Back to Journals » Drug Design, Development and Therapy » Volume 20
Medicinal Plants That Modulate Nitric Oxide Synthase Activity: Implications in Inflammation and Oxidative Stress
Authors Prayoga DK, Aulifa DL ![]() , Budiman A, Jiranusornkul S, Levita J
, Budiman A, Jiranusornkul S, Levita J ![]()
Received 18 November 2025
Accepted for publication 19 February 2026
Published 13 March 2026 Volume 2026:20 582323
DOI https://doi.org/10.2147/DDDT.S582323
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Anastasios Lymperopoulos
Deshanda Kurniawan Prayoga,1,* Diah Lia Aulifa,2,* Arif Budiman,3,* Supat Jiranusornkul,4,* Jutti Levita5,*
1Doctoral Program in Pharmacy, Faculty of Pharmacy, Universitas Padjadjaran, Sumedang, West Java, Indonesia; 2Department of Pharmaceutical Analysis and Medicinal Chemistry, Faculty of Pharmacy, Universitas Padjadjaran, Sumedang, West Java, Indonesia; 3Department of Pharmaceutics and Pharmaceutical Technology, Faculty of Pharmacy, Universitas Padjadjaran, Sumedang, West Java, Indonesia; 4Department of Pharmaceutical Science, Faculty of Pharmacy, Chiang Mai University, Chiang Mai, Thailand; 5Department of Pharmacology and Clinical Pharmacy, Faculty of Pharmacy, Universitas Padjadjaran, Sumedang, West Java, Indonesia
*These authors contributed equally to this work
Correspondence: Deshanda Kurniawan Prayoga; Jutti Levita, Email [email protected]; [email protected]
Abstract: Nitric oxide (NO) plays a central role in diverse physiological processes, such as cardiovascular tone, neurotransmission, immune defense, and cellular apoptosis. NO synthase (NOS) is the enzyme that catalyzes the conversion of L-arginine to L-citrulline and NO. Three isoforms of NOS are (1) neuronal NOS (nNOS), which regulates synaptic plasticity, memory formation, and cerebral blood flow, (2) endothelial NOS (eNOS), which supports cardiovascular homeostasis by preventing platelet aggregation and leukocyte adhesion, and (3) inducible NOS (iNOS), which contributes to inflammation and tissue damage, when in excessive production. Thus, targeting iNOS without interfering with the beneficial actions of nNOS and eNOS remains a major therapeutic challenge. Given this dual nature of NO in health and disease, it is important to understand how medicinal plants and their phytochemicals modulate NO pathways. A systematic search of Scopus and PubMed databases was performed for studies published in the last ten years. Screening by title, abstract, and full text yielded 35 eligible articles examining medicinal plants and phytochemicals that influence NO signaling pathways. Preclinical evidence indicates that phytochemicals restore antioxidant defenses and suppress excessive NO and oxidative stress under inflammatory conditions while preserving or enhancing endothelial NO bioavailability. Emerging clinical findings further suggest significantly greater insulin-stimulated NO production and eNOS activation without activating iNOS or promoting inflammatory responses. Their ability to regulate NO production underscores their potential in the development of botanical drugs targeting iNOS while preserving the physiological roles of nNOS and eNOS. This review provides an overview of the potential of medicinal plants to alleviate inflammation and oxidative stress through modulation of NO production.
Keywords: clinical study, iNOS, medicinal plants, NOS, phytochemicals
Introduction
Nitric oxide (NO) is an important and versatile neurotransmitter crucial in various physiological processes. It is synthesized from the amino acid L-arginine by nitric oxide synthase (NOS), a heme-containing monooxygenase.1,2 Three distinct isoforms of NOS have been identified: neuronal NOS (nNOS), endothelial NOS (eNOS), and inducible NOS (iNOS).3–5 Among these, nNOS and eNOS are constantly expressed in endothelial cells and neurons, respectively, producing small amounts of NO in a calcium/calmodulin (Ca2+/CaM)-dependent manner.6,7 In contrast, iNOS is not continuously expressed, while it is induced under specific conditions for a rapid action, such as exposure to bacterial lipopolysaccharide (LPS), and generates NO in much higher concentrations, approximately 100–1000-fold, for prolonged periods.8,9
The physiological roles of NO depend on its source. NO derived from nNOS supports neuronal plasticity, memory formation, and regulation of cerebral blood flow,10 while NO derived from eNOS contributes to cardiovascular homeostasis by inhibiting platelet aggregation, leukocyte adhesion, and migration.4,11 However, excessive or dysregulated NO production, particularly through iNOS, leads to harmful effects. Overexpression or uncontrolled activation of iNOS is associated with inflammation, tissue injury, and neurodegenerative disorders.6,8,12,13 For example, during chronic inflammation, excessive NO contributes to tissue damage in arthritis and respiratory diseases.14,15 Additionally, oxidative stress, which occurs when there is an imbalance between the excessive production of reactive oxygen species (ROS) and the insufficient amount of endogenous antioxidant enzymes, can affect the function of NO by reducing its availability or promoting its inactivation.16,17 Among inflammatory mediators, cytokines, chemokines, and transcription factors primarily regulate discrete upstream signaling events that reflect only specific aspects of the inflammatory cascade.18,19 In contrast, NO lies downstream of these pathways, as the pro-inflammatory mediator iNOS, because this enzyme is activated by pro-inflammatory cytokines like tumor necrosis factor-alpha (TNF-α), interleukin-1 beta (IL-1β), and interferon-gamma (IFN-γ), and a hallmark of M1 macrophage activation, leading to sustained NO production that directly contributes to oxidative stress and tissue injury. At the same time, constitutive NO signaling maintains endothelial integrity, vascular tone, and immune homeostasis. This dual and context-dependent role positions NO as an integrative regulator of both inflammatory damage and physiological protection, making its modulation a more comprehensive therapeutic target than individual inflammatory mediators.20 While NO plays a crucial protective role, the dysregulation of its production due to oxidative stress can worsen the progression of various diseases, emphasizing the complex and multifaceted nature of NO in human health and disease.21
A major challenge in the development of synthetic iNOS inhibitors is the limited clinical translation of promising preclinical findings, as the known iNOS inhibitors (1400W, N-3-aminomethylbenzyl acetamidine; GW274150, 1-iminoethyl aminoethyl-L-homocysteine; L-NIL, L-Nω-1-iminoethyl lysine; and GW273629, 3-2-1-iminoethyl amino ethyl sulphonyl-L-alanine) have not yet been approved for human use.8 Beyond methodological and assay limitations, the context-dependent and dual roles of both iNOS and NO in disease processes, together with the distinct functions and tissue distributions of NOS isoforms, complicate the therapeutic targeting.22 In oncology, for example, iNOS overexpression may promote tumor progression. Yet, NO also enhances chemosensitivity, creates a therapeutic paradox, and limits the safety of a broad NOS inhibition. These limitations highlight the need for alternative strategies that can modulate NO signaling without interfering with its inhibition.22–24 Considering these perspectives, medicinal plants provide an interesting opportunity. Long before synthetic drugs were manufactured, folklore communities used natural resources to maintain health and cure illness, and these remedies remain vital in many regions. The World Health Organization estimated that more than 80% of individuals in developing countries use traditional medicine.25,26
Medicinal plants and functional foods are already recognized for treating diseases such as wounds, inflammation, diabetes, and even cancer. In particular, inflammation is strongly associated with the overproduction of NO.27,28 Activated microglia in neuronal cells and activated macrophages during non-neuronal inflammation are key contributors to this increase, which has been identified as both a biomarker and a causative factor in secondary tissue damage.29–31 The therapeutic effects of medicinal plant extracts and their bioactive compounds are often attributed to their ability to inhibit the activity of iNOS or their free radical scavenging properties. Nitric oxide and singlet oxygen (1O2) are potent free radicals involved in lipid peroxidation, a key process in the pathogenesis of chronic inflammation.27,32 Recent studies have highlighted the potential of plant extracts to modulate NO synthesis and release in LPS-exposed cells.33–35 Accumulated evidence across experimental systems has shown that many medicinal plants regulate NO signaling through diverse mechanisms. For example, an in vitro study in LPS‐exposed RAW 264.7 macrophages showed that Amomum tsao-ko of family Zingiberaceae exerts dual biological actions by (1) inhibiting iNOS expression and (2) enhancing heme oxygenase‐1 (HO‐1) expression at the protein and mRNA levels. Intriguingly, a further study in LPS‐induced endotoxaemic mice indicated that pretreatment with the extract decreased NO serum levels and liver iNOS expression, and increased HO‐1 expression and survival rates.36 Clinical observation further supports this pathway, as preparations from Curcuma longa have been reported to normalize NO levels in inflammatory conditions. Collectively, these findings provide promising evidence for their anti-inflammatory effects.37
Given this dual nature of NO in health and disease, understanding how medicinal plants and their phytochemicals modulate NO pathways is of considerable interest. This review provides a comprehensive analysis of the effects of medicinal plants on NO production, with particular emphasis on their roles in modulating immune responses, regulating oxidative stress, and inhibiting pro-inflammatory cytokines. This review thoroughly investigates how bioactive compounds derived from medicinal plants influence the NO pathway and their potential in managing oxidative stress-induced inflammation. Furthermore, the review assesses the clinical potential of medicinal plants as complementary or alternative therapies for diseases characterized by excessive inflammation and oxidative stress. It also highlights the relationship between medicinal plants and NO production, and identifies key research gaps and future directions for studying medicinal plants and NO regulation. This emphasizes the need for more rigorous clinical trials and the development of innovative plant-based therapeutics.
Methods
Search Strategy
Initially, the search included references published from 2015 to 2025. Articles were searched in Scopus and PubMed databases by using the keywords medicinal plants, inhibit, suppress, nitric oxide production, nitric oxide synthase, preclinical studies, and clinical trials. Nonetheless, additional phrases relating to topics of interest were also employed. Throughout the search, synonyms and related terms were used interchangeably as the keywords. Boolean operators such as “AND” and “OR” were employed to refine the search and include or exclude specific terms. Specifically, the PubMed and Scopus databases were searched until July 2025. The entire search strategy utilized a combination of medical subject heading terms, as stated in Table S1. The documents that met the inclusion criteria focus on plants that specifically affect NO production, rather than just general effects on ROS or antioxidant activity. Finally, 35 articles were selected.
