Back to Journals » Journal of Pain Research » Volume 18
Microglia’s Phenotypic Heterogeneity in Pain Pathogenesis and Electroacupuncture Analgesia: Mechanisms and Therapeutic Potential
Authors Ji D, Lyu Z, Erasmus S, Wu Y ![]() , Li X, Chen X, Fang J, Du J, Fang J, Zhou J, Liang Y
, Li X, Chen X, Fang J, Du J, Fang J, Zhou J, Liang Y ![]()
Received 8 June 2025
Accepted for publication 28 October 2025
Published 11 November 2025 Volume 2025:18 Pages 6007—6022
DOI https://doi.org/10.2147/JPR.S545420
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Professor E Alfonso Romero-Sandoval
Dingru Ji,1,* Zhengyi Lyu,2,* Shannah Erasmus,3,* Yuxin Wu,1 Xingzhen Li,1 Xue Chen,1 Junfan Fang,1 Junying Du,1 Jianqiao Fang,1 Jie Zhou,4 Yi Liang1
1Key Laboratory of Acupuncture and Neurology of Zhejiang Province, Department of Neurobiology and Acupuncture Research, The Third Clinical Medical College, Zhejiang Chinese Medical University, Hangzhou, People’s Republic of China; 2Department of Acupuncture and Chinese Tuina, Sir Run Run Shaw Hospital, Zhejiang University School of Medicine, Hangzhou, People’s Republic of China; 3International Education College, Zhejiang Chinese Medical University, Hangzhou, People’s Republic of China; 4Department of Acupuncture and Moxibustion, The Third Affiliated Hospital of Zhejiang Chinese Medical University, Hangzhou, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yi Liang, Key Laboratory of Acupuncture and Neurology of Zhejiang Province, Department of Neurobiology and Acupuncture Research, The Third Clinical Medical College, Zhejiang Chinese Medical University, Hangzhou, People’s Republic of China, Tel +86 0571-86633328, Fax +86 571-86658056, Email [email protected] Jie Zhou, Department of Acupuncture and Moxibustion, The Third Affiliated Hospital of Zhejiang Chinese Medical University, Hangzhou, People’s Republic of China, Tel +8615968895450, Fax +86 0571-88393516, Email [email protected]
Abstract: Pain is a complex physiological phenomenon driven by dynamic microglial(MG) regulation in the CNS. Aberrant microglial activation mediates neuroinflammation, neuronal hyperexcitability, and synaptic plasticity, exhibiting spatiotemporal and sex-specific heterogeneity. While early pharmacological interventions show time-limited efficacy, electroacupuncture (EA) emerges as a potent non-pharmacological strategy that disrupts microglia-neuron crosstalk via multiple pathways. Critically, EA’s efficacy depends on intervention timing, with early application preventing microglial active in neuropathic models. Recent advances reveal CD11c+ microglia as dual regulators of pain resolution and relapse—clearing myelin debris acutely but driving recurrence via MMP-9-mediated synaptic remodeling. This review synthesizes microglial heterogeneity, stage-specific interventions, and EA’s translational potential in rebalancing microglial phenotypes for precision pain management.
Keywords: microglia, pain, morphology, polarization, inflammation
Introduction
Pain is recognized as the fifth vital sign in humans. Chronic pain resulting from delayed healing of acute pain significantly impacts the patients’ quality of life. According to the United States Centers for Disease Control and Prevention, 11% to 40% of the world’s population suffers from chronic pain.1 The occurrence and progression of chronic pain are closely related to microglia.2 As one of the important immune cells in the nervous system, microglia can be activated or even further polarized by exogenous stimuli, the morphology of microglia changes continuously and has different effects on the neurons. A clinical study showed that the expression of translocator protein (TSPO), a marker of glial activation, was significantly higher in the brains of chronic low back pain patients, and was negatively correlated with the expression of interleukin-6 at the thalamic level. It indicated that TSPO is involved in the analgesic and anti-inflammatory effects of patients with chronic low back pain.3,4 Experimental studies have confirmed that microglia activation and polarization play a crucial role in pain modulation. Targeted inhibition of M1 microglia or promotion of M2 microglia expression has been shown to effectively alleviate pain.5 For instance, oral low-dose naltrexone, an opioid receptor antagonist, can inhibit microglial activation through Toll-like receptor 4 (TLR4) and suppress the expression of M1-type microglia, thereby reducing the release of central pro-inflammatory factors in patients with fibromyalgia and alleviating their pain.6 Although numerous studies suggest that promoting M2 microglial polarization benefits pain reduction, the precise role of different microglial subtypes in pain modulation, and the relationship between microglial activation, polarization, and the pain process remain unclear.
The treatment of chronic pain primarily relies on pharmacological therapy, including non-steroidal anti-inflammatory drugs (NSAIDs), opioids, analgesics, antidepressants, and antiepileptic drugs. However, conventional drug treatments have certain limitations, with inevitable side effects on the digestive tract and immune system,1,5 and may even contribute to an increased incidence of chronic pain.7 The multimodal and multidisciplinary approach to pain management advocates combining pharmacological and non-pharmacological therapies, a concept that is increasingly recognized by clinicians and patients.1 Among the non-pharmacological treatments, electroacupuncture is considered an important therapeutic method from Traditional Chinese Medicine (TCM) and has been shown to provide effective pain relief in various conditions.8 Animal studies indicate that EA can down-regulate Cyclooxygenase-2 (COX2) expression in the spinal cord and inhibit microglial activation, thereby alleviating neuropathic pain induced by autologous nucleus pulposus transplantation.9 Furthermore, it is well established that EA exerts its analgesic effects by promoting the polarization of microglia towards M2 type. In the CFA-induced inflammatory pain model, EA inhibits M1 microglia polarization and promotes M2 microglial polarization by upregulating G protein-coupled receptor kinase 2 (GRK2) expression in the spinal dorsal horn.10 It is notable that early intervention targeting microglial activation is of paramount importance for effectively inhibiting pain. Once pain has developed, suppressing microglia alone fails to achieve the adequate analgesia; and at this time, intervention targeting microglial polarization should be implemented.
Early studies mainly focused on the exploration of M1 and M2 types of microglia. With the advancement of science and the deepening of research on microglia, current studies now pay more attention to the heterogeneity of microglia. Furthermore, recent advancements have disclosed the dual functions of microglial subpopulations, such as CD11c+ cells, in both pain alleviation and recurrence, emphasizing the necessity of precise, stage-specific interventions. Recent studies on human microglial cells have achieved notable advancements. Mass spectrometry-based cell analysis has revealed that human TDP-43 (hTDP-43) can promote the generation of a distinct microglial subset characterized by high CD11c expression and enhanced phagocytic capacity, a process mediated by the TREM2 signaling pathway. Furthermore, using mass spectrometry (MS) and surface plasmon resonance (SPR), direct molecular interactions between TDP-43 and TREM2 were confirmed in vitro, in vivo, and in postmortem tissue samples from patients with amyotrophic lateral sclerosis (ALS). This discovery provides novel insights into the immune regulatory mechanisms underlying human neurodegenerative disorders. This review integrates the current knowledge regarding the microglia activation mechanisms, activation heterogeneity, and related therapeutic approaches, while simultaneously exploring new strategies targeting microglial dynamics for pain management. This review elaborates on the dual role of microglial phenotypic heterogeneity in both the pathogenesis and alleviation of pain. Based on its spatiotemporal dynamics and sex-based differences, it proposes future directions for stage-specific precision therapeutic strategies, with a particular emphasis on electroacupuncture (EA) therapy.
M1-Type Microglia Activated Pain
Pain, as a complex physiological and pathological phenomenon, is closely associated with the dynamic regulation of microglia within the central nervous system. Activated microglia can be polarized into pro-inflammatory (M1) and anti-inflammatory (M2) phenotypes depending on stimulation type, microenvironment, duration of stimulation, and the balance between neurotoxic substances and neuronutrients.11 These phenotypes are closely associated with pain-related disease states. The M1 phenotype represents the classical activation pathway of microglia and is typically triggered by lipopolysaccharide (LPS), proinflammatory cytokines such as interferon- γ (IFN-γ) and tumor necrosis factor-α (TNF- α), which are secreted by TH1 cells and astrocytes, as well as by trauma-induced cell debris. Once activated, M1 microglia releases pro-inflammatory cytokines, including interleukin IL-6, IL-12, IL-18, and inducible nitric oxide synthase (iNOS). They also express surface markers such as CD86, CD14, CD16, CD32 and CD42. Activation of M1 microglia promotes cytotoxicity, acute immune response, and severe neuroinflammation.12 In contrast, M2 phenotype represents an alternative activation pathway that promotes anti-inflammatory and neuroprotective functions. In the spinal cord, Kv1.3 channels promote neuropathic pain by driving M1 microglial polarization and activating NLRP3 inflammasome.13 Morphine, a widely used opioid drug for cancer pain, presents a major clinical challenge due to morphine tolerance. Experimental studies have demonstrated that prolonged morphine administration induces microglial M1 to polarization, which in turn contributes to the development of morphine tolerance.14 Similarly, paclitaxel-induced M1 polarization of spinal microglia triggers RIP3/ MLKL-regulated necroptosis, leading peripheral neuropathic pain15 M2 microglia release anti-inflammatory cytokines (eg, IL-4, IL-10, etc)., growth factors (eg, insulin-like growth factor I, fibroblast growth factor), and neurotrophic mediators (eg, glial cell-derived neurotrophic factor, brain-derived neurotrophic factor, nerve growth factor).12,16,17 M2 microglia also express surface markers such as CD163 and CD206. Functionally, M2 microglia inhibit inflammation, promote cell proliferation and phagocytosis of cell debris, and facilitates tissue repair.12,16,17
Research indicates that the abnormal activation of MG mediates the transmission of inflammatory signals, the remodeling of neuronal excitability, and the alteration of synaptic plasticity through multiple pathways, serving as the core mechanism driving the occurrence and persistence of pain. The activation process is multidimensionally regulated by pro-inflammatory factors, damage-associated molecular patterns(DAMPs), and neurotrophic factors. Microglial activation plays a central role in pain pathogenesis, by integrating neuroinflammation, synaptic plasticity, and neuronal hyperexcitability through dynamic interactions with cytokines, DAMPs, and neurotrophic factors It also demonstrates temporal, spatial, and gender heterogeneity. Moreover, the pro-inflammatory (M1) and anti-inflammatory (M2) phenotypes of dynamic transitions at different stages of pain, further complicating intervention strategies. Intriguingly, antagonizing microglia at different time points yields distinct outcomes, and only early intervention in microglial activation may effectively suppress the occurrence of pain.
