Back to Journals » Neuropsychiatric Disease and Treatment » Volume 12
Memory deficit in patients with schizophrenia and posttraumatic stress disorder: relational vs item-specific memory
Received 16 January 2016
Accepted for publication 26 February 2016
Published 10 May 2016 Volume 2016:12 Pages 1157—1166
DOI https://doi.org/10.2147/NDT.S104384
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Wai Kwong Tang
Wookyoung Jung,1 Seung-Hwan Lee1,2
1Clinical Emotions and Cognition Research Laboratory, Department of Psychiatry, Inje University, Ilsan-Paik Hospital, 2Department of Psychiatry, Inje University, Ilsan-Paik Hospital, Goyang, Korea
Abstract: It has been well established that patients with schizophrenia have impairments in cognitive functioning and also that patients who experienced traumatic events suffer from cognitive deficits. Of the cognitive deficits revealed in schizophrenia or posttraumatic stress disorder (PTSD) patients, the current article provides a brief review of deficit in episodic memory, which is highly predictive of patients’ quality of life and global functioning. In particular, we have focused on studies that compared relational and item-specific memory performance in schizophrenia and PTSD, because measures of relational and item-specific memory are considered the most promising constructs for immediate tangible development of clinical trial paradigm. The behavioral findings of schizophrenia are based on the tasks developed by the Cognitive Neuroscience Treatment Research to Improve Cognition in Schizophrenia (CNTRICS) initiative and the Cognitive Neuroscience Test Reliability and Clinical Applications for Schizophrenia (CNTRACS) Consortium. The findings we reviewed consistently showed that schizophrenia and PTSD are closely associated with more severe impairments in relational memory compared to item-specific memory. Candidate brain regions involved in relational memory impairment in schizophrenia and PTSD are also discussed.
Keywords: schizophrenia, posttraumatic stress disorder, episodic memory deficit, relational memory, item-specific memory, prefrontal cortex, hippocampus
Introduction
Virtually every conscious thought and act we possess are based on our ability to remember past events – episodic memory.1,2 Memory performance is the strongest predictor of functional outcome and, therefore, research on the assessment and treatment of episodic memory disorders has received considerable attention.3,4 Disturbance in such cognitive ability is highly likely to bring about critical dysfunctions in everyday activities. In fact, episodic memory deficits have been identified as a key area of impairment in schizophrenia.5–7
Considering the importance in daily life and clinical settings, research on episodic memory impairments in schizophrenia needs a more tangible development of clinical trial paradigms, because current medications have only a modest effect.1 As part of an effort to improve clinical paradigms, the Cognitive Neuroscience Treatment Research to Improve Cognition in Schizophrenia (CNTRICS) initiative was organized, and its first aim was to focus on episodic memory. Cognitive Neuroscience Test Reliability and Clinical Applications for Schizophrenia (CNTRACS) Consortium also chose episodic memory as one of the core domains to improve functional outcomes in schizophrenia.
A consensus was attained in CNTRICS that measures of relational and item-specific memory are the most promising constructs for immediate translational development,1 as existing behavioral and imaging data provide consistent evidence that relational memory in schizophrenia is disproportionately impaired, whereas item-specific memory is relatively spared. Relational memory is a type of memory that focuses on the associations between items and is defined as the processes involved in the memory for stimuli/events and how they are associated with coincident context, stimuli, or events, whereas item-specific memory is based on the distinctiveness of specific items and is defined as the processes involved in memory for individual stimuli or elements, irrespective of contemporaneously presented context or elements.1
Episodic memory impairment appears to be a problem not only for schizophrenia but also for posttraumatic stress disorder (PTSD). One of the core symptoms of PTSD is intrusive re-experiencing. Re-experiencing symptoms are assumed to occur due to disturbances in the way trauma memories are encoded, organized, and retrieved in memory processing.8 Such disturbances are closely related to the lack in the defining feature of episodic memory – the awareness that the content of the memory is something from the past – so that they appear cause the “here and now” symptom in PTSD, that is, the feeling that the sensations are experienced in the present rather than a memory from the past.8 In light of the notion that episodic memory refers to the capacity to recall specific personal experiences, episodic memory impairments in PTSD are as severe as those in schizophrenia.
Of the several theories of PTSD, a cognitive model proposed by Ehlers and Clark9 explains the phenomenon of intrusive re-experiencing in PTSD as associative learning. According to the model, strong stimulus–stimulus and stimulus–response associations are formed for traumatic materials in PTSD. During traumatic experiences (unconditioned stimulus), strong conditioning causes the unconditioned emotional responses to the trauma and the stimuli (conditioned stimulus) present shortly before or during the trauma to be associated. The conditioned emotional responses overgeneralize more broadly to stimuli and situations that are similar to the original trauma, thus explaining that emotional and physiological responses can be triggered by a wide range of stimuli.8–10 That is, such a tendency to form aberrant associations may bring about intrusive re-experiencing in PTSD.
