Back to Journals » OncoTargets and Therapy » Volume 11
MicroRNA-18a promotes hepatocellular carcinoma proliferation, migration, and invasion by targeting Bcl2L10
Authors Wang X, Lu J, Cao J, Ma B, Gao C, Qi F ![]()
Received 20 July 2018
Accepted for publication 10 October 2018
Published 9 November 2018 Volume 2018:11 Pages 7919—7934
DOI https://doi.org/10.2147/OTT.S180971
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr XuYu Yang
Xiaodong Wang,1 Jian Lu,1 Jisen Cao,2 Bozhao Ma,1 Chao Gao,1 Feng Qi1
1Department of General Surgery, Tianjin Medical University General Hospital, Tianjin, China; 2Department of Hepatobiliary Surgery, Tianjin Third Center Hospital, Tianjin, China
Background: Hepatocellular carcinoma (HCC) is known to feature several microRNA dysregulations. This study aimed to determine and investigate the prognostic value of microRNA (miRNA/miR)-18a and its role in regulating the progression of HCC.
Methods: miR-18a expressions in human HCC tissues, pair-matched adjacent normal liver tissues as well as in HCC cell lines were determined by quantitative real-time PCR. The prognostic value of miR-18a was determined using Kaplan–Meier survival analysis and multivariable Cox regression assay. The ability of miR-18a in promoting HCC progression was verified in vitro.
Results: miR-18a expressions in HCC tissues and cells were more than twice those of the normal control group (P<0.05). miR-18a expression was associated with the alpha-fetoprotein (AFP) level, TNM stage, tumor size, and intrahepatic vascular invasion (P<0.05). Kaplan–Meier survival analysis revealed that HCC patients with high expression of miR-18a possessed a more unfavorable prognosis (log-rank P<0.001). Overexpression of miR-18a promoted cell apoptosis and proliferation, induced S phase transition, as well as enhanced the migration and invasion ability of HCC cells. miR-18a was found to directly target the downstream molecule Bcl2L10. Furthermore, overexpressing Bcl2L10 was able to partly reverse the promoting effects of miR-18a on HCC cell progression.
Conclusion: miR-18a may serve as a prognostic biomarker of HCC as it is demonstrated to carry out a decisive role in HCC progression by promoting HCC cell invasion, migration, and proliferation through targeting Bcl2L10.
Keywords: miR-18a, hepatocellular carcinoma, Bcl2L10, apoptosis, cell cycle
Introduction
Hepatocellular carcinoma (HCC) is a frequently encountered malignancy throughout the world.1 More than 70% of the world’s new cases of HCC occur in Asia each year, and of these new cases, more than 50% of cases occur in China.2 At present, early diagnosis, surgical resection, and gene therapy are primary HCC treatment modalities.3,4 However, HCC still makes up the second most common cause of cancer-related mortality, resulting in high morbidity and mortality because there is no effective treatment so far.1,4,5 Although the diagnosis and treatment technology of HCC have developed considerably, the cure rate of HCC is still meager. Therefore, exploring the molecular mechanisms of HCC progression plays an important role in understanding HCC and developing appropriate treatment strategies.
In recent years, gene therapy has become an intense focus of research. Transporting tumor suppressor microRNAs (miRNAs) or inhibitors of some miRNAs via nanocarriers may be a new option for the treatment of cancer.6 However, at present, the mechanism by which miRNAs play a regulatory role in tumors needs to be further studied. miRNAs represent a distinct group of non-coding RNA molecules and regulate gene expression.7 A growing amount of evidence has shown that miRNA dysregulation is related to many human diseases and the proliferation and metastasis of various tumors, including HCC,8,9 breast cancer,10,11 lung cancer,12,13 colorectal cancer,14,15 and gastric cancer.16,17 Many miRNAs in HCC show deranged profiles, for example, microRNA 135a (miRNA/miR), miR-33a, miR-122, miR-18a, and miR-31.8,18,19 Studies have shown that HCC cell invasion and migration can be promoted by miR-135a through regulation of expression fork-head box O1 expressions.18 miR-33a downregulation correlates to chemotherapy resistance, tumorigenesis, poor patient prognosis.20,21 Several lines of evidence indicate that miRNA may possess significant regulatory effects on the development of HCC. The underlying mechanisms for the effects of miRNAs on HCC progression requires further investigation.
Dysregulation of miR-18a occurs in various tumors. It should be noted that miR-18a appears to exhibit different functions in different tumors. For instance, miR-18a can inhibit the progression of colorectal cancer by restraining K-Ras expression.22 miR-18a suppresses T24 cell proliferation by targeting Dicer.23 Otherwise, it has been reported that high circulating miR-18a expression in patients with non-small-cell lung cancer experienced poor prognoses.24 miR-18a promotes HCC cell migration and invasion through inhibiting Dicer I expression in vitro.25 Collectively, the role of miR-18a in tumors remains controversial. A study by Zhang et al26 indicated that miR-18a may augment HCC proliferation. However, data regarding the role and prognostic value of miR-18a in HCC patients are scarce.
Therefore, the current investigation seeks to clarify the effects and prognostic value of miR-18a on HCC progression. In addition, potential mechanisms underlying miR-18a-mediated promotion HCC progression are also studied.
Materials and methods
Patients and tissue specimens
This study included 123 patients with HCC diagnosed pathologically between January 2008 and December 2012 in Tianjin Third Center Hospital. None of the patient received preoperative therapy. Immediately after the samples were taken, the specimens were frozen in liquid nitrogen and stored at −80°C. All tissue specimens (both HCC tissues and adjacent healthy non-cancerous tissues) were harvested simultaneously. This study was approved by the Medical Ethics Committee of Tianjin Medical University (Ethical No TMUHMEC2015007), with the protocols in strict accordance to the Declaration of Helsinki. Written informed consent was sought from each patient prior to the procedure. After surgery, all patients included in our study were followed-up, and the survival information of the patients was obtained by telephone.
Cell culture and transfection
Human HCC cell lines HepG2 and SMMC7721 and normal liver cell line LO2 were purchased commercially from Tianjin Medical University Cancer Institute and Hospital. DMEM or RPMI-1640 media (Corning Incorporated, Corning, NY, USA) containing 10% FBS (Hyclone, Logan, UT, USA) were used to maintain cells. Cells and media were kept in a 5% CO2 environment in a incubator at 37°C. The medium was replaced every 2 days. miR-18a mimic (Ribobio, Guangzhou, China), miR-18a inhibitor (Ribobio), and Bcl2L10-overexpressing plasmids (Genechem, Shanghai, China) were used to transfect HCC cell lines in order to regulate miR-18a and Bcl2L10 expressions.
Luciferase reporter system
The target of miR-18a was predicted by TargetScan 7.2 (Supplementary material for TargetScan) and miRPathDB database (Supplementary material for miRPathDB). First, in the prediction results of TargetSacn 7.2 and miRPathDB database, the results of the coincidence of the two databases or the genes with the evidence level of “strong” in the miRPathDB results are considered as possible targets for miR-18a. These targets were then imported into TargetScan 7.2 to further identify the relationship between miR-18a and the target gene by searching for possible upstream miRNAs. Bcl2L10 is one of the possible targets of miR-18a. To confirm the relationship between miR-18a and Bcl2L10, we performed a luciferase activity assay. The luciferase reporter system was constructed by Genechem as previously described.27 Briefly, introduction of mutagenesis into the miR-18a binding site of Bcl2L10 mRNA was performed with the QuikChange Lightning Site-Directed Mutagenesis Kit (Stratagene, San Diego, CA, USA). A pGL3 luciferase vector (Promega Corporation, Fitchburg, WI, USA) was used to house the 3′-UTR fragment of wild-type (Wt) and mutant (Mut) Bcl2L10 mRNA. Subcloning was carried out with PCR. The vector was co-transfected with miR-18a mimic into HCC cell lines with transfection-enhancing reagent containing 5 μg/mL polybrene (Genechem) in 96-well plates for 12 hours. HCC cell lines were then cultured for 72 hours and lysed for subsequent luciferase assay, which was normalized against expression of renilla (Promega Corporation).
RNA extraction and quantitative real-time PCR (qRT-PCR)
Total RNA extraction was performed with an RNA extraction kit (QIAGEN China Co. Ltd., Shanghai, China). About 1 μg of total RNA was added to a 20 μL system. The GoScript Reverse Transcription system (Promega Corporation) enabled us to reverse transcribe Bcl2L10 mRNA, and miR-18a using the stem-loop method (QIAGEN China Co. Ltd.). Relative expressions were normalized against U6 and GAPDH. The primer sequences were as follows: Bcl2L10: forward, 5′-TGGAGAAAACAGCTGGTCCA-3′, reverse, 5′-AACACTGGCCAAATCACCAC-3′; miR-18a: forward, 5′-GATAGCAGCACAGAAATATTGGC-3′, universal primer, 5′-TGGTGTCGTGGAGTCG-3′; GAPDH: forward, 5′-TGCACCACCAACTGCTTAGC-3′, reverse, 5′-GGCATGGACTGTGGTCATGAG-3′; U6: 5′-CTCGCTTCGGCAGCACA-3′, reverse, 5′-AACGCTTCACGAATTTGCGT-3′. PCR amplification was performed for 40 cycles, and the relative levels were measured in triplicate using the 2−ΔΔCq method.
