Back to Journals » Cancer Management and Research » Volume 12
MicroRNA-584 Impairs Cellular Proliferation and Sensitizes Osteosarcoma Cells to Cisplatin and Taxanes by Targeting CCN2
Authors Li L, Kong X, Zang M, Hu B, Fang X, Gui B, Hu Y
Received 18 January 2020
Accepted for publication 31 March 2020
Published 15 April 2020 Volume 2020:12 Pages 2577—2587
DOI https://doi.org/10.2147/CMAR.S246545
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Kenan Onel
Li Li,1,2,* Xiang’an Kong,2,* Mousheng Zang,2 Bin Hu,2 Xing Fang,2 Binjie Gui,1 Yong Hu1
1Department of Orthopedics, The First Affiliated Hospital of Anhui Medical University, Hefei, Anhui, People’s Republic of China; 2Department of Orthopedics, The Second People’s Hospital of Hefei, The Affiliated Hefei Hospital of Anhui Medical University, Hefei, Anhui, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yong Hu Tel +86-551-62923605
Email [email protected]
Background: Osteosarcoma (OS), an aggressive malignant neoplasm, exhibits osteoblastic differentiation. Cisplatin (DDP) and taxanes are among the most effective drugs for OS patients. Nevertheless, the drug resistance remains a main limitation to efficacious chemotherapy in OS. The current report sets to explore the biological function of microRNA-584 (miR-584) and the potential mechanism underlying OS cells resistance to these two drugs.
Materials and Methods: The expression profiles of miR-584 and connective tissue growth factor (CTGF, CCN2) in OS tissue samples and cell lines were tested by means of reverse transcription-quantitative polymerase chain reaction and Western blot. U2OS and MG63 cell lines were delivered with miR-584 mimic alone or plus CCN2 to excavate theirs functions by cell counting kit-8 and EdU, flow cytometric analysis, as well as transwell assay, severally. Western bot analysis was conducted to examine the expression of IκBα, pIκBα, NF-κB and pNF-κB. Dual-luciferase reporter gene assay was carried out to assess the targets of miR-584.
Results: The downregulation of miR-584 was identified in OS tissues and cells, which was closely linked to the dismal prognosis of OS patients. Overexpression of miR-584 repressed cell viability, migration as well as invasion, potentiated apoptosis and sensitized OS cells to DDP and taxanes. Mechanism investigation specified a direct targeting relationship between CCN2 and miR-584 in OS.
Conclusion: In conclusion, miR-584 has the potency to act as a therapeutic maneuver for OS mainly by inducing the chemosensitivity of OS cells to DDP and taxanes.
Keywords: osteosarcoma, microRNA-584, CCN2, the IκBα/NF-κB signaling pathway, chemoresistance
Introduction
Osteosarcoma (OS), the most relevant malignancy of the bone, is believed to initiate from mesenchymal stem cells, and prior to the introduction of chemotherapy, the outcome of OS patients was unsatisfactory, with the survival rate less than 20% before the 1970s.1 The advance in effective chemotherapies for OS in the 1980s culminated in a radical improvement in survival.2 At this time, the drugs commonly used in the neoadjuvant treatment of OS include doxorubicin, cisplatin (DDP), methotrexate and etoposide.3 Among numerous antitumor agents, taxanes is widely applied for treating breast and pancreatic cancers, and the efficacy of taxanes for treating OS was previously revealed to be a great success in high-grade OS and recurrent or metastatic soft-tissue sarcomas.4 However, increasing chemoresistance was saw in a great number of OS patients in the past few years, therefore the study of the underlying mechanisms regarding the chemosensitivity of OS cells is of great importance to enhance the survival rate.5
The dysregulation of microRNAs (miRNAs), small noncoding RNAs, have been observed in many human pathologies, and most prominently, in carcinogenesis.6 miR-584 was predicted to be dysregulated in primary OS samples by online MiRWalk and Kyoto Encyclopedia of Genes and Genomes pathway databases.7 Stratification indicated an association between lower expression of miR-584-5p with the disease progression in Ewing sarcoma.8 miR-584-5p has been reported to potentiated the vincristine response in medulloblastoma.9 Moreover, connective tissue growth factor (CTGF, CCN2) is a member of the CCN family, which is consisted of six proteins correlated to extracellular matrix which function critically in wound healing, fibrosis as well as cancers.10 In OS, CCN2 has been revealed to act as a target to cope with metastasis and angiogenesis.11 In breast cancer cells, short hairpin RNA-regulated knockdown of CCN2 reversed Taxol resistance evoked by transcriptional co-activator with PDZ-binding motif.12 Moreover, the activation of nuclear factor-kappa B (NF-κB) was participated in CCN2-modulated metastasis in OS.13 Besides, classical NF-κB could be repressed by upregulation of the IκBα super suppressor in OS cells.14 Bioinformatics analysis provided forceful evidences that there were potential binding sites between miR-584 and CCN2. Hence, we assumed miR-584 could regulate OS chemoresistance via CCN2. Hence, we examined miR-584 expression profiles in OS tissues and cells in this work and studied the therapeutic relevance of miR-584 on OS and the regulatory axis of miR-584/CCN2/IκBα/NF-κB.
Materials and Methods
Ethical Statements
The study was permitted by the Institutional Review Board of the First Affiliated Hospital of Anhui Medical University and followed the Declaration of Helsinki. Every participant provided a written informed consent.
Tissue Samples
Between April 2013 and September 2013, 37 OS patients diagnosed as primary OS by pathology and underwent surgery in First Affiliated Hospital of Anhui Medical University were enrolled. Prior to sampling, no patients received any anti-tumor treatments. A total of 37 pairs of OS tissues and adjacent paracancerous tissues were collected during surgery and immediately frozen in liquid nitrogen for further analysis. Demographic and clinicopathological characteristics of patients were recorded and summarized in Table 1. Besides, the patients were followed up every three months for 5 years after treatment, and we tracked their survival information by telephone.