Data Extraction Process
The study selection process was conducted systematically in two phases following the removal of duplicate articles from the initial search results. In the first phase, the first author (DKP) independently screened the titles and abstracts of all identified records using the web-based tool Rayyan.ai (https://new.rayyan.ai/). Articles that met the initial criteria were then moved to the second phase for a thorough full-text assessment. To be eligible, articles needed to be preclinical studies (either in vitro or in vivo) or clinical studies published in English between 2015 and 2025. These studies were required to evaluate extracts or isolated compounds from medicinal plants and report outcomes related to NO production, specifically in the context of inflammatory conditions, oxidative stress, or immune response. Articles were excluded if they were reviews, commentaries, or if their primary focus was on reactive oxygen species (ROS) rather than NO production. Key data extracted from each article included: medicinal plant family, collection location and year, parts of the plant used, extraction method and solvent, classes of metabolites or specific phytoconstituents, cell line used, inducement method, and exposure time for in vitro studies; as well as animal species, negative and positive controls, and research duration for in vivo studies; and study type, control, and duration for clinical trials. Based on these criteria, a total of 30 articles were selected for in-depth analysis.
Data Synthesis
Due to the significant variability among the included studies and the differing outcome measures, we conducted a narrative synthesis of the evidence. The findings were qualitatively summarized and organized thematically based on the reported results. This synthesis includes dedicated sections on NO production and its underlying mechanisms in immune responses, nitrosative and pro-inflammatory signaling, to provide a more comprehensive context and address gaps for a thorough review. This holistic approach is a valuable resource for researchers, integrating biological context with a systematic review of the most recent evidence.
Characteristics of Included Studies
The characteristic of the included studies was systematically extracted with particular emphasis on plant-derived formulations and phytochemicals that modulate the nitric oxide synthase (NOS) pathway. Studies were classified into three primary categories, including in vitro, in vivo, and clinical investigations, to enable structured evaluation of evidence across experimental and translational contexts. For each study, we recorded descriptive parameters, including the study design, population or biological model, intervention and inducer, and the principal outcome assessed. This includes parameters such as levels of nitrate and nitrite in clinical studies, cellular NO output assessed through the Griess assay, and iNOS mRNA or protein levels in preclinical models.
Comprehensive tables were constructed to summarize these features, delineating both the type of study and the principal methodological approaches employed. This structured documentation served two purposes: (i) to enhance transparency regarding data extraction and synthesis, and (ii) to facilitate subsequent thematic analysis across biological domains such as immune regulation and nitrosative stress in NO production.
Results
NO plays a multifaceted role in the body, influencing both innate and adaptive immune responses, oxidative stress, and triggering the pro-inflammatory cytokines.6,38–40 Studies considered these plant-based extracts or phytochemicals work synergistically as inhibitors of NO production, which can protect against cardiovascular, gastrointestinal, and other diseases by modulating the immune responses, antioxidants, and inhibiting the pro-inflammatory cytokines. A total of 35 articles were included. Among these, 26 reported in vitro studies and 6 reported in vivo experiments. Four articles incorporated both in vitro and in vivo preclinical approaches, while 3 articles presented clinical studies. Of these studies, we found that many plants were described to have effects on NO production. Overall, many medicinal plants demonstrably influence NO biology, yet only a few have been translated into therapies explicitly aimed at inhibiting or elevating NO production. However, a limitation of this review is the predominance of preclinical studies, with only a small number of clinical investigations. This imbalance may limit the direct clinical applicability of the findings and highlights the need for further clinical validation.
In vitro Studies
In vitro experimental studies employed various cell line models to investigate, including RAW 264.7 macrophages, HaCaT keratinocytes, BV-2 microglial cells, EOC-20 mouse microglia cells, and IC-21 mouse macrophage cells. These models were commonly stimulated with LPS, a well-characterized bacterial endotoxin known for its potent ability to activate immune signaling pathways.40,41 LPS exposure leads to systemic release of pro-inflammatory cytokines such as TNF-α, IL-1β, IL-6, and IL-10, which are essential for pathogen clearance, induce free radicals, and immune responses.42
NO production is typically assessed within macrophage and endothelial cell systems using the Griess assay, a widely recognized method for the indirect quantification of NO through the spectrophotometric detection of nitrite (NO2−), a stable degradation product of NO autoxidation.43,44 The results were consistent with the pharmacological targets of the test compounds, such as agents directed toward iNOS activity generally reduced nitrite/NOx levels, while polyphenols directed toward endothelial cells frequently enhanced NO signaling or increased measurements of iNOS and NO.45
Beyond these mechanistic insights, findings from the in vitro studies illustrated that phytoconstituents exerted dual biological effects, significantly inhibited NO production while simultaneously enhancing antioxidant capacity to neutralize free radicals generated during LPS exposure (Table 1). Given that reactive oxygen species (ROS) serve as secondary messengers in macrophages, amplifying host defense, gene transcription, and apoptosis, excessive ROS production can lead to uncontrolled inflammatory cascades.46 Several studies reported that these natural compounds reduced the overproduction of pro-inflammatory cytokines in LPS-stimulated macrophages by downregulating protein and mRNA expression of TNF-α, IL-1β, IL-6, and IL-10.47–49 These cytokines exhibit potent chemotactic and vasoactive properties, with TNF-α playing a pivotal role in the inflammatory response. It stimulates further cytokine release, promotes angiogenesis, activates NF-κB signalling, and stimulates NO synthesis. Likewise, IL-1β works synergistically with TNF-α to induce fever, foster coagulation abnormalities, and perpetuate a sustained inflammatory cascade through autocrine and paracrine mechanisms.48,50–52 Overall, natural extracts enriched with these phytoconstituents consistently reduced iNOS expression and suppressed NO release in ROS 17/2.8 cells activated by a combination of cytokines, including TNF-α and IL-1β.53,54 These observations suggest that the data presented in Table 1, offering a clear overview of the in vitro studies of phytochemical extracts that regulate NO signaling and modulate oxidative stress and inflammatory pathways, underscoring their potential as therapeutic agents in inflammation management.
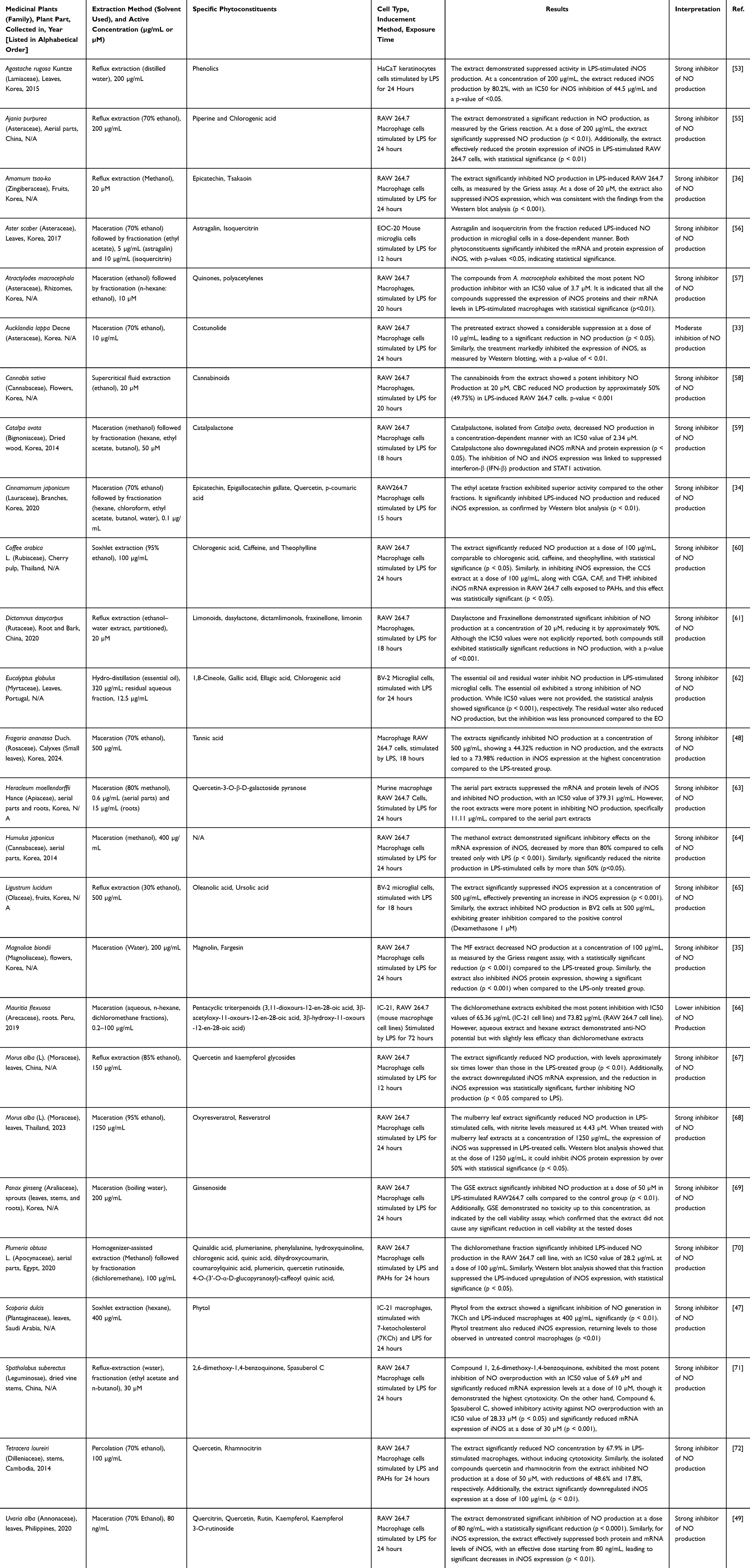 |
Table 1 Effects of Medicinal Plants on NO Production via in vitro Studies |
In vivo Studies
In vivo studies prove phytoconstituents can suppress NO production under inflammatory conditions. Murine models were most frequently employed, using inflammation induced by carrageenan, ethanol, LPS, or sodium nitroprusside (SNP) to induce systemic or localized inflammation. These models are well established for mimicking acute and chronic inflammatory responses, as they reliably trigger systemic overproduction of NO through upregulation of iNOS.73–75 However, treatment with phytochemical-rich extracts, fractions, or purified compounds consistently resulted in significant decreases in serum nitrite (NOx) and mRNA iNOS expression levels, indicating a direct inhibitory effect on NO synthesis.
