Back to Journals » Cancer Management and Research » Volume 12
miR-144-3p Contributes to the Development of Thyroid Tumors Through the PTEN/PI3K/AKT Pathway
Authors Cao HL, Gu MQ, Sun Z, Chen ZJ
Received 29 May 2020
Accepted for publication 28 August 2020
Published 9 October 2020 Volume 2020:12 Pages 9845—9855
DOI https://doi.org/10.2147/CMAR.S265196
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Seema Singh
Hui-Ling Cao,1,* Ming-Qiang Gu,2,* Zhuo Sun,3 Zhong-Jian Chen2
1Department of Head and Neck Surgery, Chengwu Hospital Affiliated to Shandong First Medical University, Heze, Shandong, 274200, People’s Republic of China; 2Department of General Surgery, Chengwu Hospital Affiliated to Shandong First Medical University, Heze, Shandong, 274200, People’s Republic of China; 3Department of Oncology, Chengwu Hospital Affiliated to Shandong First Medical University, Heze, Shandong, 274200, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Ming-Qiang Gu
Department of General Surgery, Chengwu Hospital Affiliated to Shandong First Medical University, Heze 274200, Shandong, People’s Republic of China
Tel +86-13573028527
Email [email protected]
Purpose: To explore the expression and related mechanism of miR-144-3p and PTEN in thyroid cancer (TC).
Patients and Methods: From February 2018 to November 2019, 62 patients with TC who received treatment in Chengwu Hospital Affiliated to Shandong First Medical University were collected. TC cells and human normal thyroid HTori-3 cells were purchased. The miR-144-3p-inhibitor, miR-144-3p-mimics, empty vector plasmid (miRNA-NC), si-PTEN and sh-PTEN were transfected into B-CPAP and HTh-7 cells. The expressions of miR-144-3p and PTEN in the specimens were tested by qRT-PCR (qP). WB was used to detect the expression of Bcl-2, APR3, N-cadherin, Slug and Bax proteins in the cells. The cell proliferation was detected by MTT, and the cell invasion was tested by Transwell. The apoptosis was detected by flow cytometry (FC).
Results: miR-144-3p was highly expressed and PTEN was weakly expressed in the patients’ tissues. The AUC of miR-144-3p and PTEN was > 0.8. miR-144-3p and PTEN were related to TNM stage, lymph node metastasis and differentiation degree of TC patients. The B-CPAP and HTh-7 with the greatest expression differences were selected for transfection. The expression of miR-144-3p in miR-144-3p-inhibitor group was significantly lower than that in NC group (P< 0.01), and that in miR-144-3p-mimics group was significantly higher than that in NC group (p < 0.01). The expression of PTEN in si-PTEN group was significantly lower than that in NC group (P< 0.01), while that in sh-PTEN group was significantly higher than that in NC group (P< 0.01). Silencing miR-144-3p and overexpressing PTEN could inhibit cell proliferation, invasion and promote apoptosis. WB detection uncovered that silencing the miR-144-3p expression and overexpressing PTEN could inhibit the PI3K, Akt, p-AKT, Bcl-2, APR3 and cyclinD1 proteins and promote the up-regulation of Bax expression. Rescue experiments revealed that the cell proliferation, invasion and apoptosis were not different from NC after co-transfection of miR-144-3p-mimics+sh-PTEN and miR-144-3p-inhibitor+si-PTEN into B-CPAP and HTh-7.
Conclusion: Inhibition of miR-144-3p expression can up-regulate PTEN and affect cell proliferation, invasion and apoptosis, which may be a potential therapeutic target for TC.
Keywords: miR-144-3p, PTEN/PI3K/AKT, EMT, thyroid cancer, biological mechanism
Introduction
Thyroid cancer (TC), as an endocrine disease, is the most common thyroid malignant tumor.1,2 Although TC is usually localized, TC is aggressive during blood metastases, leading to an increased risk of cancer death.3,4 Despite the progress of molecular diagnostic and targeting therapy in recent years, there is still a lack of efficient biomarkers for the prophylaxis, diagnosis, metastasis and prognosis of TC. It is conducive to conclude the potential mechanism of TC development to develop a more appropriate clinical treatment plan for TC.5–7 Some studies have revealed that specific miRNA can regulate the cellular functions of different tumors. Although studies on functional miRNAs in endocrine cancer are still insufficient, TC, as the most common study object in endocrine tumors, has been revealed that PTEN expression is decreased in TC.8–10 The development and progression of TC are related to the activation of the protein expression signal pathway associated with epithelial-mesenchymal transformation. Studies on the diagnosis or treatment of TC by miRNA-targeted signaling pathway have shown a broad development prospect.11,12
The PTEN/PI3K/AKT pathway plays a vital role in cancer development. Its changes are crucial for the prevention and treatment of tumor tissue development and related diseases.13,14 PTEN is an important phosphatase that is widely involved in the PTEN/PI3K/AKT signal pathway. The anticancer activity of PTEN may be caused by lipid dephosphorylation on the plasma membrane. Studies have shown that the proliferation of some endocrine tumor cells is inhibited, which is related to the increase of PTEN.15 However, the molecular mechanism by which miR-144-3p affects the development of thyroid tumors through the PTEN/PI3K/AKT pathway is still unclear.
Therefore, we explored the molecular mechanism of miR-144-3p in the development of thyroid tumors through the PTEN/PI3K/AKT pathway by detecting the miR-144-3p expression of TC, with a view to finding reliable tumor markers and potential drug action targets for clinical diagnosis and prognosis of TC.
Patients and Methods
From February 2018 to November 2019, 62 Patients with TC Who Visited Our Hospital and Underwent Surgery Were Collected
Inclusion criteria: Patients were diagnosed as TC by pathology, cytology and imaging;16 Patients with TC did not receive relevant preoperative chemotherapy, immunotherapy, radiation and other anti-tumor treatment.
