Back to Journals » OncoTargets and Therapy » Volume 8
miR-32 functions as a tumor suppressor and directly targets SOX9 in human non-small cell lung cancer
Authors Zhu D, Chen H, Yang X, Chen W, Wang L, Xu J, Yu L
Received 8 August 2014
Accepted for publication 24 November 2014
Published 20 July 2015 Volume 2015:8 Pages 1773—1783
DOI https://doi.org/10.2147/OTT.S72457
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Jianmin Xu
This paper has been retracted
Dan Zhu, Hui Chen, Xiguang Yang, Weisong Chen, Linying Wang, Jilin Xu, Long Yu
Department of Respiratory Medicine, Jinhua Municipal Central Hospital, Jinhua, People’s Republic of China
Purpose: MicroRNA-32 (miR-32) is dysregulated in certain human malignancies and correlates with tumor progression. However, its expression and function in non-small cell lung cancer (NSCLC) remain unclear. Thus, the aim of this study was to explore the effects of miR-32 expression on NSCLC tumorigenesis and development.
Methods: Using real-time quantitative reverse-transcription polymerase chain reaction (qRT-PCR), we detected miR-32 expression in NSCLC cell lines and primary tumor tissues. The association of miR-32 expression with clinicopathological factors and prognosis was also analyzed. Then, the effects of miR-32 expression on the biological behavior of NSCLC cells were investigated. Finally, the potential regulatory effect of miR-32 on SOX9 expression was confirmed.
Results: miR-32 expression levels were significantly downregulated in NSCLC compared with the corresponding noncancerous lung tissues (P<0.001). In addition, decreased miR-32 expression was significantly associated with lymph node metastasis (P=0.002), advanced tumor/nodes/metastasis (TNM) classification stages (P<0.001), and shorter overall survival (P<0.001). Multivariate regression analysis corroborated that downregulated miR-32 expression was an independent unfavorable prognostic factor for NSCLC patients. In vitro studies demonstrated that miR-32 overexpression reduced A549 cell proliferation, migration, and invasion, and promoted apoptosis. Furthermore, SOX9 was confirmed as a direct target of miR-32, using a luciferase reporter assay.
Conclusion: These findings indicate that miR-32 may act as a tumor suppressor in NSCLC and could serve as a novel therapeutic agent for miR-based therapy.
Keywords: prognosis, proliferation, apoptosis, invasion, migration
Introduction
With approximately 1.5 million new cases diagnosed yearly, lung cancer is one of the most common neoplasias and the leading cause of cancer-related deaths worldwide.1 Despite recent advances in clinical and experimental oncology, the prognosis of lung cancer remains unfavorable. Non-small cell lung cancer (NSCLC) accounts for more than 80% of all lung cancers, and its 5-year survival rate is only approximately 15%. As in other cancers, NSCLC development is a multistep process involving the accumulation of genetic and epigenetic changes. Previous studies have demonstrated diverse genetic alterations in NSCLC; however, the molecular mechanisms underlying NSCLC carcinogenesis and progression are highly complex, and the further identification of new candidate molecules that participate in these processes is important for improving the diagnosis, prevention, and treatment of this disease.
MicroRNAs (miRs) are a class of short (approximately 22 nucleotides in length), endogenous, single-stranded, nonprotein-coding RNAs that directly bind to the 3′ untranslated regions (3′ UTRs) of target messenger RNAs (mRNAs), leading to mRNA degradation or translational suppression.2 miRs are involved in many different biological processes, including cell growth, apoptosis, development, differentiation, and endocrine homeostasis.3 Accumulating studies also suggest that miRs play essential roles in the biology of human cancers, which may provide a new and promising way to treat cancer.4–6 The dysregulation of miR expression has frequently been reported and is closely associated with tumor initiation, promotion, and progression. miR-32 has been implicated in the pathogenesis of several human malignancies. miR-32 expression is upregulated in colorectal cancer,7 kidney cancer,8 prostate cancer,9 and multiple myeloma,10 and acts as a potential oncogene in these tumors; however, miR-32 expression is also significantly decreased in gastric cancer and osteosarcoma, and acts as a candidate tumor suppressor.11,12 Notably, Yanaihara et al revealed miR-32 downregulation in lung cancer by miR microarray analysis.13 In their study, these authors detected the expression levels of 352 miRs in 104 pairs of lung cancer and normal adjacent tissue, and identified 43 miRs that were differentially expressed, including miR-32. However, the correlation between miR-32 dysregulation and the clinicopathological characteristics of NSCLC has not yet been evaluated, and the biological roles of miR-32 and its direct functional targets in NSCLC remain poorly understood.
SOX9, which is a high-mobility-group box transcription factor, is a key regulator of developmental processes, including male sex determination, chondrogenesis, neurogenesis, and neural crest development.14,15 Recent cogent evidence has provided a link between SOX9 and cancer development,16 and SOX9 upregulation has been reported in breast cancer,17 colorectal cancer,18 prostate cancer,19 and lung adenocarcinoma,20 where its expression correlated with malignancy and overall patient survival. Intriguingly, several miRs, including miR-124,21 miR-145,22 and miR-1247,23 participate in the regulation of SOX9 activity in different tissues; however, the potential regulatory effect of miR-32 on SOX9 expression in NSCLC has not been confirmed.
In the present study, we examined miR-32 expression in NSCLC tissues and cell lines, using real-time quantitative reverse-transcription polymerase chain reaction (qRT-PCR). The association of miR-32 levels with clinicopathologic features and prognosis was also analyzed. Furthermore, we investigated the effects of miR-32 expression on the proliferation, apoptosis, invasion, and migration of NSCLC cells. Finally, SOX9 was identified as a direct target of miR-32, using a luciferase reporter assay.
