Back to Journals » Breast Cancer: Targets and Therapy » Volume 18
MLN51 (Metastatic Lymph Node Gene 51)/CASC3 (Cancer Susceptibility Candidate Gene 3), A Putative Tumour Suppressor in Breast Cancer, the Clinical and Therapeutic Connections
Authors Cong B, Martin TA, Cao X, Ruge FS, Li AX, Mansel RE, Wang Y, Jiang WG
Received 6 October 2025
Accepted for publication 5 January 2026
Published 14 January 2026 Volume 2026:18 565199
DOI https://doi.org/10.2147/BCTT.S565199
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Pranela Rameshwar
Binbin Cong,1,2 Tracey A Martin,2 Xiaoshan Cao,1,2 Fiona S Ruge,2 Amber Xinyu Li,2 Robert E Mansel,3 Yongsheng Wang,1 Wen G Jiang2
1Breast Cancer Centre, Shandong Cancer Hospital and Institute, Shandong First Medical University and Shandong Academy of Medical Sciences, Jinan, Shandong, People’s Republic of China; 2Cardiff China Medical Research Collaborative, Division of Cancer & Genetics, Cardiff University School of Medicine, Cardiff, UK; 3Wales Breast Centre, University Llandough Hospital, Cardiff, UK
Correspondence: Wen G Jiang, Email [email protected]
Introduction: The Metastatic Lymph Node Gene 51 Protein (MLN51), also known as the Cancer Susceptibility Candidate Gene 3 Protein, was first identified in malignant breast tumours. It has been shown to be a key component of the exon junction complex. This study aimed to evaluate the prognostic value of MLN51 expression in breast cancer.
Methods: MLN51 expression was evaluated in a Cardiff clinical breast cancer cohort (n = 127) and in The Cancer Genome Atlas breast cancer database. The MLN51 protein was detected by immunohistochemistry. Two groups of breast cancer cell line models were created, namely MLN51 expression activation by transcriptional activation in MLN51 negative cells, and MLN51 knockdown by shRNA in MLN51 positive cells. Cell models were used to study the effect of the expression of the MLN51 gene in their behavior.
Results: High expression of the MLN51 transcript was significantly correlated with favourable overall survival and disease free survival (p = 0.004 and p = 0.001, respectively). Patients who responded to chemotherapy had significantly higher MLN51 levels than those who resisted treatment (p = 0.00054), and this connection was more profound in ERBB2 negative type. Following activation of MLN51 in MDA MB-231cells, there was a significant decrease in the growth rate. In contrast, the knockdown of MLN51 in SKBR3 cells significantly increased cell growth. High expression of MLN51 following transcriptional activation increases the sensitivity to chemotherapy and anti-ERBB2 target therapy.
Conclusion: MLN51 expression is an independent predictor of disease prognosis, patient survival, and treatment response in breast cancer patients.
Keywords: breast cancer, MLN51, CASC3, disease prognosis, therapy response
Introduction
Breast cancer is the most common malignant tumour in female around the world has become the leading cancer in the population in 2020, seriously threatening the health of women.1,2 In China, breast cancer represents a major and growing public-health challenge.2 Due to the characteristics of invasion and migration of cancer cells, a part of early stage breast cancer even receives completed treatments and finally undergoes distant migration, which transforms to an advanced stage.2 The cause of death from breast cancer is distant metastasis, such as in the brain, lung, liver, and bone, which can lead to multiple organ failure.3 The common route of metastasis is lymphatic, which leads to lymph node involvement and a poor prognosis.4 The current challenges include the detection and treatment of involvement at the first opportunity in breast cancer, which could effectively reduce the risk of distant metastasis.
Metastatic Lymph Node Gene 51 Protein (MLN51), also known as Cancer Susceptibility Candidate Gene 3 Protein (CASC3) or Barentsz (BTZ), was first identified in a complementary deoxyribonucleic acid (cDNA) library of malignant breast tumours.5–7 Using differential screening of cDNA libraries from metastatic lesions of breast cancer and normal mammary tissues, a number of new gene transcripts were identified, including MLN51. MLN51 was found in metastatic lymph node tissues, but not in normal mammary or fatty tissues. The coding region for MLN51 was assigned to chromosome 17 q12 – q21.3, which harbours other potential oncogenes including ERBB2, MLN50, and MLN64. In some studies, MLN51 was found to be amplified in HER2 positive/ER-negative SKBR3 breast cancer cells but not in other breast cancer cell lines tested.6 Subsequently, MLN51 protein was found in SKBR3, MCF-7, MDA MB-453, the mammary epithelial cell line HBL100, and cells of other tumour origins, including ovarian cancer (SK-OV-3), leukaemia Jurkat cells, and approximately 30% of breast tumours in a small cohort study, unrelated to tumour grade and histological type (n = 50).5 In suspected co-amplification of MLN51 and HER2 in breast cancer, it was found that in HER2 positive breast cancer cells, overexpressed MLN51 (HER2-positive/MLN51-positive) appears to result in a reduction of P-bodies compared to HER2-positive/MLN51-negative breast cancers and normal mammary tissues.8 Although the exact roles of P-bodies and stress granules in cancer are yet to be fully established, they are clearly related to cancer and cancer development.9 In contrast, in a larger The Cancer Genome Atlas (TCGA) cohort, MLN51 was found to be a favourable prognostic indicator in patients with breast cancer.10
It has been reported that MLN51 is one of the core components of the EJC (core components of the exon junction complex, an important structure formed around the nuclear speckles for post-splicing events of mRNA and a translation enhancer11) and also includes other proteins such as eIF4AIII, Y14, and Magoh.12–14 Beyond the EJC, MLN51 also plays a distinct role in the assembly of cytoplasmic aggregates known as stress granules (SG) in various cell types, including breast cancer cells,15 and is an essential component of P-bodies, which are cytoplasmic granules involved in post-transcriptional regulation of mRNA and mRNA decay.8 However, it has been recognised that MLN51 is a very different member of the EJC complex and can activate mRNA isoforms targeted by nonsense-mediated decay, possibly by stimulating mRNA decay and endonucleolytic cleavage at the termination codon.16
In addition to breast cancer, studies on cancer are also difficult to perform. In gastric cancer, MLN51 is one of the few genes on chromosome 17 that has an increased copy number.17 However, if the loci harbouring the MLN51 coding gene (17q21.1~q21.2) are lost, patients have a significantly worse survival,18 tentatively arguing for the tumour suppressive role of MLN51. Interestingly, in murine gastrointestinal models, combination therapy with gemcitabine and anti-PDL1 antibody was able to eradicate subpopulations of cancer cells with MLN51 frameshift mutations.19
In non-cancer-related cells, MLN51 was found to be key to the granulocyte-macrophage colony-stimulating factor (GM-CSF)-induced proliferation of synoviocytes associated with rheumatoid arthritis.20 The brain-specific miRNA miR-128 targets MLN51 in neuronal cells that represses nonsense-mediated decay (NMD).21 It is also a highly useful marker for bone marrow stromal cells (BMSCs)22 and ascending aorta.23 miR-124-1 was also shown to target MLN51 in hepatocellular carcinoma.24 MLN51 is involved in viral entry into cells and can markedly reduce viral accumulation. This is interestingly independent of the NMD pathways as it was proposed.25 The study also identified MLN51 condensates in the cytoplasmic region and that the MLN51 condensates do not have a p-body or stress granule structure, further implicating that the MLN51 protein may act differently from its associated EJC function. It has also been recently shown that MLN51 is required for embryogenesis but is not essential for brain development in mice, suggesting that MLN51 may act in a manner depending on the tissue type.26
MLN51 therefore appears to be an important regulator of the mRNA metabolism. However, it is unclear whether MLN51 has clinical value in assessing the progression of breast cancer and, indeed, in most cancer types. Limited information regarding the role of MLN51 in breast cancer remains controversial. Here, we explored the expression of MLN51 in a clinical cohort in comparison with that in TCGA database. We also created breast cancer cell models by either transcriptionally activating MLN51 expression in MLN51 negative breast cancer cells or by knocking down MLN51 expression in MLN51 positive breast cancer cells. These models were subjected to biological and pharmaceutical tests to determine the effect of MLN51 on growth, cellular migration, and cell response to chemotherapeutic drugs. Here, we report that MLN51 is a favourable prognostic factor for patients with breast cancer and is a favourable indicator of patient response to chemotherapy. Our cell models showed that high levels of MLN51 (by way of transcriptional activation) exhibited slower growth and were more sensitive to chemotherapy drugs. Conversely, low levels of MLN51 following knockdown were associated with a faster cell growth rate and increased resistance to chemotherapy drugs.