The Three Primary Factors Driving M1-Type Microglial Activation
In their resting state, microglia perform monitoring functions. Once exogenous or endogenous stimuli are detected, microglia respond rapidly by releasing inflammatory factors, chemokines, and other signaling molecules. These amplify pain signals and remodel neuronal synaptic plasticity, thereby mediating abnormal neuronal excitation. Currently, substances capable of influencing microglial activation can be categorized into three types: pro-inflammatory cytokines, damage-associated molecular patterns (DAMPs), and neurotrophic factors.
Microglial Activation Mediated by Pro-Inflammatory Cytokines and Hyperalgesia
IL-1β serves as a key driver of microglial activation in the spinal cord. Following spinal cord injury or nerve damage, IL-1β released by damaged cells binds to TLR4 receptors on the surface of microglia, activating the NF-κB signaling pathway and inducing microglia polarization towards the pro-inflammatory (M1) type. This results in the release of secondary inflammatory mediators such as TNF-α and IL-6, intensifying excitatory synaptic transmission in spinal dorsal horn neurons and ultimately leading to hyperalgesia.18,19
TNF-α sustains activation of microglia through both autocrine and paracrine loops. In the spinal dorsal horn, TNF-α upregulates the expression of Nav1.3 (a specific type of sodium channel) in peripheral nerves, thereby lowering the pain threshold.20 Additionally, TNF-α and IL-6 synergistically activate the COX2-PGI2 pathway in spinal microglia, promoting the synthesis of prostaglandin E2 (PGE2) and exacerbating neuroinflammation-related pain. In the central nervous system, TNF-α indirectly influences pain processing by regulating hippocampal synaptic plasticity in Alzheimer’s disease models, but the spinal cord remains the principal site for driving pain.21
Microglial Activation Triggered by DAMPs and Its Association with Pain
ATP, as a typical DAMP, mediates the rapid activation of microglia via the purinergic receptor P2X7R. ATP released by damaged neurons in the spinal dorsal horn activates P2X7R on microglia, inducing the opening of cation channels and the assembly of inflammasomes (such as NLRP3), promoting the cleavage and release of the IL-1β precursor, forming a positive feedback loop of “damage - ATP release - microglial activation”.22 Pain receptors on the microglial membrane, represented by P2X4R, play a crucial role in the occurrence of pain. Studies have demonstrated that microglial activation mediates the development of various types of pain,23 with their newly formed branches carrying pain receptors, inducing microglial activation in the central nervous system and causing hyperalgesia. P2X4R is a non-selective cation channel trimer activated by ATP and is predominantly expressed on the surface of microglia in the central nervous system. Research indicates that when exposed to noxious stimuli, the large amount of ATP released in the body is recognized by P2X4R on surveillance microglia, stimulating the activation of the core pain signaling pathway mediated by brain-derived neurotrophic factor (BDNF) release. This pathway exhibits an increase in intracellular chloride de-inhibition in nociceptive neurons in the spinal dorsal horn, thereby weakening the excitability of inhibitory neurons and facilitating pain transmission. The changes in signal transmission from P2X4R+ microglia to nociceptive transmission neurons may account for the main symptoms of human neuropathic pain.24 In various types of pain, including chronic inflammatory pain, peripheral neuropathic pain, post-herpetic pain, and cancer pain, upregulated expression of P2X4R in the central nervous system has been identified to induce microglial activation, and this phenomenon occurs exclusively in male animal models, while the pain of female animals is not regulated by microglia.24 S100B, as a calcium-binding protein, is secreted by astrocytes and microglia following spinal cord injury. It activates the STAT3 pathway by binding to the receptor RAGE, promoting the polarization of microglia to the M1 type and intensifying the local inflammatory response in the spinal cord.19 In the central nervous system, S100B drives the release of IL-1β and TNF-α from microglia through the PARP-1 pathway, but its regulatory effect on chronic pain may depend on the degree of primary sensitization in the spinal dorsal horn.25
Bidirectional Regulation of Neurotrophic Factors and Microglial Activation
The role of BDNF in pain is region-specific: in the spinal dorsal horn, BDNF activates the ERK-CREB pathway via the TRKB receptor, promoting microglial proliferation and enhancing their response to noxious signals, directly contributing to the formation of pain sensitization;26 while in the central nervous system, BDNF can indirectly alleviate pain associated with neurodegenerative diseases by inhibiting the excessive activation of senescence-related microglia.27 This bidirectional regulatory mechanism suggests that targeting the BDNF/TRKB pathway may emerge as a promising strategy for precise intervention in different types of pain.
M2-Type Microglia Alleviates Pain
Pharmacological Therapies Related to Microglia Alleviate Pain
Minocycline is a well-established inhibitor of microglial activation. Early studies suggested that microglia played a predominantly detrimental role in the development and maintenance of pain, prompting researchers to explore targeted inhibition of microglial activation as a potential therapeutic strategy for pain relief.28 Minocycline-mediated suppression of microglia activation has been considered an effective approach for the treatment of chronic pain.29 However, subsequent research revealed a critical limitation: minocycline’s efficacy depends on the timing of administration. It was effective only when administered before the onset of a noxious stimulus. If the painful stimulus persisted, re-injection of minocycline could fail to reverse the pain.30 In the SNI model, intrathecal minocycline administration effectively prevented pain in the early stages; however, even continuous injection during the later stages of SNI does not fully halt pain progression.30
Extensive evidence suggests that M2-type microglia facilitate pain relief. In neuropathic pain models, DKK3 inhibits microglial polarization and neuroinflammation by modulating the ASK-1/JNK/p-38 pathway. This mechanism promote M2 polarization and reduces the production pro-inflammatory factors.31 Similarly, paeonol facilitates the transformation of M1 to M2 microglia in the spinal dorsal horn via the RhoA/p38 MAPK pathway.
Clinical trials provide evidence that enhancing M2 microglial activity can effectively alleviate fibromyalgia. Low-dose oral naltrexone has been shown to reduce pain scores in patients with fibromyalgia while downregulating pro-inflammatory factors secreted by M1 microglia in plasma.32 Similarly, aquatic exercise has been found to relieve muscle fiber pain, increase IL-10 secretion from M2 microglia, contributing to an anti-inflammatory response.33 Thus, therapeutic strategies targeting microglia polarization hold promise as a novel approach for nociceptive pain management.
In the central nervous system, microglial cells can transform from an M1 pro-inflammatory state to an M2 anti-inflammatory state, releasing large amounts of anti-inflammatory factors, facilitating nerve repair, and contributing to pain relief.31 Therefore, modulating the balance of microglial polarization is crucial for effective pain management.
Mannosylated clodronate (dichloromethylene diphosphonate; Cl2MDP) can effectively deplete M2 microglia.34 Recent studies have highlighted the critical role of M2 microglia in inflammatory pain. In the mouse model of acute inflammatory pain induced by complete Freund’s adjuvant (CFA), researchers performed targeted immune intervention via intrathecal injection of mannosylated clodronate liposomes and found that this treatment significantly augmented the persistent pain phenotype in Pi16-positive mice. Specifically, via immunofluorescence co-localization analysis, it was confirmed that these mannosylated nanoparticles selectively eliminated the CD206+ anti-inflammatory macrophage subset and contributed to the persistence of inflammatory pain. These findings demonstrate that M2 microglia plays a key role in pain in pain modulation.
