can directly modulate the host’s immune and inflammatory responses. Wastyk et al found that a diet rich in fermented foods (a source of probiotics) was more effective than a high-fiber diet at reducing multiple in-flammatory markers, including interleukin-6 (IL-6).109 This is highly relevant, as inflammation is a central mechanism of nerve damage in DPN. Furthermore, SCFAs produced from prebiotic fermentation, particularly butyrate, have been shown to in-hibit the NLRP3 inflammasome,98 a mechanism of inflammation control that par-allels the action of ketones.
Most importantly, these diets exert their effects by producing neuroactive postbi-otics, primarily SCFAs. When beneficial bacteria ferment dietary fibers (prebiotics), they produce large amounts of SCFAs like butyrate, propionate, and acetate.110 These molecules not only serve as energy for intestinal cells but also enter the circula-tion and can influence the nervous system through various pathways.111,112 A critical animal study by Sheval et al provided direct evidence for this, showing that prebiotics (galactooligosaccharides and resistant starch) effectively protected against high-fat diet-induced peripheral neuropathy and mechanical hypersensitivity.113 This suggests that promoting endogenous SCFA production through prebiotics is a key neuroprotective mechanism of microbiota-targeted diets.
The effects of these interventions can be monitored through biomarkers. In addi-tion to traditional inflammatory markers like hs-CRP and IL-6, circulating levels of SCFAs are emerging as a promising biomarker. Research by Müller et al indicates that circulating, rather than fecal, SCFA levels are closely related to metabolic markers like insulin sensitivity.114
In summary, microbiota-targeted diets represent a sophisticated, mecha-nism-based intervention. By using prebiotics, probiotics, and fermented foods to op-timize the gut ecosystem, this strategy can systematically improve gut barrier function, reduce systemic inflammation, and generate neuroprotective metabolites, offering a precise and promising new avenue for the prevention and management of DPN.115,116
Intermittent Fasting
Intermittent fasting (IF) is a unique dietary intervention that focuses on when to eat, rather than what to eat. It creates regular, cyclical periods of fasting, with common approaches including time-restricted eating (TRE) and alternate-day fasting.117 This strategy does not prescribe specific foods but instead aims to reset metabolic and cel-lular repair processes by altering the timing of nutrient intake.
A growing body of evidence points toward potential metabolic benefits of IF. Several randomized controlled trials (RCTs) in humans have indicated that TRE may improve insulin sensitivity, lower blood pressure, and reduce oxidative stress, even without weight loss.118 Other RCTs have confirmed its effectiveness for weight loss and for improving cardiometabolic markers in adults with obesity.119 For pa-tients with type 2 diabetes, systematic reviews and meta-analyses provide high-level evidence that IF effectively improves glucose, lipids, and insulin resistance.120,121 While direct clinical trials on DPN are still in their infancy, these findings are highly relevant, as improving these upstream metabolic drivers is a foundational strategy for mitigating nerve damage.
The neuroprotective potential of IF extends beyond metabolic improvement, with its most compelling and unique mechanism being the potent induction of autophagy. Autophagy is a critical intracellular process for clearing and recycling damaged com-ponents, such as dysfunctional mitochondria and misfolded proteins.122 In the pathogenesis of DPN, neurons are burdened by these damaged elements due to glu-cotoxicity and oxidative stress. A groundbreaking study by Alirezaei et al was the first to demonstrate that even short-term fasting induces profound autophagy in neurons.123 By activating this process, IF may help neurons clear harmful metabolic by-products and repair damaged mitochondria, thereby promoting cellular repair and enhancing neuronal survival.124
Furthermore, IF, particularly TRE, helps synchronize nutrient intake with the body’s internal circadian rhythms. Metabolism is tightly regulated by the biological clock, and eating at misaligned times can disrupt this rhythm and exacerbate metabolic stress.125,126 A study by Jamshed et al directly linked early time-restricted feeding to improved 24-hour glucose levels and beneficial changes in markers of the circadian clock, autophagy, and inflammation.127 Furthermore, preclinical evidence suggests that fasting has the potential to promote the regeneration of peripheral nerves and may upregulate levels of neurotrophic factors128 and increase levels of neurotrophic factors like brain-derived neurotrophic factor (BDNF), which is essential for neuronal survival, growth, and plasticity.129
In conclusion, intermittent fasting is an intriguing conceptual intervention that may target DPN through multiple, potentially synergistic pathways. Its hypothesized ability to stimulate neuronal autophagy offers a novel theoretical angle for nerve repair that is less prominent in other dietary patterns. However, research in this area is still in its early stages. Well-designed RCTs are urgently needed to evaluate the efficacy and safety of IF in patients with established diabetic peripheral neuropathy130,131 (Figure 2).