Exclusion criteria: Patients were complicated with liver cirrhosis and coagulation dysfunction were excluded; General clinical data were incomplete; Patients did not cooperate with follow-up; The predicted survival period was less than 1 month; Patients who lost contact during follow-up were excluded. The test was ratified by Chengwu Hospital Affiliated to Shandong First Medical University Ethics Committee, and the informed consent form was signed in advance. This study was conducted according to the international guidelines of Helsinki Declaration.
Main Instruments and Reagents
B-CPAP, HTh-7, BHT101, 8305C, HTh-7 human TC cell line and normal human normal thyroid HTori-3 cell line (BNCC100390, BNCC100396, BNCC100391, BNCC100387, BNCC100401, BNCC338687) were purchased from Beina Biology. There was ABI Stepone Plus real-time fluorescent quantitative PCR apparatus, Lipofectamine™2000 transfection test kit, Trizol extraction test kit, Annexin V/PI apoptosis assay kit (Invitrogen, Carlsbad, CA, USA), Horseradish peroxidase (HRP) labeled goat anti-mouse secondary antibody (R&D Systems, Minneapolis, MN, USA), SYBR Green PCR Master Mix (Applied Biosystems, Waltham, MA, America), MTT kit (Beyotime Biotechnology Co., Ltd., Product No.: C0009) and FACSCanto FC (Becton Dickinson, Franklin Lakes, NJ, America). All sequences of primer were compounded by Sangon Biotech Co., Ltd., Shanghai, China.
Detection Methods
Cell Culturing and Transfection Test
TC cell strains were transfected into DMEM medium containing 10% fetal bovine tissue and penicillin-streptomycin mixture, and cultivated in a cell incubator with 5% CO2 and 37°C constant temperature and saturated humidity. The Lipofectamine™2000 transfection kit was used to transfect miR-144-3p-inhibitor, miR-144-3p-mimics and control plasmid (NC), respectively. Primers were transfected into the cells with the maximal difference in miR-144-3p expression. After transfection for 6 hours, the original culture solution was replaced with a culture solution containing 10% fetal bovine tissue for further culturing. qP was used to verify the transfection efficiency of cells. The siRNA primers used for PTEN were as follows: GGTCAAGTGAAGACGACAA.
qP Test
In tissues and cells, mRNA expression was detected by qP. The total RNA in tissues was obtained with Trizol reagent and it was dissolved in 20μL DEPC water. Then, the total RNA was reverse transcribed by reverse transcription test kit. The synthesized c DNA was applied as a template for qP amplification. Three multiple pores were set for each sample for three repeated tests. miR-144-3p used U6 as internal parameter, and PTEN used GAPDH as internal parameter. After reaction, the amplification and dissociation curves of Real-Time PCR were determined. The relative quantification of the target gene was analyzed by 2-ΔCt. (Table 1)
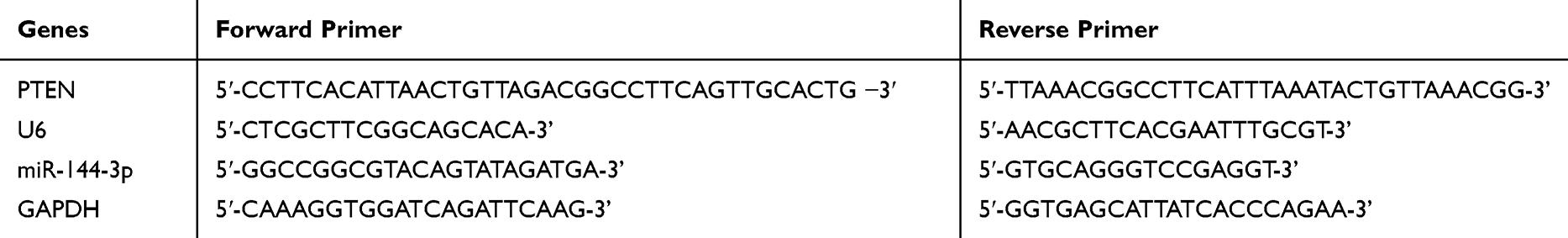 |
Table 1 Primer Sequences of PTEN, miR-144-3p and Their Internal Parameter |
Western Blot (WB) Detection
The lysed cells were collected and transferred to a centrifuge tube, and centrifuged at a temperature of 12000×g for 10min at 4°C. The supernatant was obtained as the protein sample. The protein level was tested by BCA. Finally, it was developed in the darkroom. The protein bands were scanned and the gray values were analyzed by Quantity One (Molecular Devices Corp, The Bay Area, CA, USA).
Cells Growth Assay
MTT was used to detect cell viability. After transfection for 24 hours, the cells were collected. The cells’ OD value in each group was analyzed by spectrophotometer at 570mm wavelength.
Transwell Invasion Experiment
Matrigel glue was coated in Transwell chamber and allowed to stand at 37°C for 30min. Cells were resuspended in a non-tissue DMEM medium. Under the light microscope, the cells passing through the basal membrane of the chamber were figured to indicate the cell invasion activity.
Apoptosis Experiments
After transfection for 48 h, the cells were assimilated through 0.25% trypsinase, washed twice by PBS, and resuspended with 100μL AnnexinV combining buffer to configure as 1×106 cells/mL suspension liquid. 5μL Annexin-V/FITC aqueous solution was added and cultivated at 4°C for 15min. The PI staining solution (5μL) was put and cultivated at 4°C for 5min. FC was used for detection. The test was repeated for 3 times to take the mean value.
Statistical Methods
SPSS 19.0 (SPSS, Inc, Chicago, IL, USA) was employed for statistical assay. Normal distribution data were represented as mean number ± standard deviation (meas±SD). The Inter-group measurement data were compared by independent sample t-test. The data of numerous time points were compared by repeated measure ANOVA. Bonferroni was used for back-test. One-way ANOVA was applied for multi-group comparison of mean values, and LSD-t-test was applied afterwards. The receiver operating characteristic (ROC) curve was applied to estimate the diagnosis value. Pearson test was employed for correlation. The difference was statistically significant with P<0.05.
