Back to Journals » OncoTargets and Therapy » Volume 9
Metastasis-associated lung adenocarcinoma transcript 1 promotes the proliferation of chondrosarcoma cell via activating Notch-1 signaling pathway
Authors Xu F, Zhang Z, Fang Y, Li X, Sun Y, Xiong C, Yan L, Wang Q
Received 6 November 2015
Accepted for publication 20 January 2016
Published 13 April 2016 Volume 2016:9 Pages 2143—2151
DOI https://doi.org/10.2147/OTT.S100003
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Faris Farassati
Fengqin Xu,1,* Zhi-qiang Zhang,2,* Yong-chao Fang,2 Xiao-lei Li,2 Yu Sun,2 Chuan-zhi Xiong,2 Lian-qi Yan,2 Qiang Wang2
1Department of Orthopaedics, Hongquan Hospital, 2Department of Orthopaedics, Subei People’s Hospital, Yangzhou, Jiangsu Province, People’s Republic of China
*These authors contributed equally to this work
Background: Metastasis-associated lung adenocarcinoma transcript 1 (MALAT-1) is identified to be overexpressed in several cancers. However, the role of MALAT-1 in chondrosarcoma is poorly understood.
Methods: The expression of MALAT-1 and Notch-1 signaling pathway was detected in chondrosarcoma tissues and chondrosarcoma cells by quantitative real-time polymerase chain reaction (qRT-PCR) and Western blot. 3-(4,5-Dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide (MTT) assay was performed to examine the cell viability of chondrosarcoma cells transfected with si-MALAT-1 or pcDNA-MALAT-1. Then the expression of Notch-1 signaling pathway was detected when MALAT-1 was upregulated or downregulated in chondrosarcoma cells. A subcutaneous chondrosarcoma cells xenograft model was used to confirm the effect of MALAT-1 on tumor growth in vivo.
Results: We found the increased expression of MALAT-1 and Notch-1 signaling pathway in chondrosarcoma tissue and cells. MALAT-1 promoted the proliferation of chondrosarcoma cells. In addition, MALAT-1 activated the Notch-1 signaling pathway at posttranscriptional level in chondrosarcoma cells. Meanwhile, overexpression of Notch-1 reversed the effect of si-MALAT-1 on the proliferation of chondrosarcoma cells. Finally, we found that MALAT-1 promoted the tumor growth in a subcutaneous chondrosarcoma cells xenograft model, which confirmed the promoted effect of MALAT-1 on the tumor growth in vivo.
Conclusion: Taken together, our study demonstrated that MALAT-1 promoted the proliferation of chondrosarcoma cell via activating Notch-1 signaling pathway.
Keywords: MALAT-1, cell proliferation, chondrosarcoma, Notch-1, molecular pathophysiology
Introduction
Chondrosarcoma, along with myeloma and osteosarcoma, is a major malignancy of the bone.1 Pathologically, there are four kinds of chondrosarcoma: conventional, dedifferentiated, mesenchymal, and clear cell chondrosarcomas.2 Conventional chondrosarcomas, as the approximate 85% of all chondrosarcomas, have been found to be highly resistant to chemo- and radiotherapy because of their extracellular matrix and poor vascularity.2 In addition, the majority of chondrosarcomas grow slowly and rarely metastasize. However, the molecular pathophysiology of chondrosarcomas is not fully understood.
Various prosurvival pathways play important roles in biological processes. Among them, the Notch signaling pathway is increasingly being studied as a novel mechanism for tumorigenesis. Interestingly, Notch can serve as the apparently opposite functions in tumor development, such as an oncogene or a tumor suppressor, a repressor, or inducer of terminal differentiation.3 To date, there are four Notch receptors (Notch 1–4) identified in mammals, with five corresponding ligands, including Delta-like-1, Delta-like-3, Delta-like-4, Jagged-1, and Jagged-2.4 Overexpression of the Notch receptors and/or their ligands has now been identified in multiple cancers, including breast, brain, prostate, and ovarian.5–9 Recently, Notch signaling pathway was found to be involved in the apoptosis of osteosarcoma cells.10 According to the functional role of Notch signaling during cartilage development, Notch-1 is supposed to be a marker for chondrogenic progenitor cells.11
Metastasis-associated lung adenocarcinoma transcript 1 (MALAT-1) is a long noncoding RNA ubiquity expressed in various tissues and highly conserved in mammals.12 With respect to its function, MALAT-1 participates in the regulation of gene expressions, alternative splicing, and cell cycle.13 MALAT-1 was originally found to be overexpressed in patients at high risk for metastasis of non-small-cell lung tumors (NSCLC).14 Subsequently, various studies showed that MALAT-1 was overexpressed in several cancers, such as colon, prostate, breast, and oral squamous cell carcinoma.15–19 These findings indicated the oncogenic function of MALAT-1. However, the role of MALAT-1 in chondrosarcoma is poorly understood.
In the present study, we found that MALAT-1 and Notch signaling pathway were closely associated with the proliferation of chondrosarcoma cell. By manipulating the expression of MALAT-1 in chondrosarcoma cell, we have demonstrated the unambiguous role of MALAT-1 in tumorigenesis in vivo. Most importantly, we have identified Notch signaling pathway as the downstream target that is responsible for MALAT-1 function in chondrosarcoma cell proliferation.
