Back to Journals » Journal of Experimental Pharmacology » Volume 17
Etlingera Elatior Inflorescence Extract Mitigates Acute Gastric Ulcers by Suppressing the Expression of Inducible Nitric Oxide Synthase in Ethanol-Induced Wistar Rats
Authors Prayoga DK, Aulifa DL ![]() , Budiman A, Levita J
, Budiman A, Levita J ![]() , Jiranusornkul S
, Jiranusornkul S
Received 17 March 2025
Accepted for publication 7 June 2025
Published 17 June 2025 Volume 2025:17 Pages 343—357
DOI https://doi.org/10.2147/JEP.S524517
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Prof. Dr. Abdelwahab Omri
Deshanda Kurniawan Prayoga,1,* Diah Lia Aulifa,2,* Arif Budiman,3,* Jutti Levita,4,* Supat Jiranusornkul5,*
1Doctoral Program in Pharmacy, Faculty of Pharmacy, Padjadjaran University, Sumedang, West Java, Indonesia; 2Department of Pharmaceutical Analysis and Medicinal Chemistry, Faculty of Pharmacy, Padjadjaran University, Sumedang, Indonesia; 3Department of Pharmaceutics and Pharmaceutical Technology, Faculty of Pharmacy, Padjadjaran University, Sumedang, Indonesia; 4Department of Pharmacology and Clinical Pharmacy, Faculty of Pharmacy, Padjadjaran University, Sumedang, Indonesia; 5Department of Pharmaceutical Science, Faculty of Pharmacy, Chiang Mai University, Chiang Mai, Thailand
*These authors contributed equally to this work
Correspondence: Jutti Levita, Department of Pharmacology and Clinical Pharmacy, Faculty of Pharmacy, Padjadjaran University, Sumedang, West Java, 46363, Indonesia, Email [email protected]
Background: Ethanol consumption by oral route can induce gastric cell necrosis and vascular damage due to excessive reactive oxygen species (ROS) production. ROS activates the translocation of NF-κB to the nucleus and increases iNOS expression, an enzyme that catalyzes nitric oxide (NO) production, eventually disrupting gastric protective mechanisms and contributing to ulcer formation. Etlingera elatior inflorescence, which has been traditionally used to alleviate stomach discomfort, was reported to possess anti-inflammatory activity. However, such a study on the downregulation of the iNOS expression is lacking.
Purpose: To confirm the gastro-protective and anti-ulcer effects of the E. elatior inflorescence (EEIE) extract via downregulation of iNOS expression in ethanol-induced gastric ulcer rats.
Methods: The fresh inflorescence petals were collected from West Java, Indonesia, and were extracted using 70% ethanol. The effects of 5 days of oral administration of EEIE were studied in ethanol-induced adult male Wistar rats by examining the weight gain percentage, feed residue, stomach ratio index, and microscopic and macroscopic evaluation of gastric mucosal damage. Subsequently, the iNOS expression in the rats’ stomach was Western blotted by employing β-actin as the loading control.
Results: EEIE reduced weight gain percentage but did not show a significant difference in feed residue. EEIE increased the stomach ratio index, and at a dose of 625 mg/kg BW, significantly reduced the ulcer index, providing 100% protection (p < 0.05) to rats while decreasing inflammatory cell infiltration. EEIE inhibited the expression of iNOS in both cleaved and full-length forms at this dose.
Conclusion: Etlingera elatior inflorescence may have the potential to mitigate acute gastric ulcers by downregulating the expression of iNOS in ethanol-induced Wistar rats. However, further studies are needed to confirm its role in alleviating chronic ulcer models.
Keywords: acute inflammation, etlingera, gastric ulcers, iNOS, medicinal plants, zingiberaceae
Introduction
Gastric ulcer disease is a significant global health issue affecting millions of people. The lifetime prevalence in the general population is estimated to be between 5% and 10%, with rates as high as 20% among adults in Asia.1–6 According to a survey, ulcers are formed 3–4 times more likely in men than women, primarily at older ages.7 Geographical variations in gastric ulcer-related mortality have confirmed the risk factors such as genetics, Helicobacter pylori infection, smoking, excessive alcohol consumption, long-term use of non-steroidal anti-inflammatory drugs (NSAIDs), and Zollinger–Ellison syndrome. Although severe complications for gastric and duodenal ulcers were evidenced to be caused by H. pylori and NSAIDs, alcohol, when over-consumed, could increase acid secretion and lead to the impairment of the stomach’s protective barrier.8,9
Ethanol is absorbed by the gastric cells and is metabolized in the liver by cytochrome P450 subfamily 2E1 (CYP2E1), which is involved in the production of reactive oxygen species (ROS), predominantly via electron leakage to oxygen to form the superoxide (O2•−) radical.10,11 Excessive ROS production, including ONOO−, NO•,2•− H2O2, HO•, and HClO, triggers oxidative stress, which contributes to gastric ulcers, progressive intestinal mucosa damage and exacerbates tissue inflammation, while prolonged neutrophil activation further accelerates the recruitment of monocytes, which differentiate into macrophages at the injury site, perpetuating the inflammatory response.12,13 Ethanol-induced ROS activates the nuclear translocation of NF-κB p65, a key transcription factor in inflammation, thus, via a transcription process in the nucleus, leads to the increase of iNOS expression and, subsequently, excessive nitric oxide (NO) production.14–16 While NO plays a role in immune defense, its overproduction due to unchecked NF-κB activation exacerbates oxidative stress, promoting mucosal damage instead of healing.17–19 This excessive NO disrupts gastric protective mechanisms, contributing to ulcer formation and inflammation.20,21
The management of gastric ulcer disease often relies on conventional treatments, though these can have significant limitations.22–25 For instance, discontinuing proton pump inhibitors (PPIs) may lead to rebound acid hypersecretion (RAHS), where acid production exceeds pre-treatment levels.23,26,27 Although antihistamine H2 receptor and PPI therapies are well-established for treating gastric ulcers, they do not prevent recurrence and may cause drug interactions in some patients.22,25,26 Patients undergoing long-term therapy with antibiotics may develop increased resistance of H. pylori, due to bacterial genetic mutations that alter the antibiotics or inhibit drug activation in the cells.28 As a result, interest in alternative therapies from natural sources is increasing.
The World Health Organization (WHO) reports that 80–85% of the global population depends on natural products for health management.29–32 Etlingera elatior, known as torch ginger, is traditionally used to relieve stomach discomfort and has been studied for treating stomach ulcers.33,34 E. elatior inflorescence has been reported for its bioactive phytoconstituents, including anthocyanins, flavonoids (quercetin), terpenoids, steroids, tannins, glycosides, and saponins.35–37 Flavonoids such as quercetin have shown radical scavenging activity towards ROS by reducing the violet 2,2-diphenyl-1-picrylhydrazyl (DPPH) to the yellow DPPH-H.38 Furthermore, quercetin suppressed the nuclear factor-kappa B (NF-κB) signaling pathway,39,40 inhibited cyclooxygenase-2 (COX-2),41 and regulated mitogen-activated protein kinases (MAPKs), which are crucial mediators of inflammation,42 and gastroprotective benefits.42
The phytoconstituents of E. elatior can bind with critical amino acids on iNOS (using PDB ID: 4NOS) based on an in silico study, and moderately inhibit NO production in an in vitro study, with an IC50 = 24.714 µg/mL, respectively.43 Furthermore, in vivo gastric model studies delineated that E. elatior flower extract inhibited inflammation related to gastric ulcers by downregulating NF-κB-p65 expression in Wistar rats with acetic-acid-induced ulcers,36 and E. elatior fruit extract significantly reduced IL-1ß and caspase-3 (p < 0.05) levels in sepsis model Balb/C mice.44 Considering that inhibition of NF-κB likely reduces iNOS expression, as NF-κB is a primary transcription factor that stimulates iNOS during inflammation, and there is a lack of reports on iNOS inhibitory activity by E. elatior, this study aims to investigate the gastro-protective and anti-ulcer effects of the ethanol extract of E. elatior inflorescence (EEIE) via downregulation of iNOS expression in ethanol-induced acute gastric ulcer rats. Moreover, it supports evidence of the pharmacological role of natural compounds in inflammatory disease management.