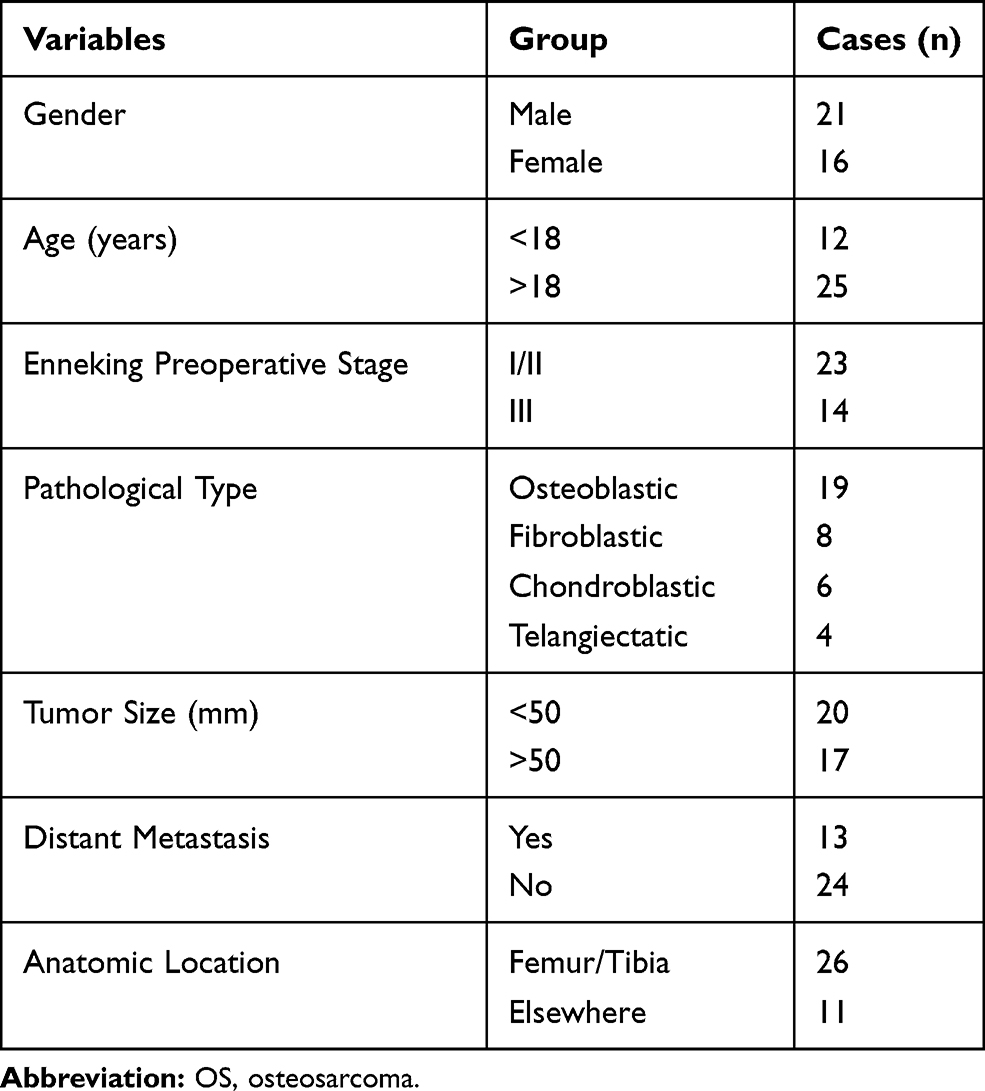 |
Table 1 The Clinicopathological Features of Patients with OS |
Oligonucleotide Microarray
OS samples and matched paracancerous samples from three OS sufferers were obtained to analyze differentially expressed miRNAs, which was performed as previously reported.15
Reverse Transcription-Quantitative Polymerase Chain Reaction (RT-qPCR)
A Trizol kit (TAKARA, Japan) was employed to extract total RNA in the tissues and cells in strict accordance with the instructions. Subsequently, a reverse transcription kit (TAKARA, Japan) was used for reverse transcription. Following, qPCR was conducted by the SYBR Green PCR Master Mix (Invitrogen Inc., Carlsbad, CA, USA). U6 was applied as the internal reference for miR-584, while GAPDH for CCN2. The sequence of primers used in the experiment is shown in Table 2.
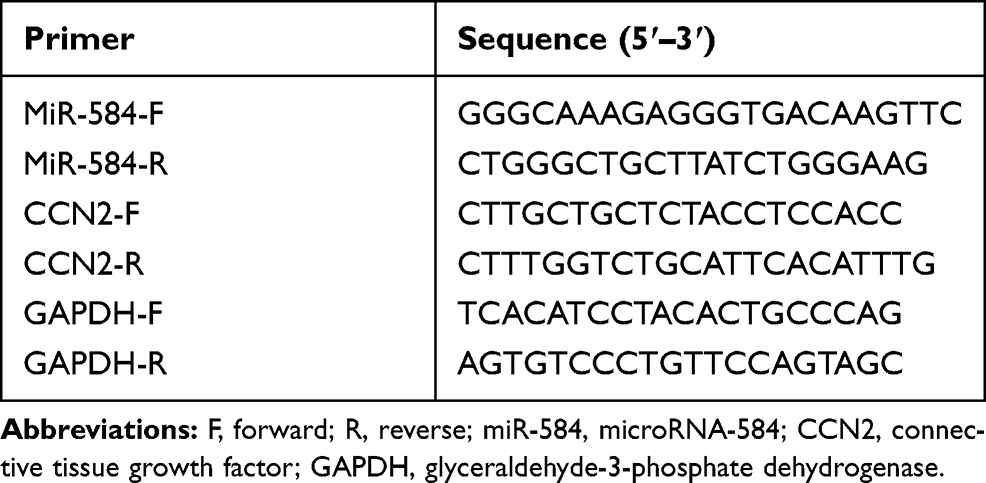 |
Table 2 Prime Sequences for Reverse Transcription-Quantitative Polymerase Chain Reaction |
Cell Culture and Treatment
Human OS cell lines (U2OS, SOSP-M, SaOS2, MG63) and human fetal osteoblastic cells (hFOB) were from American Type Culture Collection (Rock Ville, MD, USA). All cell lines were cultivated using Dulbecco’s modified Eagle’s medium containing 10% fetal bovine serum (Gibco, Greenland, N.Y., USA). MiR-584 mimic and corresponding Mock, pcDNA-CCN2 and corresponding empty were produced by Genepharma Bioengineering Co., Ltd. (Shanghai, China). Lipofectamine 2000 (Invitrogen) was employed for the transfection following the manufacturer’s protocols.
Detection of Cell Activity
CCK-8 (Shanghai Sangon Biological Engineering Technology & Services Co., Ltd., Shanghai, China) was utilized strictly in accordance with the instructions. The cell proliferation was evaluated by 5-Ethynyl-2ʹ-deoxyuridine (EdU) assay, and all experimental steps were carried out according to a previous report.16 The levels of apoptosis were measured by propidium iodide (PI)/Annexin-V-labeled-flow cytometry and Hoechst 33258 staining as previously reported.17
Transwell assays were carried out to determine the migration and invasion abilities of U2OS and MG63 cells.16
Detection of Resistance of U2OS and MG63 Cells
U2OS and MG63 were exposed to different concentrations of DDP and taxanes, respectively. After that, the cell survival rate was tested by a CCK-8 cytotoxicity assay following the instructions provided by the manufacturer. Then, the cells treated with 2.5 μM DPP or Taxanes were detected by PI/Annexin-V-labeled flow cytometry and Hoechst 33258 staining following the steps described above.
Dual-Luciferase Reporter Assay
The wild-type (WT) and a complementary mutation (MT) of 3ʹUTR of CCN2 mRNA were created onto a pGL3 luciferase reporter vector (Promega Corporation). Subsequently, the constructed luciferase reporter vector was delivered into H293T cells together with miR-584 mimic or mimic control via lipofectamine 2000 (Invitrogen). Finally, a dual-luciferase report kit (Promega) was applied for the evaluation of the luciferase activity.
Western Blot Assays
The expression of CCN2 and IκBα/NF-κB signaling pathway-related proteins in U2OS and MG63 cells was detected by Western blot as described previously.18 The following primary antibodies were used: CCN2 (1:3000, ab5097), IκBα (1:5000, ab32518), pIκBαS32 (1:5000, ab92700), NF-κB (1:5000, ab32536), pNF-κBS536 (1:2000, ab86299). The blots were then probed with a goat anti-rabbit horseradish peroxidase-linked antibody to IgG (1:50,000, ab6070). All antibodies aforementioned were from Abcam (Cambridge, UK).