Wound scratch assay and transwell assay
The abilities of cells to migrate and invade were assessed by wound scratch assay and transwell assay with Matrigel (BD Biosciences, San Jose, CA, USA). For the wound scratch assay, the transfected cells were seeded in six-well plates and allowed to proliferate until 90% confluence was achieved. Serum starved media (1% FBS) was used to culture cells overnight. The next morning, the culture medium was aspirated and cells washed with PBS. Wounds were created with three parallel scratches drawn on the surface of the monolayer cells with a 1-mL pipette tip. Debris was removed through a PBS rinse, and cells were left to culture for another 24 hours. The distance between the wound edges were measured after the 24 hours. For the transwell assay, 200 μL of serum-free media containing 1×105 cells were added into the upper chamber, and 600 μL of 10% FBS medium was added to the bottom chamber. After a 24 hour incubation period at 37°C, Matrigel was removed and cells that penetrated the Matrigel and reached the basement membrane were fixed in 4% paraformaldehyde and underwent a 10-minute staining with 0.1% crystal violet. An inverted microscope was used to count the cells (Olympus Corporation, Tokyo, Japan).
Cell proliferation assay
A 96-well plate was used to seed the HepG2 and SMMC7721 cells at a density of 2×104 cells/well. The proliferation of HCC cells was measured on days 0–5. Before the test, each well received the addition of 10 μL of cell counting assay-8 (CCK-8) solution (Dojindo Laboratories, Kumamoto, Japan). Absorbance at 450 nm was measured by a microplate reader (Bio-Rad Laboratories Inc., Hercules, CA, USA) after cells were left to incubate for 3 hours at 37°C.
Flow cytometry analysis
The impact of miR-18a on cell apoptosis and the cell cycles of HepG2 and SMMC7721 cells were analyzed with flow cytometry. For apoptosis experiments, cells were harvested and resuspended in 200 μL PBS. Propidium iodide (PI) and FITC-labeled Annexin V (BD Biosciences) were then added into the mixture and incubated for 30 minutes at room temperature. For cell cycle assay, the harvested cells were fixed in 70% pre-cooled ethanol. The samples were taken before DNA staining, and the supernatant was removed by centrifugation. After the samples were treated with RNAase (0.1 mg/mL; Sigma-Aldrich Co., St Louis, MO, USA) for 30 minutes, 1 mL of PI (50 μg/mL; Sigma-Aldrich Co.) was added, and flow cytometry analyses was performed to assess the distribution of cells in each phase of the cell cycle.
Western blot analysis
Western blot was carried out as described in our previous study.28 Briefly, total protein of HCC cells and tissues together with healthy liver tissues were lysed with RIPA lysate (containing protease inhibitors and phosphatase inhibitors, 1%) in order to extract the total protein. About 40 μg of total protein in each sample was added to a sample well of a 10% sodium dodecyl sulfate-polyacrylamide gel (SDS-PAGE) and electrophoresed. Separated proteins were then blotted onto a polyvinylidene fluoride membrane (EMD Millipore, Billerica, MA, USA). Membranes containing the imprinted protein were blocked with 0.5% bovine serum albumin for 2 hours, then the membrane was incubated with anti-Bcl2L10 (1:1,500; Cell Signaling Technology [CST], Danvers, MA, USA) or anti-β-actin (1:3,000; CST) antibodies overnight at 4°C. The next day, the membranes were rinsed thrice and further left to incubate with diluted secondary horseradish peroxidase-marked antibodies (1:3,000; CST) for 1 hour at room temperature. An enhanced chemiluminescence detection kit (EMD Millipore) was used to detect immunoreactive protein bands. Images were captured by ImageJ (National Institute of Mental Health, Bethesda, MR, USA).
Immunohistochemistry assay
The HCC tissues and adjacent healthy tissues were fixed in 4% paraformaldehyde, embedded in paraffin and sliced into 4 μm sections. Dehydration was carried out with xylene and a gradient of ethanol solution. Distilled water was used to wash the sections before endogenous peroxidase blocking was carried out with a 3% H2O2 solution. The heated sodium citrate solution (92°C–95°C, 10 nM, pH 6.0) was used to repair the antigens for 5 minutes. After rinsing the sections twice with PBS, the sections were exposed to 1% goat serum at room temperature for 2 minutes to block the sections. Rabbit antihuman Bcl2L10 antibody (1:150; Cell Signaling Technology) was applied onto the sections and left overnight to incubate at 4°C. Phosphate-buffered saline was used to wash the sections for three times prior to the addition of the secondary antibody. Freshly prepared diaminobenzidine was then added to each section after an hour of incubation at room temperature. The final step involved re-staining with hematoxylin and cell dehydration and sealing. A light microscope (Leica Microsystems, Wetzlar, Germany) was used to visualize slides, and brown particles in the cytoplasm or cytomembranes were taken to indicate positive staining.
Statistical analysis
All data were expressed in terms of mean ± SD. The relationship between miR-18a and clinical pathology features of patients with HCC was analyzed by the chi-squared test. Kaplan–Meier and log-rank tests were carried out to determine how varying miR-18a expression levels affected HCC patient survival. The prognostic value of miR-18a was determined by Cox regression analysis. Continuous data were analyzed using a t-test or one-way ANOVA. The correlation between miR-18a and Bcl2L10 was analyzed utilizing Spearman or Pearson correlation analysis. All statistical analyses were carried out with the SigmaPlot software (SPSS 19.0; IBM Corporation, Armonk, NY, USA). A P-value of <0.05 was taken to indicate statistical significance. All values are the representative of three separate experiments.
Results
Overexpression of miR-18a in HCC tissues and HCC cell lines
We sought to document the variations in miR-18a expression among HCC tissues, pair-matched adjacent healthy tissues, and HCC cells. Analysis revealed that the HCC cells and tissues possessed higher miR-18a expressions compared to the control groups (P<0.001) (Figure 1A and B).
