Back to Journals » Neuropsychiatric Disease and Treatment » Volume 16
MiRNA-532-5p Regulates CUMS-Induced Depression-Like Behaviors and Modulates LPS-Induced Proinflammatory Cytokine Signaling by Targeting STAT3
Authors Yan X, Zeng D, Zhu H, Zhang Y, Shi Y, Wu Y, Tang H, Li D
Received 24 February 2020
Accepted for publication 25 October 2020
Published 12 November 2020 Volume 2020:16 Pages 2753—2764
DOI https://doi.org/10.2147/NDT.S251152
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Yu-Ping Ning
Xue Yan,* Dehao Zeng,* He Zhu, Yijing Zhang, Yuying Shi, Yingxiu Wu, Hongmei Tang, Detang Li
Pharmaceutical Department, The First Affiliated Hospital of Guangzhou University of Chinese Medicine, Guangzhou, Guangdong Province 510405, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Xue Yan Tel +86-020-36598864
Email [email protected]
Background: It is known that miR-532-5p is critical for neuronal differentiation. However, the role of miR-532-5p in depression remains unknown. This study aimed to investigate the role and mechanism of miR-532-5p in major depressive disorder (MDD).
Methods: The depression mice model was established by chronic unpredictable mild stress (CUMS) and confirmed by forced swimming test (FST) and sucrose preference test (SPT). The role of miR-532-5p in MDD was detected by tail suspension test (TST), FST, SPT and SIT. QRT-PCR was used to detect the expression of miR-139-5p in hippocampus and BV-2 microglia of mice. ELISA and Western blotting were used to detect the expression of the nitric oxide synthase (NOS), proinflammatory cytokines (IL-6, IL-1β, TNF-α, and MCP-1) and transcriptional activator 3 (STAT3). Luciferase reporter assay was used to verify the downstream target genes of miR-532-5p.
Results: MiR-532-5p was significantly reduced in the hippocampus of mice treated with CUMS. Overexpression of miR-532-5p significantly reduced CUMS-induced depression-like behaviors and suppressed the expression of IL-6, IL-1β, TNF-α and MCP-1. MiR-532-5p directly targeted signal transducers and STAT3 in BV2 cells. In addition, overexpression of miR-532-5p restrained the raise of inducible NOS and IL-6, IL-1 β, TNF-α and MCP-1 in LPS-exposed BV2 cells.
Conclusion: This study indicates that miR-532-5p plays an important role in CUMS-induced depression-like behaviors by targeting STAT3, and miR-532-5p may be a potential target for MDD therapy.
Keywords: miR-532-5p, STAT3, major depressive disorder, inflammation
Introduction
Major depressive disorder (MDD) is one of the most prevalent forms of mental disorders that affect millions of individuals worldwide1 and exerts substantial public health and financial costs to society. Although the symptoms of MDD have been widely acknowledged and recognized by clinicians,2,3 due to the lack of effective targets for diagnosis and treatment, the incidence rate and mortality rate of MDD increase year by year.4 Hence, the well understanding of specific molecular mechanisms involved in the development of laryngeal cancer is more urgent and will contribute to identify efficient targets for the diagnosis and treatment.
MiRNAs specifically bind to the UTR of the corresponding mRNAs and regulate the transcription of mRNAs.5 At present, miRNAs are well known to play critical roles in maintaining normal physiological state.6 Moreover, related studies have also shown that differential expression of miRNAs play a part in the occurrence of depression.7–9 More and more evidences show that many immunoreactive cells in central nervous system, especially microglia, play a key role in the occurrence of depression by influencing neural plasticity.10 MiRNAs play a key role in the maturation of the nervous system, regulate the activation of microglia, and regulate the occurrence and development of central nervous system diseases.11 Many research results showed that miR-532-5p was closely related to the occurrence of many diseases, and its expression and role in different diseases were also different.12 At present, the function of miR-532-5p in depression is not clear.
MiRNAs play a biological role by regulating downstream target genes.13 STAT3 plays a critical role in maintaining the tot potency of embryonic stem cells and regulating immunity.14 STAT3 knockout mice can cause embryo death, and their abnormal expression is closely related to many diseases. STAT3 can stimulate cell proliferation and survival in most cases, but it can also induce programmed cell death in certain cases, such as breast cell degeneration.15 The specific function performed by STAT3 is closely related to the tissue and cell type in which STAT3 is located. Microglia recently has been considered to be participated in the pathogenesis of MDD.16 Existing evidence has showed that in microglia and macrophages, high expression of STAT3 inhibits the excessive activation of microglia.17 Some studies have suggested that the secretion of inflammatory cytokines by the activated immune system is related to the occurrence of depression.18 There is a certain relationship between the secretion of inflammatory cytokines by the activated immune system.19 It is speculated that the effect of antidepressant therapy is achieved by inhibiting proinflammatory cytokines and promoting the secretion of anti-inflammatory cytokines. Microglia can activate microglia to synthesize and secrete anti-inflammatory cytokines and neurotrophic factors, which are beneficial to the repair of neuron damage, thus alleviating the development of depression. Therefore, we speculated that miR-532-5p might be related to the progression of MDD through Stat3 and inflammatory factors. This study was to explore the role and mechanism of miR-532-5p in MDD.
Materials and Methods
Animals
Healthy C57BL/6J male mice was obtained from Beijing Vital River Laboratory (China), SPF grade, 10–12 weeks old, body weight 23–26 g were selected. The mice were fed adaptively for one week with temperature of 21–25°C, humidity of 50–60% and illumination of 12 h, during which they were fed with water and food normally. All procedures were approved by The First Affiliated Hospital of Guangzhou University of Chinese Medicine and carried out in accordance with the guidelines published by China Animal Protection Association (regulations on the administration of experimental animals). For euthanasia, animals were deeply anesthetized with 3% sodium pentobarbital (30 mg/kg body weight, Sigma Chemical Co., St. Louis, MO, USA) through intraperitoneal injection and sacrificed by cervical dislocation.
