Back to Journals » Clinical Interventions in Aging » Volume 20
MicroRNA-874-3p is a Potential Contributor to Primary Hyperparathyroidism-Induced Osteoporosis
Authors Cheng K, Bai R, Li M, Wei Y, Li Z, Zhao X, Cao R, Wang Z, Tan S, Zha Y, Jiang X ![]() , Lu S
, Lu S ![]()
Received 2 June 2025
Accepted for publication 4 October 2025
Published 18 November 2025 Volume 2025:20 Pages 2079—2089
DOI https://doi.org/10.2147/CIA.S538129
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Zhi-Ying Wu
Kaiyuan Cheng,1,* Ruifeng Bai,2,* Minjuan Li,1 Yongjie Wei,3 Zhigang Li,3 Xian Zhao,4 Renwei Cao,4 Zhongyu Wang,4 Shen Tan,5 Yejun Zha,1 Xieyuan Jiang,1,6 Shuai Lu1,6
1Department of Orthopedic Trauma, Beijing Jishuitan Hospital, Capital Medical University, Beijing, People’s Republic of China; 2Department of Clinical Laboratory, Beijing Jishuitan Hospital, Capital Medical University, Beijing, People’s Republic of China; 3State Key Laboratory of Environmental Criteria and Risk Assessment, Chinese Research Academy of Environmental Sciences, Beijing, People’s Republic of China; 4Department of Orthopedics Trauma, Beijing Jishuitan Hospital, Peking University Fourth School, Beijing, People’s Republic of China; 5Department of General Surgery, Beijing Jishuitan Hospital, Capital Medical University, Beijing, People’s Republic of China; 6Beijing Research Institute of Traumatology and Orthopaedics, Beijing, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Shuai Lu, Department of Orthopedic Trauma, Beijing Jishuitan Hospital, Capital Medical University, Beijing, People’s Republic of China, Email [email protected] Xieyuan Jiang, Department of Orthopedic Trauma, Beijing Jishuitan Hospital, Capital Medical University, Beijing, People’s Republic of China, Email [email protected]
Background: Dysregulation of microRNAs contributes to bone diseases. However, the microRNAs involved in primary hyperparathyroidism (PHPT)-induced osteoporosis remain unknown.
Methods: The parathyroid tissue samples were obtained from PHPT patients with or without osteoporosis (n = 5/group) during parathyroid resection and subjected to high throughput microRNA sequencing. The differentially expressed microRNAs were identified and further verified using qRT-PCR. Alizarin Red Staining was performed to detected the osteogenic differentiation. Gain- and loss-of-function assays were performed to investigate the role of miR-874-3p, which was upregulated in PHPT patients with osteoporosis, in human mesenchymal stem cells (hMSCs) undergoing osteoblastic differentiation.
Results: We identified 32 significantly upregulated and 18 significantly downregulated microRNAs in PHPT patients with osteoporosis. miR-874-3p was increased in PHPT osteoporosis patients, meanwhile, miR-874-3p in parathyroid tissue and peripheral blood extracellular vesicles of PHPT osteoporosis mice were increased. The miR-874-3p level was remarkably elevated in hMSCs grown in osteogenic medium. Overexpression of miR-874-3p repressed the hMSC osteogenic differentiation and reduced the osteogenic marker expression in hMSCs, whereas miR-874-3p inhibitor showed a contrasting effect. The results of the dual luciferase reporting system showed that miR-874-3p could reduce the luciferase activity of wild-type FTO-WT-3 ‘-UTR. However, there was no significant change in the luciferase activity of the mutant compared with the control group.
Conclusion: MiR-874-3p might specifically binds to FTO suppress osteogenic differentiation of hMSCs, thereby contributing to the development of osteoporosis in PHPT patients.
Keywords: microRNAs, primary hyperparathyroidism, osteoporosis, next-generation sequencing, mesenchymal stem cells
Introduction
Primary hyperparathyroidism (PHPT) is an endocrine disorder characterized by hypercalcemia resulting from overproduction of parathyroid hormone (PTH).1 PHPT represents the third most common endocrine disorder, with an incidence ranging from 0.4 to 82 cases per 100,000 individuals worldwide.2,3 PTH stimulates osteoblast differentiation into osteoclasts by acting on PTH receptors in osteoblasts, leading to osteopenia, osteoporosis, and even cyst formation and fibrosis.4 Osteoporosis is the most common bone disease related to PHPT, with a prevalence in PHPT varying from 39% to 62.9%.5–7 Approximately over 80% of patients with osteoporosis have no clinical manifestations until there is a fracture.8 Thus, delayed diagnosis of PHPT-induced osteoporosis is common, leading to decreased quality of life as well as increased disability and mortality.9 Osteoporosis seriously affects the health of the elderly. Currently, the dual-energy X-ray absorptiometry (DXA) measuring bone mineral density (BMD) represents the gold standard for osteoporosis diagnosis.10 However, predicting fracture based on BMD value is unreliable.11,12 Therefore, it is critical to identify new biomarkers for early screening, diagnosis, and treatment of PHPT osteoporosis before fractures occur.
The small non-coding microRNAs are post-transcriptional regulators of multiple physiological and pathological processes through base pairing with mRNA targets, playing important roles in bone metabolism.13 Owing to the high measurability and stability in body fluid, microRNAs are considered promising biomarkers for predicting and monitoring osteoporosis.14 Studies have identified over 50 microRNA families that are associated with osteoporosis, such as miR-21, miR-100, and miR-23-3p.15–17 Although some studies have investigated the microRNA signatures in secondary osteoporosis caused by glucocorticoid, autoimmune disorders, and type 2 diabetes,18–20 the microRNA signatures of PHPT-induced osteoporosis remain unknown.