Material and Methods
Preparation of E. Elatior Inflorescence Extract (EEIE)
The extraction process was performed by modifying a previously described procedure.36,43,45 The fresh Etlingera elatior inflorescence petals were collected in West Java, Indonesia, and were authenticated by Arifin Surya Dwipa Irsyam (https://www.scopus.com/authid/detail.uri?authorId=57211286941; https://herbarium.sith.itb.ac.id/profil-kurator/) at the School of Life Sciences and Technology, Bandung Institute of Technology, Bandung, West Java, Indonesia (https://herbarium.sith.itb.ac.id/koleksi/). The materials were verified as Etlingera elatior (Jack) R.M.Sm. (Zingiberaceae) (document number 74/IT1.C11.2/TA.00/2024), with characteristics that align with those documented in prior references. The plant materials were thoroughly sorted to exclude damaged parts. The selected petals were washed under running water to remove dirt and other contaminants, dried in the oven at 45–50°C for 24 h, ground, and sieved using a Mesh-12 sieve. The coarse powder (1300 g) was soaked in 70% ethanol for 24 h at room temperature at a 1:10 (w/v) ratio. The extract was filtered, and the residue was extracted repeatedly for 3×24 h. The solvent was evaporated in a vacuum rotary evaporator (Büchi) at 50 °C until a viscous extract of EEIE was yielded (20.2% w/w).
Animal Handling and Ethical Approval
Twenty-five male Wistar albino rats weighing 200–220 g, aged 6–8 weeks, were bred in the Animal Facility of the School of Life Sciences and Technology, West Java, Indonesia. In this study, we used male rats, which are known for their more consistent behavior compared to females, particularly when considering the hormonal fluctuations associated with the female estrous cycle. Interestingly, even female rats without an active estrous cycle exhibit behaviour that is comparable in reliability to male rats, suggesting that their variability does not exceed that of males.46
The rats were transferred to the Universitas Padjadjaran Animal Hospital, randomly divided into five groups, and acclimatized in cages (32 × 22×16 cm3, 5 animals per cage) in a room set under a standard laboratory environment (25–27°C with a 12-h-light/12-h-dark cycle) for approximately 5 days before treatment. They were given standard animal feed (containing carbohydrates, vegetable protein, and vegetable fat) and water freely during the acclimatization and experimental periods. The health condition and behavior of the rats were observed and recorded daily. All the experimental protocols strictly followed The Guide for the Care and Use of Laboratory Animals (NRC 2011; eighth edition) (https://grants.nih.gov/grants/olaw/guide-for-the-care-and-use-of-laboratory-animals.pdf) and are approved by the Ethics Committee, Faculty of Medicine, Universitas Padjadjaran, Indonesia (Document No. 641/UN6.KEP/EC/2024).
Experimental Study Design and Treatment
The in vivo study in Wistar rats was performed as follows: (1) the normal group (treated with standard feed and sodium carboxymethyl cellulose 0.5% suspension); (2) the negative control (was ethanol-induced and treated with sodium carboxymethyl cellulose 0.5% suspension); (3) the positive control (was ethanol-induced and treated with quercetin 20 mg/kg BW suspended in sodium carboxymethyl cellulose 0.5%, orally by using oral gavage feeding);47 (4) the treatment group dose 1 (was ethanol-induced and given EEIE dose of 375 mg/kg BW suspended in sodium carboxymethyl cellulose 0.5%, orally by using oral gavage feeding); and (5) the treatment group dose 2 (was ethanol-induced and given EEIE dose of 625 mg/kg BW suspended in sodium carboxymethyl cellulose 0.5%, orally by using oral gavage feeding). After 5 days of intervention, the rats in groups 2 to 5 were given 1 mL of 70% ethanol orally to induce acute gastric ulcers.36,48,49 One hour after the induction, the rats were sacrificed using 0.3 mL ketamine anesthesia intraperitoneally by trained personnel, and death was confirmed by determining cardiac and respiratory arrest.50,51 The stomach of each rat was excised, removed, weighed, rapidly frozen, and stored at −80°C for further use.52 The ratio of the organs per BW of the rats was calculated.
Weight Gain Evaluation
The initial body weight of each rat was recorded and weighed every day until the end of the experimental period to obtain the final body weight. The percentage of weight gain was calculated using the following formulas:
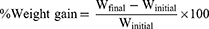
Feed Residues Evaluation
Feed residue was collected throughout the experimental period. The predetermined amount of feed was weighed and provided to each animal. After 24 hours, the remaining uneaten feed was carefully collected and weighed; this process was repeated daily.53
Stomach Index Evaluation
The stomach of rats of each group was collected, weighed, washed with water, followed by cold phosphate buffer saline, and dried using filter paper, to assess potential pathological alterations resulting from the ethanol-induction.36,52 The stomach index was calculated using a formula: Stomach index (g/kg) = stomach weight (g)/BW (kg) × 1000, and compared to that of the negative control group.
Macroscopic Evaluation of Gastric Mucosal Damage
The stomach was exposed and opened longitudinally to score the gastric damage.54 The ulcer index was measured using the following scores described by Reddy et al (2012), as follows:55
A score of 0.0 represents a normal stomach; a score of 0.5 represents a red coloration; a score of 1.0 represents ulcer spots; a score of 1.5 represents a hemorrhage streak; a score of 2.0 represents deep ulcers; and a score of 3.0 represents perforation.
The ulcer index was calculated using the following equation:

Where the UI is the ulcer index, UN is the average number of ulcers per animal, and US is the average severity score.
Moreover, the % protection was calculated using the following equation:

Microscopic Evaluation of Gastric Mucosal Damage
The histopathology tissue preparation process of the stomach includes the stages of fixation, dehydration, clearing, infiltration, embedding, sectioning, and mounting on glass slides as follows: the stomach of the rats was fixed with 10% v/v buffered formalin solution (containing 0.01 M phosphate buffer saline at pH 7.2), subsequently sliced to 10 µM thickness, embedded in paraffin, and was stained using hematoxylin and eosin to observe the infiltration of inflammatory cells under a light microscope (Olympus CX21) coupled with a microscope camera (OptiLab Microscope Digital), with 40x and 100x magnifications. The number of inflammatory cells was calculated per 1000x objective lens visibility.52,56,57
Western Blot Analysis
The Western blot analysis was performed following the procedure outlined in the iNOS antibody (NB300-605, Novus-Biologicals) manual instructions and according to Rosdianto et al, with modifications.52 The dissected stomach of the rats was weighed and extracted in lysis buffer (containing Tris-HCl solution to pH 7.6, 150 mm NaCl, 1% NP-40, 1% sodium deoxycholate, 0.1% SDS) and sample buffer (Tris-HCl solution pH 6.8, glycerol, β-mercaptothion, bromophenol blue, distilled water, and SDS) resulting in a 25 g of each gastric protein. The proteins were heat-denatured at 95°C for 5 min, poured into a 10% polyacrylamide gel, and run at 75 V for 2 h. The sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) separated the proteins (12 µg/lane) and was transferred to a nitrocellulose membrane for 30 min at room temperature, and blocked for 1 h in 5% skim milk. The membrane was incubated with iNOS polyclonal antibody (Novus Biologicals, diluted in 1:200), β-actin antibody HRP conjugate (ThermoFisher Scientific, diluted in 1:1000 in phosphate-buffered saline 0.1% Tween® 20), washed, and re-incubated with secondary antibody, IRDye® 800CW goat anti-rabbit and IRDye® 800CW goat anti-mouse (LICORBio, diluted in 1:15000 in TBST). The signals were developed using enhanced chemiluminescence reagent (GE Healthcare), and the band intensities were determined using ImageJ software (https://imagej.nih.gov/ij/). Bands were stripped and re-probed using an anti-β-actin mouse monoclonal antibody as the internal control to monitor the protein level.
Statistical Analysis
The in vivo data were presented as mean ± SD (n = 5) and statistically analyzed using IBM SPSS Statistics 26.0 for Windows 11. Power analysis was conducted with G*Power 3.1.9.7, using an effect size (dz) of 0.6, α error probability of 0.05 (5%), and a power (1-β error probability) of 0.8. The significance level was set at 5% (p = 0.05), though this value can be adjusted depending on the research question. The effect was assessed using a two-tailed approach.58,59 The total sample size calculated was 24. However, for hypothesis testing, the sample size (calculated by Federer’s formula) should generally be equal to or greater than the required sample size (power size) to ensure adequate statistical power.60
Data homogeneity was evaluated using Levene’s test. The % weight gain was statistically analyzed using a paired t-test with a p-value of 0.05, indicating a significant result. Significant differences between groups were analyzed using one-way analysis of variance (ANOVA) with a p-value of < 0.05 indicating a significant result, followed by the Dunnett post hoc test or Games-Howell (if the data was not homogeneous) (CI 95%). Data were visualized using the GraphPad Prism Free trial version.