Materials and Methods
Materials and Cell Lines
Human breast cancer cell lines SKBR3, MDA468, MDA-MB-231, T47D, MCF7, ZR751, BT474, HCC1419, MRC5, HECV, and MAD361 were obtained from the European Collection for Authenticated Cell Culture (ECACC, Porton Down, Salisbury, UK). The cell lines of MDA468, MAD361, T47D, MCF7, ZR751, MRC5, HECV, and MDA-MB-231 were cultured in Dulbecco’s modified Eagle’s medium (DMEM)-F12 medium, and SKBR3, BT474, and HCC1419 were cultured in Roswell Park Memorial Institute (RPMI) 1640 medium in a 5% CO2 incubator at 37°C. Both media were supplemented with 10% heat-inactivated, non-USA origin, sterile-filtered foetal calf serum (FCS, F9665, Sigma-Aldrich, Poole, Dorset, UK) and 1% stabilised antibiotic antimycotic solution (100×), including 10,000 units penicillin, 10 mg streptomycin and 25ug amphotericin B per mL (A5955, Sigma-Aldrich, Pooled, Dorset, UK).
Breast Cancer Clinical Cohort Data (Cardiff University)
Breast cancer tissues (n = 127) and normal breast tissues (n = 34) were collected and processed after the operation. Postoperative histology and molecular pathology results were collected from the medical records and verified by a consultant pathologist. All patients agreed to tissue and clinical data collection procedures. The study adhered to the principles outlined in the Declaration of Helsinki, including all subsequent amendments, and received approval after ensuring that all tissues and clinical data were utilized with informed patient consent. The research protocol underwent thorough review and was subsequently approved by the Bro Taf Health Authority Local Research Ethics Committee (Reference No.: 01/4303). Informed consent was secured from each individual participant. However, the necessity for obtaining informed consent was waived in this instance, given the retrospective nature of the study and the utilization of anonymized patient data. The median postoperative follow-up period was 120 months. MLN51 expression in tumour and normal tissues was determined by real-time quantitative polymerase chain reaction (QPCR).27
The Cancer Genome Atlas (TCGA) Breast Cancer RNA Sequencing Data
The relationship between MLN51 expression and survival in breast cancer was analysed using TCGA database (https://www.cancer.gov/tcga). In addition, the association between MLN51 expression and the response of patients to chemotherapy was analysed using this database. The responders were complete response (CR) and partial response (PR) based on the response evaluation criteria in solid tumors (RECIST1.1). Non-responders were stable disease (SD) and progressive disease (PD). The RNA sequencing dataset comprised 493 lymph node-negative and 587 lymph node-positive cases in a cohort of primary breast cancers.
Establishment of MLN51 Knockdown Cell Model
The human MLN51-specific shRNA plasmid (sc-93943) was obtained from Santa Cruz Biotechnology (Santa Cruz, TX, USA). SKBR3 and MDA468 breast cancer cells were transfected with MLN51-specific shRNA plasmids in T25 flasks for 24 hours. After transfection and validation, cells were subjected to selection with 1 μg/mL puromycin (Fisher Scientific, Oxford, UK) and, once tested for the success of genetic modification, were routinely maintained in maintenance medium (with 0.1 μg/mL puromycin).
Establishment of MLN51 Over-Expression Cell Model by Way of Transcription Activation
A human MLN51 CRISPR transcription activation system, which comprised three plasmids, each carrying a different selection marker, allowing the transcriptional activation of MLN51 (sc-411606), was also purchased from Santa Cruz Biotechnology (Santa Cruz, TX, USA). MDA MB-231 and MDA MB-361 breast cancer cells were transfected with the human MLN51 CRISPR activation plasmids in T25 flasks for 24 h. After transfection and validation, the cells were subjected to selection using triple antibiotics, including 2 μg/mL puromycin (Fisher Scientific, Oxford, UK), 200 µg/mL Hygromycin B (Fisher Scientific, Oxford, UK), and 1 µg/mL Blasticidin S HCl (Fisher Scientific, Oxford, UK). Once tested for the success of genetic modification, they were routinely maintained in maintenance medium (with 0.1 μg/mL puromycin, 100 µg/mL Hygromycin B, 0.05 µg/mL Blasticidin S HCl).
Isolation of Total RNA and Reverse Transcription
Total RNA was isolated separately from breast cancer cells and frozen tissues using TRI reagent (Sigma-Aldrich) and one-step reverse transcribed into cDNA using the Promega Reverse Transcription kit (Promega, Dorset, UK), according to the manufacturer’s instructions. PCR was performed in a SimpliAmp Thermal Cycler (Applied Biosystems, Paisley, UK) using GoTaq Green Master Mix (Promega, Dorset, UK) in a final volume of 12 μL. GAPDH was used as an endogenous control for each sample and the PCR products were visualised on 1.5% agarose gels stained with SYBR Safe (Invitrogen).28
Immunohistochemical Staining
Immunohistochemical staining was performed using breast cancer tissue microarray (TMA). The primary antibody used was an anti-MLN51 mouse monoclonal antibody (1:50 dilution; sc-376186; Santa Cruz Biotechnology, UK). The secondary antibody solution consisted of 100 μL biotinylated antibody stock at a 5 mL dilution (Vectastain Universal Elite ABC Kit, PK-6200, Vector Laboratories, Peterborough, UK). The presence of cancer cells was assessed by determining the intensity and distribution of the MLN51 staining. The intensity of MLN51 staining was determined using ImageJ software by one researcher at the host laboratory. Staining intensity was categorised into four levels: 0, negative; 1, weak; 2, moderate; and 3, strong.28
Quantitative Gene Transcript Analysis
RNA extraction from breast samples was performed using AB Gene Total RNA Isolation Reagent (Advanced Biotechnologies Ltd., Altrincham, Cheshire, UK). RNA was isolated from a human breast cancer cell line RNA was a TRI Reagent Kit (Sigma-Aldrich, Poole, Dorset, UK). The concentration and quality of RNA were determined by spectrophotometric measurements (WPA UV 1101; Biotech Photometer, Cambridge, UK). Complementary DNA (cDNA) was generated from 0.25 μg of each RNA sample and reverse transcribed using a transcription kit (Sigma, Poole, Dorset, UK). DNA quality was verified using β-actin primers. An iCycler IQ system (Bio-Rad, Irvine, CA, USA) was used to quantify the level (shown as copies/μL from the internal standard) of MLN51 in all samples. Breast cDNA samples were then examined for expression levels along with a set of standards and negative controls. Quantitative PCR was performed using the Amplifluor System (InterGen, Inc., Oxford, UK) and QPCR Master Mix (Abgene, Loughborough, Leicestershire, UK), in conjunction with a universal probe (UniPrimer). The conditions for real-time quantitative PCR were 95°C for 15 min, followed by 65 cycles at 95°C for 15 s, 55°C for 30 s, and 72°C for 20 s. The epithelial content within the tumours was considered to be cytokeratin 19. These methods are described in our previous study.29
Protein Preparation and Western Blot
After the cancer cells reached sufficient confluence, the medium in the flask was removed and the cell monolayer was washed twice with ice-cold PBS. Next, 5 mL PBS was added to the flask, and the cell monolayer was scraped with a sterile cell scraper from the flask surface and transferred to a universal tube. The cell suspension was centrifuged at 1800rpm for 10 min to obtain a cell pellet. The supernatant was removed, and 100 μL RIPA buffer containing protease and phosphatase inhibitors was added and suspended in the tube. Then, the cell suspension was transferred into a 1.5 mL microfuge tube and incubated at 4°C for 1 h with continuous rotation on 25rpm Labinoco rotating wheel (Wolf Laboratories, York, UK). After incubation, the cell lysate was centrifuged at 13000rpm for 15 minutes. The supernatant was transferred into a fresh tube and stored at −20°C until further use.
