Back to Journals » OncoTargets and Therapy » Volume 13
miR-27b Suppresses Tongue Squamous Cell Carcinoma Epithelial–Mesenchymal Transition by Targeting ITGA5
Authors Li T, Wu Q, Liu D, Wang X
Received 18 September 2020
Accepted for publication 31 October 2020
Published 17 November 2020 Volume 2020:13 Pages 11855—11867
DOI https://doi.org/10.2147/OTT.S281211
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Arseniy Yuzhalin
Tao Li,1 Qian Wu,2 Duanqin Liu,1 Xuxia Wang1
1Department of Oral and Maxillofacial Surgery, School and Hospital of Stomatology, Cheeloo College of Medicine, Shandong University & Shandong Key Laboratory of Oral Tissue Regeneration & Shandong Engineering Laboratory for Dental Materials and Oral Tissue Regeneration, Jinan, Shandong, People’s Republic of China; 2Department of Nursing, School and Hospital of Stomatology, Cheeloo College of Medicine, Shandong University, Jinan, Shandong, People’s Republic of China
Correspondence: Xuxia Wang
Department of Oral and Maxillofacial Surgery, School and Hospital of Stomatology, Cheeloo College of Medicine, Shandong University & Shandong Key Laboratory of Oral Tissue Regeneration & Shandong Engineering Laboratory for Dental Materials and Oral Tissue Regeneration, No. 44-1 Wenhua Road West, Jinan, Shandong 250012, People’s Republic of China
Fax +86 53188382923
Email [email protected]
Background: MicroRNA27b-3p (miR-27b) has been reported to be dysregulated in multiple types of human cancer. However, the expression levels, biological roles, and underlying mechanism of miR-27b in tongue squamous cell carcinoma (TSCC) remain to be elucidated.
Methods: Bioinformatics analyses and quantitative real-time PCR (qRT-PCR) were used to determine miR-27b expression in TSCC tissues and cell lines. The influence of miR-27b overexpression or inhibition on TSCC cell proliferation, migration, and invasion in vitro, and on tumor growth in vivo, was explored via CCK8, colony formation, wound healing, and transwell assays, and in xenograft tumors in nude mice, respectively. Luciferase reporter assays, qRT-PCR, and Western blotting were performed to clarify the potential mechanisms involving miR-27b in TSCC cells.
Results: miR-27b was significantly downregulated in TSCC tissues and cell lines, and its expression was correlated with cancer status. Overexpression of miR-27b led to diminished proliferation, migration, and invasion, and notably reduced tumor growth in vivo. Bioinformatics analysis followed by luciferase reporter assays demonstrated that miR-27b expression was inversely correlated with that of integrin subunit α 5 (ITGA5)and that miR-27b directly bound to the 3ʹ-untranslated region of ITGA5 in TSCC cells. The bioinformatics analysis also indicated that ITGA5 was upregulated in TSCC and that its expression was correlated with epithelial–mesenchymal transition (EMT) and poor prognosis. Moreover, we found that miRNA-27b could reverse ITGA5-induced promotion of TSCC cell proliferation and migration. Finally, we demonstrated that regulation of miR-27b expression in TSCC may result in alterations in the expression of ITGA5 and EMT-related marker genes at the mRNA and protein levels.
Conclusion: These results indicate that miR-27b hampers TSCC proliferation and migration via suppressing the EMT process by targeting ITGA5. These findings support consideration of miR-27b/ITGA5 as a valuable marker for the metastatic potential of TSCC, or as a therapeutic target for the treatment of TSCC.
Keywords: miR-27b, epithelial–mesenchymal transition, integrin subunit alpha 5, bioinformatics analysis, tongue squamous cell carcinoma
Introduction
Head and neck squamous cell carcinoma (HNSCC) is a common malignancy worldwide. Its 5-year survival rate has not significantly improved for decades and remains at about 63%.1 Reasons for this include delayed disease detection and intervention, and a dearth of therapeutic options for advanced HNSCC. Tongue squamous cell carcinoma (TSCC), a major subtype of HNSCC, is common in southern Asia. It is also estimated that there were approximately 17,110 new TSCC cases in 2018 in the United States alone.2 The main risk factors for TSCC are largely related to lifestyle, including tobacco and alcohol use and diet. Viral infections and lack of vitamins and trace elements are also important etiological factors.3
MicroRNAs (miRNAs) are small endogenous non-coding RNAs of 18–25 nucleotides in length, which may target and regulate the expression of mRNA at the post-transcriptional level through binding to the 3ʹ-untranslated region (3ʹ-UTR).4,5 A large number of studies have proved that miRNAs can serve as therapeutic targets and diagnostic biomarkers. For example, mechanistic studies have demonstrated that miR-205 inhibits metastatic traits of triple-negative breast cancer cells and tumor metastasis by downregulating integrin subunit α5 (ITGA5).6 miR-27b has been reported to serve as an oncogene or a tumor suppressor during tumor progression, depending on the different cellular contexts of tumors.7,8 ITGA5 has been predicted to be a possible target gene of miR-27b based on multiple databases. However, the potential mechanisms by which miR-27b may regulate ITGA5 expression in patients with TSCC have not yet been fully investigated.
Changes in the genome and tumor microenvironment are the essential alterations that lead to the malignant progression of TSCC.9 Integrins are transmembrane proteins on the cell surface that mediate communications among different cells, or between cells and the extracellular matrix (ECM). They form heterodimeric complexes composed of α and β subunits that can sense changes in the ECM and trigger a range of cellular responses by forming a physical connection between the inside and outside of a cell.10 The ITGA5 gene is located on chromosome 12q11-q13 and encodes a fibronectin receptor protein, ITGA5. A member of the integrin family, ITGA5 often combines with ITGB1 to form integrin α5β1, which serves as a receptor for cell differentiation, cell development, and migration.11 Altered expression of ITGA5 has been demonstrated in certain cancers, including colorectal, breast, and oral cancers.12–14
In this study, we confirmed that miR-27b was significantly downregulated while ITGA5 was significantly upregulated in TSCC. Then, we showed that miR-27b inhibited the proliferation, migration, and invasion of TSCC cells in vitro and suppressed tumor growth in vivo. Further mechanistic studies revealed that miR-27b suppressed EMT by targeting ITGA5.
Materials and Methods
TSCC Data Sets for Bioinformatic Analysis
In the present study, RNA sequencing and miRNA sequencing data for TSCC (primary site: other and unspecified parts of tongue; 126 tumor cases and 13 normal controls; workflow type: HTSeq counts) and corresponding clinical information were downloaded from the official website of The Cancer Genome Atlas (TCGA) (https://portal.gdc.cancer.gov/). The data obtained from TCGA were used to analyze the differences in ITGA5 expression between TSCC and normal tissues, as well as for survival analysis, gene set enrichment analysis (GSEA), and weighted gene correlation network analysis (WGCNA). The expression pattern of miR-27b in TSCC was also investigated using the TCGA data. Non-coding RNA profiling array data (GSE137865, GSE124566) were obtained from the Gene Expression Omnibus (GEO, https://www.ncbi.nlm.nih.gov/geo/). The GSE137865 data set comprised a total of 18 samples, including paired TSCC tissues, tissues adjacent to cancer, and contralateral normal tissues. The GSE124566 dataset consisted of 10 paired normal and TSCC samples.