Results
Effects of EEIE on Weight Gain and Feed Residue of Ethanol-Induced Wistar Rats
At the end of the study (D10), rats in all groups demonstrated an increase in body weight (BW) with % weight gain values ranging from 1.55% for rats treated with an EEIE at a dose of 625 mg/kg BW to the highest value of 8.69% for rats in the negative control group or placebo. Ethanol oral inducement resulted in a remarkable alteration in the BW. It is confirmed that treating the rats with quercetin at 20 mg/kg BW and all doses of EEIE resulted in a significantly lower % weight gain (p < 0.05) compared to the rats in the negative control group (Table 1).
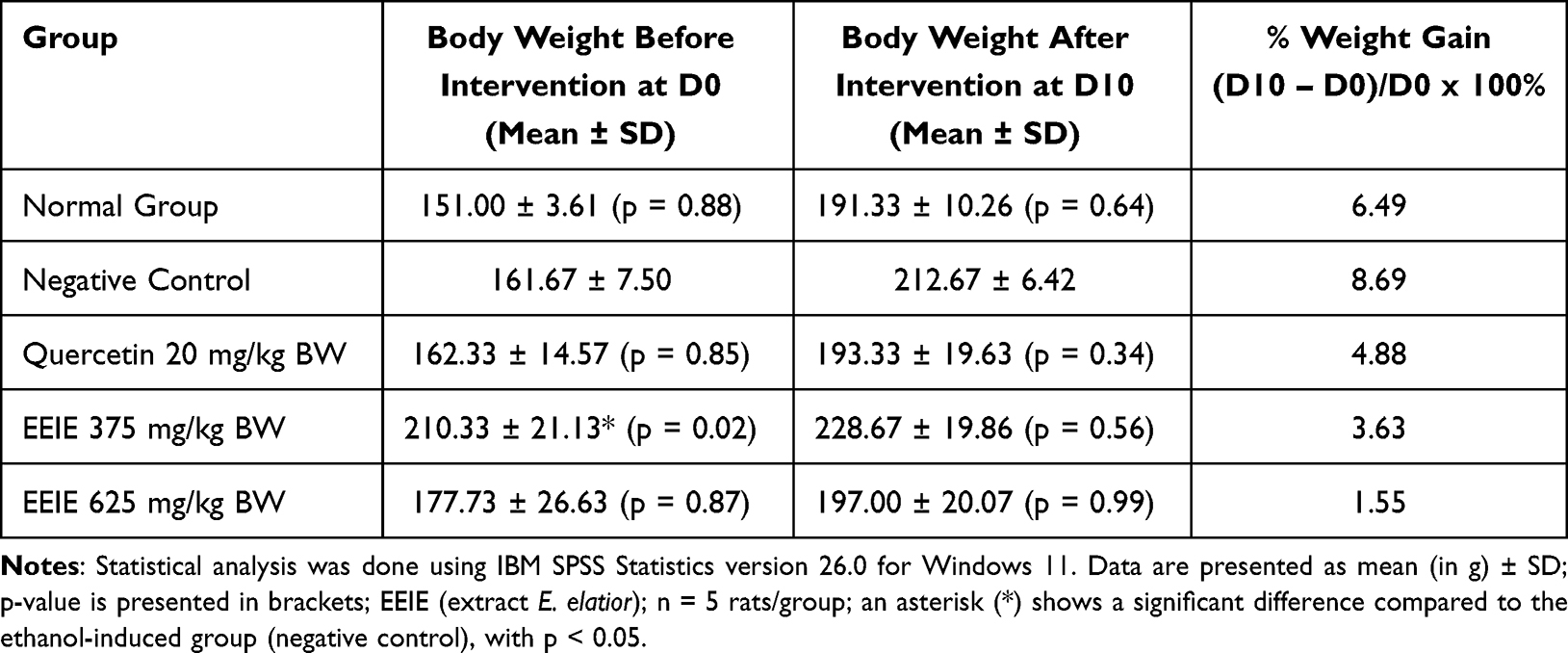 |
Table 1 Effects of EEIE on Weight Gain (%) of Ethanol-Induced Wistar Rats |
The feed residue observation (Figure 1 and Table 2) revealed a decrease in feed residue, ranging from 21.47 to 41.86% in all groups, with the highest % decrease belonging to the group of rats treated with EEIE at a dose of 625 mg/kg BW. Nevertheless, no significant difference is found compared to the negative control group, indicating that EEIE may slightly alter the appetite or intake patterns of the animals. It should be taken into noted that on D6 and D7, rats treated with EEIE at a dose of 625 mg/kg BW have finished off their feed, similar to those treated with quercetin 20 mg/kg BW (Figure 1), demonstrating a temporary increase in appetite. However, these differences in short-term consumption did not lead to significant overall changes in feed residue across the study period.
 |
Table 2 Effects of EEIE on the Feed Residue of Ethanol-Induced Wistar Rats |
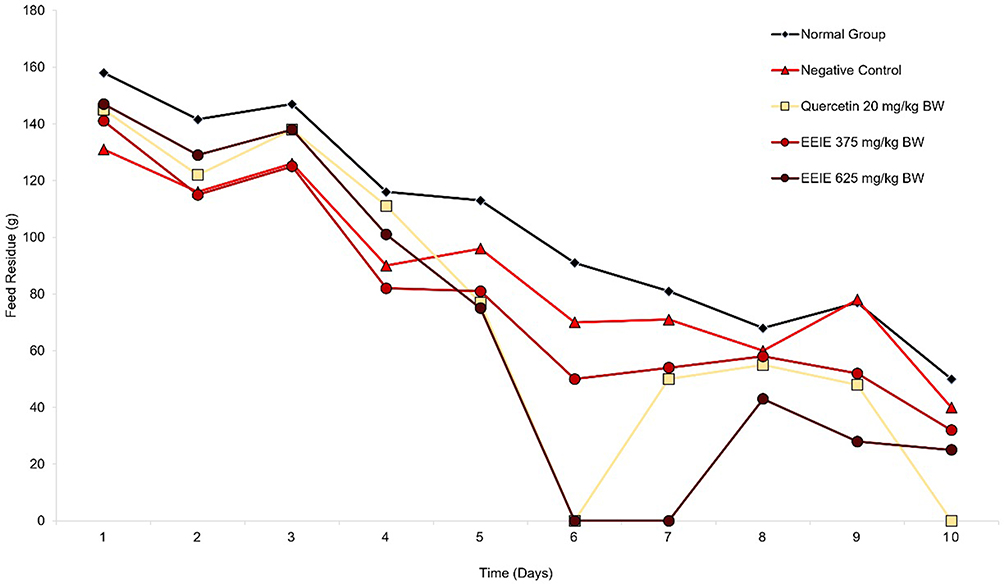 |
Figure 1 Effects of EEIE on the feed residue of ethanol-induced Wistar rats for 10 days. |
Effects of EEIE on Stomach Index, Ulcer Index, and % Protection of Ethanol-Induced Wistar Rats
The stomach index was calculated by measuring the weight of the stomach in each group of rats relative to the BW. The effect of various doses of EEIE (p = 0.003 for EEIE dose of 375 mg/kg BW, p = 0.005 for EEIE dose of 625 mg/kg BW) on the stomach index of ethanol-induced rats (Table 3) indicated significant differences compared to that of the negative control group. Similarly, quercetin at a dose of 20 mg/kg BW significantly increased the stomach index of the ethanol-induced rats. These results suggest that ethanol-induced gastric injury leads to significant changes in the stomach weight, as indicated by the decreased stomach index (p < 0.01).