Phytochemicals, particularly those belonging to the flavonoids, polyphenols, and terpenoids, demonstrated multi-targeted effects in these models. A notable example is the pre-treatment with the dichloromethane fraction (DCM-F) of Plumeria obtusa L., which produced apparent protective effects in LPS-challenged animals. Animals that received DCM-F before induction displayed markedly lower plasma NOx concentrations compared to untreated controls. Histological analysis of target organs such as the liver and lungs showed reduced leukocyte infiltration and attenuated tissue injury. DCM-F pre-treatment suppressed iNOS expression at both protein and mRNA levels, demonstrating that inhibition occurred at the transcriptional and translational stages. In parallel, cytokine analysis revealed downregulation of TNF-α, IL-1β, and IL-6. Thus, inhibition of NO production by phytochemicals was closely associated with a broader anti-inflammatory profile in vivo.70
One complementary line of evidence was provided by the carrageenan-induced paw edema model, where administration of phytoconstituents markedly reduced paw swelling and localized NO accumulation at the site of inflammation.58,76 These effects were further associated with restoration of endogenous antioxidant defences, including superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GPx).77 The restoration of redox balance highlights the dual role of these compounds; they suppress excessive NO synthesis and neutralize ROS that exacerbate inflammatory signaling. Several studies reported reduced lipid peroxidation, indicated by lower malondialdehyde (MDA) levels, suggesting protection against oxidative damage driven by nitrosative stress.76,78
Beyond acute inflammation, investigations in chronic or repeated exposure models demonstrated sustained decreases in NO levels and consistent downregulation of iNOS expression with prolonged treatment, indicating the durability of phytochemical interventions.79 These findings confirm that phytoconstituents act as broad modulators of NO pathways and inflammatory injury in vivo. They inhibit endogenous NO production through iNOS suppression while mitigating damage from exogenous NO overload.63 Significantly, decreases in oxidative stress markers such as MDA, alongside the restoration of reduced glutathione (GSH), demonstrated their capacity to maintain cellular homeostasis.40,74 These findings, summarized in Table 2, indicate that the whole extracts, fractions, and isolated compounds generally attenuate nitric oxide overproduction by downregulating iNOS expression and reducing tissue injury in vivo.
 |
Table 2 Effects of Medicinal Plants on NO Production via in vivo Studies |
Clinical Studies
Clinical studies, although fewer than preclinical investigations, provide crucial translational evidence that phytochemicals modulate NO pathways in humans. Unlike preclinical findings, where phytoconstituents predominantly inhibited NO production, clinical outcomes are more context-dependent, demonstrating both stimulation and suppression of NO. Investigations have been conducted in diverse populations, including COVID-19 patients, elderly subjects with metabolic disorders, and hypertensive patients.80–82 Collectively, these studies indicate that herbal mixtures and phytochemical-rich extracts can modulate NO activity (Table 3).
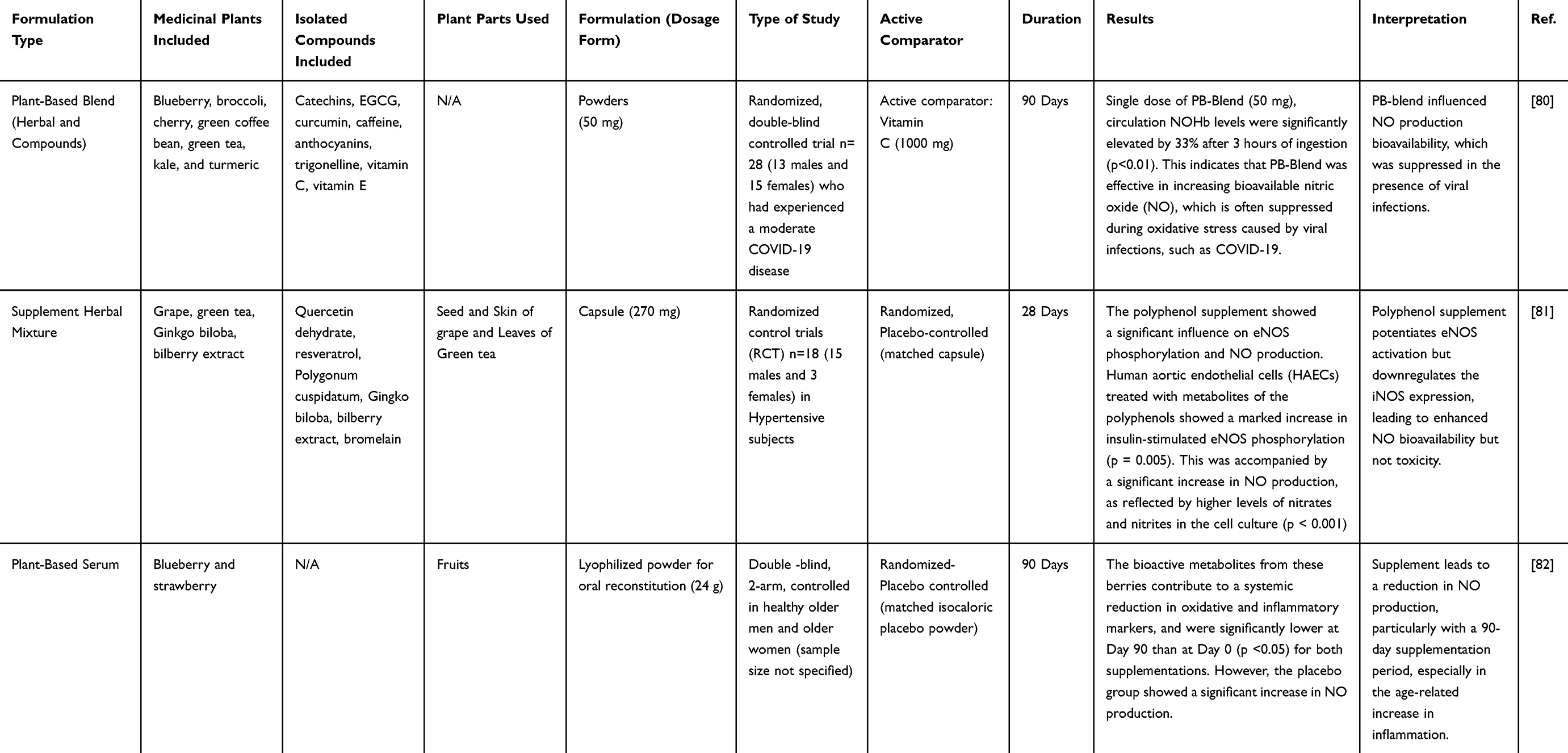 |
Table 3 Effects of Medicinal Plants on NO Production via Clinical Studies |
In several trials, supplementation with herbal formulations led to enhanced NO production, which was positively associated with improved endothelial function and immune defense. For instance, in COVID-19 patients, administering herbal mixtures traditionally used for respiratory illnesses increased circulating NO levels, a response thought to support antiviral mechanisms and vascular activity. While NO has a short half-life, it remains stable by binding to hemoglobin, forming NO-bound hemoglobin (NOHb). Therefore, measuring NOHb serves as a reliable surrogate marker for endothelial function.83 Clinical data showed that COVID-19 infection, through chronic overproduction of ROS, suppressed circulating NOHb and contributed to endothelial.80 These effects are linked to the cytokine storm, a mechanism also reported in influenza virus infections.84 A controlled trial comparing a single dose of PB-Blend to vitamin C supplementation analyzed NOHb concentrations and revealed consistently low baseline NOHb in both groups, supporting the role of ROS in depleting NO bioavailability. After supplementation, PB-Blend intake increased circulating NOHb, indicating improved endothelial activity and restoration of vascular health in subjects susceptible to oxidative stress during COVID-19 infection.80
Conversely, some clinical studies have reported reductions in NO levels following phytochemical interventions, especially in elderly populations with inflammatory or metabolic disorders.82 In a well-controlled trial, older adults received blueberry (BB) or strawberry (SB) supplementation for up to 90 days, with fasting (pre) and postprandial (post) serum samples collected and tested ex vivo. The effect was most pronounced after 90 days, with both diet groups showing a reduction in NO production after 90 days, suggesting that the absence of phytochemical supplementation could worsen LPS-driven responses. In addition to suppressing NO, serum from BB- and SB-supplemented individuals also decreased LPS-induced TNF-α release, with more pronounced effects observed in the BB. These results demonstrated that BB polyphenols, including anthocyanins, were absorbed and metabolized, producing different phenolic acid derivatives that may be contributing to the anti-inflammatory effects.82,85
In contrast, hypertensive individuals who consumed phytochemical supplementation exhibited yet another pattern, as it led to reductions in diastolic blood pressure along with significant increases in urinary nitrite and nitrate concentrations.81 These findings suggest that blood pressure reduction was linked to enhanced NO bioavailability, particularly through eNOS pathways. Mechanistic insights from studies using human aortic endothelial cells (HAECs) demonstrated that incubation with polyphenol metabolites significantly increased insulin-stimulated NO production and promoted eNOS phosphorylation.81,86–88 However, the angiotensin-converting enzyme (ACE) activity showed no significant differences between the intervention and control groups, while urinary nitrite/nitrate levels increased during supplementation.81 This supports the conclusion that enhanced NO bioavailability, rather than ACE inhibition, improved vascular function and reduced blood pressure. Overall, it is confirmed that polyphenol metabolites exhibited significantly greater insulin-stimulated NO production and eNOS activation without activating iNOS or promoting inflammatory responses, strengthening the evidence for a direct endothelial mechanism of action.
Immune Response to NO Production
NO plays a multifaceted role in the immune system, influencing both innate and adaptive responses.39,89 Its effects are complex and dual, including acting as a protective microbicidal agent against pathogens and serving as an immunoregulatory molecule that modulates the behavior of macrophages, dendritic cells, and T-lymphocytes.38,90 Through these mechanisms, NO helps eliminate invading microorganisms and shapes the balance of immune cell activation and differentiation.91,92 Due to its short half-life of only about five seconds, similar to its stability in an aqueous solution, NO must be generated rapidly and close to its target.6,93–95 This is why multiple isoforms of NOS exist, each expressed in different cell types and regulated by specific stimuli.