Statistics
All statistical data were analyzed using the SPSS 21.0 (IBM-SPSS, Inc, Chicago, IL, USA). Measurement data were depicted as mean ± standard deviation. All data were subjected to Kolmogorov–Smirnov to test normal distribution. One-way or two-way analysis of variance (ANOVA) followed by Tukey’s post hoc test was applied for data analysis. Pearson analysis was used for correlation analysis. If the p value was less than 0.05, the differences were deemed as significant.
Results
Downregulation of miR-584 in OS Is Linked to Poor Prognosis
Initially, we analyzed the miRNA expression in tumor tissues from three OS patients by microarray. After homogenization of the data, the expression heatmap of differentially expressed miRNAs was plotted, as shown in Figure 1A. RT-qPCR analysis was subsequently conducted so as to determine the expression of miR-584 in 37 OS and corresponding paracancerous tissues. MiR-584 was remarkably reduced in OS tissues relative to paracancerous tissues (Figure 1B). We then divided OS patients into high-expression and low-expression groups based on the median expression of miR-584 in OS tissue (1.76). We analyzed the correlation between miR-584 and age, gender, clinical stage, pathological type, tumor size, metastasis and survival of OS patients. We found that the expression of miR-584 was related to clinical stage, metastasis and survival rate of OS patients (Table 3). In addition, we showed that OS patients harboring reduced miR-584 expression demonstrated poorer prognosis than those with relatively high expression of miR-584 (Figure 1C).
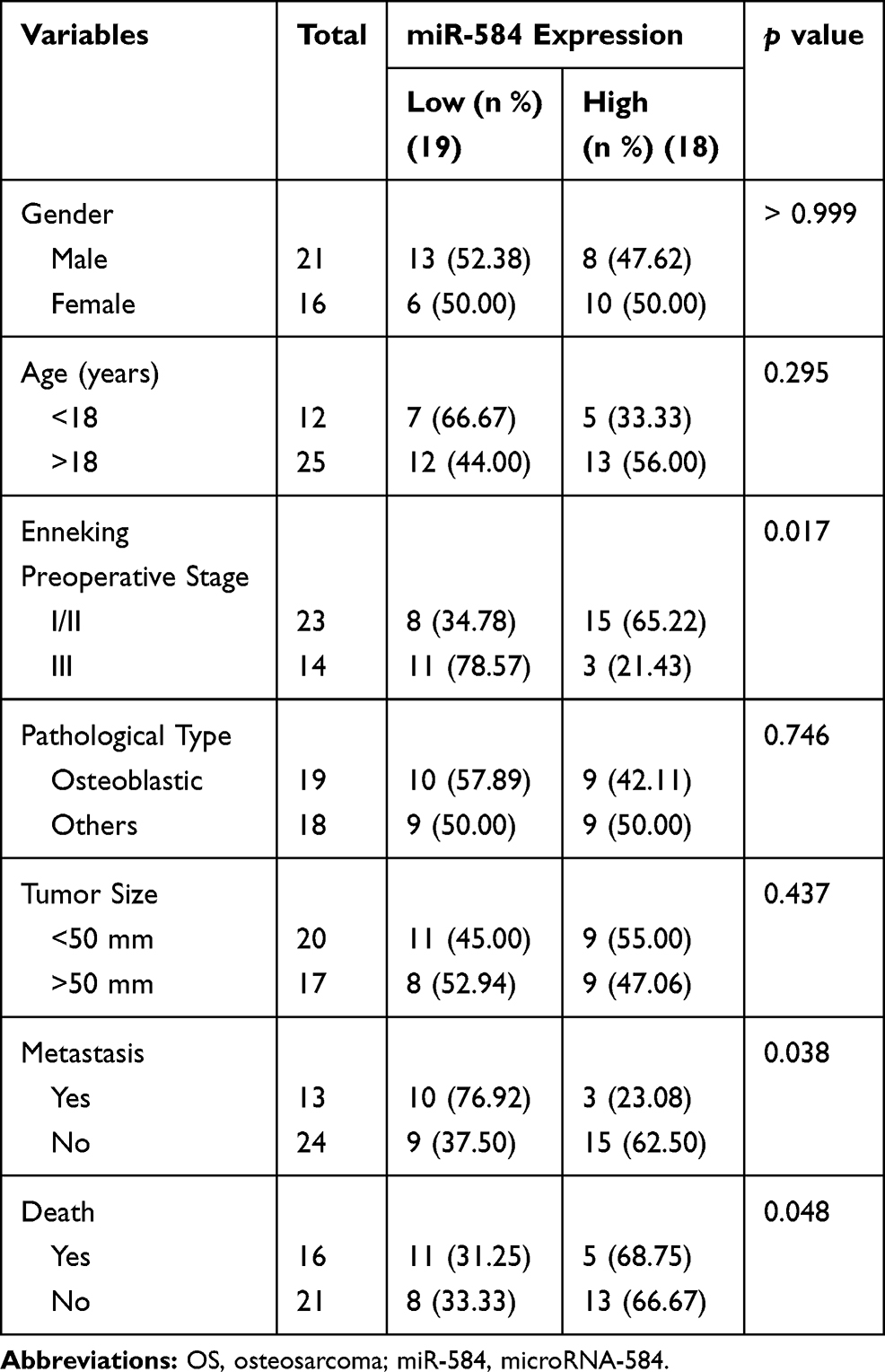 |
Table 3 Association Between miR-584 Expression and Clinicopathological Characteristics of OS Patients |
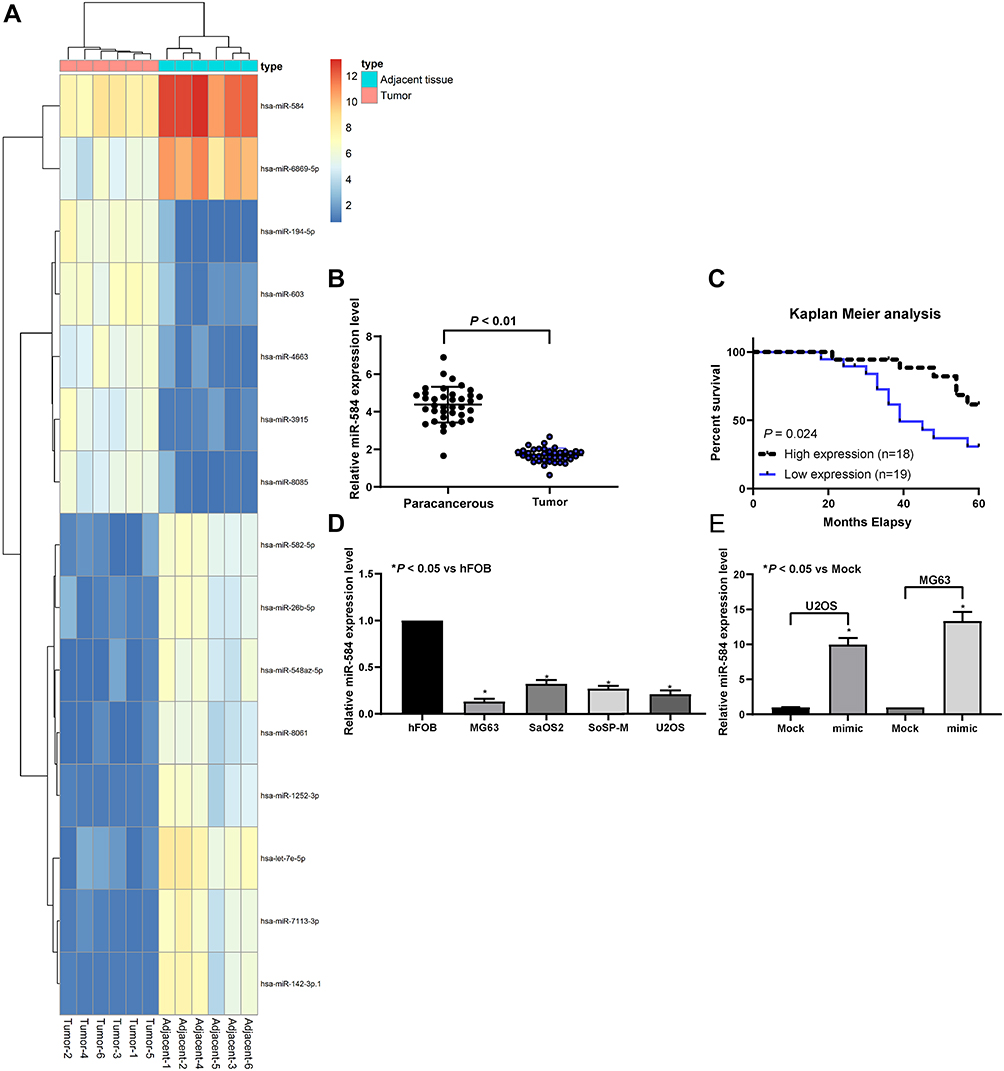 |
Figure 1 miR-584 is downregulated in OS tissues and cell lines. (A) The heatmap for 30 ectopic expressed miRNAs in OS. (B) miR-584 expression in 37 OS and paired paracancerous tissues determined by RT-qPCR. (C) The overall survival of OS patients with low and high miR-584 expression. (D) The expression of miR-584 in OS cell lines (U2OS, SoSP-M, SaOS-2, MG-63) and hFOB cells measured by RT-qPCR. (E) The expression of miR-584 in U2OS and MG63 cells transfected with miR-584 mimic or Mock plasmids determined by RT-qPCR. One-way ANOVA and Tukey’s multiple comparison test was used to determine statistical significance, *p < 0.05 vs. paracancerous tissues, hFOB cells or the Mock group. |