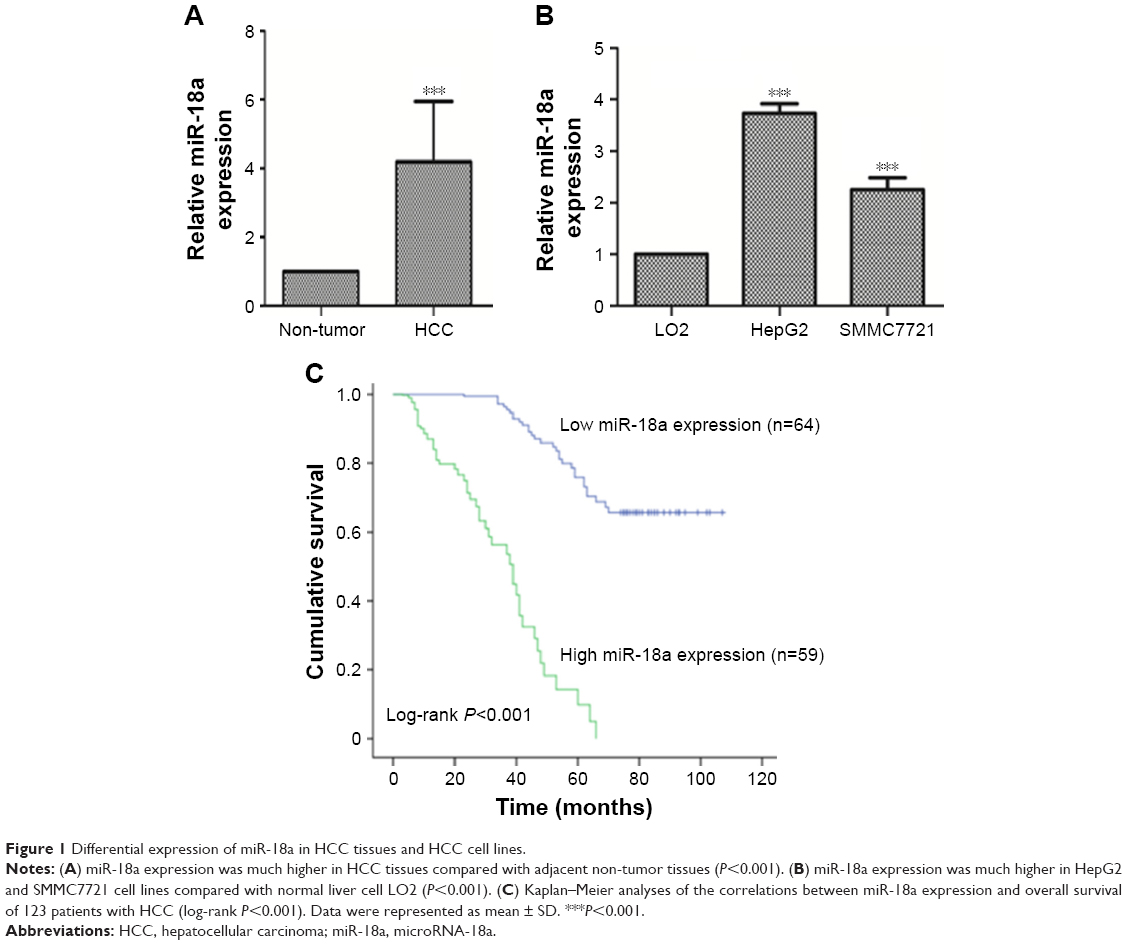 | Figure 1 Differential expression of miR-18a in HCC tissues and HCC cell lines. |
Increased miR-18a in HCC tissues was associated with disease progression and metastasis
We further sought to confirm the clinical significance of miR-18a with regard to HCC prognosis. Patients with HCC were divided into groups based on their mean relative miR-18a expression (4.193), resulting in two groups who had either low expressions (n=64) or high expressions (n=59). Chi-squared test results showed that the expression of miR-18a expression correlated with the serum level of AFP (P=0.023), tumor size (P<0.001), TNM stage (P<0.001), and intrahepatic vascular invasion (P<0.001). Age and gender were not significantly correlated to miR-18a expression levels (P>0.05) (Table 1). As shown in Figure 1C, the Kaplan–Meier survival curves indicated that patients who had raised expressions of miR-18a experienced poorer overall survival rates in contrast to those who had low expressions of miR-18a (log-rank P<0.001). Furthermore, we analyzed the effect of miR-18a expression and other clinical features on overall survival prognosis in patients with HCC by Cox regression analysis. Results indicated that miR-18a expression (Exp(B) =2.017, 95% CI =1.137–3.578, P=0.017), TNM stage (Exp(B) =4.396, 95% CI =0.164–0.527, P<0.001), and intrahepatic vascular invasion (Exp(B) =0.294, 95% CI =1.987–9.728, P<0.001) were independent prognostic factors for HCC (Table 2).
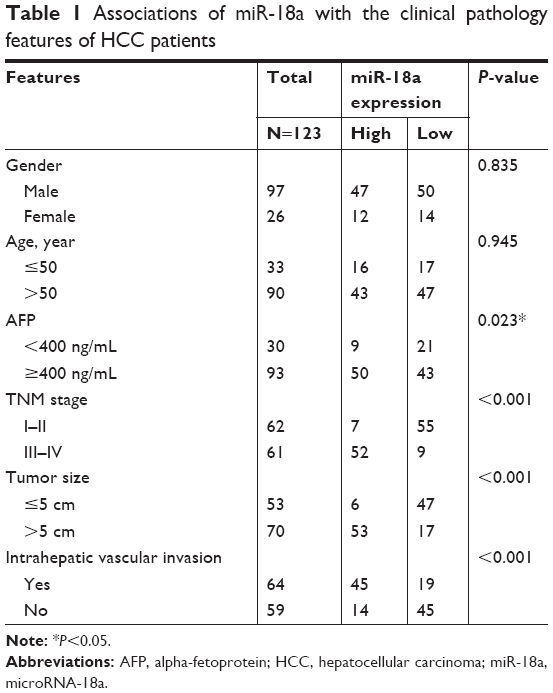 | Table 1 Associations of miR-18a with the clinical pathology features of HCC patients |
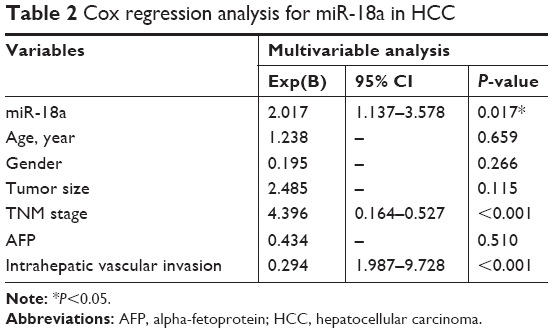 | Table 2 Cox regression analysis for miR-18a in HCC |
miR-18a promoted proliferation, migration, and invasion of HCC cells
We next investigated the in vitro effects of miR-18a on HCC cells. miR-18a mimic, inhibitor, or their negative control (NC) were used to regulate miR-18a level in HepG2 and SMMC7721 cell lines. The efficiency of miR-18a mimic and inhibitor was confirmed with qRT-PCR. As shown in Figure 2A, the miR-18a expression was raised in HCC cells transfected with the miR-18a mimic (P<0.01) but reduced in cells that were transfected with the miR-18a inhibitor (P<0.05) compared with the NC miRNA.
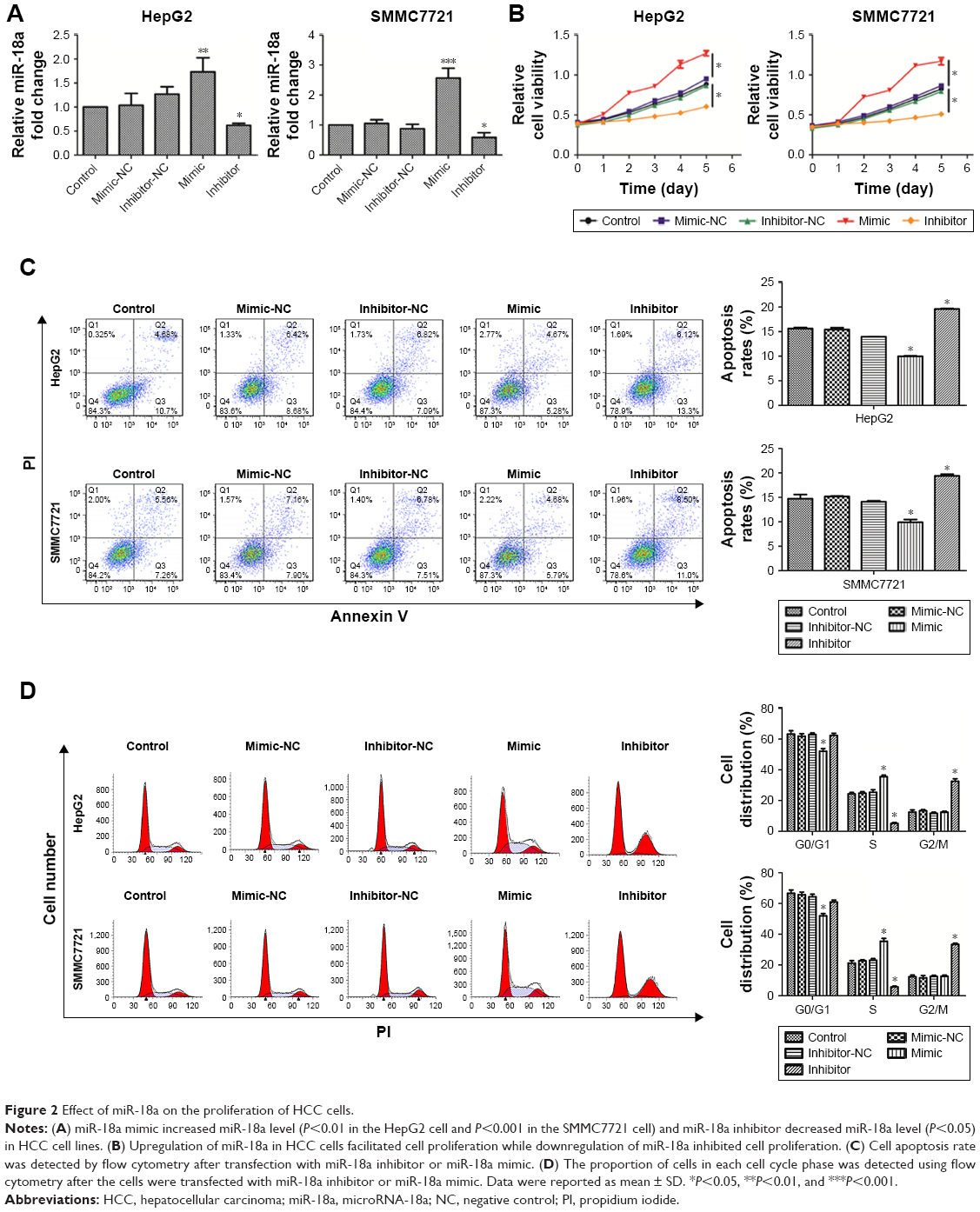 | Figure 2 Effect of miR-18a on the proliferation of HCC cells. |
A CCK-8 assay was performed to explore the impact of miR-18a expression on the proliferation of HepG2 and SMMC7721 cells. As shown in Figure 2B, overexpressing miR-18a promoted the proliferation of HepG2 and SMMC7721 HCC cells (P<0.05). In contrast, significant inhibition of cell proliferation was noted in cells transfected with miR-18a inhibitor (P<0.01). Furthermore, the effects of miR-18a on HCC cell cycle progression and apoptosis were analyzed by flow cytometry. The overexpression of miR-18a inhibited the apoptosis of HCC cell lines, while the apoptosis rate was markedly raised after miR-18a knockdown (P<0.05) (Figure 2C). Analysis of proportion of cells in each cell cycle showed that the percentage of S phase was increased and G0/G1 phase was decreased after miR-18a overexpression in HCC cell lines, whereas knockdown of miR-18a resulted in a decrease in the percentage of cells in the S phase and increased those in the G2/M phase (Figure 2D). Collectively, these data indicated that overexpression of miR-18a promoted HCC progression, and knockdown of miR-18a could repress HCC cell-induced apoptosis and proliferation.