As an m6A demethylase, FTO can regulate the differentiation direction of bone marrow mesenchymal stem cells (BMSCs) through multiple signaling pathways. On the one hand, FTO can enhance the expression of peroxisome proliferator-activated receptor γ (PPARγ) and CAAT region/enhancer binding proteins, inhibit the Wnt/β -catenin signaling pathway, thereby promoting adipogenesis, reducing the differentiation of BMSC into osteoblasts, and leading to osteopenia and osteoporosis.21 On the other hand, FTO-mediated 3 ‘-UTR demethylation of M6A-peroxisome proliferator-activated receptor mRNA reduces the stability of peroxisome proliferator-activated receptor mRNA in a YTHDF1-dependent manner, thereby affecting the differentiation and function of osteoblasts. FTO also regulates the fate of BMSC during osteoporosis through the growth differentiation factor 11-FTO-peroxisome proliferator-activated receptor γ axis, promoting their transformation into adipocytes and inhibiting bone formation.22
Next-generation sequencing-based small RNA sequencing (sRNA-seq) is currently the most powerful tool for microRNA profiling and has been used to identify differentially expressed microRNAs in osteoporosis patients.23,24 In this study, we aimed to identify osteoporosis-related microRNAs in PHPT patients. Considering that a solitary parathyroid adenoma is responsible for up to 85% of PHPT and double adenomas for 4%,25 we recruited a cohort of PHPT patients with or without osteoporosis and performed high throughput microRNA sequencing followed by bioinformatics analysis and experimental validation. Our study provides novel information about the microRNAs involved in the development of PHPT-induced osteoporosis.
Materials and Methods
Patients and Specimens
A total of 10 PHPT patients with (n = 5) or without (n = 5) osteoporosis who have undergone parathyroid resection between January 2018 and January 2020 at Beijing Jishuitan Hospital (Beijing, China) were recruited. This study was approved by the Ethics Committee of Beijing Jishuitan Hospital. All subjects provided written informed consent before the study. The inclusion criteria were: 1) parathyroid adenoma was diagnosed by cervical ultrasonography, computed tomography, and pathological examination; 2) the presence or absence of osteoporosis was confirmed by X-ray and DXA. The diagnosis of osteoporosis was made when BMD T-score was below −2.5.26 Patients with tumors, cardiovascular diseases, sepsis, or autoimmune diseases were excluded. The parathyroid tissue samples were obtained during the surgery and stored at −80 °C until use.
Animals
Male C57BL/6J mice (8–10 weeks old) were housed in a specific pathogen-free (SPF) facility at the Animal Experimental Center of Jishuitan Hospital. The animals were maintained under a 12-hour light/dark cycle at a controlled temperature of 22–26 °C, with ad libitum access to water. All procedures complied with the Regulation on the Care and Use of Experimental Animals issued by the Beijing Council on Animal Care (1996), and the protocol was approved by the Jishuitan Hospital Animal Care and Use Committee (2024–09-S004).
PHPT Model
Primary hyperparathyroidism (PHPT) was modeled by tail vein injection of an adenovirus vector carrying 7.5 × 109 Vg of the PTH gene into male C57BL/6J mice, resulting in systemic PTH overexpression and mimicking the pathophysiological features of PHPT. A control group received intraperitoneal injections of physiological saline. Three months post-injection, mice were anesthetized for the collection of parathyroid tissue and peripheral blood to assess disease development.
Total RNA Isolation, Library Construction, and Sequencing
Total RNA was isolated from the parathyroid tissue samples using TRIzol (Tiangen Biotech, Beijing, China) following the manufacturer’s protocol. The purity and quantity of RNA were determined using a NanoPhotometer® device (Implen, Camarillo, CA, USA) and a Qubit® 2.0 kit. The RNA integrity was examined using a Bioanalyzer 2100 system (Agilent Technologies, Santa Clara, CA, USA). For each sample, a total of 3 µg RNA was loaded to generate the sequencing library using a small RNA sample prep kit following the manufacturer’s instructions. After quantification and quality control, the library (1 ng/µL) was sequenced using an Illumina HiSeq 2500 sequencer (Illumina, San Diego, CA, USA). The 50-bp single-end reads were generated for microRNA sequencing.
MicroRNA Sequencing Data Analysis
Clean reads were obtained from raw reads by removing low-quality reads (sequence quality score ≤ 5), ploy-N-containing reads (ambiguous base calls > 10%), ploy-A/T/G/C-containing reads, and 5′ or 3′ adapters. The clean reads were mapped to the human genome using Bowtie25 and aligned with miRBase.27 The size distribution and nucleotide frequencies of the reads were analyzed using miREvo28 and mirdeep2.29 The reads of rRNA, tRNA, snRNA, and snoRNA were removed. The microRNA expression levels were presented as transcript per million and calculated as follows: normalized expression = mapped read count / total reads × 1,000,000.30 Differentially expressed microRNA was analyzed using DESeq R package.31 The microRNAs with |log 2 (fold change)| > 1 and q value < 0.01 were identified as differentially expressed microRNAs and were subjected to Gene ontology (GO) annotation and Kyoto encyclopedia of genes and genomes (KEGG) pathway analysis using GOseq32 and KOBAS.33
Cell Culture and Osteogenic Induction
Human mesenchymal stem cells (hMSCs) were obtained from Otwo Biotech Inc (Shenzhen, China). Cells were cultured in Dulbecco’s modified Eagle’s medium (C11995; Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS) (SH30070.03; HyClone, Logan, UT, USA). After the cells reached confluence, the medium was replaced with osteogenic medium containing 10% FBS, 50 µM L-ascorbic acid (A4403; Sigma-Aldrich, St. Louis, MO, USA), 10 mM glycerol-2-phosphate (G9422; Sigma-Aldrich), and 100 nM dexamethasone (D4902; Sigma-Aldrich).