Protein concentration was quantified using the Bio-Rad DC protein assay kit (Bio-Rad Laboratories, Hemel-Hempstead, UK). Thirty micrograms of each protein sample were transferred onto a 0.45 μm polyvinylidenedifuoride (PVDF) membrane (Millipore, Billerica, MA) after separation by 10 or 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). The membrane was blocked with 5% non-fat milk for 1 h at room temperature, then incubated with primary antibody (1:500) at 4°C overnight and secondary antibody (1:2000) for 1 h. The primary antibodies used were MLN51 (sc-376186, Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA), cleaved PARP (detecting both full-length and cleaved PARP) (eBioscience Inc., San Diego, CA, USA), p21WAF1, p53 (DO-1), and GAPDH (Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA). The results are representative of three independent experiments.28
Cell Growth Assay
Each well were seeded about 2,000 cells in 200 μL medium and the cell plates were incubated for 1, 3, and 5 days. Following incubation, cells were fixed with formalin and stained with crystal violet. The absorbance was measured after dissolving the crystal violet in 10% acetic acid at a wavelength of 540 nm using a spectrophotometre (Elx800; Bio-Tek, Bedford, UK). The growth rate was calculated using the following formula: growth rate (%) = absorbance (day 3 or day 5)/absorbance (Day1) × 100.28
Electric Cell-Substrate Impedance Sensing (ECIS) Based Cell Migration Assay
Cell solutions containing 50,000 cells with the relevant culture medium for each cell model were seeded at a final volume of 200 µL for each well into 96-well ECIS W961E electrode arrays in six replicates. The arrays were then placed on an ECIS ZTheta instrument (Applied Biophysics, Troy, New Jersey, USA). After incubating for 4 h to reach confluent monolayers, an electrical wound (2000 mA for 20 s) was created in each well using ECIS, and the impedance was measured immediately. The impedance level of each well was recorded continuously from 1000 to 64,000 Hz using the ECIS system for the next 24 h. Data were analysed using the ECIS software.28
Cytotoxicity Assays
Cell cytotoxicity assays were carried out to evaluate the effect of MLN51 on drug resistance using cell models with differentially expressed MLN51. Three thousand cells per well were seeded into 96 well plates. Serially diluted drugs were added to the cells and the plates were incubated at 37°C with 5% CO2 for 72 h. After incubation, each well was added equal amount of MTT (5 mg/mL) (Sigma-Aldrich Co., Poole, Dorset, UK) and incubated for a further 4 hour at 37°C with 5% CO2. Following the removal of the medium, 100 μL of dimethyl sulfoxide (DMSO) (Sigma-Aldrich Co., Poole, Dorset, UK) was added to each well, followed by mixing for 15 min to dissolve the formazan crystals. The plates were read using a 96-well plate reader (LT4500; Wolf Laboratories, York, UK) at 540 nm. Statistical analyses were performed using Microsoft Excel and GraphPad software.
Statistical Analysis
All statistical analyses were performed using the SPSS software (version 26, SPSS, Chicago, IL, USA) and GraphPad Prism 6.0 (GraphPad Software, La Jolla, CA, USA). The Mann–Whitney test was applied for non-normally distributed data, the t-test was used for normally distributed data, and Pearson’s chi-squared test was applied to analyse nominal data. Correlations between genes were analysed using the Spearman’s test. Statistical significance was set at P < 0.05. Survival analysis was conducted using Kaplan–Meier plots.
Results
Expression of MLN51 in Breast Cancer and Its Implications in Disease Progression and Prognosis
The QPCR results of the Cardiff clinical cohort showed that the expression levels of MLN51 were not significantly different between breast cancer and normal tissues (p = 0.4655). The results of the MLN5 expression analysis in the cardiac clinical breast cancer cohort are shown in Table 1.
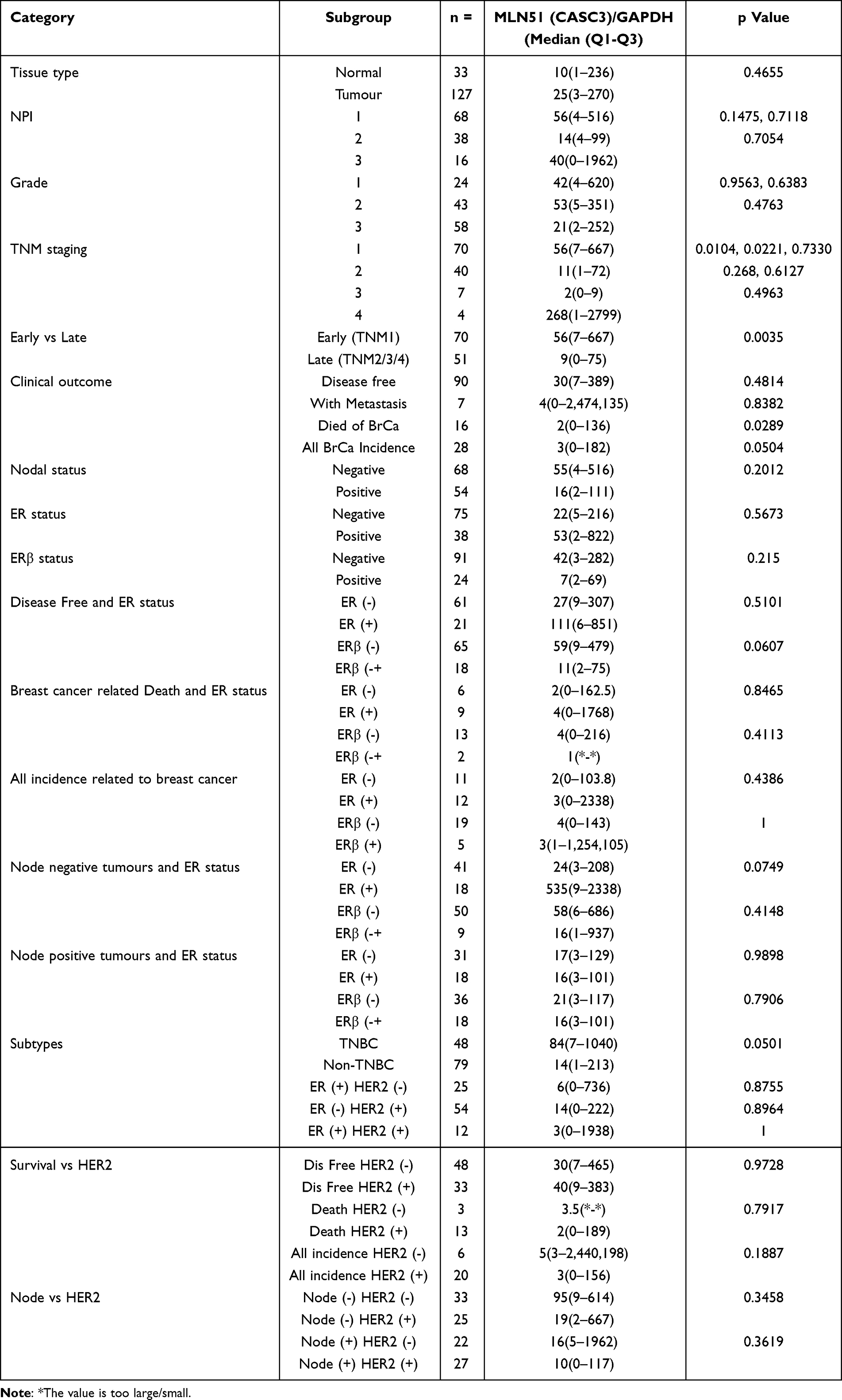 |
Table 1 MLN51 Gene Expression Analyzed in Cardiff Clinical Breast Cancer Cohort |
MLN51 and Clinical Outcomes
In the Cardiff clinical breast cancer database, high expression levels of MLN51 were a significant and independent predictive factor for overall survival ([OS] p = 0.004; Hazard Ratio [HR], 0.262]) and recurrence-free survival (RFS) (p = 0.001, HR = 0.264) (Figure 1). A favourable prediction of OS was significantly associated with lymph node-positive tumours (p = 0.004, HR = 0.194) compared to lymph node-negative tumours (p = 0.143, HR = 0.233). Positive ER expression was significantly associated with OS (p = 0.007) compared to ER-negative tumours (p = 0.2), and HER2 (+) was significantly associated with OS (p = 0.007) compared to HER2 (-) tumours (p = 0.655). A significant association with OS was also observed in TNBC (p = 0.002) and ER (−) HER2 (+) (p = 0.002) tumours (Figure 1).