As briefly mentioned in the Introduction, considerable progress has been observed on the memory impairments of patients with schizophrenia through the efforts taken by CNTRICS and CNTRACS. In contrast, comparatively little research has been done on memory impairments in PTSD, because only recently PTSD has been classified under mental disorders.11 Through this brief review, we aimed to seek the evidence on whether patients with PTSD also have a disproportionate impairment in relational memory, assuming that relational and item-specific memories would be also promising for immediate translational development in PTSD as well as in schizophrenia. Here, the terms “associative memory” and “relational memory” are usually used interchangeably. We have used the term “associative memory” for PTSD review because associative memory seems to be the more common term in the literature pertaining to PTSD.12,13 To the extent that the core symptom of PTSD is related to associative memory deficits, the evidence supporting the fact that relational and item-specific memory would be also promising for immediate translational development in PTSD, as well as in schizophrenia, could also be found.
In addition, we have presented the specific findings of the fact that schizophrenia disproportionately impairs relational memory, based upon the tasks suggested by the CNTRICS and CNTRACS:14 relational and item-specific encoding (RISE) task,15 associative inference paradigm (AIP),16,17 and levels-of-processing (LOP) effects.18
This review is organized as follows: before the main review of episodic memory impairment, we have first provided a brief overview of the mechanisms of memory formation, and then behavioral and neuroimaging findings on each task in schizophrenia are summarized in sequence. For PTSD, few experiments were conducted on relational vs item-specific memory without any specific task. Accordingly, we have provided behavioral and neuroimaging evidence focusing on a small number of studies. The results of each task in schizophrenia and PTSD are summarized in Tables 1 and 2, respectively. In addition, brain regions involved in memory deficit in schizophrenia and PTSD are compared in Table 3.
 | Table 1 Task summary and behavioral and neuroimaging findings for schizophrenia |
 | Table 2 Task summary and behavioral and neuroimaging findings for PTSD |
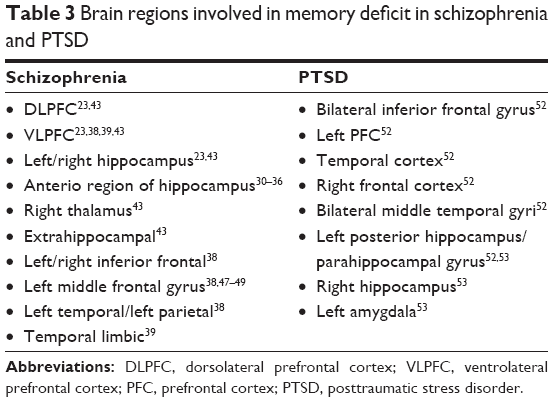 | Table 3 Brain regions involved in memory deficit in schizophrenia and PTSD |
Overview of the mechanism of memory formation
Long-term memory is generally thought to consist of two types of memory: declarative memory (also referred to as explicit memory) and nondeclarative memory (referred to as implicit or procedural memory). Declarative memory encompasses semantic memory, the memory of general knowledge and facts, and episodic memory, the memory of personally experienced events or episodes.2 Episodic memory is bound to specific contexts.
The process of memory formation involves three stages: encoding, consolidation, and retrieval. What critically determines whether an event or experience will be remembered or not is the encoding process. Encoding typically involves the initial process in which information to be remembered is transformed into a memory representation. Encoding generally falls into two categories – relational encoding and item-specific encoding. Relational encoding emphasizes associations among items, allowing information to be processed more elaborately, whereas item-specific encoding emphasizes the distinctiveness of specific items.19–21 Encoded memories are more stable over time through consolidation such that it is more likely that an event will be remembered subsequently. The final stage of memory processing is retrieval, which refers to the processes by which encoded and stored memory traces are reactivated or re-accessed.22 With the brief background in memory formation, we have proceeded to the main part of behavioral findings on episodic memory impairment in schizophrenia.
Findings for associative vs item-specific episodic memory impairment in behavioral findings of schizophrenia based on the RISE task
As explained here, formation of episodic memory is mainly composed of encoding (and consolidation) and retrieval processes; therefore, it is important to clarify the locus of the memory impairment to better understand episodic memory deficits in schizophrenia. The attempt to assess the locus has led to the development of an experimental paradigm known as the RISE task, which was designed not only to reliably measure episodic memory but also to dissociate discrete encoding and retrieval stages involved in memory impairments of patients with schizophrenia.15,23
In the RISE task, the encoding process is examined using two specific manipulations: relational and item-specific encoding. For example, the relational encoding manipulation presents pairs of items and asks subjects to decide whether one of the items could fit inside the other in real life, whereas the item-specific encoding manipulation presents a single item and asks subjects to decide whether each item is living or nonliving. The retrieval process is usually examined by two recognition formats: 1) an item-recognition task in which studied and non-studied items are randomly presented, asking whether each item is old or new 2) and an associative-recognition task in which old item pairs studied during relational encoding and new (ie, non-studied) item pairs are presented and subjects are asked whether each pair is presented together or not.
The retrieval function is generally captured by two cognitive processes: recollection and familiarity. Recollection is the conscious recall of specific contextual and event details of prior occurrences, whereas familiarity is the subjective sense of having previously encountered an item without any specific details.24,25 For example, when using a word “BIRD” to test patients, asking the patient whether the word was presented in capital letters or not is intended to measure recollection, whereas simply asking whether the word was previously presented or not, irrespective of upper or lowercase letters, is to measure familiarity.