Non-Pharmacological Therapies for Pain Management
Currently, therapeutic strategies targeting microglial polarization for pain relief are under active investigation, with the efficacy of related drugs and interventions scientifically validated. Microglia may not only respond to various stimuli but also modulate immune homeostasis in the central nervous system through multiple pathways, contributing to central sensitization. Clinical studies have demonstrated that electroacupuncture has significant analgesic effects, however, its non-neuronal mechanism remain poorly understood.8 EA builds upon traditional acupuncture by introducing electrical stimulation to acupoints, activating afferent peripheral nerve fibers and influencing the central nervous system. To elucidate how acupuncture extends its local effects to exert systemic anti-inflammatory and analgesic effects, we examined microglial changes in different pain models. Findings indicate that EA treatment inhibits microglia activation and promotes microglial polarization (Table 1 and Figure 1).
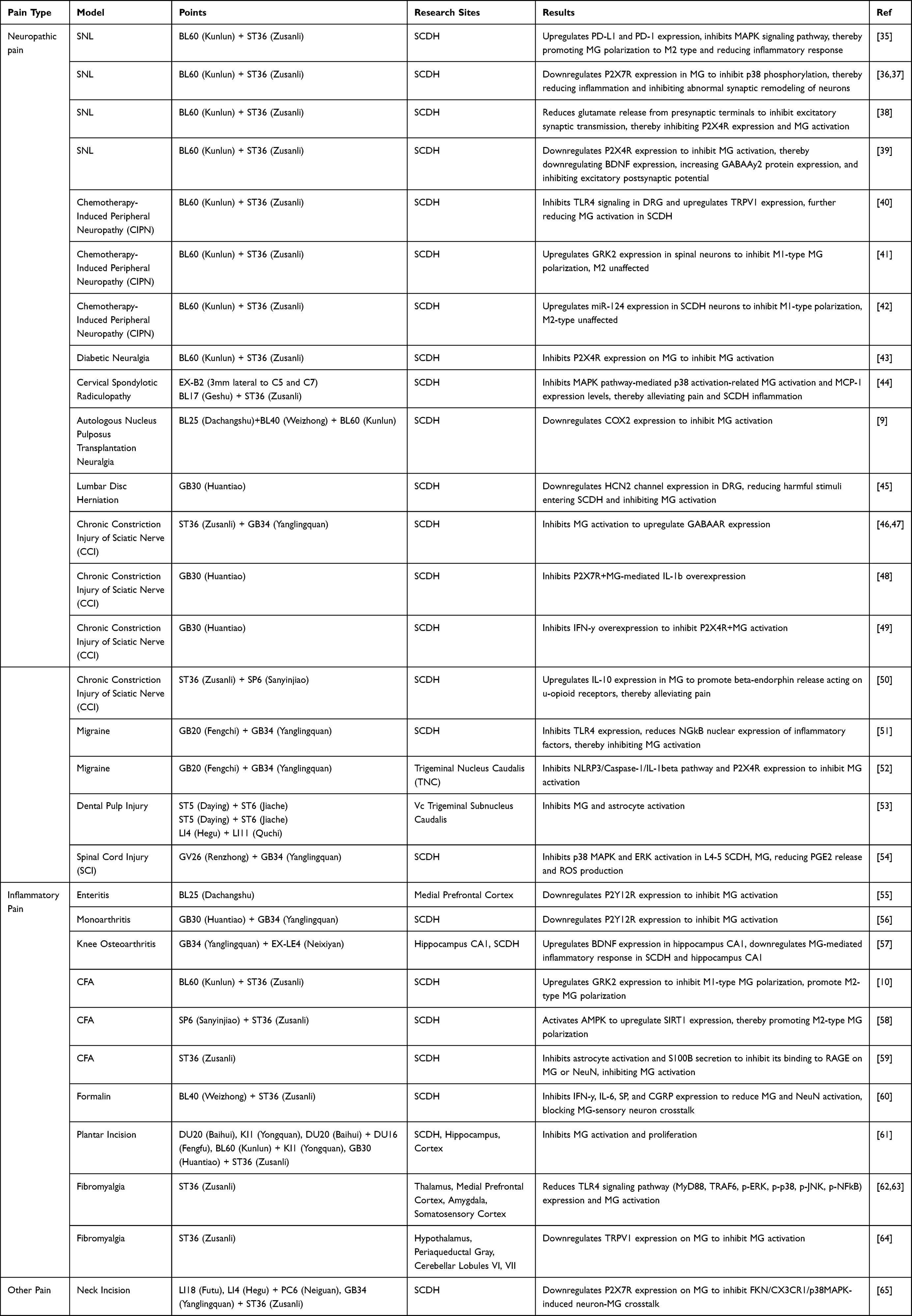 |
Table 1 Summary of EA Parameters and Microglial Modulation in Pain Models |
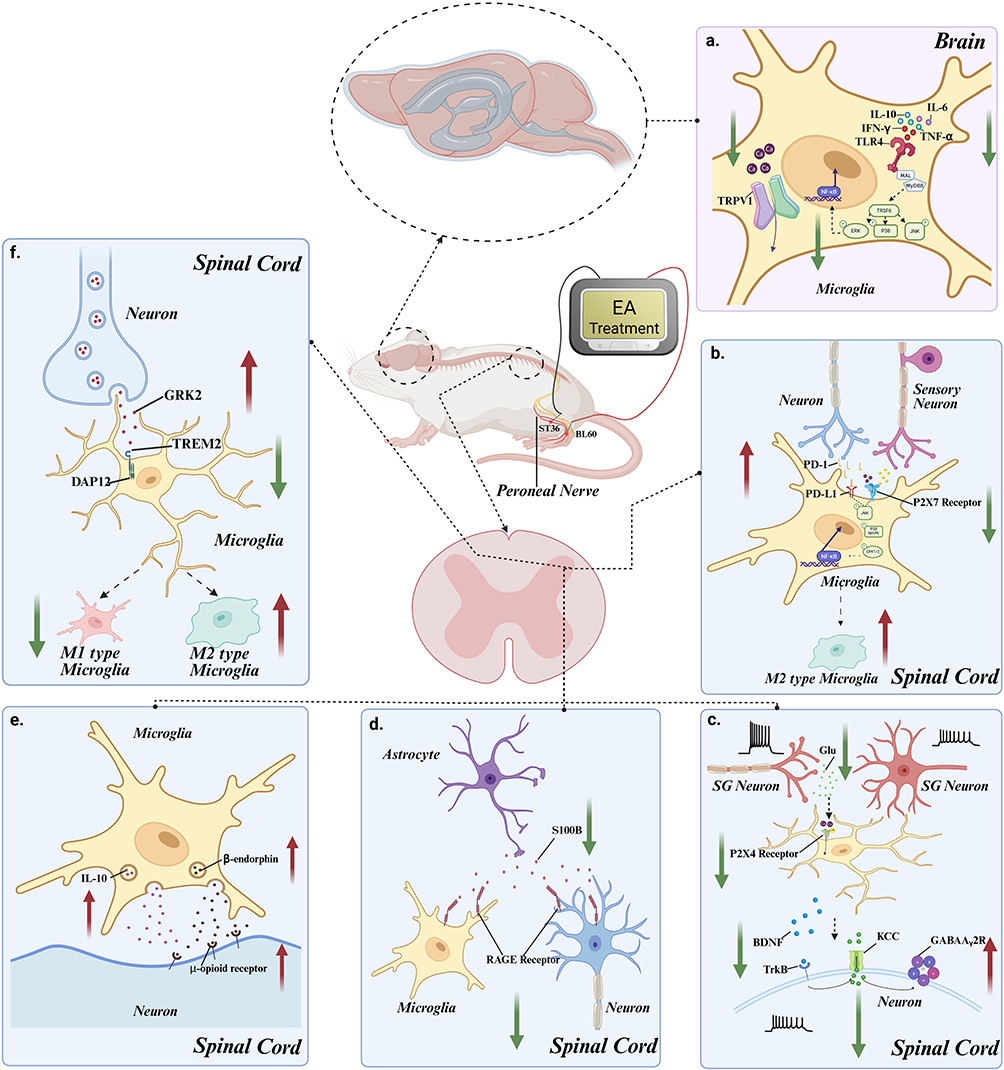 |
Figure 1 EA is involved in the analgesic mechanism through microglia. (a) Electroacupuncture reduces activation of TLR4 signaling pathway in microglia (MyD88, TRAF6, p-ERK, p-p38, p-JNK, p-NFκB) by suppressing the expression of pro-inflammatory cytokines such as IL-1β, IL-6, IFN-γ, and TNF-α in the brain; Concurrently, it inhibits microglial activation by downregulating TRPV1 ion channels activity and reducing calcium influx. (b) Electroacupuncture inhibits activation of the JNK signaling pathway (p-JNK, p38-MAPK, p-ERK1/2, NF-κB) by downregulating ATP release from adjacent sensory neurons in the spinal dorsal horn and reducing the expression of P2X7R on the microglial membrane. Additionally, it upregulates neuronal PD-1 release, thereby limiting its interaction with PD-L1 on microglia, which collectively promotes microglial polarization towards the M2 phenotype. (c) Electroacupuncture inhibits the expression of P2X4R receptors on the microglial membrane by downregulating the release of glutamate from substantia gelatinosa neuron terminals in the spinal dorsal horn his suppression reduces microglial BDNF release and subsequently activates inhibitory neurons via the TrkB signaling pathway (TrkB, KCC2, and GABAAγ2 receptors). (d) Electroacupuncture inhibits the binding of S100B, released by astrocytes, to RAGE receptors on the microglial and neuronal membranes, thereby reducing the excitability of microglia and neurons. (e) Electroacupuncture promotes the release of β-endorphin by upregulating IL-10 production in microglia, which then acts on the μ-opioid receptor on neurons to alleviate pain. (f) Electroacupuncture promotes the microglial polarization towards the M2 direction by enhancing the neuronal release of GRK2, which acts on TREM2 and DAP12 receptors on microglia. |
EA Is Involved in Analgesia by Inhibiting the Activation of Microglia
In the spared nerve injury model, EA suppresses excitatory synaptic transmission by reducing glutamate release from presynaptic terminals. This leads to downregulation of P2X4R expression in spinal cord dorsal horn, inhibition of microglial activation, and reduction of brain-derived neurotrophic factor expression. Additionally, EA treatment increases neuronal GABAA-y2 protein expression and suppresses excitatory postsynaptic potential (EPSPs).38,39 In chemotherapy-induced peripheral neuropathy (CIPN), EA inhibits Toll-like receptor 4 (TLR4) signaling in DRG while increasing transient receptor potential vanilloid 1 (TRPV1) expression, which subsequently leads to inhibition of microglia activation in SCDH.40 In diabetic peripheral neuropathic pain model, EA inhibits microglial activation by suppressing P2X4R expression on spinal microglia.43 Similarly, in a rat model of cervical spondylotic radiculopathy, EA alleviates pain and reduces SCDH inflammation by inhibiting microglial activation via the MAPK pathway-mediated p38 activation and suppressing monocyte chemoattractant protein-1 (MCP-1) expression.44 Furthermore, EA has been shown to suppress spinal microglial activation while upregulating γ-aminobutyric acid type A receptor (GABA-AR) expression, contributing to its analgesic effects.46 Additionally, EA upregulates IL-10 expression in microglia and promotes the β-endorphin release, which acts on μ-opioid receptors to alleviate neuropathic pain caused by chronic constriction injury (CCI) of the sciatic nerve.50 In complete Freund’s adjuvant -induced inflammation model, EA inhibits S100B activation in astrocytes, thereby preventing its binding to receptor for advanced glycation end products (RAGE) on microglia and NeuN-expressing neurons, ultimately suppressing microglial activation.59 In a formalin-induced inflammatory pain model, EA reduces microglia-neuron interactions and disrupts sensory neuron crosstalk by downregulating the expression of interferon-γ (IFN-γ), interleukin-6 (IL-6), substance P (SP), and calcitonin gene-related peptide (CGRP).60 Furthermore, in the formalin-induced inflammatory pain model, EA alleviates pain by downregulating of TLR4 signaling pathway components and inhibit microglial activation in the thalamus, medial prefrontal cortex, amygdala, and somatosensory cortex.62,63 