Overexpression and Knockdown of microRNA
The mimics or inhibitor of miR-874-3p and corresponding negative controls were purchased from RiboBio (Guangzhou, China) and transfected into hMSCs cultured in serum-free Opti-Minimum Essential medium (31985062; Thermo Fisher Scientific) at a final concentration of 200 nM using Lipofectamine RNAiMAX™ (13778150; Thermo Fisher Scientific), as previously described.34 At 24 h after transfection, cells were grown in osteogenic medium for 6 days.
Quantitative Real-Time PCR (qRT-PCR)
Total RNA was isolated using TRIzol (15596018; Thermo Fisher Scientific), followed by cDNA synthesis using the miRcute Plus miRNA First-Strand cDNA Synthesis Kit (KR211; TIANGEN BIOTECH) according to the manufacturer’s instruction. cDNA was amplified using the Luna universal qPCR master mix (M3003L; NEB, Ipswich, MA, USA). The primers were summarized in Table 1. For microRNAs, amplification and detection were performed using the CFX96 Touch Real-Time PCR detection system (Bio-Rad, Hercules, CA, USA) and specific primers (RiboBio). U6 was used as an internal control.
 |
Table 1 Primers for Quantitative Real-Time PCR |
Statistical Analysis
Data were presented as the mean ± standard deviation. Statistical analysis was performed using GraphPad Prism 5 (GraphPad, San Diego, CA, USA). The intergroup comparisons were performed using the two-tailed unpaired Student’s t-test. A P value < 0.05 was considered statistically significant.
Results
Identification of Differentially Expressed microRNAs in PHPT Patients with Osteoporosis
About 90% of reads ranged between 21 to 24 nucleotides in length (Supplementary Figure 1A). Pearson correlation analysis showed that the correlation within the group was greater than that between the groups (Supplementary Figure 1B). Position 1 of the reads contained the highest percentages of uridines compared with other positions (Supplementary Figure 1C). These data conform to the characteristics of mammalian microRNAs.35 Taken together, these results suggest an appropriate sampling strategy and reliable data quality of this study.
By comparing the microRNA expression profiles between PHPT patients with and without osteoporosis, we identified 32 significantly upregulated and 18 significantly downregulated microRNAs in PHPT patients with osteoporosis. Hierarchical clustering assay showed that patients in each group displayed similar microRNA expression patterns (Figure 1A). GO analysis showed that the differentially expressed microRNAs target genes were mainly termed as “metabolic process”, “intracellular” component, and “binding” function (Figure 1B). KEGG pathway enrichment analysis revealed that these microRNAs were predominantly enriched in the “apoptosis” pathway (Figure 1C). In differential microRNAs, mir-874-3p target genes are involved in MAPK, Wnt/β-catenin pathways related to osteoporosis, may regulate the progression of osteoporosis.
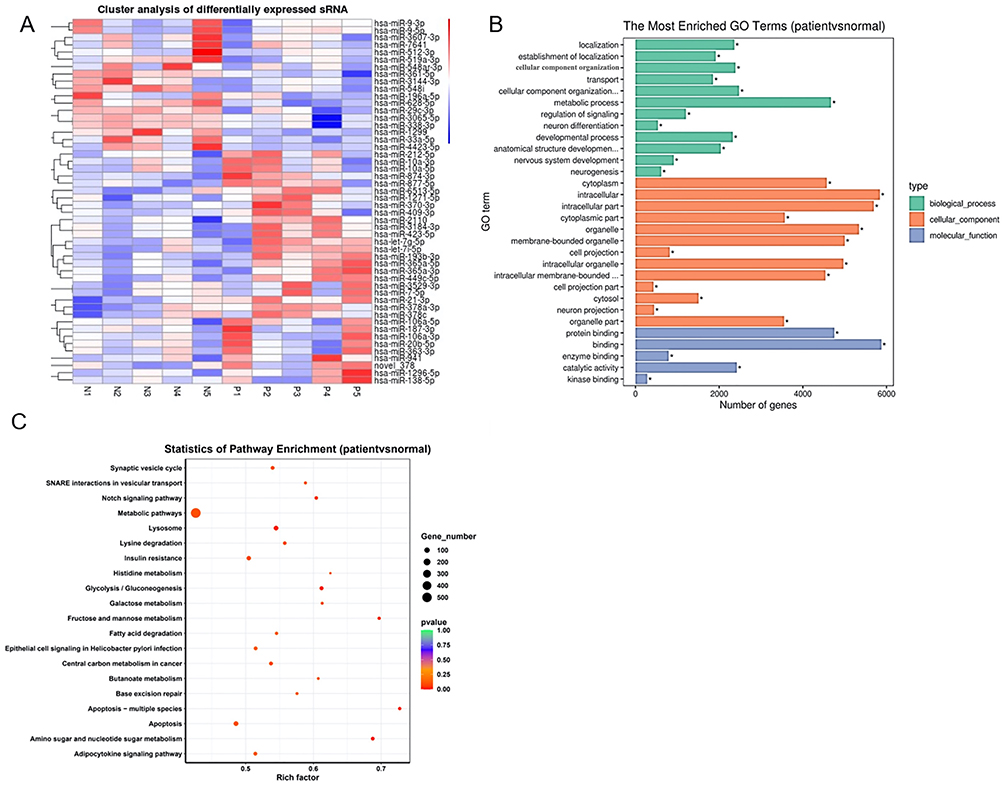 |
Figure 1 Identification of differentially expressed microRNAs in patients with primary hyperparathyroidism (PHPT)-induced osteoporosis. A total of 10 PHPT patients with (n = 5) or without (n = 5) osteoporosis were recruited. (A) Heatmap of hierarchical clustering assay of differentially expressed microRNAs. Upregulated and downregulated microRNAs (adjusted P-value < 0.05) are highlighted in red and blue, respectively. n = 5. (B) Gene ontology annotation of differentially expressed microRNAs. The microRNAs were classified into three functional categories. Twelve biological process annotations, thirteen cell component annotation, and five molecular function annotations are shown. (C) Kyoto encyclopedia of genes and genomes (KEGG) pathway analysis. Top 20 KEGG pathways are shown. *P < 0.05. |
MiR-874-3p Inhibits Osteogenic Marker Expression of hMSCs
To explore the role of miR-874-3p in osteogenic differentiation in hMSCs, we performed gain- and loss-of-function assays. Compared with those in uninduced hMSCs, the mRNA levels of osteogenic markers alkaline phosphatase (ALP) and osteocalcin 2 (OCN 2) were dramatically increased in hMSCs at 14 days after growing in osteogenic medium (Figure 2A), along with a substantial decrease in miR-874-3p level (Figure 2B). Compared with corresponding negative controls, miR-874-3p mimics significantly suppressed the mRNA expression of ALP, OCN2, and Runx2 in hMSCs cultured in osteogenic medium (Figure 2C), whereas miR-874-3p inhibitor exhibited contrasting effects (Figure 2E). In addition, the results of Alizarin Red Staining showed that miR-874-3p mimics significantly inhibited the ostogenic differentiation of hMSCs (Figure 2D), inhibition of miR-874-3p significantly increase the ostogenic differentiation of hMSCs (Figure 2E). These results suggest that miR-874-3p might inhibit osteogenic differentiation of hMSCs, and thus contributing to the development of osteoporosis in patients with PHPT.