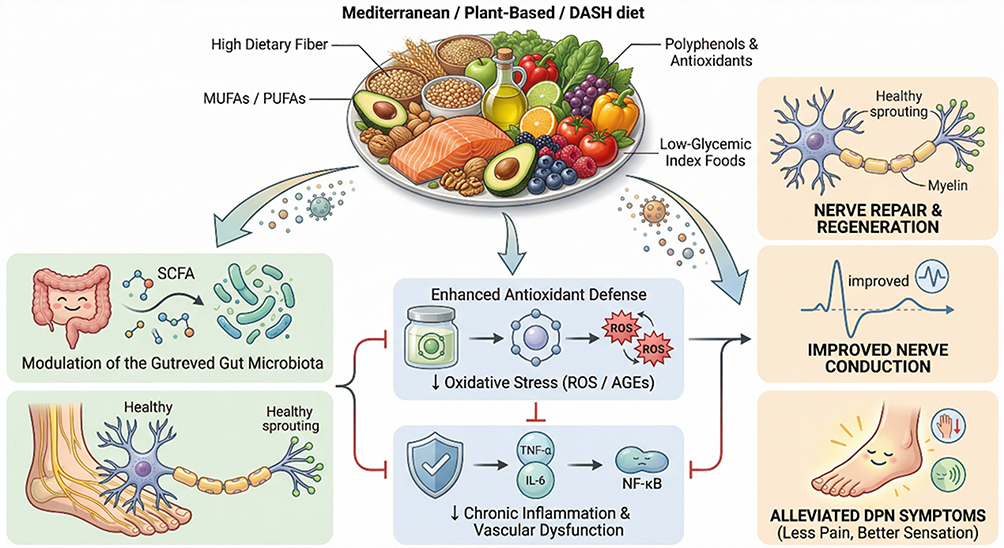 |
Figure 2 Neuroprotective mechanisms of healthy dietary patterns in DPN. Healthy dietary patterns (e.g., Mediterranean, DASH, plant-based) modulate the gut-nerve axis and promote SCFA production via bioactive nutrients like fiber and antioxidants. These interventions inhibit neuroinflammation and oxidative stress (e.g., ROS and AGEs), facilitating nerve fiber regeneration and improving nerve conduction velocity (NCV), ultimately alleviating neuropathic pain and sensory symptoms. Abbreviations: DPN, diabetic peripheral neuropathy; SCFA, short-chain fatty acids; ROS, reactive oxygen species; AGEs, advanced glycation end products; NCV, nerve conduction velocity. |
Discussion
The pathogenesis of diabetic peripheral neuropathy (DPN) is driven by a complex network of interconnected metabolic, oxidative, and inflammatory pathways.4–6 This review establishes that dietary patterns are a pivotal, modifiable factor capable of influencing these core mechanisms. Healthy dietary patterns, such as the Mediterra-nean and plant-based diets, appear to confer neuroprotection by targeting multiple pathways simultaneously.33,56 Their rich composition of antioxidants and an-ti-inflammatory compounds counteracts the damage from oxidative stress and chronic inflammation, which is reflected in the improvement of relevant biomarkers.132 Conversely, the Western dietary pattern, high in saturated fats and refined carbohy-drates, likely exacerbates DPN by fueling these detrimental pathways.73,74,133
Despite compelling mechanistic evidence, a critical gap exists in the literature: the profound lack of high-quality intervention studies targeting patients already diag-nosed with DPN. Most research focuses on primary prevention, leaving it unclear whether dietary modification can serve as a therapeutic strategy to slow progression or alleviate symptoms in those already suffering from neuropathy. Therefore, there is an urgent need to shift the research focus toward this population. Well-designed randomized controlled trials are required to build an evidence base for dietary inter-ventions as a form of secondary prevention and to develop more precise, personalized nutritional therapies.
Abbreviations
The following abbreviations are used in this article: AGEs, Advanced Glycation End Products; AR, Aldose Reductase; CGRP, Calcitonin Gene-Related Peptide; DAG, Diacylglycerol; DN, Diabetic Nephropathy; DPN, Diabetic Peripheral Neuropathy; DSPN, Diabetic Sensorimotor Polyneuropathy; GSH, Reduced Glutathione; HBP, Hexosamine Biosynthesis Pathway; HDL-C, High-Density Lipoprotein Cholesterol; hs-CRP, High-Sensitivity C-Reactive Protein; IENF, Intraepidermal Nerve Fiber Density; IL, Interleukin; IR, Insulin Resistance; IRS, Insulin Receptor Substrate; Lp(a), Lipoprotein(a); MD, Mediterranean Diet; MUFA, Monounsaturated Fatty Acids; NAD+, Nicotinamide Adenine Dinucleotide; NADPH, Reduced Nicotinamide Adenine Dinucleotide Phosphate; NF-κB, Nuclear Factor-κB; NLR, Neutrophil-to-Lymphocyte Ratio; O-GlcNAc, O-linked N-acetylglucosamine; oxLDL, Oxidized Low-Density Lipoprotein; PAI-1, Plasminogen Activator Inhibitor-1; PARP, Poly(ADP-ribose) Polymerase; PKC, Protein Kinase C; RAGE, Receptor for Advanced Glycation End Products; ROS, Reactive Oxygen Species; SAA, Serum Amyloid A; Scr, Serum Creatinine; SDH, Sorbitol Dehydrogenase; SFA, Saturated Fatty Acids; SUA, Serum Uric Acid; T2DM, Type 2 Diabetes Mellitus; TCSS, Toronto Clinical Scoring System; TG, Triglycerides; TGF-β, Transforming Growth Factor-β; TIR, Time in Range; TNF-α, Tumor Necrosis Factor-alpha; TSH, Thyroid-Stimulating Hormone; TyG, Triglyceride-glucose; UACR, Urinary Albumin-to-Creatinine Ratio; UDP-GlcNAc, Uridine Diphosphate N-acetylglucosamine; VEGF, Vascular Endothelial Growth Factor; WMR, White Blood Cell Count to Mean Platelet Volume Ratio.
Data Sharing Statement
Data sharing is not applicable to this article as new data were created.
Acknowledgments
During the preparation of this article, the author(s) used Google’s Gemini for the purposes of language refinement. The authors have reviewed and edited the output and take full responsibility for the content of this publication.