Databases and R Packages
The TargetScan (http://www.targetscan.org/vert_72/), MiRanda (http://www.microrna.org/microrna/home.do), and miRDB (http://www.mirdb.org/) databases were used to predict that miR-27b might be a target of ITGA5. The expression status of ITGA5 in TSCC was validated using the Oncomine (https://www.oncomine.org/) and Human Protein Atlas (www.proteinatlas.org) databases. The co-expression relationship of ITGA5 and miR-27b in TSCC was explored using ENCORI (http://starbase.sysu.edu.cn/index.php). Survival analysis for ITGA5 in TSCC patients used the “survival” and “survminer” R packages. GSEA was performed with software from the GSEA database (https://www.gsea-msigdb.org/gsea/index.jsp), using the expression level of ITGA5 as a phenotype label to reveal significant differences (false discovery rate <0.01, P<0.05) in enrichment in the MSigDB Collection (c2.cp.biocarta and hall. v6.1.symbols). WGCNA was performed using the “WGCNA” R package following the instructions at https://horvath.genetics.ucla.edu/html/Coexpression-Network/Rpackages/WGCNA/.
Cell Culture
Normal gingival epithelial cells and human TSCC cell lines (CAL 27, SCC-4, and SCC-9) were obtained from the American Type Culture Collection (Manassas, VA, USA). Normal gingival epithelial cells were cultured in minimum essential media supplemented with 10% fetal bovine serum (FBS; Invitrogen, Carlsbad, CA, USA) and 1% penicillin–streptomycin (Gibco, Thermo Fisher Scientific, Inc., Waltham, MA, USA). CAL 27 cells were cultured in Dulbecco’s modified Eagle medium (DMEM, Sigma-Aldrich, St. Louis, MO, USA) with 10% FBS and 1% penicillin–streptomycin, and SCC-9 and SCC-4 cells were cultured in DMEM F-12 medium containing 10% FBS, 1% penicillin–streptomycin, and 400 ng/mL hydrocortisone at 37°C in a humidified tissue culture incubator with 5% CO2. All cells were HPV free.
Transfection
The miR-27b mimic and inhibitor and a negative control for the mimic (mimic-NC) were purchased from GenePharma (Shanghai, China). The ITGA5 recombinant plasmid and negative control plasmid were obtained from Origene (Suzhou, China). For transfection, cells were plated in six-well plates with antibiotic-free culture medium at a density of 7×105 cells per well. The next day, cells were transfected with the appropriate miRNA or plasmid using Lipofectamine 2000 reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. Cells in the mock group were incubated in the presence of transfection reagents only, whereas the miR-NC group cells were transfected with mimic-NC.
RNA Extraction and Quantitative Real-Time PCR (qRT-PCR)
Total RNA of cells was isolated and purified using a miRNeasy Mini Kit (Qiagen, Maryland, USA) according to the manufacturer’s instructions. Reverse transcription was performed using a PrimeScript RT reagent kit (Takara, Otsu, Japan), and qRT-PCR was performed using SYBR Green dye (TaKaRa, Japan). The 2−ΔΔCq method was used to analyze the relative quantitative data, which were normalized to glyceraldehyde 3-phosphate dehydrogenase (GAPDH). The primers are shown in Table 1.
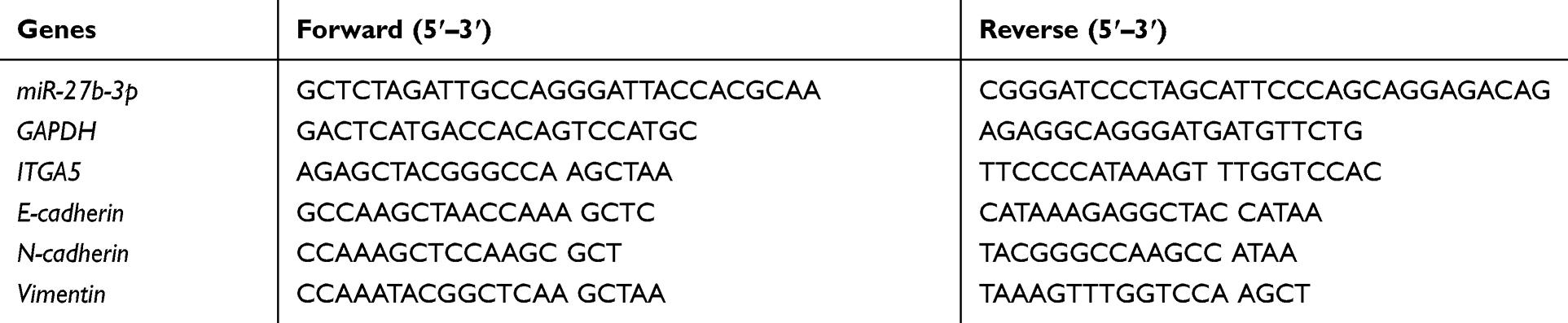 |
Table 1 Primers Used for Real‐time PCR |
Luciferase Reporter Assay
TargetScan was used to predict that ITGA5 was a target gene of miR-27b. The predicted binding sequences on the 3ʹ-UTR of ITGA5 mRNA were mutated from 5ʹ-CACUGUGA-3ʹ to 5ʹ-GAGAGUCT-3ʹ in SCC-9 cells. Subsequently, either the wild type or the mutant 3ʹ-UTR of ITGA5 mRNA was inserted into the dual-luciferase reporter vectors, which were designated WT or MUT, respectively. For the luciferase assay, Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) was used to co-transfect the miR-27b mimic and the WT or MUT plasmid into SCC-9 cells. Next, luciferase activity was measured using a Dual-Luciferase Reporter Assay System (Promega, Madison, WI, USA).
Cell Counting Kit-8 (CCK-8) Assay
Cell proliferation ability was ascertained using CCK-8 assays. In brief, 2000 cells were incubated in 96-well plates, and the absorbance was evaluated after culture for 6, 24, 48, 72, or 96 h using CCK-8 (Dojindo, Kuma-moto, Japan) with a Model 680 Microplate Reader (Bio-Rad, Hercules, CA, USA). The optical density value in each well was determined at a wavelength of 450 nm.
Xenograft Tumor Formation Assay
Six-week-old female nude mice were purchased from Charles River Laboratories (Beijing, China) and divided randomly into a miR-NC and a mimic group. Transfected SSC-9 cells (1×106 cells suspended in 100 μL serum-free Ham’s F12 nutrient medium) were injected subcutaneously on both sides of the backs of nude mice to establish the xenotransplantation model. The experiment was terminated after 4 weeks of culture, the mice were sacrificed, and the xenografts were dissected out. The weights of the xenograft tumors were recorded. All animal experiments were approved by the Animal Care and Use Committee of Shandong Key Laboratory of Oral Tissue Regeneration (reference number: GR2018017) and carried out in accordance with the guidance of Animal Protection Law of the People’s Republic of China-2018 for experimental animals.
