Back to Journals » Cancer Management and Research » Volume 12
MiR-139-5p-ZEB1 is a Molecular Regulator of Growth, Invasion, and Epithelial-to-Mesenchymal Transition of Cervical Cancer
Authors Sun J, Wang S, Liu P, Liu Y
Received 13 June 2020
Accepted for publication 15 September 2020
Published 10 December 2020 Volume 2020:12 Pages 12723—12733
DOI https://doi.org/10.2147/CMAR.S267634
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Chien-Feng Li
Jinrui Sun,1 Shanshan Wang,2 Ping Liu,1 Yulan Liu3
1Department of Gynecology, Shanxi Provincial People’s Hospital, Taiyuan, Shanxi Province 030012, People’s Republic of China; 2Department of Cardiology, Yidu Central Hospital of Weifang City, Weifang, Shandong, People’s Republic of China; 3Department of Gynecology, Maternal and Child Health Hospital of Hubei Province, Wuhan, Hubei 430070, People’s Republic of China
Correspondence: Yulan Liu
Department of Gynecology, Maternal and Child Health Hospital of Hubei Province, NO. 745 Wuluo Road, Hongshan, Wuhan, Hubei 430070, People’s Republic of China
Tel +86-15698784852
Email [email protected]
Objective: To verify that miR-139-5p-zinc finger E-box-binding homeobox 1 (ZEB1) is a molecular regulator of the biological function and epithelial–mesenchymal transition (EMT) of cervical cancer (CC) cells.
Methods: Cancerous tissues, corresponding paracancerous tissues, and serum were sampled from patients with CC. MiR-139-5p and ZEB1 in tissue specimens, serum specimens, and purchased CC cell lines were quantified, and Pearson correlation coefficient was adopted for correlation analysis of miR-139-5p in clinical specimens. Receiver operating characteristic (ROC) curves were adopted to analyze the diagnostic value of miR-139-5p and ZEB1 for CC. The expression of genes in CC cells was changed by transfection. The proliferation, colony formation, invasion, and apoptosis of cells were determined, and the protein level of EMT markers (N-cadherin, vimentin, and E-cadherin) was also quantified. Moreover, the targeting relationship between miR-139-5p and ZEB1 was determined.
Results: Our data showed that the expression of miR-139-5p decreased greatly in CC tissues, and it also significantly decreased in the serum, while the expression of serum ZEB1 was opposite. In addition, the miR-139-5p expression in CC tissues was positively correlated with that in serum, while serum miR-139-5p was negatively correlated with serum ZEB1. The areas under the curves (AUCs) of the two for identifying CC were 0.923 and 0.890, respectively. Both up-regulation of miR-139-5p and down-regulation of ZEB1 suppressed the colony formation, proliferation, invasion, and EMT of CC cells, and intensified their apoptosis. Moreover, miR-139-5p negatively regulated the transcription of ZEB1, and down-regulation of the former could reverse the molecular regulatory effects of down-regulating ZEB1 on the above biological behaviors of CC cells.
Conclusion: The above data imply that miR-139-5p-ZEB1 axis may be the key to curbing the progression of CC.
Keywords: cervical cancer, miR-139-5p, ZEB1, growth, invasion, epithelial–mesenchymal transition
Introduction
Cervical cancer (CC) is a gynecologic tumor with poor prognosis, which may deteriorate to brain metastasis or bone metastasis.1,2 According to global data, there were 570,000 new cases with CC and up to 311,000 cases dying of the disease in 2018.3 At present, the exact mechanism of the progression of CC remains unclear, and biological indicators for the diagnosis of CC remain to be found,4,5 so it is crucial for the management of CC to study the mechanism of CC progression and find potential screening indicators.
MicroRNA (miRNA), as a small non-coding RNA molecule, is a molecular regulator of the growth, invasion and epithelial–mesenchymal transition (EMT) of various tumors including CC.6–8 MiRNA is found to have regulatory effect in various gynecologic tumors. For example, in endometrial carcinoma, miR-136 regulates the development of tumor cells by regulating NOTCH3 under the regulation of Circ_pumilio homolog 1 (PUM1),9 and miR-653-5p accelerates the malignant growth of CC under the sponge of DGUOK-AS1.10 In the present study, we focused on the expression and role of miR-139-5p in CC, which reportedly serves as a tumor inhibitor in the disease.11 It has been found that miR-139-5p can regulate the pathological processes of glioma such as metastasis, infiltration, and EMT.12 We found a potential targeting relationship between miR-139-5p and zinc finger E-box-binding homeobox 1 (ZEB1) based on an online target gene prediction website. Moreover, it has been uncovered that both miR-139-5p and ZEB1 have targeted regulatory effects on breast cancer (BC), colorectal cancer (CRC), as well as glioblastoma. For example, miR-139-5p-ZEB1 axis affects the malignant biological behaviors of BC cells under the regulation of TTN-AS1, and it also affects the pathological changes of CRC cells under the regulation of HLA complex P5 (HCP5).13–15 However, ZEB1 is reported abnormally up-regulated in cases with CC, which affects the proliferation, metastatic behavior, and EMT of CC cells.16,17
In the present study, we inferred that miR-139-5p-ZEB1 was a molecular regulator of the growth, invasion, and EMT of CC, and thus we conducted relevant studies to verify it.
Materials and Methods
Sample Collection
This study was approved by the Ethics Committee of Shanxi Provincial People’s Hospital, and written consent forms were obtained from all participants. Cancerous tissues and corresponding paracancerous tissues were sampled from99 patients with CC (CC group) during operation from April 2016 to April 2020, and serum was also sampled from them before operation. All the patients were diagnosed with CC based on pathology.18 In addition, serum was sampled from 50 healthy individuals in physical examination during the same period as a healthy control group (HC group). All patients with CC had not received any surgery or drug treatment before sample collection, and all sampled specimens were stored at −80°C for later analysis. Moreover, the clinical staging of the patients was evaluated according to the International Federation of Gynecology and Obstetrics (FIGO) criteria,19 and the histological grading of them is also evaluated in Table 1.