CUMS Program
The CUMS procedure in this study was performed as described with a slight modification.20 In brief, mice in the CUMS group were subjected to various stressors for 5 weeks. The stressors included (1) food or water deprivation for 24 h, (2) overnight illumination, (3) absence of sawdust in the cage for 24 h, (4) moistened sawdust with water for 24 h, (5) cold or hot swimming at 8°C or 40°C, respectively, for 5 min, (6) tail nipping (1 cm from the tip of the tail), (7) physical restraint for 6 h, and (8) 45° tilt of the cage for 12 h. One of the above eight stimuli was given randomly every day.
Drug Administration
MiR-532-5p mimic (miR-532-5p) and miR-532-5p mimic-negative control (NC) were purchased from RiBoBio (Shanghai, China). MiR-532-5p/NC (Entranster ™ -in vivo, Engreen, China) was injected into mouse hippocampus. The mice were randomly divided into NC + control, miR-532-5p + control, NC + CUMS control and miR-532-5p + CUMS control, 6 mice per group. To evaluate the effect of miR-532-5p overexpression on CUMS-induced microglia activation, 1 week after injection, mice were exposed to a CUMS or control protocol for 5 weeks and then subjected to behavioral tests. FST was performed after SPT and the schedule of the experimental process is shown in Figure 1A.
 |
Figure 1 The expression level of miR532-5p in hippocampal tissue of CUMS-induced mouse depression model. (A) Timeline of experimental procedures in CUMS-induced mouse depression model. Results of behavioral test: SPT (B), FST (C), TST (D) and SIT (E). (F) MiRNA-532-5p expression levels in hippocampus of CUMS mice. n = 6. ** p < 0.01. |
Sucrose Preference Test (SPT)
All food and water were taken at 5:00 pm on the same day. At 8:00 the next morning, sucrose preference test conducted for 1 h. Two identical water bottles were placed on each cage, one was sugar water (2% sucrose solution), and the other was water, with a volume of 100 mL. After 1h, all water bottles were removed, the volume of liquid consumed was measured in the water bottle, and the sugar water preference rate (sugar water consumption volume/total consumption volume × 100%) was calculated. It was generally believed that the decrease in sugar water preference rate reflected the lack of pleasure in rodent models.21
Forced Swim Test (FST)
FST was usually used to assess rodent behavioral despair to screen for antidepressant activity of the drug. 25°C water was poured in a plastic bucket high enough. After mice were put in plastic buckets, the swimming time, struggling time and immobility time of the mice were recorded within 5 min. Swimming time refers to the time spent with the upper body or head of the mouse above the water. Struggling time referred to the time that the mouse climbs continuously along the wall of the plastic barrel. Immobility time, which was defined as minimal movements of both limbs and tails that were required to keep the rat in the water.22
Tail Suspension Test (TST)
One-third of the tail tip of the mouse was suspended in the suspended tail box, and the head of the mouse was facing the lens about 10 cm away from the bottom of the box. The latency of immobility within 6 min and the duration of immobility within 4 min were recorded by the camera system. Immobility meant that the animal gave up the active struggle and the body was in a state of suspension and immobility. The test was conducted with the lowest background noise.
Social Interaction Test (SIT)
Sit experiments were performed on the mice as described in the literature.23 The mice were placed in a separate measuring cage and allowed to explore freely for 120 min. A male juvenile mouse (4–5 weeks old) was then placed in a cage to record the time spent socializing between them during the test.
Real-Time PCR
Total RNA was extracted by TRIzol reagent (Haigene, Haerbin, China). Then, viiATM 7 real-time PCR system was detected by qRT-PCR. The expression level of miR-532-5p was calculated using 2−ΔΔCT method. The expression levels of miRNA and lncRNA were standardized by GADPH, respectively. The method of qRT-PCR was performed with reference.24 The primer sequences are described in Table 1.
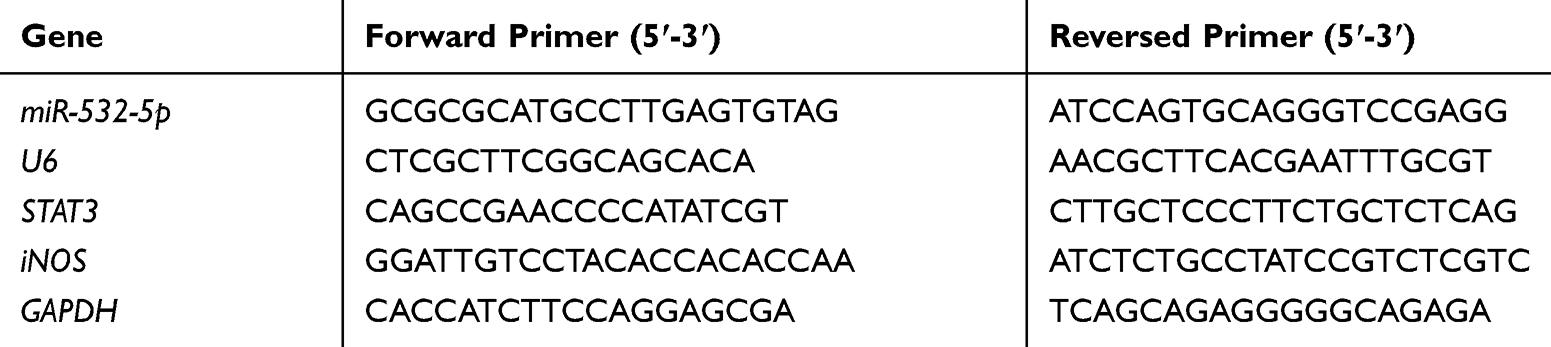 |
Table 1 Sequences of Primers Used in qRT-PCR |
Cell Culture and Treatment
The mouse BV-2 microglial cell line was obtained from the Cell Bank of the Chinese Academy of Sciences (Beijing, China), and the mouse HT22 hippocampal neuron cell line was obtained from the BeNa Culture Collection (Beijing, China). Bovine serum was cultured in RPMI 1640 medium. All cells were incubated at 37°C in a cell incubator containing 5% CO2. MiR-532-5p inhibitor/miR-532-5p inhibitor negative control (NC inhibitor) and STAT3 siRNA (si-STAT3)/si-STAT3 negative control (si-NC) were obtained from RiBoBio (Shanghai, China). RNAiMax and Lipofectamine 3000 with Plus Reagent (Thermo Fisher Scientific) cells were used to transiently transfect. Cells were harvested 24 h after transfection.