Wound Healing Assay
Transfected cells were seeded into six-well plates at a density of 1 × 105 cells/well in a medium containing 10% FBS. After the cells reached 100% density, a 20 μL pipette was used to scratch the surface to make a wound. The cells were then washed twice with phosphate-buffered saline to remove cell debris and incubated for 48 h in fresh medium. Wounds were analyzed using the ImageJ software.
Transwell Assays
Migration and invasion were assessed using transwell plates. For the invasion assay, Matrigel-coated transwell chambers (8 μm pore size, 24 wells) were used: 5×104 cells were resuspended in 250 μL plain medium without FBS in the upper chamber, while the lower chamber was filled with 700 µL complete medium with 10% FBS. After incubation for 2 days, the upper chamber was fixed with 100% methanol and stained with 0.1% crystal violet. For the migration assay, 5×104 cells were plated on the chambers, and transmembrane cells were estimated under a microscope (Nikon, Tokyo, Japan) at 100× magnification.
Colony Formation Assay
After 24 h of transfection, CAL 27 cells were initially seeded into six-well plates at a density of 1000 cells per well and maintained in a medium containing 10% FBS, which was refreshed every 2 days. After incubation for approximately 2 weeks at 37°C in 5% CO2, the cells were fixed with 4% paraformaldehyde for 15 min and stained with 0.1% crystal violet for 15 min.
Western Blotting
Cells were digested with 0.25% trypsin and collected by centrifugation (1000 g), then lysed in RIPA buffer for 30 min. Equal amounts of protein extract were separated via 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to polyvinylidene fluoride membranes (Millipore, Billerica, MA, USA). After membrane blocking, primary antibodies (ITGA5: ab226816, N-cadherin: ab18203, E-cadherin: ab15148, vimentin: ab92547, GAPDH: ab181602) were added, followed by overnight incubation at 4°C. Finally, the membranes were incubated with the respective horseradish peroxidase-conjugated secondary antibodies for 1 h. Protein bands of interest were visualized using enhanced chemiluminescence reagents (Millipore, Burlington, MA, USA).
Statistical Analysis
All data are presented as mean±standard deviation from at least three independent experiments. SPSS 16.0 (SPSS Inc., Chicago, IL, USA) was used to analyze the data. Student’s t-tests or one-way analysis of variance was used to compare differences between two or more groups. The association between ITGA5 expression and overall survival of TSCC patients was assessed using a Kaplan–Meier curve and Log rank test. Results were considered to be statistically significant if the p-value did not exceed 0.05.
Results
miR-27b is Downregulated in TSCC and Correlated with TSCC
miR-27b expression levels were downregulated in cancer tissues compared with paired normal controls in the TCGA-TSCC samples (Figure 1A). The same expression pattern was observed in the TSCC samples from the GSE124566 and GSE137865 data sets (Figure 1B–C). miR-27b expression was then detected in three human TSCC cell lines (SCC-4, SCC-9, and CAL 27) and normal oral epithelial cells. The qRT-PCR results showed that miR-27b expression was significantly lower in SCC-9 and CAL 27 cells compared with normal oral epithelial cells (Figure 1D). WGCNA was used to show that miR-27b was one of the non-coding genes in the blue module that was significantly related to the status of TSCC (Figure 1E–F).
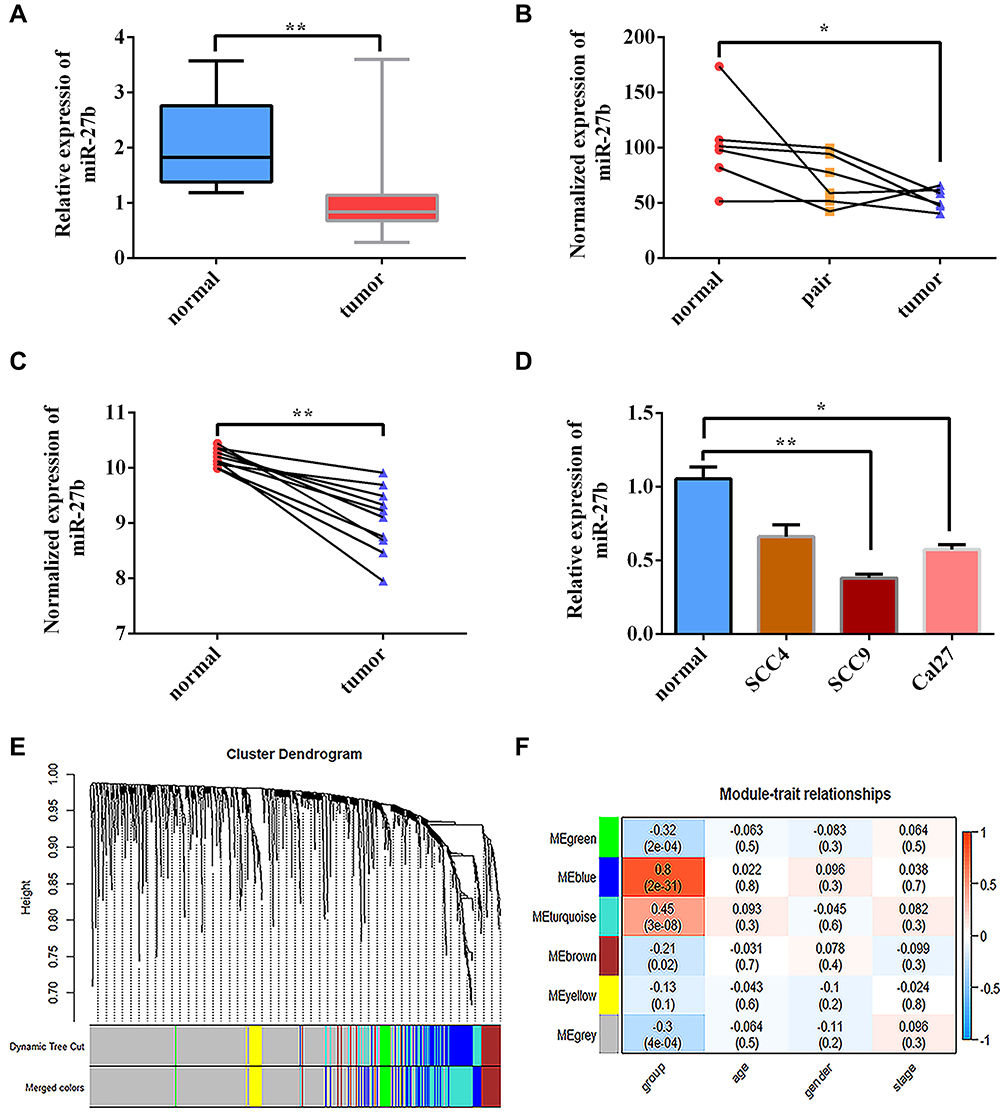 |
Figure 1 miR-27b expression is downregulated in TSCC cancer tissues and associated with TSCC cancer status. (A) Expression of miR-27b was lower in TCGA-TSCC samples compared with normal controls. (B, C) Expression of miR-27b was lower in TSCC cancer samples compared with normal controls based on GEO data sets GSE137865 and GSE124566. (D) Relative expression of miR-27b in TSCC cell lines was significantly reduced compared with that in normal gingival epithelial cells according to qRT-PCR. (E) Identification of gene co-expression modules in TSCC via hierarchical average linkage clustering. (F) Heatmap of the correlations between module eigengenes and phenotypic characteristics of TSCC. *Represents P<0.05, **Represents P<0.01. |
miR-27b Suppresses the Proliferation, Migration, and Invasion of TSCC in vitro
To explore the effects of miR-27b on the proliferation of TSCC, transfection of a miR-27b mimic or miR-27b inhibitor was used to up- or downregulate miR-27b expression in TSCC cell line SCC-9 (Figure 2A). The CCK8 assay showed that the proliferation of SCC-9 cells was inhibited by the miR-27b mimic and promoted by the miR-27b inhibitor compared with the mock group cells (Figure 2B). Next, we investigated the effects of miR-27b on TSCC migration and invasion. The results of the wound healing assay showed that CAL 27 cell migration was inhibited by the miR-27b mimic and promoted by the miR-27b inhibitor (Figure 2C–D). Cell migration and invasion ability were detected by transwell assays. The results showed that the miR-27b mimic significantly reduced the numbers of migrating and invading CAL 27 cells, whereas the miR-27b inhibitor had the converse effect (Figure 2E–G). These results indicate that miR-27b can inhibit proliferation, migration, and invasion of TSCC cells.