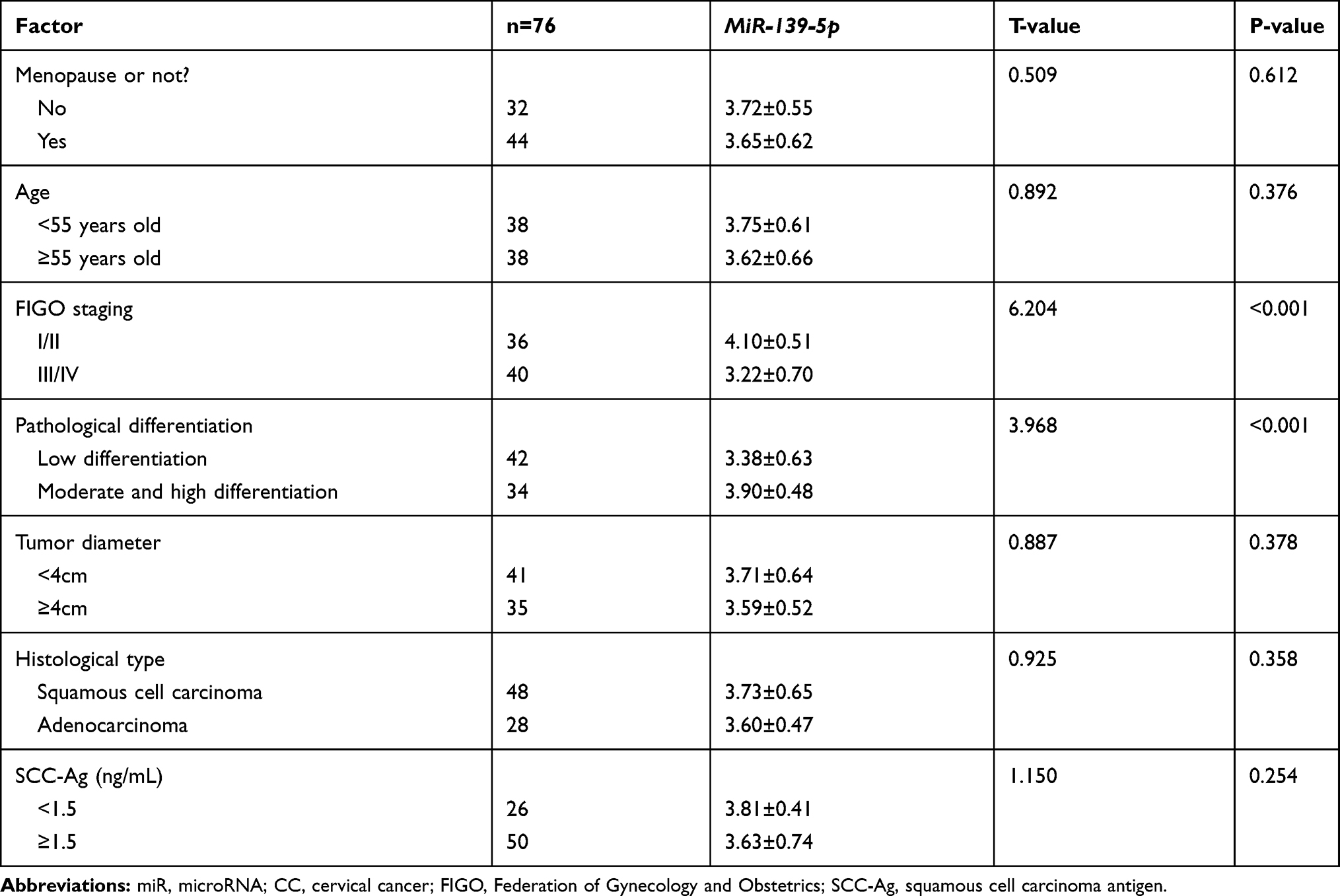 |
Table 1 Relationship Between MiR-139-5p and Pathological Data of Patients with CC [Mean±SD] |
Cell Culturing
Human normal cervical epithelial cells (HUCEC) (FE1528, Qiming Biotechnology Co., Ltd., Shanghai, China) and human CC cells (HeLa 229, H1HeLa, C-33 A, and MS751) (YBCC102035, YBCC101654, YBCC101021, and YBCC100277, Yubo Biological Technology Co., Ltd., Shanghai, China) were incubated in dulbecco’s modified eagle medium (DMEM; PM150220B, Yaji Biotechnology Co., Ltd., Shanghai, China) supplemented with 10% phosphate buffer saline (PBS) under 5% CO2 at 37°C.
Cell Transfection
Transfectants used in this study included miR-139-5p mimics (miR-139-5p), inhibition sequence (inhibitor), miR negative control (miR-NC), targetedly overexpressed sequence of ZEB1 (ZEB1), targetedly inhibited sequence of ZEB1 (si-ZEB1), and si-negative control (si-NC), which were mainly purchased from Shanghai Huishen Biological Technology Co., Ltd. CC cells were transfected with above transfectants, respectively using a cell transfection kit (YSRIBIO-C5838, Yansheng Industrial Co., Ltd., Shanghai, China) in strict accordance with the kit instructions. After48 h, the transfected cells were harvested for later analysis.
Real-Time Polymerase Chain Reaction (PCR)
Total RNA was extracted from harvested tissues, serum, as well as cells by a TRIzol kit (R523-200, Spectral Experimental Equipment Technology Co., Ltd., Dongguan, China), followed by detection of its purity and concentration with an ultraviolet spectrophotometer (SPCC, Spectral Experimental Equipment Technology Co., Ltd., Dongguan, China). Subsequently, total RNA was reversely transcribed into cDNA and amplified. The data in this assay were analyzed using the 2−ΔΔct, with β-Actin as internal reference for ZEB1, Caspase-3, Bax, and Bcl-2, and U6 as internal reference for miR-139-5p.
Western Blot
Protein in tissues or cells was isolated by RIPA buffer (HLIT0050, Haling Biotechnology Co., Ltd., Shanghai, China), and then ionized and transferred to a membrane. Subsequently, the membrane was sealed with blocking solution (BH-DB6564, Bohu Biotechnology Co., Ltd., Shanghai, China) for 1 h, followed by incubation with primary antibodies (N-cadherin, vimentin, E-cadherin, ZEB1, and β-Actin) at 4°C overnight. Afterwards, the membrane was subjected to 1-h incubation with secondary antibody at 4°C. All antibodies were purchased from Shanghai Xuanya Biotechnology Co., Ltd. Finally, the protein strips were visualized and analyzed by Quantity One software (ECL-0013, EASYBIO Technology Co., Ltd., Beijing, China).
Cell Proliferation
Cell proliferation was determined using a MTT kit (RF(m)11,473, Qiming Biotechnology Co., Ltd., Shanghai, China) as follows: The cells were seeded into a 96-well plate at 1000 cells/well, incubated at 37°C for 3 d, and then added with 20μL MTT solution (2.5 mg/mL), followed by 3-h incubation. Subsequently, each well was added with 150 μL dimethyl sulfoxide, and the optical density of cells in each group at 570 mm was detected using an ELx808LBS microplate reader (ZEPING Bioscience & Technologies Co., Ltd., Beijing, China).
Colony Formation Assay
CC cells (2×103 cells) were seeded into a 6-well plate. After14 d, the cells were immobilized with 4% paraformaldehyde (PFA; M002, Gefan Biotechnology Co., Ltd., Shanghai, China) at room temperature for 10–20 minutes, and then dyed with 0.05% crystal violet for 10–20 minutes. Finally, colonies were counted under a microscope.