How miR-18a affected HCC cell migration and invasion were investigated with a wound scratch assay and transwell assay. The open wound area was quantified as a “mean % migration distance.” As shown in Figure 3A and B, elevated miR-18a expressions markedly raised HCC cell invasion and migration (all P<0.05), whereas downregulation of miR-18a decreased the invasion and migration of HepG2 and SMMC7721 cells. These findings indicate that HCC progression may be regulated by miR-18a through stimulation of HCC cell metastasis (all P<0.05).
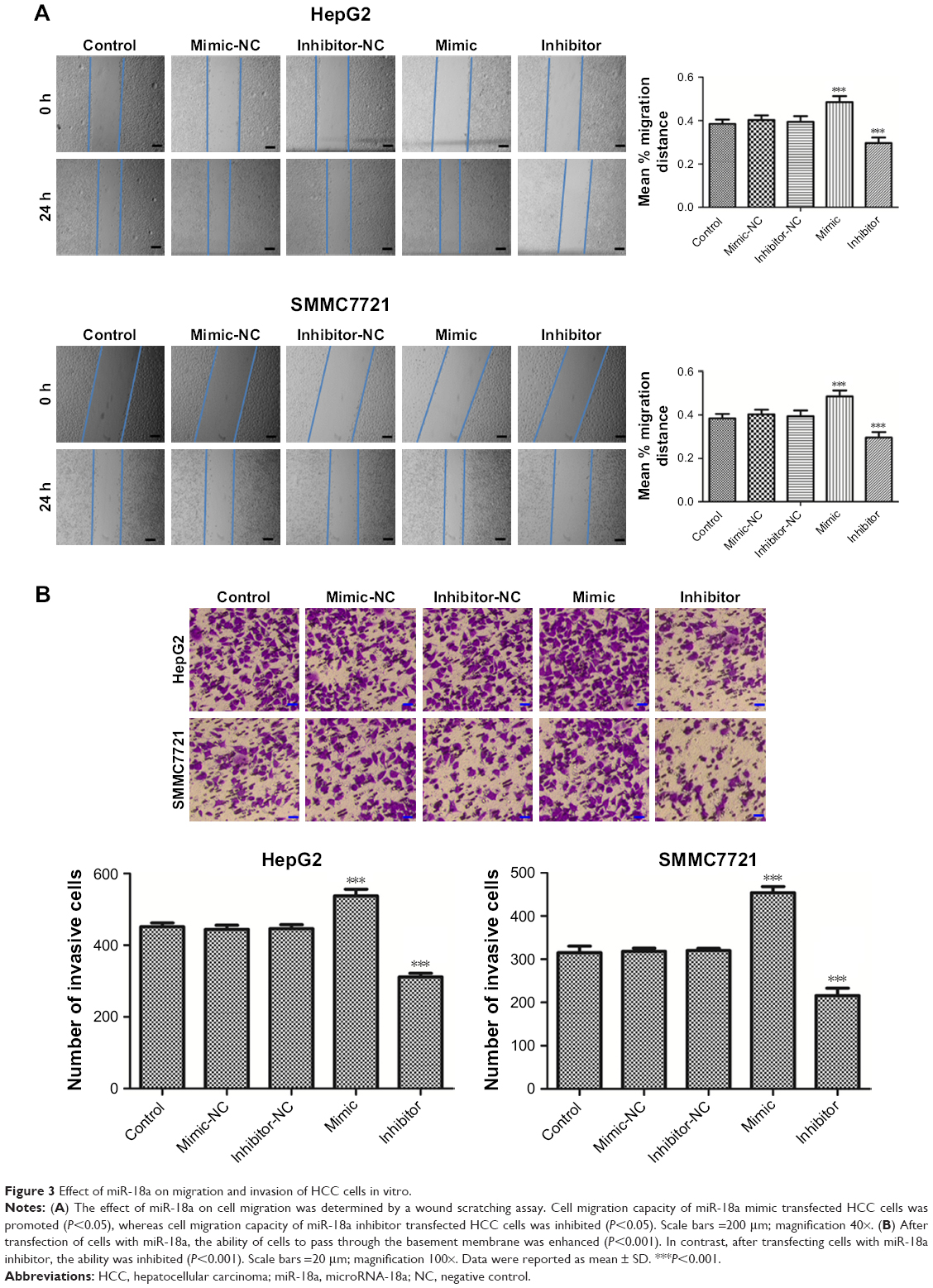 | Figure 3 Effect of miR-18a on migration and invasion of HCC cells in vitro. |
Bcl2L10 was a direct target of miR-18a in HCC cells
To explain how miR-18a promoted HCC progression, TargetScan 7.2 and miRPathDB databases were used to identify potential miR-18a gene targets. The prediction results revealed that Bcl2L10 was a potential miR-18a target in human tissues. A luciferase reporter assay was then carried out to determine if Bcl2L10 is a direct target of miR-18a in HCC cells. The 3′-UTR regions of Bcl2L10 containing the predicted binding site of miR-18a (Bcl2L10 3′UTR-Wt) or the mutant site (Bcl2L10 3′UTR-Mut) were cloned into a luciferase vector (Figure 4A). Luciferase activity results showed that luciferase activities of Bcl2L10-3′-UTR-Wt reporter miR-18a mimic were significantly attenuated in both cell lines (P<0.001), while no inhibitory effects on the luciferase activity of Bcl2L10-3′-UTR-Mut reporter in HCC cells was seen (P>0.05) (Figure 4B).