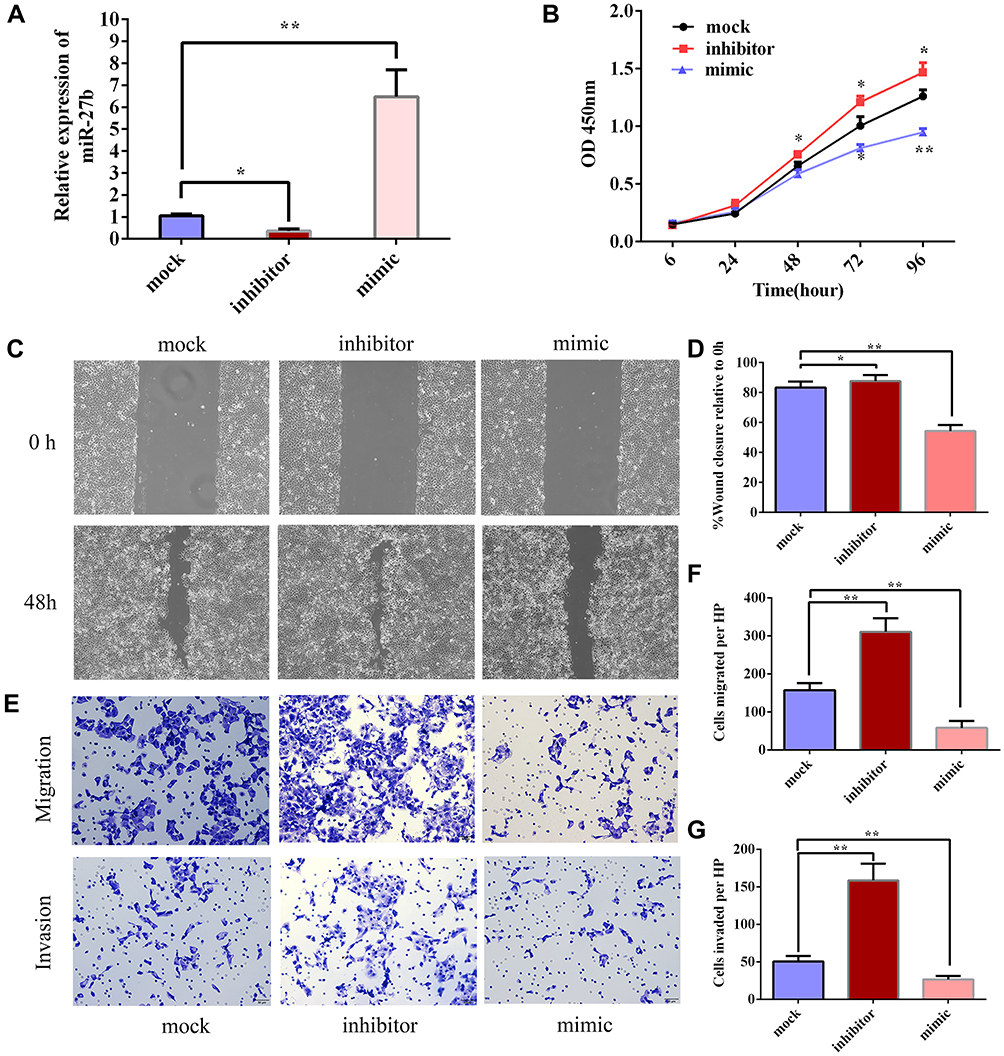 |
Figure 2 miR-27b suppresses the proliferation, migration, and invasion of TSCC in vitro. (A) miR-27b mimic or inhibitor was transfected into SCC-9 cells to up- or downregulate miR-27b, respectively. (B) CCK8 assay revealed that miR-27b inhibited proliferation of SCC-9 cells. (C, D) Wound healing assay was performed in CAL 27 cells after transfection with miR-27b NC, mimic, and inhibitor. (E–G) Transwell assay was performed in CAL 27 after transfection with miR-27b NC, mimic, and inhibitor. *Represents P<0.05, **Represents P<0.01. |
ITGA5 is a Direct Target of miR-27b in TSCC Cells
ITGA5 was predicted to be targeted by miR-27b by TargetScan, MiRanda, and miRDB, with the predicted binding site located at positions 190–197 on the 3ʹ-UTR of the ITGA5 mRNA (Figure 3A). To validate the binding of miR-27b to this site, the binding sequence was mutated from 5ʹ-CACUGUGA-ʹ3 to 5ʹ-GAGAGUCT-3ʹ and then inserted into luciferase vectors. Compared with miR-NC, the miR-27b mimic reduced the luciferase activity of SCC-9 cells transfected with the WT ITGA5 3ʹ-UTR; however, the luciferase activity of cells transfected with the MUT ITGA5 3ʹ-UTR showed no significant alteration (Figure 3B). The bioinformatics analysis indicated that there was a negative correlation between the expression levels of ITGA5 and miR-27b in TCGA-HNSC (Figure 3C).