Results
Expression of miR-144-3p and PTEN in TC Tissues
The expressions of miR-144-3p and PTEN in TC tissues and cell strains were quantitatively tested by qP. These results indicated that miR-144-3p in TC tissues was significantly enhanced and the expression of PTEN was significantly declined compared with corresponding non-tumor normal tissues. ROC assay revealed that the area under the miR-144-3p curve was greater than 0.8. On the basis of the median expression of miR-144-3p, patients were separated into high-expression group and low-expression group of miR-144-3p. These results uncovered that the expression of miR-144-3p was correlated with TNM stage, lymphatic metastasis and differentiation of TC patients. Compared with HTori-3 in normal human normal thyroid cells, the expression of miR-144-3p in TC cells was markedly enhanced while PTEN was significantly decreased. After correlation analysis, the expressions of miR-144-3p and PTEN were negatively correlated in TC tissues. In addition, miR-144-3p and PTEN showed the most significant changes in B-CPAP and HTh-7 cells, so B-CPAP and HTh-7 cells were selected as the study objects in this paper (Figure 1).
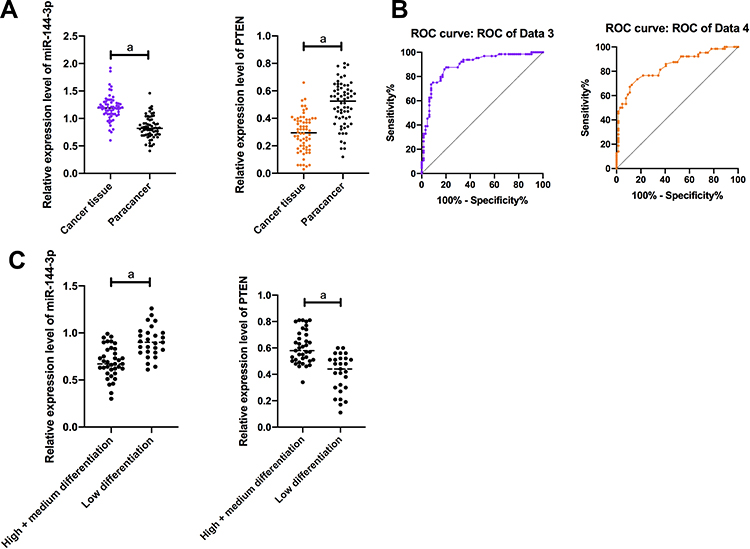 |
Figure 1 Expression and clinical value of miR-144-3p and PTEN in TC patients. (A) The expression level of miR-144-3p in TC tissues was significantly higher than that in adjacent tissues. The expression level of PTEN in TC tissues was significantly lower than that in adjacent tissues. a means P < 0.001. (B) AUC of miR-144-3p was >0.8; AUC of PTEN was >0.8. a means P<0.001. (C) The expression level of miR-144-3p in patients with low differentiation was significantly higher than that in patients with medium and high differentiation. The expression level of PTEN in patients with low differentiation was significantly lower than that in patients with medium and high differentiation. aMeans P < 0.001. |
Expression of miR-144-3p and PTEN in Cells and Their Influences on Cell Biological Functions
(1) qP was applied to detect the expression of miR-144-3p in the cell strains of each group, and it was concluded that the expression of miR-144-3p in B-CPAP, BHT101, 8305C, HTh-7 and HTh-7 cells was significantly increased compared with the thyroid cells of HTori-3 normal subjects (P<0.05). We selected B-CPAP and HTh-7 cells with the maximal difference in expression for transfection. Compared with thyroid cells of HTori-3 normal subjects, the expression of miR-144-3p in B-CPAP, BHT101, 8305C, HTh-7 and HTh-7 cells was obviously enhanced (P<0.05). We selected B-CPAP and HTh-7 with the maximal expression difference for transfection. The miR-144-3p in miR-144-3p-inhibitor group (MIG) was obviously lower than that in NC group (NG), and the expression of miR-144-3p in miR-144-3p-mimics group (MMG) was obviously higher than that in NG (P<0.01). MTT results indicated that the proliferation ability in MIG was obviously lower than that in NG, and the proliferation ability in MMG was higher than that in NG. FC uncovered that the apoptosis rate in MIG was significantly lower than that in NG (P<0.001), and the apoptosis rate in MMG was significantly lower than that in NG (P<0.001). Transwell experiment results indicated that the cell invasion ability in MIG was obviously lower than that in NG, and the cell invasion ability in MMG was obviously higher than that in NG (P<0.001). (Figure 2)
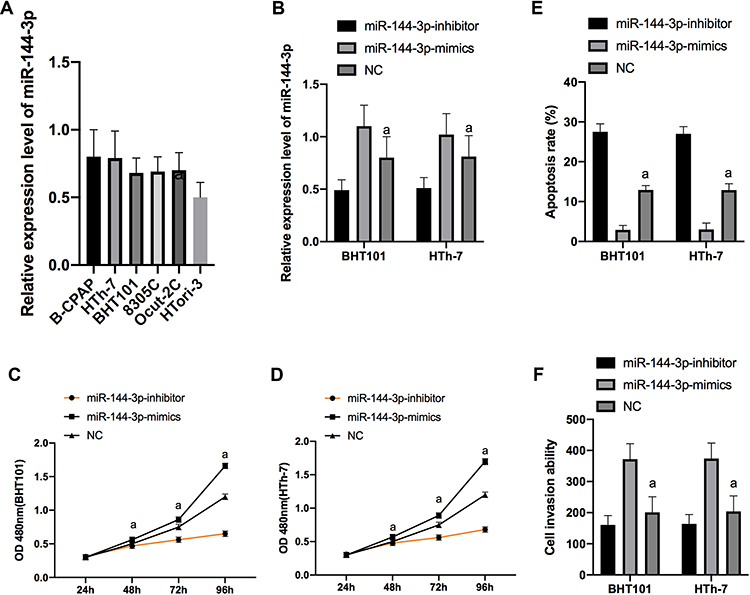 |
Figure 2 Expression of miR-144-3p in cells and its effect on cell biological functions. Expression of miR-144-3p in cell line in each group (A). Expression of miR-144-3p after transfection of B-CPAP and HTh-7 cells (B). Proliferation of BHT101 (C) and HTh-7 (D) cells after transfection. Apoptosis of B-CPAP and HTh-7 cells after transfection (E). Invasion of B-CPAP and HTh-7 cells after transfection (F). |
(2) qP was used to detect the PTEN expression of the cell strains in each group, and it was concluded that the expression of PTEN in B-CPAP and HTh-7 cells was significantly down-regulated compared with the thyroid cells of HTori-3 normal subjects (P<0.05). We selected B-CPAP and HTh-7 with the maximal difference in expression for transfection. PTEN expression in si-PTEN group (siPG) was obviously lower than that in NG (P<0.01), and PTEN expression in sh-PTEN group (shPG) was obviously higher than that in NG. CCK-8 results revealed that the proliferation ability in shPG was markedly lower than that in NG, and the proliferation ability in the siPG was markedly higher than that in the NG. The FC results uncovered that the apoptosis rate in shPG was markedly higher than that in NG, and that of the siPG was obviously lower than that of the miR-NC group (MNG) (P<0.001). Transwell experiment results indicated that the cell invasion ability in shPG was markedly lower than that of NG (P<0.001), and that of siPG was markedly higher than that of MNG (Figure 3).