Subsequently, miR-584 was remarkably diminished in OS cells contrast to hFOB cells (Figure 1D). Then, RT-qPCR analysis confirmed that miR-584 mimic enhanced miR-584 expression in both U2OS and MG-63 cell lines (Figure 1E). All in all, downregulation of miR-584 may have a significant relevance in OS.
MiR-584 Hinders OS Cell Viability, Migration and Invasion, Yet Promoting Apoptosis
The subsequent CCK-8 and EdU staining displayed that miR-584 mimic lowered the U2OS and MG63 cell viability and proliferation (Figure 2A and B). Subsequently, through PI/Annexin V-flow cytometry and Hoechst 33258 staining, we found that miR-584 mimic promoted the apoptosis of both cell lines (Figure 2C and D). Also, miR-584 reduced the migration and invasion of U2OS as well as MG63 cell lines (Figure 2E and F).
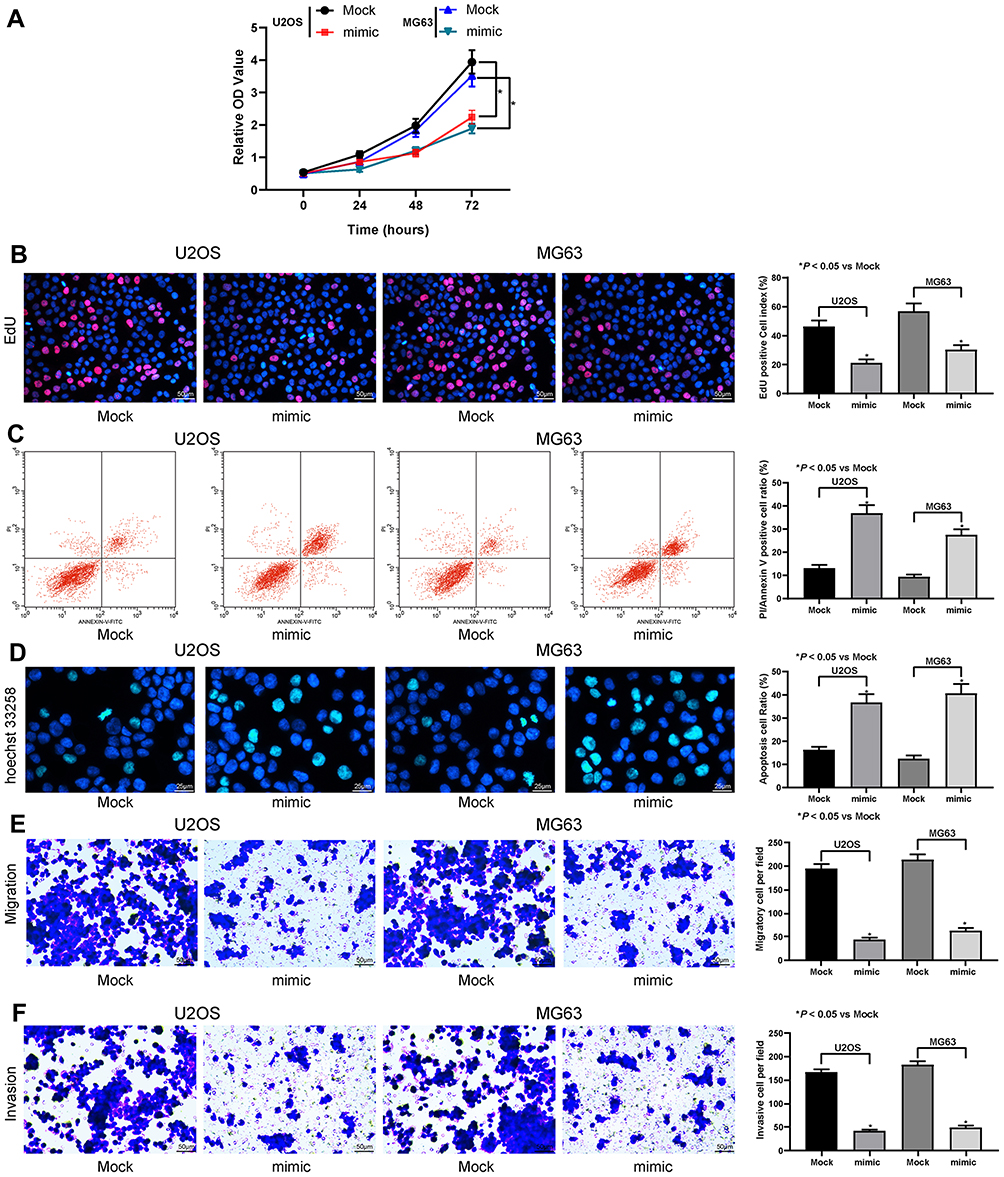 |
Figure 2 miR-584 inhibits OS cell viability. (A) The optical density (OD) value of U2OS and MG63 cells on the 0th, 24th, 48th and 72nd h after transfection determined by CCK-8 assays. (B) The EdU staining of U2OS and MG63 cells. (C) U2OS and MG63 cell apoptosis determined by PI/Annexin V flow cytometry. (D) Hoechst 33258 staining of U2OS and MG63 cells. (E) U2OS and MG63 cell migration assessed by transwell assay. (F) U2OS and MG63 cell invasion assessed by transwell assay. One-way ANOVA and Tukey’s multiple comparison test was used to determine statistical significance, * p < 0.05 vs. the Mock group. |
MiR-584 Enhances Drug Sensitivity of OS Cells to DDP and Taxanes
Subsequently, our CCK-8 cytotoxicity assay showed that miR-584 mimic increased the drug sensitivity of U2OS and MG63 cells to DDP and taxanes (Figure 3A). Then, we found that after miR-584 mimic treatment, U2OS and MG63 cells under exposure to 2.5 μM DDP and 2.5 μM Taxanes demonstrated induced apoptosis (Figure 3B and C). Therefore, miR-584 downregulation may link to a chemoresistant phenotype of OS cells.