A successful retrieval in the RISE task is determined by the type of recognition task and the type of cognitive process involved in retrieval. A successful associative recognition test depends more on recollection because the associative recognition test targets only studied, familiar items and measures the ability to discriminate studied and non-studied (ie, newly rearranged) pairs, so that recollection is a more valid test measure. In contrast, a successful item recognition test depends more on familiarity because the item recognition test covers both studied and non-studied items and measures the ability to discriminate new from old items, which is possible by simply knowing whether the item is familiar or not.23
Ragland et al15 found that item recognition accuracy in healthy controls was somewhat higher following item-specific encoding than following relational encoding. Between-group comparisons revealed that recognition memory in schizophrenia patients was spared as in healthy controls when item-specific encoding and familiarity-based retrieval were required together. However, the recognition performance in patients declined when relational encoding and familiarity-based retrieval were required, and also declined in the case of recollection-based retrieval, regardless of the type of encoding. The worst decline in patients was revealed under the condition in which both relational encoding and recollection-based retrieval were required together. In conclusion, the RISE task highlights to elucidate the nature of memory impairment in schizophrenia. Whereas conscious recollection and associative recognition are severely impaired in schizophrenia, familiarity is significantly impaired depending on the way in which patients encode information.
Neuroimaging findings for the RISE task
It is well known that the prefrontal cortex (PFC) and medial temporal lobe (MTL) make a critical contribution to episodic memory.23,26,27 Ragland et al23 used functional magnetic resonance imaging (fMRI) during RISE task performance to test the hypothesis that schizophrenia disproportionately affects PFC and MTL subregions during relational encoding and retrieval relative to item-specific memory processes.
The behavioral findings of relational memory impairment were replicated.15 The fMRI data showed that these memory deficits were linked to reduced activation in dorsolateral prefrontal cortex (DLPFC) during relational encoding and to reduced activation in left and right hippocampus (HI) during successful recognition following relational encoding.
The findings imply that schizophrenia might have disrupted PFC and HI subregions and mnemonic processes. Patients were most impaired following relational encoding, which demanded engagement of the DLPFC during encoding and left and right HI during retrieval. In contrast, patients were relatively less impaired when engagement of the ventrolateral prefrontal cortex was required to encode item-specific information.23
In summary, the results provided evidence of the subregions of the PFC and MTL that contributed to encoding and retrieval processes by comparing healthy and schizophrenia subjects.23 Activity increases in the ventrolateral prefrontal cortex when goal-relevant features of items are activated or inhibited in order to induce successful item recognition, whereas activity increases in the DLPFC when relational processing between items is required so that retrieval of relational information or relational recognition is facilitated.28 Behavioral and neuroimaging results of the RISE task emphasized that a more integrated and refined account of episodic memory in schizophrenia should not overlook the importance of disentangling discrete encoding and retrieval processes.
Behavioral findings for the AIP
The AIP, which was launched by the CNTRICS initiative,14 was also designed to investigate the disproportionate impairment of relational memory in schizophrenia. The AIP tests the ability to infer new associations from novel situations based on associations of studied items; therefore, their hypothesis was that schizophrenia patients would have difficulty recognizing previously studied paired associates and making a relational binding of studied items. In the initial version,16 35% of the subjects with schizophrenia were excluded due to poor performance. As such, the modified version included a smaller number of training pairs, additional feedback, and clear instructions to make the task easier.
In the modified version,17 both schizophrenia patients and healthy controls were trained on three sets of paired associates of houses (H) and faces (F). All subjects completed three study-test sessions on 10 pairs in a fixed order (H-F1, H-F2, F3-F4). For example, in the H-F1 pair, subjects were presented with one wood house–blonde woman pair during the study and then presented with a two-alternative forced-choice task in which the same blonde woman, the same wood house, and a new brick house were presented. Subjects were asked to choose the house that matched the blonde woman. After training, each group was tested on a non-inferential pair (ie, already studied during training, such as the F3-F4 pair) and on an inferential pair (ie, a novel pair, such as the F1-F2 pair). The results showed that schizophrenia patients were more impaired on inferential pairs (F1-F2) than non-inferential pairs (F3-F4) compared to healthy controls, which confirms that relational memory is impaired in schizophrenia.
In addition to their main finding, another important message in this study is that the researchers succeeded in improving the feasibility of the task by remedying the shortcomings from the previous study. They seem to provide a good example for the iterative process of implementing experimental designs from the cognitive neuroscience literature in schizophrenia research.29
Neuroimaging findings for the AIP
To date, neuroimaging data for the AIP with schizophrenia patients are not yet available. The majority of fMRI studies with healthy subjects show that the anterior region of the HI appears to be involved in the relational memory task.30–36 Based on the current findings with healthy controls, future research should aim to obtain structural and functional neuroimaging data for insight into the neural correlates of memory deficit in schizophrenia.
Behavioral findings for the LOP effects
Another experimental task that is used to examine memory impairment in schizophrenia patients is the LOP paradigm,37 which assumes that the memory trace is influenced by the type of encoding strategy – deep vs shallow encoding. The deep encoding is a strategy that helps the information to be processed in a more semantic and elaborate way, such as deciding whether the word presented is concrete or abstract, whereas the shallow encoding is a strategy that leads to focus more on physical characteristics of the information, such as deciding if the word presented is lowercase or uppercase. The basic mechanism of the paradigm is that the memory trace is more strengthened as the encoding strategy moves from shallow (ie, perceptual processing) to deep processing (ie, semantically associated).