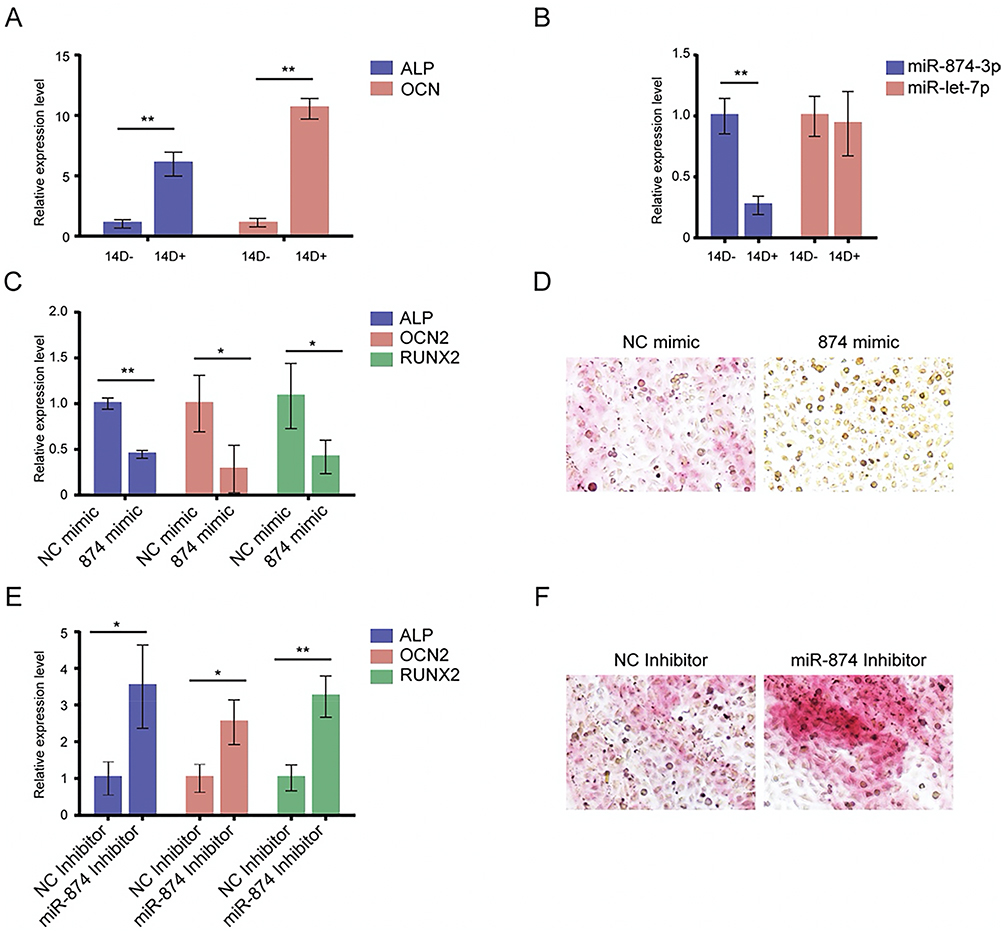 |
Figure 2 MiR-874-3p inhibits osteogenic marker expression of hMSCs. (A) qRT-PCR was performed to determine the mRNA levels of ALP, OCN 2 followed by osteogenic induction in osteogenic medium for 14 days. Data are expressed as the mean ± SD. *P < 0.05, ** P < 0.01, n = 5. ALP, alkaline phosphatase; OCN 2, osteocalcin 2; RUNX2, Runt-related transcription factor 2. (B) qRT-PCR was performed to determine the mRNA levels of let-7p, and miR-874-3p. (C) hMSCs were transfected with miR-874-3p mimics, qRT-PCR was performed to determine the mRNA levels of ALP, OCN 2. (D) Alizarin Red Staining was performed to detected the osteogenic differentiation, x100. Data are expressed as the mean ± standard deviation (SD). n = 5. ALP, alkaline phosphatase; OCN 2, osteocalcin 2; D, days. (E) miR-874-3p inhibitor, or corresponding negative controls for 24 h, qRT-PCR was performed to determine the mRNA levels of ALP, OCN 2. (F) Alizarin Red Staining was performed to detect the osteogenic differentiation, x100. |
Peripheral Blood Extracellular Vesicles miR-874-3p Was Increased in PHPT Osteoporosis Patients
microRNA sequencing results of peripheral blood extracellular vesicles of PHPT patients showed that miR-874-3p in peripheral blood extracellular vesicles of PHPT patients with osteoporosis was significantly up-regulated compared with non-osteoporosis patients (Figure 3C). KEGG results showed that upregulated microRNA was involved in the synthesis and secretion of parathyroid hormone (Figure 3D). The expression of miR-874-3p in serum extracellular vesicles of PHPT patients with osteoporosis and non-osteoporosis was detected by qRT-PCR, and the trend was consistent with the microRNA sequencing results of peripheral blood extracellular vesicles (Figure 3E). It is suggested that miR-874-3p may be released through extracellular vesicles.