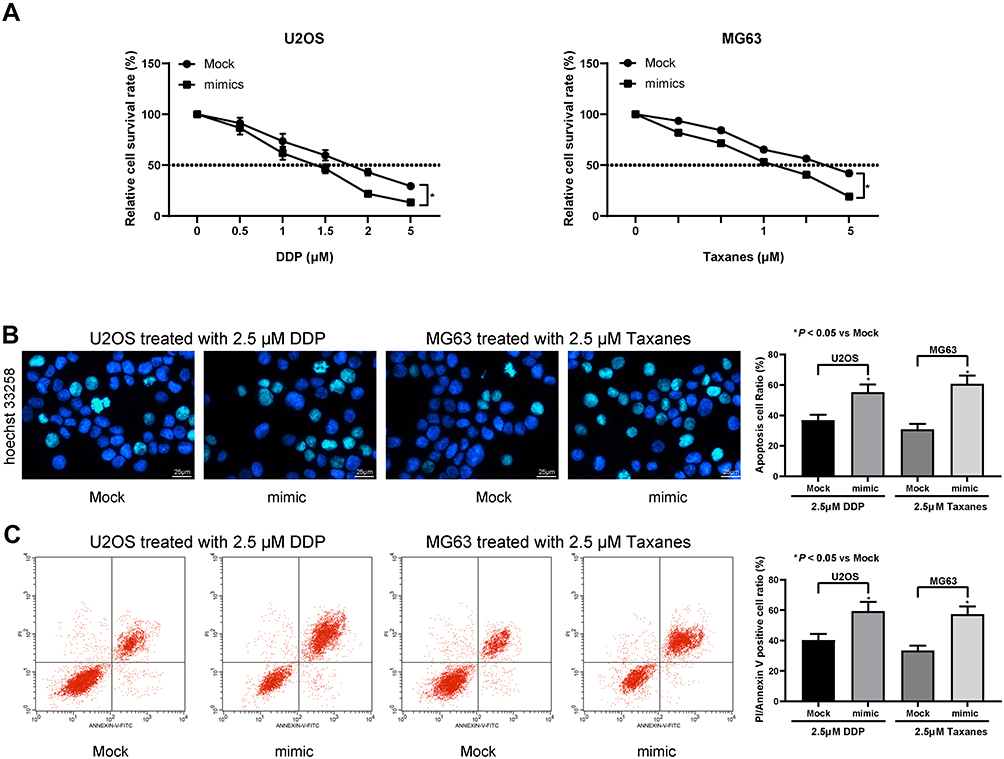 |
Figure 3 miR-584 promotes OS cell drug sensitivity. (A) U2OS and MG63 cell survival rate under different concentrations of DPP and Taxanes. (B) Hoechst 33258 staining of U2OS and MG63 cells. (C) U2OS and MG63 cell apoptosis evaluated by PI/Annexin V-flow cytometry. One-way ANOVA and Tukey’s multiple comparison test was used to determine statistical significance, *p < 0.05 vs. the Mock group. |
CCN2 May Interact with miR-584 in OS Cells
In order to probe the mechanisms how miR-584 reverted the malignant phenotype of OS cells, we firstly carried out bioinformatics analysis to predict the putative target genes of miR-584. Among which, CCN2, an oncogene reported in OS, has been screened out.13 Therefore, we employed a dual-luciferase reporter gene assay to evaluate whether CCN2 could be targeted by miR-584 in OS cells (Figure 4A). Then we examined the mRNA expression of CCN2 in 37 OS patients by RT-qPCR. CCN2 was found to be promoted in OS tissues versus paracancerous tissues and negatively correlated with the expression of miR-584 (Figure 4B and C). Furthermore, we potentiated miR-584 expression in U2OS and MG63 cells to assess whether CCN2 expression altered as a consequence. Delivery of miR-584 remarkably diminished CCN2 expression at both mRNA and protein levels in U2OS and MG63 cells (Figure 4D and E).
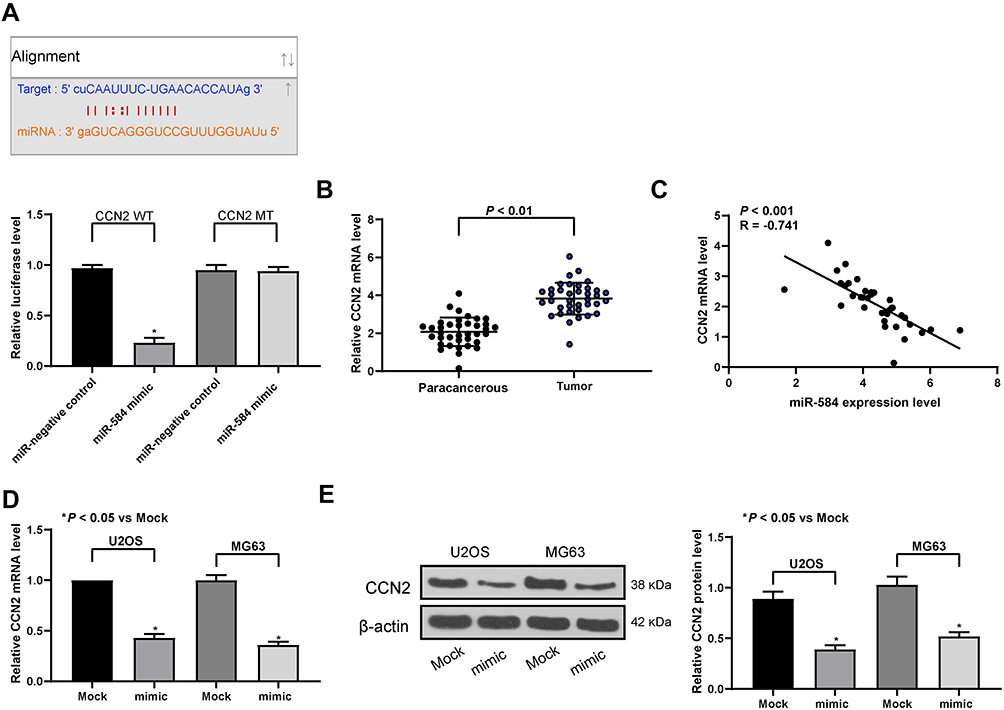 |
Figure 4 miR-584 targets and negatively regulates CCN2 in OS cells. (A) The binding relationship between miR-584 and CCN2 mRNA predicted by StarBase and tested by dual-luciferase assays. (B) CCN2 mRNA expression in 37 OS and paired paracancerous tissues determined by RT-qPCR. (C) The negative correlation between miR-584 expression and CCN2 mRNA expression evaluated by Pearson’s correlation analysis. (D) The mRNA expression of CCN2 in U2OS and MG63 cells determined by RT-qPCR. (E) The protein expression of CCN2 in U2OS and MG63 cells determined by and Western blot assays. One-way ANOVA and Tukey’s multiple comparison test or paired t-test was used to determine statistical significance, *p < 0.05 vs. the Mock group or paracancerous tissues. |
CCN2 Attenuates the Action of miR-584 on OS Cells
With an aim to further elucidate whether the decline in CCN2 expression in response to miR-584 was accountable for the inhibition of the malignant aggressiveness of OS cells, we promoted CCN2 expression in U2OS and MG63 cells overexpressing miR-584. Functional assays discovered that the effects of miR-584 overexpression on U2OS and MG63 cell viability and apoptosis were partially counteracted by CCN2 restoration (Figure 5A and B). Additionally, the abilities of cell invasion and migration were markedly enhanced by CCN2 in U2OS and MG63 cell overexpressing miR-584 (Figure 5C and D). Consequently, miR-584 exerted tumor-suppressive properties in OS cells by reducing CCN2.