Ragland et al tested the LOP effect on word recognition in schizophrenia.18 Their hypothesis was that schizophrenia patients would have difficulty encoding semantic information and thus would show a reduced LOP effect. During deep encoding, schizophrenia and healthy subjects were asked to decide if the word presented was concrete or not, and during shallow encoding, both groups of subjects were asked to decide if the word presented was in uppercase letters. For the recognition test, words were randomly chosen from the two encoding conditions. Subjects decided whether the word was in the encoding list. Contrary to the hypothesis, even schizophrenia patients, as well as healthy controls, showed a robust LOP effect in that the deep encoding strategy helped schizophrenia patients to better recognize the studied words. The implication of these findings is that semantic knowledge is intact in schizophrenia and hence schizophrenia patients could benefit from organizational strategies associated with semantic encoding.18,38,39
The other finding that schizophrenia patients have intact semantic knowledge was examined in false memory using the Deese–Roediger–McDermott paradigm,40 in which subjects were presented with a list of target words (eg, bed, dream, pillow, and rest) that relate to a critical lure (eg, sleep) that was never presented during the study. When tested, subjects produced robust false memory for the critical lure they did not study. From the paradigm, false memory was thought to stem from implicit processing of semantic association.40,41
Paz-Alonso et al42 tested false memory in schizophrenia using the Deese–Roediger–McDermott paradigm. They used two types of word list including high associative strength, which is more likely to produce false memory, and low associative strength, which is less likely to produce false memory. Schizophrenia patients and healthy subjects studied word lists under two different conditions: a standard condition (ie, simply required to remember the word) and a relational condition (ie, required to think about the relationship between the words). Patients showed impaired true memory (ie, recognizing the words in the list) when relational encoding was explicitly required compared to healthy controls, which means patients were impaired in their ability to explicitly engage semantic relational strategies to improve true memory. In contrast, patients showed the same pattern as healthy controls (ie, increase in false memory) when the implicit semantic strength of the word lists was increased.
The results imply that the automatic spread of activation within the semantic network is intact in schizophrenia patients42 and also that the ability to encode semantic knowledge that is spared in schizophrenia appears sufficient to support robust LOP effects.39 The results are further consistent with the previous findings that showed that schizophrenia patients appear to have a fundamental deficit in their ability to encode and process relational information of episodic knowledge.
Neuroimaging findings for the LOP effects
Heckers et al43 attempted to dissociate the dysfunctions in the DLPFC and HI involved in the memory impairment of schizophrenia patients using positron emission tomography (PET). As the usual LOP paradigm, perceptual encoding (counting the number of T-junctions in the letters of each word) and semantic encoding (counting the number of meanings for each word) were compared. During test session, the subjects were asked to complete three-letter word stems (eg, DRA__) of words presented either shallowly encoded or semantically encoded. Their hypothesis was that activation of the PFC would reflect the effort of retrieval and that HI activation would be observed during successful memory retrieval.
The PET data from healthy subjects showed that recollection of semantically encoded words was associated with right hippocampal activation, reflecting a conscious recollective process, while recollection of perceptually encoded words was associated with PFC activation (Brodmann area [BA] 8), reflecting an effortful attempt to retrieve memory.44
Schizophrenia patients showed robust activations in several prefrontal areas including the right parietal cortex (BA 7), right prefrontal areas (BA 9, 10, and 11), and right thalamus during the recollection of shallowly encoded words. The more widespread activation of prefrontal areas, compared to healthy controls, might reflect the patient’s greater effort through the experiment. The researchers suggest that the patients may be engaging a strategy that employs structures distinct from those commonly activated by healthy controls. The patients did not show activation in HI during conscious recollection. Instead, they showed significant activation in the extrahippocampal areas associated with attention (BA 40) and retrieval effort (BA 10), which might reflect the effort for compensating the failed recruitment of the HI.43 This study provided the first evidence of impaired hippocampal function in schizophrenia during episodic memory retrieval. The researchers suggest that a failure to modulate hippocampal activity based on environmental contingencies is connected to abnormal recruitment during conscious recollection and also that hippocampal dysfunction in schizophrenia might also be involved in the occurrence of psychotic symptoms contributing to poor prognosis and inadequate response to treatment.43
Neuroimaging data of LOP effects were also found with nonverbal materials. Bonner-Jackson et al38 examined the subregions of the brain involved in encoding deep information (abstract vs concrete judgments for the words presented), shallow information (“Does the first or last letter of the word come first in the alphabet?”), and face (sex judgments) employing fMRI. The behavioral results supported the previous findings of LOP effects, confirming deep encoding as an efficient encoding strategy in both healthy subjects and in those with schizophrenia. During encoding and retrieving the face stimuli, healthy subjects made more correct judgments than subjects with schizophrenia.
PFC activation occurred during deep encoding both in both healthy subjects and those with schizophrenia.45,46 Schizophrenia patients also showed robust activation in other brain regions associated with deep encoding, including the left inferior frontal (BA 45), right inferior frontal (BA 45), and left middle frontal gyrus (BA 10).47–49 The researchers claim that this additional activation reflects that schizophrenia patients need additional regions of the PFC to process deep encoding strategies, whereas healthy subjects need a relatively smaller subset of the left prefrontal regions.38
Brain activation differed depending on the material type used during encoding. Activation in the left PFC regions, and left temporal and parietal regions occurred during encoding words rather than faces, in both schizophrenia and healthy subjects. The patients also exhibited greater activation in the right PFC regions during word encoding compared to face encoding.38 In this study, the authors provided further evidence supporting the fact that patients with schizophrenia can benefit from advantageous encoding conditions.