Enzyme-Linked Immunosorbent Assay (ELISA)
The kit method was used to detect the levels of cytokines in cell culture supernatant and hippocampus extract, including interleukin (IL) −6, IL-1β, monocyte chemotactic protein 1 (MCP-1) and tumor necrosis factor-α (TNF-α). ELISA kit was purchased from Nanjing Jinbai Biotechnology Co., Ltd. (Nanjing, China). Mouse TNF-α ELISA kit (DY410-05) was purchased from R&D Systems (Minneapolis, Minnesota, USA).
Luciferase Activity Assays
StarBase v2.0 (http://starbase.sysu.edu.cn/) was used to analyze the potential binding sites of miR-532-5p and STAT3. Wild-type STAT3 containing putative binding sites of miR-532-5p −3ʹ-UTR (WT) and mutant STAT3-3ʹ-UTR (MT) were constructed. The report vector containing WT or MT STAT3 3ʹ-UTR was co-transfected into BV-2 cells with miR-532-5p mimic or NC by Lipofectamine 3000. The luciferase activity was measured using a dual luciferase assay system (Promega) at 48 h after transfection.
Western Blot
Total proteins were extracted from the collected transfected cells using RIPA solution (GenePharma). To achieve protein denaturing, all protein samples were incubated in boiling water for 5 min, and the protein concentration quantified using the BCA Protein Assay Kit. Following by electrophoresis performed using 10% SDS-PAGE gel. Proteins were transferred to PVDF membranes, and PBS containing 5% non-fat milk was used to block the membranes for 2 h at 22°C. Then, it was incubated with rabbit anti-inducible nitric oxide synthase (iNOS) (1:1000; YBio, Shanghai, China); STAT3 (1:1000; YBio, Shanghai, China) and GAPDH (1:5000; YBio, Shanghai, China) at 4°C overnight. Then, a horseradish peroxidase-conjugated secondary antibody (1:5000; Abcam) was added for incubation at room temperature 1 h. Pierce ECL Western Blotting Substrate (Thermo Fisher Scientific) was then used to incubate with the membranes for 15 min at 22°C. Image J v1.46 software was used to process all data.25
Statistical Methods
All data were analyzed by SPSS 19.0 statistical software. The results of data analysis were showed as mean ± standard deviation (mean ± SD). Data analysis in multiple groups was used one-way ANOVA and LSD test. P < 0.05 indicated the difference was significant.
Results
The Expression of miR-532-5p Decreased in the Hippocampus of CUMS Mouse
To evaluate the expression of miR-532-5p in depression, depression mouse model was established by CUMS method and confirmed by behavior examinations. As shown in Figure 1, we found that the sucrose preference was significantly reduced (t-test, t(10) = 5.301, p = 0.006) (p < 0.01, Figure 1B), the immobility time in FST (t-test, t(10) = −5.249, p = 0.006) and TST (t-test, t(10) = −4.937, p = 0.008) was significantly increased (p < 0.01, Figure 1C and D), and the interaction time in SIT was decreased in mice exposed to CUMS compared with mice in control group (t-test, t(10) =5.006, p = 0.007) (p < 0.01, Figure 1E). These results suggest that the mouse CUMS model was successfully constructed. We therefore detected the expression of miR-532-5p and found that it was significantly decreased in mice exposed to CUMS compared with mice in control group (t-test, t (10) =7.052, P = 0.002) (p < 0.01, Figure 1F).
Overexpression of miR-532-5p Improved Depressive-Like Behaviors Induced by CUMS
To evaluate the role of miR-532-5p in depressive, miR-532-5p overexpression was carried out in mice (t-test, t (10) = −14.850, P = 0.000) (p <0.01, Figure 2A). The results showed that the overexpression of miR-532-5p have no effect on the sucrose preference in SPT (ANOVA, F(3,20) = 14.852, p = 0.001; LSD test, t =0.460, p = 0.616) (p > 0.01, Figure 2B), the immobility time in FST (ANOVA, F(3,20) = 10.868, p = 0.003; LSD test, t =−0.352, p = 0.774) (p > 0.01, Figure 2C) and TST (ANOVA, F(3,20) = 20.073, p = 0.000; LSD test, t =0.482, p = 0.689) (p > 0.01, Figure 2D) and the interaction time in SIT (ANOVA, F(3,20) = 19.504, p = 0.000; LSD test, t =−0.350, p = 0.718) (p > 0.01, Figure 2E) in control group of mice, suggesting that miR-532-5p overexpression did not change the depressive-like behaviors of control mice. However, overexpression of miR-532-5p significantly increased the sucrose preference in SPT (ANOVA, F(3,20) = 14.852, p = 0.001; LSD test, t =−5.558, p = 0.002) (p < 0.01, Figure 2B) and the interaction time in SIT (ANOVA, F(3,20) = 19.504, p = 0.000; LSD test, t =−4.517, p = 0.011) (p < 0.01, Figure 2E) and decreased the immobility time in FST (ANOVA, F(3,20) = 10.868, p = 0.003; LSD test, t =3.212, p = 0.033) (p < 0.01, Figure 2C) and TST (ANOVA, F(3,20) = 20.073, p = 0.000; LSD test, t =4.871, p = 0.001) (p < 0.01, Figure 2D) in mice exposed to CUMS, suggesting that miR-532-5p overexpression ameliorated depressive-like behaviors in mice exposed to CUMS. Taken together, these results suggest that miR-532-5p plays an important role in depressive.