Materials and methods
Patients and clinical specimens
Paired NSCLC and adjacent noncancerous lung tissues (pathologically confirmed normal tissues and >2 cm away from the cancer tissues) were obtained from 115 patients who received curative resection of NSCLC at Jinhua Municipal Central Hospital (Jinhua, Zhejiang Province, People’s Republic of China) from January 2007 to December 2009. These tissues were flash-frozen in liquid nitrogen immediately after resection and stored at -80°C until use. None of the patients received neoadjuvant chemo- or radiotherapy before surgery. The patient characteristics are shown in Table 1. Follow-up information was available for all patients. Overall survival (OS) was defined as the time from the day of operation to death or, for living patients, to the date of the last follow-up visit. This study was approved by the Research Ethics Committee of our hospital, and written informed consent was obtained from each patient.
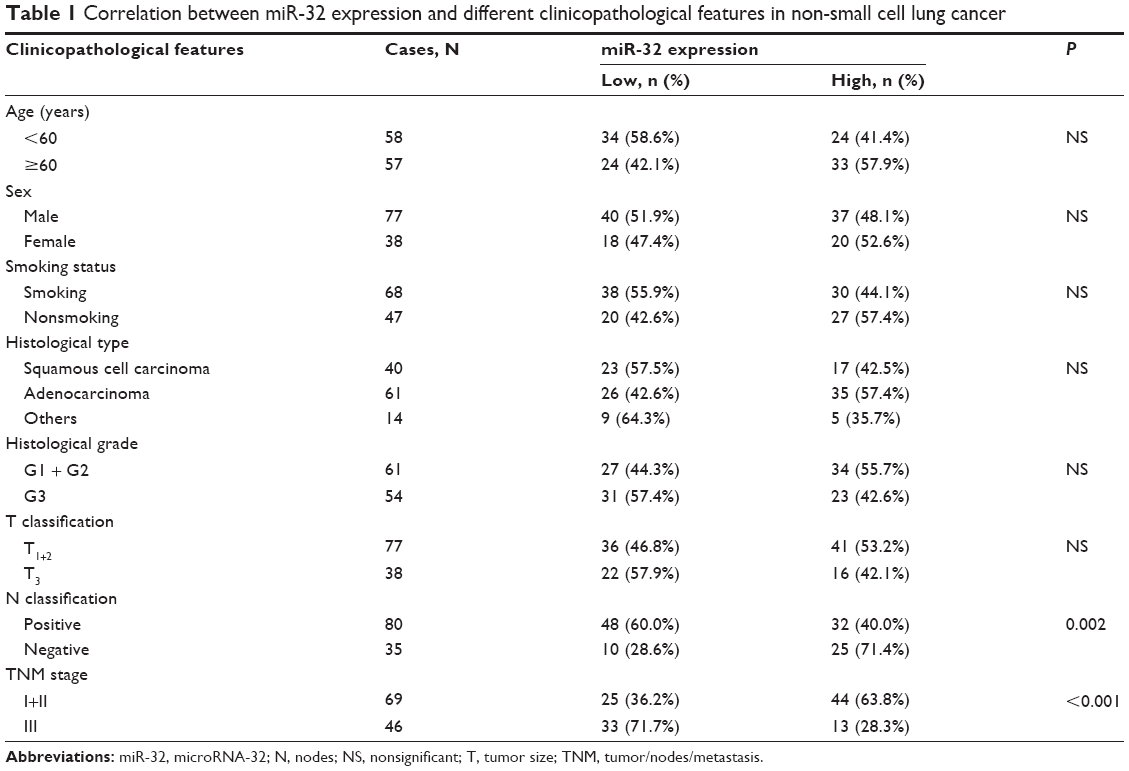 | Table 1 Correlation between miR-32 expression and different clinicopathological features in non-small cell lung cancer |
Cell lines and miR transfection
Four NSCLC cell lines (A549, H460, 95D, and HCC827) and normal lung epithelial cells were purchased from American Type Culture Collection (ATCC) (Manassas, VA, USA) and maintained in Roswell Park Memorial Institute (RPMI) 1640 medium (Invitrogen®; Life Technologies Corp, Carlsbad, CA, USA) supplemented with 10% heat-inactivated fetal bovine serum (FBS), 100 units/mL of penicillin G sodium, and 100 μg/mL streptomycin sulfate. All of the cells were incubated at 37°C in a humidified atmosphere with 5% CO2.
For RNA transfection, the cells were seeded into each well of 24-well plates, incubated overnight, and then transfected with mature miR-32 mimics, miR-32 inhibitors (anti-miR-32), or negative control (miR-NC or anti-miR-NC; GenePharma, Shanghai, People’s Republic of China) using Lipofectamine® 2000 (Invitrogen; Life Technologies Corp) according to the manufacturer’s instructions.
RNA extraction and qRT-PCR
Total RNA was isolated using TRIzol® reagent (Invitrogen; Life Technologies Corp) according to the manufacturer’s instructions. The reverse reaction was performed using 100 ng of total RNA at the templete with looped primers. qRT-PCR was performed using the standard TaqMan® miRNA assay protocol on an ABI7500 Real-Time PCR System, with the following cycling conditions: 95°C for 10 minutes, followed by 40 cycles of 95°C for 15 seconds, 60°C for 30 seconds, and 74°C for 5 seconds. U6 small nuclear RNA was used as an internal control. The threshold cycle (Ct) was defined as the fractional cycle number at which the fluorescence passed the fixed threshold. Each sample was measured in triplicate, and the relative amount of miR-32 to U6 was calculated using the equation:
Relative amount of miR-32 = 2−ΔCt | (1) |
where
ΔCt = (CtmiR-32 – CtU6) | (2) |
MTT assay
After transfection, NSCLC cells were harvested, seeded into 96-well culture plates at a density of 2,000 cells in 200 μL/well, and incubated at 37°C. At different time points (24, 48, 72, or 96 hours), 100 μL of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) solution (0.5 mg/mL) (Sigma-Aldrich Corp, St Louis, MO, USA) was added to each well, and the plates were incubated for another 4 hours. Then, the MTT solution was removed, and 150 μL of dimethyl sulfoxide (DMSO) was added to each well to stop the reaction. The plates were gently shaken on a swing bed for 10 minutes, and spectrometric absorbance, at 490 nm, was measured using a microplate reader. This experiment was run in triplicate for each sample.