Additionally, EA can downregulate transient receptor potential vanilloid 1 (TRPV1) expression on microglia in the hypothalamus, periaqueductal gray matter, and the VI and VII lobules of the cerebellum, thereby inhibiting microglial activation.64 In an incision-induced inflammatory pain model, EA suppresses microglial activation in SCDH, hippocampus and cortex, thereby alleviating pain caused by plantar incision.61 Furthermore, EA downregulates P2X7R expression on the spinal cord microglia and inhibit the fractalkine (FKN)/CX3CR1/p38MAPK signaling pathway, which mediates neuron-microglia crosstalk, thereby relieving neck incision pain.65 These findings suggest that EA inhibits the activation of microglia, which in turn modulates dialogue between microglia and neurons, contributing to its analgesic effects.
EA Regulates Microglia Polarization to M2 to Mediate Analgesia
In the spinal nerve ligation (SNL) model, EA stimulation at ST36+BL60 acupoints increases mechanical and thermal pain threshold while preserving myelin structure integrity. At the spinal cord level, EA may regulate excitability of primary afferent neurons via programmed death-ligand 1 (PD-L1), thereby inhibiting the mitogen-activated protein kinase signaling pathway, promoting the transition of activated M1 microglia to M2 microglia, reducing pro-inflammatory factor release, and ultimately achieving analgesia.36,37 Additionally, EA upregulates the expression of G protein-coupled receptor kinase 2 (GRK2) expression and miR-124 expression in spinal cord neurons, collectively inhibiting M1 polarization without affecting M2 microglia.42 In CFA-induced inflammatory pain model, EA can inhibits M1 microglia polarization and promotes M2 microglial polarization by upregulating GRK2 expression in the spinal dorsal horn.10 EA also activates AMP-activated protein kinase (AMPK) and increase sirtuin 1 (SIRT1) expression, further enhancing M2 microglial polarization.58
Electroacupuncture demonstrates considerable promise in modulating microglia and managing pain; however, its clinical translation faces several important limitations and unresolved issues. Although preclinical studies consistently indicate that EA can suppress M1 polarization of microglia, promote an M2 phenotype, and alleviate pain, there remains a relative scarcity of high-quality, large-sample randomized controlled trials (RCTs). This is particularly evident in the lack of long-term efficacy and safety evaluations across different pain types, such as neuropathic and inflammatory pain. Furthermore, significant gaps exist in the standardization of EA treatment parameters—including optimal stimulation frequency (eg, 2 Hz vs 100 Hz), waveform, session duration, and treatment course—which hinders both reproducibility and broad clinical application. More importantly, while mechanistic research on EA has largely been conducted in animal models, it remains unclear whether the same microglia-neuron interaction pathways operate in humans, underscoring the need for more direct neuroimmunological evidence.
Compared to pharmacological interventions, EA offers potential advantages such as holistic regulation, fewer side effects, and non-dependence. Pharmacological agents (eg, minocycline, P2X4R antagonists) typically have well-defined targets and clear mechanisms of action. However, they also face challenges related to blood-brain barrier penetration, systemic toxicity, and drug resistance. Notably, many drugs effective in male animal models have failed in clinical trials due to sex-specific differences or insufficient efficacy. In contrast, as a non-pharmacological intervention, EA modulates bodily functions through multiple targets and pathways. It not only influences microglial polarization but may also regulate the local inflammatory milieu, endogenous opioid systems, and autonomic nerve activity, reflecting its systemic mode of action. That said, pharmacotherapy holds advantages in dose controllability, ease of administration, and standardization, whereas the efficacy of EA is more dependent on practitioner skill and experience. Additionally, the accessibility and acceptability of EA are influenced by cultural, regional, and healthcare resource disparities.
Therefore, future research should prioritize rigorously designed clinical trials to validate the efficacy and safety of EA. Advanced techniques such as radiomics, immunohistochemistry, and molecular biology should be employed to elucidate its mechanisms of action in humans. Moreover, exploring combination strategies that integrate EA with low-dose pharmacotherapy may synergistically enhance therapeutic outcomes while reducing drug-related side effects, offering novel integrated approaches for pain management.
Heterogeneity of Microglia
Spatiotemporal Heterogeneity
Morphological changes serve as key indicators of microglial activation and reflect their functional status. In 1932, Pio del Rio-Hortega systematically introduced microglia to the scientific community and classified them according to their morphological characteristics, distinguishing between the resting state and amoeboid degeneration. Notably, the amoeboid structure is a hallmark of microglial activation.66 As the resident macrophages of the central nervous system, microglia exhibit remarkable morphological plasticity, enabling their involvement in range of pathological processes, including pain (Figure 2).