Materials and methods
Clinical samples
Our study consisted of 20 chondrosarcoma tissue samples and 20 adjacent noncancerous tissues taken from the Subei People’s Hospital, People’s Republic of China. The pathological types of chondrosarcoma were conventional chondrosarcoma in 17 cases and myxoid chondrosarcoma in three cases. Histologically, they were grade II–III. Among them, lymph node metastasis occurred in four cases. Written informed consent was obtained from all patients, and the study protocols were approved by the ethics committee of Subei People’s Hospital. All tissue specimens were immediately frozen in liquid nitrogen after surgery and stored at −80°C until RNA isolation, and diagnosis of chondrosarcoma was verified by pathologists.
Western blotting
Western blotting assay was performed to examine the protein levels of Notch-1, Hes-1, Hey-1, and Hey-2. Proteins were separated by SDS-PAGE (sodium dodecyl sulfate polyacrylamide gel electropheresis) and electroblotted to polyvinylidene difluoride membranes (Millipore, Milford, MA, USA) according to manufacturer’s manual. After blocking with 5% nonfat dry-milk, membranes were incubated overnight at 4°C with the primary antibodies: Notch-1 (1:1,000), Hes-1 (1:2,000), Hey-1 (1:2,000) and Hey-2 (1:2,000), and β-actin (1:1,000). Afterward, the membrane was washed with PBS (phosphate buffer saline)/0.05% Tween-20, and incubated with antimouse or antirabbit secondary antibody conjugated to horseradish peroxidase for 30 minutes at room temperature. The enhanced chemiluminescence system was used to visualize the stained bands according to the manufacturer’s protocol. For the chondrosarcoma and the normal tissue, we selected three independent samples of chondrosarcoma and normal tissue, respectively, for the Western blotting assay.
Quantitative real-time PCR
The total RNA from treated cells was isolated by Trizol (Invitrogen, Carlsbad, CA, USA) and purified by RNeasy Mini Kit and RNase-free DNase Set (Qiagen, Chatsworth, CA, USA) according to the manufacturer’s protocols. First strand cDNA was synthesized with the PrimeScript™ RT Kit (Takara Biotechnology Co, Dalian, People’s Republic of China). MALAT-1 and Notch-1 expression were detected by both semi-quantitative polymerase chain reaction (PCR) and quantitative PCR (qPCR) using PrimeScript™ PCR Master Mix (Takara Biotechnology Co) and an ABI 7500 real-time PCR system. Glyceraldehyde phosphate dehydrogenase was used as an internal control. The assay was run in triplicate for each sample.
RNA pull-down assay
Biotin RNA Labeling Mix (Roche, Mannheim, Germany) and T7 RNA polymerase were used for in vitro transcription of biotin-labeled MALAT-1, the products of which were purified with RNeasy Mini Kit (Roche). Radio-immunoprecipitation assay buffer was utilized to prepare the cell lysate and then the cell lysate was incubated with the biotinylated RNA biotin-labeled RNAs at 4°C for 4 hours. The washed streptavidin agarose beads were added into each binding reaction to incubate for 1 hour at room temperature. Beads were washed five times and the retrieved proteins were run on SDS-PAGE gels for Western blot.
Cell viability assay
JJ012 and CH2879 cells were seeded in 96-well plates and transfected with si-MALAT-1 or pcDNA-MALAT-1, respectively. Then, 10 μL of 3-(4,5-dimethylthiazolyl-2) 2,5-diphenyltetrazolium bromide (MTT, 5 mg/mL, Sigma-Aldrich, Shanghai, People’s Republic of China) was added to each well and incubated for 4 hours at 37°C. After 4 hours, 100 μL of solubilization buffer was added to each well and optical density was then measured at a wavelength of 490 nm. Each MTT assay was performed in quadruplicate and repeated three times.
Cell culture and transfection
Chondrosarcoma cell lines CH2879, L835, JJ012 and cartilage cells TC28a2 were cultured in RPMI1640 (Invitrogen Life-Technologies, Scotland, UK) supplemented with 1% L-glutamax, 1% penicillin/streptomycin (P/S) (100 U/mL), and 10% heat-inactivated fetal calf serum (Invitrogen Life-Technologies). Cells were grown at 37°C in a humidified incubator with 95% air and 5% CO2. For in vitro study, JJ012 was transfected with siRNA (GenePharma, Shanghai, People’s Republic of China) against MALAT-1 to knock down MALAT-1 or pcDNA-Notch-1 to overexpress Notch-1 using Lipofectamine2000 transfection reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. CH2879 cells were transfected with siRNA (GenePharma) against Notch-1 to knock down Notch-1 or pcDNA-MALAT-1 to overexpress MALAT-1 using Lipofectamine2000 transfection reagent. siRNA-control and pcDNA were used as a negative control.
Determining the effect of MALAT-1 on tumor growth in vivo
All animal procedures were conducted according to the guidelines approved by the China Association of Laboratory Animal Care. Four weeks old male BALB/c-nu/nu mice (Slac Laboratory Animal, Shanghai, People’s Republic of China) were kept in the animal facilities at the Subei People’s Hospital under specific pathogen-free conditions and randomly divided into four groups (six mice per group). Cultured JJ012 cells, which were transfected with si-MALAT-1 or si-control, and CH2879 cells, which were transfected with pcDNA-MALAT-1 or pcDNA, were suspended in 0.2 mL PBS and then injected into the right flank of mice. The mice were kept in a pathogen-free environment. The tumor size was measured in two orthogonal directions using calipers at 8th, 12th, 16th, 20th, 24th, and 28th day, and the tumor volume (mm3) was estimated using the equation length × (width)2 ×0.5.
Statistical analysis
Each experiment was repeated at least three times. All the experimental data are presented as mean ± SD. The differences among mean values were statistically analyzed by a t-test. All statistical analyses were performed using SPSS13.0 software (SPSS, Chicago, IL, USA). P<0.05 was considered statistically significant.