 |
Figure 3 Peripheral blood extracellular vesicles miR-874-3p was increased in patients with PHPT osteoporosis (A) Through transmission electron microscopy (TEM) and nano-size analysis (NTA), there was no difference in the morphology and particle size of extracellular vesicles between the control group and the PHPT group. (B) Flow chart of the Construction Method of strand specific Small RNA Library. (C) RNA-sequence volcano map (C-map) showed that miR-874-3p expression was different between the PHPT group and the control group. (D)The expression difference of miR-874-3p in PHPT group and control group KEGG pathway analysis showed that the differentially expressed genes between PHPT group and control group were mainly concentrated in autophagy and lysosomal related pathways. (E) qRT-PCR was performed to determine the mRNA levels of miR-874-3p. ** P < 0.01. |
miR-874-3p in Parathyroid Tissue of PHPT Osteoporosis Mice and miR-874-3p, PTH and Gcm2 Genes in Peripheral Blood Extracellular Vesicles Were Increased
Compared with control mice, the expression of miR-874-3p in the parathyroid tissue of PHPT osteoporosis mice was significantly increased (Figure 4A). The levels of miR-874-3p, PTH and Gcm2 in peripheral blood extracellular vesicles (Figure 4B) and bone marrow MSC extracellular vesicles (Figure 4C) of model mice were significantly higher than those of normal control mice. It is suggested that miR-874-3p may be mediated by extracellular vesicles, but its uptake by bone marrow MSC needs to be further clarified.
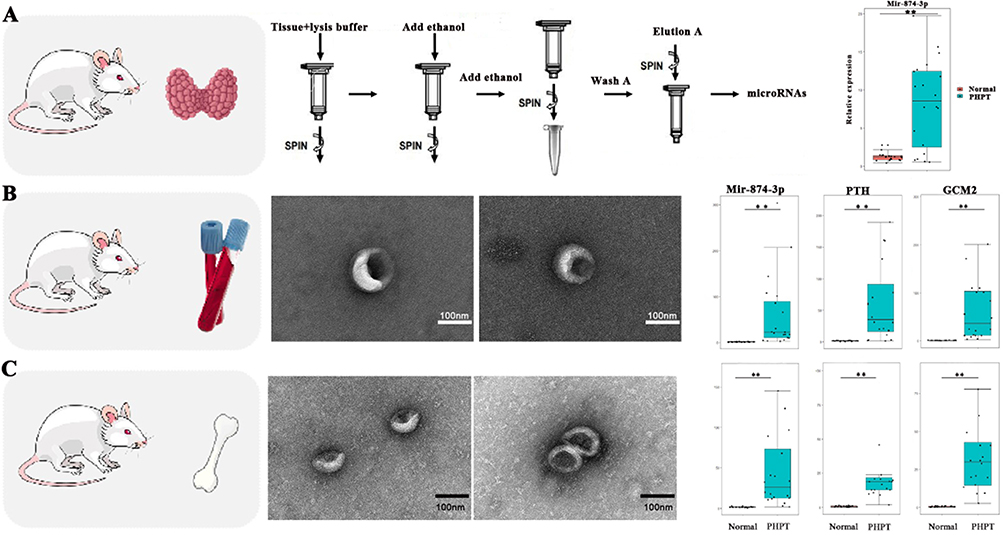 |
Figure 4 miR-874-3p in parathyroid tissue of PHPT osteoporosis mice and miR-874-3p, Pth and Gcm2 genes in peripheral blood extracellular vesicles were increased (A) Expression of miR-874-3p in parathyroid tissue of PHPT osteoporosis mice. (B) Expression of miR-874-3p, Pth and Gcm2 in peripheral blood extracellular vesicles of mice in each group. (C) Expression of miR-874-3p, Pth and Gcm2 in bone marrow MSC extracellular vesicles of mice. ** P < 0.01. |
miR-874-3p Specifically Binds to FTO and Inhibits Its Expression
Bioinformatics related software predicted the targeting relationship between miR-874-3p and fat mass and obesity-associated protein (FTO) (Figure 5A). The results of the dual luciferase reporting system showed that miR-874-3p could reduce the luciferase activity of wild-type FTO-WT-3 ‘-UTR. However, there was no significant change in the luciferase activity of the mutant compared with the control group (Figure 5B). Western Blot showed that overexpression of miR-874-3p decreased the level of FTP protein (Figure 5C). miR-874-3p was found to inhibit the loss of osteogenic differentiation by siRNA knockdown of FTO (Figure 5D-5F). These results indicated that miR-874-3p inhibited osteogenic differentiation by regulating FTO expression.