 |
Figure 2 Overexpression of miR-532-5p improved CUMS-induced depression behaviors. (A) RT-PCR detected miR-532-5p expression levels in hippocampus. The effects of miR-532-5p on depression-like behavior in CUMS mice Impact: SPT (B), FST (C), TST (D) and SIT (E). n = 6. * p < 0.05, ** p < 0.01. |
Overexpression of miR-532-5p Improved Neuroinflammation
It is known that the secretion of inflammatory cytokines by the activated immune system is related to the occurrence of depression. To investigate the role of miR-532-5p in neuroinflammation, we further detected the inflammatory cytokines in mice exposed to CUMS. Compared with the control group, the expression of iNOS in the hippocampus of CUMS mice was increased significantly (ANOVA, F(3,20) = 60.002, p = 0.000; LSD test, t =−11.757, p = 0.000) (p < 0.01, Figure 3A and B), and overexpression of miR-532-5p significantly inhibited the increased expression of iNOS (ANOVA, F(3,20) = 60.002, p = 0.000; LSD test, t =7.821, p = 0.000) (p < 0.01, Figure 3A and B). The expression of IL-6, IL-1β, TNF-α and MCP-1 in hippocampus of CUMS mice were significantly increased (IL-6, ANOVA, F(3,20) = 72.672, p = 0.000; LSD test, t =−11.763, p = 0.000; IL-1.0 ANOVA, F(3,20) = 29.012, p = 0.000; LSD test, t =−7.531, p = 0.000; TNF-., ANOVA, F(3,20) = 21.476, p = 0.000; LSD test, t =−7.485, p = 0.000; MCP-1, ANOVA, F(3,20) = 99.092, p = 0.000; LSD test, t =−7.485, p = 0.000)(p < 0.01, Figure 3B–E), while miR-532-5p overexpression significantly inhibited the increased expression of cytokines (IL-6, ANOVA, F(3,20) = 72.672, p = 0.000; LSD test, t =4.555, p = 0.010; IL-1β, ANOVA, F(3,20) = 29.012, p = 0.000; LSD test, t = 4.717, p = 0.001; TNF-α, ANOVA, F(3,20) = 21.476, p = 0.000; LSD test, t =3.145, p = 0.035; MCP-1, ANOVA, F(3,20) = 99.092, p = 0.000; LSD test, t =3.600, p = 0.023) (p < 0.05, p < 0.01, Figure 3B–E). Taken together, these results suggest that overexpression of miR-532-5p improves neuroinflammation.
 |
Figure 3 MiR-532-5p overexpression inhibited CUMS-induced hippocampal neuroinflammation in mice. (A) Effect of miR-532-5p overexpression on iNOS levels in CUMS mice. (B–E) MiR-532-5p overexpression inhibited CUMS-induced hippocampal cytokine expression. ELISA assay analyzed the levels of IL-6 (B), IL-1β (C) MCP-1 (D) and TNF-α (E). N = 6. * p < 0.05, ** p < 0.01. |
Overexpression of miR-532-5p Inhibited Microglial Activation in vitro
Microglia is known to be participated in the pathogenesis of MDD. To further investigate the underlying mechanism of miR-532-5p in depression, we detected the microglial activation in BV-2 cells after LPS stimulation. The results showed that the expression of miR-532-5p in BV2 cells after LPS stimulation was significantly reduced compared with the control group (12 h, ANOVA, F (2,6) = 53.391, p = 0.000; LSD test, t =6.934, p = 0.000; 24 h, ANOVA, F(2,6) = 53.391, p = 0.000; LSD test, t =8.773, p = 0.000) (p < 0.01, Figure 4A). As shown in Figure 4B (ANOVA, F(3,8) = 38.698, p = 0.000; LSD test, t =−8.905, p = 0.000) and 4C (ANOVA, F(3,8) = 74.173, p = 0.000; LSD test, t =−11.065, p = 0.000), the expression of iNOS in BV2 cells induced by LPS was significantly increased (p < 0.01, Figure 4B and C), and overexpression of miR-532-5p significantly inhibited the increased expression of iNOS in LPS-induced BV2 cells (mRNA, ANOVA, F(3,8) = 38.698, p = 0.000; LSD test, t =5.852, p = 0.000; protein, ANOVA, F(3,8) = 74.173, p = 0.000; LSD test, t =8.894, p = 0.000) (p < 0.01, Figure 4B and C). Meanwhile, the expression of proinflammatory cytokines (IL-6, IL-1β, TNF-α and MCP-1) were significantly increased in LPS-induced BV2 cells (IL-6, ANOVA, F(3,8) = 57.549, p = 0.000; LSD test, t =−10.891, p = 0.000; IL-1β, ANOVA, F(3,8) = 64.137, p = 0.000; LSD test, t =−10.773, p = 0.000; TNF-α, ANOVA, F(3,8) = 58.350, p = 0.000; LSD test, t =−10.790, p = 0.000; MCP-1, ANOVA, F(3,8) = 89.036, p = 0.000; LSD test, t =−12.494, p = 0.000) (p < 0.01, Figure 4D–G), whereas overexpression of miR-532-5p significantly inhibited the increased proinflammatory cytokines (IL-6, IL-1β, TNF-αand MCP-1) in LPS-induced BV2 cells (IL-6, ANOVA, F(3,8) = 57.549, p = 0.000; LSD test, t =3.582, p = 0.023; IL-1β, ANOVA, F(3,8) = 64.137, p = 0.000; LSD test, t =5.696, p = 0.000; TNF-α, ANOVA, F(3,8) = 58.350, p = 0.000; LSD test, t =6.751, p = 0.000; MCP-1, ANOVA, F(3,8) = 89.036, p = 0.000; LSD test, t =4.983, p = 0.000) (p < 0.05, p < 0.01, Figure 4D–G). Altogether, these results suggest that overexpression of miR-532-5p inhibits microglial activation in vitro.