Results
The expression of MALAT-1 and protein levels of Notch-1 signaling pathway in chondrosarcoma tissues and controls
To investigate the effect of MALAT-1 and Notch-1 signaling pathway on the pathology of chondrosarcoma, first of all, we examined the expression of MALAT-1 and Notch-1, Hes-1, Hey-1, and Hey-2 in the chondrosarcoma tissues, and the control tissues. As shown in Figure 1A, the mRNA level of MALAT-1 was increased in the chondrosarcoma tissues compared to that of in the controls. Additionally, the protein level of Notch-1 was enhanced in the chondrosarcoma tissues, as well as that of its target proteins Hes-1, Hey-1, and Hey-2 (Figure 1B).
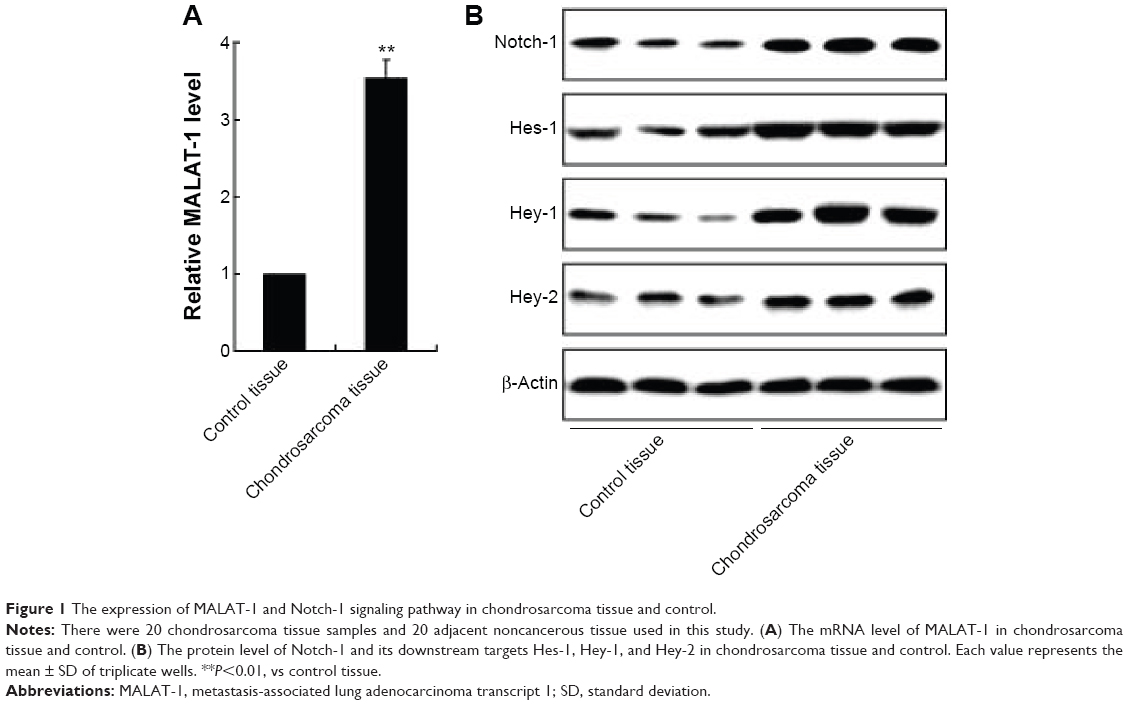 | Figure 1 The expression of MALAT-1 and Notch-1 signaling pathway in chondrosarcoma tissue and control. |
The expression of MALAT-1 and Notch-1 signaling pathway in chondrosarcoma cells and controls
To verify the dysregulation of MALAT-1 and Notch-1 signaling pathway in chondrosarcoma, we detected the expression of MALAT-1 and Notch-1, Hes-1, Hey-1, and Hey-2 in chondrosarcoma cells (CH2879, L835, JJ012), and TC28a2 as the control cells. It has been shown that the mRNA level of MALAT-1 was higher in chondrosarcoma cells than that in control cells (Figure 2A). Moreover, the mRNA level of MALAT-1 was highest in JJ012 among the three kinds of chondrosarcoma cells. Consistent with the results in chondrosarcoma tissue, the protein levels of Notch-1 as well as Hes-1, Hey-1, and Hey-2 were significantly enhanced in chondrosarcoma cells (Figure 2B). These data indicated that MALAT-1 and Notch-1 signaling pathway might be involved in the pathology of chondrosarcoma.