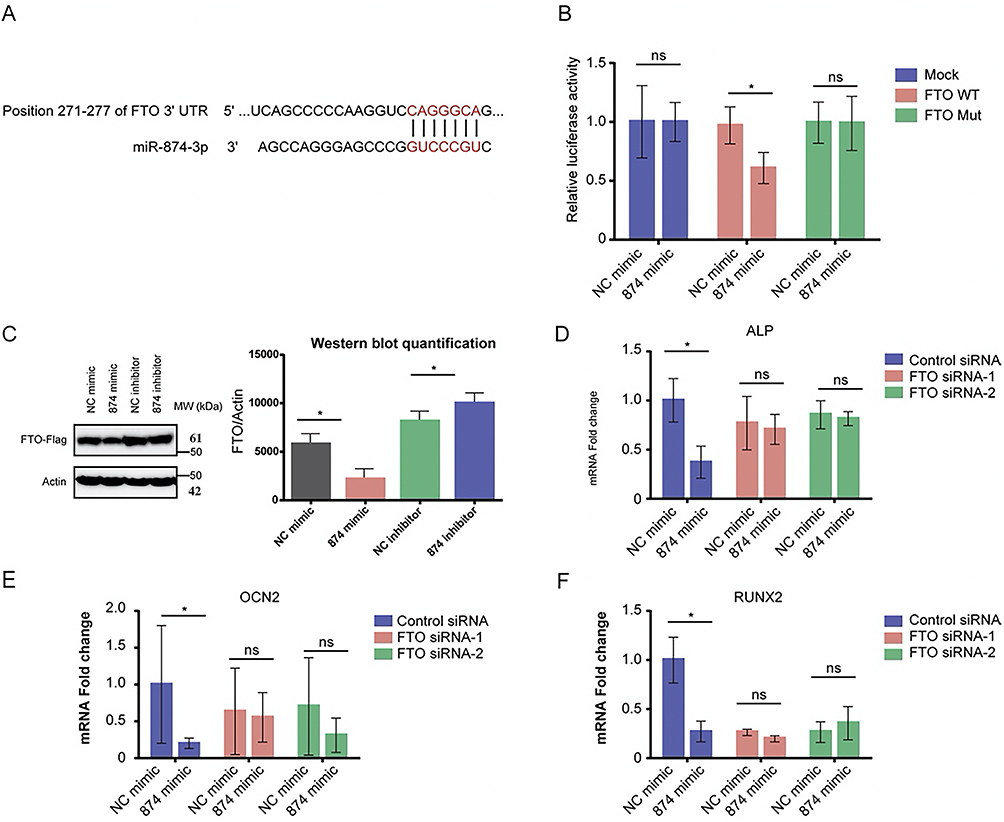 |
Figure 5 miR-874-3p specifically binds to FTO and inhibits its expression (A) Targeting relationship between miR-874-3p and FTO. (B) Mutant luciferase activity and control level. (C) Western Blot analysis of FTP protein levels. (D) mRNA of ALP after siRNA knockdown of FTO. (E) mRNA level of OCN2 with siRNA knockdown of FTO. (F) mRNA level of RUNX2 after siRNA knockdown of FTO. *P < 0.05, ns>0.05. |
Discussion
In this study, we identified 32 significantly upregulated and 18 significantly downregulated microRNAs in PHPT patients with osteoporosis compared with those in PHPT patients without osteoporosis. Of these differentially expressed microRNAs, miR-874-3p was substantially upregulated in hMSCs undergoing osteoblastic differentiation. Overexpression of miR-874-3p suppressed mRNA expression of osteogenic markers in hMSCs cultured in osteogenic medium, whereas the inhibitor of miR-874-3p showed contrasting results. These findings suggest that miR-874-3p might inhibit osteogenic differentiation of hMSCs, serving as a potential diagnostic marker and therapeutic target for PHPT-induced osteoporosis.
Of the 50 differentially expressed microRNAs in PHPT patients with osteoporosis, 12 microRNAs were randomly selected and verified by qRT-PCR. Among these microRNAs, miR-1299, miR-193b-3p, let-7i-5p, miR-10a-3p, miR-138-5p, miR-363-3p, miR-365a-3p, miR-4423-5p, miR-33a-5p, miR-3607-3p, and miR-874-3p were upregulated, whereas miR-512 was downregulated, in PHPT patients with osteoporosis. Consistent with our findings, studies have linked some of these microRNAs with osteogenesis. Jia et al have shown that miR-193b-3p and let-7i-5p were significantly upregulated in adipose-derived stem cells during osteogenic differentiation.36 Zhang et al have demonstrated that let-7i-5p promotes osteogenic differentiation by targeting casein kinase 2-interacting protein 1, playing an important role in fracture healing.37 These findings suggest that these microRNAs and downstream targets form a complex network in the regulation of osteogenesis.
In this study, we demonstrated that miR-874-3p was significantly upregulated in PHPT patients with osteoporosis compared with those in patients without osteoporosis. miR-874-3p promotes mouse osteoblast differentiation, as evidence by increased ALP activity, mineralization, and Runx2 promoter activity, via repressing histone deacetylase 1 expression.38 These findings collectively suggest that miR-874-3p is closely associated with bone metabolism. However, the roles of miR-874-3p in osteogenic differentiation of hMSCs remain unexplored. In this study, we demonstrated that miR-874-3p mimics suppressed the mRNA expression of ALP, OCN2, and Runx2, whereas miR-874-3p inhibitor showed contrasting results, suggesting that miR-874-3p might inhibit osteogenic differentiation of hMSCs, and thus contributes to the development of PHPT-induced osteoporosis. However, the direct role of miR-874-3p in osteogenesis needs further investigation. Each microRNA may suppress multiple mRNA targets.