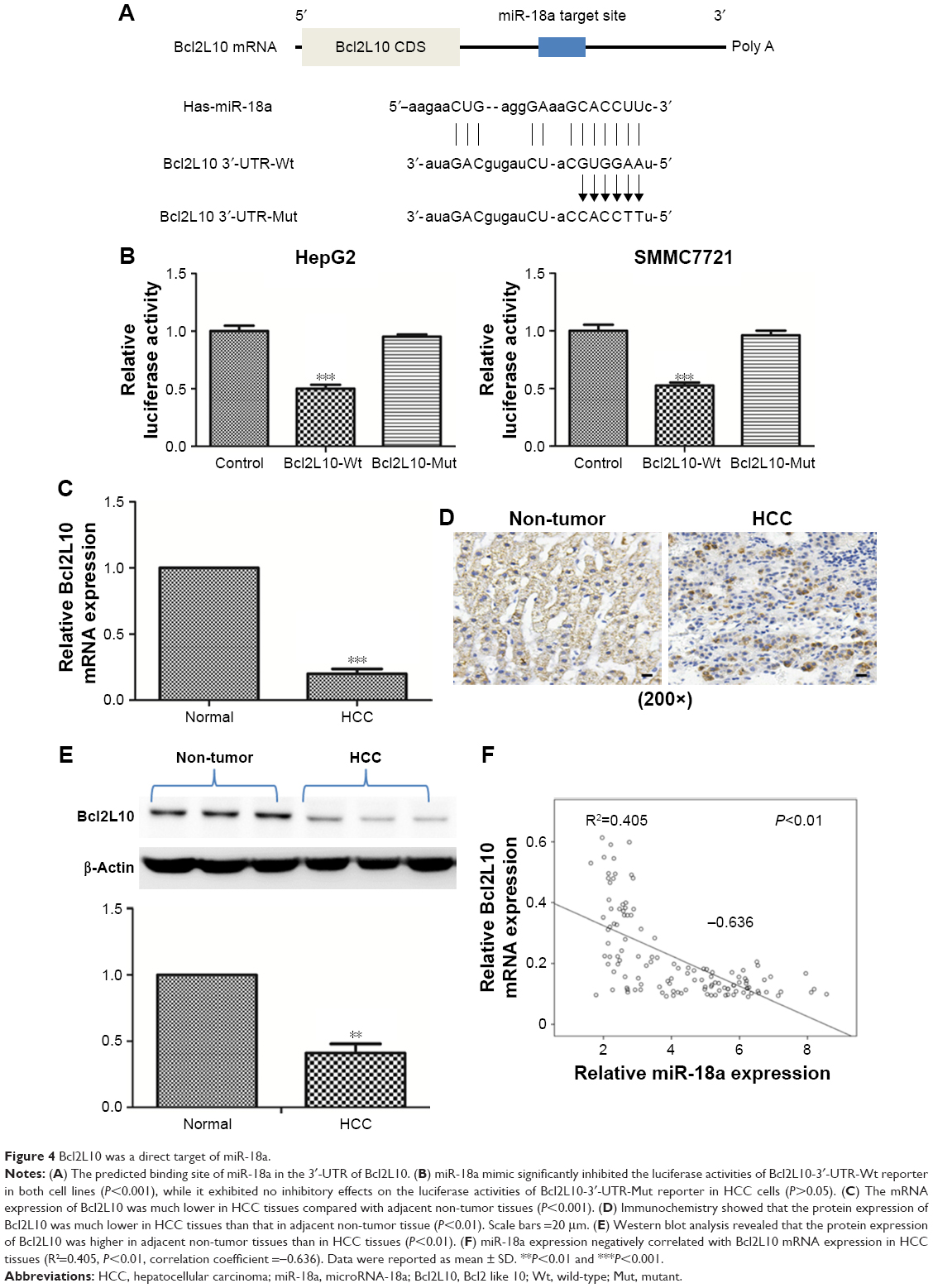 | Figure 4 Bcl2L10 was a direct target of miR-18a. |
Besides, we detected mRNA and protein level of Bcl2L10 in HCC tissues. Figure 4C shows that the mRNA level of Bcl2L10 was higher in adjacent no-tumor tissues than in HCC tissues (P<0.01). In addition, we determined the protein levels of Bcl2L10 by immunohistochemistry and Western blot. As shown in Figure 4D and E, Bcl2L10 protein was markedly diminished in HCC tissues (P<0.01). Spearman correlation analysis showed that the Bcl2L10 mRNA expression correlated negatively with miR-18a expressions in HCC tissues (Figure 4F).
miR-18a inhibits Bcl2L10 expression in HCC cells
The results of luciferase reporter assay have suggested that miR-18a may directly target Bcl2L10 in HCC cells. Therefore, to validate how miR-18a affected Bcl2L10 expressions in HCC cells, Bcl2L10 mRNA and protein levels in transfected HCC cells were examined by qRT-PCR and Western blot. We confirmed that overexpressing miR-18a strongly decreased Bcl2L10 mRNA and protein level in HepG2 and SMMC7721 cells (P<0.01) (Figure 5A and B). Conversely, when miR-18a was knocked down, the expression of Bcl2L10 was raised in both the cell lines (P<0.01), indicating that miR-18a may promote the progression of HCC by inhibiting the expression of Bcl2L10.
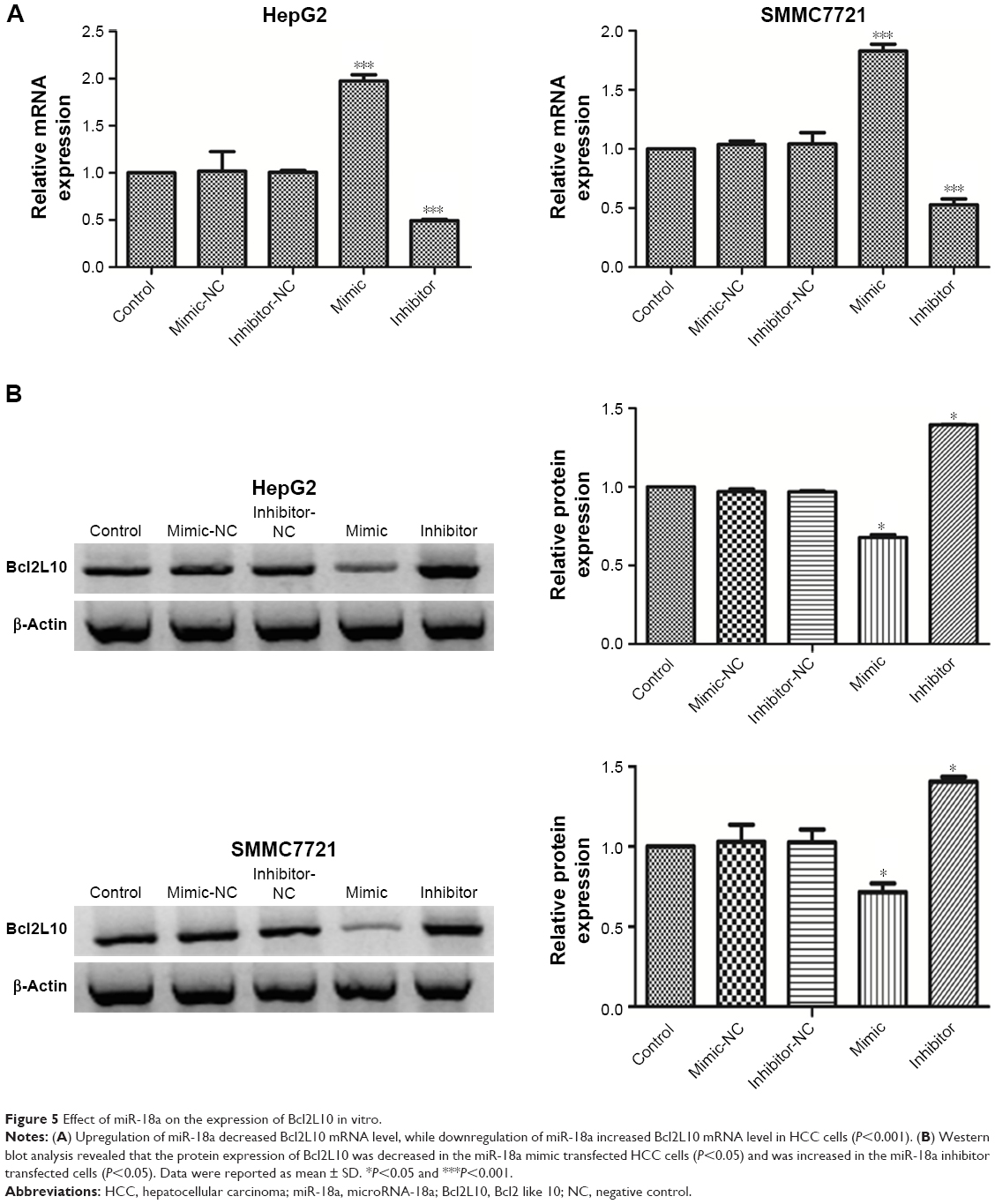 | Figure 5 Effect of miR-18a on the expression of Bcl2L10 in vitro. |
Overexpression of Bcl2L10 partially reversed the effect of miR-18a in HCC cells
To test whether Bcl2L10 can offset the effect of miR-18a in HCC cells, we transfected Bcl2L10 plasmids to the HCC cell lines and further analyzed the proliferative, invasive, and migratory abilities of HCC cell lines. Figure 6A shows that the mRNA and protein expressions of Bcl2L10 markedly increased in the two cell lines after transfected with Bcl2L10 plasmids (P<0.001).