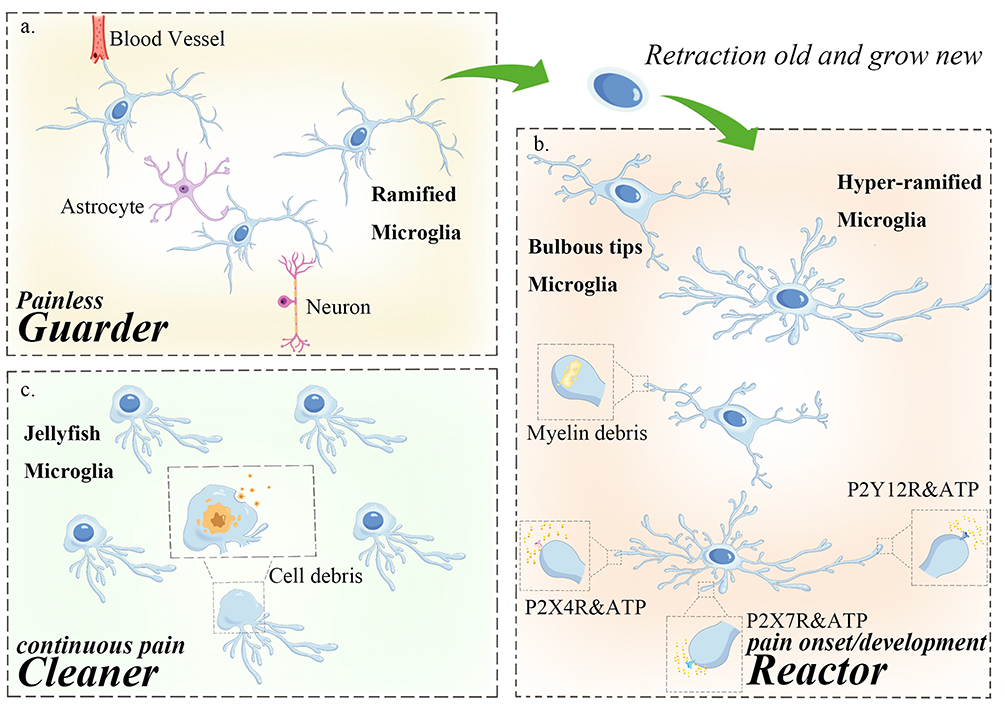 |
Figure 2 At different stages of pain, microglia undergo distinct morphological changes and assume diverse functional roles. (a) Painless state (healthy state): microglia exhibit a branched “surveillance type” morphology. Their peripheral processes continuously extend retract, while the cell body remains stationary. These slender branches interact with surrounding peripheral neurons, blood vessels, and astrocytes, enabling microglia to constantly monitor the state of the central nervous system. (b) Pain onset: microglia detect acute injury in the central nervous system and migrate to the injured site within 30–60 minutes. In response to an increase in peripheral ATP concentration, microglia rapidly retract their original branches and extend new ones with specific receptors. The terminal ends of these branches develop a bulb-like structure, allowing microglia to analyze subtle in ATP fluctuations and generate precise responses. This process leads to neurons activation, facilitating the transmission of pain signals to the brain and the perception of pain. Pain development: some microglia branch terminals engulf small myelin debris and apoptotic cells, forming a spheroid-chain-like structure. Meanwhile, other microglia extend additional branches, continuously monitoring changes in the surrounding environment to facilitate timely manner. (c) Pain maintenance: prolonged pain exposure leads microglia to phagocytosed large amounts of cellular debris, resulting in an enlarged cell body and shortened branches, resembling a jellyfish-like structure. At this stage, microglia primarily function as scavengers. |
The activation of microglia in the spinal dorsal horn is characterized by an “immediate response to injury”, relying on rapid signaling pathways such as ATP-P2X4R and IL-1β-TLR4, which directly participate in the primary processing of pain signals; while the activation of central microglia (eg, in the brain) is more closely related to chronic neuroinflammation, indirectly influencing pain perception by regulating the descending analgesic pathway or emotion-related brain regions.18,20 Notably, factors such as TNF-α and IL-1β released by spinal microglia can be transported to the brain via the circulatory system or retrograde axonal transport, activating central immune cells and forming a “spinal cord - brain” inflammatory loop, amplifying the central processing of pain signals.20
The role of microglial activation in driving the occurrence and maintenance of pain has been demonstrated in multiple models. Microglia are present throughout all stages of pain, however, distinct microglial subtypes are expressed at different stages, highlighting their heterogeneity, which has become a focus of attention in recent years. A 2022 study67 utilized single-cell RNA sequencing (scRNA-seq) technology to systematically reveal the spatiotemporal heterogeneity of microglia in the spinal cords of mice after peripheral Spared Nerve Injury (SNI). Notably, this study was the first to link the ApoE gene to the sex-specific modulation of chronic pain. In the SNI model, during the acute pain phase (3–7 days), microglia rapidly transition from a homeostatic phenotype (Cluster 1–6) to an inflammatory phenotype. The increase in Cluster 9 in males marks the peak of acute inflammation. Recent evidence has demonstrated that the central renin-angiotensin system (RAS) participates in oxidative stress and neuroinflammation. The angiotensin-converting enzyme/angiotensin II/angiotensin receptor-1 (ACE/Ang II/AT1R) axis enhances inflammation by inducing excessive activation of NOX2 in microglia and augmenting the production of reactive oxygen species (ROS).68 By 14 days post-SNI, the expression of inflammatory genes declines, and metabolic pathways (such as mitochondrial ATP synthesis and lipid metabolism) predominate. At this juncture, ApoE emerges as the most significantly upregulated gene in microglia of both male and female mice, and its protein level persistently elevates in microglia of the spinal dorsal horn. Apolipoprotein E (ApoE) can mediate the transportation of cholesterol and triglycerides, thereby maintaining lipid homeostasis in the central nervous system. Simultaneously, it plays a vital role in the nervous system by participating in the formation and repair of synapses and regulating the phenotype of microglia, thereby influencing neuroimmune responses. In Alzheimer’s disease related studies, it has been unequivocally discovered that ApoE is closely associated with microglia. The TREM2-APOE pathway constitutes the primary regulatory factor of microglial functional phenotypes in neurodegenerative diseases and serves as a potential new target for restoring the homeostasis of microglia.69,70
Gender Heterogeneity
In addition to spatiotemporal heterogeneity, microglia also possess sex-based heterogeneity. ScRNA-seq analysis revealed that microglia in the mouse spinal cord can be divided into 11 distinct subpopulations, among which Cluster9 characterized by high expresses inflammation-related genes (such as IL-1β, TNF-α) and interferon signaling pathways. This cluster of genes was significantly upregulated in male mice on day 3 after SNI, while no significant expression differences were observed in female and male mice on day 14 and 5 months. These findings suggest that the transcriptional profile of this subpopulation of cells supports phenotypic conversion through enhanced protein synthesis capacity, playing a key role in acute pain sensitization after nerve injury in male mice. At this time, the subtypes of microglia in female mice were primarily associated with metabolic and endocytic processes. Microglial proliferation is one hallmark of their activation. Although both male and female mice exhibited microglial proliferation in the spinal cord after SNI modeling, the proliferation rate of males in the acute phase was significantly higher than that of females (68.8% for males vs 46.6% for females), However, no differences were found in the total number of microglia between male and female mice. Concurrently, studies have shown that intrathecal injection of minocycline only reverses neuropathic pain in male mice and has no therapeutic effect in female mice.71 It is hypothesized that the comparable total number of microglia in the spinal cord of female mice after SNI may be attributed to the reduced apoptosis of microglia in female mice.
The Subtype of CD11c As a Heterogeneity Phenotypic Is Involved in Pain Recurrence
Recent studies no longer limit the classification of microglia to the traditional M1 and M2 types but instead detail them into more distinctive subtypes based on their different expressed factors. These subtypes exhibit varying expressions at different disease stages and even show opposite effects, highlighting the necessity of precise regulation based on specific microglial subtypes. CD11c, as a newly discovered subtype, is receiving extensive attention from the field of pain research. The latest research72 has revealed a spatiotemporal-specific regulatory mechanism of the CD11c+ microglial cell subset in the resolution and recurrence of neuropathic pain. In the peripheral nerve injury (PNI) model, mechanical hyperalgesia in the mouse’s foot persisted for at least 27 days, while the number of CD11c+ microglial cells in the spinal dorsal horn (SDH) remained continuously elevated for up to35 days post-injury, showing a “lagged activation” characteristic. This subset gradually expanded after the onset of pain hypersensitivity (approximately 7 days post-PNI) and remained resident long-term even after the recovery of the pain threshold (28 days after PNI). Temporal clearance experiments using genetic editing techniques demonstrated that clearing spinal CD11c+ microglial cells on day 7 post-PNI had no significant effect on pain recovery (mechanical withdrawal threshold changed by less than 10%); however, clearing this subset during the pain resolution period (day 14 post-PNI), prolonged pain hypersensitivity by 1.8 times (p < 0.001). More notably, clearing CD11c+ cells after complete pain recovery (day 35 post-PNI) re-induced pain hypersensitivity (recurrence rate >75%), and effect associated with complement C1q/C3 cascade activation and abnormal neuronal synaptic pruning.
Mechanistic studies have shown that CD11c+ microglia clear myelin debris from the axonal terminals of Aβ fibers in the spinal dorsal horn via the phagolysosomal system (electron microscopy revealed a 3.2-fold increase in the density of myelin remnants within phagocytic vesicles). Concurrently, these microglia secrete anti-inflammatory cytokines (such as IL-10 and TGF-β) and synaptic protective molecules (such as SPARC). However, their dual roles are stage-dependent in pathology: in the early phase, they promote repair by clearing nerve debris, but prolonged retention may lead to the release matrix metalloproteinase 9 (MMP-9) to degrade the extracellular matrix, exposing cryptic pain-related antigenic epitopes (such as NGF-TrkA). Single-cell sequencing further revealed that during the relapse phase, CD11c+ cells highly express genes associated with phagocytosis (Trem2, Ctsd) and pain sensitization factors (Cxcl1, Bdnf), and form a positive feedback loop with the CX3CR1-CX3CL1 axis of C fibers terminals.