In addition to its signaling functions, NO contributes directly to host defense. At high concentrations, it exerts cytotoxic effects similar to ROS. Polymorphonuclear neutrophils (PMNs) release both NO and ROS to kill microbes.96,97 In neutrophils, the NADPH oxidase complex plays a central role. The membrane-bound subunits gp91phox (NOX2/CYBB) and p22phox (CYBA) combine with cytosolic components (p47phox, p67phox, p40phox) to form a functional oxidase complex upon activation.98–101 The GTP-binding protein Rac2 (Ras-related C3 botulinum toxin substrate 2) enhances this process, leading to the formation of robust superoxide or singlet oxygen (1O2).102–104 The reaction of 1O2 with NO can rapidly and site-specifically generate a nitrite pool through the pathways. In an iron-rich and oxidative setting, nitrite may also arise from NO interacting with hypervalent metal complexes. Under the acidic conditions of the phagosome, nitrite can be further oxidized to NO2, either directly or through protonation, providing antimicrobial activity.105,106
Macrophages are another primary source of NO. Classical M1 macrophages, activated by toll-like receptor (TLR) ligands or cytokines such as IFN-γ, express high levels of iNOS.39,107 The NO generated by iNOS is strongly pro-inflammatory and cytotoxic, enabling macrophages to inactivate or destroy infectious agents.108,109 Simultaneously, iNOS activity regulates the polarization of macrophages, influencing the M1/M2 balance through transcription factors such as IRF5 (Interferon regulatory factor 5).107,110 Interestingly, NO can also act as a negative regulator of immunity. Studies show that NO suppresses IL-12 production in dendritic cells and macrophages, thereby modulating Th1 responses. T cell-derived iNOS has also been implicated in limiting Th17 differentiation.110–112 Mechanistic evidence demonstrates that NO donors and the nitration of transcription factors like RORγt can inhibit IL-17 production, while iNOS-deficient T-cells lead to more severe colitis in animal models.113
Preclinical studies indicate that phytochemicals can regulate NO in immune cells. In vitro, LPS stimulation of macrophages or microglia induced strong upregulation of iNOS, NO release, and cytokine secretion (TNF-α, IL-1β, IL-6). Phytochemical extracts attenuated these responses by downregulating iNOS expression, reducing nitrite accumulation, and suppressing cytokine release.47–49 In vivo, plant fractions lowered systemic NOx levels, reduced leukocyte infiltration, and decreased circulating cytokines in models challenged with LPS or carrageenan.36,58 Clinical findings further support this evidence. In elderly subjects, long-term supplementation with blueberry or strawberry extracts reduced LPS-induced NO and TNF-α production in ex vivo microglial assays. These reductions correlated with decreased iNOS expression in immune cells and lower systemic pro-inflammatory cytokines, indicating improved immune regulation.85 Conversely, phytochemicals showed stimulatory effects in conditions where NO deficiency contributes to impaired host defense. In COVID-19 patients, herbal mixtures elevated circulating NO and increased hemoglobin-bound NO (NOHb), which partially restored impaired antiviral and endothelial responses.80 In hypertensive subjects, polyphenol supplementation enhanced eNOS phosphorylation and insulin-stimulated NO release, improving vascular function without triggering iNOS-driven inflammation.83
Nitrosative Stress in NO Production
Under normal physiological conditions, the balance between oxidative and nitrosative stress is tightly regulated by antioxidant enzymes and scavenging systems that neutralize ROS and reactive nitrogen species (RNS).114–116 Key endogenous defenses include the glutathione (GSH) and thioredoxin (Trx) systems, bolstered by selenoenzymes such as glutathione peroxidases (GPx) and thioredoxin reductases (TrxR). These pathways typically prevent cellular injury by managing damage caused by ROS and RNS.117,118 However, in chronic inflammatory states or diseases such as sickle cell disease (SCD), diabetes, and neurodegeneration, this balance is disrupted, resulting in excessive production of NO and RNS, particularly peroxynitrite (formed from the reaction of NO with superoxide).114,115
Nitrosative stress can damage proteins, lipids, and nucleic acids, impairing mitochondrial function by blocking components of the respiratory chain. The sensitivity of specific cell types to peroxynitrite varies based on their antioxidant capacity, with neurons tending to be more vulnerable than astrocytes. This vulnerability contributes to the role of nitrosative stress in neurological conditions such as Alzheimer’s disease, Parkinson’s disease, multiple sclerosis, Huntington’s disease, and ischemia-reperfusion injury.119,120 Protein tyrosine nitration is a crucial biochemical marker of this process; accumulation of nitrotyrosine has been observed in inflammatory diseases, including diabetic complications like retinopathy.121,122 In these conditions, factors such as vascular endothelial growth factor (VEGF) are upregulated, partly driven by ROS and NO, thereby connecting nitrosative stress to vascular pathology.123 Additionally, high-intensity exercise can lead to transient increases in ROS and RNS, unsettling the oxidant-antioxidant balance and potentially worsening cellular stress when antioxidant defenses are insufficient.124
Evidence from preclinical studies indicates that phytochemicals can help mitigate nitrosative stress in inflammatory conditions. In macrophages stimulated with LPS, elevated NO production and peroxynitrite formation were reduced by plant extract treatments, which downregulated iNOS expression, decreased NOx levels, and suppressed markers of lipid peroxidation like MDA.36,60 Similar effects were observed in animal models of carrageenan-induced inflammation or sodium nitroprusside challenge, where phytochemical supplementation lowered tissue NO levels, restored antioxidant enzyme activity (SOD, CAT, GPx), and limited nitrosative injury.53,70,78
Clinical findings support these results. In elderly subjects, long-term berry supplementation reduced systemic nitrosative stress, as shown by lower MDA levels and improved antioxidant capacity.85 Serum from these participants also decreased LPS-induced NO and TNF-α production in ex vivo assays, indicating diminished iNOS activity. In hypertensive patients, polyphenol supplementation enhanced endothelial NO bioavailability through the phosphorylation of eNOS, while keeping nitrosative stress markers within normal ranges.83 This demonstrates selective enhancement of protective NO pathways without triggering iNOS toxicity. In COVID-19 patients, herbal mixtures that increased circulating NO also improved redox balance, with higher levels of hemoglobin-bound NO (NOHb) stabilizing NO and preventing the accumulation of harmful peroxynitrite.80
Discussion
Mechanistically, the three NOS isoforms frame how NO signalling interacts across systems. eNOS maintains vascular tone and homeostasis,125 while iNOS, upregulated during immune activation, generates higher nitric oxide levels that can kill pathogens but also injure host tissue if unchecked.126 This polarity explains the therapeutic window for NO modulation. Lowering NO levels helps when hyperinflammation, high cytokines, and nitrotyrosine indicate peroxynitrite injury, because suppressing iNOS limits oxidative damage.6 Raising NO is useful when endothelial function is impaired, for example, in hypertension, metabolic syndrome, aging, or post-viral vascular stress, where supporting eNOS improves perfusion and microvascular function.80,83,85
At the molecular level, the active iNOS enzyme functions as a homodimer.45 iNOS induction varies by cell type and species because multiple signaling pathways and transcription factors converge on its promoter.127 In addition, NO, depending on the cell type, along with protein kinase A, protein kinase C, p42/p44 mitogen-activated protein kinase/extracellular signal-regulated kinases (MAPK/ERK) pathway.128 The nuclear factor-κB (NF-κB) serves as a pivotal regulator of iNOS expression, responding to both activators and inhibitors. Various stimuli, including LPS, IL-1β, TNF-α, and oxidative stress, have been shown to induce iNOS expression by activating NF-κB. Pharmacologic inhibition of iNOS expression by glucocorticoids, transforming growth factor family members, antioxidants, and phosphatidylcholine-specific phospholipase C inhibitors is closely tied to suppression of NF-κB activation through mechanisms that include blocking nuclear translocation, reducing transactivation, or increasing IκB isoforms.129,130
Medicinal plants have long been recognized as a rich source of bioactive compounds that influence NO pathways.28,131,132 Across experimental systems, diverse class of compounds, including flavonoids, phenolics, terpenoids, and alkaloids, demonstrate bidirectional regulation of NO homeostasis. Under inflammatory conditions characterized by excessive iNOS activation, these phytochemicals consistently suppress iNOS expression, reduced nitric oxide overproduction, and limit downstream nitrosative stress and cytokine amplification. Conversely, in settings of endothelial dysfunction where NO bioavailability is impaired, several plant-derived polyphenols enhance eNOS activity and promote protective NO signalling, thereby restoring vascular function. This context-dependent modulation highlighting a unifying pharmacological principle in which medicinal plants do not simply inhibit NO formation but instead normalize NO balance according to physiological demand. Evidence from clinical studies, particularly those involving polyphenol-rich fruits such as blueberry, strawberry, grape, strawberry, and etc, further supports improvements in endothelial function and eNOS activation, although these findings are largely derived from mixed formulation and remain limited in number. Collectively, these observations underscore the therapeutic potential of plant-derived compounds while emphasizing the need for more standardized and mechanistically targeted clinical investigation.
Dietary polyphenols intersect with this biology by acting on NO signaling and metabolism. Evidence indicates improvements in eNOS expression and activity, reduced eNOS uncoupling, and downregulated iNOS expression. Flavonoids were the largest group contributing to NO pathway modulation in the compiled results, consistent with their anti-inflammatory and antioxidant properties. Structural features are important for activity, including a planar ring with unsaturation at C2–C3 and hydroxyl groups at the 3′ and 4′ positions of the B ring, which are critical for anti-inflammatory effects.133 Although mechanisms differ across compounds, several patterns recur. Quercetin shows anti-inflammatory effects by inhibiting JNK and ERK, which limits MAPK, AP-1, and NF-κB activity.134–136 Kaempferol inhibits the NF-κB pathway and reduces IL-6, IL-1β, COX-2, NOS, and TNF-α.137 Phenolic compounds can also shift cellular metabolism from glycolysis toward mitochondrial oxidative phosphorylation by enhancing mitochondrial biogenesis and efficiency, often through PGC-1α (peroxisome proliferator-activated receptor gamma coactivator 1-α) and Sirtuin 1.130 This shift improves ATP production and redox balance, reduces ROS, and lowers downstream inflammatory signaling, which together reduce iNOS expression and nitric oxide overproduction while helping maintain redox homeostasis.138,139 Notably, the strawberry calyx. In LPS-stimulated RAW 264.7 macrophages, 70% ethanol extracts of Fragaria ananassa Duch. calyx significantly inhibited protein and mRNA expression of iNOS and COX-2 and reduced pro-inflammatory cytokines TNF-α and IL-6.48 The extract attenuated MAPK signaling by lowering phosphorylated ERK and JNK, with a more modest reduction in phosphorylated p38, indicating regulation of inflammatory responses through MAPK pathway control.140 It also reduced phosphorylated p65 and phosphorylated IκB, consistent with inhibition of NF-κB signaling triggered by LPS-TLR4 activation, and thereby suppressed transcription of IL-6, TNF-α, iNOS, and COX-2.141,142 Together, these findings support a dual action in which plant-derived constituents limit upstream inflammatory signaling while also constraining downstream enzymatic drivers of NO and prostaglandin E2 production.48
Furthermore, synthesis of the preclinical data revealed that only four plants, namely A. tsao-ko, C. sativa, M. biondii, and P. obtuse, demonstrated consistent activity across both in vitro and in vivo models. These studies reported reproducible reductions in NO production and iNOS expression, indicating mechanistic support for their anti-inflammatory effects. However, none of these candidates have yet progressed to well-controlled clinical trials, underscoring a substantial gap between preclinical promise and clinical validation. This pattern suggests that only a small subset of medicinal plants has undergone multi-level validation, highlighting the need for more standardized preclinical to clinical progression.