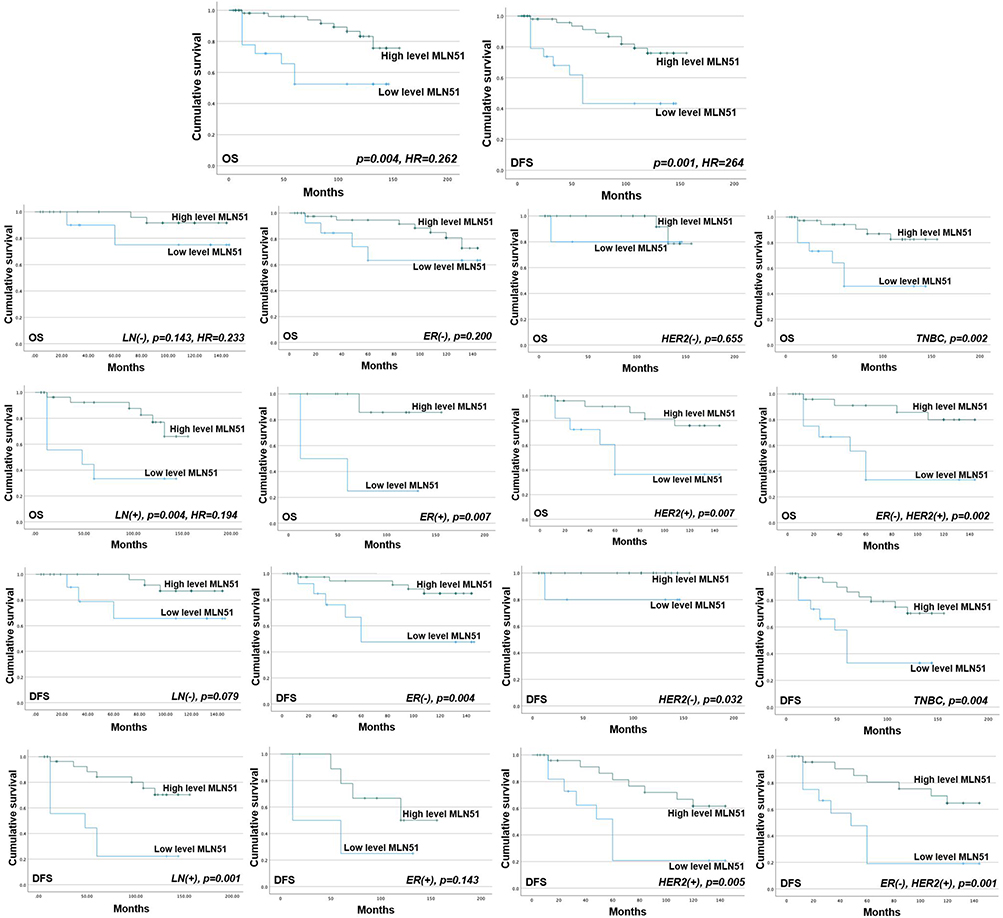 |
Figure 1 MLN51 transcript expression and the survival of the patients in Cardiff clinical breast cancer database. |
The favourable prediction of RFS was associated with lymph node-positive tumours (p = 0.001) compared to lymph node-negative tumours (p = 0.079) (Figure 1). Negative ER expression was significantly associated with RFS (p = 0.004) compared to ER-positive tumours (p = 0.143), and HER2 (+) was significantly associated with RFS (p = 0.005) compared to HER2 (-) tumours (p = 0.032). A significant association with RFS was also observed in TNBC (p = 0.004) and ER (−) HER2 (+) (p = 0.001) tumours.
In TCGA breast cancer database, high expression levels of MLN51 were also significantly correlated with favourable OS (p = 0.0011, HR = 0.68) and favourable RFS (p < 0.0001, HR = 0.74) (Figure 2). HER2 (-) was significantly associated with OS (p = 0.0057) compared with HER2 (+) tumours (p = 0.32). Furthermore, both positive and negative ER expression were significantly associated with RFS (p = 0.0066 and p = 0.0063, respectively), and HER2 (+) and HER2 (−) were significantly associated with RFS (p = 0.00086 and p < 0.001, respectively). A significant association with RFS was also observed in patients with TNBC (P = 0.0062) (Figure 3). The prognostic value of MLN51 was independent of the other clinical and pathological factors (Table 2).