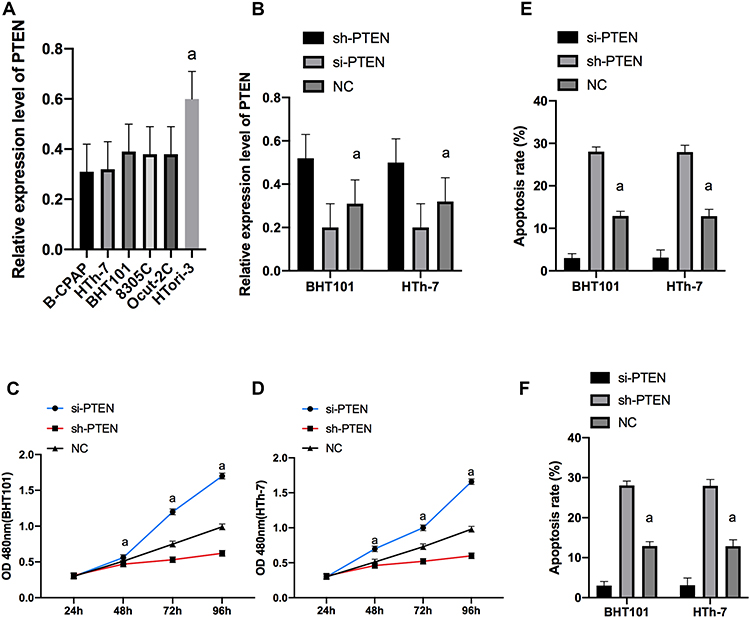 |
Figure 3 Expression of PTEN in cells and its effect on cell biological functions. Expression of PTEN in cell line in each group (A). Expression of PTEN after transfection of B-CPAP and HTh-7 cells (B). Proliferation of BHT101 (C) and HTh-7 (D) cells after transfection. Apoptosis of B-CPAP and HTh-7 cells after transfection (E). Invasion of B-CPAP and HTh-7 cells after transfection (F). |
Effects of miR-144-3p and PTEN Expressions on PTEN/PI3K/AKT Pathway and Apoptotic Protein
WB indicated that the expressions of PI3K, Akt, p-Akt, Bcl-2, APR3 and cyclinD1 proteins were significantly up-regulated and Bax was markedly declined after transfection of miR-144-3p-mimics and si-PTEN compared with transfected NC cells. After transfection of miR-144-3p-inhibitor and sh-PTEN, the PI3K, Akt, p-Akt, Bcl-2, APR3 and cyclinD1 protein expression was markedly declined, while the expression of Bax was markedly enhanced compared with transfected NC cells (p <0.001). (Figure 4)
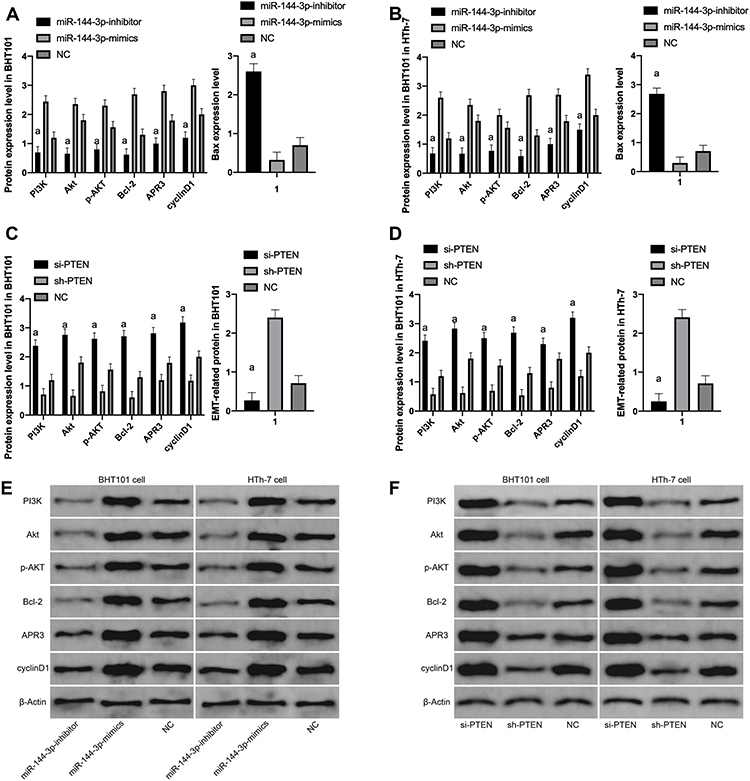 |
Figure 4 Effect of miR-144-3p and PTEN on PTEN/PI3K/AKT pathway and apoptotic protein. (A) Compared with NC, the expression of PI3K, Akt, p-AKT, Bcl-2, APR3 and cyclinD1 proteins in transfected miR-144-3p-inhibitor cells was significantly down-regulated and the expression of Bax was significantly up-regulated, while the expression of PI3K, Akt, p-AKT, Bcl-2, APR3, cyclinD1 protein in transfected miR-144-3p-mimics cells was significantly up-regulated and the expression of Bax was significantly down-regulated. (B) Compared with NC, the expression of PI3K, Akt, p-AKT, Bcl-2, APR3 and cyclinD1 proteins in transfected miR-144-3p-inhibitor cells was significantly down-regulated and the expression of Bax was significantly up-regulated, while the expression of PI3K, Akt, p-AKT, Bcl-2, APR3, cyclinD1 protein in transfected miR-144-3p-mimics cells was significantly up-regulated and the expression of Bax was significantly down-regulated. (C) Compared with NC, the expression of PI3K, Akt, p-AKT, Bcl-2, APR3 and cyclinD1 proteins in transfected si-PTEN cells was significantly up-regulated and the expression of Bax was significantly down-regulated, while the expression of PI3K, Akt, p-AKT, Bcl-2, APR3, cyclinD1 protein in transfected sh-PTEN cells was significantly down-regulated and the expression of Bax was significantly up-regulated. (D) Compared with NC, the expression of PI3K, Akt, p-AKT, Bcl-2, APR3 and cyclinD1 proteins in transfected si-PTEN cells were significantly up-regulated and the expression of Bax was significantly down-regulated, while the expression of PI3K, Akt, p-AKT, Bcl-2, APR3 and cyclinD1 proteins in transfected sh-PTEN cells were significantly down-regulated and the expression of Bax was significantly up-regulated. (E) WB map; (F) WB map. |