Despite encouraging preclinical findings, the clinical translation of medicinal plants targeting nitric oxide signalling remains limited. Substantial variability in extraction methods, phytochemical composition, and dosing regimens contributes to inconsistent outcomes, while many herbal preparations lack adequate chemical and pharmacokinetic characterization. Most in vitro studies rely heavily on LPS-induced inflammatory models, which provide experimental consistency but restrict generalizability and fail to reflect the complexity of chronic inflammatory conditions. Animal studies are fewer and methodologically heterogeneous, often emphasizing broad inflammatory markers such as cytokines or oxidative stress parameters. This imbalance reduces mechanistic insight into NO-specific pathways. Differences in experimental design, dosing strategies, and outcome measures also restrict comparability and weaken causal interpretation. As a result, although animal data are encouraging of nitric oxide modulation in therapeutic effects remains insufficiently defined. Furthermore, the clinical translational studies represent the most significant gap. Few human studies directly assess NO-related outcomes following plant-based interventions. Many trials use complex multi-component formulations rather than standardized single plant extracts, making attribution of effects to specific phytochemicals difficult. Variability in formulations, populations, and endpoints further limits reproducibility. Consequently, current clinical findings should be interpreted cautiously, and definitive conclusions regarding therapeutic efficacy cannot yet be established.
Collectively, the evidence demonstrates a clear translational gradient. Bridging this gap will require standardized extract preparation, consistent dosing protocols, clearly defined NO-specific endpoints, and well-designed randomized control trials (RCTs). Such improvements are essential to determine whether modulation of NO signalling by medicinal plants can produce reliable clinical benefits.
Limitations of the Study
Although the present review provides comprehensive insights into the effects of medicinal plants on NO production, several limitations must be acknowledged that could affect the interpretation of the results. Firstly, preclinical models often use acute LPS or SNP challenges; this could be a discrepancy between in vitro and in vivo models that remains a concern. The conditions in cell cultures do not always replicate the complexities of the in vivo system, where factors such as the pharmacokinetics of bioactive compounds may vary. Secondly, although this study uses human trials, they often rely on indirect measures such as plasma NOx, which do not distinguish between harmful iNOS activity and protective eNOS signaling. Furthermore, these studies remain short, small in scale, and rarely stratify participants by baseline NO status or endothelial function, even though this likely dictates the direction of response. In addition, herbal mixtures are frequently under-characterized with insufficient data on phytochemical composition, pharmacokinetics, and dose-response relationships. Additional research is needed to better understand the interactions between the various bioactive compounds of plants and their individual contributions to therapeutic outcomes. Although many studies reported significant iNOS inhibition, the statistical robustness varied considerably. Several investigations relied on small sample sizes, limited replication, and incomplete reporting of variance or statistical testing, which may reduce confidence in the estimated effects. In addition, heterogeneity in experimental designs and outcome measures limits cross-study comparability. Moreover, further studies should focus on mechanism-anchored randomized controlled trials that stratify participants by endothelial function and nitrosative stress at baseline. Harmonized biomarker panels that include endothelial function tests, nitrotyrosine, HbNO, cytokines, and antioxidant enzyme activity will be critical. Linking phytochemical dose and plasma metabolites to pharmacodynamic effects will help establish causality, while longer-term studies are needed to assess durability and safety.
Conclusion
Taken together, accumulating evidence indicates that many medicinal plants exert anti-inflammatory and antioxidant effects through modulation of NO signaling and improve NO bioavailability. Rather than acting solely as inhibitors, these phytochemicals appear to normalize nitric oxide homeostasis by reducing excessive NO production during inflammatory states while restoring deficient NO levels in vascular dysfunction. This bidirectional regulation provides a coherent mechanistic links phytochemicals and improvements in inflammation, oxidative stress, and endothelial health, suggesting a shared mechanistic basis underlying their therapeutic potential. However, the current evidence remains largely preclinical and heterogeneous, and well-controlled clinical trials remain limited, warranting cautious interpretation of translational relevance. Future studies should prioritize standardized interventions, robust biomarkers, and precision-targeted strategies to ensure safe and effective translation of phytochemical-based NO modulation into clinical practice.
Acknowledgments
The authors thank (1) the Directorate of Higher Education of the Ministry of Education and Culture of the Republic of Indonesia for supporting the tuition fee and doctoral research of the first author, and (2) the Rector of Universitas Padjadjaran via the Directorate of Research, Downstream, and Community Engagement for facilitating the APC. The APC is funded by Universitas Padjadjaran through the Indonesian Endowment Fund for Education (LPDP) on behalf of the Indonesian Ministry of Higher Education, Science and Technology, and managed under the EQUITY Program (Contract No. 4303/ B3/DT.03.08/2025 and 3927/UN6. RKT/HK.07.00/2025). The authors also thank all reviewers and editors for their valuable reviews, remarks, and suggestions, which helped improve the quality and clarity of this manuscript.
The present study was conducted in the framework of the dissertation project of the first author at the Doctoral Program in Pharmacy, Faculty of Pharmacy, Universitas Padjadjaran, West Java, Indonesia.
Disclosure
The authors declared that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
1. Kurhaluk N, Tkaczenko H. L-Arginine and nitric oxide in vascular regulation—experimental findings in the context of blood donation. Nutrients. 2025;17(4):665. doi:10.3390/nu17040665
2. Wu G, Meininger CJ, McNeal CJ, Bazer FW, Rhoads JM. Role of L-Arginine in Nitric Oxide Synthesis and Health in Humans. In: Amino Acids in Nutrition and Health. Advances in Experimental Medicine and Biology; 2021:167–21. doi:10.1007/978-3-030-74180-8_10
3. Rochette L, Lorin J, Zeller M, et al. Nitric oxide synthase inhibition and oxidative stress in cardiovascular diseases: possible therapeutic targets? Pharmacol Ther. 2013;140(3):239–257. doi:10.1016/j.pharmthera.2013.07.004
4. Reis PA, de ACFG, Maron‐Gutierrez T, Silva AR, Neto HC de CF. Role of Nitric Oxide Synthase in the Function of the Central Nervous System under Normal and Infectious Conditions. In: Nitric Oxide Synthase - Simple Enzyme-Complex Roles. InTech. 2017. doi:10.5772/67816
5. Günther M, Al Nimer F, Gahm C, Piehl F, Mathiesen T. iNOS-mediated secondary inflammatory response differs between rat strains following experimental brain contusion. Acta Neurochir. 2012;154(4):689–697. doi:10.1007/s00701-012-1297-1
6. Förstermann U, Sessa WC. Nitric oxide synthases: regulation and function. Eur Heart J. 2012;33(7):829–837, 837a–837d. doi:10.1093/eurheartj/ehr304
7. Kone BC, Kuncewicz T, Zhang W, Yu ZY. Protein interactions with nitric oxide synthases: controlling the right time, the right place, and the right amount of nitric oxide. Am J Physiol Physiol. 2003;285(2):F178–F190. doi:10.1152/ajprenal.00048.2003
8. Cinelli MA, Do HT, Miley GP, Silverman RB. Inducible nitric oxide synthase: regulation, structure, and inhibition. Med Res Rev. 2020;40(1):158–189. doi:10.1002/med.21599
9. MacMicking J, wen XQ, Nathan C. Nitric oxide and macrophage function. Annu Rev Immunol. 1997;15(1):323–350. doi:10.1146/annurev.immunol.15.1.323
10. Toda N, Okamura T. The pharmacology of nitric oxide in the peripheral nervous system of blood vessels. Pharmacol Rev. 2003;55(2):271–324. doi:10.1124/pr.55.2.3
11. Schuman EM, Madison DV. Nitric oxide and synaptic function. Annu Rev Neurosci. 1994;17(1):153–183. doi:10.1146/annurev.ne.17.030194.001101
12. Yuste JE, Tarragon E, Campuzano CM, Ros-Bernal F. Implications of glial nitric oxide in neurodegenerative diseases. Front Cell Neurosci. 2015;9:322. doi:10.3389/fncel.2015.00322
13. MacMicking JD, Nathan C, Hom G, et al. Altered responses to bacterial infection and endotoxic shock in mice lacking inducible nitric oxide synthase. Cell. 1995;81(4):641–650. doi:10.1016/0092-8674(95)90085-3
14. Behera J, Nagarajan S, Saran U, et al. Nitric oxide restores peripheral blood mononuclear cell adhesion against hypoxia via NO‐cGMP signalling. Cell Biochem Funct. 2020;38(3):319–329. doi:10.1002/cbf.3502
15. Menzies-Gow A, Mansur AH, Brightling CE. Clinical utility of fractional exhaled nitric oxide in severe asthma management. Eur Respir J. 2020;55(3):1901633. doi:10.1183/13993003.01633-2019
16. Aranda-Rivera AK, Cruz-Gregorio A, Arancibia-Hernández YL, Hernández-Cruz EY, Pedraza-Chaverri J. ROS and oxidative stress: an overview of basic concepts. Oxygen. 2022;2(4):437–478. doi:10.3390/oxygen2040030
17. Lambeth JD, Neish AS. Nox enzymes and new thinking on reactive oxygen: a double-edged sword revisited. Annu Rev Pathol Mech Dis. 2014;9(1):119–145. doi:10.1146/annurev-pathol-012513-104651
18. Michalak KP, Michalak AZ. Understanding chronic inflammation: couplings between cytokines, ROS, NO, Cai 2+, HIF-1α, Nrf2 and autophagy. Front Immunol. 2025;16:1558263. doi:10.3389/fimmu.2025.1558263
19. Manful CF, Fordjour E, Ikumoinein E, Abbey L, Thomas R. Therapeutic strategies targeting oxidative stress and inflammation: a narrative review. BioChem. 2025;5(4):35. doi:10.3390/biochem5040035
20. Kim ME, Lee JS. Advances in the regulation of inflammatory mediators in nitric oxide synthase: implications for disease modulation and therapeutic approaches. Int J Mol Sci. 2025;26(3). doi:10.3390/ijms26031204
21. Ziegler K, Kunert AT, Reinmuth-Selzle K, et al. Chemical modification of pro-inflammatory proteins by peroxynitrite increases activation of TLR4 and NF-κB: implications for the health effects of air pollution and oxidative stress. Redox Biol. 2020;37:101581. doi:10.1016/j.redox.2020.101581
22. Wong V, Wai C, Lerner E. Nitric oxide inhibition strategies. Futur Sci OA. 2015;1(1). doi:10.4155/fso.15.35
23. Sullivan R, Graham C. Chemosensitization of cancer by nitric oxide. Curr Pharm Des. 2008;14(11):1113–1123. doi:10.2174/138161208784246225
24. Fitzpatrick B, Mehibel M, Cowen RL, Stratford IJ. iNOS as a therapeutic target for treatment of human tumors. Nitric Oxide. 2008;19(2):217–224. doi:10.1016/j.niox.2008.05.001
25. World Health Organization. Integrating Traditional Medicine in Health Care. 2023. Available from: https://www.who.int/southeastasia/news/feature-stories/detail/integrating-traditional-medicine.