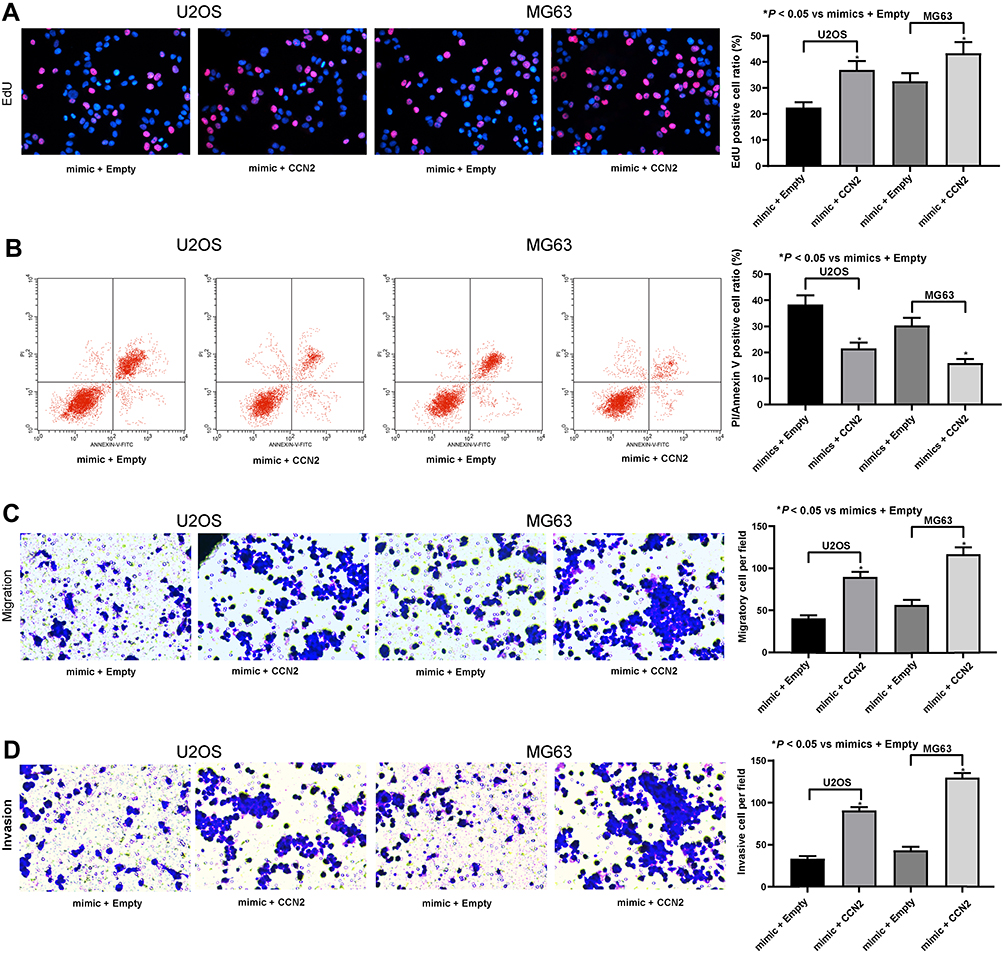 |
Figure 5 CCN2 overexpression attenuates the effects of miR-584 mimic on OS cells. (A) EdU staining of U2OS and MG63 cells. (B) U2OS and MG63 cell apoptosis determined by PI/Annexin V-flow cytometry. (C) U2OS and MG63 cell migration evaluated by transwell assay. (D) U2OS and MG63 cell invasion evaluated by transwell assay. One-way ANOVA and Tukey’s multiple comparison test was used to determine statistical significance, *p < 0.05 vs. the Mock group. |
miR-584 Impairs the IκBα/NF-κB Pathway by Targeting CCN2 in vitro in OS
CCN2 has been underscored to promote OS migration and the expression of MMP-2 and MMP-9 via the IκBα/NF-κB pathway.19 Hence, protein levels of IκBα, pIκBα, NF-κB, pNF-κB in U2OS and MG63 cells after co-transfection with miR-584 mimic plus pcDNA-CCN2 or the empty plasmid vector were examined by Western blot assays. miR-584 remarkably decreased pIκBα and pNF-κB expression in U2OS and MG63 cells, whereas total IκBα and NF-κB was barely affected. Of note, CCN2 overexpression partially counteracted the downregulation in pIκBα and pNF-κB expression induced by miR-584 (Figure 6). All these data indicate that miR-584 leads to the IκBα/NF-κB signaling deficits in OS cells by downregulating CCN2.