Identification of miR-144-3P Target Genes
To further verify the connection of miR-144-3p with PTEN, the downstream target gene of miR-144-3p was predicted by Targetscan6.2. Then, it was found that there was a targeted binding locus of miR-144-3p with PTEN. For this reason, we tested double luciferase activity and found that the luciferase activity of PTEN-3ʹUTR-WT was significantly decreased after overexpression of PTEN (P<0.001), but it had no impact on PTEN-3ʹUTR-Mut luciferase activity (P>0.05). PCR detection found that PTEN expression in B-CPAP and HTh-7 cells was significantly down-regulated after miR-144-3p-mimics transfection, and PTEN in B-CPAP and HTh-7 cells was significantly up-regulated after miR-144-3p-inhibitor transfection (Figure 5).
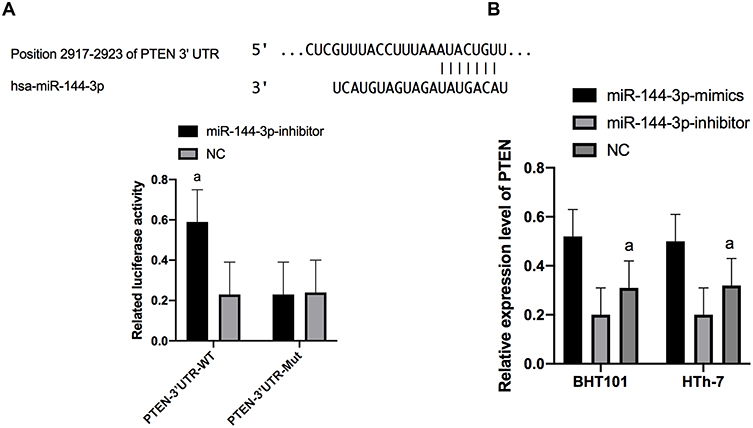 |
Figure 5 Detection of double luciferase activity. (A) There was a binding site between miR-144-3P and PTEN, and the relative luciferase activity-double luciferase report test. (B) Expression of PTEN in transfected B-CPAP and HTh-7 cells. |
Rescue Experiments
Furthermore, miR-144-3p-mimics+sh-PTEN and miR-144-3p-inhibitor+si-PTEN were transfected into B-CPAP and HTh-7 cells to detect the biology functions of the cells. It was concluded that the proliferation, invasion and migration of cells transfected with miR-144-3p-mimics+sh-PTEN and miR-144-3p-inhibitor+si-PTEN had no difference compared with NG, and there was no difference in both groups (P>0.05). However, miR-144-3p-mimics+sh-PTEN, miR-144-3p-inhibitor+si-PTEN had significantly higher proliferation, invasion and migration capabilities than miR-144-3p-mimics, while their proliferation, invasion and migration capabilities were markedly lower than miR-144-3p-inhibitor. Western Blot test was used to detect PI3K-Akt/MAPK-ERK signal pathway and EMT related protein, and it revealed that the expression of EMT protein in miR-144-3p-mimics+sh-PTEN and miR-144-3p-inhibitor+si-PTEN had no difference compared with NG (P>0.05). However, compared with miR-144-3p-mimics, the expression of Bax protein in miR-144-3p-mimics+sh-PTEN and miR-144-3p-inhibitor+si-PTEN reduced markedly, and the expressions of PI3K, Akt, p-AKT, Bcl-2, APR3, cyclinD1 proteins were obviously increased. However, compared with miR-144-3p-inhibitor, the Bax protein expression was markedly increased and the PI3K, Akt, p-AKT, Bcl-2, APR3, cyclinD1 protein expression was markedly declined (Figure 6).