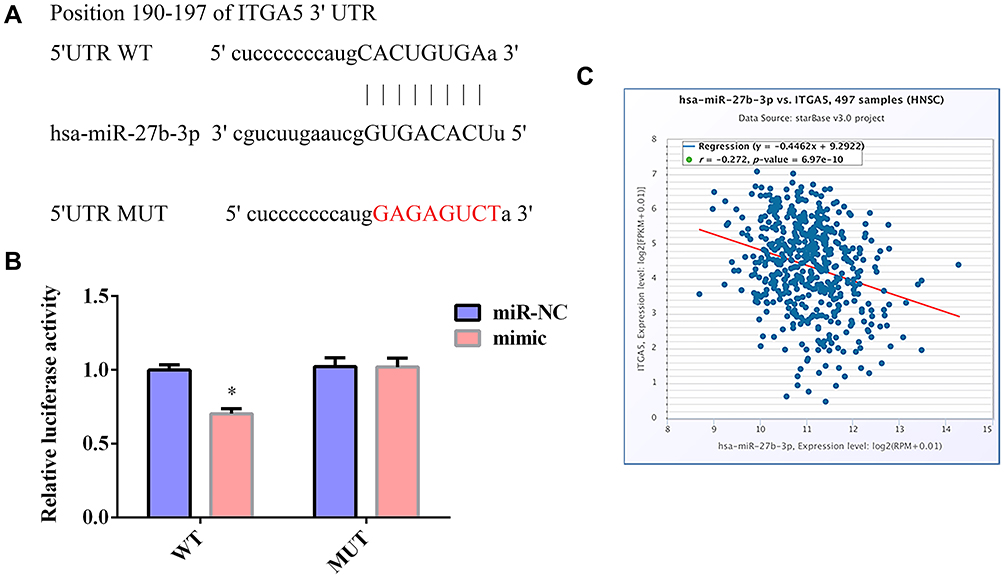 |
Figure 3 ITGA5 is the target of miR-27b in TSCC cells. (A) Binding sites between ITGA5 and miR-27b predicted by TargetScan. (B) Luciferase reporter assay indicated that miR-27b targeted the 3ʹ-UTR of ITGA5 mRNA in SCC-9 cells. WT-ITGA5 or MUT-ITGA5 was cotransfected into SCC-9 with miR-27b mimic or the corresponding negative control. (C) Expression levels of ITGA5 and miR-27b were negatively correlated in TCGA-HNSC samples. *Represents P<0.05. |
ITGA5 is Upregulated in TSCC and Correlated with Poor Prognosis and EMT
As shown in Table 2, data for 126 primary tumors, including both clinical information and gene expression data, were downloaded from TCGA. ITGA5 showed significantly higher expression in cancer tissues than in normal tissues (Figure 4A). Elevated expression of ITGA5 was significantly correlated with tumor clinical stage (stage Ⅱ vs stage Ⅳ) and metastasis (Figure 4B–C). We used Oncomine to validate these results (Figure 4D). Kaplan–Meier survival analysis showed that TSCC with high ITGA5 expression had a poorer prognosis than that with low ITGA5 expression (Figure 4E). The GSEA results indicated that the epithelial–mesenchymal transition (EMT) gene set was enriched in the ITGA5 high expression phenotype (Figure 4F). Moreover, immunohistochemistry staining results obtained from The Human Protein Atlas showed that the expression of ITGA5 at the protein level was higher in TSCC tumor tissues than in normal tongue mucosa (Figure 4G–H).
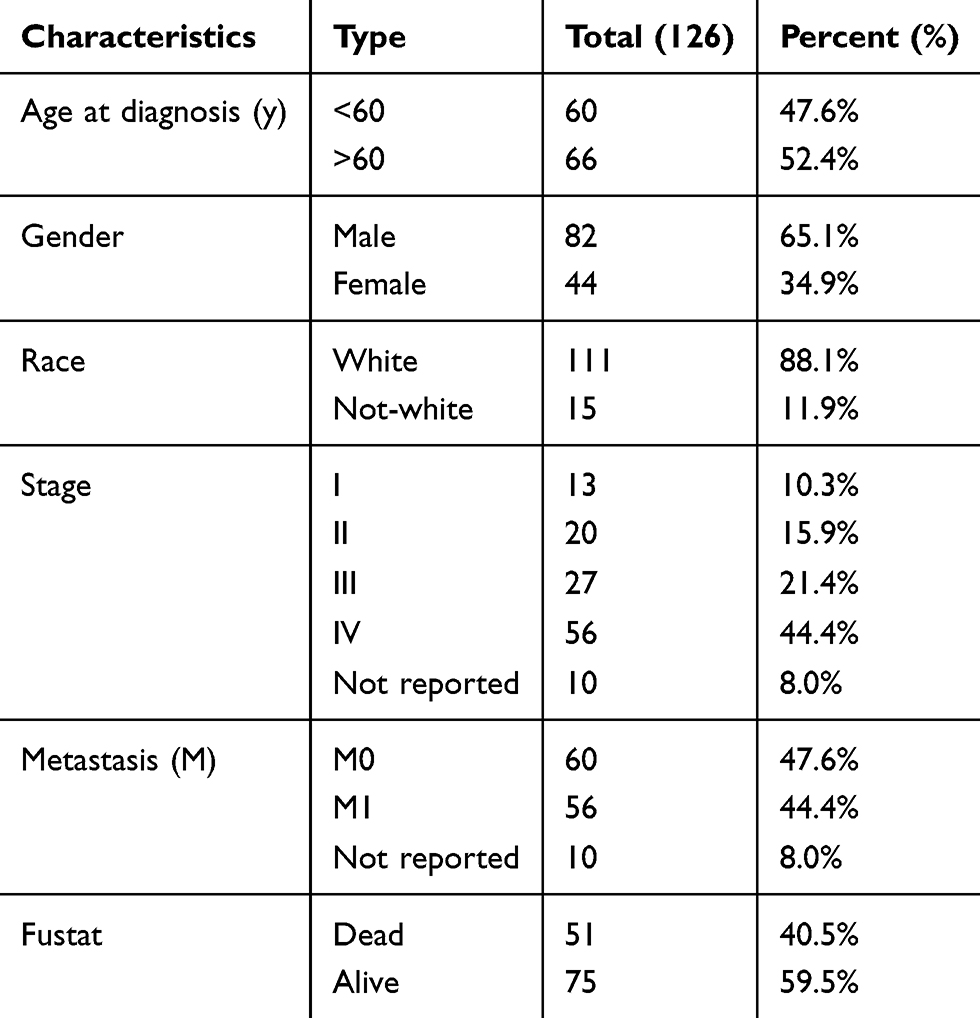 |
Table 2 Characteristics of Patients with Tongue Squamous Cell Carcinoma Based on TCGA |
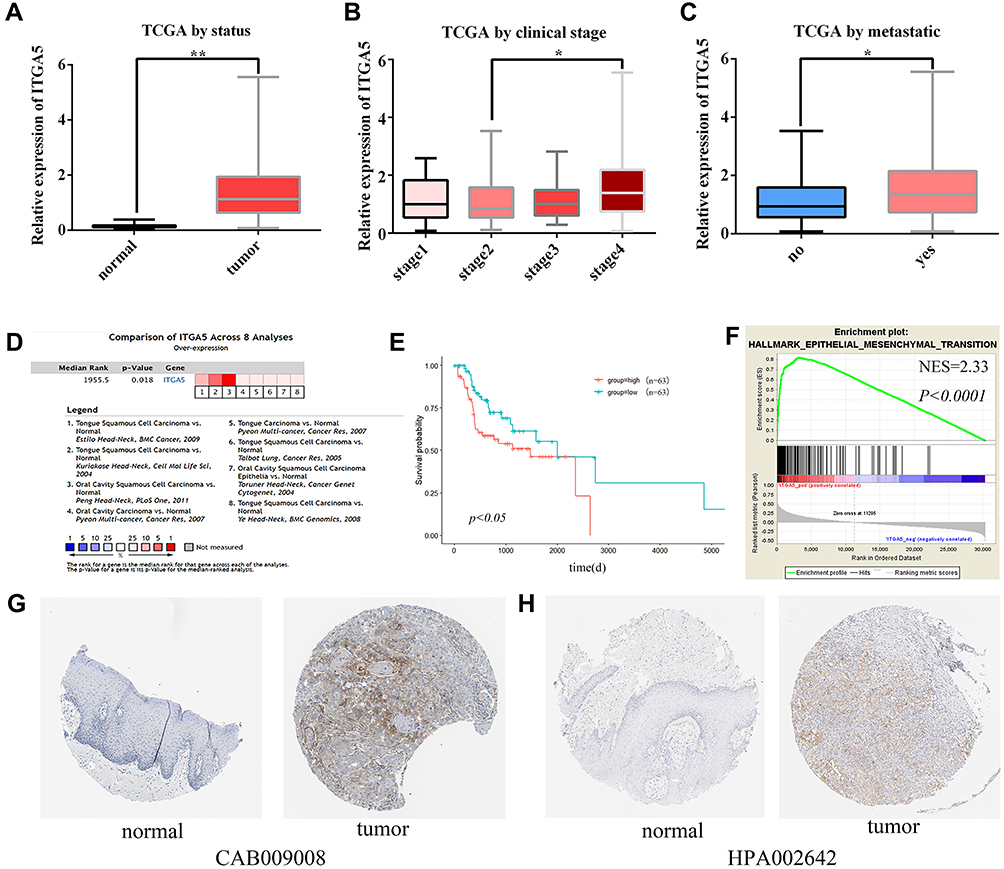 |
Figure 4 ITGA5 expression levels in TSCC. The transcriptional expression level of ITGA5 in TSCC was associated with clinicopathological characteristics including (A) cancer status, (B) clinical stage, and (C) metastases based on TCGA database. (D) Comparison of ITGA5 across eight analyses using the Oncomine database. (E) Univariate analysis of overall survival based on ITGA5 expression in TCGA TSCC. (F) GSEA results showing that EMT hallmark genes are enriched in ITGA5-high TSCC. (G, H) The upregulation of ITGA5 expression in TSCC was validated using The Human Protein Atlas database. *Represents P<0.05, **Represents P<0.01. |
miR-27b Suppressed Tumor Growth of TSCC in vivo