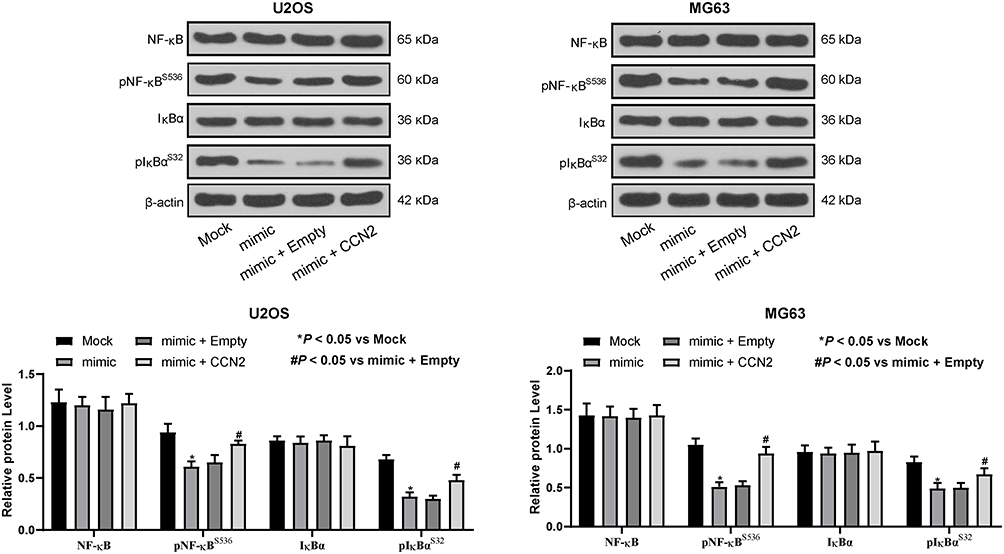 |
Figure 6 miR-584 prohibits IκBα/NF-κB signaling pathway by targeting CCN2. The extent of IκBα and NF-κB phosphorylation is inhibited by miR-584 mimic and then restored by CCN2 overexpression. Two-way ANOVA and Tukey’s multiple comparison test was used to determine statistical significance, *p < 0.05 vs. the Mock group; #p < 0.05 vs. the mimic + Empty group. |
Discussion
The conventional therapeutic regimes for OS include surgery and chemotherapy whose efficacy is ineffective due to drug resistance and the intricate microenvironment of cancer tissues.20 Herein, miR-584 was markedly diminished in OS tissues as well as cells relative to the normal controls. Additionally, miR-584 has the potency to suppress cell proliferation, migration and invasion and to enhance OS cell sensitivity to DDP and taxanes as well as apoptosis. In a word, miR-584 may function as an inhibitor in OS.
Reduced miR-584 has been monitored in quite a few cancers, involving glioma,21 colorectal cancer22 and gastric cancer.23 Moreover, we illustrated that the reduced miR-584 in OS patients was linked to poor prognosis. Poor expression of miR-584 tightly correlated to tumor size, tumor node metastasis as well as distant metastasis in non-small cell lung cancer.24 Furthermore, hampered proliferation, migration and invasion coupled with induced OS cell sensitivity were monitored after overexpression of miR-584. Likewise, the knockdown of miR-584-3p facilitated the migratory and invasive abilities of glioma cells.25 Meanwhile, Ueno et al reported that miR-584 remarkably reduced the cell motility of clear cell renal cell carcinoma.26 This study displayed that overexpression of miR-584 sensitized OS cells to DDP and taxanes.
In unearthing the mechanisms underlying the relevance of miR-584 on the promotion of drug sensitivity in OS cells, we found out that miR-584 targeted CCN2 3ʹUTR directly and conversely regulated its expression in OS cells. Our rescue experiments provided that upregulation of CCN2 might reverse the anti-proliferative, anti-migratory and anti-invasive properties of OS cells induced by miR-584 resumption. Similar to our data, CCN2 was revealed as a target of miR-26b in OS cells, and si-CCN2 hampered the expression of Smad1, ERK1/2, and MMP2, thus blocking the migration process in OS.27 Silencing of CCN2 expression through CCN2-shRNA profoundly diminished the cell migration and metastasis of OS cell both in vitro and in vivo.28 Besides, CCN2 was also upregulated by mechanical stresses; and CCN2 accumulated in mature osteocytes resulting from diminished cell projections, contributing to osteocyte death. Previously, CCN2 upregulation in breast cancer cells elicited an enhancement in resistance to doxorubicin and paclitaxel as revealed by immunohistochemical staining.29 Moreover, xenograft experiments conducted by Tsai et al suggested that CCN2 upregulation protected OS cells from doxorubicin, and the therapeutic efficacy was remarkably promoted when CCN2 was reduced.30 However, the relevance of CCN2 in chemoresistance of OS cells to DDP and taxanes was not clarified yet, which might be the direction for our further experiments.
Wu et al showed that WISP-1, a protein belonging to the CCN family, potentiated the OS cell migration by inducing the NF-κB transduction pathway.19 We now propose that miR-584 targets CCN2 and conversely modulates its expression and the downstream IκBα/NF-κB pathway in OS cells. NF-κB, a vital transcription factor in cancers, is located in the cytoplasm of inactive cells and isolates from IκB and translocates to the nucleus to modulate the transcription of genes at downstream by interacting with κB-binding sites once in response to stimuli.31 Also, increased expression of CCN2 correlated to the NF-κB pathway activation in endometrium of intrauterine adhesions.32 In addition, miR-29a prevented the NF-κB signaling pathway from activation in OS cells, and repressive effects of miR-29a on cell migration, invasion along with epithelial-mesenchymal transition were counteracted by knockdown of suppressor of cytokine signaling 1.33
Conclusion
Altogether, this study underscored that overexpressed miR-584 or silencing CCN2 slows down proliferation, migration and invasion, and potentiates chemosensitivity and apoptosis of U2OS and MG63 cells by disrupting the IκBα/NF-κB pathway. This study provides a novel way to further probe the pathogenesis of OS. Still, more efforts need to be made to authenticate the hypothesis and in vivo experiments should be carried out to substantiate our results.
Acknowledgments
This study was supported by the Natural Science Foundation Project of Anhui province (1708085MH215) and the School Fund of Anhui Medical University (2015xkj088).
Author Contributions
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; gave final approval of the version to be published; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Miwa S, Shirai T, Yamamoto N, et al. Current and emerging targets in immunotherapy for osteosarcoma. J Oncol. 2019;2019:7035045. doi:10.1155/2019/7035045
2. Anderson ME. Update on survival in osteosarcoma. Orthop Clin North Am. 2016;47(1):283–292. doi:10.1016/j.ocl.2015.08.022
3. Ferrari S, Serra M. An update on chemotherapy for osteosarcoma. Expert Opin Pharmacother. 2015;16(18):2727–2736. doi:10.1517/14656566.2015.1102226
4. Martella E, Ferroni C, Guerrini A, et al. Functionalized keratin as nanotechnology-based drug delivery system for the pharmacological treatment of osteosarcoma. Int J Mol Sci. 2018;19(11):3670. doi:10.3390/ijms19113670
5. Tsai HC, Huang CY, Su HL, Tang CH. CCN2 enhances resistance to cisplatin-mediating cell apoptosis in human osteosarcoma. PLoS One. 2014;9(3):e90159. doi:10.1371/journal.pone.0090159
6. Kushlinskii NE, Fridman MV, Braga EA. Molecular mechanisms and microRNAs in osteosarcoma pathogenesis. Biochemistry (Mosc). 2016;81(4):315–328. doi:10.1134/S0006297916040027
7. Delsin LEA, Roberto GM, Fedatto PF, et al. Downregulated adhesion-associated microRNAs as prognostic predictors in childhood osteosarcoma. Pathol Oncol Res. 2019;25(1):11–20. doi:10.1007/s12253-017-0316-1
8. Roberto GM, Delsin LEA, Vieira GM, et al. ROCK1-predictedmicroRNAs dysregulation contributes to tumor progression in ewing sarcoma. Pathol Oncol Res. 2017:1–7.