Unlike neuropathic pain, in the CFA-induced inflammatory pain model, the CD11c+ microglial cells are only transiently activated, peaking at 3 days after CFA. Eliminating of this subpopulation does not induce pain recurrence, suggesting that their function is influenced by the local microenvironment. Following nerve injury, the ongoing Wallerian degeneration releases substantial amounts of myelin lipids, which activates microglia to differentiate into the CD11c+ phenotype through TLR2/4 signaling. In contrast, the inflammatory pain model lacks such chronic injury signals. This study was the first to propose the concept of “microglial memory”, that the CD11c+ subpopulation retains its dual roles of promoting both repair and recurrence through epigenetic remodeling. This provides a theoretical basis for precise intervention in pain recurrence: targeting their spatiotemporal-specific secretome or blocking the myelin fragment-sensing pathway may offer promising strategies for preventing chronic pain relapse. However, recent studies have identified the presence of microglia in the peripheral nervous system.73 The aforementioned article only investigated the involvement of CD11c⁺ microglia within the central nervous system in pain. Interventions targeting the central nervous system remain challenging in clinical practice, particularly with respect to monitoring the expression of CD11c-related genes in the CNS. The key finding that CD11c is involved in pain recurrence suggests its potential as a therapeutic target for preventing the transition from acute to chronic pain. How to translate this meaningful discovery into clinical research represents a major challenge for the future.
Microglial activation plays a central role in pain pathogenesis, by integrating neuroinflammation, synaptic plasticity, and neuronal hyperexcitability through dynamic interactions with cytokines, DAMPs, and neurotrophic factors. The spatiotemporal and sex-based heterogeneity of microglia—evidenced by male-biased inflammatory phenotypes in acute pain and female-dominated metabolic adaptations—highlights the complexity of their functions. Therapeutic strategies must take this heterogeneity into account. Early interventions effectively suppress activation, while later stages require approaches that modulate microglial polarization or target specific microenvironmental contexts. Non-pharmacological therapies such as electroacupuncture have shown broad efficacy by disrupting microglia-neuron crosstalk and promoting reparative microglial phenotypes.
Conclusion
This review has systematically elucidated the multifaceted role of microglia in pain pathogenesis, highlighting their central function as both mediators and modulators of pain signaling. As key immune sentinels of the CNS, microglia undergo dynamic activation in response to pro-inflammatory cytokines, DAMPs, and neurotrophic factors, driving neuroinflammation, synaptic remodeling, and neuronal hyperexcitability through pathways such as TLR4/NF-κB, P2X4R/BDNF, and COX2-PGI2. This activation is not uniform but exhibits spatiotemporal and sex-based heterogeneity.
Non-pharmacological interventions, particularly electroacupuncture, demonstrate broad-spectrum efficacy by targeting microglia-neuron crosstalk. EA alleviates pain through multiple mechanisms, such as, suppressing spinal P2X4R/TLR4/MAPK signaling, enhancing GABAergic inhibition, promoting M2 polarization via AMPK/SIRT1, and disrupting neuroinflammatory loops. However, challenges remain in optimizing EA parameters to prevent the transition from acute to chronic pain.
A groundbreaking discovery lies in the dual role of the CD11c+ microglial subset. While they contribute to the resolution of acute neuropathic pain through phagocytic clearance of myelin debris and anti-inflammatory secretion, their spatiotemporal-specific behavior, persisting long after pain resolution, suggests that therapeutic strategies must balance their reparative and maladaptive roles. For example, timed blockade of CD11c+ secretome or disruption of myelin fragment sensing could prevent relapses without impairing acute repair.
The role of microglia in pain regulation exhibits considerable complexity and apparent paradox, precluding a simple binary classification as either “pro-nociceptive” or “analgesic”. Instead, their functions demonstrate marked context-dependency and spatiotemporal dynamics. Numerous studies have shown that activated microglia in neuropathic and inflammatory pain states contribute to the maintenance of hyperalgesia by releasing mediators such as IL-1β, TNF-α, and brain-derived neurotrophic factor (BDNF), which promote central sensitization. Conversely, evidence also indicates that specific subpopulations—such as CD206⁺ M2-type or CD11c⁺ cells—can exert neuroprotective and reparative effects during pain resolution phases through the secretion of anti-inflammatory factors like IL-10 and TGF-β, as well as via phagocytosis of apoptotic debris. This functional duality underscores the plasticity of microglial states and reflects their capacity to play divergent roles across different stages of pain.
Further research has revealed significant species and sex differences, posing important challenges for the translation of preclinical findings to clinical applications. For instance, microglia-mediated mechanisms—exemplified by the P2X4R–BDNF–KCC2 signaling axis—are predominant in male mice, whereas pain behaviors in female mice may be more dependent on T lymphocytes. This discovery has profound implications for the design of clinical trials and patient stratification. Moreover, the classical M1/M2 polarization paradigm proves overly simplistic in characterizing microglial phenotypes. Single-cell transcriptomic studies have identified multiple activation states, including proliferative, interferon-responsive, and disease-associated microglia (DAM)-like phenotypes, indicating that their responsive repertoire extends far beyond traditional dichotomous classifications.
Of particular interest is the dual role of the CD11c⁺ microglial subset in pain progression and resolution. On one hand, it contributes to pain relief through the clearance of myelin debris and secretion of anti-inflammatory factors; on the other hand, its prolonged persistence may lead to aberrant synaptic pruning via the release of matrix metalloproteinase-9, ultimately resulting in pain recurrence. Additionally, the phagocytic function of microglia also exhibits dual effects: moderate synaptic pruning is essential for maintaining neural circuit function, whereas excessive phagocytosis may disrupt synaptic integrity, promoting pain chronicity and concomitant cognitive impairment. In summary, microglia play a dynamic, multifaceted, and highly heterogeneous role in pain. Future research should focus on employing high-resolution techniques—such as spatiotemporal transcriptomics, single-cell sequencing, and in vivo imaging—to further decipher functional subpopulations and establish precise phenotype–function relationships across sex, species, and disease stages. These efforts will provide a transformative perspective for developing microglia-targeted pain therapeutic strategies with genuine clinical translatability.
Utilizing the heterogeneity of microglia for precise pain management. The spatial, temporal, and gender-specific heterogeneity of microglia determines the optimal timing and location for intervention in the pain generation mechanism. Our comprehensive analysis has identified three key therapeutic windows for clinical translation: 1. Activation window (0–72 hours post-injury): Minocycline becomes ineffective in alleviating pain if intervention is delayed beyond 72 hours.28 In contrast, electroacupuncture can reduce the upregulation of P2X4R by 68% within 48 hours.46,60 Therefore, early blockade of the ATP/IL-1β signaling pathway and suppression of microglial activation are recommended during this phase. 2. Polarization window (3–14 days post-injury): During this stage, activated microglia display an imbalance in their polarization states. Intervention strategies should focus on restoring microglial phenotypic homeostasis. While minocycline, as a general microglial inhibitor, fails to alleviate pain during this window, electroacupuncture—functioning as an alternative therapeutic approach—can modulate microglial polarization and produce analgesic effects. 3. Memory window (beyond 14 days): CD11c+ microglia exhibit dual roles in both tissue repair and pain recurrence. Although patients may no longer report subjective pain at this stage, CD11c+ microglia remain highly prevalent. Therefore, sustained maintenance therapy should be emphasized in clinical practice to prevent relapse.
In conclusion, microglia represent a double-edged sword in pain biology, capable of orchestrating both resolution and chronicity. Harnessing the dual nature of microglial plasticity, balancing their protective vigilance and pathological exuberance, may unlock precise therapies that transform chronic pain from an enduring burden into a reversible chapter of human resilience.
Data Sharing Statement
The datasets generated/analysed during the current study are available from the corresponding authors.
Consent for Publication
Consent for publication was obtained from the participants.
Acknowledgments
The authors acknowledge all colleagues at the Key Laboratory of Acupuncture and Neurology of Zhejiang Province, Department of Neurobiology and Acupuncture Research, the Third Clinical Medical College, Zhejiang Chinese Medical University, for their valuable discussions on this topic.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work received financial support from the National Key R&D Program of China (2024YFC3505200 and 2024YFC3505204) and the Zhejiang Provincial Natural Science Foundation of China (Grant No. LY23H270007 from JZ).
Disclosure
The authors declare that they have no conflicts in interest in this work.