A xenograft tumor formation assay was performed subcutaneously in nude mice to study the impact of miR-27b on tumor growth in vivo. The mice were euthanized after 4 weeks, and the weights of the collected tumors were measured. Compared with miR-NC tumors, the miR-27b-overexpressing tumors showed a significant reduction in weight at the time the mice were sacrificed (Figure 5A–B). The tumors harvested from the three mice in each group were used for further investigation by Western blotting and qRT-PCR. The Western blotting results confirmed that there was a marked decrease in ITGA5 at the protein level in the tumors in the miR-27b mimic group, as shown by the representative bands in Figure 5C. The qRT-PCR results confirmed that ITGA5 expression was significantly lower at the transcriptional level in the tumors in the miR-27b mimic group (Figure 5D).
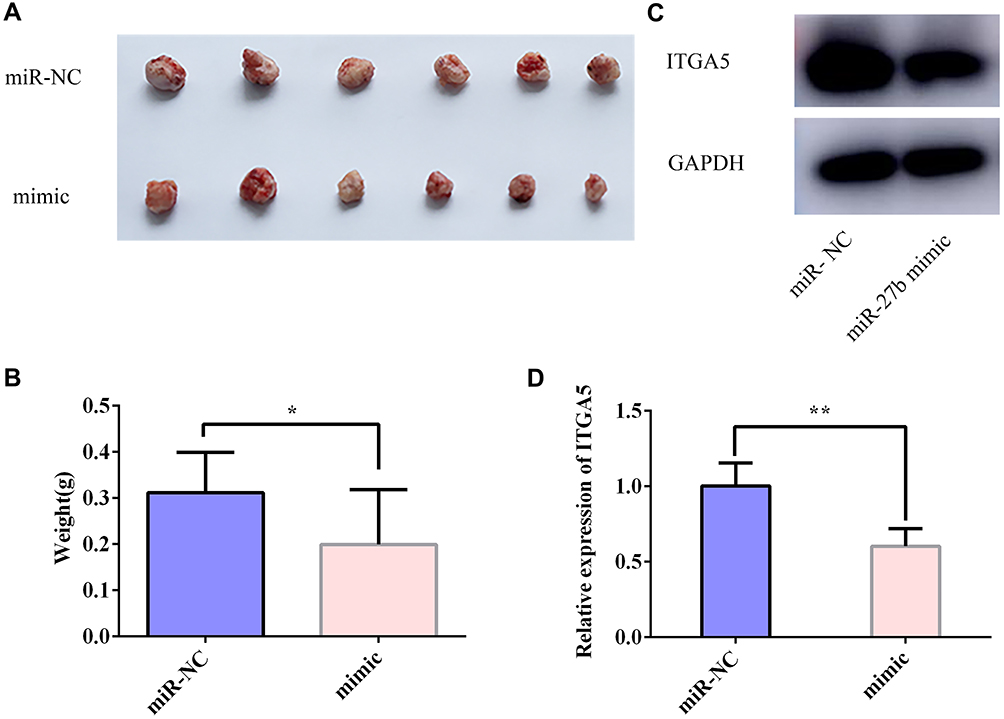 |
Figure 5 miR-27b suppressed TSCC tumor growth in vivo. (A, B) Tumor xenografts in the miR-27b mimic group were significantly smaller than those in the miR-NC group. (C, D) Total protein and RNA were extracted from xenografts and subjected to Western blotting and qRT-PCR assays to test the relative expression levels of ITGA5. *Represents P<0.05, **Represents P<0.01. |
Restoration of ITGA5 Reverses the Inhibitory Effects of miR-27b in TSCC
Western blotting showed that the ITGA protein level in TSCC was increased by transfection with the ITGA5 plasmid, but this effect could be reversed by a miR-27b mimic (Figure 6A). Colony formation assays revealed that the miR-27b mimic abolished the promotion of cell proliferation by ITGA5 (Figure 6B). Further, wound healing and transwell assays showed that the promotion of Cal 27 cell migration and invasion by ITGA5 could also be reversed by the miR-27b mimic (Figure 6C–D).
 |
Figure 6 Restoration of ITGA5 reversed the inhibitory effects of miR- 27b on TSCC cells. (A) ITGA5 protein expression in the Cal 27 cell line was detected by Western blotting after 48 h culture in different groups (NC, ITGA5+miR-27b mimic, ITGA5). Colony formation (B), wound healing (C), and transwell (D) assays were performed in Cal 27 cells after transfection with NC, ITGA5+ miR-27b mimic, and ITGA5. |
miR-27b Targets ITGA5 and Suppresses EMT in TSCC
In our previous study, the GSEA results indicated that changes in the expression of ITGA5 might lead to the enrichment of EMT-related gene sets in TSCC. We speculated that miR-27b might be able to reduce the expression levels of EMT-related markers by targeting ITGA5. Here, the qRT-PCR and Western blotting results revealed that the miR-27b mimic suppressed the expression of ITGA5, N-cadherin, and vimentin in SCC-9 cells, whereas the expression of E-cadherin was increased (Figure 7A–B). These results indicate that miR-27b inhibits EMT by regulating the expression of ITGA5.