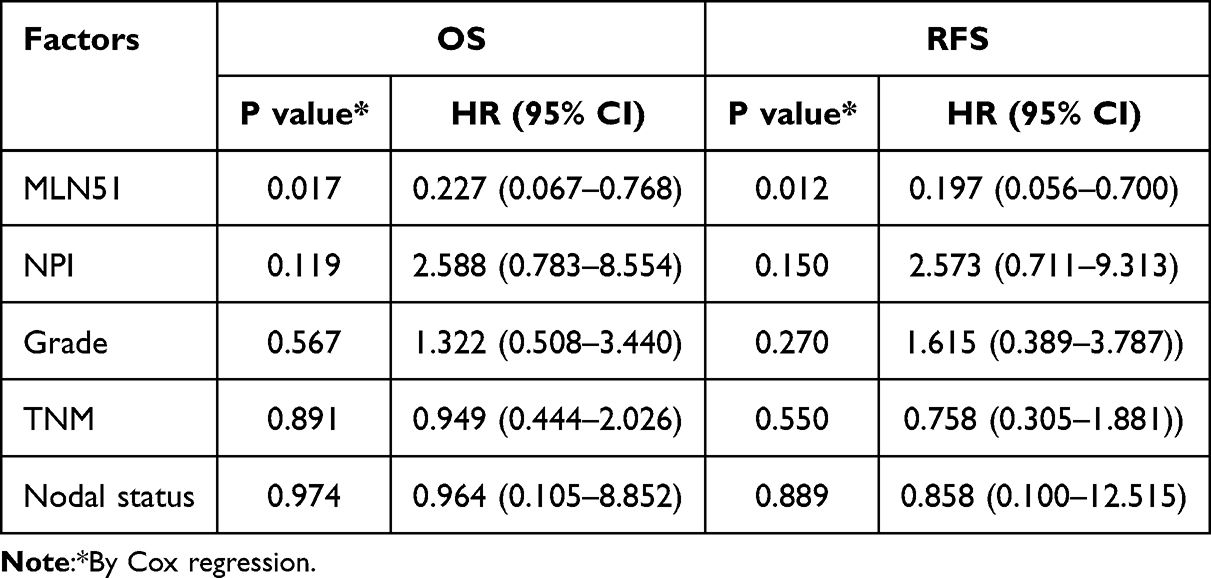 |
Table 2 Results of Multivariant Analysis with MLN51 and Clinical Factors by Cox Regression |
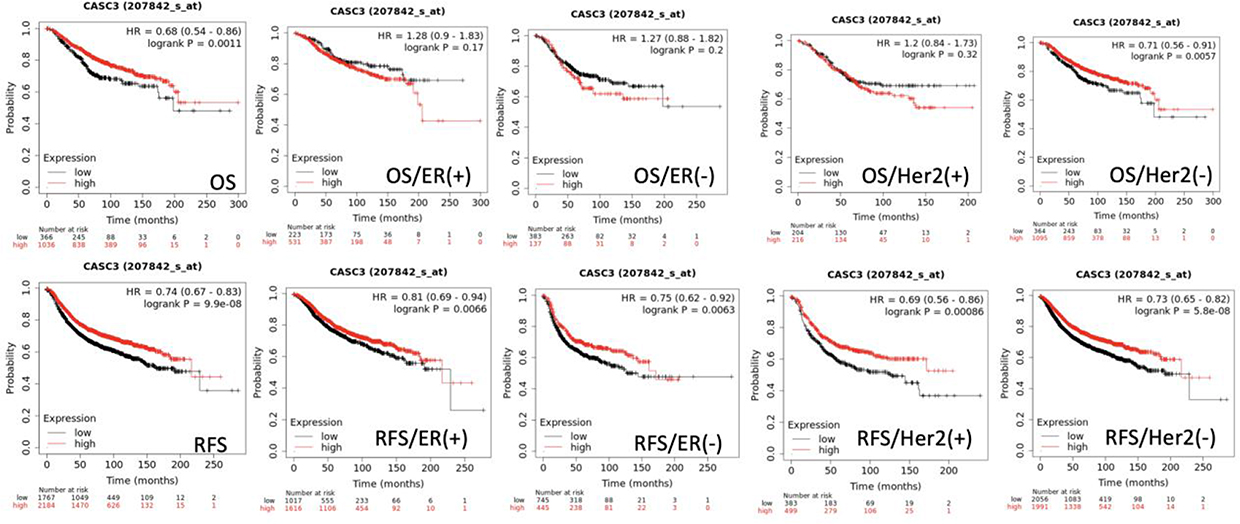 |
Figure 2 Survival analysis results of MLN51 expression levels and clinical association factors in TCGA (genechip dataset). |
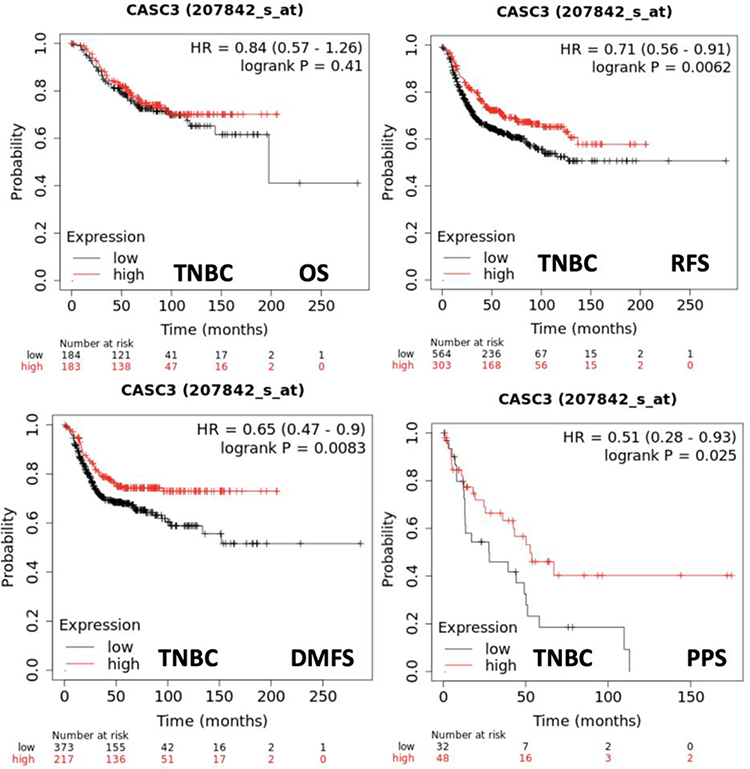 |
Figure 3 Correlation between MLN51 expression and survival probabilities in TNBC subtypes from TCGA (genechip dataset). |
MLN51 and Drug Responses of Patients with Breast Cancer
In TCGA breast cancer database, MLN51 expression levels were significantly higher in responders than in non-responders (p = 0.00054). High MLN51 expression was more sensitive to drug treatment than MLN51 low expression in the analysis of five-year RFS (AUC = 0.592, p = 0.00021, Figure 4). Subgroup analysis showed MLN51 high expression was associated with a better pathological response, regardless of ER (-, p = 0.021), ER (+, p = 0.00059), and HER2 (-, 0.0048) (Table 3). Furthermore, TNBC, luminal-A, and luminal-B were also found MLN51 high expression have better pathological responses than those with low expression (p = 0.0032, 0.046, and 0.014, respectively). In patients with ER (+), HER2 (-), and luminal-B subtypes, high MLN51 expression was significantly associated with 5-year RFS (p = 0.00075, 0.0015, and 0.0028, respectively). The details of MLN51 expression and drug responses are showed in Table 3.