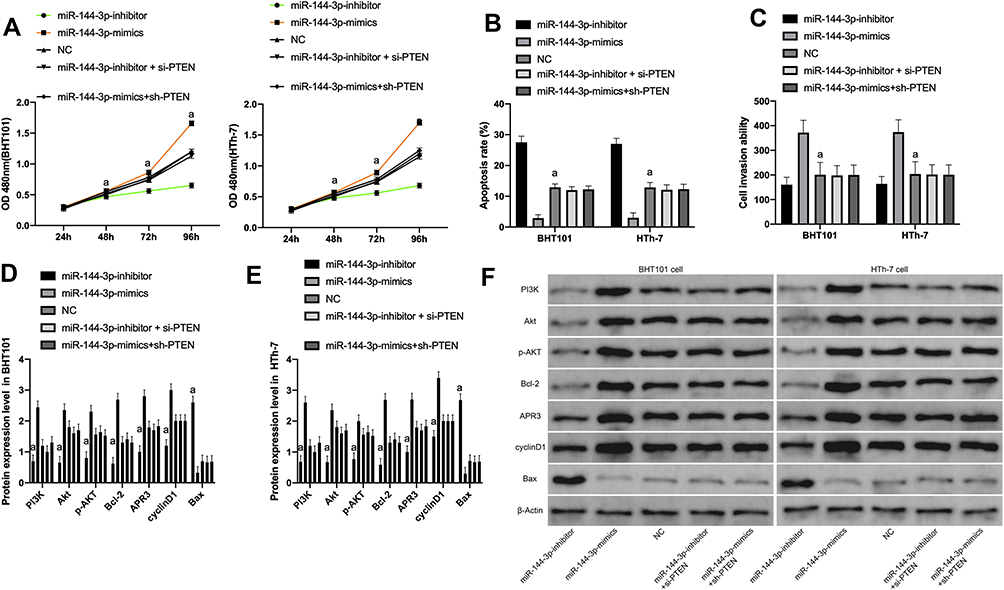 |
Figure 6 Rescue experiments. (A) Cells proliferation ability after transfection of miR-144-3p-mimics+sh-PTEN or miR-144-3p-inhibitor+si-PTEN. (B) Cells apoptosis ability after transfection of miR-144-3p-mimics+sh-PTEN or miR-144-3p-inhibitor+si-PTEN. (C) Cells invasion ability after transfection of miR-144-3p-mimics+sh-PTEN or miR-144-3p-inhibitor+si-PTEN. (D) Expression of Bax, PI3K, Akt, p-AKT, Bcl-2, APR3, cyclinD1 proteins after transfection of miR-144-3p-mimics+sh-PTEN or miR-144-3p-inhibitoror+si-PTEN. (E) It was the same as D; (F) WB map. |
Discussion
The specific miRNA expression exists in various cancer tissues of human and participates in protein coding. In recent years, the role of miRNA in regulating signaling pathways has attracted attention from clinical research and development of new molecular target therapy.17 miRNA has unique expression status in different cancer types. Related cell biology studies have found that cytoplasmic miRNA can affect the growth and differentiation of cancer cells by regulating signaling pathway related proteins.18 This test was designed to explore the biology function of miR-144-3p on TC cells by regulating PTEN/PI3K/AKT pathway, so as to offer a new theoretical foundation for the diagnosis and treatment of TC in molecular biology.
In our research, we applied qP technology to test the expression of miR-144-3p in serum of TC patients and healthy people. Then, we concluded that miR-144-3p was abnormally enhanced in serum of TC patients. We further concluded that the high expression of miR-144-3p was correlated with the differentiation degree, staging and lymphatic metastasis of TC patients through correlation analysis of clinicopathological features of patients, and the AUC of miR-144-3p was more than 0.8 by drawing ROC curve. At present, some reports have shown that miR-144-3p can target PTEN gene pairs to regulate cell biological functions.19,20 We further revealed the targeted connection of PTEN with miR-144-3p through the analysis of the TargetScan database. Clinicopathological features of PTEN and TC patients were also analyzed. It also revealed that PTEN expression was low, and the AUC was more than 0.8 by drawing ROC curve. miR-144-3p was abnormally expressed in digestive tract diseases such as colorectal carcinoma, non-small cell lung carcinoma and hepatocellular cancer, but the specific regulatory mechanism of miR-144-3p on endocrine tumor cells has not been elaborated in detail.21 miR-196a can promote the proliferation of ovarian cancer cells and weaken apoptosis by down-regulating DDX3 expression through PTEN/PI3K/AKT pathway.22 PTEN gene is a new tumor suppressor gene of metabolic products, which is weakly expressed in the development of endocrine tumor.23 Then, we detected the TCGA database and the expressions of miR-144-3p and PTEN in TC cells. The results indicated that miR-144-3p was highly expressed and PTEN was weakly expressed in TC cells, which was consistent with the results of this research, suggesting that both miR-144-3p and PTEN might play a regulatory role in TC.