26. Wake GE, Fitie GW. Magnitude and determinant factors of herbal medicine utilization among mothers attending their antenatal care at public health institutions in Debre Berhan Town, Ethiopia. Front Public Health. 2022;10. doi:10.3389/fpubh.2022.883053.
27. Gaire BP, Subedi L. Medicinal plant diversity and their pharmacological aspects of Nepal Himalayas. Pharmacogn J. 2011;3(25):6–17. doi:10.5530/pj.2011.25.2
28. Subedi L, Gaire BP, Kim SY, Parveen A. Nitric oxide as a target for phytochemicals in anti-neuroinflammatory prevention therapy. Int J Mol Sci. 2021;22(9):4771. doi:10.3390/ijms22094771
29. Giles DA, Washnock‐Schmid JM, Duncker PC, et al. Myeloid cell plasticity in the evolution of central nervous system autoimmunity. Ann Neurol. 2018;83(1):131–141. doi:10.1002/ana.25128
30. Xu Y, Gao W, Sun Y, Wu M. New insight on microglia activation in neurodegenerative diseases and therapeutics. Front Neurosci. 2023;17. doi:10.3389/fnins.2023.1308345.
31. Bagnato F, Hametner S, Yao B, et al. Tracking iron in multiple sclerosis: a combined imaging and histopathological study at 7 Tesla. Brain. 2011;134(12):3602–3615. doi:10.1093/brain/awr278
32. Martemucci G, Costagliola C, Mariano M, D’andrea L, Napolitano P, D’Alessandro AG. Free radical properties, source and targets, antioxidant consumption and health. Oxygen. 2022;2(2):48–78. doi:10.3390/oxygen2020006
33. Lim JS, Lee SH, Lee SR, et al. Inhibitory Effects of Aucklandia lappa Decne. extract on inflammatory and oxidative responses in LPS-treated macrophages. Molecules. 2020;25(6):1336. doi:10.3390/molecules25061336
34. Kim JM, Choi MH, Yang JH. Cinnamomum japonicum Siebold branch extracts attenuate NO and ROS production via the inhibition of p38 and JNK phosphorylation. Molecules. 2023;28(4):1974. doi:10.3390/molecules28041974
35. Gil TY, Jin BR, Cha YY, An HJ. Magnoliae flos downregulated lipopolysaccharide-induced inflammatory responses via NF-κB/ERK-JNK MAPK/STAT3 pathways. Mediators Inflamm. 2022;2022:1–11. doi:10.1155/2022/6281892
36. Shin JS, Ryu S, Jang DS, Cho YW, Chung EK, Lee KT. Amomum tsao-ko fruit extract suppresses lipopolysaccharide-induced inducible nitric oxide synthase by inducing heme oxygenase-1 in macrophages and in septic mice. Int J Exp Pathol. 2015;96(6):395–405. doi:10.1111/iep.12159
37. Borchard J, Mazzarella L, Spelman K. A review of medicinal plants that modulate nitric oxide activity. Altern Med Stud. 2012;2(1):6. doi:10.4081/ams.2012.e6
38. Sadaf S, Nagarkoti S, Awasthi D, et al. nNOS induction and NOSIP interaction impact granulopoiesis and neutrophil differentiation by modulating nitric oxide generation. Biochim Biophys Acta Mol Cell Res. 2021;1868(7):119018. doi:10.1016/j.bbamcr.2021.119018
39. Bogdan C. Nitric oxide synthase in innate and adaptive immunity: an update. Trends Immunol. 2015;36(3):161–178. doi:10.1016/j.it.2015.01.003
40. Soufli I, Toumi R, Rafa H, Touil-Boukoffa C. Overview of cytokines and nitric oxide involvement in immuno-pathogenesis of inflammatory bowel diseases. World J Gastrointest Pharmacol Ther. 2016;7(3):353–360. doi:10.4292/wjgpt.v7.i3.353
41. Arya P, Sharma V, Singh P, Thapliyal S, Sharma M. Bacterial endotoxin-lipopolysaccharide role in inflammatory diseases: an overview. Iran J Basic Med Sci. 2025;28(5):553–564. doi:10.22038/ijbms.2025.82302.17799
42. Heiles B. Lipopolysaccharide (LPS): Understanding the role in inflammation and disease. J Probiotics Heal. 2023;1(02):8901. doi:10.35248/2329-8901.23.11.321
43. Kleinbongard P, Rassaf T, Dejam A, Kerber S, Kelm M. Griess method for nitrite measurement of aqueous and protein-containing samples. In: Detection of Nitric Oxide; 2002:158–168. doi:10.1016/S0076-6879(02)59180-1
44. V Pinto R, Antunes F, Pires J, Silva-Herdade A, Pinto ML. A comparison of different approaches to quantify nitric oxide release from NO-releasing materials in relevant biological media. Molecules. 2020;25(11). doi:10.3390/molecules25112580
45. Serreli G, Deiana M. Role of dietary polyphenols in the activity and expression of nitric oxide synthases: a review. Antioxidants. 2023;12(1):147. doi:10.3390/antiox12010147
46. Lindsay RT, Rhodes CJ. Reactive oxygen species (ROS) in metabolic disease—don’t shoot the metabolic messenger. Int J Mol Sci. 2025;26(6):2622. doi:10.3390/ijms26062622
47. Duraisamy P, Angusamy A, Ravi S, et al. Phytol from Scoparia dulcis prevents NF-κB-mediated inflammatory responses during macrophage polarization. 3 Biotech. 2024;14(3):1–30. doi:10.1007/s13205-024-03924-9
48. Kim HM, Yoo DH, Kang JW, Lee IC, Bae JS. Anti-inflammatory effect of extract from Fragaria ananassa Duch. calyx via MAPK and NF-κB signaling pathway. J Microbiol Biotechnol. 2024;34(12):2662–2674. doi:10.4014/jmb.2409.09044
49. Notarte KIR, Quimque MTJ, Macaranas IT, et al. Attenuation of lipopolysaccharide-induced inflammatory responses through inhibition of the NF-κB pathway and the increased NRF2 level by a flavonol-enriched n-butanol fraction from Uvaria alba. ACS Omega. 2023;8(6):5377–5392. doi:10.1021/acsomega.2c06451
50. Zhao Y, Yang Y, Liu M, et al. COX-2 is required to mediate crosstalk of ROS-dependent activation of MAPK/NF-κB signaling with pro-inflammatory response and defense-related NO enhancement during challenge of macrophage-like cell line with Giardia duodenalis. PLoS Negl Trop Dis. 2022;16(4):e0010402. doi:10.1371/journal.pntd.0010402
51. Kim EA, Kim SY, Ye BR, et al. Anti-inflammatory effect of Apo-9′-fucoxanthinone via inhibition of MAPKs and NF-kB signaling pathway in LPS-stimulated RAW 264.7 macrophages and zebrafish model. Int Immunopharmacol. 2018;59:339–346. doi:10.1016/j.intimp.2018.03.034
52. Lee JE, Lee EH, Park HJ, et al. Inhibition of inflammatory responses in lipopolysaccharide-induced RAW 264.7 cells by Pinus densiflora root extract. J Appl Biol Chem. 2018;61(3):275–281. doi:10.3839/jabc.2018.039
53. Lee Y, Lim HW, Ryu IW, et al. Anti-inflammatory, barrier-protective, and antiwrinkle properties of agastache rugosa kuntze in human epidermal keratinocytes. Biomed Res Int. 2020;2020:1759067. doi:10.1155/2020/1759067
54. Oh HM, Kang YJ, Kim SH, et al. Agastache rugosa leaf extract inhibits the iNOS expression in ROS 17/2.8 cells activated with TNF-α and IL-1β. Arch Pharm Res. 2005;28(3):305–310. doi:10.1007/BF02977797
55. Wang X, Dongzhi Z, Li Y, et al. Ajania purpurea extract attenuates LPS-induced inflammation in RAW264.7 cells and peritonitis mice. Biol Pharm Bull. 2022;45(12):b22–00388. doi:10.1248/bpb.b22-00388
56. Kim EH, Shim YY, Lee HI, Lee S, Reaney MJT, Chung MJ. Astragalin and isoquercitrin isolated from Aster scaber suppress LPS-induced neuroinflammatory responses in microglia and mice. Foods. 2022;11(10). doi:10.3390/foods11101505
57. Jeong D, zhi DG, Lee HJ, Ryu JH. Anti-Inflammatory compounds from atractylodes macrocephala. Molecules. 2019;24(10):1–11. doi:10.3390/molecules24101859
58. Hong M, Kim JH, Han JH, et al. In vitro and in vivo anti-inflammatory potential of cannabichromene isolated from Hemp. Plants. 2023;12(23):1–13. doi:10.3390/plants12233966
59. Kim HY, Han AR, Kil YS, Seo EK, Jin CH. Anti-inflammatory effects of catalpalactone isolated from catalpa ovata in LPS-induced RAW264.7 cells. Molecules. 2019;24(7):1–12. doi:10.3390/molecules24071236
60. Preedalikit W, Chittasupho C, Leelapornpisid P, Duangnin N, Kiattisin K. Potential of coffee cherry pulp extract against polycyclic aromatic hydrocarbons in air pollution induced inflammation and oxidative stress for topical applications. Int J Mol Sci. 2024;25(17):1–20. doi:10.3390/ijms25179416
61. Chen Y, Ruan J, Sun F, et al. Anti-inflammatory limonoids from Cortex dictamni. Front Chem. 2020;8(February):1–12. doi:10.3389/fchem.2020.00073
62. Moreira P, Macedo J, Matos P, et al. Effect of bioactive extracts from Eucalyptus globulus leaves in experimental models of Alzheimer’s disease. Biomed Pharmacother. 2024:181. doi:10.1016/j.biopha.2024.117652.