Cell Invasion
Collected cells were transferred to a upper compartment coated with Matrigel (356,234, Haoyang Biotechnology Co., Ltd., Shanghai, China), and medium with 20% FBS was added into the lower compartment. The insert was incubated at 37°C for 24 h, and then the cells were immobilized with PFA for 10 min after being washed with PBS three times. Then the cells were dyed with 0.5% crystal violet. Finally, invasive cells were counted under a microscope.
Cell Apoptosis
Cell apoptosis was detected by a cell apoptosis kit (BW3302, Biomiga Medical Technology Co., Ltd., Hangzhou, China) as follows: The transfected cells were trypsinized, and then the cells were washed with PBS twice, and then mixed with 100 μL binding buffer to produce 1*106cells/mL suspension. The suspension was mixed with 10 μL AnnexinV-FITC and 10 μL PI in sequence, cultured at room temperature in the dark for 5 min, and finally determined with a DxFLEX flow cytometer (Beckman Coulter Trading (China) Co., Ltd., Shanghai, China).
Luciferase Determination
Cells were transferred to a 24-well plate. After 24 h, ZEB1 wild-type (ZEB1-Wt) and ZEB1 mutant (ZEB1-Mut) were constructed by a dual luciferase reporter (DLR) gene assay kit (SLDL-100, Bei Nuo Biotechnology Co., Ltd., Shanghai, China) and transfected into cells. After 48 h, the luciferase activity of the cells was determined.
Xenotransplantation Tumor Model of Mouse
We purchased 12 female BALB/c nu/nu nude mice (4 weeks old and 18–22 g in weight) from Cavens Experimental Animal Co., Ltd., Changzhou, China, and raised them in specific pathogen-free conditions under 12 h light/12 h dark cycle and studied them. The animal study was approved by the Animal Ethics Committee of our hospital (Shanxi Provincial People’s Hospital), and it was carried out in strict accordance with the Laboratory animal—Guideline for ethical review of animal welfare (Standardization Administration of the People’s Republic of China) for protecting animals. First, the mice were fed for 1 week, and then randomly divided into two groups (MOCK group and miR-139-5p group). H1HeLa cells transfected with miR-NC or miR-139-5p mimics (107 cells per mouse) were injected subcutaneously into the left armpit of mice in the two groups. On the 21st day, mice were anesthetized with chloral hydrate (A4988, Shifeng Biological Technology Co., Ltd., Shanghai, China) and euthanized (decapitated), and their tumors were removed, measured and weighed.
Statistical Analyses
In the present study, all assays were repeated three times and the collected data were analyzed statistically and visualized into figures via GraphPad 6. Data differences were analyzed using the independent t-test, one-way ANOVA, LSD-t-test, and Bonferroni post hoc test. In addition, receiver operating characteristic (ROC) curves were drawn to analyze the area under the curve (AUC) of miR-139-5p in diagnosing CC, and Pearson correlation analysis was carried out to analyze the relationship between miR-139-5p expression in the serum of patients with CC and that in cancerous tissues of the patients, and the correlation between serum miR-139-5p and serum ZEB1. P< 0.05 indicates a remarkable difference.
Results
MiR-139-5p Was Down-Regulated in Cancerous Tissues and Serum of Patients with CC, While ZEB1 Was Up-Regulated in Them
For the purpose of verifying our assumption, we quantified miR-139-5p in the cancerous tissues and serum specimens from patients with CC, finding that miR-139-5p was lowly expressed in both specimens. The results implied that miR-139-5p may serve as a cancer suppressor in CC. Further investigation revealed that the miR-139-5p expression in the two specimens was positively correlated, suggesting that miR-139-p may have a certain clinical value as a non-invasive serum indicator. We also found that the expression of ZEB1 in the two samples was significantly opposite to that of miR-139-5p in them, and the expression of miR-139-5 and ZEB1 in serum was significantly negatively correlated. Then, we drew ROC curves of miR-139-p and ZEB1 in identifying CC as serum indicators, and found that the AUC of miR-139-5 for diagnosing CC was as high as 0.923, while that of ZEB1 was 0.890, indicating that they may be used as auxiliary indicators for CC screening Figure 1.
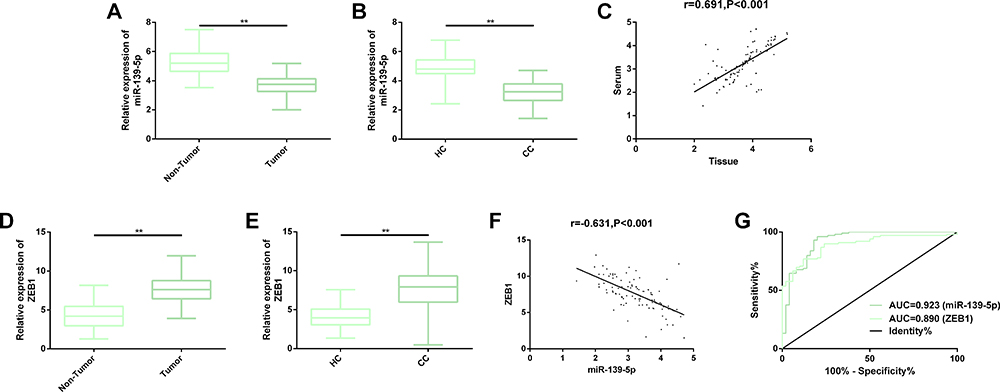 |
Figure 1 Expression of miR-139-5p and ZEB1 in the cancerous tissues and serum of patients with CC. (A, B) MiR-139-5p was down-regulated in both cancerous tissues and serum of patients with CC. (C) The expression of miR-139-5p in cancerous tissues was strongly positively correlated with that in serum. (D, E) The expression of ZEB1 was up-regulated in cancerous tissues and serum of patients with CC. (F) MiR-139-5p and ZEB1 in serum were negatively correlated. (G) AUC of serum miR-139-5p in identifying CC exceeded 0.900, and that of serum ZEB1 for identifying CC exceeded 0.850. Abbreviations: miR, microRNA; CC, cervical cancer; AUC, area under the curve; HC, healthy control; ZEB1, zinc finger E-box-binding homeobox 1. Note: **Indicates that in terms of inter-group comparison, P<0.01. |
MiR-139-5p Was Linked to the FIGO Staging and Pathological Differentiation of Patients with CC
With the aim of exploring the potential correlation between miR-139-5p and pathological parameters ofCC patients, we analyzed the relative expression of miR-139-5p in patients with different pathological parameters, finding that miR-139-5p was not strongly linked to menopause, age, tumor diameter, histological grading, as well as squamous cell carcinoma antigen (SCC-Ag) (all P>0.05), but low miR-139-5p expression was significantly linked to high FIGO staging and low differentiation (both P<0.05). The results imply that miR-139-5p may be helpful to predict FIGO staging and differentiation of patients in Table 1.