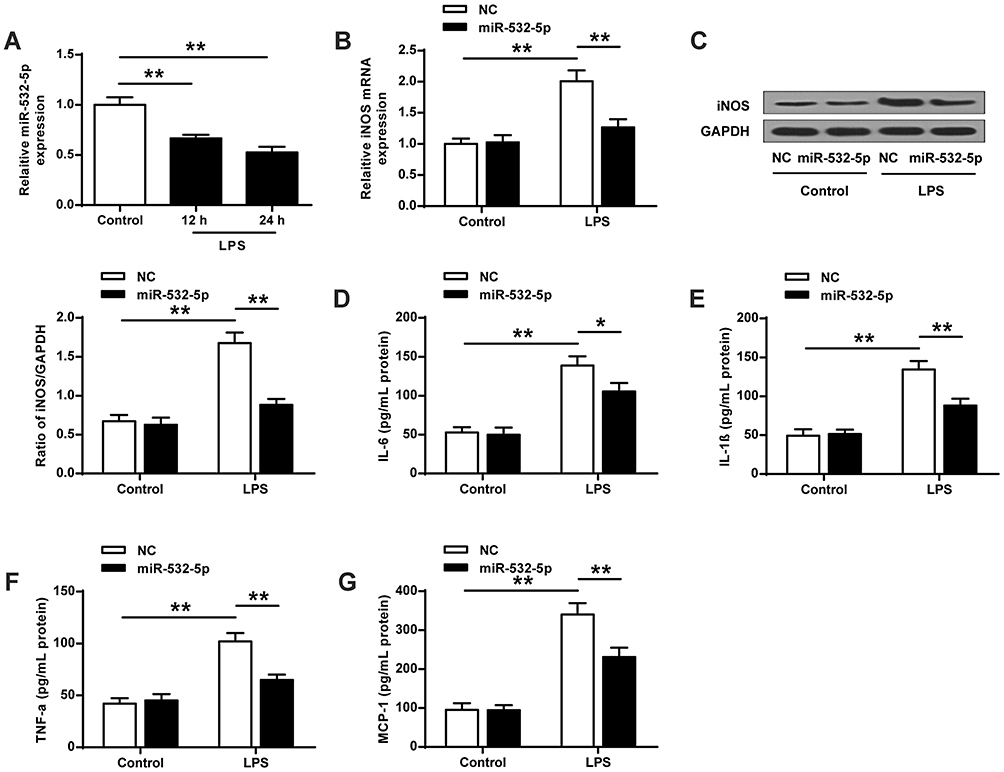 |
Figure 4 MiR-532-5p expression inhibited microglial activation in vitro. (A) LPS (100 ng/mL) treatment reduced miR-532-5p expression, q-PCR detected miR-532- 5B expression level. (B and C) LPS-induced iNOS mRNA and protein expression levels in BV2 cells after miR-532-5p transfection. MiR-532-5p transfected cells inhibited LPS-induced expression of cytokines in BV2 cells. The expression level of cytokines IL-6 (D), IL-1β (E), TNF-α (F) and MCP-1 (G) was determined by ELISA. N = 3. * p < 0.05, * * p < 0.01. |
MiR-532-5p Regulated STAT3 Expression by Directly Targeting Its 3ʹUTR
Next, the target of miR-532-5p in MDD was explored. We predicted it through the online prediction tool Starbase v2.0, and STAT3 was identified as a potential target of miR-532-5p (Figure 5A). The results of luciferase reporter assay showed that the luciferase activity of pGL3-REPOR-STAT3-WT was reduced by the miR-532-5p mimic (ANOVA, F(3,8) = 54.559, p = 0.000; LSD test, t =11.555, p = 0.000) (p < 0.01, Figure 5B), but the luciferase activity of pGL3-REPOR-STAT3-mut did not change significantly (ANOVA, F(3,8) = 54.559, p = 0.000; LSD test, t = 0.815, p = 0.395). In addition, in BV-2 cells, as shown in Figure 5C and D, miR-532-5p overexpression significantly reduced the mRNA (ANOVA, F(3,8) = 146.661, p = 0.000; LSD test, t =10.873, p = 0.000) and protein (ANOVA, F(3,8) = 124.902, p = 0.000; LSD test, t = 6.112, p = 0.001) expression of STAT3 in miR-532-5p mimic group compared with NC group, while miR-532-5p inhibition significantly increased the mRNA (ANOVA, F(3,8) = 146.661, p = 0.000; LSD test, t =−10.453, p = 0.000) and protein (ANOVA, F(3,8) = 124.902, p = 0.000; LSD test, t =−11.686, p = 0.000) expression of STAT3 in miR-532-5p inhibitor group compared with NC group (Figure 5C and D). Moreover, miR-532-5p overexpression significantly reduced the mRNA (ANOVA, F (3,8) = 49.212, p = 0.000; LSD test, t =6.213, p = 0.000) and protein protein (ANOVA, F(3,8) = 81.912, p = 0.000; LSD test, t =9.202, p = 0.000) expression of STAT3 in CUMS-induced mice transfected with miR-532-5p mimic compared with CUMS-induced mice transfected with NC (Figure 5E and F), but had no effect on the mRNA and protein expression of STAT3 in control mice. These results demonstrate that miR-532-5p may perform its biological function through STAT3.