63. Jang HY, Lee SO. Heme oxygenase 1-mediated anti-inflammatory effect of extract from the aerial part of Heracleum moellendorffii hance. Foods. 2023;12(17). doi:10.3390/foods12173309
64. Lim H, Noh JR, Kim YH, et al. Anti-atherogenic effect of Humulus japonicus in apolipoprotein E-deficient mice. Int J Mol Med. 2016;38(4):1101–1110. doi:10.3892/ijmm.2016.2727
65. Kim YJ, Park SY, Koh YJ, Lee JH. Anti-neuroinflammatory effects and mechanism of action of Fructus ligustri lucidi extract in BV2 microglia. Plants. 2021;10(4):688. doi:10.3390/plants10040688
66. Apaza Ticona L, Sánchez Sánchez-Corral J, Montoto Lozano N, Prieto Ramos P, Sánchez ÁR. Study of pentacyclic triterpenes from lyophilised aguaje: anti-inflammatory and antioxidant properties. Int J Mol Sci. 2024;25(17). doi:10.3390/ijms25179615
67. Lin Z, Gan T, Huang Y, et al. Anti-inflammatory activity of mulberry leaf flavonoids in vitro and in vivo. Int J Mol Sci. 2022;23(14). doi:10.3390/ijms23147694
68. Suriyaprom S, Srisai P, Intachaisri V, et al. Antioxidant and anti-inflammatory activity on LPS-stimulated RAW 264.7 macrophage cells of white Mulberry (Morus alba L.) Leaf extracts. Molecules. 2023;28(11):4395. doi:10.3390/molecules28114395
69. Hwang SJ, Wang JH, Lee JS, et al. Ginseng Sprouts attenuate mortality and systemic inflammation by modulating TLR4/NF-κB signaling in an LPS-induced mouse model of sepsis. Int J Mol Sci. 2023;24(2). doi:10.3390/ijms24021583
70. Eloutify YT, El-Shiekh RA, Ibrahim KM, et al. Bioactive fraction from Plumeria obtusa L. attenuates LPS-induced acute lung injury in mice and inflammation in RAW 264.7 macrophages: LC/QToF-MS and molecular docking. Inflammopharmacology. 2023;31(2):859–875. doi:10.1007/s10787-023-01144-w
71. Liu XY, Zhang YB, Yang XW, et al. Anti-inflammatory activity of some characteristic constituents from the vine stems of Spatholobus suberectus. Molecules. 2019;24(20):1–16. doi:10.3390/molecules24203750
72. Lee JA, Shin JY, Hong SS, et al. Tetracera loureiri extract regulates lipopolysaccharide-induced inflammatory response via nuclear factor-κB and mitogen activated protein kinase signaling pathways. Plants. 2022;11(3):284. doi:10.3390/plants11030284
73. Tucureanu MM, Rebleanu D, Constantinescu CA, et al. Lipopolysaccharide-induced inflammation in monocytes/macrophages is blocked by liposomal delivery of Gi-protein inhibitor. Int J Nanomed. 2018;13:63–76. doi:10.2147/IJN.S150918
74. Zhou Y, Wang Y, Yu D, et al. Sodium nitroprusside improves circulatory failure in rabbit acute pulmonary embolism combined with shock model possibly by enhancing NO release and inhibiting TLR4/NF-кB/HIF-1α signaling pathway. Front Physiol. 2025;16:1573405. doi:10.3389/fphys.2025.1573405
75. Prayoga D, Aulifa D, Budiman A, Levita J, Jiranusornkul S. Etlingera elatior inflorescence extract mitigates acute gastric ulcers by suppressing the expression of inducible nitric oxide synthase in ethanol-induced wistar rats. J Exp Pharmacol. 2025;17:343–357. doi:10.2147/JEP.S524517
76. Zarev Y, Marinov L, Momekova D. Exploring phytochemical composition and in vivo anti-inflammatory potential of grape seed oil from an alternative source after traditional fermentation processes: implications for phytotherapy. Plants. 2023;12:2795.
77. Salem S, Leghouchi E, Soulimani R, Bouayed J. Reduction of paw edema and liver oxidative stress in carrageenan-induced acute inflammation by Lobaria pulmonaria and Parmelia caperata, lichen species, in mice. Int J Vitamin Nutr Res. 2021;91:143–151.
78. Shen J, Ma X, He Y, Wang Y, Zhong T, Zhang Y. Anti-inflammatory and anti-oxidant properties of Melianodiol on DSS-induced ulcerative colitis in mice. PeerJ. 2022;10. doi:10.7717/peerj.14209.
79. Kim H, Hong J, Lee J, et al. Curcuma aromatica Salisb. protects from acetaminophen-induced hepatotoxicity by regulating the Sirt1/HO-1 signaling pathway. Nutrients. 2023;15(4). doi:10.3390/nu15040808
80. Fink B, Hunter JM, Pietrzkowski Z, et al. A plant-based dietary supplement exhibits significant effects on markers of oxidative stress, inflammation, and immune response in subjects recovering from respiratory viral infection: a randomized, double-blind clinical study using Vitamin C as a positive control. Int J Mol Sci. 2025;26(11):5209. doi:10.3390/ijms26115209
81. Biesinger S, Michaels HA, Quadros AS, et al. A combination of isolated phytochemicals and botanical extracts lowers diastolic blood pressure in a randomized controlled trial of hypertensive subjects. Eur J Clin Nutr. 2016;70(1):10–16. doi:10.1038/ejcn.2015.88
82. Rutledge GA, Fisher DR, Miller MG, Kelly ME, Bielinski DF, Shukitt-Hale B. The effects of blueberry and strawberry serum metabolites on age-related oxidative and inflammatory signaling in vitro. Food Funct. 2019;10(12):7707–7713. doi:10.1039/c9fo01913h
83. Nemzer BV, Pietrzkowski Z, Hunter JM, Robinson JL, Fink B. A betalain-rich dietary supplement, but not PETN, increases vasodilation and nitric oxide: a comparative, single-dose, randomized, placebo-controlled, blinded, crossover pilot study. J Food Res. 2020;10(1):26. doi:10.5539/jfr.v10n1p26
84. Gu Y, Zuo X, Zhang S, et al. The mechanism behind influenza virus cytokine storm. Viruses. 2021;13(7):1362. doi:10.3390/v13071362
85. Sandhu A, Miller MG, Shukitt‐Hale B, Edirisinghe I, Burton‐Freeman B. Metabolic fate of blueberry anthocyanins after chronic supplementation in healthy older adults. FASEB J. 2017;31(S1). doi:10.1096/fasebj.31.1_supplement.646.20
86. Barona J, Aristizabal JC, Blesso CN, Volek JS, Fernandez ML. Grape polyphenols reduce blood pressure and increase flow-mediated vasodilation in men with metabolic syndrome. J Nutr. 2012;142(9):1626–1632. doi:10.3945/jn.112.162743
87. Kapil V, Milsom AB, Okorie M, et al. Inorganic nitrate supplementation lowers blood pressure in humans. Hypertension. 2010;56(2):274–281. doi:10.1161/HYPERTENSIONAHA.110.153536
88. Webb AJ, Patel N, Loukogeorgakis S, et al. Acute blood pressure lowering, vasoprotective, and antiplatelet properties of dietary nitrate via bioconversion to nitrite. Hypertension. 2008;51(3):784–790. doi:10.1161/HYPERTENSIONAHA.107.103523
89. Kim JH, Choi MS. Nitric oxide signal transduction and its role in skin sensitization. Biomol Ther. 2023;31(4):388–394. doi:10.4062/biomolther.2023.101
90. Kashfi K, Kannikal J, Nath N. Macrophage reprogramming and cancer therapeutics: role of iNOS-derived NO. Cells. 2021;10(11):3194. doi:10.3390/cells10113194
91. Mishra BB, Rathinam VAK, Martens GW, et al. Nitric oxide controls the immunopathology of tuberculosis by inhibiting NLRP3 inflammasome–dependent processing of IL-1β. Nat Immunol. 2013;14(1):52–60. doi:10.1038/ni.2474
92. Mao K, Chen S, Chen M, et al. Nitric oxide suppresses NLRP3 inflammasome activation and protects against LPS-induced septic shock. Cell Res. 2013;23(2):201–212. doi:10.1038/cr.2013.6
93. Kraus R, Maier E, Gruber M, Wittmann S. Impact of nitric oxide on polymorphonuclear neutrophils’ function. Biomedicines. 2024;12(10):2353. doi:10.3390/biomedicines12102353
94. Farah C, Michel LYM, Balligand JL. Nitric oxide signalling in cardiovascular health and disease. Nat Rev Cardiol. 2018;15(5):292–316. doi:10.1038/nrcardio.2017.224
95. Naseem K. The role of nitric oxide in cardiovascular diseases. Mol Aspects Med. 2005;26(1–2):33–65. doi:10.1016/j.mam.2004.09.003
96. Gajecki D, Gawryś J, Szahidewicz-Krupska E, Doroszko A. Role of erythrocytes in nitric oxide metabolism and paracrine regulation of endothelial function. Antioxidants. 2022;11(5):943. doi:10.3390/antiox11050943
97. Cyr AR, Huckaby LV, Shiva SS, Zuckerbraun BS. Nitric oxide and endothelial dysfunction. Crit Care Clin. 2020;36(2):307–321. doi:10.1016/j.ccc.2019.12.009
98. Groemping Y, Lapouge K, Smerdon SJ, Rittinger K. Molecular basis of phosphorylation-induced activation of the NADPH oxidase. Cell. 2003;113(3):343–355. doi:10.1016/S0092-8674(03)00314-3
99. Nunes P, Demaurex N, Dinauer MC. Regulation of the NADPH oxidase and associated ion fluxes during phagocytosis. Traffic. 2013;14(11):1118–1131. doi:10.1111/tra.12115
100. Nisimoto Y, Motalebi S, Han CH, Lambeth JD. The p67 activation domain regulates electron flow from NADPH to flavin in flavocytochromeb 558. J Biol Chem. 1999;274(33):22999–23005. doi:10.1074/jbc.274.33.22999
101. Zeng MY, Miralda I, Armstrong CL, Uriarte SM, Bagaitkar J. The roles of NADPH oxidase in modulating neutrophil effector responses. Mol Oral Microbiol. 2019;34(2):27–38. doi:10.1111/omi.12252
102. Lehman HK, Segal BH. The role of neutrophils in host defense and disease. J Allergy Clin Immunol. 2020;145(6):1535–1544. doi:10.1016/j.jaci.2020.02.038