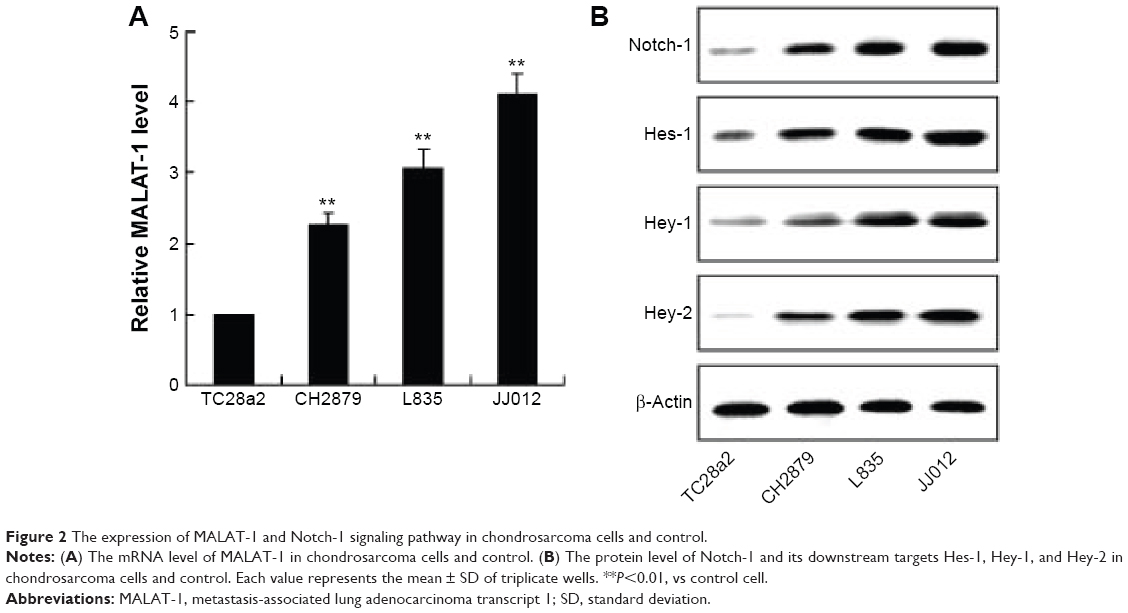 | Figure 2 The expression of MALAT-1 and Notch-1 signaling pathway in chondrosarcoma cells and control. |
The effect of MALAT-1 on the proliferation of chondrosarcoma cells
Next, JJ012 cells were transfected with si-MALAT-1 to obtain MALAT-1 knockdown, and the cell viability of JJ012 cells were detected. As shown in Figure 3A, the mRNA level of MALAT-1 was significantly decreased by si-MALAT-1, which also greatly suppressed the cell viability of JJ012 cells. Then CH2879 cells were transfected with pcDNA-MALAT-1 to overexpress MALAT-1. It has been shown that the cell viability of CH2879 cells transfected with pcDNA-MALAT-1 was greatly enhanced compared with that of CH2879 cells transfected with pcDNA (Figure 3B). These data indicated that MALAT-1 promoted the proliferation of chondrosarcoma cells.
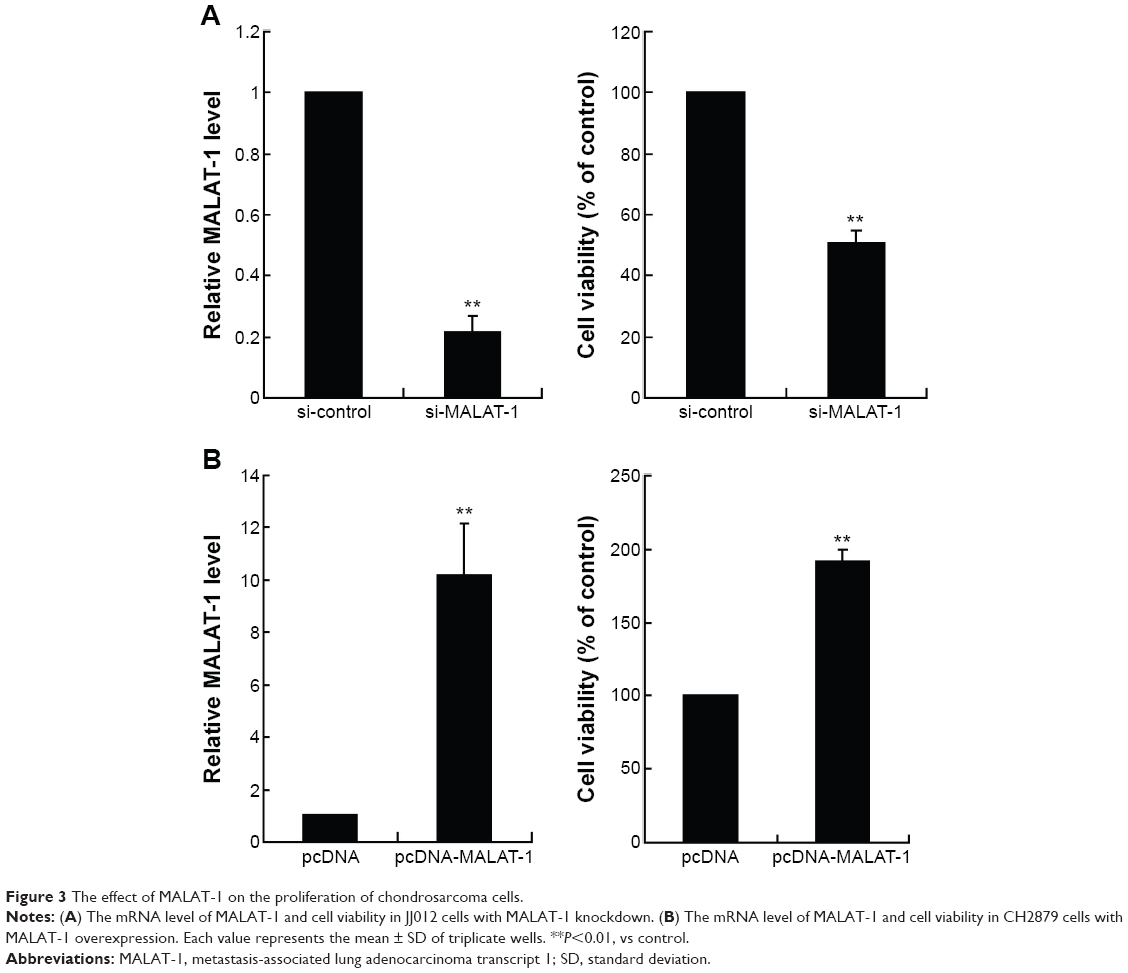 | Figure 3 The effect of MALAT-1 on the proliferation of chondrosarcoma cells. |
The effect of MALAT-1 on the expression of Notch-1 in chondrosarcoma cells
To investigate the regulated effect of MALAT-1 on the Notch-1 signaling pathway, JJ012 cells were transfected with si-MALAT-1, then the mRNA level of Notch-1 and the protein level of Notch-1 and Hes-1, Hey-1, and Hey-2 were detected. As shown in Figure 4A, si-MALAT-1 exhibited no significant effect on the mRNA level of Notch-1 whereas the protein levels of Notch-1 and Hes-1, Hey-1, and Hey-2 were reduced by si-MALAT-1. Cycloheximide (CHX) is a well-known inhibitor of protein biosynthesis in vivo and in vitro. si-MALAT-1 further accelerated the degradation of Notch-1 induced by CHX (Figure 4B). Moreover, the expression of Notch-1 signaling pathways was examined in CH2879 cells transfected with pcDNA-MALAT-1. Overexpression of MALAT-1 enhanced the protein level of Notch-1 and Hes-1, Hey-1, and Hey-2 (Figure 4C). To further elucidate the relationship between MALAT-1 and Notch-1, RNA pull-down assay was performed and the results indicated that MALAT-1 could bind to Notch-1 (Figure 4D). These results indicated that MALAT-1 could activate the Notch-1 signaling pathway at posttranscriptional level.