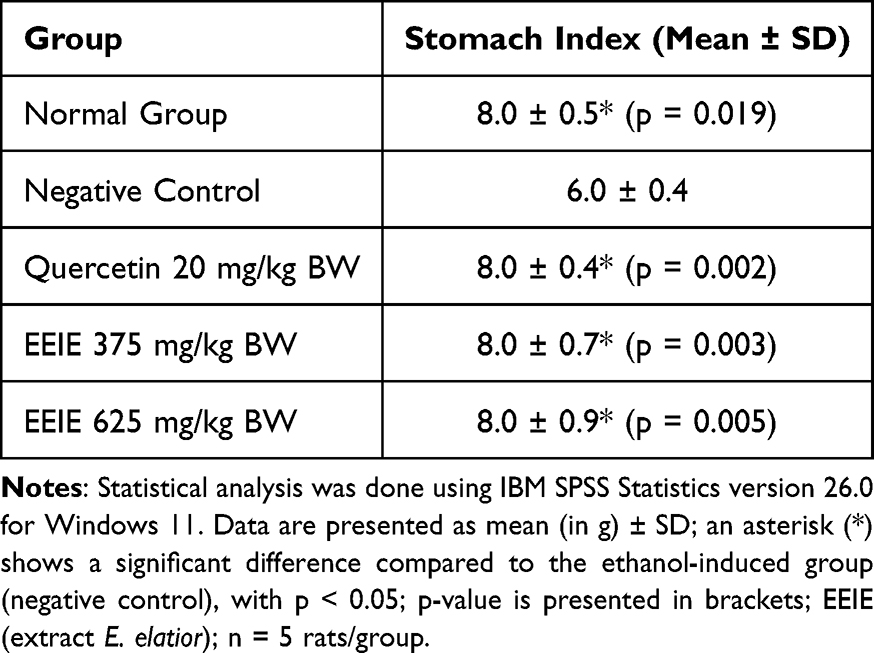 |
Table 3 Effects of EEIE on the Stomach Index of Ethanol-Induced Wistar Rats |
Treatment with 70% ethanol (5 mL/kg BW) caused superficial or deep erosions and bleeding in the stomach, with the ulcer index of 14.50 ± 0.44 (p = 0.001). The effects of EEIE on the ulcer index and % protection of ethanol-induced rats (Table 4) indicated that only the EEIE dose of 625 mg/kg BW and quercetin 20 mg/kg BW significantly inhibited gastric ulcer, as shown by the lowest ulcer index and 100% protection. EEIE dose of 375 mg/kg BW indicated 23.27% protection.
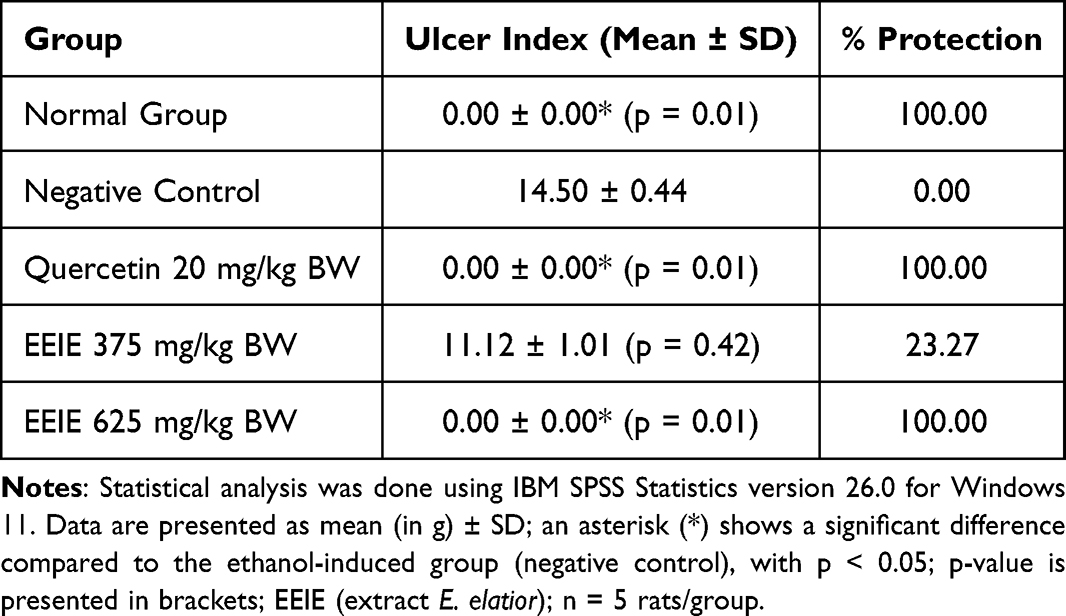 |
Table 4 Effects of EEIE on the Ulcer Index and % Protection of Ethanol-Induced Wistar Rats |
Effects of EEIE on Macroscopic and Microscopic Gastric Mucosal Damage
Significant tissue damage and visible hemorrhagic mucosal lesions were macroscopically observed in the stomach of the ethanol-treated rats (the negative control group). The ethanol-induced rats treated with quercetin at 20 mg/kg BW and an EEIE dose of 625 mg/kg BW did not show ulcers (Figure 2). In contrast, treatment with an EEIE dose of 375 mg/kg BW did not give satisfying results, as proven by the presence of a few ulcers.
 |
Figure 2 Effects of EEIE on macroscopic observation of the gastric mucosa of ethanol-induced Wistar rats: normal group (upper left), negative control group (upper middle), quercetin 20 mg/kg BW group (upper right), EEIE 375 mg/kg BW (lower left), and EEIE 625 mg/kg BW (lower right). The ulcer spots are shown by black arrows. |
Microscopic evaluation exhibited the presence of inflammatory cells in all the gastric mucosa of all ethanol-induced rats (Table 5). Interestingly, rats in the normal group also showed the presence of a few inflammatory cells (3.13 ± 0.67), although statistically, there were no significant differences in the number of inflammatory cells in each group compared to the negative control, which is probably caused by the small sample size.
 |
Table 5 Effects of EEIE on the Number of Inflammatory Cell Infiltration in the Gastric Tissue of Ethanol-Induced Wistar Rats |
Effects of EEIE on the Expression of iNOS by Western Blot Analysis
The effects of EEIE on the expression of inducible nitric oxide synthase (iNOS) are presented in Figure 3. Oral inducement with ethanol stimulates the expression of full-length iNOS at 130 kDa, as shown in the negative control. Interestingly, the expression of cleaved iNOS at 70 kDa is observed in all groups of rats, including the normal control. Only EEIE at a dose of 625 mg/kg BW, not the dose of 375 mg/kg BW, reduced the expression of both cleaved and full-length iNOS, although not statistically significant due to the small sample size. The inhibitory activity of EEIE at a dose of 625 mg/kg BW towards iNOS expression is stronger than that of quercetin. Quercetin (the positive control or standard drug) at 20 mg/kg BW suppressed the expression of both cleaved and full-length iNOS, but not significant. Densitometric analysis of the Western blot bands is presented in Figure 4.