Detection of apoptosis by flow cytometry
Apoptosis was detected by flow cytometric analysis. Briefy, the cells were washed and resuspended at a concentration of 1×106 cells/mL. Then, the cells were stained with annexin V and propidium iodide using an annexin V apoptosis detection kit. After incubation at room temperature in the dark for 15 minutes, cell apoptosis was analyzed on a FACSCalibur™ (BD Biosciences, Franklin Lakes, NJ, USA).
Transwell® invasion assay
The invasion assay was performed using 24-well Transwell chambers (8 μm; Corning Inc, Corning, NY, USA). After transfection, tumor cells were resuspended in serum-free RPMI-1640 medium, and 2×105 cells were seeded into the upper chambers, which were covered with 1 mg/mL Matrigel®. Then, 0.5 mL of RPMI-1640 containing 10% FBS was added to the bottom chambers. Following a 24-hour incubation, the cells on the upper surface of the membrane were scrubbed off, and the invaded cells were fixed with 95% ethanol, stained with 0.1% crystal violet, and counted under a light microscope.
Scratch migration assays
Scratch migration assays were performed to observe the influence of miR-32 expression on NSCLC cell migration. When cells transfected with miR-32 mimics, miR-32 inhibitors, or negative controls were grown to confluence, the cell monolayer was scratched with a cell-scratch spatula. After the cells were incubated under standard conditions for 24 hours, images of the scratches were captured using a digital camera system coupled to a microscope.
Luciferase reporter assays
The pGL3-reporter luciferase vector was used to construct the pGL3-SOX9 or pGL3-SOX9-mut vectors. The pGL3-SOX9-mut vector was built with SOX9 that underwent site-directed mutagenesis of the miR-32 target site, using a Stratagene Quik-Change® Site-Directed Mutagenesis Kit (Stratagene; Agilent Technologies, Santa Clara, CA, USA). For the luciferase reporter assay, cells were cultured in 24-well plates and transfected with the plasmids and miR-32 mimics using Lipofectamine 2000. At 24 hours after transfection, luciferase activity was measured using the Dual Luciferase Reporter Assay System (Promega Corporation, Fitchburg, WI, USA). Firefly luciferase activity was normalized to Renilla luciferase activity for each transfected well.
Western blot analysis
Protein lysates were separated by 10% sodium dodecyl sulfate (SDS)-polyacrylamide gel electrophoresis and transferred to nitrocellulose membranes. After blocking, the membranes were incubated with purified rabbit anti-SOX9 antisera at 4°C overnight. The next day, the membranes were washed with phosphate-buffered saline (PBS) and then incubated with peroxidase-conjugated goat anti-rabbit IgG. Immunodetection was conducted with enhanced chemiluminescence (ECL) reagents (Pierce; Thermo Fisher Scientific Inc, Waltham, MA, USA) and exposed on an X-ray film. β-Actin was used as an internal reference for relative quantification.
Statistics
All statistical analyses were performed using the SPSS 16.0 software package (SPSS Inc., Chicago, IL, USA). The data were presented as the mean ± standard deviation (SD). Differences between groups were analyzed using Student’s t-test or the chi-square test. Relations between miR-32 expression and SOX9 protein levels were explored by Pearson correlation analysis. Survival curves were constructed using the Kaplan–Meier method and compared by log-rank tests. To evaluate independent prognostic factors associated with survival, a multivariate Cox proportional hazards regression analysis was used. P<0.05 was considered significant.
Results
Decreased expression of miR-32 in NSCLC and its correlation with SOX9 levels
The expression levels of miR-32 in NSCLC tissues, corresponding noncancerous samples, human NSCLC cell lines A549, H460, 95D, and HCC827, and normal lung epithelial cells were detected by qRT-PCR and normalized to U6 small nuclear RNA. As shown in Figure 1A, the results indicated that the expression levels of miR-32 were significantly lower in NSCLC specimens (8.2±1.9) compared with those in the corresponding adjacent noncancerous tissues (19.2±4.0) (P<0.001). The miR-32 expression levels were also clearly downregulated in the four NSCLC cell lines (Figure 1B). Because A549 cells exhibited the lowest miR-32 expression, whereas 95D cells expressed relatively high levels of miR-32 expression among the four NSCLC cell lines, these two cell lines were selected for mature miR-32 mimic or miR-32 inhibitor transfection and for further studies.