Fat mass and obesity-associated protein (FTO) is a chromosomal N6-methyladenosine (m6A) demethylase involved in the metabolism of various types of RNA within cells. Studies have shown that FTO is significantly upregulated during the differentiation of BMSC into osteoblasts, and inhibition of FTO expression will hinder the osteogenic differentiation of BMSC. The preliminary biogenic analysis suggested that miR-874-3p might have a targeting relationship with FTO, and the applicant conducted cell experiments to explore the relationship between the two. The results suggest that miR-874-3p can suppress the expression of FTO-LUc in luciferase reporting system detection, indicating that miR-874-3p can specifically bind to FTO UTR sequence. Further experimental results showed that miR-874-3p inhibited FTO expression. These results indicated that miR-874-3p inhibited osteogenic differentiation by regulating FTO expression.
However, this study still has limitations. Firstly, the regulatory effect of the miR-874-3p has only been preliminarily verified through in vitro cell experiments so far, and its correlation has not yet been confirmed in animal models; Secondly, the key targets regulated by m6A modification downstream of FTO have not been fully clarified, and whether there are other potential pathways of action for miR-874-3p also remains to be explored.
In conclusion, our data suggested that miR-874-3p was dramatically increased in hMSCs grown in osteogenic medium. Overexpression of miR-874-3p suppressed osteogenic marker expression, whereas miR-874-3p inhibitor enhanced osteogenic marker expression in hMSCs. Therefore, targeting miR-874-3p might serve as a potential therapeutic strategy to prevent or treat osteoporosis for patients with PHPT.
Ethics Statement
The studies involving human participants were reviewed and approved by the Institutional Review Board of Beijing Jishuitan Hospital (Reg. No. 201905–01). Our study complies with the Declaration of Helsinki. The patients/participants provided their written informed consent to participate in this study.
Funding
This work was supported by grants from National Natural Science Foundation of China Youth Fund (82302659), Project supported by Beijing Jishuitan Research Funding (KYYC202301), Beijing Municipal Health Commission (BJRITO-RDP-2024), Beijing Municipal Public Welfare Development and Reform Pilot Project for Medical Research Institutes (JYY2023-11), Beijing Municipal Public Welfare Development and Reform Pilot Project for Medical Research Institutes (JYY2023-8).
Disclosure
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Bilezikian JP, Bandeira L, Khan A, Cusano, Cusano NE. Hyperparathyroidism. Lancet. 2018;391(10116):168–178. doi:10.1016/S0140-6736(17)31430-7
2. Yeh MW, Ituarte PH, Zhou HC, et al. Incidence and prevalence of primary hyperparathyroidism in a racially mixed population. J Clin Endocrinol Metab. 2013;98(3):1122–1129. doi:10.1210/jc.2012-4022
3. Wermers RA, Khosla S, Atkinson EJ, et al. Incidence of primary hyperparathyroidism in Rochester, Minnesota, 1993-2001: an update on the changing epidemiology of the disease. J Bone Miner Res. 2006;21(1):171–177.
4. Qiu T, Xian L, Crane J, et al. PTH receptor signaling in osteoblasts regulates endochondral vascularization in maintenance of postnatal growth plate. J Bone Miner Res. 2015;30(2):309–317.
5. Viccica G, Cetani F, Vignali E, Miccoli M, Marcocci C. Impact of vitamin D deficiency on the clinical and biochemical phenotype in women with sporadic primary hyperparathyroidism. Endocrine. 2017;55(1):256–265. doi:10.1007/s12020-016-0931-8
6. Walker MD, Cong E, Lee JA, et al. Vitamin D in primary hyperparathyroidism: effects on clinical, biochemical, and densitometric presentation. J Clin Endocrinol Metab. 2015;100(9):3443–3451. doi:10.1210/jc.2015-2022
7. Cipriani C, Biamonte F, Costa AG, et al. Prevalence of kidney stones and vertebral fractures in primary hyperparathyroidism using imaging technology. J Clin Endocrinol Metab. 2015;100(4):1309–1315. doi:10.1210/jc.2014-3708
8. Walker MD, Silverberg SJ. Primary hyperparathyroidism. Nat Rev Endocrinol. 2018;14(2):115–125. doi:10.1038/nrendo.2017.104
9. Sozen T, Ozisik L, Basaran NC. An overview and management of osteoporosis. Eur J Rheumatol. 2017;4(1):46–56. doi:10.5152/eurjrheum.2016.048
10. Morgan SL, Prater GL. Quality in dual-energy X-ray absorptiometry scans. Bone. 2017;104:13–28. doi:10.1016/j.bone.2017.01.033
11. Marshall D, Johnell O, Wedel H. Meta-analysis of how well measures of bone mineral density predict occurrence of osteoporotic fractures. BMJ. 1996;312(7041):1254–1259. doi:10.1136/bmj.312.7041.1254
12. Small RE. Uses and limitations of bone mineral density measurements in the management of osteoporosis. MedGenMed. 2005;7(2):3.