 |
Figure 5 MiR-532-5p regulated STAT3 expression level by directly targeting the 3ʹUTR of STAT3. (A) MiR-532-5p and STAT3 binding sites. (B) Luciferase reporter assay was used for detection of luciferase activity. (C) The mRNA expression level of STAT3 in cells treated with miR-532-5p mimic. (D) The protein expression level of STAT3 in cells treated with miR-532-5p mimic. (E) The mRNA expression level of STAT3 in mice treated with miR-532-5p mimic. (F) The protein expression level of STAT3 in mice treated with miR-532-5p mimic. N = 6. ** p < 0.01. |
Role of miR-532-5p/STAT3 Axis in LPS-Induced Microglial Activation
To further investigate the role of STAT3 in BV-2 cells after LPS stimulation, we firstly detected the expression of STAT3 in BV-2 cells after LPS stimulation. The results showed that LPS significantly increased the mRNA (t-test, t (4) = −11.609, p = 0.000) and protein expression (t-test, t (4) = −9.613, p = 0.001) of STAT3 compared with the control group (p <0.01, Figure 6A and B). Compared with the si-NC group, the mRNA (t-test, t (4) = 8.941, p = 0.001) and protein expression (t-test, t (4) = 9.999, p = 0.001) of STAT3 in the si-STAT3 group were significantly reduced (p < 0.01, Figure 6C and D). As shown in Figure 6E, compared with the si-NC group, LPS significantly increased the expression of iNOS (ANOVA, F (5,12) = 42.455, p = 0.000; LSD test, t =−12.007, p = 0.000), while si-STAT3 significantly inhibited the expression of iNOS (ANOVA, F(5,12) = 42.455, p = 0.000; LSD test, t =8.895, p = 0.000) (p < 0.01, Figure 6E). Co-transfection of si-STAT3 with miR-532-5p inhibitor reversed the effect of si-STAT3 on the expression of iNOS in BV2 cells (ANOVA, F (5, 12) = 42.455, p = 0.000; LSD test, t =−3.350, p = 0.029) (p < 0.05, Figure 6E). These results suggest that miRNA-532 modulates microglial activation by targeting STAT3.
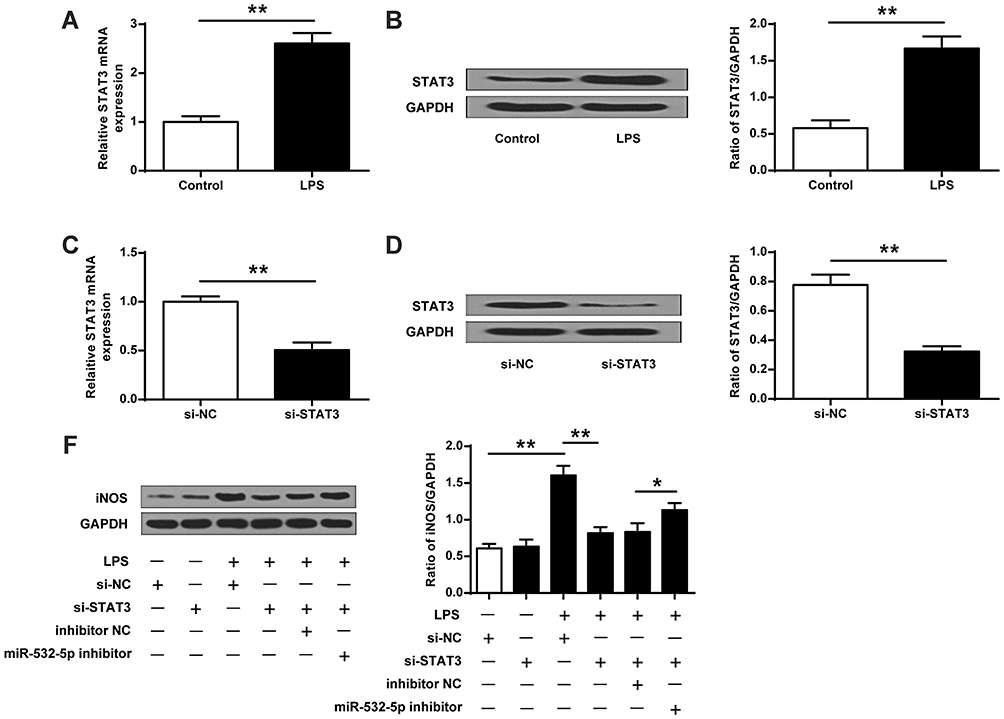 |
Figure 6 The role of miR-532-5p/STAT3 in LPS-induced microglia activation. (A and B) Detection of mRNA and protein expression of STAT3 in BV2 cells with or without stimulating by LPS. (C and D) STAT3 siRNA reduced the mRNA and protein expression of STAT3. (E) The effect of si-STAT3 and miR-532-5p on the expression of iNOS in BV2 cells treated with or without LPS. N = 3. * p < 0.05, ** p < 0.01. |
Discussion
Symptoms of depression include cognitive impairment and forgetfulness, which suggest abnormal synaptic function in the pathophysiology of MDD.26 Studies have shown that MDD is accompanied by other diseases, such as cardiovascular disease, osteoporosis, diabetes, cerebral ischemia and so on.27 However, the current understanding of its molecular mechanism is limited, and there are no effective treatments with low adverse reactions.28 Recently, a large number of studies have shown that cytokines secreted by brain microglia cells after activation are also related to mood disorders, and the development of new antidepressant drugs targeting them is becoming a research hotspot in this field.29
The CUMS depression model is the closest to the type of depression induced by external stimuli in humans, and has good credibility and validity.30 This study found that sucrose preference was reduced in CUMS mice, immobility time in FST and TST mice was increased, and the SI time is shortened. These results indicated that the model mice had decreased exercise capacity, interest and pleasure, increased hopelessness and helpless behavior. The above results were consistent with previous studies, indicating that the depression model was successfully established.
More and more studies have found that miRNAs are associated with a variety of mental diseases, such as schizophrenia, autism, drug dependence, and bipolar disorder.31 Some studies have found that the plasma level of miR-144-5p in patients with depression is reduced, and its expression level is related to the manifestation of depressive symptoms.32 It has been reported that miRNA is closely related to depression and participates in the process of hippocampal neuron generation. MiR-532-5p as a member of miRNA, there are differences in the role in different diseases.12 From a structural point of view, the hippocampus is part of the limbic system and develops nerve fiber connectivity with emotion-related brain regions, for instance, the prefrontal cortex and amygdala. Nevertheless, it has been widely reported that there is a significant reduction in hippocampal volume in depression patients.33 So, we detected the expression and role of miR-144-5p in this depression mouse model. Our study found that the expression of miR-532-5p in the hippocampus of mice was significantly decreased in CUMS group. Among the CUMS group mice, the sucrose preference index of the mice in the miR-532-5p group was reduced, the immobility time in TST and FST and the interaction time in SIT was prolonged. These results suggest that miR-532-5p may play important role in the development of depression.