References
1. Cohen SP, Vase L, Hooten WM. Chronic pain: an update on burden, best practices, and new advances. Lancet Lond Engl. 2021;397(10289):2082–2097. doi:10.1016/S0140-6736(21)00393-7
2. Atta AA, Ibrahim WW, Mohamed AF, Abdelkader NF. Microglia polarization in nociplastic pain: mechanisms and perspectives. Inflammopharmacology. 2023;31(3):1053–1067. doi:10.1007/s10787-023-01216-x
3. Ml L, Db C, A O, et al. Evidence for brain glial activation in chronic pain patients. Brain J Neurol. 2015;138(Pt 3). doi:10.1093/brain/awu377
4. Albrecht DS, Normandin MD, Shcherbinin S, et al. Pseudoreference regions for glial imaging with 11C-PBR28: investigation in 2 clinical cohorts. J Nucl Med off Publ Soc Nucl Med. 2018;59(1):107–114. doi:10.2967/jnumed.116.178335
5. Singh KP, Verma N. Teratogenic potential of third-generation antiepileptic drugs: current status and research needs. Pharmacol Rep PR. 2019;71(3):491–502. doi:10.1016/j.pharep.2019.01.011
6. Hummig W, Baggio DF, Lopes RV, Dos Santos SMD, Ferreira LEN, Chichorro JG. Antinociceptive effect of ultra-low dose naltrexone in a pre-clinical model of postoperative orofacial pain. Brain Res. 2023;1798:148154. doi:10.1016/j.brainres.2022.148154
7. Parisien M, Lima LV, Dagostino C, et al. Acute inflammatory response via neutrophil activation protects against the development of chronic pain. Sci Transl Med. 2022;14(644):eabj9954. doi:10.1126/scitranslmed.abj9954
8. Zhang R, Lao L, Ren K, Berman BM. Mechanisms of acupuncture-electroacupuncture on persistent pain. Anesthesiology. 2014;120(2):482–503. doi:10.1097/ALN.0000000000000101
9. Ji LL, Guo MW, Ren XJ, Ge DY, Li GM, Tu Y. Effects of electroacupuncture intervention on expression of cyclooxygenase 2 and microglia in spinal cord in rat model of neuropathic pain. Chin J Integr Med. 2017;23(10):786–792. doi:10.1007/s11655-016-2606-y
10. Chen Y, Zhou Y, Li XC, et al. Neuronal GRK2 regulates microglial activation and contributes to electroacupuncture analgesia on inflammatory pain in mice. Biol Res. 2022;55(1):5. doi:10.1186/s40659-022-00374-6
11. Du L, Zhang Y, Chen Y, Zhu J, Yang Y, Zhang HL. Role of microglia in neurological disorders and their potentials as a therapeutic target. Mol Neurobiol. 2017;54(10):7567–7584. doi:10.1007/s12035-016-0245-0
12. Munder M. Arginase: an emerging key player in the mammalian immune system. Br J Pharmacol. 2009;158(3):638–651. doi:10.1111/j.1476-5381.2009.00291.x
13. Spinal voltage-gated potassium channel Kv1.3 contributes to neuropathic pain via the promotion of microglial M1 polarization and activation of the NLRP3 inflammasome - PubMed. Available from: https://pubmed.ncbi.nlm.nih.gov/36440534/.
14. Tu H, Chu H, Guan S, et al. The role of the M1/M2 microglia in the process from cancer pain to morphine tolerance. Tissue Cell. 2021;68:101438. doi:10.1016/j.tice.2020.101438
15. Dong Yang M, Ming Jie W, Hui Zhou L, et al. Spinal microglial M1 polarization contributes paclitaxel-induced neuropathic pain by triggering cells necroptosis. J Biochem Mol Toxicol. 2024;38(3):e23669. doi:10.1002/jbt.23669
16. Wang LX, Zhang SX, Wu HJ, Rong XL, Guo J. M2b macrophage polarization and its roles in diseases. J Leukoc Biol. 2019;106(2):345–358. doi:10.1002/JLB.3RU1018-378RR
17. Jurga AM, Paleczna M, Kuter KZ. Overview of general and discriminating markers of differential microglia phenotypes. Front Cell Neurosci. 2020;14:198. doi:10.3389/fncel.2020.00198
18. Sato A, Ohtaki H, Tsumuraya T, et al. Interleukin-1 participates in the classical and alternative activation of microglia/macrophages after spinal cord injury. J Neuroinflammation. 2012;9(1):65. doi:10.1186/1742-2094-9-65
19. Duan K, Liu S, Yi Z, et al. S100-β aggravates spinal cord injury via activation of M1 macrophage phenotype. J Musculoskelet Neuronal Interact. 2021;21(3):401–412.
20. Liu Y, Zhou LJ, Wang J, et al. TNF-α differentially regulates synaptic plasticity in the hippocampus and spinal cord by microglia-dependent mechanisms after peripheral nerve injury. J Neurosci off J Soc Neurosci. 2017;37(4):871–881. doi:10.1523/JNEUROSCI.2235-16.2016
21. Kuno R, Wang J, Kawanokuchi J, Takeuchi H, Mizuno T, Suzumura A. Autocrine activation of microglia by tumor necrosis factor-alpha. J Neuroimmunol. 2005;162(1–2):89–96. doi:10.1016/j.jneuroim.2005.01.015
22. Walz W, Ilschner S, Ohlemeyer C, Banati R, Kettenmann H. Extracellular ATP activates a cation conductance and a K+ conductance in cultured microglial cells from mouse brain. J Neurosci off J Soc Neurosci. 1993;13(10):4403–4411. doi:10.1523/JNEUROSCI.13-10-04403.1993
23. Halievski K, Ghazisaeidi S, Salter MW. Sex-dependent mechanisms of chronic pain: a focus on microglia and P2X4R. J Pharmacol Exp Ther. 2020;375(1):202–209. doi:10.1124/jpet.120.265017
24. Beggs S, Trang T, Salter MW. P2X4R+ microglia drive neuropathic pain. Nat Neurosci. 2012;15(8):1068–1073. doi:10.1038/nn.3155
25. Xu J, Wang H, Won SJ, Basu J, Kapfhamer D, Swanson RA. Microglial activation induced by the alarmin S100B is regulated by poly(ADP-ribose) polymerase-1. Glia. 2016;64(11):1869–1878. doi:10.1002/glia.23026
26. Activation of microglial cells triggers a release of brain-derived neurotrophic factor (BDNF) inducing their proliferation in an adenosine A2A receptor-dependent manner: A2A receptor blockade prevents BDNF release and proliferation of microglia - PubMed. Available from: https://pubmed.ncbi.nlm.nih.gov/23363775/.
27. Wu SY, Pan BS, Tsai SF, et al. BDNF reverses aging-related microglial activation. J Neuroinflammation. 2020;17(1):210. doi:10.1186/s12974-020-01887-1
28. Chen LQ, Lv XJ, Guo QH, et al. Asymmetric activation of microglia in the hippocampus drives anxiodepressive consequences of trigeminal neuralgia in rodents. Br J Pharmacol. 2023;180(8):1090–1113. doi:10.1111/bph.15994
29. Zhou YQ, Liu DQ, Chen SP, et al. Minocycline as a promising therapeutic strategy for chronic pain. Pharmacol Res. 2018;134:305–310. doi:10.1016/j.phrs.2018.07.002
30. Activated microglia contribute to convergent nociceptive inputs to spinal dorsal horn neurons and the development of neuropathic pain - PubMed. Available from: https://pubmed.ncbi.nlm.nih.gov/25784443/.