Ragland et al39 tested the hypothesis that frontotemporal abnormalities in schizophrenia may be caused by the failure in encoding semantic information using fMRI. That is, their main interest was examining either whether the use of semantic strategy could provide not only a cognitive benefit, such as facilitating successful retrieval, but also a neurological benefit, such as normalizing brain function during encoding, or whether there exist residual abnormalities that cannot be sufficiently explained by encoding strategies.
The behavioral results replicated the LOP effects in both schizophrenia and healthy groups. The neuroimaging results showed that semantic encoding accompanied normalized frontotemporal function in both groups. It was observed that schizophrenia patients could engage the left DLPFC (BA 47) during semantic processing, indicating that there was no group difference in the left DLPFC. However, residual patterns of temporal-limbic overactivation during encoding and less robust right anterior prefrontal activation during retrieval in patients suggest that schizophrenia patients have other processing challenges during encoding and retrieval that are independent of top–down frontally mediated semantic control. In addition, a novel finding in the study was that hippocampal overactivation occurred during encoding in spite of unimpaired left prefrontal response in the schizophrenia group, which suggests that hippocampal hyperactivity may be separate from top–down prefrontal control.39
This finding added to the growing evidence that patients with schizophrenia can form and maintain semantic representations when they are provided with organizational cues and further that left hemisphere overactivation during encoding is associated with a residual disturbance in distributed mnemonic or semantic processing in schizophrenia.39
Taken together, the behavioral and neuroimaging findings we reviewed establish the behavioral and neural basis of episodic memory impairment in schizophrenia. The findings suggest that the RISE, AIP, and LOP paradigms can successfully detect functionally and neuroanatomically specific deficits in relational memory processes and related DLPFC and HI dysfunctions in schizophrenia across different experimental methods and imaging environments.
Behavioral findings for associative vs item-specific episodic memory impairment in patients with PTSD
It has been shown that nontrauma-related episodic memory impairment is as severe as trauma-related memory impairment in PTSD.50 A recent meta-analysis of memory impairment in PTSD patients concluded that there exists robust memory impairment of emotionally neural information in PTSD patients.51 Specifically, they found that impairment of verbal memory was more severe than that of visual memory. However, the specific locus of episodic memory deficits in PTSD patients has not been clarified yet.
Guez et al13 assumed that the propensity to make inappropriate associations would be related to fundamental deficits in associative memory in PTSD. As we explained in the introduction, the tendency to make inappropriate associations might be responsible for the symptom to overgeneralize from traumatic events to emotionally neutral materials or for intrusive thoughts to be triggered by very different contexts from the traumatic situations.9
Guez et al13 examined associative and item-specific memory processing in PTSD and healthy subjects. In addition, they included patients with acute stress disorder (ASD) to examine the effect of acute vs chronic traumatic stress on episodic memory impairment and to rule out methodological issues, such as comorbidity, variation in time elapsed after the trauma, and medications. Only ASD patients with no comorbidity, a relatively short or homogeneous time since the traumatic event, and no medical treatment were included.
All the subjects learned pairs of unrelated emotionally neutral words or pictures, and they were tested on both item-specific and associative recognition tasks. The results showed that both chronic PTSD and ASD patients were disproportionately impaired in associative recognition, whereas item recognition seemed to be intact. The recognition accuracy was obtained by hit–false alarm scores, where a hit was defined as responding yes to a target and a false alarm was defined as responding yes to a lure. The large decline in associative recognition seemed to reflect a relative increase in false alarms when erroneously responding to rearranged pairs.
Guez et al12 further tested the time course of associative memory impairment with ASD patients. Their main interest was whether impairment in associative memory could be resolved along with an improvement in posttraumatic symptoms. Patients with ASD and healthy subjects were tested using the same procedure as in their previous study, the only exception being that they were tested twice: once within 2 weeks of the traumatic event and again at 10–12 weeks after the event. They used the Clinician-Administered PTSD Scale (CAPS) score to assess the presence of posttraumatic stress symptoms.
There was a significant decrease in the total CAPS score. The ASD patients showed more impaired performance in associative memory relative to item memory, regardless of whether the stimuli were words or pictures. The key finding was that the impairment in associative memory diminished with the time lag, only for the verbal materials. After 10–12 weeks, performance in associative memory was improved for the verbal material, but it remained impaired for the visual stimuli. This result may imply the answer to whether the impairment in associative memory would reflect a preexisting cognitive vulnerability to ASD or be a consequence of it. The key is at the nature of stimuli. Whereas a deficit in verbal processing is symptom-relevant and thus may be considered as a consequence of ASD, a deficit in visual processing is symptom-irrelevant and may be considered as a predisposition to ASD.12
To summarize, these studies emphasized that traumatic stress is linked to a deficit in associative memory by demonstrating that abnormal associations observed in PTSD patients may stem from their tendency to falsely associate unrelated stimuli. They further demonstrated that associative memory impairment in ASD patients could be resolved with symptom resolution for verbal pairs but not for visual ones.