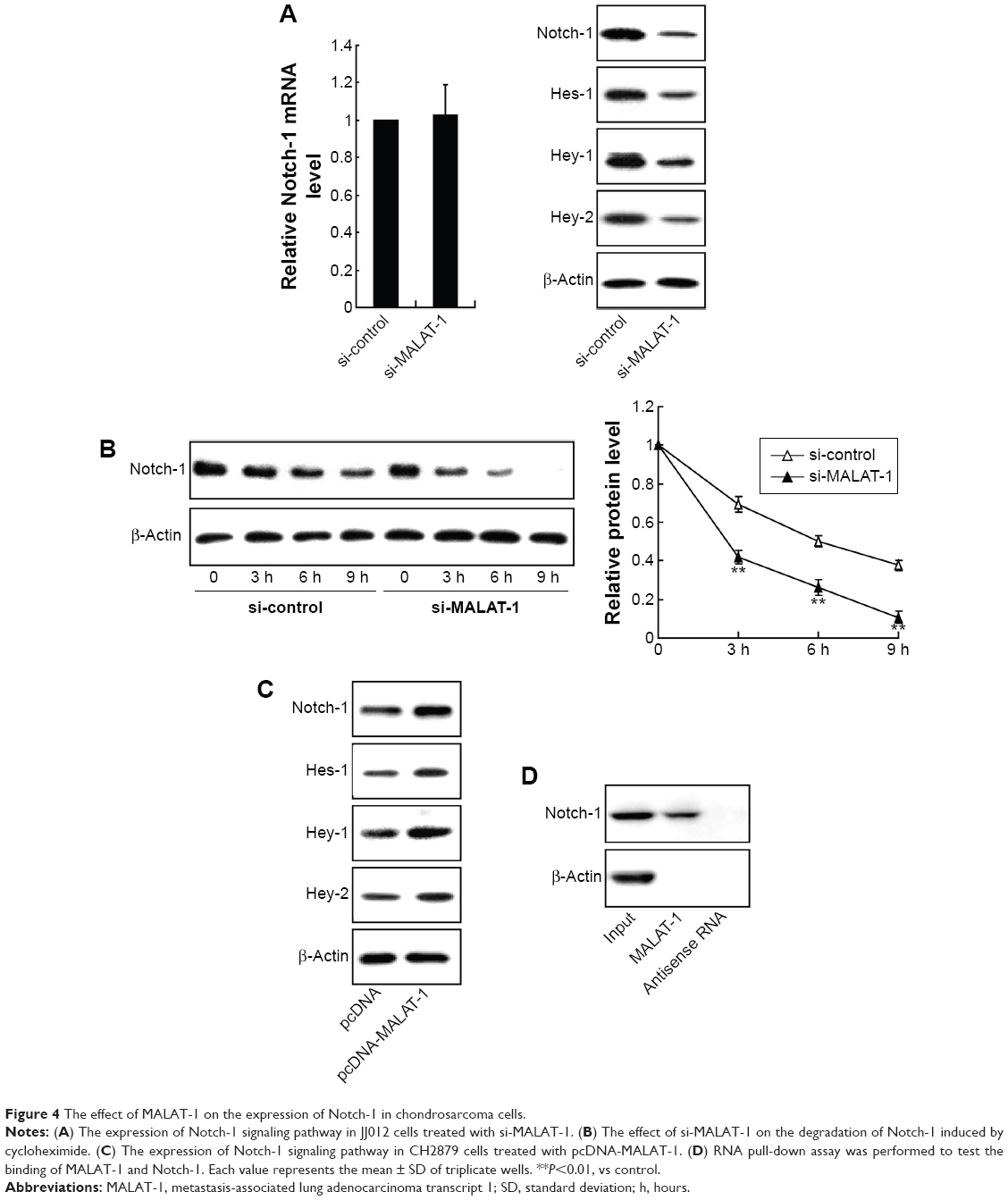 | Figure 4 The effect of MALAT-1 on the expression of Notch-1 in chondrosarcoma cells. |
The effect of Notch-1 on the proliferation of chondrosarcoma cells regulated by MALAT-1
According to the aforementioned experiments, we knew that MALAT-1 could promote the proliferation of chondrosarcoma cells. Here, we investigated the effect of Notch-1 on the proliferation of chondrosarcoma cells regulated by MALAT-1. JJ012 cells were transfected with si-MALAT-1 and pcDNA or si-MALAT-1 and pcDNA-Notch-1. As shown in Figure 5A, the cell viability of JJ012 transfected with si-MALAT-1 and pcDNA-Notch-1 was highly increased. When CH2879 cells were transfected with pcDNA-MALAT-1 and si-Notch-1, the cell viability was lower than that of cells transfected with pcDNA-MALAT-1 and si-control (Figure 5B). These data indicated that Notch-1 overexpression reversed the effect of si-MALAT-1 on the proliferation of chondrosarcoma cells.