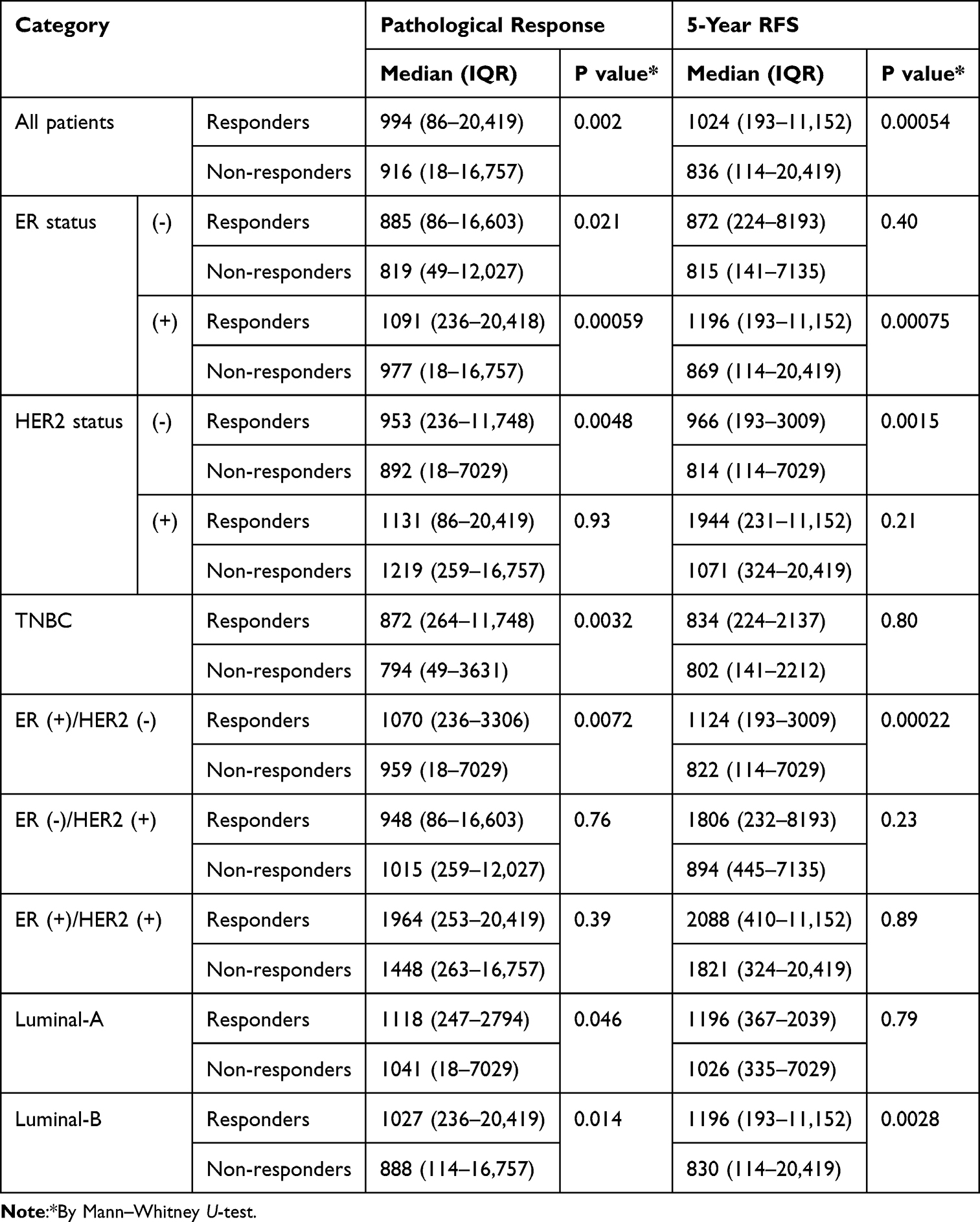 |
Table 3 MLN51 Expression and Patient’s Responses to Chemotherapies in TCGA Breast Cancer Database |
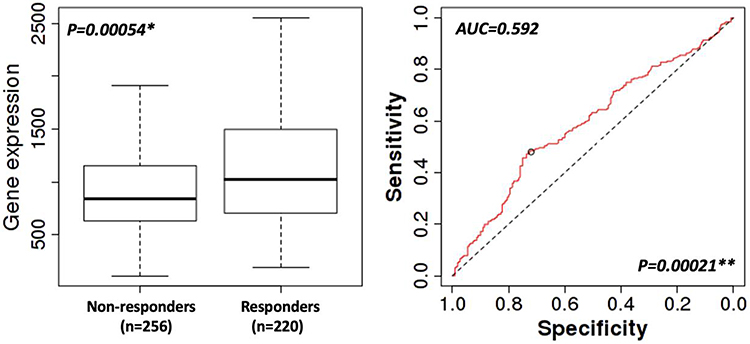 |
Figure 4 Five-year RFS response to chemotherapies. * by Mann–Whitney U-test; ** by ROC model. |
MLN51 Expression in Human Breast Cancer Cell Lines and Creation of Cell Models with Differential Expression of MLN51
Positive expression of MLN51 was observed in the SKBR3, MDA468, BT474, ZR751, and MCF7 breast cancer cell lines. Low expression of MLN51 was observed in the MDA231, MDA361, and HCC1419 cell lines (Figure 5).
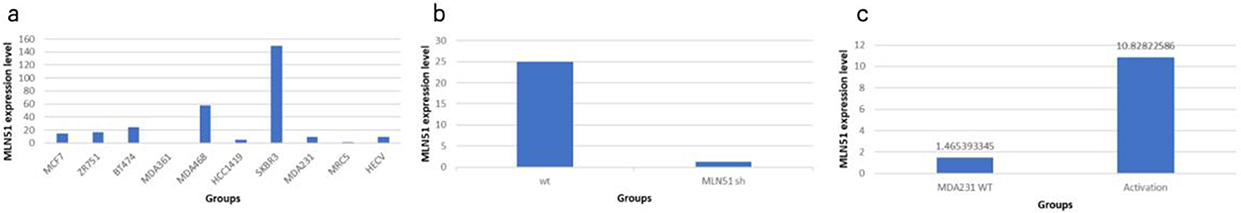 |
Figure 5 MLN51 expression in different breast cancer cell lines (a) and validation of SKBR3 MLN51 knockdown (b) MDA231 MLN51 activation (c). |
SKBR3, MDA468, and MCF7 breast cancer cells were chosen to knock down MLN51 using shRNA, and MDA231, MDA361, and MCF7 were chosen to express MLN51 via transcriptional activation. The validation study showed that the MLN51 knockdown rate was 94.9% in SKBR3 cells and the activation rate of MLN51 was 86.5% in MDA231 (Figure 5).
MLN51 Expression and Cell Growth Rate
In the functional test, the growth rate of MDA MB-231 cells was decreased after MLN51 activation. In contrast, the rate of SKBR3 expression increased after MLN51 knockdown (Figure 6).
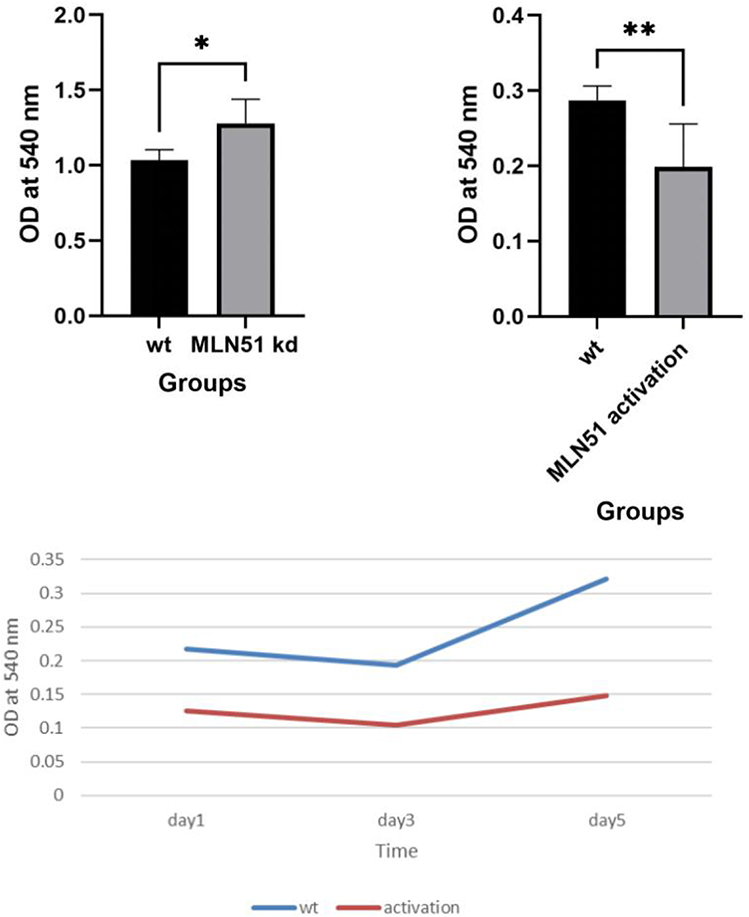 |
Figure 6 Proliferation test of SKBR3 MLN51 knockdown, MDA231 MLN51 activation. * p < 0.05, ** p < 0.01. |
MLN51 Expression and Cell Response to Chemotherapy Drugs
In the cell toxicity test, the viability of MDA231 MLN51 cells significantly decreased after treatment with docetaxel (DOC), methotrexate (MTX), and gemcitabine (GEM) (Figure 7). When combined with a broad-spectrum HER2 inhibitor drug, neratinib, the cytotoxicity of DOC and GEM was significantly increased in MLN51 activated MDA MB-231 cells compared with that without the HER2 inhibitor. Thus, high expression of MLN51 could increase sensitivity to chemotherapy ± target therapy and improve the response to therapy.