Neuroimaging findings for associative vs item-specific episodic memory impairment in PTSD
Although many studies have examined memory impairment in PTSD, no neuroimaging study has yet directly compared associative vs item-specific memory impairment in PTSD. Therefore, we have only reviewed neuroimaging evidence of associative memory, focusing on the findings of Geuze et al52 and Shin et al.53
Geuze et al52 examined the neural correlates of associative memory in veterans with PTSD using fMRI. Unrelated word pairs were presented during an encoding block, alternating with a control block in which two successive figures were presented. The subjects were required to memorize the word pairs during encoding and to repeat the two figures during the control block to minimize rehearsal. For the retrieval procedure, the first word in each pair was presented and the subjects were required to recall the second word and to repeat one of the two figures presented during the control block.
The behavioral results showed that veterans with PTSD (the PTSD group) were impaired in associative memory compared to veterans without PTSD (the control group). The fMRI data exhibited that during encoding the PTSD group showed underactivation in the bilateral inferior frontal gyri and left PFC, whereas they showed overactivation in the temporal cortex compared to the control group. In the retrieval results, the PTSD group exhibited underactivation in the right frontal cortex, the bilateral middle temporal gyri, and the left posterior HI/parahippocampal gyrus compared to the control group. These neuroimaging results provided the first fMRI evidence of subregions in brain associated with associative memory impairment in patients with PTSD.52
In general, findings of activity in both the frontal lobe and temporal lobe reflect that the frontal cortex is strongly connected to the HI.54–56 Sweatt’s study further provides the evidence supporting the fact that both the HI and PFC are responsible for associative memory in PTSD.57
Shin et al53 used PET to examine the relationship between memory deficits and hippocampal function in PTSD. Firefighters with PTSD (the PTSD group) and firefighters without PTSD (the control group) were required to encode non-emotional words presented during deep and shallow encoding. During the PET scanning session, all the subjects were required to complete a word stem task, in which subjects were given three-letter word stems and asked to complete each stem with a word that they had previously encoded either deeply or shallowly.
The PET data showed that the PTSD group revealed a slight increase in regional cerebral blood flow (rCBF) in the left HI during deep vs shallow encoding compared to the control group. The finding stemmed from relatively elevated rCBF during shallow encoding in the PTSD group. In addition, collapsing across deep and shallow encoding, the PTSD group revealed an elevated rCBF in bilateral HI and left amygdala compared to the control group. The authors found that symptom severity was positively correlated with rCBF in the HI and parahippocampal gyrus.53 They further demonstrated that the PTSD group had significantly smaller right hippocampal volumes relative to the control group. In summary, the researchers suggest that firefighters with PTSD exhibited an abnormal rCBF response in the HI during conscious recollection of non-emotional material and also that this abnormal response appears to be driven by relatively elevated hippocampal rCBF.
The results we just reviewed emphasized the role of both the HI and the PFC in associative memory in PTSD. Future research should be conducted to elucidate associative vs item-specific memory impairment in PTSD with various experimental methods, imaging environments, and survivor groups.
Conclusion
We reviewed the evidence of disproportionate relational memory impairment in people with schizophrenia and PTSD. Several tasks proposed by the CNTRICS initiative and the CNTRACS Consortium have played an important role as reliable and valid measures of item-specific and relational memory in schizophrenia. Experiments using these tasks have provided consistent evidence that patients with schizophrenia are disproportionately impaired in relational memory compared to healthy control subjects. Research on PTSD has also consistently reported that a disproportionate impairment in relational memory is observed in patients with PTSD. In addition, we reviewed the neural correlates and patterns of abnormal brain activation of the selective deficit in relational memory in patients with schizophrenia and PTSD. There has been consistent evidence supporting the fact that PFC and HI are critical for processing relational memory.
Given the relative scarcity of experimental studies of associative memory in PTSD and, in particular, taking into account the theoretical and therapeutic importance of the disorder, considerable further work is required. Recent research has reported that the number of veterans with PTSD has increased at a greater rate since 2005 than the number of veterans with other mental disorders.58 Anyone who has experienced a traumatic event, on the battlefield or elsewhere, can develop PTSD. That is why research into associative memory disorder in PTSD is important. It also could be a key to resolving the PTSD symptoms such as intrusive and distressing memories or thoughts, which are thought to be based on the tendency of aberrant associations.
Some studies have demonstrated that patients with schizophrenia have surprisingly intact abilities, such as semantic memory and the LOP effects. It would be helpful to scrutinize which ability is impaired or intact in schizophrenia in order to improve therapeutic interventions. This type of research should also be extended to other psychiatric disorders, including PTSD.
Acknowledgments
This research was supported by the Brain Research Program of the National Research Foundation of Korea funded by the Ministry of Science, ICT, and Future Planning (NRF-2015M3C7A1028252) and by a grant from the Korea Science and Engineering Foundation funded by the Korean government (NRF-2015R1A2A2A).
Disclosure
The authors report no conflicts of interests in this work.