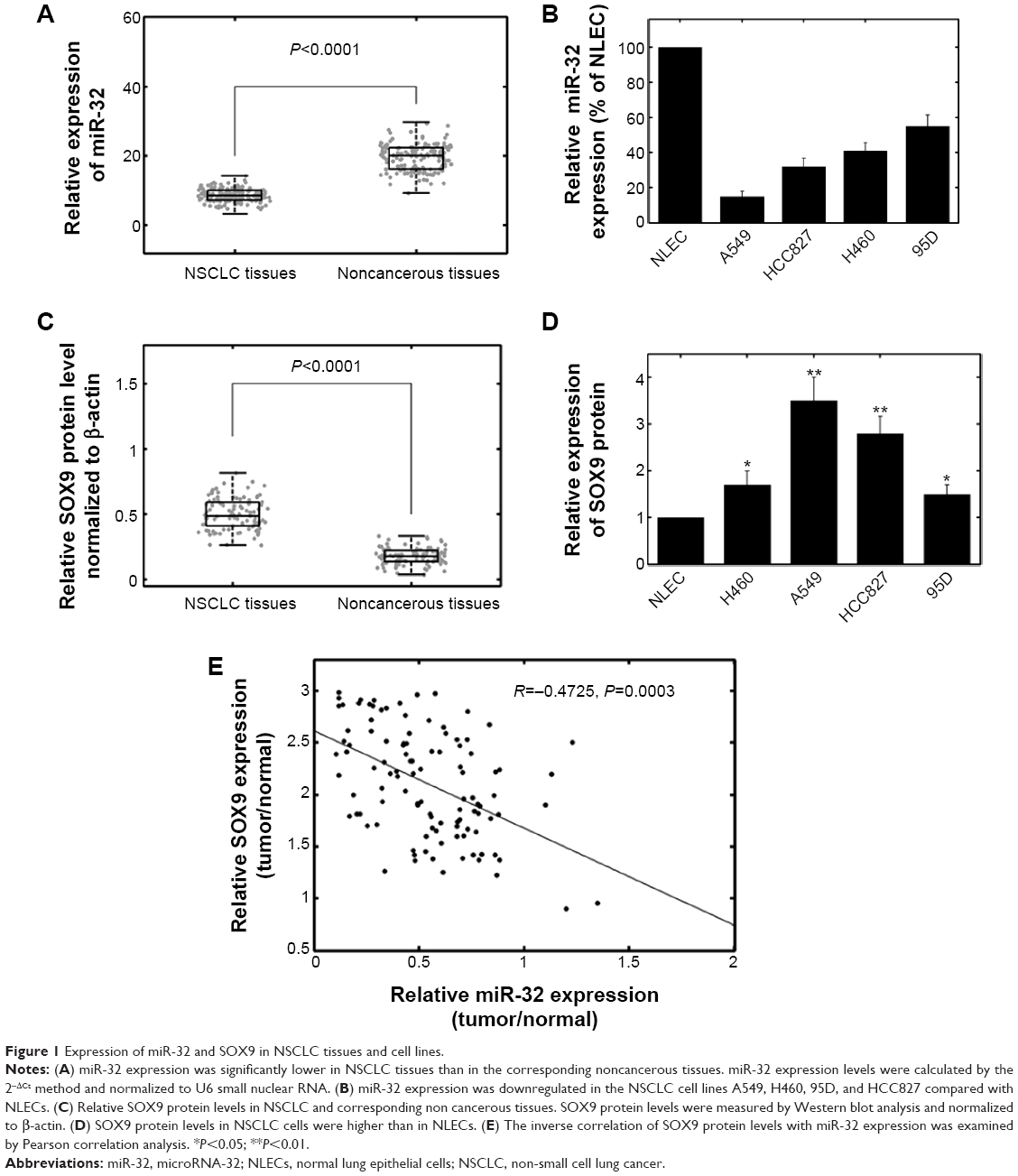 | Figure 1 Expression of miR-32 and SOX9 in NSCLC tissues and cell lines. |
SOX9 protein levels were detected by western blot analysis in clinical specimens and cell lines. The results showed that the SOX9 protein levels were higher in tumor samples than in the adjacent normal tissues (P<0.001) (Figure 1C). SOX9 protein levels were also higher in NSCLC cells than in normal lung epithelial cells (Figure 1D). In addition, we observed an obvious inverse correlation (R=-0.4725, P=0.0003) between SOX9 levels and miR-32 expression in NSCLC tumor tissues (Figure 1E).
miR-32 expression and clinicopathologic features in NSCLC
The associations of miR-32 expression with various clinicopathological parameters of NSCLC tissues are summarized in Table 1. Using the median miR-32 expression in all 115 NSCLC patients as a cutoff, the patients were divided into a high-miR-32-expression group and a low-miR-32-expression group. As shown in Table 1, the miR-32 expression levels were lower in samples with lymph node metastasis (P=0.002) and advanced tumor/nodes/metastasis (TNM) classification stage (P<0.001). No significant differences between miR-32 expression and patient age, sex, smoking status, cell types, tumor (T) stage, or tumor differentiation were observed.
Downregulation of miR-32 confers poor prognosis in NSCLC patients
Then, we evaluated whether miR-32 expression had prognostic potential for the OS of NSCLC patients. Using the Kaplan–Meier method and log-rank test, we found that the survival rate of patients with high miRNA-32 expression was higher than that of patients with low miRNA-32 expression (P<0.001) (Figure 2). In addition, the survival rates was also higher for those patients with negative nodes (N) classification (P=0.022) and early TNM stages (P<0.001) (Table 2).