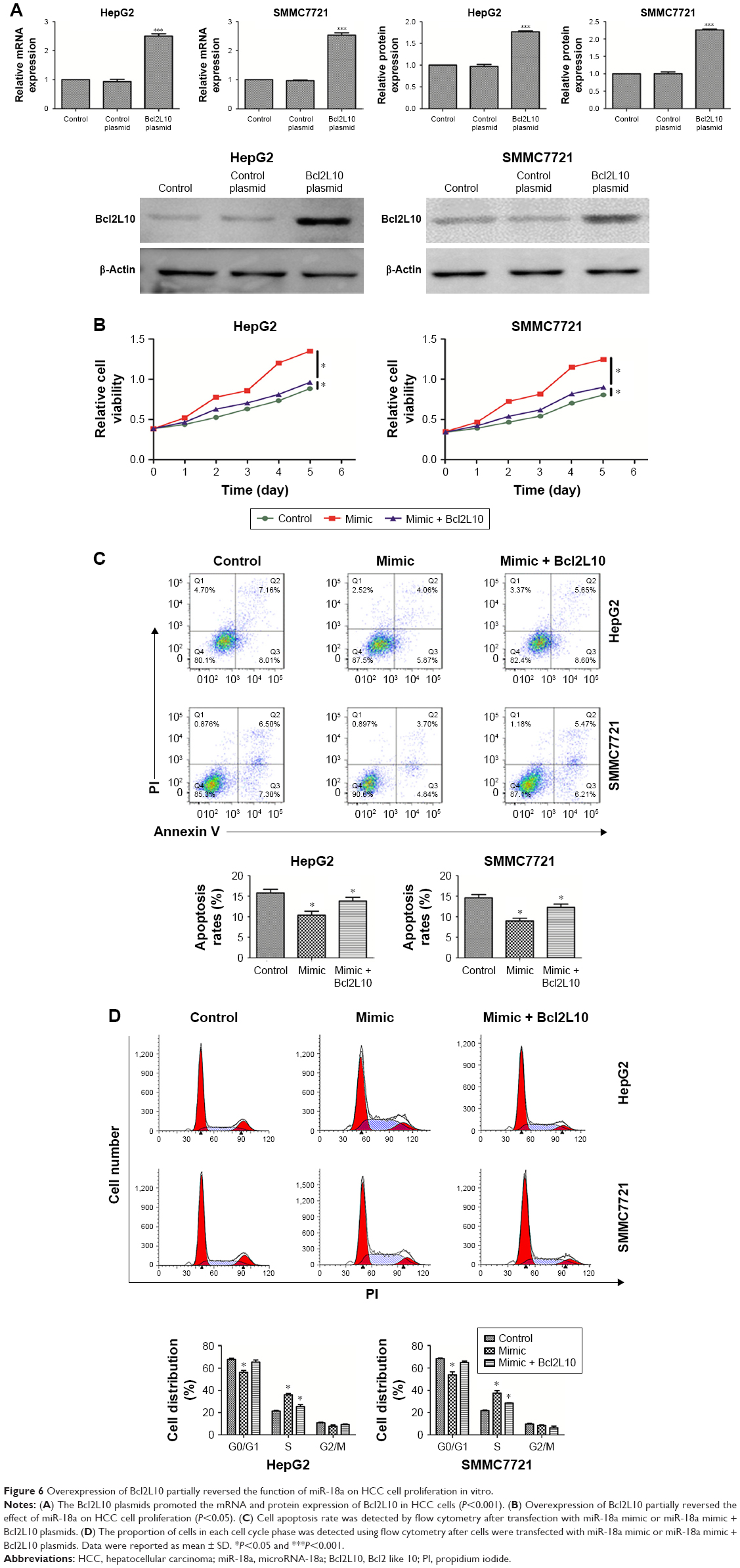 | Figure 6 Overexpression of Bcl2L10 partially reversed the function of miR-18a on HCC cell proliferation in vitro. |
A CCK-8 assay was carried out to detect the effect of Bcl2L10 on the proliferation of miR-18a mimic transfected HCC cells. Figure 6B demonstrates that an overexpression of Bcl2L10 in miR-18a transfected HCC cells significantly diminished HCC cell proliferation, in comparison with miR-18a transfected cells. However, Bcl2L10 cannot completely reverse the pro-proliferative effect of miR-18a on HCC cells. Moreover, increased Bcl2L10 expressions in miR-18a transfected HCC cells induced apoptosis of HCC cells (P<0.05) (Figure 6C) and reduced the proportion of cells in the S phase (P<0.05) (Figure 6D).
Lastly, we sought to document the impact of Bcl2L10 on invasion and migration of HepG2 and SMMC7721 cells. In contrast to the miR-18a mimic group, the invasive and migratory abilities of HepG2 and SMMC7721 cells in the Bcl2L10+ miR-18a mimic group were inhibited (P<0.05), but were still higher than the control group (Figure 7A and B), indicating that Bcl2L10 can partially reverse pro-migratory and pro-invasive properties of miR-18a on HCC cells.
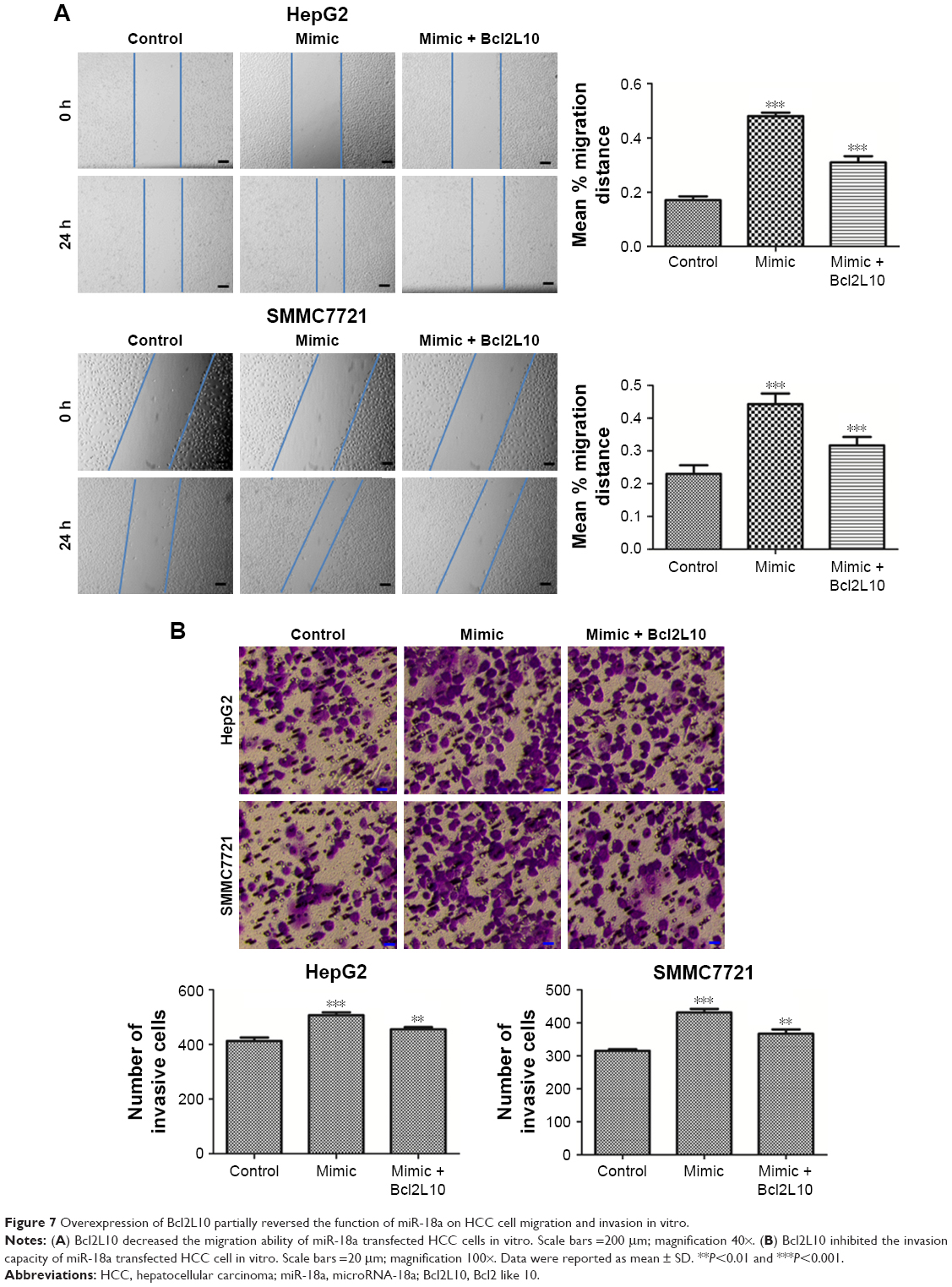 | Figure 7 Overexpression of Bcl2L10 partially reversed the function of miR-18a on HCC cell migration and invasion in vitro. |
Discussion
miRNA dysregulation occurs in several diseases, especially tumors. Therefore, miRNAs may become potential targets for the development of future chemotherapeutic medications. Although there are still many problems to be resolved in miRNA-based targeted therapy, such as low transfection efficiency and high toxicity, a novel nanomaterial carrier-based research has made great progress in overcoming these hurdles.29,30 Therefore, studying the clinical significance and function of miRNA along with its possible mechanisms of action may contribute greatly toward the gene therapy in cancer. Many studies have demonstrated that miRNA dysregulation plays an essential role in HCC tumorigenesis and progression.31–34 Although miRNA overexpression or downexpression have both been proved to contribute to HCC tumorigenesis and progression, the prognostic value of miRNAs and potential mechanisms still need to be further studied. Here in our present study, we focus on miR-18a, a potential screening biomarker for HCC.35 We demonstrated that elevated expressions of miR-18a in HCC tissues was related to cancer progression and poor patient prognosis. In vitro experiments revealed miR-18a to be a promotor for HCC cell invasion, migration, and proliferation by inhibiting Bcl2L10, indicating that miR-18a possesses a decisive function in dictating the progression of HCC.