References
Ranganath C, Minzenberg MJ, Ragland JD. The cognitive neuroscience of memory function and dysfunction in schizophrenia. Biol Psychiatry. 2008;64(1):18–25. | ||
Tulving E. Episodic and semantic memory. In: Tulving E, Donaldson W, editors. Organization of Memory. New York: Academic Press; 1972: 381–403. | ||
Green MF. What are the functional consequences of neurocognitive deficits in schizophrenia? Am J Psychiatry. 1996;153(3):321–330. | ||
Milev P, Ho BC, Arndt S, Andreasen NC. Predictive values of neurocognition and negative symptoms on functional outcome in schizophrenia: a longitudinal first-episode study with 7-year follow-up. Am J Psychiatry. 2005;162(3):495–506. | ||
Aleman A, Hijman R, de Haan EH, Kahn RS. Memory impairment in schizophrenia: a meta-analysis. Am J Psychiatry. 1999;156(9):1358–1366. | ||
Cirillo MA, Seidman LJ. Verbal declarative memory dysfunction in schizophrenia: from clinical assessment to genetics and brain mechanisms. Neuropsychol Rev. 2003;13(2):43–77. | ||
Heinrichs RW, Zakzanis KK. Neurocognitive deficit in schizophrenia: a quantitative review of the evidence. Neuropsychology. 1998;12(3):426–445. | ||
Ehlers A, Hackmann A, Michael T. Intrusive re-experiencing in post-traumatic stress disorder: phenomenology, theory, and therapy. Memory. 2004;12(4):403–415. | ||
Ehlers A, Clark DM. A cognitive model of posttraumatic stress disorder. Behav Res Ther. 2000;38(4):319–345. | ||
Wegerer M, Blechert J, Kerschbaum H, Wilhelm FH. Relationship between fear conditionability and aversive memories: evidence from a novel conditioned-intrusion paradigm. PLoS One. 2013;8(11):e79025. | ||
Ehlers A, Clark DM. Post-traumatic stress disorder: the development of effective psychological treatments. Nord J Psychiatry. 2008;62:11–18. | ||
Guez J, Cohen J, Naveh-Benjamin M, et al. Associative memory impairment in acute stress disorder: characteristics and time course. Psychiatry Res. 2013;209(3):479–484. | ||
Guez J, Naveh-Benjamin M, Yankovsky Y, Cohen J, Shiber A, Shalev H. Traumatic stress is linked to a deficit in associative episodic memory. J Trauma Stress. 2011;24(3):260–267. | ||
Ragland JD, Cools R, Frank M, et al. CNTRICS final task selection: long-term memory. Schizophr Bull. 2009;35(1):197–212. | ||
Ragland JD, Ranganath C, Barch DM, et al. Relational and Item-Specific Encoding (RISE): task development and psychometric characteristics. Schizophr Bull. 2012;38(1):114–124. | ||
Armstrong K, Kose S, Williams L, Woolard A, Heckers S. Impaired associative inference in patients with schizophrenia. Schizophr Bull. 2012;38(3):622–629. | ||
Armstrong K, Williams LE, Heckers S. Revised associative inference paradigm confirms relational memory impairment in schizophrenia. Neuropsychology. 2012;26(4):451–458. | ||
Ragland JD, Moelter ST, McGrath C, et al. Levels-of-processing effect on word recognition in schizophrenia. Biol Psychiatry. 2003;54(11):1154–1161. | ||
Hunt RR, Einstein GO. Relational and item-specific information in memory. J Verbal Learning Verbal Behav. 1981;20:497–514. | ||
Hunt RR, McDaniel MA. The enigma of organization and distinctiveness. J Mem Lang. 1993;32:421–445. | ||
Bower GH. Organizational factors in memory. Cogn Psychol. 1970;1:18–46. | ||
Tulving E, Thomson DM. Encoding specificity and retrieval processes. Psychol Rev. 1973;80(5):352–373. | ||
Ragland JD, Ranganath C, Harms MP, et al. Functional and neuroanatomic specificity of episodic memory dysfunction in schizophrenia: a functional magnetic resonance imaging study of the relational and item-specific encoding task. JAMA Psychiatry. 2015;72(9):909–916. | ||
Gardiner JM. Functional aspects of recollective experience. Mem Cognit. 1988;16(4):309–313. | ||
Yonelinas AP. The nature of recollection and familiarity: a review of 30 years of research. J Mem Lang. 2002;46(3):441–517. | ||
Blumenfeld RS, Ranganath C. Prefrontal cortex and long-term memory encoding: an integrative review of findings from neuropsychology and neuroimaging. Neuroscientist. 2007;13(3):280–291. | ||
Race E, Keane MM, Verfaellie M. Medial temporal lobe damage causes deficits in episodic memory and episodic future thinking not attributable to deficits in narrative construction. J Neurosci. 2011;31(28):10262–10269. | ||
Blumenfeld RS, Ranganath C. Dorsolateral prefrontal cortex promotes long-term memory formation through its role in working memory organization. J Neurosci. 2006;26(3):916–925. | ||
Luck SJ, Gold JM. The construct of attention in schizophrenia. Biol Psychiatry. 2008;64(1):34–39. | ||
Chua EF, Schacter DL, Rand-Giovannetti E, Sperling RA. Evidence for a specific role of the anterior hippocampal region in successful associative encoding. Hippocampus. 2007;17(11):1071–1080. | ||