 |
Figure 3 The effects of EEIE (a) on the relative expression of cleaved iNOS (70 kDa) in the gastric mucosa of ethanol-induced Wistar rats normalized by beta-actin; (b) on the relative expression of full-length iNOS (130 kDa) in the gastric mucosa of ethanol-induced Wistar rats normalized by beta-actin; (c) the Western blot electropherogram cleaved bands of iNOS (70 kDa) and the full-length bands of iNOS (130 kDa) normalized by beta-actin. Statistical analysis was done using IBM SPSS Statistics version 26.0 for Windows 11. The * indicates a significant difference compared with the negative control (p < 0.05), and ** indicates a significant difference compared with the negative control (p < 0.01) (calculated by one-way ANOVA followed by post hoc Dunnett test). 1 = Normal Control; 2 = Negative Control; 3 = Quercetin 20 mg/kg BW; 4 = EEIE at a dose of 375 mg/kg BW; 5 = EEIE at a dose of 625 mg/kg BW. |
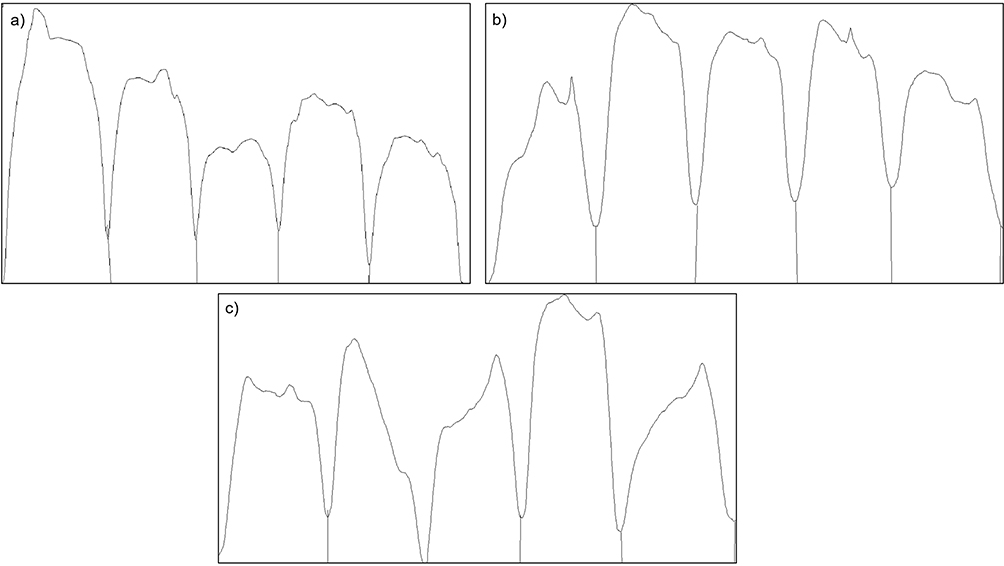 |
Figure 4 The densitogram of the Western blot bands of (a) β-actin, (b) cleaved iNOS, and (c) full-length iNOS. The bands were analyzed using ImageJ (https://imagej.net/ij/). |
Discussion
The present study aims to investigate the effects of Etlingera elatior extract on iNOS in ethanol-induced acute gastric injury in rats. Excessive ethanol consumption is considered one of the risk factors for gastric ulcers in humans; its use in experimental animals allows for the evaluation of cytoprotective activity. Ethanol can damage the stomach and readily penetrate and digest the gastric wall through proteolytic and hydrolytic actions, generating ROS and reducing blood circulation.54,61 The ethanol-induced gastric model is used in animals to study and develop new anti-ulcer medications.62 Ethanol exposure leads to necrotic damage in the gastric tissues, followed by infiltration of inflammatory cells. It also disrupts the protective mechanism of the stomach by decreasing the secretion of bicarbonate and mucus. Additionally, ethanol impairs gastric blood flow and triggers oxidative stress by elevating malondialdehyde levels while depleting glutathione reserves.56,63,64 With previous findings, administration of 80% ethanol solution by intragastric gavage produced marked damage in the gastric mucosa of rats, characterized mainly by elongated macroscopic lesions with intense hemorrhaging and hyperemia, as well as loss of mucus.65
Ethanol has been shown to increase the expression of iNOS while decreasing the gene expression of eNOS in gastric tissue. The elevated levels of NO, whose production is catalyzed by iNOS, contribute to tissue damage in the gut under inflammatory conditions.66 Previous findings by Nagai et al highlighted that NO generated by iNOS plays a crucial role in developing gastric ulcers.67 The activation of iNOS expression has been linked to gastric ulcers and chronic ulcerative colitis in affected patients, indicating that excessive NO production can adversely affect the progression of these conditions.18,68
Our study found that EEIE can effectively maintain BW during a five-day intervention, indicating a significant impact on controlling weight gain. Additionally, C3G, a phytoconstituent in EEIE, has been described to possess anti-obesity properties, leading to lower consumption levels than the control group.69 However, this weight reduction alone is insufficient to classify it as “anti-obesity”; robust, long-term evidence demonstrating substantial and consistent decreases in the body mass index (BMI) is necessary. Furthermore, differences in short-term consumption did not lead to significant changes in feed residue throughout the study period. These findings are consistent with previous work by Ruiz et al, who delineated that high doses of quercetin did not affect the feeding behavior of the rats, nor did they disrupt overall feed intake.70 Consequently, any physiological or metabolic changes observed in the intervention groups cannot be attributed to variations in food consumption. Instead, the physiological alterations, such as reduced weight gain and metabolic improvements noted in the intervention group, were primarily linked to the specific effects of C3G rather than differences in feed intake. The absence of significant discrepancies in feed consumption further supports the notion that the intervention directly influenced metabolic processes without impacting appetite or feeding behavior.69
Macroscopic analysis of the gastric tissues provided further evidence of the protective effects of EEIE. In the ethanol-treated group, extensive mucosal damage was observed, characterized by elongated hemorrhagic lesions, hyperemia, and a significant loss of mucus. Ethanol administration can increase ROS levels while depleting glutathione (GSH) levels in the gastric mucosa.71 The gastroprotective activity of EEIE is attributed to its phytoconstituents, which exhibit strong antioxidant properties. The antioxidant activity of flavonoids, particularly quercetin, primarily arises from the presence of the o-dihydroxy ring on ring B (catechol), which is further enhanced by the conjugation of double and triple bonds with the 4-oxo function. Additionally, the hydroxyl groups at positions 3, 5, and 7 of its structure significantly contribute to its pharmacological effects.72,73
Furthermore, C3G plays an important role in the gastroprotective effects of EEIE. Research has shown that C3G and its bioactive phenolic metabolites, including protocatechuic acid (PCA), vanillic acid (VA), and ferulic acid (FA), can enhance the antioxidant enzyme system by increasing the activities of manganese-dependent superoxide dismutase (MnSOD) and glutathione (GSH).74,75 Cyanidin and its derivatives demonstrate antioxidant effects mainly through their ability to donate hydrogen atoms or electrons, thereby neutralizing free radicals. The diphenyl structure in the B ring is crucial for this activity, while hydroxyl groups at specific positions, such as 3′ and 4′, further enhance the antioxidant capacity by stabilizing free radicals.76,77
The present study showed that EEIE at doses of 375 and 625 mg/kg reduced the histopathologic changes and the number and size of gastric ulcers induced by ethanol in the rats. However, the analysis of stomach tissue showed some overall reduction of damage, although not significant. Neutrophils, a type of immune cell, infiltrated the stomach of rats in the negative control group, positive control group, and those treated with EEIE at a dose of 675 mg/kg, suggesting the occurrence of inflammation. In addition, our results showed that quercetin at a dose of 20 mg/kg caused a slight inflammation in the gastric mucosa. This aligns with previous studies that described that higher doses of quercetin could significantly lower cell survival.36,52,78 The granulocyte neutrophils are produced in the bone marrow and, when activated by various cytokines, migrate through the vascular endothelium to the site of inflammation.79 Once activated, they release their granule proteins. These granulocyte neutrophils are also referred to as polymorphonuclear neutrophils or polymorphs.80
The Western blot experiment confirmed that iNOS was activated in ethanol-induced rats, and EEIE had reduced the expression of this enzyme, consistent with previous studies indicating that ethanol exposure leads to oxidative stress and inflammatory responses in the gastric mucosa. Ethanol stimulates the expression of full-length iNOS at 130 kDa, as shown in the negative control. Interestingly, the expression of cleaved iNOS at 70 kDa is observed in all groups of rats, including the normal control. Only EEIE at a dose of 625 mg/kg BW, not the dose of 375 mg/kg BW, reduced the expression of both cleaved and full-length iNOS, although not statistically significant, suggesting the potential protective role in gastric mucosal injury.
The NF-κB signaling pathway, which is crucial in iNOS gene transcription, most likely contributes to the activation of iNOS expression.81,82 Our findings demonstrated that ethanol exposure significantly upregulated iNOS expression; the 130 kDa band corresponds to the full-length, active form of iNOS, while the 70 kDa band represents a cleaved form of the enzyme that contributes to oxidative stress and inflammatory damage in gastric tissues. Interestingly, immunoblotting analysis revealed a predominant iNOS band at 70 kDa, which suggests a cleaved version of the enzyme rather than the expected full-length 130 kDa protein.
Previous reports indicate that iNOS can undergo proteolytic cleavage, resulting in fragments around 70 kDa. Such cleavage often occurs near the calmodulin-binding site, affecting the enzyme’s activity and stability.83 This observation highlights a potential instability or degradation of iNOS under pathological conditions, which may have functional implications in ethanol-induced gastric injury.
The full-length and cleaved iNOS forms suggest a dynamic regulation of iNOS activity in response to ethanol-induced gastric injury. However, in this study, EEIE is shown to inhibit both forms. The dual inhibition of both forms raises questions about the balance of NO signaling, while inhibiting the cleaved form may reduce harmful effects, it is essential to consider how this affects the overall regulation of NOS and the physiological outcomes. However, EEIE may be affecting the translation process of iNOS, leading to reduced levels of the full-length protein. This could be due to a direct effect on the transcription, resulting in decreased synthesis of iNOS overall.84 Furthermore, the degradation of iNOS can be influenced by various factors, including specific inhibitors that promote the formation of the monomeric form of iNOS, which is more susceptible to ubiquitination and subsequent degradation.85 By reducing the levels of iNOS, E. elatior may eventually decrease NO production, thereby limiting excessive inflammation and promoting a more stable gastric environment,86 thus leading to reduced ulcer formation and improved healing of existing ulcers.