miR-18a dysregulation is associated with a myriad of conditions and functions as a crucial mediator of tumor progression.24,36–39 For example, miR-18a is associated with breast cancer resistance to trastuzumab40 and promoting tumor progression.41,42 These results may indicate that miR-18a is an essential miRNA in tumors. Existing literature has demonstrated an increase in miR-18a levels in HCC cell lines and tissues, further supporting the functions of miR-18a in HCC progression.26,43 However, the prognostic value has rarely been reported until now. As miR-18a may act as a biomarker in HCC, we first explored the association between miR-18a expression and clinical characteristics of HCC patients. We collected HCC tissues and pair-matched adjacent healthy liver tissues of 123 patients with primary HCC and detected miR-18a expression using qRT-PCR. Similarly, the results of our research show that miR-18a expressions were raised in HCC tissues in contrast to healthy cells. Moreover, miR-18a expressions were found to be linked with AFP level, TNM stage, tumor size, and intrahepatic vascular invasion. Since HCC tissues demonstrate a marked increase in miR-18a expressions, we further explored the effect of miR-18a expression on patient prognosis. Kaplan–Meier analysis showed that patients with higher expression of miR-18a had significantly shorter survival time compared to low-expressing miR-18a HCC patients. miR-18a was found to be an independent prognostic factor for patients with HCC as shown by multivariable COX regression analysis.
Tumor proliferation and metastasis are essential for tumor progression. Consistent with previous studies, miR-18a expression was much higher in HCC cell lines compared with normal liver cells LO2. To explore the in vitro function of miR-18a in HCC in vitro, miR-18a mimic and miR-18a inhibitor were used to regulate miR-18a expression in HCC cells. HCC cell invasion, migration, and proliferation were analyzed after being transfected with either a mimic or an inhibitor. Overexpressed miR-18a significantly promoted the proliferation, invasion, and migration abilities of SMMC7721 and HepG2 cells. Conversely, knockdown miR-18a inhibited cell invasion, migration, and proliferation. In addition, overexpression of miR-18a can inhibit the apoptosis of HepG2 and SMMC7721 cells and promote more cells to undergo the S phase transition, whereas knockdown expression of miR-18a can promote the apoptosis of HepG2 and SMMC7721 and induce G2/M arrest of HCC cell lines. Collectively, our findings support our hypothesis that miR-18a is able to promote HCC progression.
Many studies have demonstrated that miRNAs mainly participate in post-transcriptionally regulated gene expression by attaching to the 3′-UTR region of mRNA, leading to translational repression or mRNA degradation. Therefore, we hypothesized that miR-18a might affect HCC cell proliferation, migration, and invasion through targeting of downstream genes expression. Potential targets of miR-18a were predicted with TargetScan 7.2 and miRPathDB databases, which revealed Bcl2L10 to be a potential miR-18a target in human tissues. Some studies have revealed that Bcl2L10 may have a tumor suppressing function. For instance, Bcl2L0 binds to IRBIT and promotes cell apoptosis.44 In gastric cancer, Bcl2L10 inhibits gastric cancer proliferation and induces apoptosis through a variety of mechanisms.45 Previous studies have reported that HCC cell migration and proliferation may be attenuated by Bcl2L10.46 We considered that miR-18a might affect the biological behavior of HCC by affecting the expression of Bcl2L10. A luciferase reporter assay was done to confirm that miR-18a directly targeted Bcl2L10 in HCC cells. Results demonstrated that miR-18a directly targeted Bcl2L10 in HCC cells and can be post-transcriptionally inhibited by miR-18a. To confirm that miR-18a could regulate Bcl2L10 expression, we quantified Bcl2L10 protein and mRNA expressions in HCC tissues and miR-18a mimic-transfected HCC cell lines. Both Bcl2L10 protein and mRNA expressions in HCC tissues were decreased in contrast to healthy tissues. Correlation analysis demonstrated miR-18a levels to be negatively correlated with Bcl2L10 mRNA expressions. Similarly, in HepG2 and SMMC7721 cells, miR-18a mimic transfection of SMMC7721 and HepG2 cells resulted in much lower protein and mRNA expressions of Bcl2L10. These findings indicate that Bcl2L10 was targeted directly by miR-18a in HCC. Inhibiting Bcl2L10 expression by miR-18a may be a potential promotor for HCC progression.
To further confirm if Bcl2L10 represented a credible downstream mediator of miR-18a in promoting HCC cell proliferation and migration, we further transferred Bcl2L10 plasmids to HCC cells to induce Bcl2L10 expression in miR-18a transfected HCC cells. Results showed that Bcl2L10 partially reversed miR-18a effects on cell invasion, migration, and proliferation. Together, miR-18a may promote HC cell invasion, migration, and proliferation by acting on Bcl2L10. However, these results do not entirely explain the only partial reversal of miR-18a effects by Bcl2L10. One possible reason is that miR-18a may act on multiple targets simultaneously. For example, Liu et al have reported that miR-18a might promote HCC proliferation and metastasis by targeting KLF4.47 Their research shows that HCC cell migration and proliferation is promoted by miR-18a through targeting of the KLF4 molecule. KLF4 is a transcription factor of the KLF family with a zinc finger structure and plays an important role in cell differentiation and development. In addition to inducing HCC cell migration and proliferation, miR-18a also inhibited apoptosis and initiated cell cycle transition. Overexpression of Bcl2L10 can promote apoptosis of HepG2 and SMMC7721 cells. In conclusion, we found that miRNA may act on different targets and exert different biological effects and we discovered a new target of miR-18a in HCC cells in our current investigation. Moreover, this may also suggest why Bcl2L10 only partially reverses the effect of miR-18a in HCC cell lines. The function of miR-18a in HCC still need to be further studied.
Conclusion
We discovered a new target for miR-18a to regulate the prognosis of HCC. Increased miR-18a expression predicts a poor prognosis of patients with HCC. miR-18a augments HCC metastasis and proliferation through Bcl2L10. Overexpression of Bcl2L10 partially reverses the effect of miR-18a on HCC. Therefore, reducing the expression of miR-18a in HCC cells or increasing the expression of its downstream target gene Bcl2L10 may both be effective methods for treating HCC.
Acknowledgments
The National Natural Science Foundation of China (8157020853) and “Thirteenth Five” comprehensive investment discipline construction project supported this work. Xiaodong Wang and Jian Lu are co-first authors for this study.
Disclosure
The authors report no conflicts of interest in this work.