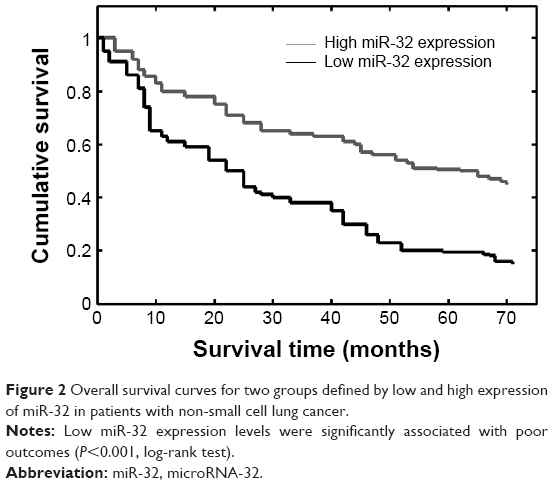 | Figure 2 Overall survival curves for two groups defined by low and high expression of miR-32 in patients with non-small cell lung cancer. |
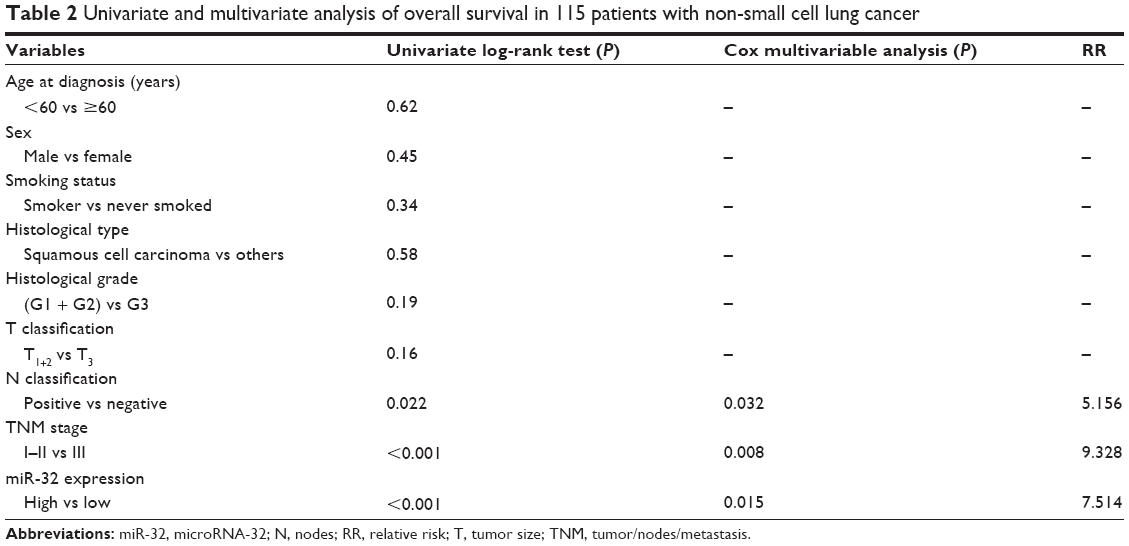 | Table 2 Univariate and multivariate analysis of overall survival in 115 patients with non-small cell lung cancer |
Multivariate Cox regression analysis using the aforementioned significant parameters revealed that miR-32 expression (relative risk [RR] 7.514; P=0.015), lymph node metastasis (RR 5.156; P=0.032), and TNM stage (RR 9.328; P=0.008) were independent prognostic markers for the OS of NSCLC patients (Table 2).
Effects of miR-32 expression on the biological behaviors of NSCLC cells
To selectively overexpress or downregulate miR-32, mature miR-32 mimics or miR-32 inhibitors were transfected into A549 or 95D cells, respectively. qRT-PCR analysis confirmed increased miR-32 expression after miR-32 mimic transfection and decreased miR-32 expression following miR-32 inhibitor transfection (Figure 3A). The MTT assay indicated that cell proliferation was significantly impaired in A549 cells transfected with miR-32 mimics, whereas the proliferation of 95D cells increased in miR-32 inhibitor-transfected cells compared with controls (Figure 3B).
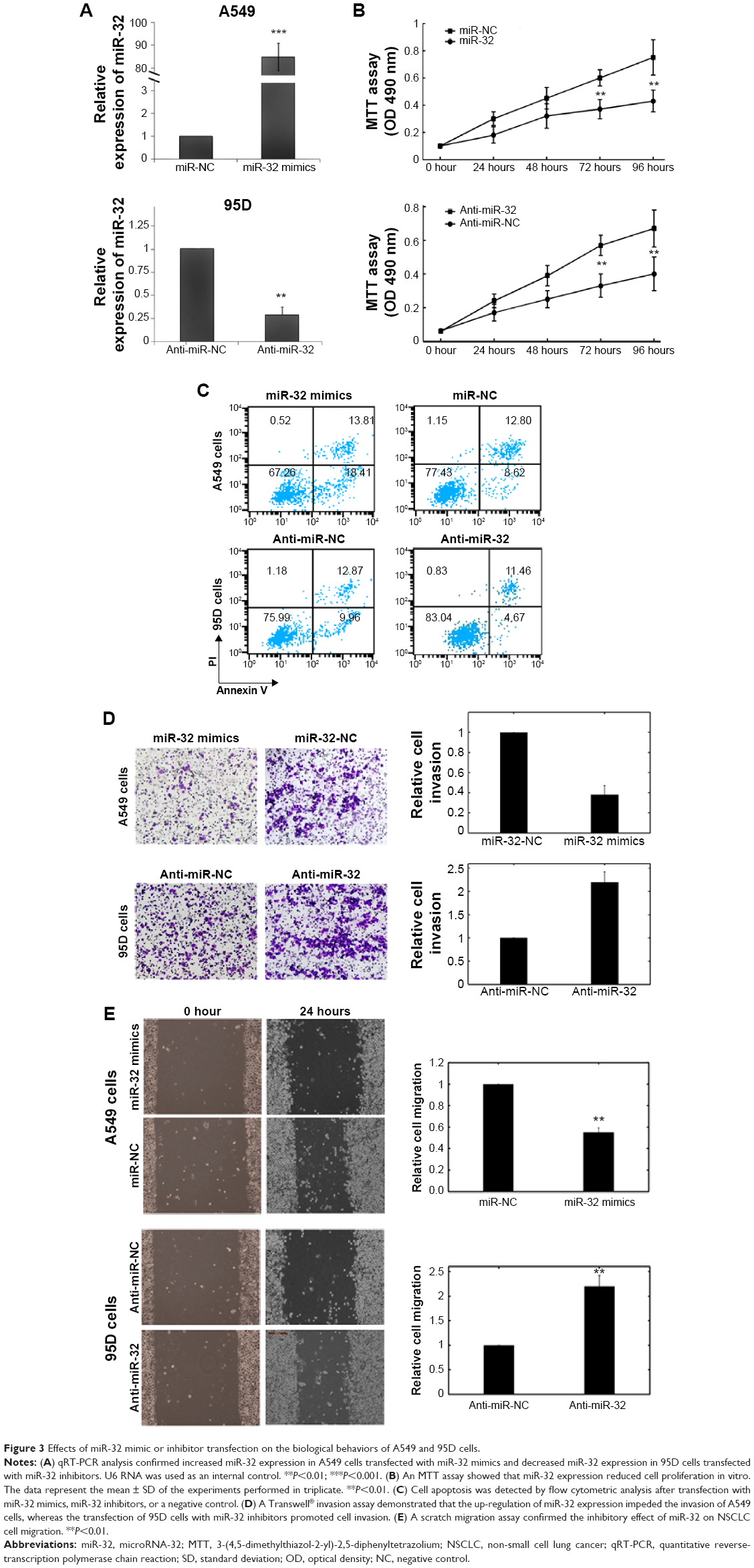 | Figure 3 Effects of miR-32 mimic or inhibitor transfection on the biological behaviors of A549 and 95D cells. |
Flow cytometry was employed to determine the effect of miR-32 on cell apoptosis. The proportion of apoptotic A549 cells transfected with miR-32 mimics was significantly higher than that of the negative control group. Moreover, miR-32 downregulation reduced 95D cell apoptosis (Figure 3C).