31. Zhang LQ, Gao SJ, Sun J, et al. DKK3 ameliorates neuropathic pain via inhibiting ASK-1/JNK/p-38-mediated microglia polarization and neuroinflammation. J Neuroinflammation. 2022;19(1):129. doi:10.1186/s12974-022-02495-x
32. Parkitny L, Younger J. Reduced pro-inflammatory cytokines after eight weeks of low-dose naltrexone for fibromyalgia. Biomedicines. 2017;5(2):16. doi:10.3390/biomedicines5020016
33. Ortega E, Bote ME, Giraldo E, García JJ. Aquatic exercise improves the monocyte pro- and anti-inflammatory cytokine production balance in fibromyalgia patients. Scand J Med Sci Sports. 2012;22(1):104–112. doi:10.1111/j.1600-0838.2010.01132.x
34. Miron VE, Boyd A, Zhao JW, et al. M2 microglia and macrophages drive oligodendrocyte differentiation during CNS remyelination. Nat Neurosci. 2013;16(9):1211–1218. doi:10.1038/nn.3469
35. Wu Q, Zheng Y, Yu J, et al. Electroacupuncture alleviates neuropathic pain caused by SNL by promoting M2 microglia polarization through PD-L1. Int Immunopharmacol. 2023;123:110764. doi:10.1016/j.intimp.2023.110764
36. Liang Y, Du JY, Qiu YJ, Fang JF, Liu J, Fang JQ. Electroacupuncture attenuates spinal nerve ligation-induced microglial activation mediated by p38 mitogen-activated protein kinase. Chin J Integr Med. 2016;22(9):704–713. doi:10.1007/s11655-015-2045-1
37. Wu Q, Yue J, Lin L, et al. Electroacupuncture may alleviate neuropathic pain via suppressing P2X7R expression. Mol Pain. 2021;17:1744806921997654. doi:10.1177/1744806921997654
38. Zheng Y, Jia C, Jiang X, et al. Electroacupuncture effects on the P2X4R pathway in microglia regulating the excitability of neurons in the substantia gelatinosa region of rats with spinal nerve ligation. Mol Med Rep. 2021;23(3):175. doi:10.3892/mmr.2020.11814
39. Zheng Y, Zhou Y, Wu Q, et al. Effect of electroacupuncture on the expression of P2 × 4, GABAA γ 2 and long-term potentiation in spinal cord of rats with neuropathic pain. Brain Res Bull. 2020;162:1–10. doi:10.1016/j.brainresbull.2020.04.020
40. Li Y, Yin C, Li X, et al. Electroacupuncture alleviates paclitaxel-induced peripheral neuropathic pain in rats via suppressing TLR4 signaling and TRPV1 upregulation in sensory neurons. Int J Mol Sci. 2019;20(23):5917. doi:10.3390/ijms20235917
41. Ma X, Chen Y, Li XC, et al. Spinal neuronal GRK2 contributes to preventive effect by electroacupuncture on cisplatin-induced peripheral neuropathy in mice. Anesth Analg. 2022;134(1):204–215. doi:10.1213/ANE.0000000000005768
42. Li XC, Chen H, Chen Y, et al. Spinal neuronal miR-124 inhibits microglial activation and contributes to preventive effect of electroacupuncture on chemotherapy-induced peripheral neuropathy in mice. J Immunol. 2023;212(3):410–420. doi:10.4049/jimmunol.2300539
43. Qu S, Wang H, Hu Q, et al. Electroacupuncture may alleviate diabetic neuropathic pain by inhibiting the microglia P2X4R and neuroinflammation. Purinergic Sig. 2023:1–5. doi:10.1007/s11302-023-09972-9
44. Shi T, Liu Y, Ji B, et al. Acupuncture relieves cervical spondylosis radiculopathy by regulating spinal microglia activation through MAPK signaling pathway in rats. J Pain Res. 2023;16:3945–3960. doi:10.2147/JPR.S419927
45. Qin Q, Chen Z, Fan W, Li J, Liao L, Li Y. Hyperpolarization-activated cyclic nucleotide-gated 2 contributes to electroacupuncture analgesia on lumbar disc herniation-induced radicular pain through activation of microglia in spinal dorsal horn. J Tradit Chin Med. 2022;42(3):372–378. doi:10.19852/j.cnki.jtcm.2022.03.004
46. Li S, Jiang X, Wu Q, et al. Electroacupuncture suppresses CCI-induced neuropathic pain through GABAA receptors. Evid-Based Compl Altern Med ECAM. 2022;2022:4505934. doi:10.1155/2022/4505934
47. Tu WZ, Li SS, Jiang X, et al. Effect of electro-acupuncture on the BDNF-TrkB pathway in the spinal cord of CCI rats. Int J Mol Med. 2018;41(6):3307–3315. doi:10.3892/ijmm.2018.3563
48. Xu J, Chen XM, Zheng BJ, Wang XR. Electroacupuncture relieves nerve injury-induced pain hypersensitivity via the inhibition of spinal P2X7 receptor-positive microglia. Anesth Analg. 2016;122(3):882–892. doi:10.1213/ANE.0000000000001097
49. Chen XM, Xu J, Song JG, Zheng BJ, Wang XR. Electroacupuncture inhibits excessive interferon-γ evoked up-regulation of P2X4 receptor in spinal microglia in a CCI rat model for neuropathic pain. Br J Anaesth. 2015;114(1):150–157. doi:10.1093/bja/aeu199
50. Ali U, Apryani E, Wu HY, Mao XF, Liu H, Wang YX. Low frequency electroacupuncture alleviates neuropathic pain by activation of spinal microglial IL-10/β-endorphin pathway. Biomed Pharmacother. 2020;125:109898. doi:10.1016/j.biopha.2020.109898
51. Zhou M, Pang F, Liao D, He X, Yang Y, Tang C. Electroacupuncture at Fengchi(GB20) and Yanglingquan(GB34) ameliorates paralgesia through microglia-mediated neuroinflammation in a rat model of migraine. Brain Sci. 2023;13(4):541. doi:10.3390/brainsci13040541
52. Zhou M, Pang F, Liao D, et al. Electroacupuncture improves allodynia and central sensitization via modulation of microglial activation associated P2X4R and inflammation in a rat model of migraine. Mol Pain. 2024;20:17448069241258113. doi:10.1177/17448069241258113
53. Ballon Romero SS, Lee YC, Fuh LJ, Chung HY, Hung SY, Chen YH. Analgesic and neuroprotective effects of electroacupuncture in a dental pulp injury model-a basic research. Int J Mol Sci. 2020;21(7):2628. doi:10.3390/ijms21072628
54. Choi DC, Lee JY, Lim EJ, Baik HH, Oh TH, Yune TY. Inhibition of ROS-induced p38MAPK and ERK activation in microglia by acupuncture relieves neuropathic pain after spinal cord injury in rats. Exp Neurol. 2012;236(2):268–282. doi:10.1016/j.expneurol.2012.05.014
55. Li Y, Zhang H, Yang J, et al. P2Y12 receptor as a new target for electroacupuncture relieving comorbidity of visceral pain and depression of inflammatory bowel disease. Chin Med. 2021;16(1):139. doi:10.1186/s13020-021-00553-9
56. Wang J, Song W, Zhang Y, et al. Electroacupuncture alleviates pain by suppressing P2Y12R-dependent microglial activation in monoarthritic rats. Neurochem Res. 2024;49(5):1268–1277. doi:10.1007/s11064-024-04114-y
57. Jie Z, Feng Y, Pu-wei Y, et al. Effect of electroacupuncture on the expression of Iba-1 in spinal dorsal horn and hippocampus of chronic pain rats with knee osteoarthritis. Acupunct Res. 2023;48(5):431–437. doi:10.13702/j.1000-0607.20211398
58. Nan FB, Gu YX, Wang JL, Chen SD. Electroacupuncture promotes macrophage/microglial M2 polarization and suppresses inflammatory pain through AMPK. NeuroReport. 2024;35(6):343. doi:10.1097/WNR.0000000000002005
59. Liao HY, Hsieh CL, Huang CP, Lin YW. Electroacupuncture attenuates CFA-induced inflammatory pain by suppressing Nav1.8 through S100B, TRPV1, opioid, and adenosine pathways in mice. Sci Rep. 2017;7(1):42531. doi:10.1038/srep42531
60. Liu Q, Liu Y, Bian J, Li Q, Zhang Y. The preemptive analgesia of pre-electroacupuncture in rats with formalin-induced acute inflammatory pain. Mol Pain. 2019;15:1744806919866529. doi:10.1177/1744806919866529
61. Yang L, Ding W, Dong Y, et al. Electroacupuncture attenuates surgical pain-induced delirium-like behavior in mice via remodeling gut microbiota and dendritic spine. Front Immunol. 2022;13:955581. doi:10.3389/fimmu.2022.955581
62. Tsai ST, Yang CC, Liao HY, Lin YW. Electroacupuncture reduces fibromyalgia pain via neuronal/microglial inactivation and toll-like receptor 4 in the mouse brain: precise interpretation of chemogenetics. Biomedicines. 2024;12(2):387. doi:10.3390/biomedicines12020387
63. Electroacupuncture attenuates fibromyalgia pain via toll-like receptor 4 in the mouse brain - PubMed. Available from: https://pubmed.ncbi.nlm.nih.gov/37240805/.
64. Liao HY, Lin YW. Electroacupuncture reduces cold stress-induced pain through microglial inactivation and transient receptor potential V1 in mice. Chin Med. 2021;16(1):43. doi:10.1186/s13020-021-00451-0
65. Gao YH, Li CW, Wang JY, et al. Effect of electroacupuncture on the cervicospinal P2X7 receptor/fractalkine/CX3CR1 signaling pathway in a rat neck-incision pain model. Purinergic Sig. 2017;13(2):215–225. doi:10.1007/s11302-016-9552-1
66. Prinz M, Jung S, Priller J. Microglia biology: one century of evolving concepts. Cell. 2019;179(2):292–311. doi:10.1016/j.cell.2019.08.053
67. Tansley S, Uttam S, Ureña Guzmán A, et al. Single-cell RNA sequencing reveals time- and sex-specific responses of mouse spinal cord microglia to peripheral nerve injury and links ApoE to chronic pain. Nat Commun. 2022;13(1):843. doi:10.1038/s41467-022-28473-8
68. Zhang W, Jiao B, Yu S, et al. Spinal AT1R contributes to neuroinflammation and neuropathic pain via NOX2-dependent redox signaling in microglia. Free Radic Biol Med. 2025;227:143–156. doi:10.1016/j.freeradbiomed.2024.12.004
69. Wang S, Sudan R, Peng V, et al. TREM2 drives microglia response to amyloid-β via SYK-dependent and -independent pathways. Cell. 2022;185(22):4153–4169.e19. doi:10.1016/j.cell.2022.09.033
70. The TREM2-APOE pathway drives the transcriptional phenotype of dysfunctional microglia in neurodegenerative diseases - PubMed. Available from: https://pubmed.ncbi.nlm.nih.gov/28930663/.
71. Different immune cells mediate mechanical pain hypersensitivity in male and female mice - PubMed. Available from: https://pubmed.ncbi.nlm.nih.gov/26120961/.
72. Kohno K, Shirasaka R, Yoshihara K, et al. A spinal microglia population involved in remitting and relapsing neuropathic pain. Science. 2022;376(6588):86–90. doi:10.1126/science.abf6805
73. Wu Z, Wang Y, Chen WW, et al. Peripheral nervous system microglia-like cells regulate neuronal soma size throughout evolution. Cell. 2025;188(8):2159–2174.e15. doi:10.1016/j.cell.2025.02.007
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.