References
Siegel RL, Miller KD, Jemal A. Cancer statistics, 2018. CA Cancer J Clin. 2018;68(1):7–30. | ||
Ashtari S, Pourhoseingholi MA, Sharifian A, Zali MR. Hepatocellular carcinoma in Asia: Prevention strategy and planning. World J Hepatol. 2015;7(12):1708–1717. | ||
Li W, Man W, Guo H, Yang P. Clinical study of transcatheter arterial chemoembolization combined with microwave ablation in the treatment of advanced hepatocellular carcinoma. J Cancer Res Ther. 2016;12(Suppl):C217–C220. | ||
Takeyama H, Beppu T, Higashi T, et al. Impact of surgical treatment after sorafenib therapy for advanced hepatocellular carcinoma. Surg Today. 2018;48(4):431–438. | ||
Okinaga H, Yasunaga H, Hasegawa K, Fushimi K, Kokudo N. Short-Term Outcomes following Hepatectomy in Elderly Patients with Hepatocellular Carcinoma: An Analysis of 10,805 Septuagenarians and 2,381 Octo- and Nonagenarians in Japan. Liver Cancer. 2018;7(1):55–64. | ||
Liang C, Sun W, He H, et al. Antitumor effect of a new nano-vector with miRNA-135a on malignant glioma. Int J Nanomedicine. 2018;13:209–220. | ||
di Leva G, Garofalo M, Croce CM. MicroRNAs in cancer. Annu Rev Pathol. 2014;9:287–314. | ||
Mourad L, El-Ahwany E, Zoheiry M, et al. Expression analysis of liver-specific circulating microRNAs in HCV-induced hepatocellular Carcinoma in Egyptian patients. Cancer Biol Ther. 2018;19(5):400–406. | ||
Zhang X, Zhang X, Wang T, et al. MicroRNA-26a is a key regulon that inhibits progression and metastasis of c-Myc/EZH2 double high advanced hepatocellular carcinoma. Cancer Lett. 2018;426:98–108. | ||
Evans-Knowell A, Larue AC, Findlay VJ. MicroRNAs and Their Impact on Breast Cancer, the Tumor Microenvironment, and Disparities. Adv Cancer Res. 2017;133:51–76. | ||
Zhuang C, Yuan Y, Song T, et al. miR-219a-5p inhibits breast cancer cell migration and epithelial-mesenchymal transition by targeting myocardin-related transcription factor A. Acta Biochim Biophys Sin. 2017;49(12):1112–1121. | ||
Arab A, Karimipoor M, Irani S, et al. Corrigendum to “Potential circulating miRNA signature for early detection of NSCLC” [Cancer Genetics 216–217 (2017) 150–158]. Cancer Genet. 2018 Epub Jun 15. | ||
Catacchio I, Scattone A, Silvestris N, Mangia A. Immune Prophets of Lung Cancer: The Prognostic and Predictive Landscape of Cellular and Molecular Immune Markers. Transl Oncol. 2018;11(3):825–835. | ||
Slaby O, Svoboda M, Michalek J, Vyzula R. MicroRNAs in colorectal cancer: translation of molecular biology into clinical application. Mol Cancer. 2009;8:102. | ||
Xu P, Wang J, Sun B, Xiao Z. Integrated analysis of miRNA and mRNA expression data identifies multiple miRNAs regulatory networks for the tumorigenesis of colorectal cancer. Gene. 2018;659:44–51. | ||
Zhao G, Jiang T, Liu Y, et al. Droplet digital PCR-based circulating microRNA detection serve as a promising diagnostic method for gastric cancer. BMC Cancer. 2018;18(1):676. | ||
Zhang F, Xu W, Liu J, et al. Optimizing miRNA-module diagnostic biomarkers of gastric carcinoma via integrated network analysis. PLoS One. 2018;13(6):e0198445. | ||
Zeng YB, Liang XH, Zhang GX, et al. miRNA-135a promotes hepatocellular carcinoma cell migration and invasion by targeting forkhead box O1. Cancer Cell Int. 2016;16:63. | ||
Xie RT, Cong XL, Zhong XM, et al. MicroRNA-33a downregulation is associated with tumorigenesis and poor prognosis in patients with hepatocellular carcinoma. Oncol Lett. 2018;15(4):4571–4577. | ||
Chang W, Zhang L, Xian Y, Yu Z. MicroRNA-33a promotes cell proliferation and inhibits apoptosis by targeting PPARα in human hepatocellular carcinoma. Exp Ther Med. 2017;13(5):2507–2514. | ||
Meng W, Tai Y, Zhao H, et al. Downregulation of miR-33a-5p in Hepatocellular Carcinoma: A Possible Mechanism for Chemotherapy Resistance. Med Sci Monit. 2017;23:1295–1304. | ||
Tsang WP, Kwok TT. The miR-18a* microRNA functions as a potential tumor suppressor by targeting on K-Ras. Carcinogenesis. 2009;30(6):953–959. | ||
Tao J, Wu D, Li P, Xu B, Lu Q, Zhang W. microRNA-18a, a member of the oncogenic miR-17–92 cluster, targets Dicer and suppresses cell proliferation in bladder cancer T24 cells. Mol Med Rep. 2012;5(1):167–172. | ||
Xu X, Zhu S, Tao Z, Ye S. High circulating miR-18a, miR-20a, and miR-92a expression correlates with poor prognosis in patients with non-small cell lung cancer. Cancer Med. 2018;7(1):21–31. | ||
Zhang X, Yu B, Zhang F, Guo Z, Li L. microRNA-18a Promotes Cell Migration and Invasion Through Inhibiting Dicer l Expression in Hepatocellular Carcinoma In Vitro. Chin Med Sci J. 2017;32(1):34–33. | ||
Zhang YY, Yang LW, Liu J, Sun YQ. MicroRNA-18a targets IRF2 and CBX7 to promote cell proliferation in hepatocellular carcinoma. Oncol Res. Epub 2018 Jan 31. | ||
Ding M, Bowman L, Castranova V. Luciferase reporter system for studying the effect of nanoparticles on gene expression. Methods Mol Biol. 2012;906:403–414. | ||
Chen D, Li Y, Wang X, et al. Generation of regulatory dendritic cells after treatment with paeoniflorin. Immunol Res. 2016;64(4):988–1000. | ||
Li Y, Duo Y, Bi J, et al. Targeted delivery of anti-miR-155 by functionalized mesoporous silica nanoparticles for colorectal cancer therapy. Int J Nanomedicine. 2018;13:1241–1256. | ||
Küçüktürkmen B, Bozkir A. Development and characterization of cationic solid lipid nanoparticles for co-delivery of pemetrexed and miR-21 antisense oligonucleotide to glioblastoma cells. Drug Dev Ind Pharm. 2018;44(2):306–315. | ||
Xie F, Yuan Y, Xie L, et al. miRNA-320a inhibits tumor proliferation and invasion by targeting c-Myc in human hepatocellular carcinoma. Onco Targets Ther. 2017;10:885–894. | ||
Yu M, Lin Y, Zhou Y, et al. MiR-144 suppresses cell proliferation, migration, and invasion in hepatocellular carcinoma by targeting SMAD4. Onco Targets Ther. 2016;9:4705–4714. | ||
Liu W, Hu J, Zhou K, et al. Serum exosomal miR-125b is a novel prognostic marker for hepatocellular carcinoma. Onco Targets Ther. 2017;10:3843–3851. | ||
Liu W, Kang L, Han J, et al. miR-342-3p suppresses hepatocellular carcinoma proliferation through inhibition of IGF-1R-mediated Warburg effect. Onco Targets Ther. 2018;11:1643–1653. | ||
Li L, Guo Z, Wang J, Mao Y, Gao Q. Serum miR-18a: a potential marker for hepatitis B virus-related hepatocellular carcinoma screening. Dig Dis Sci. 2012;57(11):2910–2916. | ||
Zhang N, Zhang H, Liu Y, et al. SREBP1, targeted by miR-18a-5p, modulates epithelial-mesenchymal transition in breast cancer via forming a co-repressor complex with Snail and HDAC1/2. Cell Death Differ. Epub 2018 Jul 9. | ||
Yang H, Li W, Zhang Y, et al. Regulatory role of miR-18a to CCN2 by TGF-β1 signaling pathway in pulmonary injury induced by nano-SiO2. Environ Sci Pollut Res Int. 2018;25(1):867–876. | ||
Liang Q, Zhang G, Wang J, Sheng S. Diagnostic Value of MicroRNA-18a for Gastric Cancer: a Meta-Analysis. Clin Lab. 2018;64(1):177–184. | ||
Li X, Yu T, Shan H, et al. lncRNA PFAL promotes lung fibrosis through CTGF by competitively binding miR-18a. FASEB J. 2018;32(10):5285–5297. | ||
Zhu HY, Bai WD, Ye XM, Yang AG, Jia LT. Long non-coding RNA UCA1 desensitizes breast cancer cells to trastuzumab by impeding miR-18a repression of Yes-associated protein 1. Biochem Biophys Res Commun. 2018;496(4):1308–1313. | ||
Zhang Y, Qian W, Feng F, et al. Upregulated lncRNA CASC2 may inhibit malignant melanoma development through regulating miR-18a-5p/RUNX1. Oncol Res. Epub 2018 Feb 8. | ||
Zhang G, Han G, Zhang X, et al. Long non-coding RNA FENDRR reduces prostate cancer malignancy by competitively binding miR-18a-5p with RUNX1. Biomarkers. 2018;23(5):435–445. | ||
Mou T, Zhu D, Wei X, et al. Identification and interaction analysis of key genes and microRNAs in hepatocellular carcinoma by bioinformatics analysis. World J Surg Oncol. 2017;15(1):63. | ||
Bonneau B, Ando H, Kawaai K, Hirose M, Takahashi-Iwanaga H, Mikoshiba K. IRBIT controls apoptosis by interacting with the Bcl-2 homolog, Bcl2l10, and by promoting ER-mitochondria contact. Elife. 2016;5:e19896. | ||
Xu JD, Cao XX, Long ZW, et al. BCL2L10 protein regulates apoptosis/proliferation through differential pathways in gastric cancer cells. J Pathol. 2011;223(3):400–409. | ||
Bai Y, Wang J, Han J, et al. BCL2L10 inhibits growth and metastasis of hepatocellular carcinoma both in vitro and in vivo. Mol Carcinog. 2017;56(3):1137–1149. | ||
Liu L, Cai X, Liu E, Tian X, Tian C. MicroRNA-18a promotes proliferation and metastasis in hepatocellular carcinoma via targeting KLF4. Oncotarget. 2017;8(40):68263–68269. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.