A Transwell invasion assay was performed to investigate whether miR-32 had a direct influence on NSCLC cell invasion. As shown in Figure 3D, the upregulation of miR-32 expression impeded the invasion of A549 cells compared with the control. Conversely, the transfection of 95D cells with miR-32 inhibitors promoted cell invasion. The scratch migration assay also confirmed the inhibitory effect of miR-32 expression on NSCLC cell migration (Figure 3E).
SOX9 is the target gene of miR-32
Using the bioinformatics software miRWalk for target gene prediction, SOX9 was identified as a potential target of miR-32. The predicted binding of miR-32 with the SOX9 3′ UTR is illustrated in Figure 4A. To further confirm that SOX9 is a direct target of miR-32 in NSCLC, first, we transfected miR-32 mimics into A549 cells and found that miR-32 mimics significantly reduced SOX9 protein levels in these cells (Figure 4B). Then, we created pGL3-SOX9 and pGL3-SOX9-mut plasmids. The results of the reporter assay revealed that the transfection of miR-32 mimics triggered a marked decrease in the luciferase activity of the pGL3-SOX9 plasmid in A549 cells, without changing the luciferase activity of pGL3-SOX9-mut (Figure 4C). These data indicated that SOX9 was a direct target of miR-32 in NSCLC.
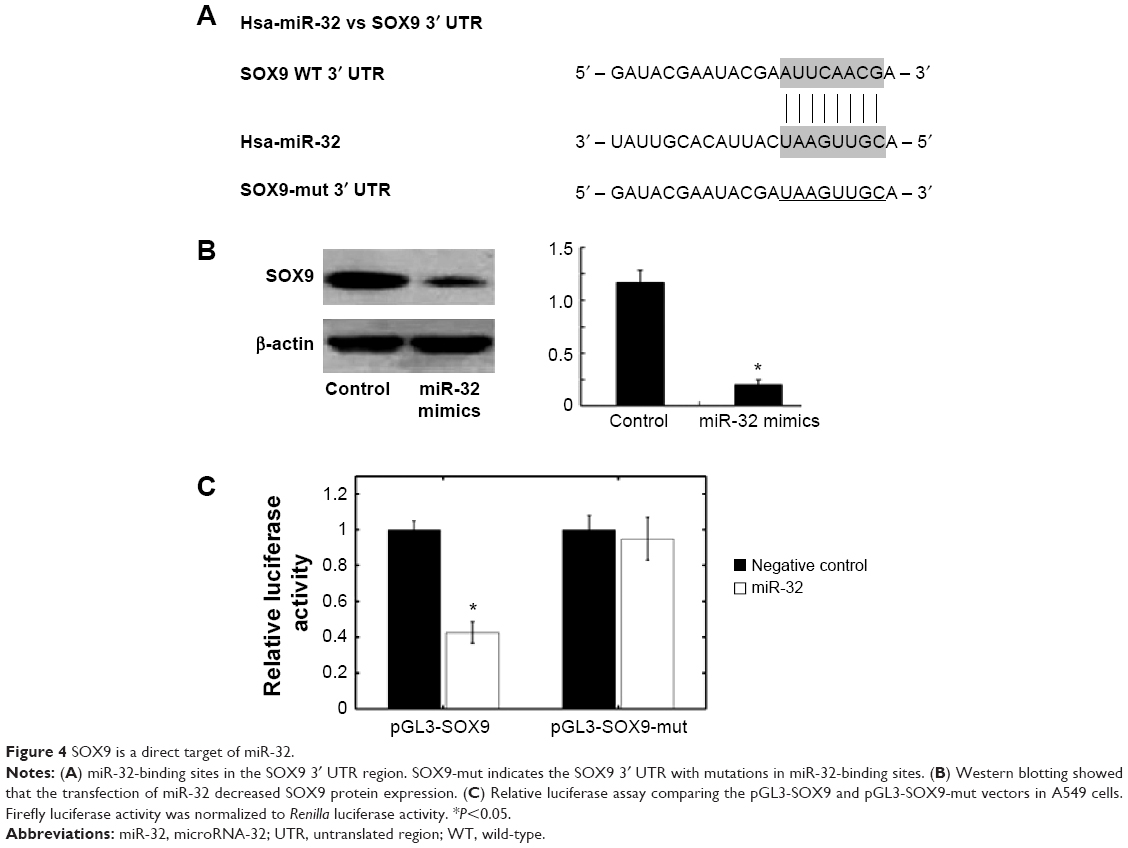 | Figure 4 SOX9 is a direct target of miR-32. |
Discussion
The dysregulation of miRs is involved in the tumorigenesis and progression of various types of tumors; however, their potential roles in NSCLC remain unclear. In the present study, we found that miR-32 was downregulated in NSCLC cell lines and in primary tumor samples. Decreased miR-32 expression significantly correlated with aggressive clinicopathological features and poor OS. Moreover, in vitro functional assays demonstrated that the modulation of miR-32 expression affected NSCLC cell proliferation, apoptosis, invasion, and migration. Finally, SOX9 was identified as a direct target of miR-32. To our knowledge, this study is the first to analyze the clinical significance and biological function of miR-32 in NSCLC.