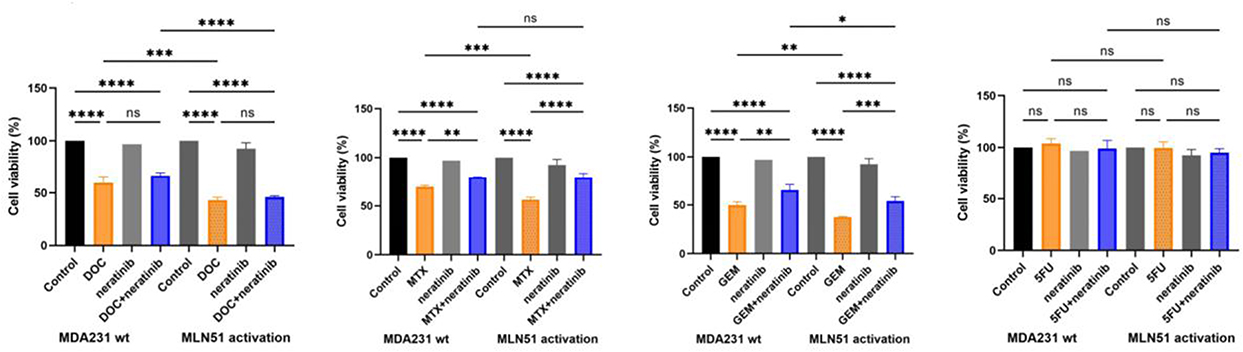 |
Figure 7 Toxicity test of MDA231 wt and MLN51 activation with DOC, MTX, GEM, 5FU ± neratinib.* p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, ns = non-significant. |
Discussion
MLN51 is a tumour suppressor gene that plays an important role in the modification of cancer cell growth. MLN51 was first detected in the metastatic lymph node.5 However, we found that the expression of MLN51 was no different between normal breast tissue and breast cancer tissue (p = 0.4655) in the Cardiff clinical cohort data. The expression level of MLN51 was significantly higher in early stage breast cancer than in late-stage breast cancer (p = 0.0035) in the clinical breast cancer cohort. Analysis of clinical survival results showed that MLN51 was associated with patient prognosis. Low MLN51 expression could be considered a predictor of breast cancer death (p = 0.0289) in the clinical outcome. More important, MLN51 high expression means good survival and low expression indicates poor survival in patients with lymph node metastasis. Furthermore, the predicted vale of MLN51 high for better survival was observed for TNBC and ER (-) HER2 (+) breast cancer subtypes.
We also explored the clinical prognostic value of MLN51 in TCGA breast cancer database. MLN51 expression was found to be an independent factor for survival prognosis compared to NPI, issue grade, TNM stage, and nodal status by COX regression. High expression of MLN51, indicating better OS, was observed in the HER2 (-) subgroup which differed from the analysis results of the Cardiff clinical breast cancer cohort data. The survival prognosis of HER2-positive breast cancer is affected by anti-HER2 targeted therapy, and trastuzumab reduces the risk of death from early stage breast cancer by 33%. The difference in treatments between the two groups may have led to these differences. However, high expression MLN51 meaning better RFS was found in all subgroups, which is consistent with our clinical database. The analysis results from TCGA database and the clinical case database provide an important reference for us to carry out basic research.
We designed and prepared cell models to examine the molecular, cellular, and therapeutic responses to different levels of MLN51 expression. The results of the cell function test showed that upregulated expression level of MLN51 inhibited the growth rate of MDA MB-231 cells which showed MLN51 low expression but downregulated expression of MLN51 promoted the proliferation of SKBR3 cells which showed MLN51 high expression. These results further confirm that MLN51 is a tumour suppressor gene that is important for the modification of cancer cell growth. Thus, MLN51 may serve as a reference factor for breast cancer prognosis, providing crucial clues regarding recurrence and metastasis in clinical practice. This will help clinicians to make more accurate judgments about the high-risk conditions of patients with MLN51 different expression level.
In the cell toxicity test, we found that the status of MLN51 could influence its therapeutic effect in different cell lines, especially in triple-negative and HER2 positive breast cancer cell lines. The viability of MDA231 MLN51 cells significantly decreased after treatment with cytotoxic drugs. When combined with a broad-spectrum HER2 inhibitor drug, neratinib, the cytotoxicity of DOC and GEM was significantly increased in MLN51 activated MDA MB-231 cells compared with that without the HER2 inhibitor. Thus, high expression of MLN51 could increase sensitivity to chemotherapy ± target therapy and improve the response to therapy. The significance of this study lies in its potential of MLN51 to provide effective biological predictive biomarkers for evaluating the efficacy of neoadjuvant therapy for breast cancer.
Neoadjuvant therapy has been widely used in patients with early stage breast cancer and can provide better drug sensitivity evaluation for this group of patients and offer downstaging opportunities for breast and axillary preservation in some patients.30 Although neoadjuvant therapy achieves a higher pathological complete response (pCR) rate in triple-negative and HER2-positive breast cancers, a subset of patients develops resistance leading to disease progression, indicating suboptimal efficacy of neoadjuvant therapy for these individuals. However, biological markers that can effectively predict neoadjuvant therapy are lacking. NFIX inhibits the proliferation of breast cancer cells, but its significance in cell cycle regulation by cyclin-dependent kinases has not been identified, especially in neoadjuvant therapy.31 Currently, MLN51 co-amplifies HER2 in breast cancer cells, leading to increased HER2 protein expression and the subsequent activation of downstream signalling pathways.
Conclusions
The expression level of MLN51 has been established as an independent prognostic factor that influences disease outcome, patient survival, and therapeutic response in breast cancer. Higher levels of MLN51 expression were correlated with better disease prognosis and increased sensitivity to treatment compared to lower expression levels. The expression level of MLN51 might be an essential testing item in clinical samples to assess prognosis and guide treatment. Future research should focus on validating the predictive role of MLN51 in the context of neoadjuvant therapy for breast cancer.