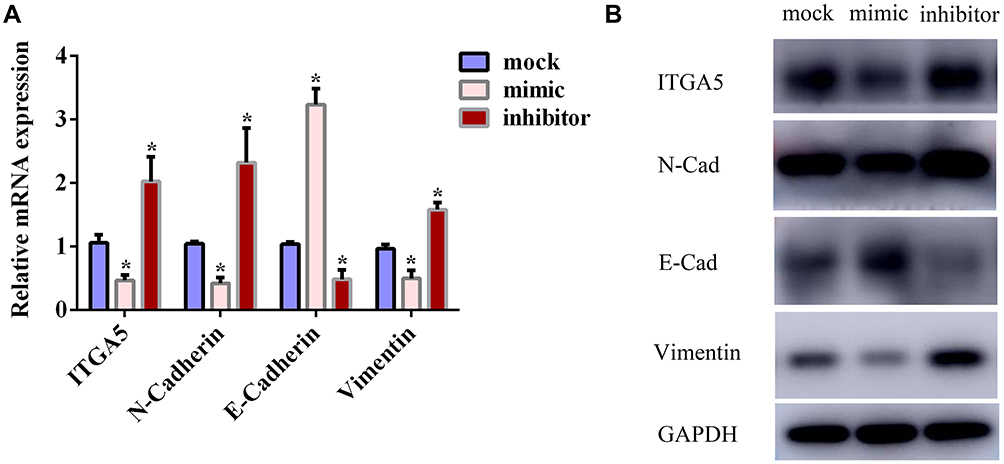 |
Figure 7 miR-27b suppressed EMT of tongue squamous cell carcinoma by targeting ITGA5. (A) Relative mRNA levels of ITGA5 and EMT markers in each group. (B) Western blotting for ITGA5 and EMT markers in each group. *Represents P<0.05. |
Discussion
Emerging evidence reveals that miRNAs function as both tumor suppressors and oncogenes in cells. Dysregulation of miRNAs has strong associations with human oncogenesis, metastasis, and drug resistance.15 Therefore, studying the mechanisms by which miRNAs participate in the occurrence and development of cancer may help to find effective targets for treatment. The poor prognosis of TSCC patients is due to distant metastasis of cancer cells. Identifying tumor-suppressing miRNAs that regulate new metastatic pathways and genes that promote metastasis may improve our understanding of TSCC progression and metastasis. A recent study found that multiple miRNAs are significantly dysregulated in TSCC, including miR-21, miR-493, and miR-27b.16–18 In the present study, we focused on miR-27b based on bioinformatics analysis. As miRNAs can target different transcripts, miR-27b was reported to have conflicting roles in different tumor cells. Previous reports indicated that miR-27b expression was elevated in triple-negative breast cancer and served as a carcinogenic miRNA.7 However, it was also reported to inhibit proliferation and metastasis in ER-positive breast cancer.19 In lung cancer, miR-27b was reported to act as a tumor suppressor by inhibiting cancer growth, EMT, and chemoresistance via targeting Snail1.20 We found that miR-27b was downregulated in TSCC, and its restoration significantly inhibited cancer cell proliferation, migration, and invasion. However, the mechanism of miR-27b in TSCC and its interactions with gene expression have not yet been fully explored. Possible target genes of miR-27b were searched using bioinformatics analysis and online miRNA target gene prediction platforms, including miRanda, TargetScan, and miRDB. ITGA5 was identified as a potential target of miR-27b and confirmed by luciferase assay.
ITGA5, a key member of the integrin family, has critical roles in vital cellular processes. Integrins are cell-surface receptors that mediate the connections between cells and their external environment, including the ECM. They are heterodimeric cell-surface adhesion receptors, generally consisting of non-covalently linked alpha and beta subunits. Many reports have revealed that integrin expression levels are correlated with prognosis in several cancer types, including glioblastoma, cervical squamous cell cancer, ovarian cancer, gastric cancer, and oral cancer.21,22 Integrins can promote cancer growth, metastasis, tumor angiogenesis, and resistance to radiotherapy and chemotherapy by activating growth receptors and downstream cellular signals.23 It has been reported that increased ITGA5 expression predicts poor outcomes in non-small-cell lung cancer.24 ITGA5 has been found to be a direct target gene of several miRNAs, including miR-205, miR-148b, miR-145-5p, miR-330-5p, and miR-17.6,25–27 Owing to their target genes, diversity, and tissue type specificity, the specific regulatory functions of miRNAs have yet to be fully delineated.
Bioinformatics analysis using high-throughput RNA sequencing data from TCGA demonstrated that ITGA5 was upregulated in TSCC and that increased expression of ITGA5 was associated with advanced clinical pathologic characteristics (clinical stage, metastasis) and poor prognosis. To further investigate the functions of ITGA5 in TSCC, we performed GSEA using the TCGA-TSCC data. The results showed that EMT hallmark genes were differentially enriched in the ITGA5 high expression phenotype. This suggested that ITGA5 might promote EMT and serve as a potential marker of prognosis in TSCC.
Excessive epithelial cell proliferation and angiogenesis are hallmarks of the initiation and early growth of primary epithelial cancers.28 The subsequent acquisition of invasiveness is thought to herald the beginning of the last stages of the multi-step process that leads to metastatic dissemination.29 EMT associated with metastasis is known to be an initiation process related to intravasation. In the EMT process, the epithelial cell type loses its original morphology and attains the morphology of a mesenchymal cell type. Many studies have proposed the activation of an EMT program as the critical mechanism for the acquisition of malignant phenotypes by epithelial cancer cells.30 The EMT program is also linked to the residence of cells in stem-cell-like states in some epithelial tissues. In our study, we proved that ITGA5 was a target of miR-27b and confirmed a direct interaction between miR-27b and the ITGA5 3ʹ-UTR. We found that overexpression of miR-27b led to a suppression of TSCC cell proliferation ability both in vitro and in vivo, as well as a reduction in migration and invasion in vitro. Related mechanism research confirmed that overexpression of miR-27b may decrease mesenchymal markers and increase epithelial markers owing to the targeting of ITGA5. These results provide evidence that miR-27b can constrain the growth of TSCC at least in part via inhibiting ITGA5.
Conclusions
miR-27b may target ITGA5 and inhibit proliferation, migration, and invasion of oral squamous cell carcinoma cells, which could activate the EMT process and metastatic pathways. This study provides new insights into the roles of miR-27b in TSCC progression and metastasis, which may be helpful for the development of treatment strategies in the future.