The tumor-suppressor functions of miR-32, which is on chromosome band Xq26.2, have been shown in human osteosarcoma and gastric cancer. Xu et al found that miR-32 was significantly downregulated in osteosarcoma tissues compared with the adjacent normal tissues. In vitro studies demonstrated that miR-32 mimics suppressed proliferation and invasion, whereas its antisense oligonucleotides promoted the proliferation and invasion of Saos-2 and U2OS osteosarcoma cells.12 Zhang et al revealed that the upregulation of miR-32 expression significantly inhibited the proliferation capabilities and decreased the migration and invasion capabilities of SGC-7901 gastric cancer cells.11
In contrast to the aforementioned antitumor properties, miR-32 also acts as an oncogene in several cancers. miR-32 overexpression in colorectal carcinoma cells enhanced cell proliferation, migration, and invasion, and reduced cell apoptosis.7 High miR-32 levels in colorectal carcinoma patients were significantly associated with lymph-node and distant metastasis, as well as with poor OS.24 Increased miR-32 expression has also been reported in kidney cancer and prostate cancer,8,25 and miR-32 was shown to be androgen-regulated and overexpressed in castration-resistant prostate cancer.26 In addition, pretreatment with anti-miR-32 oligonucleotides sensitized acute myelogenous leukemia (AML) cells to arabinocytosine, a chemotherapeutic drug used to treat human AML, via the induction of cell apoptosis.27 Taken together, these findings indicate that the role of miR-32 in human malignancies may be multifaceted, depending on the involved tissue.
miRs clearly execute their oncogenic or tumor suppressor functions by regulating the expression of target genes.28 Several targets of miR-32 have been confirmed in recent research, including B-cell translocation gene 2 (BTG2),26 phosphatase and tensin homolog (PTEN),7 and tumor necrosis factor receptor-associated factor (TRAF).29 SOX9, which is a tumor-promoting gene that is upregulated in various tumor types, has been identified as a target gene of several miRs. Zhou et al corroborated high SOX9 expression in both NSCLC tumor tissues and cell lines.30 Further analysis indicated that the upregulation of SOX9 expression significantly correlated with advanced tumor stages and shorter OS times. Using a luciferase reporter assay, our study demonstrated that SOX9 is a direct target of miR-32 in NSCLC. However, a “one to one” connection between miRs and target mRNAs does not exist, as an average miR can have more than 100 targets.31 Conversely, several miRs can converge on a single transcript target.32 SOX9 is not the only miR-32 target dysregulated in NSCLC. Other functional targets of miR-32, such as PTEN and TRAF,33,34 also modulate NSCLC pathogenesis. Therefore, the potential regulatory circuitry afforded by miR-32 is enormous, and the actual mechanisms by which miR-32 influences NSCLC progression require further clarification.
In summary, our study revealed that the expression level of miR-32 significantly decreased in NSCLC and was associated with tumor development. Low miR-32 expression may imply a poor prognosis. The antitumor effects of miR-32 were also observed in the functional analysis, and SOX9 was confirmed as a direct target of miR-32. These findings suggest that miR-32 may act as a tumor suppressor in NSCLC and could be a potential candidate for miR-based therapy against NSCLC.
Disclosure
The authors report no conflicts of interest in this work.