Our findings are in agreement with previous studies demonstrating iNOS under pathological conditions. For example, a report indicated that naringenin, a flavonoid compound, significantly reduced the protein expression of iNOS in gastric tissues exposed to ethanol.82 Taken together, the mechanism of EEIE against ethanol-induced gastric injury by downregulating proinflammatory mediators, including NF-κB and iNOS, also enhances antioxidant defenses by increasing superoxide dismutase (SOD) and decreasing malondialdehyde (MDA) content. Nevertheless, despite the promising findings, our study has certain limitations. The mechanism underlying iNOS degradation and its impact on NO bioavailability remains fully unelucidated. Therefore, investigating the EEIE interactions with other pro-inflammatory mediators, cytokines, and chemokines, such as COX-2, prostaglandins, interleukins, and tumor necrosis factors, will provide a broader understanding of its therapeutic potential.
Conclusion
Our findings revealed that oxidative stress and inflammation may play critical roles in the pathogenesis of ethanol-induced acute gastric ulcers. Oral exposure to 70% ethanol significantly upregulated iNOS expression, as shown by the 130 kDa band that corresponds to the full-length, active form of iNOS, while the 70 kDa band represents a cleaved form of the enzyme that contributes to oxidative stress and inflammatory damage in gastric tissues. Pretreatment with the ethanol extract of Etlingera elatior inflorescence (EEIE) has demonstrated potential anti-inflammatory activity by inhibiting the development of gastric ulcers through the suppression of both forms of inducible nitric oxide synthase (iNOS). While inhibiting the cleaved form of iNOS may reduce harmful effects, it is essential to consider how this affects the overall regulation of NOS and the physiological outcomes. Additionally, EEIE did not significantly alter the BW but significantly increased the stomach index, similar to that of quercetin, which further confirms its safety. This research contributes to advancing plant-based therapies for the treatment of acute gastric ulcers. However, it is not suitable for assessing chronic effects or long-term healing, and with only five animals per group (n = 5), the statistical power may be limited. These were considered limitations and opened the chance for further studies. Another limitation of this study is that the levels of NO were not measured. Additionally, further assessment of its toxicity and bioavailability, followed by therapeutic applications in clinical settings, will be essential to ensure the efficacy and safety of EEIE, as well as evaluating potential interactions with conventional ulcer treatments will be crucial for establishing EEIE as a viable alternative therapy for gastric ulcers.
Acknowledgments
The authors thank (1) the Directorate of Higher Education of the Ministry of Education and Culture of the Republic of Indonesia and (2) the Rector of Universitas Padjadjaran, West Java, Indonesia. The present study was conducted in the framework of the dissertation project of the first author at the Doctoral Program in Pharmacy, Faculty of Pharmacy, Universitas Padjadjaran, West Java, Indonesia.
Funding
The first author’s tuition fee and living cost are funded by the Directorate of Higher Education of the Ministry of Education and Culture of the Republic of Indonesia via the PMDSU (Program Magister Doktor Sarjana Unggul) Scholarship years 2023 and 2024. The purchase of the plant materials and the solvent for extraction were funded by the Rector of Universitas Padjadjaran via the Directorate of Research and Community Engagement in the scheme of Academic-Leadership Research Grant under a contract document number 1479/UN6.3.1/PT.00/2024 of Prof. Dr. Jutti Levita. The purchase of the iNOS antibody (Item No. NB300-605, Novus-Biologicals), the Wistar rats, the ethical approval, the microscopic analysis, and the Western blot analysis were funded by the PMDSU Research Grant under a contract document number 3923/UN6.3.1/PT.00/2024 of Prof. Dr. Jutti Levita. The APC was supported by the Rector of Universitas Padjadjaran via the Directorate of Research and Community Engagement.
Disclosure
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
1. Zelickson MS, Bronder CM, Johnson BL, et al. Helicobacter pylori is not the predominant etiology for peptic ulcers requiring operation. Am Surg. 2011;77(8):1054–1060. doi:10.1177/000313481107700827
2. Lanas A, Chan FKL. Peptic ulcer disease. Lancet. 2017;390(10094):613–624. doi:10.1016/S0140-6736(16)32404-7
3. Kruk ME, Gage AD, Joseph NT, Danaei G, García-Saisó S, Salomon JA. Mortality due to low-quality health systems in the universal health coverage era: a systematic analysis of amenable deaths in 137 countries. Lancet. 2018;392(10160):2203–2212. doi:10.1016/S0140-6736(18)31668-4
4. Kavitt RT, Lipowska AM, Anyane-Yeboa A, Gralnek IM. Diagnosis and treatment of peptic ulcer disease. Am J Med. 2019;132(4):447–456. doi:10.1016/j.amjmed.2018.12.009
5. Abbasi-Kangevari M, Ahmadi N, Fattahi N, et al. Quality of care of peptic ulcer disease worldwide: a systematic analysis for the global burden of disease study 1990–2019. PLoS One. 2022;17(8):e0271284. doi:10.1371/journal.pone.0271284
6. Xie X, Ren K, Zhou Z, Dang C, Zhang H. The global, regional and national burden of peptic ulcer disease from 1990 to 2019: a population-based study. BMC Gastroenterol. 2022;22(1):58. doi:10.1186/s12876-022-02130-2
7. Hosseini E, Poursina F, van de Wiele T, Safaei HG, Adibi P. Helicobacter pylori in Iran: a systematic review on the association of genotypes and gastroduodenal diseases. J Res Med Sci. 2012;17(3):280.
8. Jones MP. The role of psychosocial factors in peptic ulcer disease: beyond Helicobacter pylori and NSAIDs. J Psychosom Res. 2006;60(4):407–412. doi:10.1016/j.jpsychores.2005.08.009
9. Yadav S, Pandey A, Mali SN. From lab to nature: recent advancements in the journey of gastroprotective agents from medicinal chemistry to phytotherapy. Eur J Med Chem. 2024;272:116436. doi:10.1016/j.ejmech.2024.116436
10. Taha MME, Salga MS, Ali HM, Abdulla MA, Abdelwahab SI, Hadi AHA. Gastroprotective activities of Turnera diffusa Willd. Ex Schult. revisited: role of arbutin. J Ethnopharmacol. 2012;141(1):273–281. doi:10.1016/j.jep.2012.02.030
11. Wasman SQ, Mahmood AA, Salehhuddin H, Zahra AA, Salmah I. Cytoprotective activities of Polygonum minus aqueous leaf extract on ethanol-induced gastric ulcer in rats. J Med Plants Res. 2010;4(24):2658–2665. doi:10.5897/jmpr09.412
12. Bourgonje AR, Feelisch M, Faber KN, Pasch A, Dijkstra G, van Goor H. Oxidative stress and redox-modulating therapeutics in inflammatory bowel disease. Trends Mol Med. 2020;26(11):1034–1046. doi:10.1016/j.molmed.2020.06.006
13. Prame Kumar K, Nicholls AJ, Wong CHY. Partners in crime: neutrophils and monocytes/macrophages in inflammation and disease. Cell Tissue Res. 2018;371(3):551–565. doi:10.1007/s00441-017-2753-2
14. Cederbaum AI, Lu Y, Wu D. Role of oxidative stress in alcohol-induced liver injury. Arch Toxicol. 2009;83(6):519–548. doi:10.1007/s00204-009-0432-0
15. Badr AM, Nf ELO, Ali RA. The implication of the crosstalk of Nrf2 with NOXs, and HMGB1 in ethanol-induced gastric ulcer: potential protective effect is afforded by Raspberry Ketone. PLoS One. 2019;14(8):e0220548. doi:10.1371/journal.pone.0220548
16. Park J, Min JS, Kim B, et al. Mitochondrial ROS govern the LPS-induced pro-inflammatory response in microglia cells by regulating MAPK and NF-κB pathways. Neurosci Lett. 2015;584:191–196. doi:10.1016/j.neulet.2014.10.016
17. Youn GS, Lee KW, Choi SY, Park J. Overexpression of HDAC6 induces pro-inflammatory responses by regulating ROS-MAPK-NF-κB/AP-1 signaling pathways in macrophages. Free Radic Biol Med. 2016;97:14–23. doi:10.1016/j.freeradbiomed.2016.05.014