MiR-139-5p Could Inhibit the Growth, Invasion, and EMT of CC Cells
In order to explore the potential anticancer effect of miR-139-5p in CC, we analyzed its effect on the biological behaviors of CC cells. First of all, we found that miR-139-5p was generally expressed at low levels in CC cells, especially in H1HeLa and C-33 A cells, so we mainly studied H1HeLa and C-33A cells. We transfected miR-139-5p mimics and miR-139-5p inhibition sequence into H1HeLa and C-33 A cells to overexpress and underexpress miR-139-5p in them, respectively. The cellular function test revealed that down-regulating miR-139-5p accelerated the malignant proliferation, invasion, colony formation, and EMT of CC cells, inhibited the apoptosis rate, decreased the levels of pro-apoptotic factors (Caspase-3 and Bax), and increased the level of anti-apoptotic factor (Bcl-2), but the above results were significantly reversed after miR-139-5p was up-regulated: The malignant behaviors of CC cells were strongly suppressed and the cell apoptosis was induced. The results implied that regulating miR-139-5p could control the malignant development of CC cells, and miR-139-5p had certain cancer inhibiting effect in CC Figure 2.
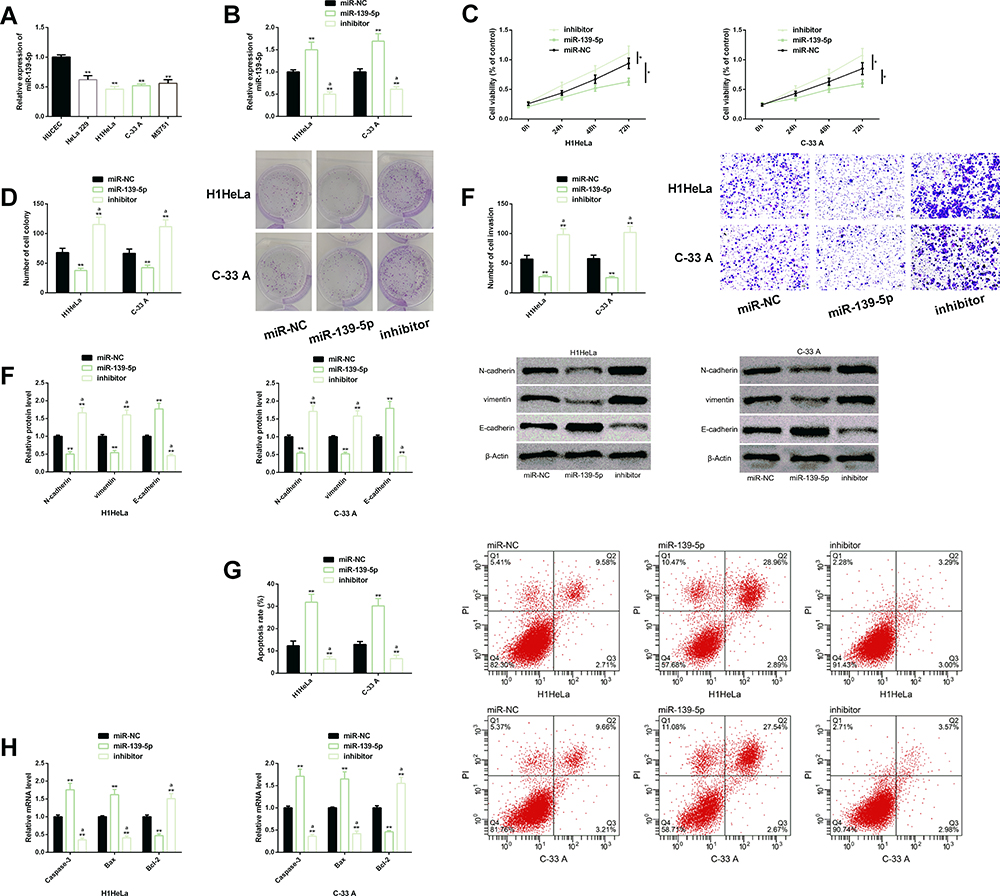 |
Figure 2 Effects of miR-139-5p on the biological function of CC cells. (A) MiR-139-5p was significantly down-regulated in CC cells. (B) Transfection efficiency of miR-139-5p. (C–F) Up-regulation of miR-139-5p significantly inhibited the malignant proliferation, colony formation, invasion, EMT of CC cells. Protein profiling of EMT-related factors of CC cells. (G) Up-regulation of miR-139-5p significantly promoted the apoptosis rate of CC cells. Its cell flow cytometry profiling. (H) Down-regulation of ZEB1 increased the expression of Caspase-3 and Bax, but inhibited the expression of Bcl-2. Abbreviations: miR, microRNA; CC, cervical cancer; EMT, epithelial–mesenchymal transition; NC, negative control; PI, propidium iodide. Notes: In terms of inter-group comparison or comparison with miR-NC, *Indicates P<0.05 and **Indicates P<0.01; in terms of comparison with miR-139-5p, aIndicates P<0.01 in terms of comparison with miR-139-5p. |
MiR-139-5p Negatively Affected the Transcription and Protein Level of ZEB1
For the purpose of further understanding the mechanism of miR-139-5p in CC, we also analyzed its potential downstream targets. Based on TargetScan (http://www.targetscan.org/vert_72/), we found potential binding locus between miR-139-5p and ZEB1, so we carried out analysis for further verification. The DLR revealed that miR-139-5p mimics down-regulated only ZEB1-Wt (not ZEB1-Mut), and the Western blot assay showed that miR-139-5p negatively regulated the transcription and protein level of ZEB1, which suggested that miR-139-5p could regulate ZEB1 negatively (Figure 3).