13. Cheng VK, Au PC, Tan KC, Cheung CL. MicroRNA and human bone health. JBMR Plus. 2019;3(1):2–13. doi:10.1002/jbm4.10115
14. Hackl M, Heilmeier U, Weilner S, Grillari J. Circulating microRNAs as novel biomarkers for bone diseases - Complex signatures for multifactorial diseases? Mol Cell Endocrinol. 2016;432:83–95. doi:10.1016/j.mce.2015.10.015
15. Seeliger C, Karpinski K, Haug AT, et al. Five freely circulating miRNAs and bone tissue miRNAs are associated with osteoporotic fractures. J Bone Miner Res. 2014;29(8):1718–1728. doi:10.1002/jbmr.2175
16. Kelch S, Balmayor ER, Seeliger C, Vester H, Kirschke JS, van Griensven M. miRNAs in bone tissue correlate to bone mineral density and circulating miRNAs are gender independent in osteoporotic patients. Sci Rep. 2017;7(1):15861. doi:10.1038/s41598-017-16113-x
17. Yavropoulou MP, Anastasilakis AD, Makras P, Tsalikakis DG, Grammatiki M, Yovos JG. Expression of microRNAs that regulate bone turnover in the serum of postmenopausal women with low bone mass and vertebral fractures. Eur J Endocrinol. 2017;176(2):169–176. doi:10.1530/EJE-16-0583
18. Belaya ZE, Grebennikova TA, Melnichenko GA, et al. Effects of endogenous hypercortisolism on bone mRNA and microRNA expression in humans. Osteoporos Int. 2018;29(1):211–221. doi:10.1007/s00198-017-4241-7
19. Ding Q, Zhou H, Yun B, et al. Interleukin-13 inhibits expression of cyp27b1 in peripheral cd14+ cells that is correlated with vertebral bone mineral density of patients with ulcerative colitis. J Cell Biochem. 2017;118(2):376–381. doi:10.1002/jcb.25646
20. Moayeri A, Mohamadpour M, Mousavi SF, Shirzadpour E, Mohamadpour S, Amraei M. Fracture risk in patients with type 2 diabetes mellitus and possible risk factors: a systematic review and meta-analysis. Ther Clin Risk Manag. 2017;13:455–468. doi:10.2147/TCRM.S131945
21. Chen A, Zhang Z VX, Sze Q, et al. Targeting the oncogenic m6A demethylase FTO suppresses tumourigenesis and potentiates immune response in hepatocellular carcinoma. Gut. 2025;74(1):90–102. doi:10.1136/gutjnl-2024-331903
22. Yang Y, Ren J, Zhang J, Shi H, Wang J, Yan Y. FTO ameliorates doxorubicin-induced cardiotoxicity by inhibiting ferroptosis via P53-P21/Nrf2 activation in a HuR-dependent m6A manner. Redox Biol. 2024;70:103067. doi:10.1016/j.redox.2024.103067
23. Pantaleao LC, Ozanne SE. Small RNA sequencing: a technique for miRNA Profiling. Methods Mol Biol. 2018;1735:321–330.
24. Gu H, Wu L, Chen H, et al. Identification of differentially expressed microRNAs in the bone marrow of osteoporosis patients. Am J Transl Res. 2019;11(5):2940–2954.
25. Mendoza-Moreno F, Rodriguez-Pascual A, Diez-Gago MR, et al. Primary hyperparathyroidism due to parathyroid adenoma originated from supernumerary gland. Case Rep Otolaryngol. 2018;2018:6482546. doi:10.1155/2018/6482546
26. Akkawi I, Zmerly H. Osteoporosis: current Concepts. Joints. 2018;6(2):122–127. doi:10.1055/s-0038-1660790
27. Kozomara A, Griffiths-Jones S. miRBase: annotating high confidence microRNAs using deep sequencing data. Nucleic Acids Res. 2014;42:D68–73. doi:10.1093/nar/gkt1181
28. Wen M, Shen Y, Shi S, Tang T. Tang T. miREvo: an integrative microRNA evolutionary analysis platform for next-generation sequencing experiments. BMC Bioinf. 2012;13:140. doi:10.1186/1471-2105-13-140
29. Friedlander MR, Mackowiak SD, Li N, Chen W, Rajewsky N. miRDeep2 accurately identifies known and hundreds of novel microRNA genes in seven animal clades. Nucleic Acids Res. 2012;40(1):37–52. doi:10.1093/nar/gkr688
30. Zhou L, Chen J, Li Z, et al. Integrated profiling of microRNAs and mRNAs: microRNAs located on Xq27.3 associate with clear cell renal cell carcinoma. PLoS One. 2010;5(12):e15224. doi:10.1371/journal.pone.0015224
31. Storey JD. The positive false discovery rate: a Bayesian interpretation and the q-value. Ann Stat. 2003;31:2013–2035. doi:10.1214/aos/1074290335
32. Young MD, Wakefield MJ, Smyth GK, Oshlack A. Gene ontology analysis for RNA-seq: accounting for selection bias. Genome Biol. 2010;11(2):R14. doi:10.1186/gb-2010-11-2-r14
33. Mao X, Cai T, Olyarchuk JG, Wei L. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics. 2005;21(19):3787–3793. doi:10.1093/bioinformatics/bti430
34. Ren G, Sun J, Li MM, Zhang YD, Li RH, Li YM. MicroRNA-23a-5p regulates osteogenic differentiation of human bone marrow-derived mesenchymal stem cells by targeting mitogen-activated protein kinase-13. Mol Med Rep. 2018;17(3):4554–4560. doi:10.3892/mmr.2018.8452
35. Kim H, Kim J, Kim K, Chang H, You K, Kim VN. Bias-minimized quantification of microRNA reveals widespread alternative processing and 3’ end modification. Nucleic Acids Res. 2019;47(5):2630–2640. doi:10.1093/nar/gky1293
36. Jia B, Zhang Z, Qiu X, et al. Analysis of the miRNA and mRNA involved in osteogenesis of adipose-derived mesenchymal stem cells. Exp Ther Med. 2018;16(2):1111–1120. doi:10.3892/etm.2018.6303
37. Zhang Y, Cheng W, Han B, et al. Let-7i-5p functions as a putative osteogenic differentiation promoter by targeting CKIP-1. Cytotechnology. 2021;73(1):79–90. doi:10.1007/s10616-020-00444-1
38. Kushwaha P, Khedgikar V, Sharma D, et al. MicroRNA 874-3p exerts skeletal anabolic effects epigenetically during weaning by suppressing hdac1 expression. J Biol Chem. 2016;291(8):3959–3966. doi:10.1074/jbc.M115.687152
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Curculigoside is a Promising Osteoprotective Agent for Osteoporosis: Review
Du J, Qin W, Wen F, Liu Y, Zhang Q, Liu W, Huang C, Feng Z, Pan Z, Gu E
Drug Design, Development and Therapy 2025, 19:3323-3336
Published Date: 28 April 2025