18. Li W, Huang H, Niu X, Fan T, Mu Q, Li H. Protective effect of tetrahydrocoptisine against ethanol-induced gastric ulcer in mice. Toxicol Appl Pharmacol. 2013;272(1):21–29. doi:10.1016/j.taap.2013.05.035
19. Raish M, Ahmad A, Ansari MA, et al. Momordica charantia polysaccharides ameliorate oxidative stress, inflammation, and apoptosis in ethanol-induced gastritis in mucosa through NF-kB signaling pathway inhibition. Int J Biol Macromol. 2018;111:193–199. doi:10.1016/j.ijbiomac.2018.01.008
20. Frank S, Pfeilschifter J, Madlener M, Werner S. Induction of inducible nitric oxide synthase and its corresponding tetrahydrobiopterin-cofactor-synthesizing enzyme GTP-cyclohydrolase I during cutaneous wound repair. J Invest Dermatol. 1998;111(6):1058–1064. doi:10.1046/j.1523-1747.1998.00434.x
21. Lundberg JON, Lundberg JM, Alving K, Weitzberg E. Nitric oxide and inflammation: the answer is blowing in the wind. Nat Med. 1997;3(1):30–31. doi:10.1038/nm0197-30
22. Lambert AA, Lam JO, Paik JJ, Ugarte-Gil C, Drummond MB, Crowell TA. Risk of community-acquired pneumonia with outpatient proton-pump inhibitor therapy: a systematic review and meta-analysis. PLoS One. 2015;10(6):e0128004. doi:10.1371/journal.pone.0128004
23. Molnar T. Pathogenesis of ulcerative colitis and Crohn’s disease: similarities, differences and a lot of things we do not know yet. J Clin Cell Immunol. 2014;05(04). doi:10.4172/2155-9899.1000253
24. Probert CSJ, Dignass AU, Lindgren S, Oudkerk Pool M, Marteau P. Combined oral and rectal mesalazine for the treatment of mild-to-moderately active ulcerative colitis: rapid symptom resolution and improvements in quality of life. J Crohn’s Colitis. 2014;8(3):200–207. doi:10.1016/j.crohns.2013.08.007
25. Scally B, Emberson JR, Spata E, et al. Effects of gastroprotectant drugs for the prevention and treatment of peptic ulcer disease and its complications: a meta-analysis of randomised trials. Lancet Gastroenterol Hepatol. 2018;3(4):231–241. doi:10.1016/S2468-1253(18)30037-2
26. Haastrup P, Paulsen MS, Begtrup LM, Hansen JM, Jarbol DE. Strategies for discontinuation of proton pump inhibitors: a systematic review. Fam Pract. 2014;31(6):625–630. doi:10.1093/fampra/cmu050
27. Shaw DH. Drugs acting on the gastrointestinal tract. In: Pharmacology and Therapeutics for Dentistry. Elsevier. 2017:404–416. doi:10.1016/B978-0-323-39307-2.00028-X
28. Hasanuzzaman M, Bang CS, Gong EJ. Antibiotic resistance of Helicobacter pylori: mechanisms and clinical implications. J Korean Med Sci. 2024;39(4):e44. doi:10.3346/jkms.2024.39.e44
29. Gupta M, Kapoor B, Gupta R, Singh N. Plants and phytochemicals for treatment of peptic ulcer: an overview. South Afr J Bot. 2021;138:105–114. doi:10.1016/j.sajb.2020.11.030
30. Kuna L, Jakab J, Smolic R, Raguz-Lucic N, Vcev A, Smolic M. Peptic ulcer disease: a brief review of conventional therapy and herbal treatment options. J Clin Med. 2019;8(2):179. doi:10.3390/jcm8020179
31. Saini P, Kumar N, Kumar S, et al. Bioactive compounds, nutritional benefits and food applications of colored wheat: a comprehensive review. Crit Rev Food Sci Nutr. 2021;61(19):3197–3210. doi:10.1080/10408398.2020.1793727
32. Kapoor B, Kaur G, Gupta M, Gupta R. Sub-chronic safety evaluation of aqueous extract of Alangium salvifolium (L.f.) Wangerin leaves in rats. J Adv Pharm Technol Res. 2017;8(3):108–113. doi:10.4103/japtr.JAPTR_69_17
33. Juwita T, Puspitasari IM, Levita J. Torch ginger (Etlingera elatior): a review on its botanical aspects, phytoconstituents and pharmacological activities. Pakistan J Biol Sci. 2018;21(4):151–165. doi:10.3923/PJBS.2018.151.165
34. Prayoga D, Aulifa D, Budiman A, Levita J. Plants with anti-ulcer activity and mechanism: a review of preclinical and clinical studies. Drug Des Devel Ther. 2024;18:193–213. doi:10.2147/DDDT.S446949
35. Ghasemzadeh A, Jaafar HZE, Rahmat A, Ashkani S. Secondary metabolites constituents and antioxidant, anticancer and antibacterial activities of Etlingera elatior (Jack) R.M.Sm grown in different locations of Malaysia. BMC Complement Altern Med. 2015;15(1). doi:10.1186/s12906-015-0838-6
36. Juwita T, Pakpahan WHP, Puspitasari IM, Saptarini NM, Levita J. Anti-inflammatory activity of etlingera elatior (Jack) R.M. Smith flower on gastric ulceration-induced Wistar rats. Pakistan J Biol Sci. 2020;23(9):1193–1200. doi:10.3923/pjbs.2020.1193.1200
37. Noordin L, Wan Ahmad WAN, Muhamad Nor NA, Abu Bakar NH, Ugusman A. Etlingera elatior flower aqueous extract protects against oxidative stress-induced nephropathy in a rat model of type 2 diabetes. Evid Based Complement Altern Med. 2022;2022:1–12. doi:10.1155/2022/2814196
38. Krajarng A, Chulasiri M, Watanapokasin R. Etlingera elatior Extract promotes cell death in B16 melanoma cells via down-regulation of ERK and Akt signaling pathways. BMC Complement Altern Med. 2017;17(1):415. doi:10.1186/s12906-017-1921-y
39. Ahn HI, Jang HJ, Kwon OK, et al. Quercetin attenuates the production of pro-inflammatory cytokines in H292 human lung epithelial cells infected with Pseudomonas aeruginosa by modulating ExoS production. J Microbiol Biotechnol. 2023;33(4):430–440. doi:10.4014/jmb.2208.08034
40. Ruiz PA, Braune A, Hölzlwimmer G, Quintanilla-Fend L, Haller D. Quercetin inhibits TNF-induced NF-κB transcription factor recruitment to proinflammatory gene promoters in murine intestinal epithelial cells. J Nutr. 2007;137(5):1208–1215. doi:10.1093/jn/137.5.1208
41. Bastin A, Teimouri M, Faramarz S, Shabani M, Doustimotlagh AH, Sadeghi A. In vitro and molecular docking analysis of quercetin as an anti-inflammatory and antioxidant. Curr Pharm Des. 2023;29(11):883–891. doi:10.2174/1381612829666230330084043
42. Hsieh HL, Yu MC, Cheng LC, et al. Quercetin exerts anti-inflammatory effects via inhibiting tumor necrosis factor-α-induced matrix metalloproteinase-9 expression in normal human gastric epithelial cells. World J Gastroenterol. 2022;28(11):1139–1158. doi:10.3748/wjg.v28.i11.1139
43. Prayoga DK, Pitaloka DAE, Aulifa DL, et al. Phytochemical analysis of Etlingera elatior inflorescence extract (nutritional composition, total anthocyanin, the levels of vitamin C, cyanidin-3-glucoside, and quercetin) and computational study towards inducible nitric oxide synthase. J Exp Pharmacol. 2025;17:123–141. doi:10.2147/JEP.S505658
44. Nurhayatun E, Purwanto B, Soetrisno S, et al. The beneficial effect of the ethanolic extract Etlingera elatior on IL-1ß and caspase-3 levels in sepsis mice model. J Appl Pharm Sci. 2023;13(06):116–120. doi:10.7324/JAPS.2023.90410
45. Jackie T, Haleagrahara N, Chakravarthi S. Antioxidant effects of Etlingera elatior flower extract against lead acetate-induced perturbations in free radical scavenging enzymes and lipid peroxidation in rats. BMC Res Notes. 2011;4(1). doi:10.1186/1756-0500-4-67
46. Becker JB, Prendergast BJ, Liang JW. Female rats are not more variable than male rats: a meta-analysis of neuroscience studies. Biol Sex Differ. 2016;7(1):34. doi:10.1186/s13293-016-0087-5
47. Alkushi AGR, Elsawy NAM. Quercetin attenuates, indomethacin-induced acute gastric ulcer in rats. Folia Morphol. 2017;76(2):252–261. doi:10.5603/FM.a2016.0067
48. Silva LP, De Angelis CD, Bonamin F, et al. Terminalia catappa L.: a medicinal plant from the Caribbean pharmacopeia with anti-Helicobacter pylori and antiulcer action in experimental rodent models. J Ethnopharmacol. 2015;159:285–295. doi:10.1016/j.jep.2014.11.025
49. Bento EB, Júnior FEB, de Oliveira DR, et al. Antiulcerogenic activity of the hydroalcoholic extract of leaves of Annona muricata Linnaeus in mice. Saudi J Biol Sci. 2016;25(4):609–621. doi:10.1016/j.sjbs.2016.01.024