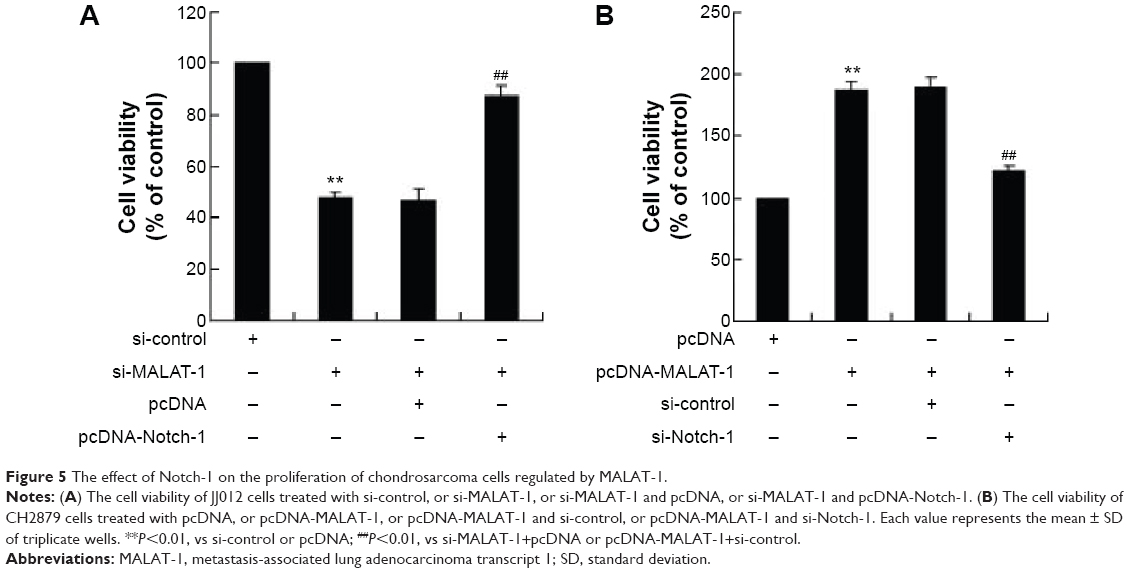 | Figure 5 The effect of Notch-1 on the proliferation of chondrosarcoma cells regulated by MALAT-1. |
MALAT-1 promotes the tumor growth in a subcutaneous chondrosarcoma cells xenograft model
To determine the effect of MALAT-1 in vivo, we evaluated its effect in a nude mouse xenograft model of JJ012 cells or CH2879 cells. JJ012 cells were transfected with si-MALAT-1 or si-control and subcutaneously inoculated into the nude mice (six mice per group). Then, the tumor volume was measured. Si-MALAT-1 inhibited tumor growth significantly after inoculation for 28 days (Figure 6A). CH2879 cells transfected with pcDNA-MALAT-1 or pcDNA were subcutaneously inoculated into the nude mice (six mice per group) and the tumor volume was measured. As shown in Figure 6B, the tumor volume was increased in xenograft tumors of mice treated with CH2879 cells containing pcDNA-MALAT-1 compared with the control. Results in vivo further confirmed the promoted effect of MALAT-1 on the tumor growth.
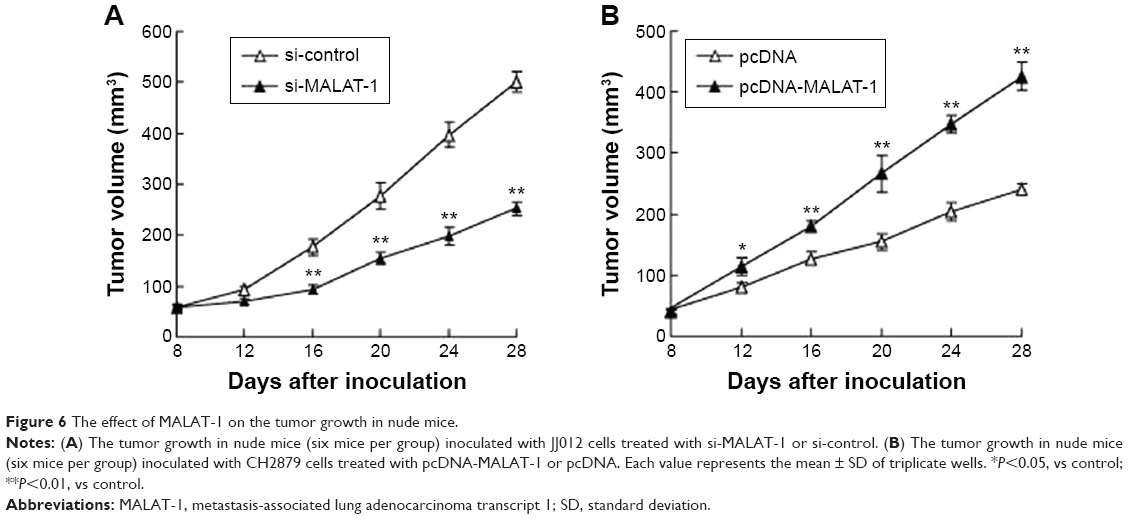 | Figure 6 The effect of MALAT-1 on the tumor growth in nude mice. |
Discussion
In this study, we have demonstrated the high expression of MALAT-1 and Notch-1 signaling pathway in chondrosarcoma tissue and cells. MALAT-1 could promote the proliferation of chondrosarcoma cells. In addition, MALAT-1 activated the Notch-1 signaling pathway at posttranscriptional level in chondrosarcoma cells. Meanwhile, Notch-1 overexpression reversed the effect of si-MALAT-1 on the proliferation of chondrosarcoma cells. Finally, we found that MALAT-1 promoted the tumor growth in a subcutaneous chondrosarcoma cells xenograft model, which confirmed the promoted effect of MALAT-1 on the tumor growth in vivo.