In cell experiment, human TC cell strains B-CPAP and HTh-7 and normal human thyroid cells were compared, and we found that miR-144-3p was highly expressed and PTEN was weakly expressed in TC cell strains. Subsequently, we treated the miR-144-3p and PTEN expression levels in B-CPAP and HTh-7 cells through silencing and over-expression. We further transfected miR-144-3p-mimics, miR-144-3p-inhibitor, sh-PTEN and si-PTEN sequences into B-CPAP and HTh-7. By observing the cell biological function, we found that the cell proliferation and invasion ability were significantly controlled after the inhibition of miR-144-3p expression and over-expression of PTEN. The development and progression of tumor were closely related to EMT. The development of EMT would promote the biological function changes of tumor cells. This suggested that PTEN could be used as a potential target for the therapy of TC. Inhibiting the expression of miR-144-3p could control proliferation, invasion and EMT of TC cells. miR-144-3p and KRAS expression in CRC tumors and cells was up-regulated, while the expression of miR-944 was down-regulated. Besides, some researches have revealed that knockdown of miR-144-3p gene can hinder tumor growth, and miR-144-3p can reduce the proliferation of colorectal carcinoma cell and accelerate the apoptosis of colorectal cancer cell by inhibiting PI3K/AKT pathway.24 Studies by Liu et al have also uncovered that miR-144-3p can target pairwise box gene 8 (PAX8) to influence the biological function of TC cells. This further suggests the impact of miR-144-3p changes on TC. However, it is not clear how miR-144-3p affects the biology function of TC cells.25
Studies have shown that PTEN/PI3K/AKT signal pathway affects the proliferation and apoptosis of cells, and the overexpressing miR-218 inactivates PTEN/PI3K/AKT pathway by targeting Runx2 during the growth of TC cells, thus inhibiting the occurrence of thyroid papillary carcinoma.26 In this study, we observed PTEN/PI3K/AKT related proteins by inhibiting or over-expressing miR-144-3p and treating PTEN in TC cells. These results uncovered that the Bcl-2, APR3, N-cadherin and Slug proteins expression in cells with low expression of miR-144-3p or high expression of PTEN were markedly declined and the expression of Bax was significantly enhanced, while the results of PTEN high expression or miR-144-3p low expression were opposite. This suggested that miR-144-3p could regulate PTEN to inhibit the activation of PTEN/PI3K/AKT and promote the apoptosis of cancer cells. miR-144-3p can directly target the 3ʹ- untranslated region of multiple myeloma cell stroma to epithelial transformation factor and inhibit the expression of PI3K/AKT downstream signal pathway.27 Some reports on apoptotic proteins in TC cells have also indicated that APR3, Bcl-2, cyclinD1, and Bax play key roles in cell cycle and apoptosis in tumorigenesis.28 The expression rate of APR3 and cyclinD1 was linked to TNM stage and cell cycle of tumor.29 However, the expression of Bcl-2/and Bax related genes was correlated with the number of apoptotic cells, and the smaller the Bcl-2/Bax value, the more apoptotic cells.30
At the end of the study, rescue experiments indicated that after co-transfecting sh-PTEN+miR-144-3p-mimics, si-PTEN+miR-144-3p-inhibitor into B-CPAP and HTh-7, the biological function changes of the cells were not different from those of MNG. However, compared with miR-144-3p-inhibitor, the proliferation, invasion and migration capabilities were enhanced, while miR-144-3p-mimics were opposite. This implied that miR-144-3p could target PTEN regulation. For this reason, we further determined the connection between PTEN and miR-144-3p through double luciferase report. These results indicated that the luciferase activity of miR-144-3p-3ʹ UT Wt was significantly increased after PTEN over-expression, but it had no influence on miR-144-3p-3ʹ UTR Mut luciferase activity. The PTEN expression in transfected miR-144-3p-inhibitor was markedly enhanced, which suggested that there was a targeted regulatory connection of miR-144-3p with PTEN. Therefore, we believed that inhibiting miR-144-3p expression could promote PTEN expression, thus affecting the biological function of TC cells.
In our research, we have revealed that miR-144-3p was highly expressed in TC, and over-expression of miR-144-3p could control PTEN-mediated cell growth. However, there are still some limitations in this study. For example, we can further conduct colony, wound healing assays and related animal experiments to further prove the role of miR-144-3p in thyroid cancer. Regarding the regulatory network of PTEN, whether it can affect the development and progression of tumors through other ways still needs to be further studied. Therefore, we hope to explore the regulatory network of PTEN through bioinformatics analysis in future studies, so as to provide more evidence for our experiments.
To sum up, miR-144-3p can adjust PTEN/PI3K/AKT signal pathway by targeting PTEN to change the growth and apoptosis of TC cells. It is expected to become a potential clinical therapeutic target for TC. miR-144-3p may be a useful marker for diagnosis and prognosis evaluation of disease.
Conclusion
To sum up, miR-144-3p can adjust PTEN/PI3K/AKT signal pathway by targeting PTEN to change the growth and apoptosis of TC cells. It is expected to become a potential clinical therapeutic target for TC. miR-144-3p may be a useful marker for diagnosis and prognosis evaluation of disease.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Lortet‐Tieulent J, Franceschi S, Dal Maso L, et al. Thyroid cancer “epidemic” also occurs in low‐and middle‐income countries. Int J Cancer. 2019;144(9):2082–2087. doi:10.1002/ijc.31884
2. Klaus A, Fathi O, Tatjana TW, et al. Expression of hypoxia-associated protein HIF-1α in follicular thyroid cancer is associated with distant metastasis. Pathol Oncol Res. 2018;24(2):289–296. doi:10.1007/s12253-017-0232-4
3. Lippi F, Picone A. Differentiated thyroid cancer: new strategy of treatment. EC Endocrinol Metab Res. 2018;3:222–224.
4. Lortet-Tieulent J, Vaccarella S. International and subnational variation thyroid cancer incidence and mortality over 2008–2012. Revue d’Épidémiologie et de Santé Publique. 2018;66:S254. doi:10.1016/j.respe.2018.05.053
5. Landa I, Pozdeyev N, Korch C, et al. Comprehensive genetic characterization of human thyroid cancer cell lines: a validated panel for preclinical studies. Clin Cancer Res. 2019;25(10):3141–3151. doi:10.1158/1078-0432.CCR-18-2953
6. Xhikola M, Young G, Sharma A, et al. SAT-585 presence of metastatic medullary thyroid cancer without a distinct thyroid nodule: a diagnostic challenge. J Endocrine Soc. 2019;3(Supplement_1):SAT–585. doi:10.1210/js.2019-SAT-585
7. Kitahara C, Pfeiffer R, Sosa J, et al. OR27-4 the impact of overweight and obesity on us trends in papillary thyroid cancer incidence. J Endocrine Soc. 2019;3(Supplement_1):OR27–4. doi:10.1210/js.2019-OR27-4
8. Ramírez-Moya J, Santisteban P. miRNA-directed regulation of the main signaling pathways in thyroid cancer. Front Endocrinol. 2019;10:430. doi:10.3389/fendo.2019.00430
9. Jankovic-Miljus J, Wert-Lamas L, Guillen-Sacoto MA, et al. Profiling circulating microRNAs as serum biomarkers for thyroid cancer recurrence before and after recombinant human TSH stimulation. In 21st European Congress of Endocrinology. BioScientifica. 2019;63.