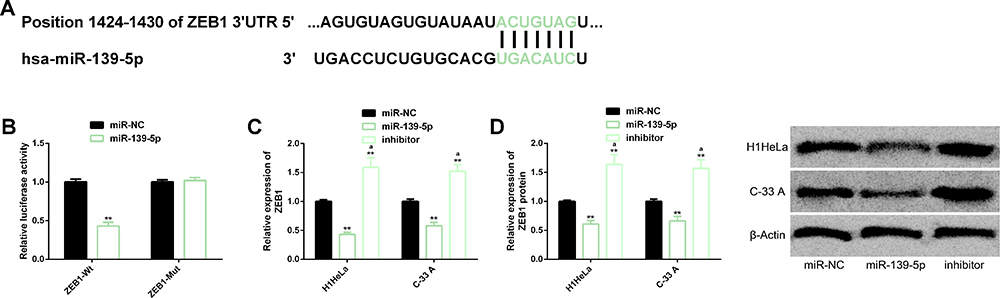 |
Figure 3 Targeting regulatory relationship between miR-139-5p and ZEB1. (A) Binding locus between miR-139-5p and ZEB1. (B) DLR result. (C, D) Up-regulation of miR-139-5p down-regulated the transcription and protein level of ZEB1. Its protein profiling. Abbreviations: miR, microRNA; ZEB1, zinc finger E-box-binding homeobox 1; DLR, dual luciferase reporter; NC, negative control; Wt, wild type; Mut, mutant. Notes: **Indicates P<0.01 in terms of comparison with the miR-NC group. aIndicates P<0.01 in terms of comparison with the miR-139-5p group. |
ZEB1 Was Up-Regulated in CC Cells and Could Promote the Growth, Invasion, and EMT of CC Cells
We also explored whether ZEB1 had influence on the malignant function of CC cells. First of all, we quantified ZEB1 in CC cells, and found that it was generally abnormally up-regulated, indicating that it may also be involved in the malignant development of CC cells. Further analysis of cell behaviors showed that up-regulation of ZEB1 promoted the malignant proliferation, colony formation, invasion, and EMT of CC cells, strongly inhibited cell apoptosis, decreased the expression of Caspase-3 and Bax, and increased the expression of Bcl-2, while knock-down of ZEB1 significantly inhibited the malignant function of CC cells and intensified their apoptosis. The results implied that both down-regulating ZEB1 and up-regulating miR-139-5p had significant anti-tumor effects on CC cells (Figure 4).
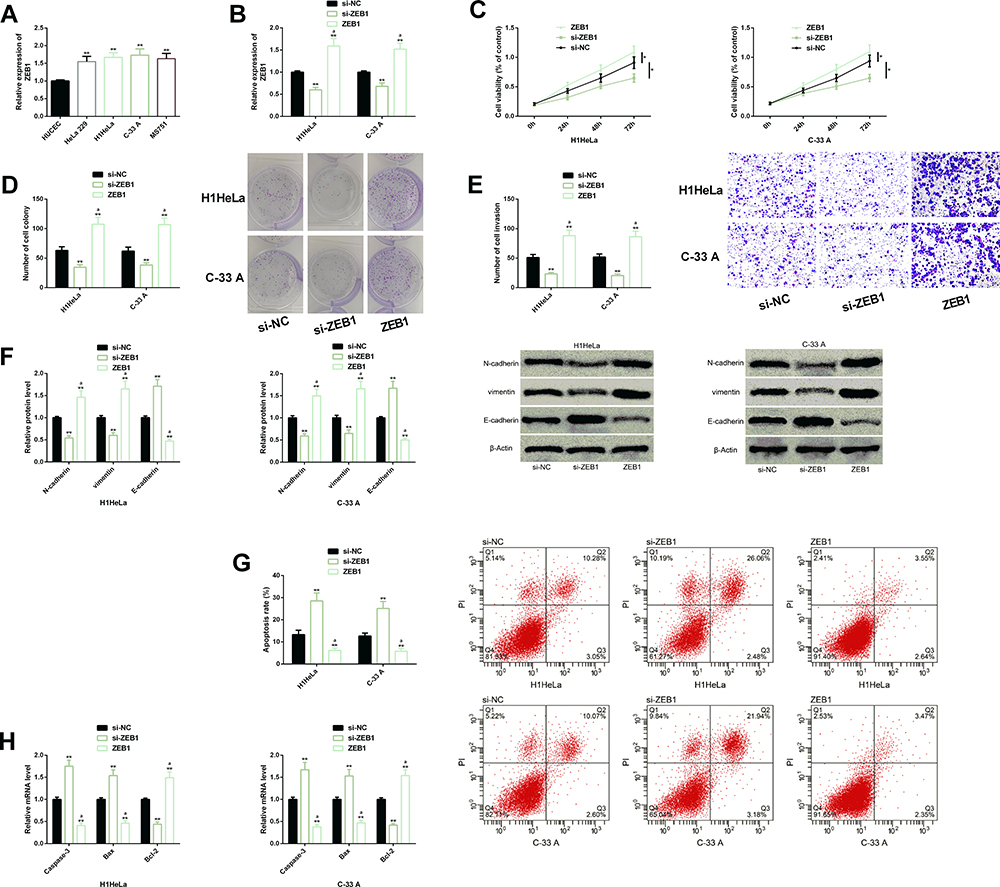 |
Figure 4 Effects of ZEB1 on the biological function of CC cells. (A) ZEB1 was generally highly expressed in CC cells. (B) Transfection efficiency of ZEB1. (C–F) Down-regulation of ZEB1 significantly inhibited the malignant proliferation, colony formation, invasion, EMT of CC cells. Protein profiling of EMT-related factors. (G) Down-regulation of ZEB1 significantly promoted the apoptosis rate of CC cells. Its cell flow cytometry profiling. (H) Down-regulation of ZEB1 increased the expression of Caspase-3 and Bax, but inhibited the expression of Bcl-2. Abbreviations: ZEB1, zinc finger E-box-binding homeobox 1; CC, cervical cancer; EMT, epithelial–mesenchymal transition; si, short interfering; NC, negative control; PI, propidium iodide; Bax, Bcl-2-associated X protein; Bcl-2, B-cell lymphoma-2. Notes: In terms of inter-group comparison or comparison with si-NC, *Indicates P<0.05 and **Indicates P<0.01; in terms of comparison with ZEB1, aIndicates P<0.01 |
Down-Regulating MiR-139-5p Could Weaken the Anti-Tumor Activity of Transfecting Si-ZEB1 in CC Cells
We carried out a co-transfection experiment about miR-139-5p and ZEB1, finding that down-regulating miR-139-5p could reverse the inhibiting effect of down-regulating ZEB1 on the malignant proliferation, invasion, colony formation, as well as EMT of CC cells and induction of it on the cell apoptosis (including the effect on apoptotic factor), which implied that miR-139-5p regulated the biological behaviors of CC cells through targeting ZEB1 (Figure 5).