103. Abo A, Webb MR, Grogan A, Segal AW. Activation of NADPH oxidase involves the dissociation of p21 rac from its inhibitory GDP/GTP exchange protein (rhoGDI) followed by its translocation to the plasma membrane. Biochem J. 1994;298(3):585–591. doi:10.1042/bj2980585
104. Reeves EP, Lu H, Jacobs HL, et al. Killing activity of neutrophils is mediated through activation of proteases by K+ flux. Nature. 2002;416(6878):291–297. doi:10.1038/416291a
105. Thomas DD, Heinecke JL, Ridnour LA, et al. Signaling and stress: the redox landscape in NOS2 biology. Free Radic Biol Med. 2015;87:204–225. doi:10.1016/j.freeradbiomed.2015.06.002
106. Jyoti A, Singh AK, Dubey M, et al. Interaction of inducible nitric oxide synthase with rac2 regulates reactive oxygen and nitrogen species generation in the human neutrophil phagosomes: implication in microbial killing. Antioxid Redox Signal. 2014;20(3):417–431. doi:10.1089/ars.2012.4970
107. Xue Q, Yan Y, Zhang R, Xiong H. Regulation of iNOS on immune cells and its role in diseases. Int J Mol Sci. 2018;19(12):3805. doi:10.3390/ijms19123805
108. Bogdan C, Röllinghoff M, Diefenbach A. The role of nitric oxide in innate immunity. Immunol Rev. 2000;173(1):17–26. doi:10.1034/j.1600-065X.2000.917307.x
109. Giordano D, Li C, Suthar MS, et al. Nitric oxide controls an inflammatory-like Ly6ChiPDCA1+ DC subset that regulates Th1 immune responses. J Leukoc Biol. 2010;89(3):443–455. doi:10.1189/jlb.0610329
110. Zhang R, Li Q, Chuang PY, et al. Regulation of pathogenic Th17 cell differentiation by IL-10 in the development of glomerulonephritis. Am J Pathol. 2013;183(2):402–412. doi:10.1016/j.ajpath.2013.05.001
111. Yang J, Zhang R, Lu G, et al. T cell–derived inducible nitric oxide synthase switches off TH17 cell differentiation. J Exp Med. 2013;210(7):1447–1462. doi:10.1084/jem.20122494
112. Xiong H, Zhu C, Li F, et al. Inhibition of interleukin-12 p40 transcription and NF-κB activation by nitric oxide in murine macrophages and dendritic cells. J Biol Chem. 2004;279(11):10776–10783. doi:10.1074/jbc.M313416200
113. Obermajer N, Wong JL, Edwards RP, et al. Induction and stability of human Th17 cells require endogenous NOS2 and cGMP-dependent NO signaling. J Exp Med. 2013;210(7):1433–1445. doi:10.1084/jem.20121277
114. Anele UA, Burnett AL. Nitrergic mechanisms for management of recurrent priapism. Sex Med Rev. 2015;3(3):160–168. doi:10.1002/smrj.56
115. Wood KC, Granger DN. Sickle cell disease: role of reactive oxygen and nitrogen metabolites. Clin Exp Pharmacol Physiol. 2007;34(9):926–932. doi:10.1111/j.1440-1681.2007.04639.x
116. Burnett AL, Musicki B, Jin L, Bivalacqua TJ. Nitric oxide/redox-based signalling as a therapeutic target for penile disorders. Expert Opin Ther Targets. 2006;10(3):445–457. doi:10.1517/14728222.10.3.445
117. Arnér ESJ, Holmgren A. Physiological functions of thioredoxin and thioredoxin reductase. Eur J Biochem. 2000;267(20):6102–6109. doi:10.1046/j.1432-1327.2000.01701.x
118. Arteel GE, Sies H. The biochemistry of selenium and the glutathione system. Environ Toxicol Pharmacol. 2001;10(4):153–158. doi:10.1016/S1382-6689(01)00078-3
119. Heales SJR, Bolaños JP, Stewart VC, Brookes PS, Land JM, Clark JB. Nitric oxide, mitochondria and neurological disease. Biochim Biophys Acta - Bioenergy. 1999;1410(2):215–228. doi:10.1016/S0005-2728(98)00168-6
120. Arora R, Kumar A, Gupta R, Maurya PK. Clinical role of oxides in Neuropsychiatric and Neurodegenerative disorders. In: Oxides for Medical Applications. Elsevier; 2023:359–378. doi:10.1016/B978-0-323-90538-1.00011-X
121. Pacher P, Beckman JS, Liaudet L. Nitric oxide and peroxynitrite in health and disease. Physiol Rev. 2007;87(1):315–424. doi:10.1152/physrev.00029.2006
122. Kowluru RA, Abbas SN, Odenbach S. Reversal of hyperglycemia and diabetic nephropathy. J Diabetes Complications. 2004;18(5):282–288. doi:10.1016/j.jdiacomp.2004.03.002
123. Keenan HA, Costacou T, Sun JK, et al. Clinical factors associated with resistance to microvascular complications in diabetic patients of extreme disease duration. Diabetes Care. 2007;30(8):1995–1997. doi:10.2337/dc06-2222
124. Tota Ł, Matejko B, Morawska-Tota M, et al. Changes in oxidative and nitrosative stress indicators and vascular endothelial growth factor after maximum-intensity exercise assessing aerobic capacity in males with type 1 diabetes mellitus. Front Physiol. 2021;12. doi:10.3389/fphys.2021.672403
125. Tran N, Garcia T, Aniqa M, Ali S, Ally A, Nauli SM. Endothelial nitric oxide synthase (eNOS) and the cardiovascular system: in physiology and in disease states. Am J Biomed Sci Res. 2022;15(2):153–177.
126. Farahani A, Farahani A, Kashfi K, Ghasemi A. Inducible nitric oxide synthase (iNOS): more than an inducible enzyme? Rethinking the classification of NOS isoforms. Pharmacol Res. 2025;216:107781. doi:10.1016/j.phrs.2025.107781
127. Kleinert H, Schwarz PM, Förstermann U. Regulation of the expression of inducible nitric oxide synthase. Biol Chem. 2003;384(10–11). doi:10.1515/BC.2003.152
128. Zhang W, Liu HT. MAPK signal pathways in the regulation of cell proliferation in mammalian cells. Cell Res. 2002;12(1):9–18. doi:10.1038/sj.cr.7290105
129. Vannini F, Kashfi K, Nath N. The dual role of iNOS in cancer. Redox Biol. 2015;6:334–343. doi:10.1016/j.redox.2015.08.009
130. Sahakyan N. Plant-derived phenolics as regulators of nitric oxide production in microglia: mechanisms and therapeutic potential. Med Gas Res. 2025;16(2):169–175. doi:10.4103/mgr.MEDGASRES-D-25-00058
131. Forte M, Conti V, Damato A, et al. Targeting nitric oxide with natural derived compounds as a therapeutic strategy in vascular diseases. Oxid Med Cell Longev. 2016;2016:7364138. doi:10.1155/2016/7364138
132. Pradubyat N, Wunnakup T, Praparatana R, et al. Evaluation of antioxidant and anti-inflammatory properties, bioactive compound profiling, and molecular mechanisms of a multicomponent Thai herbal formulation. Phytomed Plus. 2024;4(4):100662. doi:10.1016/j.phyplu.2024.100662
133. Choy KW, Murugan D, Leong XF, Abas R, Alias A, Mustafa MR. Flavonoids as natural anti-inflammatory agents targeting nuclear factor-kappa B (NFκB) signaling in cardiovascular diseases: a mini review. Front Pharmacol. 2019;10. doi:10.3389/fphar.2019.01295
134. Serafini M, Peluso I, Raguzzini A. Flavonoids as anti-inflammatory agents. Proc Nutr Soc. 2010;69(3):273–278. doi:10.1017/S002966511000162X
135. Cunha TM, Roman-Campos D, Lotufo CM, et al. Morphine peripheral analgesia depends on activation of the PI3Kγ/AKT/nNOS/NO/K ATP signaling pathway. Proc Natl Acad Sci. 2010;107(9):4442–4447. doi:10.1073/pnas.0914733107
136. Sachs D, Cunha FQ, Ferreira SH. Peripheral analgesic blockade of hypernociception: activation of arginine/NO/cGMP/protein kinase G/ATP-sensitive K + channel pathway. Proc Natl Acad Sci. 2004;101(10):3680–3685. doi:10.1073/pnas.0308382101
137. Salaritabar A, Darvishi B, Hadjiakhoondi F, et al. Therapeutic potential of flavonoids in inflammatory bowel disease: a comprehensive review. World J Gastroenterol. 2017;23(28):5097. doi:10.3748/wjg.v23.i28.5097
138. Zhang S, Gao L, Liu X, Lu T, Xie C, Jia J. Resveratrol attenuates microglial activation via SIRT1‐SOCS1 pathway. Evid Based Complement Altern Med. 2017;2017(1). doi:10.1155/2017/8791832
139. Dragone T, Cianciulli A, Calvello R, Porro C, Trotta T, Panaro MA. Resveratrol counteracts lipopolysaccharide-mediated microglial inflammation by modulating a SOCS-1 dependent signaling pathway. Toxicol Vitr. 2014;28(6):1126–1135. doi:10.1016/j.tiv.2014.05.005
140. Shin MH, Kim J, Lim SA, Kim J, Lee KM. Current insights into combination therapies with MAPK inhibitors and immune checkpoint blockade. Int J Mol Sci. 2020;21(7):2531. doi:10.3390/ijms21072531
141. Zhou Y, Gu C, Zhu Y, et al. Pharmacological effects and the related mechanism of scutellarin on inflammation-related diseases: a review. Front Pharmacol. 2024:15. doi:10.3389/fphar.2024.1463140.
142. Zheng Y, Wei K, Jiang P, et al. Macrophage polarization in rheumatoid arthritis: signaling pathways, metabolic reprogramming, and crosstalk with synovial fibroblasts. Front Immunol. 2024:15. doi:10.3389/fimmu.2024.1394108.
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Etlingera Elatior Inflorescence Extract Mitigates Acute Gastric Ulcers by Suppressing the Expression of Inducible Nitric Oxide Synthase in Ethanol-Induced Wistar Rats
Prayoga DK, Aulifa DL, Budiman A, Levita J, Jiranusornkul S
Journal of Experimental Pharmacology 2025, 17:343-357
Published Date: 17 June 2025