Author Contributions
Ruoxiang Sun: Conceptualization, methodology, formal analysis, investigation, data curation, writing—original draft preparation, visualization. Xu Li: Methodology, validation, writing—review and editing. Xianping Liu: Methodology, validation, writing—review and editing. Dongjie Guo: Formal analysis, data curation, writing—review and editing. Yuanqin Chen: Formal analysis, data curation, writing—review and editing. Yu Zhao: Investigation, data curation, writing—review and editing. Yanxiang Yin: Investigation, data curation, writing—review and editing. Xingyue Zhai: Conceptualization, validation, resources, writing—review and editing, supervision, project administration, funding acquisition. All authors gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This research was funded by the Natural Science Foundation of Liaoning Province, grant number 2023-MS-271, for the project “A study on the mechanism of oyster polysaccharides in regulating type 2 diabetes through the gut microbiota-bile acid pathway”, 2023.
Disclosure
The authors declare no conflicts of interest.
References
1. Savelieff MG, Elafros MA, Viswanathan V, Jensen TS, Bennett DL, Feldman EL. The global and regional burden of diabetic peripheral neuropathy. Nature Reviews Neurology. 2025;21(1):17–20. doi:10.1038/s41582-024-01041-y
2. ElSayed NA, Aleppo G, Bannuru RR, et al. 12. Retinopathy, Neuropathy, and Foot Care: standards of Care in Diabetes—2024. Diabetes Care. 2024;47:S231–S243. doi:10.2337/dc24-S012
3. International Diabetes Federation. IDF Diabetes Atlas.
4. Lu X, Xie Q, Pan X, et al. Type 2 diabetes mellitus in adults: pathogenesis, prevention and therapy. Signal Transduct Target Ther. 2024;9:110. doi:10.1038/s41392-024-01824-0
5. Soni K, Srivastava DK, Kumar S, Bajpai PK, Shukla SK, Prakash S. Prevalence of Diabetic Peripheral Neuropathy among Type 2 Diabetes Mellitus patients and its associated risk factors. Indian J Commun Health. 2024;36:208–214. doi:10.51910/ijch.v36i2.645
6. Dubský M, Sojáková D, Fejfarová V, Jude EB. Diabetic Peripheral Neuropathy: new Diagnostics and Treatment Perspectives. J Clin Med. 2025;14:456. doi:10.3390/jcm14020456
7. García-Redondo AB, Aguado A, Briones AM, Salaices M. NADPH oxidases and vascular remodeling in cardiovascular diseases. Pharmacol Res. 2016;114:110–120. doi:10.1016/j.phrs.2016.10.021
8. Vistoli G, De Maddis D, Cipak A, Zarkovic N, Carini M, Aldini G. Advanced glycoxidation and lipoxidation end products (AGEs and ALEs): an overview of their mechanisms of formation. Free Radical Research. 2013;47(sup1):3–27. doi:10.3109/10715762.2013.815348
9. Hashim M, Akhtar J, Khan MI, Ahmad M, Islam A, Ahmad A. Diabetic neuropathy: an overview of molecular pathways and ProtectiveMechanisms of phytobioactives. Endocr Metab Immune Disord. Drug Targets. 2024;24:758–776. doi:10.2174/1871530323666230510153328
10. Pan D, Xu L, Guo M. The role of protein kinase C in diabetic microvascular complications. Front Endocrinol. 2022;13:973058. doi:10.3389/fendo.2022.973058
11. Dutra RC, Bicca MA, Segat GC, et al. The antinociceptive effects of the tetracyclic triterpene euphol in inflammatory and neuropathic pain models: the potential role of PKCε. Neuroscience. 2015;303:126–137. doi:10.1016/j.neuroscience.2015.06.056
12. Martín-Guerrero SM, Casado P, Hijazi M, et al. PARP-1 activation after oxidative insult promotes energy stress-dependent phosphorylation of YAP1 and reduces cell viability. Biochem. J. 2020;477:4491–4513. doi:10.1042/BCJ20200540
13. Wu S, Li X, Gao F, De Groot JF, Koul D, Yung WKA. PARP-mediated PARylation of MGMT is critical to promote repair of temozolomide-induced O6-methylguanine DNA damage in glioblastoma. Neuro-Oncoll. 2021;23:920–931. doi:10.1093/neuonc/noaa283
14. Rendra E, Riabov V, Mossel DM, Sevastyanova T, Harmsen MC, Kzhyshkowska J. Reactive oxygen species (ROS) in macrophage activation and function in diabetes. Immunobiology. 2019;224:242–253. doi:10.1016/j.imbio.2018.11.010
15. Obrosova IG, Drel VR, Pacher P, et al. Oxidative-nitrosative stress and poly(ADP-ribose) polymerase (PARP) activation in experimental diabetic neuropathy. Diabetes. 2005;54:3435–3441. doi:10.2337/diabetes.54.12.3435
16. Shen G, Zhou J, Chen Y, et al. A study on the correlation between serum uric acid and diabetic peripheral neuropathy in patients with type 2 diabetes. Chin J Diabetes. 2023;31:764–768. doi:10.3969/j.issn.1006-6187.2023.10.008
17. Zhao Z, Xu J, Hong J. Study on the interaction between serum uric acid and pancreatic β-cell secretory function in patients with type 2 diabetic peripheral neuropathy. Chin J Diabetes. 2024;32:813–820. doi:10.3969/j.issn.1006-6187.2023.11.006
18. Guo L, Hua F, Liu T. Study on the interaction of glycated hemoglobin and dyslipidemia on peripheral neuropathy in diabetic patients. Chin J Diabetes. 2024;32:595–600. doi:10.3969/j.issn.1006-6187.2023.08.007
19. Zhang C, Xue L, Liu X, Song Y. Correlation study between EMG parameters and metabolic indicators in DPN patients. J Jinzhou Med Univ. 2025;46:88–94.