Giovanello KS, Schnyer DM, Verfaellie M. A critical role for the anterior hippocampus in relational memory: evidence from an fMRI study comparing associative and item recognition. Hippocampus. 2004;14(1):5–8. | ||
Heckers S, Zalesak M, Weiss AP, Ditman T, Titone D. Hippocampal activation during transitive inference in humans. Hippocampus. 2004;14(2):153–162. | ||
Jackson O 3rd, Schacter DL. Encoding activity in anterior medial temporal lobe supports subsequent associative recognition. Neuroimage. 2004;21(1):456–462. | ||
Kirwan CB, Stark CE. Medial temporal lobe activation during encoding and retrieval of novel face-name pairs. Hippocampus. 2004;14(7):919–930. | ||
Preston AR, Shrager Y, Dudukovic NM, Gabrieli JD. Hippocampal contribution to the novel use of relational information in declarative memory. Hippocampus. 2004;14(2):148–152. | ||
Staresina BP, Davachi L. Selective and shared contributions of the hippocampus and perirhinal cortex to episodic item and associative encoding. J Cogn Neurosci. 2008;20(8):1478–1489. | ||
Craik FI, Lockhart RS. Levels of processing: a framework for memory research. J Verbal Learning Verbal Behav. 1972;11(6):671–684. | ||
Bonner-Jackson A, Haut K, Csernansky JG, Barch DM. The influence of encoding strategy on episodic memory and cortical activity in schizophrenia. Biol Psychiatry. 2005;58(1):47–55. | ||
Ragland JD, Gur RC, Valdez JN, et al. Levels-of-processing effect on frontotemporal function in schizophrenia during word encoding and recognition. Am J Psychiatry. 2005;162(10):1840–1848. | ||
Roediger HL, McDermott KB. Creating false memories: remembering words not presented in lists. J Exp Psychol Learn Mem Cogn. 1995;21(4):803–814. | ||
McDermott K, Roediger HL. Attempting to avoid illusory memories: robust false recognition of associates persists under conditions of explicit warnings and immediate testing. J Mem Lang. 1998;39:508–520. | ||
Paz-Alonso PM, Ghetti S, Ramsay I, et al. Semantic processes leading to true and false memory formation in schizophrenia. Schizophr Res. 2013;147(2–3):320–325. | ||
Heckers S, Rauch SL, Goff D, et al. Impaired recruitment of the hippocampus during conscious recollection in schizophrenia. Nat Neurosci. 1998;1(4):318–323. | ||
Schacter DL, Alpert NM, Savage CR, Rauch SL, Albert MS. Conscious recollection and the human hippocampal formation: evidence from positron emission tomography. Proc Natl Acad Sci U S A. 1996;93(1):321–325. | ||
Buckner RL, Wheeler ME. The cognitive neuroscience of remembering. Nat Rev Neurosci. 2001;2(9):624–634. | ||
Wagner AD, Schacter DL, Rotte M, et al. Building memories: remembering and forgetting of verbal experiences as predicted by brain activity. Science. 1998;281(5380):1188–1191. | ||
Fletcher PC, Stephenson CM, Carpenter TA, Donovan T, Bullmorel ET. Regional brain activations predicting subsequent memory success: an event-related fMRI study of the influence of encoding tasks. Cortex. 2003;39(4–5):1009–1026. | ||
Kapur S, Craik FI, Tulving E, Wilson AA, Houle S, Brown GM. Neuroanatomical correlates of encoding in episodic memory: levels of processing effect. Proc Natl Acad Sci U S A. 1994;91(6):2008–2011. | ||
Otten LJ, Henson RN, Rugg MD. Depth of processing effects on neural correlates of memory encoding: relationship between findings from across- and within-task comparisons. Brain. 2001;124(Pt 2):399–412. | ||
Wolfe J, Charney DS. Use of neuropsychological assessment in posttraumatic stress disorder. Psychol Assessment. 1991;3:573–580. | ||
Brewin CR, Kleiner JS, Vasterling JJ, Field AP. Memory for emotionally neutral information in posttraumatic stress disorder: a meta-analytic investigation. J Abnorm Psychol. 2007;116(3):448–463. | ||
Geuze E, Vermetten E, Ruf M, de Kloet CS, Westenberg HG. Neural correlates of associative learning and memory in veterans with posttraumatic stress disorder. J Psychiatr Res. 2008;42(8):659–669. | ||
Shin LM, Shin PS, Heckers S, et al. Hippocampal function in posttraumatic stress disorder. Hippocampus. 2004;14(3):292–300. | ||
Squire LR, Knowlton B, Musen G. The structure and organization of memory. Annu Rev Psychol. 1993;44:453–495. | ||
Taylor JG, Horwitz B, Shah NJ, Fellenz WA, Mueller-Gaertner HW, Krause JB. Decomposing memory: functional assignments and brain traffic in paired word associate learning. Neural Netw. 2000;13:923–940. | ||
Rolls E. Memory systems in the brain. Annu Rev Psychol. 2000;51:599–630. | ||
Sweatt JD. Hippocampal function in cognition. Psychopharmacolog (Berl). 2004;174:99–110. | ||
Hermes ED, Rosenheck RA, Desai R, Fontana AF. Recent trends in the treatment of posttraumatic stress disorder and other mental disorders in the VHA. Psychiatr Serv. 2012;63(5):471–476. |
 © 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.