Mounting data are indicating that major depression is related to diverse functions of the immune system.34 Several observations indicate that cytokine concentrations might also relate to the intensity of depressive manifestations.35 Excessive secretion of cytokines, acute phase reaction, increase of monocyte and neutrophil count in peripheral blood and increase of autoantibody titer.36 IL-6, IL-1β, TNF-α, MCP-1 and iNOS are typical cellular inflammatory factors. This study found that miR-532-5p overexpression prevented the increase of cytokine upregulation after stress/LPS. CUMS and LPS treatment can raise IL-6, IL-1β, TNF- α and MCP-1 expression. The results confirmed that miR-532 overexpression prevented the increase of cytokine upregulation after stress/LPS.
In recent years, it has been reported that miRNA targets regulatory related pathways and affects the development of disease.37 STAT3 belongs to the STAT family and is a transcription factor that can regulate gene expression.38 STAT can stimulate the production of epidermal growth factors Response. When STAT3 is phosphorylated, it will cause STAT3 dimerization, translocate to the nucleus, bind to DNA, and participate in the body’s inflammatory response.39 At present, the role of STAT3 in depression was unclear. This study found that STAT3 was a potential target of miR-532-5p. In addition, miR-532-5p transfected BV2 cells reduced the expression level of STAT3, while miR-532-5p inhibitors had the opposite result. LPS increased the expression level of STAT3. In addition, si-STAT3 significantly inhibited the expression of iNOS. Co-transfection of si-STAT3 and miR-532-5p inhibitor in BV2 cells reversed the effect of si-STAT3 on the expression levels of iNOS. These results indicate that miR-532 −5p can regulate hippocampal neuronal cell damage by STAT3.
Conclusion
MiR-532-5p can regulate hippocampal inflammatory signaling through STAT3, and down-regulation of miR-532-5p may be involved in depression development by abrogating the expression of stress/LPS-stimulated cytokines.
Funding
This work was supported by the Scientific Research Project of Traditional Chinese Medicine Bureau of Guangdong Province, 20191121.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Wang QZ, Shelton RC, Dwivedi Y. Interaction between early-life stress and FKBP5 gene variants in major depressive disorder and post-traumatic stress disorder: a systematic review and meta-analysis. J Affect Disord. 2018;225:422–428. doi:10.1016/j.jad.2017.08.066
2. Medeiros GC, Seger L, Grant JE, Tavares H. Major depressive disorder and depressive symptoms in intermittent explosive disorder. Psychiatry Res. 2018;262:209. doi:10.1016/j.psychres.2018.02.001
3. Baumeister SE, Leitzmann MF, Bahls M, et al. Associations of leisure-time and occupational physical activity and cardiorespiratory fitness with incident and recurrent major depressive disorder, depressive symptoms, and incident anxiety in a general population. J Clin Psychiatry. 2017;78(1):E41–E47. doi:10.4088/JCP.15m10474
4. Janssenkallenberg H, Schulz H, Kluge U, et al. Acculturation and other risk factors of depressive disorders in individuals with Turkish migration backgrounds. BMC Psychiatry. 2017;17(1):264. doi:10.1186/s12888-017-1430-z
5. Zhou H, Rigoutsos I. MiR-103a-3p targets the 5ʹ UTR of GPRC5A in pancreatic cells. RNA. 2014;20(9):1431–1439. doi:10.1261/rna.045757.114
6. Lu TX, Rothenberg ME. MicroRNA. J Allergy Clin Immunol. 2018;141(4):1202–1207. doi:10.1016/j.jaci.2017.08.034
7. Ming Y, Zhou D, Hui Z, et al. MSDD: a manually curated database of experimentally supported associations among miRNAs, SNPs and human diseases. Nucleic Acids Res. 2017;(D1):D1.
8. Lopizzo N, Zonca V, Cattane N, Pariante C, Cattaneo A. miRNAs in depression vulnerability and resilience: novel targets for preventive strategies. J Neural Transm. 2019;126.
9. Lin -C-C, Tsai M-C, Lee C-T, Sun M-H, Huang T-L. Antidepressant treatment increased serum miR-183 and miR-212 levels in patients with major depressive disorder. Psychiatry Res. 2018;270.
10. Hellwig S, Brioschi S, Dieni S, Frings L, Biber K. Altered microglia morphology and higher resilience to stress-induced depression-like behavior in CX3CR1-deficient mice. Brain Behav Immun. 2015;55.
11. Brites D, Fernandes A. Neuroinflammation and depression: microglia activation, extracellular microvesicles and microRNA dysregulation. Front Cell Neurosci. 2015;9:476.
12. Griesing S, Kajino T, Tai M, et al. Abstract 2529: TTF-1/NKX2-1 induced miR-532-5p targets KRAS and MKL2 oncogenes and causes apoptosis in lung adenocarcinoma cells. Cancer Res. 2017;77:2529.