20. Casadei G, Filippini M, Brognara L. Glycated hemoglobin (HbA1c) as a biomarker for diabetic foot peripheral neuropathy. J Pers Med. 2022;12:1563. doi:10.3390/jpm12101563
21. Coppini DV, Spruce MC, Thomas P, Masding MG. Established diabetic neuropathy seems irreversible despite improvements in metabolic and vascular risk markers—a retrospective case–control study in a hospital patient cohort. Diabet Med. 2006;23:1016–1020. doi:10.1111/j.1464-5491.2006.01945.x
22. Wang D, Shi X, Deng X, et al. A study on the correlation between time in range of glucose and diabetic complications and comorbidities in patients with type 2 diabetes. Chin J Diabetes. 2023;31:486–490. doi:10.3969/j.issn.1006-6187.2023.07.002
23. Rosales-Hernandez A, Cheung A, Podgorny P, Chan C, Toth C. Absence of clinical relationship between oxidized low density lipoproteins and diabetic peripheral neuropathy: a case control study. Lipids Health Dis. 2014;13:32. doi:10.1186/1476-511X-13-32
24. Zhang Y, Jian J, Lou X, Zhu T. Correlation between Toronto Clinical Scoring System scores and inflammatory status in patients with diabetic peripheral neuropathy. Chin J Diabetes. 2019;27:352–356. doi:10.3969/j.issn.1006-6187.2019.05.006
25. Wei W, Zhao S, Lü X, Dong J, Mao H. Correlation analysis of diabetic peripheral neuropathy and inflammatory factors. Chin J Diabetes. 2020;28:326–330. doi:10.3969/j.issn.1006-6187.2020.05.002
26. Fritzen AM, Lundsgaard AM, Jordy AB, et al. New nordic diet–induced weight loss is accompanied by changes in metabolism and AMPK signaling in adipose tissue. J Clin Endocrinol Metab. 2015;100:3509–3519. doi:10.1210/jc.2015-1886
27. Rumora AE, Lograsso G, Hayes JM, et al. The divergent roles of dietary saturated and monounsaturated fatty acids on nerve function in murine models of obesity. J Neurosci. 2019;39:3770–3781. doi:10.1523/JNEUROSCI.2951-18.2019
28. Martín-Peláez S, Fito M, Castaner O. Mediterranean diet effects on type 2 diabetes prevention, disease progression, and related mechanisms. A review. Nutrients. 2020;12:2236. doi:10.3390/nu12082236
29. Krznarić Ž, Karas I, Ljubas Kelečić D, Vuletić M. The Mediterranean and nordic diet: a review of differences and similarities of two sustainable, health-promoting dietary patterns. Front Nutr. 2021;8:683678. doi:10.3389/fnut.2021.683678
30. De Mello VDF, Schwab U, Kolehmainen M, et al. A diet high in fatty fish, bilberries and wholegrain products improves markers of endothelial function and inflammation in individuals with impaired glucose metabolism in a randomised controlled trial: the sysdimet study. Diabetologia. 2011;54:2755–2767. doi:10.1007/s00125-011-2293-1
31. Kanerva N, Loo BM, Eriksson JG, et al. Associations of the baltic sea diet with obesity-related markers of inflammation. Annals of Medicine. 2014;46:90–96. doi:10.3109/07853890.2013.868722
32. Galbete C, Kröger J, Jannasch F, et al. Nordic diet, Mediterranean diet, and the risk of chronic diseases: the EPIC-potsdam study. BMC Med. 2018;16:99. doi:10.1186/s12916-018-1082-8
33. Zooravar D, Soltani P, Khezri S, et al. Mediterranean diet and diabetic microvascular complications: a systematic review and meta-analysis. BMC Nutr. 2025;11:66. doi:10.1186/s40795-024-00911-0
34. Qu C, Zhao J, Lai J, et al. Adherence to a Mediterranean diet is associated with a lower risk of diabetic kidney disease among individuals with hyperglycemia: a prospective cohort study. BMC Med. 2024;22:224. doi:10.1186/s12916-024-03444-1
35. Zunica-Garcia S, Blanquer-Gregori JJ, Sanchez-Ortiga R, et al. Relationship between diabetic peripheral neuropathy and adherence to the Mediterranean diet in patients with type 2 diabetes mellitus: an observational study. J Endocrinol Invest. 2024;47:2603–2613. doi:10.1007/s40618-024-02377-z
36. Landberg R, Hanhineva K. Biomarkers of a healthy nordic diet—from dietary exposure biomarkers to microbiota signatures in the metabolome. Nutrients. 2020;12(27). doi:10.3390/nu12010027
37. Roager HM, Licht TR, Poulsen SK, et al. Microbial enterotypes, inferred by the prevotella-to-bacteroides ratio, remained stable during a 6-month randomized controlled diet intervention with the new nordic diet. Appl Environ Microbiol. 2014;80:1142–1149. doi:10.1128/AEM.03541-13
38. Ghaemi F, Dehghani Firouzabadi F, Moosaie F, et al. Effects of a Mediterranean diet on the development of diabetic complications: a longitudinal study from the nationwide diabetes report of the national program for prevention and control of diabetes (NPPCD 2016-–2020). Maturitas. 2021;153:61–67. doi:10.1016/j.maturitas.2021.08.006