References
Jemal A, Siegel R, Xu J, Ward E. Cancer statistics. CA Cancer J Clin. 2010;60(5):277–300. | ||
Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell. 2009;136(2):215–233. | ||
Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116(2):281–297. | ||
Heneghan HM, Miller N, Kerin MJ. MiRNAs as biomarkers and therapeutic targets in cancer. Curr Opin Pharmacol. 2010;10(5):543–550. | ||
Romero-Cordoba SL, Salido-Guadarrama I, Rodriguez-Dorantes M, Hidalgo-Miranda A. miRNA biogenesis: biological impact in the development of cancer. Cancer Biol Ther. 2014;15(11):1444–1455. | ||
Shin VY, Chu KM. MiRNA as potential biomarkers and therapeutic targets for gastric cancer. World J Gastroenterol. 2014;20(30): 10432–10439. | ||
Wu W, Yang J, Feng X, et al. MicroRNA-32 (miR-32) regulates phosphatase and tensin homologue (PTEN) expression and promotes growth, migration, and invasion in colorectal carcinoma cells. Mol Cancer. 2013;12:30. | ||
Petillo D, Kort EJ, Anema J, Furge KA, Yang XJ, Teh BT. MicroRNA profiling of human kidney cancer subtypes. Int J Oncol. 2009;35(1):109–114. | ||
Ambs S, Prueitt RL, Yi M, et al. Genomic profiling of microRNA and messenger RNA reveals deregulated microRNA expression in prostate cancer. Cancer Res. 2008;68(15):6162–6170. | ||
Pichiorri F, Suh SS, Ladetto M, et al. MicroRNAs regulate critical genes associated with multiple myeloma pathogenesis. Proc Natl Acad Sci U S A. 2008;105(35):12885–12890. | ||
Zhang J, Kuai X, Song M, et al. microRNA-32 inhibits the proliferation and invasion of the SGC-7901 gastric cancer cell line in vitro. Oncol Lett. 2014;7(1):270–274. | ||
Xu JQ, Zhang WB, Wan R, Yang YQ. MicroRNA-32 inhibits osteosarcoma cell proliferation and invasion by targeting Sox9. Tumour Biol. 2014;35(10):9847–9853. | ||
Yanaihara N, Caplen N, Bowman E, et al. Unique microRNA molecular profiles in lung cancer diagnosis and prognosis. Cancer Cell. 2006;9(3):189–198. | ||
Chaboissier MC, Kobayashi A, Vidal VI, et al. Functional analysis of Sox8 and Sox9 during sex determination in the mouse. Development. 2004;131(9):1891–1901. | ||
Akiyama H, Chaboissier MC, Martin JF, Schedl A, de Crombrugghe B. The transcription factor SOX9 has essential roles in successive steps of the chondrocyte differentiation pathway and is required for expression of SOX5 and SOX6. Genes Dev. 2002;16(21):2813–2828. | ||
Matheu A, Collado M, Wise C, et al. Oncogenicity of the developmental transcription factor SOX9. Cancer Res. 2012;72(5):1301–1315. | ||
Müller P, Crofts JD, Newman BS, et al. SOX9 mediates the retinoic acid-induced HES-1 gene expression in human breast cancer cells. Breast Cancer Res Treat. 2010;120(2):317–326. | ||
Bruun J, Kolberg M, Nesland JM, Svindland A, Nesbakken A, Lothe RA. Prognostic significance of β-catenin, E-cadherin, and SOX9 in colorectal cancer: Results from a large population-representative series. Front Oncol. 2014;4:118. | ||
Qin GQ, He HC, Han ZD, et al. Combined overexpression of HIVEP3 and SOX9 predicts unfavorable biochemical recurrence-free survival in patients with prostate cancer. Onco Targets Ther. 2014;7:137–146. | ||
Jiang SS, Fang WT, Hou YH, et al. Upregulation of SOX9 in lung adenocarcinoma and its involvement in the regulation of cell growth and tumorigenicity. Clin Cancer Res. 2010;16(17):4363–4373. | ||
Real FM, Sekido R, Lupiáñez DG, Lovell-Badge R, Jiménez R, Burgos M. A microRNA (mmu-miR-124) prevents Sox9 expression in developing mouse ovarian cells. Biol Reprod. 2013;89(4):78. | ||
Rani SB, Rathod SS, Karthik S, Kaur N, Muzumdar D, Shiras AS. miR-145 functions as a tumor-suppressive RNA by targeting Sox9 and adducin 3 in human glioma cells. Neuro Oncol. 2013;15(10):1302–1316. | ||
Martinez-Sanchez A, Murphy CL. miR-1247 functions by targeting cartilage transcription factor SOX9. J Biol Chem. 2013;288(43):30802–30814. | ||
Wu W, Yang P, Feng X, et al. The relationship between and clinical significance of MicroRNA-32 and phosphatase and tensin homologue expression in colorectal cancer. Genes Chromosomes Cancer. 2013;52(12):1133–1140. | ||
Leite KR, Tomiyama A, Reis ST, et al. MicroRNA expression profiles in the progression of prostate cancer – from high-grade prostate intraepithelial neoplasia to metastasis. Urol Oncol. 2013;31(6):796–801. | ||
Jalava SE, Urbanucci A, Latonen L, et al. Androgen-regulated miR-32 targets BTG2 and is overexpressed in castration-resistant prostate cancer. Oncogene. 2012;31(41):4460–4471. | ||
Gocek E, Wang X, Liu X, Liu CG, Studzinski GP. MicroRNA-32 upregulation by 1,25-dihydroxyvitamin D3 in human myeloid leukemia cells leads to Bim targeting and inhibition of AraC-induced apoptosis. Cancer Res. 2011;71(19):6230–6239. | ||
Liu GF, Tang D, Li P, et al. S-1-based combination therapy vs S-1 monotherapy in advanced gastric cancer: a meta-analysis. World J Gastroenterol. 2014;20(1):310–318. | ||
Mishra R, Chhatbar C, Singh SK. HIV-1 Tat C-mediated regulation of tumor necrosis factor receptor-associated factor-3 by microRNA 32 in human microglia. J Neuroinflammation. 2012;9:131. | ||
Zhou CH, Ye LP, Ye SX, et al. Clinical significance of SOX9 in human non-small cell lung cancer progression and overall patient survival. J Exp Clin Cancer Res. 2012;31:18. | ||
Brennecke J, Stark A, Russell RB, Cohen SM. Principles of microRNA-target recognition. PLoS Biol. 2005;3(3):e85. | ||
Krek A, Grün D, Poy MN, et al. Combinatorial microRNA target predictions. Nat Genet. 2005;37(5):495–500. | ||
Zheng M, Morgan-Lappe SE, Yang J, et al. Growth inhibition and radiosensitization of glioblastoma and lung cancer cells by small interfering RNA silencing of tumor necrosis factor receptor-associated factor 2. Cancer Res. 2008;68(18):7570–7578. | ||
Liu ZL, Wang H, Liu J, Wang ZX. MicroRNA-21 (miR-21) expression promotes growth, metastasis, and chemo- or radioresistance in non-small cell lung cancer cells by targeting PTEN. Mol Cell Biochem. 2013;372(1–2):35–45. |
 © 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.