13. Wang Z, Xi J, Hao X, Deng W, Wang H. Red blood cells release microparticles containing human argonaute 2 and miRNAs to target genes of plasmodium falciparum. Emerg Microbes Infect. 2017;6(8):e75. doi:10.1038/emi.2017.63
14. Wang Y, Shen Y, Wang S, Shen Q, Zhou X. The role of STAT3 in leading the crosstalk between human cancers and the immune system. Cancer Lett. 2017;415:117–128. doi:10.1016/j.canlet.2017.12.003
15. Johnson DE. Targeting the IL-6/JAK/STAT3 signalling axis in cancer. Nat Rev Clin Oncol. 2018;15(4):234.
16. Tang J, Yu WB, Chen S, Gao ZD, Xiao BG. Microglia polarization and endoplasmic reticulum stress in chronic social defeat stress induced depression mouse. Neurochem Res. 2018;43(5):985–994. doi:10.1007/s11064-018-2504-0
17. Ohgomori T, Yamasaki R, Takeuchi H, Kadomatsu K, Kira J, Jinno S. Differential activation of neuronal and glial STAT3 in the spinal cord of the SOD1(G93A) mouse model of amyotrophic lateral sclerosis. Eur J Neurosci. 2017;46(4):2001–2014. doi:10.1111/ejn.13650
18. Rajabi M, Mohaddes G, Farajdokht F, Nayebi RS, Mesgari M, Babri S. Impact of loganin on pro-inflammatory cytokines and depression- and anxiety-like behaviors in male diabetic rats. Physiol Int. 2018;105(2):116. doi:10.1556/2060.105.2018.2.8
19. Ji YB, Bo CL, Xue XJ, et al. Association of inflammatory cytokines with the symptom cluster of pain, fatigue, depression, and sleep disturbance in chinese patients with cancer. J Pain Symptom Manage. 2017;54(6):843–852. doi:10.1016/j.jpainsymman.2017.05.003
20. Liu W, Xue X, Xia J, Liu J, Qi Z. Swimming exercise reverses CUMS-induced changes in depression-like behaviors and hippocampal plasticity-related proteins. J Affect Disord. 2018;227:126–135. doi:10.1016/j.jad.2017.10.019
21. Yu J, Li XY, Cao XD, Wu GC. Sucrose preference is restored by electro-acupuncture combined with chlorimipramine in the depression-model rats. Acupunct Electrother Res. 2006;31(3–4):223–232. doi:10.3727/036012906815844201
22. Yankelevitch-Yahav R, Franko M, Huly A, Doron R. The forced swim test as a model of depressive-like behavior. J Vis Exp. 2015;(97):52587.
23. Gjendal K, Sørensen D, Kiersgaard M, Ottesen J. Hang on: an evaluation of the hemp rope as environmental enrichment in C57BL/6 mice. Anim Welf. 2017;26(4):437–447. doi:10.7120/09627286.26.4.437
24. Bernard PS, Wittwer CT. Real-time PCR technology for cancer diagnostics. Clin Chem. 2002;48(8):1178–1185. doi:10.1093/clinchem/48.8.1178
25. Tomasello L, Cluts L, Croce CM. Experimental validation of MicroRNA targets: analysis of microRNA targets through western blotting. Methods Mol Biol. 2019;1970:341–353.
26. Radziwiłłowicz W, Lewandowska M. From traumatic events and dissociation to body image and depression symptoms – in search of self-destruction syndrome in adolescents who engage in nonsuicidal self-injury. Psychiatr Pol. 2017;51(2):283–301. doi:10.12740/PP/63801
27. Martinez P, Castro A, Alonso D, Vohringer PA, Rojas G. Effectiveness of the management of major depressive episodes/disorder in adults with comorbid chronic physical diseases: a protocol for a systematic review and meta-analysis. BMJ Open. 2017;7(7):421–426. doi:10.1136/bmjopen-2016-011249
28. Gartlehner G, Wagner G, Matyas N, et al. Pharmacological and non-pharmacological treatments for major depressive disorder: review of systematic reviews. BMJ Open. 2017;7(6):321–324. doi:10.1136/bmjopen-2016-014912
29. Hwang DS, Gu PS, Kim N, Jang YP, Oh MS. Effects of Rhei Undulati Rhizoma on lipopolysaccharide-induced neuroinflammation in vitro and in vivo. Environ Toxicol. 2017;33(7).
30. Lu J, Shao RH, Jin SY, Hu L, Tu Y, Guo JY. Acupuncture ameliorates inflammatory response in a chronic unpredictable stress rat model of depression. Brain Res Bull. 2017;128:106–112. doi:10.1016/j.brainresbull.2016.11.010
31. O’Dushlaine C, Kenny E, Heron E, et al. Molecular pathways involved in neuronal cell adhesion and membrane scaffolding contribute to schizophrenia and bipolar disorder susceptibility. Mol Psychiatry. 2011;16(3):286–292. doi:10.1038/mp.2010.7
32. Mo M, Xiao Y, Huang S, Cen L, Xu P. MicroRNA expressing profiles in A53T mutant alpha-synuclein transgenic mice and Parkinsonian. Oncotarget. 2017;8(1):15–28. doi:10.18632/oncotarget.13905
33. Schriber RA, Anbari Z, Robins RW, Conger RD, Guyer AE. Hippocampal volume as an amplifier of the effect of social context on adolescent depression. Clin Psychol Sci. 2017;5(4):632–649. doi:10.1177/2167702617699277
34. Madi Y, Yogev A, Bar-Gad M, Amital H, Amital D. serum cytokine concentrations association with the severity of symptoms iv major depression. Harefuah. 2017;156(12):753–756.
35. Liu RH, Pan JQ, Tang XE, Bing L, Ma WL. The role of immune abnormality in depression and cardiovascular disease. J Geriatr Cardiol. 2017;14(11):703–710.
36. Kang A, Xie T, Zhu D, Shan J, Zheng X, Zheng X. Suppressive effect of ginsenoside Rg3 against lipopolysaccharide-induced depression-like behavior and neuroinflammation in mice. J Agric Food Chem. 2017;65(32):6861–6869. doi:10.1021/acs.jafc.7b02386
37. Backes C, Kehl T, Stockel D, et al. miRPathDB: a new dictionary on microRNAs and target pathways. Nucleic Acids Res. 2017;45(D1):D90–D96. doi:10.1093/nar/gkw926
38. Tangye SG, Forbes LR, Leiding J, et al. Low IgE is insufficiently sensitive to guide genetic testing of STAT3 gain-of-function mutations. Clin Chem. 2017;63(9):1539–1540. doi:10.1373/clinchem.2017.273458
39. Johnston PA, Sen M, Hua Y, et al. High content imaging assays for IL-6-induced STAT3 pathway activation in head and neck cancer cell lines. Methods Mol Biol. 2018;1683:229–244.
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.