39. Salas-Salvadó J, Becerra-Tomás N, Papandreou C, et al. Dietary patterns emphasizing the consumption of plant foods in the management of type 2 diabetes: a narrative review. Adv Nutr. 2019;10:S320–S331. doi:10.1093/advances/nmy114
40. Ard JD, Grambow SC, Liu D, et al. The effect of the PREMIER interventions on insulin sensitivity. Diabetes Care. 2004;27:340–347. doi:10.2337/diacare.27.2.340
41. Shirani F, Salehi-Abargouei A, Azadbakht L. Effects of dietary approaches to stop hypertension (DASH) diet on some risk for developing type 2 diabetes: a systematic review and meta-analysis on controlled clinical trials. Nutrition. 2013;29:939–947. doi:10.1016/j.nut.2013.01.002
42. Ramesh G, Wood AC, Allison MA, et al. Associations between adherence to the dietary approaches to stop hypertension (DASH) diet and six glucose homeostasis traits in the Microbiome and Insulin Longitudinal Evaluation Study (MILES). Nutr Metab Cardiovasc Dis. 2022;32:1418–1426. doi:10.1016/j.numecd.2022.03.003
43. Golzarand M, Moslehi N, Mirmiran P, et al. Adherence to the DASH, MeDi, and MIND diet scores and the incidence of metabolically unhealthy phenotypes. Obesity Research Clin Pract. 2023;17:226–232. doi:10.1016/j.orcp.2023.05.004
44. Azadbakht L, Pour Fard NR, Karimi M, et al. Effects of the dietary approaches to stop hypertension (DASH) eating plan on cardiovascular risks among type 2 diabetic patients. Diabetes Care. 2011;34:55–57. doi:10.2337/dc10-1381
45. Azadbakht L, Surkan PJ, Esmaillzadeh A, et al. The dietary approaches to stop hypertension eating plan affects C-reactive protein, coagulation abnormalities, and hepatic function tests among type 2 diabetic patients. J Nutr. 2011;141:1083–1088. doi:10.1093/jn/nxr022
46. Weber KS, Schlesinger S, Lang A, et al. Association of dietary patterns with diabetes-related comorbidities varies among diabetes endotypes. Nutr Metab Cardiovasc Dis. 2024;34:911–924. doi:10.1016/j.numecd.2023.12.016
47. Yang Y, Liu L, Wang H, et al. Development and assessment of an early diagnostic approach for painful diabetic peripheral neuropathy using basic clinical and laboratory parameters. Exp Gerontol. 2025;209:112847. doi:10.1016/j.exger.2025.112847
48. Lien LF, Brown AJ, Ard JD, et al. Effects of PREMIER lifestyle modifications on participants with and without the metabolic syndrome. Hypertension. 2007;50:609–616. doi:10.1161/HYPERTENSIONAHA.106.082719
49. Mcmacken M, Shah S. A plant-based diet for the prevention and treatment of type 2 diabetes. J Geriatric Cardiol. 2017;14:342–354. doi:10.11909/j.issn.1671-5411.2017.05.009
50. Willett W, Rockström J, Loken B, et al. Food in the Anthropocene: the EAT-Lancet Commission on healthy diets from sustainable food systems. Lancet. 2019;393:447–492. doi:10.1016/S0140-6736(18)33179-9
51. Satija A, Bhupathiraju SN, Rimm EB, et al. Plant-based dietary patterns and incidence of type 2 diabetes in US men and women: results from three prospective cohort studies. PLoS Med. 2016;13:e1002039. doi:10.1371/journal.pmed.1002039
52. Wang Y, Liu B, Han H, et al. Associations between plant-based dietary patterns and risks of type 2 diabetes, cardiovascular disease, cancer, and mortality – a systematic review and meta-analysis. Nutr J. 2023;22:46. doi:10.1186/s12937-023-00868-x
53. Guest NS, Raj S, Landry MJ, et al. Vegetarian and vegan dietary patterns to treat adult type 2 diabetes: a systematic review and meta-analysis of randomized controlled trials. Adv Nutr. 2024;15:100294. doi:10.1016/j.advnut.2024.100294
54. Yokoyama Y, Barnard ND, Levin SM, et al. Vegetarian diets and glycemic control in diabetes: a systematic review and meta-analysis. Cardiovas Diagnosis Ther. 2014;4:373–382. doi:10.3978/j.issn.2223-3652.2014.10.04
55. Petermann-Rocha F, Parra-Soto S, Gray S, et al. Plant-based and vegetarian diets and the risk of developing type 2 diabetes in the UK Biobank: a prospective cohort study. Diabetes Care. 2021;44:984–991. doi:10.2337/dc20-2387
56. Asadi S, Shiva F, Mohtashamian A, et al. Dietary phytochemical index and its relationship with diabetic sensorimotor polyneuropathy: a case-control study. Sci Rep. 2024;14:21688. doi:10.1038/s41598-024-72602-w
57. Tomova A, Bukovsky I, Rynkiewicz R, et al. The effects of vegetarian and vegan diets on gut microbiota. Nutrients. 2019;11:474. doi:10.3390/nu11030474