Acknowledgment
Binbin Cong received grants from the Collaborative Academic Innovation Project of Shandong Cancer Hospital (GF003) for his research on breast cancer at the Cardiff University School of Medicine. The abstract of this paper was presented at the 2023 San Antonio Breast Cancer Symposium Conference name Clinical and prognostic value of MLN51 (Metastatic lymph node gene 51)/CASC3 (Cancer susceptibility candidate gene 3) in clinical breast cancer, connection with nodal and hormonal receptor status as a poster presentation with interim findings. The poster’s abstract was published in “Poster Abstracts” in Cancer Research Journal name Abstract PO5-04-10: Clinical and prognostic value of MLN51 (Metastatic lymph node gene 51)/CASC3 (Cancer susceptibility candidate gene 3) in clinical breast cancer, connection with nodal and hormonal receptor status: Hyperlink with DOI: https://doi.org/10.1158/1538-7445.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Sung H, Ferlay J, Siegel RL, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021;71(3):209–17. doi:10.3322/caac.21660
2. Siegel RL, Miller KD, Fuchs HE, et al. Cancer statistics, 2022. CA Cancer J Clin. 2022;72(1):7–33. doi:10.3322/caac.21708
3. Smolarz B, Nowak AZ, Romanowicz H. Breast cancer-epidemiology, classification, pathogenesis and treatment (Review of literature). Cancers. 2022;14(10):2569. doi:10.3390/cancers14102569
4. DeSantis C, Ma J, Bryan L, et al. Breast cancer statistics, 2013. CA Cancer J Clin. 2014;64(1):52–62. doi:10.3322/caac.21203
5. Degot S, Régnier CH, Wendling C, et al. Metastatic lymph node 51, a novel nucleo-cytoplasmic protein overexpressed in breast cancer. Oncogene. 2002;21(28):4422–4434. doi:10.1038/sj.onc.1205611
6. Tomasetto C, Régnier C, Moog-Lutz C, et al. Identification of four novel human genes amplified and overexpressed in breast carcinoma and localized to the q11-q21.3 region of chromosome 17. Genomics. 1995;28(3):367–376. doi:10.1006/geno.1995.1163
7. Macchi P, Kroening S, Palacios IM, et al. Barentsz, a new component of the Staufen-containing ribonucleoprotein particles in mammalian cells, interacts with Staufen in an RNA-dependent manner. J Neurosci. 2003;23(13):5778–5788. doi:10.1523/JNEUROSCI.23-13-05778.2003
8. Cougot N, Daguenet E, Baguet A, et al. Overexpression of MLN51 triggers P-body disassembly and formation of a new type of RNA granules. J Cell Sci. 2014;127(Pt 21):4692–4701. doi:10.1242/jcs.154500
9. Anderson P, Kedersha N, Ivanov P. Stress granules, P-bodies and cancer. Biochim Biophys Acta. 2015;1849(7):861–870. doi:10.1016/j.bbagrm.2014.11.009
10. Eini M, Parsi S, Barati M, et al. Bioinformatic investigation of micro RNA-802 target genes, protein networks, and its potential prognostic value in breast cancer. Avicenna J Med Biotechnol. 2022;14(2):154–164. doi:10.18502/ajmb.v14i2.8882
11. Daguenet E, Baguet A, Degot S, et al. Perispeckles are major assembly sites for the exon junction core complex. Mol Biol Cell. 2012;23(9):1765–1782. doi:10.1091/mbc.e12-01-0040
12. Degot S, Le Hir H, Alpy F, et al. Association of the breast cancer protein MLN51 with the exon junction complex via its speckle localizer and RNA binding module. J Biol Chem. 2004;279(32):33702–33715. doi:10.1074/jbc.M402754200
13. Ballut L, Marchadier B, Baguet A, et al. The exon junction core complex is locked onto RNA by inhibition of eIF4AIII ATPase activity. Nat Struct Mol Biol. 2005;12(10):861–869. doi:10.1038/nsmb990
14. Chazal PE, Daguenet E, Wendling C, et al. EJC core component MLN51 interacts with eIF3 and activates translation. Proc Natl Acad Sci U S A. 2013;110(15):5903–5908. doi:10.1073/pnas.1218732110
15. Baguet A, Degot S, Cougot N, et al. The exon-junction-complex-component metastatic lymph node 51 functions in stress-granule assembly. J Cell Sci. 2007;120(Pt 16):2774–2784. doi:10.1242/jcs.009225
16. Gerbracht JV, Boehm V, Britto-Borges T, et al. CASC3 promotes transcriptome-wide activation of nonsense-mediated decay by the exon junction complex. Nucleic Acids Res. 2020;48(15):8626–8644. doi:10.1093/nar/gkaa564
17. Varis A, Wolf M, Monni O, et al. Targets of gene amplification and overexpression at 17q in gastric cancer. Cancer Res. 2002;62(9):2625–2629.
18. Tomioka N, Morita K, Kobayashi N, et al. Array comparative genomic hybridization analysis revealed four genomic prognostic biomarkers for primary gastric cancers. Cancer Genet Cytogenet. 2010;201(1):6–14. doi:10.1016/j.cancergencyto.2010.04.017
19. Salewski I, Henne J, Engster L, et al. Combined gemcitabine and immune-checkpoint inhibition conquers Anti-PD-L1 resistance in low-immunogenic mismatch repair-deficient tumors. Int J Mol Sci. 2021;22(11):5990. doi:10.3390/ijms22115990
20. Jang J, Lim D-S, Choi Y-E, et al. MLN51 and GM-CSF involvement in the proliferation of fibroblast-like synoviocytes in the pathogenesis of rheumatoid arthritis. Arthritis Res Ther. 2006;8(6):R170. doi:10.1186/ar2079
21. Bruno IG, Karam R, Huang L, et al. Identification of a microRNA that activates gene expression by repressing nonsense-mediated RNA decay. Mol Cell. 2011;42(4):500–510. doi:10.1016/j.molcel.2011.04.018
22. Schildberg T, Rauh J, Bretschneider H, et al. Identification of suitable reference genes in bone marrow stromal cells from osteoarthritic donors. Stem Cell Res. 2013;11(3):1288–1298. doi:10.1016/j.scr.2013.08.015
23. Rueda-Martinez C, Lamas O, Mataró MJ, et al. Selection of reference genes for quantitative real time PCR (qPCR) assays in tissue from human ascending aorta. PLoS One. 2014;9(5):e97449. doi:10.1371/journal.pone.0097449
24. Xu L, Dai W, Li J, et al. Methylation-regulated miR-124-1 suppresses tumorigenesis in hepatocellular carcinoma by targeting CASC3. Oncotarget. 2016;7(18):26027–26041. doi:10.18632/oncotarget.8266
25. Rademacher DJ, Bello AI, May JP. CASC3 biomolecular condensates restrict turnip crinkle virus by limiting host factor availability. J Mol Biol. 2023;435(16):167956. doi:10.1016/j.jmb.2023.167956
26. Mao H, Brown HE, Silver DL. Mouse models of Casc3 reveal developmental functions distinct from other components of the exon junction complex. RNA. 2017;23(1):23–31. doi:10.1261/rna.058826.116
27. Cunnick GH, Jiang WG, Gomez KF, et al. Lymphangiogenesis quantification using quantitative PCR and breast cancer as a model. Biochem Biophys Res Commun. 2001;288(4):1043–1046. doi:10.1006/bbrc.2001.5869
28. Gao H. Electromagnetic Radiation and Its Impact on Cancer Treatment. Thesis submitted to Cardiff University for the degree of Doctor of Philosophy, 2024.
29. Parr C, Sanders AJ, Davies G, et al. Matriptase-2 inhibits breast tumor growth and invasion and correlates with favorable prognosis for breast cancer patients. Clin Cancer Res. 2007;13(12):3568–3576. doi:10.1158/1078-0432.CCR-06-2357
30. Kerr AJ, Dodwell D, McGale P, et al. Adjuvant and neoadjuvant breast cancer treatments: a systematic review of their effects on mortality. Cancer Treat Rev. 2022;105:102375. doi:10.1016/j.ctrv.2022.102375
31. Ma HY, Sun R, Tian T, et al. NFIX suppresses breast cancer cell proliferation by delaying mitosis through downregulation of CDK1 expression. Cell Death Discov. 2025;11(1):77. doi:10.1038/s41420-025-02361-8
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.