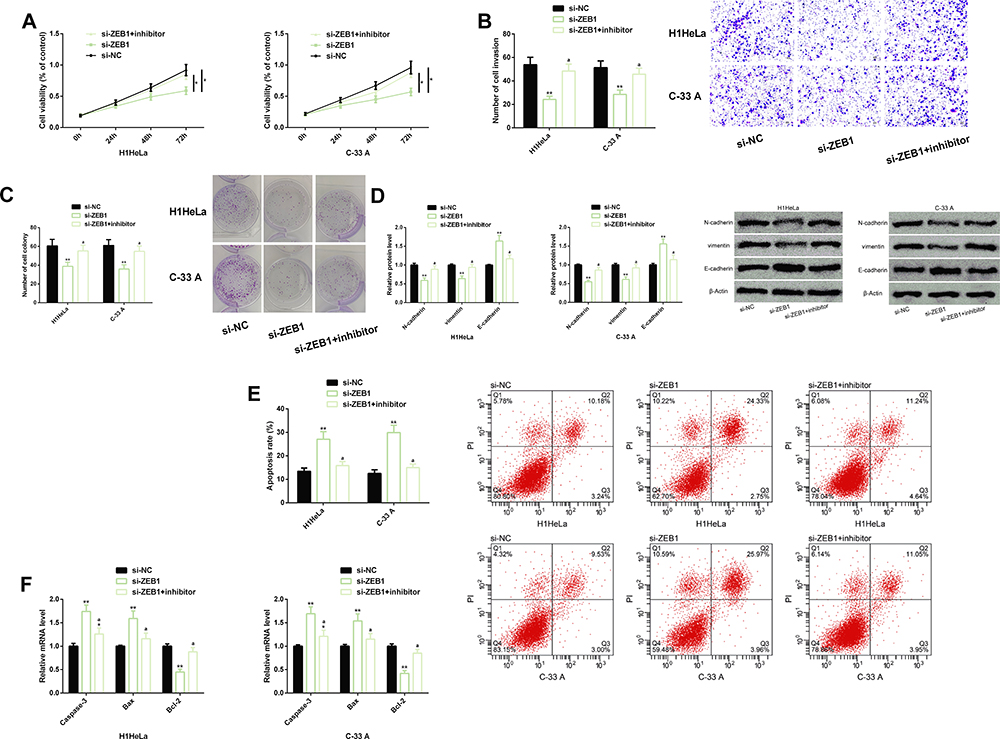 |
Figure 5 Effects of down-regulating miR-139-5p on the anti-tumor activity of si-ZEB1 in CC cells. (A–D) Inhibiting miR-139-5p could reverse the inhibiting effect of down-regulating ZEB1 on the malignant proliferation, invasion, and colony formation, and EMT of CC cells. Protein profiling of EMT-related factors. (E) Inhibiting miR-139-5p could eliminate the induction of si-ZEB1 on the apoptosis of CC cells. Cell flow cytometry profiling. (F) Inhibiting miR-139-5p could eliminate the induction of si-ZEB1 on pro-apoptotic factors (Caspase-3 and Bax) and the inhibition of it on anti-apoptotic factor (Bcl-2). Abbreviations: miR, microRNA; ZEB1, zinc finger E-box-binding homeobox 1; CC, cervical cancer; EMT, epithelial–mesenchymal transition; si, short interfering; NC, negative control; PI, propidium iodide; Bax, Bcl-2-associated X protein; Bcl-2, B-cell lymphoma-2. Notes: In terms of inter-group comparison or comparison with miR-NC, *Indicates P<0.05 and **Indicates P<0.01; in terms of comparison with miR-139-5p, aIndicates P<0.01. |
Up-Regulation of MiR-139-5p Can Inhibit Tumor Growth in CC Xenotransplantation Model Mice
To study the effect of miR-139-5p on the tumor growth of CC model in vivo, H1HeLa cells transfected with miR-NC or miR-139-5p mimetic were inoculated into the left armpit of nude mice. The results showed that at 4 weeks after inoculation, up-regulation of miR-139-5p significantly inhibited the tumor volume and mass in CC xenotransplantation model mice compared with the MOCK group (Figure 6).
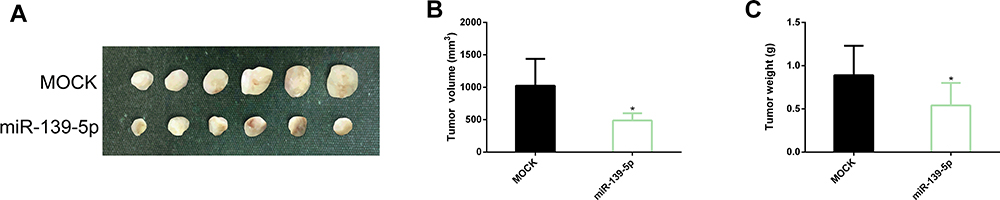 |
Figure 6 Effect of up-regulating miR-139-5p on tumor growth in CC xenotransplantation model mice. (A) Tumor experimental map of CC xenotransplantation model mice in the two groups. (B, C) Up-regulation of miR-139-5p could inhibit tumor volume and mass in CC xenotransplantation model mice. Abbreviations: miR, microRNA; CC, cervical cancer. Note: *Indicates P<0.05 vs. the MOCK group. |
Discussion
The etiology of CC is strongly linked to human papillomavirus (HPV) infection. As a fatal gynecologic tumor, CC shows a continuously increasing incidence in some countries, which results in great cancer burden to women.20,21 At present, CC is intractable, which is mainly manifested in drug resistance and lack of effective treatment strategies for advanced or recurrent CC.22 A large body of evidence has verified that the abnormal dysregulation of miRNA is linked to tumor progression,23,24 so analyzing the molecular mechanism of miRNA in CC is of great value for improving treatment schemes, prevention, and management of CC.