58. Ley SH, Hamdy O, Mohan V, et al. Prevention and management of type 2 diabetes: dietary components and nutritional strategies. Lancet. 2014;383:1999–2007. doi:10.1016/S0140-6736(14)60613-9
59. Liu Q, Sun L, Tan Y, et al. Role of iron deficiency and overload in the pathogenesis of diabetes and diabetic complications. Curr Med Chem. 2009;16:113–129. doi:10.2174/092986709787002890
60. Turner-Mcgrievy GM, Barnard ND, Scialli AR. A two-year randomized weight loss trial comparing a vegan diet to a more moderate low-fat diet. Obesity. 2007;15:2276–2281. doi:10.1038/oby.2007.270
61. Barbaresko J, Mena-Sánchez A, Noriega-Rodríguez A, et al. Dietary pattern analysis and biomarkers of low-grade inflammation: a systematic review of randomized controlled trials. Nutr Rev. 2019;77:1–19. doi:10.1093/nutrit/nuy036
62. Morris MC, Tangney CC, Wang Y, et al. MIND diet associated with reduced incidence of Alzheimer’s disease. Alzheimer’s Dementia. 2015;11:1007–1014. doi:10.1016/j.jalz.2014.11.009
63. Hosseinpour-Niazi S, Moghaddam-Amiri S, Mirmiran P, et al. Adherence to the MIND diet is associated with a lower risk of type 2 diabetes. Diab Res Clin Pract. 2021;181:109095. doi:10.1016/j.diabres.2021.109095
64. Devore EE, Kang JH, Breteler MMB, et al. Dietary intakes of berries and flavonoids in relation to cognitive decline. Ann Neurol. 2012;72:135–143. doi:10.1002/ana.23594
65. Morris MC, Wang Y, Barnes LL, et al. Nutrients and bioactives in green leafy vegetables and cognitive decline: prospective study. Neurology. 2018;90:e214–e222. doi:10.1212/WNL.0000000000004815
66. Chen C, Hayden KM, Kaufman JD, et al. Adherence to a MIND-Like Dietary Pattern, Long-Term Exposure to Fine Particulate Matter Air Pollution, and MRI-Based Measures of Brain Volume: the Women’s Health Initiative Memory Study-MRI. Environ Health Perspect. 2021;129:127008. doi:10.1289/EHP9374
67. Dhana K, James BD, Agarwal P, et al. MIND diet, common brain pathologies, and cognition in community-dwelling older adults. J Alzheimers Dis. 2021;83:683–692. doi:10.3233/JAD-210107
68. Gong D, Lai WF. Dietary patterns and type 2 diabetes: a narrative review. Nutrition. 2025;140:112905. doi:10.1016/j.nut.2025.112905
69. Shivappa N, Steck SE, Hurley TG, et al. Designing and developing a literature-derived, population-based dietary inflammatory index. Public Health Nutrition. 2014;17:1689–1696. doi:10.1017/S1368980013002927
70. Monteiro CA, Cannon G, Levy RB, et al. Ultra-processed foods: what they are and how to identify them. Public Health Nutrition. 2019;22:936–941. doi:10.1017/S1368980018003762
71. Maghsoudi Z, Ghiasvand R, Salehi-Abargouei A, et al. Empirically derived dietary patterns and incident type 2 diabetes mellitus: a systematic review and meta-analysis on prospective observational studies. Public Health Nutrition. 2016;19:230–241. doi:10.1017/S136898001500130X
72. Chen Z, Khandpur N, Desjardins C, et al. Ultra-processed food consumption and risk of type 2 diabetes: three large prospective U.S. cohort studies. Diabetes Care. 2023;46:1335–1344. doi:10.2337/dc22-1718
73. Umbaugh DS, Maciejewski JC, Wooten JS, et al. Neuronal inflammation is associated with changes in epidermal innervation in high fat fed mice. Front Physiol. 2022;13:891550. doi:10.3389/fphys.2022.891550
74. Jaroslawska J, Korytko A, Zglejc-Waszak K, et al. Peripheral neuropathy presents similar symptoms and pathological changes in both high-fat diet and pharmacologically induced pre- and diabetic mouse models. Life. 2021;11:1267. doi:10.3390/life11111267
75. Hossain MJ, Kendig MD, Wild BM, et al. Evidence of altered peripheral nerve function in a rodent model of diet-induced prediabetes. Biomedicines. 2020;8:313. doi:10.3390/biomedicines8090313
76. Wang P, Song M, Eliassen AH, et al. Optimal dietary patterns for prevention of chronic disease. Nat Med. 2023;29:719–728. doi:10.1038/s41591-023-02237-8
77. Zinöcker MK, Lindseth IA. The Western diet–microbiome-host interaction and its role in metabolic disease. Nutrients. 2018;10:366. doi:10.3390/nu10030366
78. Guo K, Figueroa-Romero C, Noureldein M, et al. Gut microbiota in a mouse model of obesity and peripheral neuropathy associated with plasma and nerve lipidomics and nerve transcriptomics. Microbiome. 2023;11:52. doi:10.1186/s40168-023-01490-6
79. Cani PD, Amar J, Iglesias MA, et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes. 2007;56:1761–1772. doi:10.2337/db06-1491
80. Christ A, Lautenbach M, Lazar M. Western diet and the immune system: an inflammatory Connection. Immunity. 2019;51:794–811. doi:10.1016/j.immuni.2019.09.020