Notch-1, as a critical moderator of the fate of all stem cells, plays a pivotal role in the differentiation of many tissues. The Notch signaling pathway is conserved from Drosophila to humans, which consists of Notch receptors, ligands, negative and positive modifiers, and transcription factors.20 Through proteolytic cleavage events and interacting with the transcription regulator C-promoter binding factor 1 (CSL, suppressor of hairless, Lag-1), Notch could activate several target genes, including several helix-loop-helix transcription factors collectively named hairy/enhancer of split (HES) and hairy and enhancer of split-related with YRPW motif (HEY).21 Notch also has been associated with the pathogenesis of several cancers.20 In ovarian cancer, depletion of Notch-1 led to the growth inhibition of cancer cells.22 siRNA targeting Notch-1 decreases glioma stem cell proliferation and tumor growth.23 Ai et al24 showed that Notch-1 might act as an oncogene, regulating the proliferation and differentiation of bladder cancer cells by inhibiting Krüppel-like factor 4.
Notch signaling inhibition is a viable strategy for the treatment of several solid and hematopoietic tumor. Numerous genetic and pharmacological strategies are available to block or silence Notch signaling for therapeutic purposes.25 In this study, we found the upregulation of Notch-1 and its target genes Hes-1, Hey-1, and Hey-2 in chondrosarcoma tissue and cells, which indicated that the Notch signaling pathway might be involved in the pathogenesis of chondrosarcoma.
Emerging evidences show that dysregulation of long non-coding RNAs is an important feature of many human diseases, including ischemic diseases, heart disease, and cancers.26 MALAT-1 is an evolutionarily highly conserved transcript and localizes in specific subcellular position. MALAT-1 has been shown to be overexpressed in many solid tumors and promotes the proliferation/migration of cancer cells.27 It is reported that MALAT-1 inhibits G2/M cell cycle arrest to promote epithelial–mesenchymal transition in pancreatic cancer.28 MALAT-1 is also a critical regulator of the metastasis phenotype of lung cancer cells.29 Mounting studies have indicated that MALAT-1 contributed to the cancer cell migration and invasion by influencing the expression of motility-related genes and disturbing the apoptosis pathways.15,30 Lai et al30 found that MALAT-1 overexpression could be used to predict tumor recurrence of hepatocellular carcinoma after liver transplantation. Based on these researches, we detected the role of MALAT-1 in chondrosarcoma cells. We found that MALAT-1 was overexpressed in chondrosarcoma cells and it promoted the proliferation of chondrosarcoma cell. To further explore the molecular mechanism of MALAT-1 promoting the cell proliferation, we examined the regulation of MALAT-1 on the expression of Notch signaling pathway. MALAT-1 could activate the Notch-1 signaling pathway at posttranscriptional level. In addition, overexpression of Notch-1 reversed the effect of si-MALAT-1 on the proliferation of chondrosarcoma cells.
Taken together, our studies demonstrated that MALAT-1 and Notch-1 were involved in the pathogenesis of chondrosarcoma. Then, increase in MALAT-1 RNA caused the boost in proliferation, which could be reversed by Notch-1 knockdown. These data indicated that MALAT-1 promoted the proliferation of chondrosarcoma cell via activating Notch-1 signaling pathway. Our findings suggested the important roles of MALAT-1 and Notch-1 signaling pathway in the proliferation of chondrosarcoma cell, which put MALAT-1 and Notch-1 forward as valid therapeutic targets to be further tested in vivo.
Disclosure
The authors report no conflicts of interest in this work.