According to clinical analysis, miR-139-5p was expressed at low levels in cancerous tissues and serum of patients with CC, while ZEB1 was expressed at high levels in them, and both serum miR-139-5p and ZEB1 had relatively high diagnostic value (AUC=0.923 and 0.890, respectively). Therefore, we believed that the two may have good diagnostic value in CC., and the former may inhibit cancer, while the latter may induce cancer. One study by Miyoshi et al25 has pointed out that serum miR-139-5p in patients with CRC is helpful to predict tumor recurrence and metastasis. In this study, further analysis showed that low miR-139-5p level was strongly linked to high FIGO staging and low pathological differentiation, indicating that miR-139-5p level also has certain predictive value for pathological parameters of patients. According to one report by Wang et al,26 low miR-139-5p level is linked to FIGO staging and lymph node metastasis of patients with ovarian cancer, which is different from the results of our study. Similarly, one study by Ji et al11 has revealed that the low miR-139-5p level was strongly linked to lymph node metastasis of patients with CC. We inferred that miR-139-5p may also be adopted to predict the prognosis of patients, but we did not get data about it in this study, so we were unable to further verify it. In addition to cases with CC, miR-139-5p is also lowly expressed in cases with non-small cell lung cancer (NSCLC), low-grade glioma in children, as well as prostate cancer, implying that miR-139-5p may have relatively extensive cancer inhibition performance.27–29 For example, it exerts anti-cancer effect in NSCLC by inhibiting oncogenic gene, c-Met, in a targeted manner, and suppresses the survival of low-grade glioma cells in children by disabling oncogenic signaling pathway conduction (PI3K-AKT pathway) in them. In addition, it also hinders the progression of prostate cancer by lowering SOX5. On the other hand, ZEB1 is not only highly expressed in cases with CC, but also overexpressed in cases with diseases such as esophageal squamous cell carcinoma, liver cancer, and cutaneous squamous cell carcinoma, which suggests that it may have extensive carcinogenicity.30–32
An increasing number of scholars have shown interest in studying the influence of miR-139-5p on gynecologic tumors, and have published many studies. For instance, Liu et al33 have reported that miR-139-5p can suppress the viability and migration of endometrial carcinoma cells through targeted inhibition on HOXA10. Liu et al34 have pointed out that miR-139-5p can suppress the progression of ovarian cancer by regulating downstream target, ROCK2, under the targeted control of TTN-AS1. Furthermore, Ji et al11 have verified that miR-139-5p can mediate TCF4 and inactivate the Wnt/β-catenin signal, thereby exerting potential therapeutic effect on CC. In our study, miR-139-5p exerted its anticancer ability in CC by inhibiting the proliferation, colony formation, invasion, and EMT of CC cells and intensifying their apoptosis, increasing the expression of pro-apoptotic factors (Caspase-3 and Bax) and decreasing the expression of anti-apoptotic factor Bcl-2. Further exploration on the mechanism of miR-139-5p revealed that it had a targeted relationship with ZEB1, and could negatively regulate the level of ZEB1, and down-regulating ZEB1 could suppress the aforementioned malignant biological behaviors of CC cells and induce apoptosis. ZEB1 has also been found to be carcinogenic in a variety of gynecological tumors. For instance, Chen et al35 have reported that ZEB1 can promote the accumulation of tumor-related macrophages in the hypoxic tumor microenvironment, thus aggravating the malignant progression of CC Cui et al36 have also reported that ZEB1 is related to the resistance of ovarian cancer cells against chemotherapy, and targeted regulation of ZEB1-SLC3A2 is conducive to strengthening cisplatin-related chemosensitivity. We down-regulated the two indexes meantime, and found that down-regulating miR-139-5p could strongly eliminate the anti-tumor activity after down-regulation of ZEB1. Namely, the inhibitory effect of transfecting si-ZEB1on the malignant progression of CC cells was greatly weakened. All data suggest that miR-139-5p can ameliorate the malignant procession of CC cells through targeted inhibition on ZEB1, and the miR-139-5p-ZEB1 axis had molecular regulatory influence on the growth, invasion, as well as EMT of CC cells. Finally, we also found through xenotransplantation assay that up-regulation of miR-139-5p could inhibit the growth of tumor in CC mice, namely reducing the volume and mass of tumor, which indicated that over-expression of miR-139-5p had a positive effect on inhibiting the development of tumor in CC mice. Based on the above research results, we believe that the development of preparations for targetedly overexpressing miR-139-5p or ZEB1 inhibitors may be helpful to the treatment of patients with CC.
Our study has confirmed the functional expression of miR-139-5p-ZEB1 in CC, but it still has some room for improvement. First of all, we can supplement the relevant research on whether miR-139-5p has upstream factors in the regulatory mechanism of CC to further expand the molecular regulatory network. In addition, we can also explore the effect of miR-139-5p on miR-139-5p-ZEB1 signal transduction pathway by analyzing whether the molecular action of the pathway has reverse influence on miR-139-5p. Furthermore, we can analyze the influence of miR-139-5p on the chemical sensitivity of CC and explore whether miR-139-5p is helpful to alleviate the drug resistance of patients. Finally, we can supplement the detection of inflammatory factors to further explore the effect of miR-139-5p on inflammation in pathological process of CC. In the future, we will gradually improve our research from the above points.
To sum up, we have proposed for the first time that miR-139-5p-ZEB1 axis can exert molecular regulation on the growth, invasion, and EMT of CC, which may provide a new direction for treating CC.
Disclosure
The authors report no conflicts of interest for this work.
References
1. Kim H, Lee KK, Heo MH, Kim JY. The prognostic factors influencing overall survival in uterine cervical cancer with brain metastasis. Korean J Intern Med. 2019;34(6):1324–1332.
2. Zhang Y, Guo X, Wang G, et al. Real-world study of the incidence, risk factors, and prognostic factors associated with bone metastases in women with uterine cervical cancer using surveillance, Epidemiology, and End Results (SEER) data analysis. Med Sci Monit. 2018;24:6387–6397. doi:10.12659/MSM.912071
3. Arbyn M, Weiderpass E, Bruni L, et al. Estimates of incidence and mortality of cervical cancer in 2018: a worldwide analysis. Lancet Glob Health. 2020;8(2):e191–e203. doi:10.1016/S2214-109X(19)30482-6
4. Yao S, Xu J, Zhao K, et al. Down-regulation of HPGD by miR-146b-3p promotes cervical cancer cell proliferation, migration and anchorage-independent growth through activation of STAT3 and AKT pathways. Cell Death Dis. 2018;9(11):1055. doi:10.1038/s41419-018-1059-y
5. Park S, Eom K, Kim J, et al. MiR-9, miR-21, and miR-155 as potential biomarkers for HPV positive and negative cervical cancer. BMC Cancer. 2017;17(1):658. doi:10.1186/s12885-017-3642-5
6. Liu S, Song L, Zeng S, Zhang L. MALAT1-miR-124-RBG2 axis is involved in growth and invasion of HR-HPV-positive cervical cancer cells. Tumour Biol. 2016;37(1):633–640. doi:10.1007/s13277-015-3732-4
7. Xin M, Qiao Z, Li J, et al. miR-22 inhibits tumor growth and metastasis by targeting ATP citrate lyase: evidence in osteosarcoma, prostate cancer, cervical cancer and lung cancer. Oncotarget. 2016;7(28):44252–44265. doi:10.18632/oncotarget.10020
8. Qu ZY, Cui GY, Shi PJ, Wang HQ. Potential suppressive functions of microRNA-504 in cervical cancer cells malignant process were achieved by targeting PAICS and regulating EMT. Arch Gynecol Obstet. 2020;302(1):173–182. doi:10.1007/s00404-020-05538-x
9. Zong ZH, Liu Y, Chen S, Zhao Y. Circ_PUM1 promotes the development of endometrial cancer by targeting the miR-136/NOTCH3 pathway. J Cell Mol Med. 2020;24(7):4127–4135. doi:10.1111/jcmm.15069
10. Wu N, Song H, Ren Y, Tao S, Li S. DGUOK-AS1 promotes cell proliferation in cervical cancer via acting as a ceRNA of miR-653-5p. Cell Biochem Funct. 2020.
11. Ji X, Guo H, Yin S, Du H. miR-139-5p functions as a tumor suppressor in cervical cancer by targeting TCF4 and inhibiting Wnt/beta-catenin signaling. Onco Targets Ther. 2019;12:7739–7748. doi:10.2147/OTT.S215796
12. Li J, Li Q, Lin L, et al. Targeting the Notch1 oncogene by miR-139-5p inhibits glioma metastasis and epithelial-mesenchymal transition (EMT). BMC Neurol. 2018;18(1):133.