10. Sloot YJE, Rabold K, Netea MG, et al. Effect of PTEN inactivating germline mutations on innate immune cell function and thyroid cancer-induced macrophages in patients with PTEN hamartoma tumor syndrome. Oncogene. 2019;38(19):3743. doi:10.1038/s41388-019-0685-x
11. Ramirez-Moya J, Wert-Lamas L, Santisteban P. MicroRNA-146b promotes PI3K/AKT pathway hyperactivation and thyroid cancer progression by targeting PTEN. Oncogene. 2018;37(25):3369.
12. Wu YR, Qi HJ, Deng DF, et al. MicroRNA-21 promotes cell proliferation, migration, and resistance to apoptosis through PTEN/PI3K/AKT signaling pathway in esophageal cancer. Tumor Biol. 2016;37(9):12061–12070. doi:10.1007/s13277-016-5074-2
13. Pan H, Li T, Jiang Y, et al. Overexpression of circular RNA ciRS‐7 abrogates the tumor suppressive effect of miR‐7 on gastric cancer via PTEN/PI3K/AKT signaling pathway. J Cell Biochem. 2018;119(1):440–446. doi:10.1002/jcb.26201
14. Pérez-Ramírez C, Cañadas-Garre M, Molina MÁ, et al. PTEN and PI3K/AKT in non-small-cell lung cancer. Pharmacogenomics. 2015;16(16):1843–1862. doi:10.2217/pgs.15.122
15. Ke TW, Wei PL, Yeh KT, et al. MiR-92a promotes cell metastasis of colorectal cancer through PTEN-mediated PI3K/AKT pathway. Ann Surg Oncol. 2015;22(8):2649–2655.
16. La Vecchia C, Malvezzi M, Bosetti C, et al. Thyroid cancer mortality and incidence: a global overview. Int J Cancer. 2015;136(9):2187–2195. doi:10.1002/ijc.29251
17. Reddy KB. MicroRNA (miRNA) in cancer. Cancer Cell Int. 2015;15(1):38. doi:10.1186/s12935-015-0185-1
18. Liang WC, Fu WM, Wong CW, et al. The lncRNA H19 promotes epithelial to mesenchymal transition by functioning as miRNA sponges in colorectal cancer. Oncotarget. 2015;6(26):22513. doi:10.18632/oncotarget.4154
19. Yuan X, Pan J, Wen L, et al. MiR-144-3p enhances cardiac fibrosis after myocardial infarction by targeting PTEN. Front Cell Dev Biol. 2019;7:249.
20. Murphy CP, Li X, Maurer V, et al. MicroRNA-mediated rescue of fear extinction memory by miR-144-3p in extinction-impaired mice. Biol Psychiatry. 2017;81(12):979–989. doi:10.1016/j.biopsych.2016.12.021
21. Li H, Shi H, Gao M, et al. Long non-coding RNA CASC2 improved acute lung injury by regulating miR-144-3p/AQP1 axis to reduce lung epithelial cell apoptosis. Cell Biosci. 2018;8(1):15. doi:10.1186/s13578-018-0205-7
22. Ni J, Chen L, Ling L, et al. MicroRNA-196a promotes cell proliferation and inhibits apoptosis in human ovarian cancer by directly targeting DDX3 and regulating the PTEN/PI3K/AKT signaling pathway. Mol Med Rep. 2020;22:1277–1284. doi:10.3892/mmr.2020.11236
23. Yehia L, Eng C. 65 YEARS OF THE DOUBLE HELIX: one gene, many endocrine and metabolic syndromes: PTEN-opathies and precision medicine. Endocr Relat Cancer. 2018;25(8):T121–T140.
24. Wu M, Huang C, Huang X, et al. MicroRNA-144-3p suppresses tumor growth and angiogenesis by targeting SGK3 in hepatocellular carcinoma. Oncol Rep. 2017;38(4):2173–2181. doi:10.3892/or.2017.5900
25. Liu C, Su C, Chen Y, et al. MiR-144-3p promotes the tumor growth and metastasis of papillary thyroid carcinoma by targeting paired box gene 8. Cancer Cell Int. 2018;18(1):54. doi:10.1186/s12935-018-0550-y
26. Han M, Chen L, Wang Y. miR-218 overexpression suppresses tumorigenesis of papillary thyroid cancer via inactivation of PTEN/PI3K/AKT pathway by targeting Runx2. Onco Targets Ther. 2018;11:6305.
27. Zhao Y, Xie Z, Lin J, et al. MiR-144-3p inhibits cell proliferation and induces apoptosis in multiple myeloma by targeting c-Met. Am J Transl Res. 2017;9:2437–2446.
28. Ding XD, Chen YW, Han LS, et al. Apoptosis related protein 3 is a lysosomal membrane protein. Biochem Biophys Res Commun. 2015;460(4):915–922. doi:10.1016/j.bbrc.2015.03.127
29. Han S, Lu Q, Wang N. Apr3 accelerates the senescence of human retinal pigment epithelial cells. Mol Med Rep. 2016;13(4):3121–3126. doi:10.3892/mmr.2016.4926
30. Kalashnikova I, Mazar J, Neal CJ, et al. Nanoparticle delivery of curcumin induces cellular hypoxia and ROS-mediated apoptosis via modulation of Bcl-2/Bax in human neuroblastoma. Nanoscale. 2017;9(29):10375–10387. doi:10.1039/C7NR02770B
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.