Acknowledgments
This study was supported by the Technology development Program of Shandong Province (2012G0021852).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Mashberg A, Samit A. Early diagnosis of asymptomatic oral and oropharyngeal squamous cancers. CA Cancer J Clin. 1995;45(6):328–351. doi:10.3322/canjclin.45.6.328
2. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. doi:10.3322/caac.21492
3. Peltanova B, Raudenska M, Masarik M. Effect of tumor microenvironment on pathogenesis of the head and neck squamous cell carcinoma: a systematic review. Mol Cancer. 2019;18(1):1–24. doi:10.1186/s12943-019-0983-5
4. Xu F, Zhou F. Inhibition of microRNA-92a ameliorates lipopolysaccharide-induced endothelial barrier dysfunction by targeting ITGA5 through the PI3K/Akt signaling pathway in human pulmonary microvascular endothelial cells. Int Immunopharmacol. 2020;78(July2019):106060. doi:10.1016/j.intimp.2019.106060
5. Shi S, Liu ZH. The role of microRNAs in podocytes. Podocytopathy. 2014;183(November):235–244. doi:10.1159/000360121
6. Xiao Y, Li Y, Tao H, et al. Integrin α5 down-regulation by miR-205 suppresses triple negative breast cancer stemness and metastasis by inhibiting the Src/Vav2/Rac1 pathway. Cancer Lett. 2018;433:199–209. doi:10.1016/j.canlet.2018.06.037
7. Eastlack SC, Dong S, Ivan C, Alahari SK. Suppression of PDHX by microRNA-27b deregulates cell metabolism and promotes growth in breast cancer. Mol Cancer. 2018;17(1):1–16. doi:10.1186/s12943-018-0851-8
8. Sun W, Li J, Zhou L, et al. The c-Myc/miR-27b-3p/ATG10 regulatory axis regulates chemoresistance in colorectal cancer. Theranostics. 2020;10(5):1981–1996. doi:10.7150/thno.37621
9. Ali J, Sabiha B, Jan HU, Haider SA, Khan AA, Ali SS. Genetic etiology of oral cancer. Oral Oncol. 2017;70:23–28. doi:10.1016/j.oraloncology.2017.05.004
10. Zheng W, Jiang C, Li R. Integrin and gene network analysis reveals that ITGA5 and ITGB1 are prognostic in non-small-cell lung cancer. Onco Targets Ther. 2016;9:2317–2327. doi:10.2147/OTT.S91796
11. Li X, Tang M, Zhu Q, Wang X, Lin Y, Wang X. The exosomal integrin α5β1/AEP complex derived from epithelial ovarian cancer cells promotes peritoneal metastasis through regulating mesothelial cell proliferation and migration. Cell Oncol. 2020;43(2):263–277. doi:10.1007/s13402-019-00486-4
12. Lu L, Xie R, Wei R, et al. Integrin α5 subunit is required for the tumor supportive role of fibroblasts in colorectal adenocarcinoma and serves as a potential stroma prognostic marker. Mol Oncol. 2019;13(12):2697–2714. doi:10.1002/1878-0261.12583
13. Li Y, Xiao Y, Lin HP, et al. In vivo β-catenin attenuation by the integrin α5-targeting nano-delivery strategy suppresses triple negative breast cancer stemness and metastasis. Biomaterials. 2019;188:160–172. doi:10.1016/j.biomaterials.2018.10.019
14. Deng Y, Wan Q, Yan W. Integrin α5/itga5 promotes the proliferation, migration, invasion and progression of oral squamous carcinoma by epithelial–mesenchymal transition. Cancer Manag Res. 2019;11:9609–9620. doi:10.2147/CMAR.S223201
15. Iorio MV, Croce CM. MicroRNAs in cancer: small molecules with a huge impact. J Clin Oncol. 2009;27(34):5848–5856. doi:10.1200/JCO.2009.24.0317
16. Spector ME, Bellile E, Amlani L, et al. Prognostic value of MiR-21: an updated meta-analysis in Head and Neck Squamous Cell Carcinoma (HNSCC). JAMA Otolaryngol Head Neck Surg. 2019;8(12):2041. doi:10.1001/jamaoto.2019.2427
17. Jiao D, Liu Y, Tian Z. microRNA-493 inhibits tongue squamous cell carcinoma oncogenicity via directly targetingHMGA2. Onco Targets Ther. 2019;12:6947–6959. doi:10.2147/OTT.S210567
18. Fukumoto I, Koshizuka K, Hanazawa T, Kikkawa N. The tumor-suppressive microRNA-23b/27b cluster regulates the MET oncogene in oral squamous cell carcinoma. Int J Oncol. 2016;49(3):1119–1129. doi:10.3892/ijo.2016.3602
19. Li X, Wu Y, Liu A, Tang X. MiR-27b is epigenetically downregulated in tamoxifen resistant breast cancer cells due to promoter methylation and regulates tamoxifen sensitivity by targeting HMGB3. Biochem Biophys Res Commun. 2016;477(4):768–773. doi:10.1016/j.bbrc.2016.06.133
20. Zhang J, Hua X, Qi N, et al. MiR-27b suppresses epithelial–mesenchymal transition and chemoresistance in lung cancer by targeting snail1. Life Sci. 2020;254:117238. doi:10.1016/j.lfs.2019.117238
21. Goldberg I, Davidson B, Reich R, et al. Alphav integrin expression is a novel marker of poor prognosis in advanced-stage ovarian carcinoma. Clin Cancer Res. 2001;7(12):4073–4079.
22. Schittenhelm J, Schwab EI, Sperveslage J, et al. Longitudinal expression analysis of αv integrins in human gliomas reveals upregulation of integrin αvβ3 as a negative prognostic factor. J Neuropathol Exp Neurol. 2013;72(3):194–210. doi:10.1097/NEN.0b013e3182851019
23. Ye IC, Byun J, Jayatilaka H, et al. Hypoxia selectively enhances integrin α5β1 receptor expression in breast cancer to promote metastasis. Mol Cancer Res. 2017;15(6):723–734. doi:10.1158/1541-7786.MCR-16-0338
24. Dingemans AC, Boogaart V, Van Den Vosse BA, Van SR, Griffioen AW, Thijssen VL. Integrin expression profiling identifies integrin alpha5 and beta1 as prognostic factors in early stage non-small cell lung cancer. Mol Cancer. 2010;9(1):152. doi:10.1186/1476-4598-9-152
25. Gong C, Yang Z, Wu F, Han L, Liu Y, Gong W. MIR 17 inhibits ovarian cancer cell peritoneal metastasis by targeting ITGA5 and ITGB1. Oncol Rep. 2016;36(4):2177–2183. doi:10.3892/or.2016.4985
26. Chen J, Chen T, Zhu Y, et al. CircPTN sponges miR-145-5p/miR-330-5p to promote proliferation and stemness in glioma. J Exp Clin Cancer Res. 2019;38(1):1–17. doi:10.1186/s13046-019-1376-8
27. Orso F, Quirico L, Virga F, et al. miR-214 and miR-148b targeting inhibits dissemination of melanoma and breast cancer. Cancer Res. 2016;76(17):5151–5163. doi:10.1158/0008-5472.CAN-15-1322
28. Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144(5):646–674. doi:10.1016/j.cell.2011.02.013
29. Das V, Bhattacharya S, Chikkaputtaiah C, Hazra S, Pal M. The basics of epithelial–mesenchymal transition (EMT): a study from a structure, dynamics, and functional perspective. J Cell Physiol. 2019;234(9):14535–14555. doi:10.1002/jcp.28160
30. Yun J, Song S, Kim H, Han S, Yi EC, Kim T. Dynamic cohesin‐mediated chromatin architecture controls epithelial–mesenchymal plasticity in cancer. EMBO Rep. 2016;17(9):1343–1359. doi:10.15252/embr.201541852
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.