13. Fang J, Huang C, Ke J, et al. lncRNA TTN-AS1 facilitates proliferation, invasion, and epithelial-mesenchymal transition of breast cancer cells by regulating miR-139-5p/ZEB1 axis. J Cell Biochem. 2020. doi:10.1002/jcb.29700
14. Yang C, Sun J, Liu W, et al. Long noncoding RNA HCP5 contributes to epithelial-mesenchymal transition in colorectal cancer through ZEB1 activation and interacting with miR-139-5p. Am J Transl Res. 2019;11(2):953–963.
15. Yue S, Wang L, Zhang H, et al. miR-139-5p suppresses cancer cell migration and invasion through targeting ZEB1 and ZEB2 in GBM. Tumour Biol. 2015;36(9):6741–6749. doi:10.1007/s13277-015-3372-8
16. Xu J, Wang H, Wang H, et al. The inhibition of miR-126 in cell migration and invasion of cervical cancer through regulating ZEB1. Hereditas. 2019;156:11.
17. Gan L, Chen Y, Liu H, Ju WH. Long non-coding RNA ZEB1-antisense 1 affects cell migration and invasion of cervical cancer by regulating epithelial-mesenchymal transition via the p38MAPK signaling pathway. Gynecol Obstet Invest. 2019;84(2):136–144. doi:10.1159/000493265
18. Kuhn L, Saidu R, Boa R, et al. Clinical evaluation of modifications to a human papillomavirus assay to optimise its utility for cervical cancer screening in low-resource settings: a diagnostic accuracy study. Lancet Glob Health. 2020;8(2):e296–e304. doi:10.1016/S2214-109X(19)30527-3
19. Canaz E, Ozyurek ES, Erdem B, et al. Preoperatively assessable clinical and pathological risk factors for parametrial involvement in surgically treated FIGO stage IB-IIA cervical cancer. Int J Gynecol Cancer. 2017;27(8):1722–1728. doi:10.1097/IGC.0000000000001060
20. Liu M, Wang Z, Liu Q, Zhu H, Xu N. Expression of micro-RNA-492 (MiR-492) in human cervical cancer cell lines is upregulated by transfection with wild-type P53, irradiation, and 5-fluorouracil treatment in vitro. Med Sci Monit. 2018;24:7750–7758. doi:10.12659/MSM.911585
21. Msyamboza KP, Phiri T, Sichali W, Kwenda W, Kachale F. Cervical cancer screening uptake and challenges in Malawi from 2011 to 2015: retrospective cohort study. BMC Public Health. 2016;16(1):806. doi:10.1186/s12889-016-3530-y
22. Rui X, Xu Y, Jiang X, Ye W, Huang Y, Jiang J. Long non-coding RNA C5orf66-AS1 promotes cell proliferation in cervical cancer by targeting miR-637/RING1 axis. Cell Death Dis. 2018;9(12):1175. doi:10.1038/s41419-018-1228-z
23. Babion I, Jaspers A, van Splunter AP, van der Hoorn IAE, Wilting SM, Steenbergen RDM. miR-9-5p exerts a dual role in cervical cancer and targets transcription factor TWIST1. Cells. 2019;9:1. doi:10.3390/cells9010065
24. Gu X, Dong M, Liu Z, Yang J, Shi Y. MiR-499a-5p inhibits proliferation, invasion, migration, and epithelial-mesenchymal transition, and enhances radiosensitivity of cervical cancer cells via targeting eIF4E. Onco Targets Ther. 2020;13:2913–2924. doi:10.2147/OTT.S241631
25. Miyoshi J, Toden S, Yoshida K, et al. MiR-139-5p as a novel serum biomarker for recurrence and metastasis in colorectal cancer. Sci Rep. 2017;7:43393. doi:10.1038/srep43393
26. Wang Y, Li J, Xu C, Zhang X. MicroRNA-139-5p inhibits cell proliferation and invasion by targeting RHO-associated coiled-coil-containing protein kinase 2 in ovarian cancer. Oncol Res. 2018;26(3):411–420. doi:10.3727/096504017X14974343584989
27. Sun C, Sang M, Li S, et al. Hsa-miR-139-5p inhibits proliferation and causes apoptosis associated with down-regulation of c-Met. Oncotarget. 2015;6(37):39756–39792. doi:10.18632/oncotarget.5476
28. Catanzaro G, Besharat ZM, Miele E, et al. The miR-139-5p regulates proliferation of supratentorial paediatric low-grade gliomas by targeting the PI3K/AKT/mTORC1 signalling. Neuropathol Appl Neurobiol. 2018;44(7):687–706.
29. Yang B, Zhang W, Sun D, et al. Downregulation of miR-139-5p promotes prostate cancer progression through regulation of SOX5. Biomed Pharmacother. 2019;109:2128–2135. doi:10.1016/j.biopha.2018.09.029
30. Zhao YL, Li JB, Li YJ, Li SJ, Zhou SH, Xia H. Capn4 promotes esophageal squamous cell carcinoma metastasis by regulating ZEB1 through the Wnt/beta-catenin signaling pathway. Thorac Cancer. 2019;10(1):24–32. doi:10.1111/1759-7714.12893
31. Qin Y, Yu J, Zhang M, Qin F, Lan X. ZEB1 promotes tumorigenesis and metastasis in hepatocellular carcinoma by regulating the expression of vimentin. Mol Med Rep. 2019;19(3):2297–2306.
32. Murata M, Ito T, Tanaka Y, Yamamura K, Furue K, Furue M. OVOL2-mediated ZEB1 downregulation may prevent promotion of actinic keratosis to cutaneous squamous cell carcinoma. J Clin Med. 2020;9(3):618. doi:10.3390/jcm9030618
33. Liu J, Li C, Jiang Y, Wan Y, Zhou S, Cheng W. Tumor-suppressor role of miR-139-5p in endometrial cancer. Cancer Cell Int. 2018;18:51. doi:10.1186/s12935-018-0545-8
34. Liu X, Li Y, Wen J, Qi T, Wang Y. Long non-coding RNA TTN-AS1 promotes tumorigenesis of ovarian cancer through modulating the miR-139-5p/ROCK2 axis. Biomed Pharmacother. 2020;125:109882. doi:10.1016/j.biopha.2020.109882
35. Chen XJ, Deng YR, Wang ZC, et al. Hypoxia-induced ZEB1 promotes cervical cancer progression via CCL8-dependent tumour-associated macrophage recruitment. Cell Death Dis. 2019;10(7):508. doi:10.1038/s41419-019-1748-1
36. Cui Y, Qin L, Tian D, et al. ZEB1 promotes chemoresistance to cisplatin in ovarian cancer cells by suppressing SLC3A2. Chemotherapy. 2018;63(5):262–271. doi:10.1159/000493864
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.