9. Abdelfattah N, Rajamanickam S, Panneerdoss S, et al. MiR-584-5p potentiates vincristine and radiation response by inducing spindle defects and DNA damage in medulloblastoma. Nat Commun. 2018;9(1):4541. doi:10.1038/s41467-018-06808-8
10. Chen PC, Cheng HC, Yang SF, Lin CW, Tang CH. The CCN family proteins: modulators of bone development and novel targets in bone-associated tumors. Biomed Res Int. 2014;2014:437096.
11. Wang LH, Tsai HC, Cheng YC, et al. CTGF promotes osteosarcoma angiogenesis by regulating miR-543/angiopoietin 2 signaling. Cancer Lett. 2017;391:28–37. doi:10.1016/j.canlet.2017.01.013
12. Lai D, Ho KC, Hao Y, Yang X. Taxol resistance in breast cancer cells is mediated by the hippo pathway component TAZ and its downstream transcriptional targets Cyr61 and CTGF. Cancer Res. 2011;71(7):2728–2738. doi:10.1158/0008-5472.CAN-10-2711
13. Hou CH, Yang RS, Tsao YT. Connective tissue growth factor stimulates osteosarcoma cell migration and induces osteosarcoma metastasis by upregulating VCAM-1 expression. Biochem Pharmacol. 2018;155:71–81. doi:10.1016/j.bcp.2018.06.015
14. Londhe P, Yu PY, Ijiri Y, et al. Classical NF-kappaB metabolically reprograms sarcoma cells through regulation of hexokinase 2. Front Oncol. 2018;8:104. doi:10.3389/fonc.2018.00104
15. Sastre D, Baiochi J, de Souza Lima IM, et al. Focused screening reveals functional effects of microRNAs differentially expressed in colorectal cancer. BMC Cancer. 2019;19(1):1239. doi:10.1186/s12885-019-6468-5
16. Liu B, Zhao H, Zhang L, Shi X. Silencing of long-non-coding RNA ANCR suppresses the migration and invasion of osteosarcoma cells by activating the p38MAPK signalling pathway. BMC Cancer. 2019;19(1):1112. doi:10.1186/s12885-019-6335-4
17. Huang L, Huang Z, Lin W, et al. Salidroside suppresses the growth and invasion of human osteosarcoma cell lines MG63 and U2OS in vitro by inhibiting the JAK2/STAT3 signaling pathway. Int J Oncol. 2019;54(6):1969–1980. doi:10.3892/ijo.2019.4781
18. Xu C, Zhang Y, Wang Q, et al. Long non-coding RNA GAS5 controls human embryonic stem cell self-renewal by maintaining NODAL signalling. Nat Commun. 2016;7(1):13287. doi:10.1038/ncomms13287
19. Wu CL, Tsai HC, Chen ZW, et al. Ras activation mediates WISP-1-induced increases in cell motility and matrix metalloproteinase expression in human osteosarcoma. Cell Signal. 2013;25(12):2812–2822. doi:10.1016/j.cellsig.2013.09.005
20. Duan R, Li C, Wang F, Yangi JC. Polymer-lipid hybrid nanoparticles-based paclitaxel and etoposide combinations for the synergistic anticancer efficacy in osteosarcoma. Colloids Surf B Biointerfaces. 2017;159:880–887. doi:10.1016/j.colsurfb.2017.08.042
21. Xu S, Zhang J, Xue H, et al. MicroRNA-584-3p reduces the vasculogenic mimicry of human glioma cells by regulating hypoxia-induced ROCK1 dependent stress fiber formation. Neoplasma. 2017;64(1):13–21. doi:10.4149/neo_2017_102
22. Huang H, Cai L, Li R, Ye L, Chen Z. A novel lncRNA LOC101927746 accelerates progression of colorectal cancer via inhibiting miR-584-3p and activating SSRP1. Biochem Biophys Res Commun. 2019;509(3):734–738. doi:10.1016/j.bbrc.2018.12.174
23. Zheng L, Chen Y, Ye L, et al. miRNA-584-3p inhibits gastric cancer progression by repressing Yin Yang 1- facilitated MMP-14 expression. Sci Rep. 2017;7(1):8967. doi:10.1038/s41598-017-09271-5
24. Zhang Y, Wang Y, Wang J. MicroRNA-584 inhibits cell proliferation and invasion in non-small cell lung cancer by directly targeting MTDH. Exp Ther Med. 2018;15(2):2203–2211. doi:10.3892/etm.2017.5624
25. Xue H, Guo X, Han X, et al. MicroRNA-584-3p, a novel tumor suppressor and prognostic marker, reduces the migration and invasion of human glioma cells by targeting hypoxia-induced ROCK1. Oncotarget. 2016;7(4):4785–4805. doi:10.18632/oncotarget.6735
26. Ueno K, Hirata H, Shahryari V, et al. Tumour suppressor microRNA-584 directly targets oncogene Rock-1 and decreases invasion ability in human clear cell renal cell carcinoma. Br J Cancer. 2011;104(2):308–315. doi:10.1038/sj.bjc.6606028
27. Duan G, Ren C, Zhang Y, Feng S. MicroRNA-26b inhibits metastasis of osteosarcoma via targeting CTGF and Smad1. Tumour Biol. 2015;36(8):6201–6209. doi:10.1007/s13277-015-3305-6
28. Tsai HC, Su HL, Huang CY, Fong YC, Hsu CJ, Tang CH. CTGF increases matrix metalloproteinases expression and subsequently promotes tumor metastasis in human osteosarcoma through down-regulating miR-519d. Oncotarget. 2014;5(11):3800–3812. doi:10.18632/oncotarget.1998
29. Wang MY, Chen PS, Prakash E, et al. Connective tissue growth factor confers drug resistance in breast cancer through concomitant up-regulation of Bcl-xL and cIAP1. Cancer Res. 2009;69(8):3482–3491. doi:10.1158/0008-5472.CAN-08-2524
30. Tsai HC, Chang AC, Tsai CH, et al. CCN2 promotes drug resistance in osteosarcoma by enhancing ABCG2 expression. J Cell Physiol. 2019;234(6):9297–9307. doi:10.1002/jcp.27611
31. Lu Y, Li F, Xu T, Sun J. Tetrandrine prevents multidrug resistance in the osteosarcoma cell line, U-2OS, by preventing Pgp overexpression through the inhibition of NF-kappaB signaling. Int J Mol Med. 2017;39(4):993–1000. doi:10.3892/ijmm.2017.2895
32. Xue X, Chen Q, Zhao G, Zhao JY, Duan Z, Zheng PS. The overexpression of TGF-beta and CCN2 in intrauterine adhesions involves the NF-kappaB signaling pathway. PLoS One. 2015;10(12):e0146159. doi:10.1371/journal.pone.0146159
33. Gong H-L, Tao Y, Mao X-Z, Song D-Y, You D, Ni J-D. MicroRNA-29a suppresses the invasion and migration of osteosarcoma cells by regulating the SOCS1/NF-κB signalling pathway through negatively targeting DNMT3B. Int J Mol Med. 2019;44(4):1219–1232. doi:10.3892/ijmm.2019.4287
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.