References
Rozeman LB, Cleton-Jansen AM, Hogendoorn PC. Pathology of primary malignant bone and cartilage tumours. Int Orthop. 2006;30(6):437–444. | ||
Gelderblom H, Hogendoorn PC, Dijkstra SD, et al. The clinical approach towards chondrosarcoma. Oncologist. 2008;13(3):320–329. | ||
Roy M, Pear WS, Aster JC. The multifaceted role of Notch in cancer. Curr Opin Genet Dev. 2007;17(1):52–59. | ||
Bolós V, Grego-Bessa J, de la Pompa JL. Notch signaling in development and cancer. Endocr Rev. 2007;28(3):339–363. | ||
Meng RD, Shelton CC, Li Y-M, et al. γ-Secretase inhibitors abrogate oxaliplatin-induced activation of the Notch-1 signaling pathway in colon cancer cells resulting in enhanced chemosensitivity. Cancer Res. 2009;69(2):573–582. | ||
Reedijk M, Odorcic S, Chang L, et al. High-level coexpression of JAG1 and NOTCH1 is observed in human breast cancer and is associated with poor overall survival. Cancer Res. 2005;65(18):8530–8537. | ||
Hopfer O, Zwahlen D, Fey M, Aebi S. The Notch pathway in ovarian carcinomas and adenomas. Br J Cancer. 2005;93(6):709–718. | ||
Santagata S, Demichelis F, Riva A, et al. JAGGED1 expression is associated with prostate cancer metastasis and recurrence. Cancer Res. 2004;64(19):6854–6857. | ||
Fan X, Mikolaenko I, Elhassan I, et al. Notch1 and notch2 have opposite effects on embryonal brain tumor growth. Cancer Res. 2004;64(21):7787–7793. | ||
Ji P, Yu L, Guo W-C, et al. Doxorubicin inhibits proliferation of osteosarcoma cells through upregulation of the Notch signaling pathway. Oncol Res. 2015;22(4):185–191. | ||
Oldershaw RA, Hardingham TE. Notch signaling during chondrogenesis of human bone marrow stem cells. Bone. 2010;46(2):286–293. | ||
Bernard D, Prasanth KV, Tripathi V, et al. A long nuclear-retained non-coding RNA regulates synaptogenesis by modulating gene expression. EMBO J. 2010;29(18):3082–3093. | ||
Cho S-F, Chang YC, Chang C-S, et al. MALAT1 long non-coding RNA is overexpressed in multiple myeloma and may serve as a marker to predict disease progression. BMC Cancer. 2014;14(1):809. | ||
Ji P, Diederichs S, Wang W, et al. MALAT-1, a novel noncoding RNA, and thymosin β4 predict metastasis and survival in early-stage non-small cell lung cancer. Oncogene. 2003;22(39):8031–8041. | ||
Guo F, Li Y, Liu Y, Wang J, Li Y, Li G. Inhibition of metastasis-associated lung adenocarcinoma transcript 1 in CaSki human cervical cancer cells suppresses cell proliferation and invasion. Acta Biochim Biophys Sin(Shanghai). 2010;42(3):224–229. | ||
Kan JY, Wu DC, Yu FJ, et al. Chemokine (C-C Motif) ligand 5 is involved in tumor-associated dendritic cell-mediated colon cancer progression through non-coding RNA MALAT-1. J Cellular Physiol. 2015;230(8):1883–1894. | ||
Wang F, Ren S, Chen R, et al. Development and prospective multicenter evaluation of the long noncoding RNA MALAT-1 as a diagnostic urinary biomarker for prostate cancer. Oncotarget. 2014;5(22):11091–11102. | ||
Jin C, Yan B, Lu Q, Lin Y, Ma L. Reciprocal regulation of Hsa-miR-1 and long noncoding RNA MALAT1 promotes triple-negative breast cancer development. Tumour Biol. Epub 2015 Dec 16. | ||
Zhou X, Liu S, Cai G, et al. Long non coding RNA MALAT1 promotes tumor growth and metastasis by inducing epithelial-mesenchymal transition in oral squamous cell carcinoma. Sci Rep. 2015;5:15972. | ||
Allenspach EJ, Maillard I, Aster JC, Pear WS. Notch signaling in cancer. Cancer Biol Ther. 2002;1(5):466–476. | ||
Chen Y, De Marco MA, Graziani I, et al. Oxygen concentration determines the biological effects of NOTCH-1 signaling in adenocarcinoma of the lung. Cancer Res. 2007;67(17):7954–7959. | ||
Rose SL, Kunnimalaiyaan M, Drenzek J, Seiler N. Notch 1 signaling is active in ovarian cancer. Gynecol Oncol. 2010;117(1):130–133. | ||
Wang J, Wang C, Meng Q, et al. siRNA targeting Notch-1 decreases glioma stem cell proliferation and tumor growth. Mol Biol Rep. 2012;39(3):2497–2503. | ||
Ai X, Jia Z, Liu S, Wang J, Zhang X. Notch-1 regulates proliferation and differentiation of human bladder cancer cell lines by inhibiting expression of Krüppel-like factor 4. Oncol Rep. 2014;32(4):1459–1464. | ||
Nickoloff BJ, Osborne BA, Miele L. Notch signaling as a therapeutic target in cancer: a new approach to the development of cell fate modifying agents. Oncogene. 2003;22(42):6598–6608. | ||
Perez DS, Hoage TR, Pritchett JR, et al. Long, abundantly expressed non-coding transcripts are altered in cancer. Hum Mol Genet. 2008;17(5):642–655. | ||
Yang M-H, Hu Z-Y, Xu C, et al. MALAT1 promotes colorectal cancer cell proliferation/migration/invasion via PRKA kinase anchor protein 9. Biochim Biophys Acta. 2015;1852(1):166–174. | ||
Jiao F, Hu H, Yuan C, et al. Elevated expression level of long noncoding RNA MALAT-1 facilitates cell growth, migration and invasion in pancreatic cancer. Oncol Rep. 2014;32(6):2485–2492. | ||
Gutschner T, Hämmerle M, Eißmann M, et al. The noncoding RNA MALAT1 is a critical regulator of the metastasis phenotype of lung cancer cells. Cancer Res. 2013;73(3):1180–1189. | ||
Lai MC, Yang Z, Zhou L, et al. Long non-coding RNA MALAT-1 overexpression predicts tumor recurrence of hepatocellular carcinoma after liver transplantation. Med Oncol. 2012;29(3):1810–1816. |
 © 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.