81. Bell KJ, Brand-Miller J, Colagiuri S. Diabetes and Insulin Resistance. In: Present Knowledge in Nutrition.
82. Zafar MI, Mills KE, Zheng J, et al. Low-glycemic index diets as an intervention for diabetes: a systematic review and meta-analysis. Am J Clin Nutr. 2019;110:891–902. doi:10.1093/ajcn/nqz162
83. Chiavaroli L, Lee D, Ahmed A, et al. Effect of low glycaemic index or load dietary patterns on glycaemic control and cardiometabolic risk factors in diabetes: systematic review and meta-analysis of randomised controlled trials. BMJ. 2021;374:n1651. doi:10.1136/bmj.n1651
84. Shourbagi S, Abbasi B, Heidari-Beni M, et al. The effect of a low-glycemic index diet on glycemic variability in patients with type 2 diabetes: a systematic review and meta-analysis of randomized controlled trials. Diab Res Clin Pract. 2021;181:109088. doi:10.1016/j.diabres.2021.109088
85. Pai YW, Lin CH, Lee IT, et al. Variability of fasting plasma glucose and the risk of painful diabetic peripheral neuropathy in patients with type 2 diabetes. Diabetes Metabolism. 2018;44:129–134. doi:10.1016/j.diabet.2017.07.001
86. Lu Y, Xing P, Cai X, et al. Prevalence and risk factors for diabetic peripheral neuropathy in type 2 diabetic patients from 14 countries: estimates of the INTERPRET-DD study. Front Public Health. 2020;8:534372. doi:10.3389/fpubh.2020.534372
87. Ceriello A. Postprandial hyperglycaemia and diabetes complications: is it time to treat? Diabetes. 2005;54:1–7. doi:10.2337/diabetes.54.1.1
88. Giacco F, Brownlee M. Oxidative stress and diabetic complications. Circulation Research. 2010;107:1058–1070. doi:10.1161/CIRCRESAHA.110.223545
89. Monnier L, Mas E, Gineton A, et al. Activation of oxidative stress by acute glucose fluctuations compared with sustained chronic hyperglycemia in patients with type 2 diabetes. JAMA. 2006;295:1681–1687. doi:10.1001/jama.295.14.1681
90. Holman RR, Cull CA, Turner RC. A randomized double-blind trial of acarbose in type 2 diabetes shows improved glycemic control over 3 years (UKPDS 44). Diabetes Care. 1999;22:960–964. doi:10.2337/diacare.22.6.960
91. Feinman RD, Pogozelski WK, Astrup A, et al. Dietary carbohydrate restriction as the first approach in diabetes management: critical review and evidence base. Nutrition. 2015;31:1–13. doi:10.1016/j.nut.2014.06.011
92. Goldenberg JZ, Day A, Brinkworth GD, et al. Efficacy and safety of low and very low carbohydrate diets for type 2 diabetes remission: systematic review and meta-analysis of published and unpublished randomized trial data. BMJ. 2021;372:m4743. doi:10.1136/bmj.m4743
93. Snorgaard O, Poulsen GM, Andersen HK, et al. Systematic review and meta-analysis of dietary carbohydrate restriction in patients with type 2 diabetes. BMJ Open Diabetes Res Care. 2017;5:e000354. doi:10.1136/bmjdrc-2016-000354
94. Enders JC, Swanson MT, Ryals JM, et al. A ketogenic diet reduces mechanical allodynia and improves epidermal innervation in diabetic mice. Pain. 2022;163:e682–e689. doi:10.1097/j.pain.0000000000002441
95. Bozkurt HS, Kilic E, Cicek M, et al. The effect of the ketogenic diet on neuropathic pain in diabetic rats. J Pain. 2021;22:1106–1114. doi:10.1016/j.jpain.2021.03.010
96. Vidali S, Aminzadeh S, Lambert B, et al. Ketogenic diet in neurological diseases: a systematic review. Front Neurosci. 2017;11:403. doi:10.3389/fnins.2017.00403
97. Newman JC, Verdin E. Ketone bodies as signaling metabolites. Trends Endocrinol Metab. 2017;28:595–608. doi:10.1016/j.tem.2017.04.008
98. Youm YH, Nguyen KY, Grant RW, et al. The ketone metabolite β-hydroxybutyrate blocks NLRP3 inflammasome–mediated inflammatory disease. Nat Med. 2015;21:263–269. doi:10.1038/nm.3804
99. Shimazu T, Hira Sche Y, Kawamura T, et al. Suppression of oxidative stress by β-hydroxybutyrate, an endogenous histone deacetylase inhibitor. Science. 2013;339:211–214. doi:10.1126/science.1227166
100. Tay J, Thompson CH, Luscombe-Marsh ND, et al. Effects of an energy-restricted low-carbohydrate, high unsaturated fat/low saturated fat diet versus a high-carbohydrate, low-fat diet in type 2 diabetes: a 2-year randomized clinical trial. Diabetes Obesity Metab. 2018;20:858–871. doi:10.1111/dom.13174
101. Brouns F. Overweight and diabetes prevention: is a low-carbohydrate-high-fat diet a good alternative? Eur J Nutr. 2018;57:1301–1312. doi:10.1007/s00394-018-1636-z
102. Kirkpatrick CF, Boloick JP, Hecht C, et al. Review of current evidence and clinical recommendations on the effects of low-carbohydrate and very-low-carbohydrate (including ketogenic) diets for the management of body weight and other cardiometabolic risk factors: a scientific statement from the National Lipid Association. J Clin Lipidol. 2019;13:689–711.e1. doi:10.1016/j.jacl.2019.08.003
103. Paoli A. Ketogenic diet for obesity: friend or foe? Int J Environ Res Public Health. 2014;11:2092–2107. doi:10.3390/ijerph110202092
104. Evert AB, Dennison M, Gardner CD, et al. Nutrition therapy for adults with diabetes or prediabetes: a consensus report. Diabetes Care. 2019;42:731–754. doi:10.2337/dci19-0014
105. Valdes AM, Walter J, Segal E, et al. Role of the gut microbiota in nutrition and health. BMJ. 2018;361:k2179. doi:10.1136/bmj.k2179
106. Gibson GR, Hutkins R, Sanders ME, et al. Expert consensus document: the International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat Rev Gastroenterol Hepatol. 2017;14:491–502. doi:10.1038/nrgastro.2017.75
107. Marco ML, Heeney D, Binding S, et al. Health benefits of fermented foods: microbiota and beyond. Curr Opin Biotechnol. 2017;44:94–102. doi:10.1016/j.copbio.2016.11.010
108. Shen CL, Wang R, Santos JM, et al. Ginger alleviates mechanical hypersensitivity and anxio-depressive behavior in rats with diabetic neuropathy through beneficial actions on gut microbiome composition, mitochondria, and neuroimmune cells of colon and spinal cord. Nutr Res. 2024;124:73–84. doi:10.1016/j.nutres.2024.02.003
109. Wastyk HC, Fragiadakis GK, Perelman D, et al. Gut-microbiota-targeted diets modulate human immune status. Cell. 2021;184:4137–4153.e14. doi:10.1016/j.cell.2021.06.019
110. Zhao L, Zhang F, Ding X, et al. Gut bacteria selectively promoted by dietary fibers alleviate type 2 diabetes. Science. 2018;359:1151–1156. doi:10.1126/science.aao5774
111. Dalile B, Van Oudenhove L, Vervliet B, et al. The role of the gut-brain axis in mental health. Eur Neuropsychopharmacol. 2019;29:1339–1350. doi:10.1016/j.euroneuro.2019.09.012
112. Silva YP, Bernardi A, Frozza RL. The role of short-chain fatty acids from gut microbiota in gut-brain communication. Front Endocrinol. 2020;11:25. doi:10.3389/fendo.2020.00025
113. Sheval V, Yorek MS, Khan A, et al. Prebiotic galactooligosaccharides and resistant starch protect against high-fat diet-induced peripheral neuropathy and mechanical hypersensitivity in male mice. Front Nutr. 2022;9:893357. doi:10.3389/fnut.2022.893357
114. Müller M, Hernández MAG, Goossens GH, et al. Circulating but not faecal short-chain fatty acids are related to insulin sensitivity, lipolysis and GLP-1 concentrations in humans. Sci Rep. 2019;9:12515. doi:10.1038/s41598-019-48762-0
115. Barengolts E. Gut Microbiota, prebiotics, probiotics, and synbiotics in management of obesity and prediabetes: a review. Endocr Pract. 2016;22:1224–1234. doi:10.4158/EP161370.RA
116. Correa-Oliveira R, Fachi JL, Vieira A, et al. Regulation of the immune response by probiotics: a new chapter in clinical and basic research. Scandinavian J Immunol. 2016;83:241–248. doi:10.1111/sji.12411
117. De Cabo R, Mattson MP. Effects of intermittent fasting on health, aging, and disease. New Engl J Med. 2019;381:2541–2551. doi:10.1056/NEJMra1905136
118. Sutton EF, Beyl R, Early KS, et al. Early time-restricted feeding improves insulin sensitivity, blood pressure, and oxidative stress even without weight loss in men with prediabetes. Cell Metab. 2018;27:1212–1221.e3. doi:10.1016/j.cmet.2018.04.010
119. Cienfuegos S, Gabel K, Kala A, et al. Effects of 4- and 6-h time-restricted feeding on weight and cardiometabolic health: a randomized controlled trial in adults with obesity. Cell Metab. 2020;32:366–378.e3. doi:10.1016/j.cmet.2020.06.018
120. Yuan X, Wang J, Yang S, et al. Effect of intermittent fasting diet on glucose and lipid metabolism and insulin resistance in patients with type 2 diabetes: a systematic review and meta-analysis. Nutrients. 2022;14:671. doi:10.3390/nu14030671
121. Wang X, Yang K, Li R, et al. Intermittent fasting versus continuous energy-restricted diet for patients with type 2 diabetes and obesity: a systematic review and meta-analysis. J Diabetes Invest. 2020;11:967–977. doi:10.1111/jdi.13218
122. Mizushima N, Levine B. Autophagy in human diseases. New Engl J Med. 2020;383:1564–1576. doi:10.1056/NEJMc2029918
123. Alirezaei M, Kemball CC, Flynn CT, et al. Short-term fasting induces profound neuronal autophagy. Autophagy. 2010;6:702–710. doi:10.4161/auto.6.6.12376
124. Bagherniya M, Butler AE, Barreto GE, et al. The effect of fasting or calorie restriction on autophagy induction: a review of the literature. Ageing Res Rev. 2018;47:183–197. doi:10.1016/j.arr.2018.08.004
125. Panda S. Circadian physiology of metabolism. Science. 2016;354:1008–1015. doi:10.1126/science.aah4967
126. Chaix A, Zarrinpar A, Panda S. Time-restricted eating to prevent and manage chronic metabolic diseases. Ann Rev Nutr. 2019;39:291–315. doi:10.1146/annurev-nutr-082018-124320
127. Jamshed H, Beyl RA, Della Manna DL, et al. Early time-restricted feeding improves 24-hour glucose levels and affects markers of the circadian clock, autophagy, and inflammation in humans. Nutrients. 2019;11:1234. doi:10.3390/nu11061234
128. Madkour MI, El-Sarha AA, Gad AM. Intermittent fasting, a possible priming tool for regeneration in the injured peripheral nerve. Exp Neurol. 2019;322:113069. doi:10.1016/j.expneurol.2019.113069
129. Mattson MP, Longo VD, Harvie M. Impact of intermittent fasting on health and disease processes. Ageing Res Rev. 2017;39:46–58. doi:10.1016/j.arr.2016.10.005
130. Antoni R, Johnston KL, Collins AL, et al. The effects of intermittent fasting on glucose and lipid metabolism.
131. Rynders CA, Thomas EA, Zaman A, et al. Effectiveness of intermittent fasting and time-restricted feeding compared to continuous energy restriction for weight loss. Nutrients. 2019;11:2442. doi:10.3390/nu11102442
132. Zheng H, Sun W, Zhang Q, et al. Proinflammatory cytokines predict the incidence of diabetic peripheral neuropathy over 5 years in Chinese type 2 diabetes patients: a prospective cohort study. EClinicalMedicine. 2021;31:100649. doi:10.1016/j.eclinm.2020.100649
133. Hinder LM, O’Brien PD, Hayes JM, et al. Dietary reversal of neuropathy in a murine model of prediabetes and the metabolic syndrome. Dis Model Mech. 2017;10:867–877. doi:10.1242/dmm.028530
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.