50. Leary S, Ridge H, et al. AVMA Guidelines for the Euthanasia of Animals: 2020. 2020.
51. Tobar Leitão SA, Soares DS, Carvas Junior N, Zimmer R, Ludwig NF, Andrades M. Study of anesthetics for euthanasia in rats and mice: a systematic review and meta-analysis on the impact upon biological outcomes (SAFE-RM). Life Sci. 2021;284:119916. doi:10.1016/j.lfs.2021.119916
52. Rosdianto AM, Puspitasari IM, Lesmana R, Levita J. Inhibitory activity of Boesenbergia rotunda (L.) Mansf. rhizome towards the expression of Akt and NF‐kappaB p65 in acetic acid‐induced Wistar rats. Evid Based Complement Altern Med. 2020;2020(1). doi:10.1155/2020/6940313
53. Manjula K, Raj M, Krishna R. Feed efficiency and serobiochemical profile of Wistar rats fed with spirulina as functional food. Curr Res Nutr Food Sci J. 2016;4(2):135–140. doi:10.12944/CRNFSJ.4.2.07
54. Adinortey MB, Ansah C, Galyuon I, Nyarko A. In vivo models used for evaluation of potential antigastroduodenal ulcer agents. Ulcers. 2013;2013:1–12. doi:10.1155/2013/796405
55. Prasanth Reddy V, Sudheshna G, Afsar SK, et al. Evaluation of anti-ulcer activity of Citrullus colocynthis fruit against pylorus ligation induced ulcers in male Wistar rats. Int J Pharm Pharm Sci. 2012;4(2):446–451.
56. Sistani Karampour N, Arzi A, Rezaie A, Pashmforoosh M, Kordi F. Gastroprotective effect of zingerone on ethanol-induced gastric ulcers in rats. Medicina. 2019;55(3). doi:10.3390/medicina55030064
57. Magro F, Langner C, Driessen A, et al. European consensus on the histopathology of inflammatory bowel disease. J Crohn’s Colitis. 2013;7(10):827–851. doi:10.1016/j.crohns.2013.06.001
58. Gietz L, Mayer B. A resampling approach for sample size estimation in animal experiments. J Int Transl Med. 2017;5(2):53–62. doi:10.11910/2227-6394.2017.05.02.01
59. Zhang X, Hartmann P. How to calculate sample size in animal and human studies. Front Med Lausanne. 2023;10:1215927. doi:10.3389/fmed.2023.1215927
60. Serdar CC, Cihan M, Yücel D, Serdar MA. Sample size, power and effect size revisited: simplified and practical approaches in pre-clinical, clinical and laboratory studies. Biochem Medica. 2021;31(1):27–53. doi:10.11613/BM.2021.010502
61. Guzmán-Gómez O, García-Rodríguez RV, Quevedo-Corona L, et al. Amelioration of ethanol-induced gastric ulcers in rats pretreated with phycobiliproteins of Arthrospira (Spirulina) maxima. Nutrients. 2018;10(6):763. doi:10.3390/nu10060763
62. Arab HH, Salama SA, Omar HA, Arafa ESA, Maghrabi IA. Diosmin protects against ethanol-induced gastric injury in rats: novel anti-ulcer actions. PLoS One. 2015;10(3):e0122417. doi:10.1371/journal.pone.0122417
63. Bhattacharyya A, Chattopadhyay R, Mitra S, Crowe SE. Oxidative stress: an essential factor in the pathogenesis of gastrointestinal mucosal diseases. Physiol Rev. 2014;94(2):329–354. doi:10.1152/physrev.00040.2012
64. El-Maraghy SA, Rizk SM, Shahin NN. Gastroprotective effect of crocin in ethanol-induced gastric injury in rats. Chem Biol Interact. 2015;229:26–35. doi:10.1016/j.cbi.2015.01.015
65. Mamache W, Benabdallah H, Hannachi A, et al. Preventing ethanol-induced stomach ulcers in rats using Senecio perralderianus leaf extract. Rev Científica la Fac Ciencias Vet. 2024;34(1):1–8. doi:10.52973/rcfcv-e34348
66. Barrachina MD, Panes J, Esplugues JV. Role of nitric oxide in gastrointestinal inflammatory and ulcerative diseases: perspective for drugs development. Curr Pharm Des. 2001;7(1):31–48. doi:10.2174/1381612013398491
67. Nagai N, Fukuhata T, Ito Y, Usui S, Hirano K. Involvement of interleukin 18 in indomethacin-induced lesions of the gastric mucosa in adjuvant-induced arthritis rat. Toxicol. 2009;255(3):124–130. doi:10.1016/j.tox.2008.10.005
68. Li Y, Wang WP, Wang HY, Cho CH. Intragastric administration of heparin enhances gastric ulcer healing through a nitric oxide-dependent mechanism in rats. Eur J Pharmacol. 2000;399(2–3):205–214. doi:10.1016/S0014-2999(00)00278-8
69. Yu RQ, Wu XY, Zhou X, Zhu J, Ma LY. Cyanidin-3-glucoside attenuates body weight gain, serum lipid concentrations and insulin resistance in high-fat diet-induced obese rats]. Zhongguo Dang Dai Er Ke Za. Zhi. 2014;16(5):534–538. doi:10.7499/j.issn.1008-8830.2014.05.018
70. Ruiz MJ, Fernández M, Picó Y, et al. Dietary administration of high doses of pterostilbene and quercetin to mice is not toxic. J Agric Food Chem. 2009;57(8):3180–3186. doi:10.1021/jf803579e
71. tan SY, Li J, Chen Y, Yang C-M, Tang H-L, Wang J-C. Yang C min, Tang H lan, Wang J chang. [Effects of glutathione on plasma heat shock protein 70 of acute gastric mucosal injury in rats exposed to positive acceleration]. Zhonghua Yi Xue Za Zhi. 2013;93(46):3708–3710.
72. Sarkar S, Sengupta A, Mukhrjee A, et al. Antiulcer potential of morin in acetic acid-induced gastric ulcer via modulation of endogenous biomarkers in laboratory animals. Pharmacologia. 2015;6(7):273–281. doi:10.5567/pharmacologia.2015.273.281
73. Kolgazi M, Ozdemir-Kumral ZN, Cantali-Ozturk C, et al. Anti-inflammatory effects of nesfatin-1 on acetic acid-induced gastric ulcer in rats: involvement of cyclo-oxygenase pathway. J Physiol Pharmacol. 2017;68(5):765–777.
74. Pereira SR, Pereira R, Figueiredo I, Freitas V, Dinis TCP, Almeida LM. Comparison of anti-inflammatory activities of an anthocyanin-rich fraction from Portuguese blueberries (Vaccinium corymbosum L.) and 5-aminosalicylic acid in a TNBS-induced colitis rat model. PLoS One. 2017;12(3):e0174116. doi:10.1371/journal.pone.0174116
75. Jiménez S, Gascón S, Luquin A, Laguna M, Ancin-Azpilicueta C, Rodríguez-Yoldi MJ. Rosa canina extracts have antiproliferative and antioxidant effects on Caco-2 human colon cancer. PLoS One. 2016;11(7):e0159136. doi:10.1371/journal.pone.0159136
76. Barreca D, Gattuso G, Laganà G, Leuzzi U, Bellocco E. C- and o-glycosyl flavonoids in sanguinello and tarocco blood Orange (Citrus sinensis (L.) Osbeck) juice: identification and influence on antioxidant properties and acetylcholinesterase activity. Food Chem. 2016;196:619–627. doi:10.1016/J.FOODCHEM.2015.09.098
77. Pesca MS, Dal Piaz F, Sanogo R, et al. Bioassay-guided isolation of proanthocyanidins with antiangiogenic activities. J Nat Prod. 2013;76(1):29–35. doi:10.1021/NP300614U
78. Tang J, Diao P, Shu X, Li L. Quercetin and quercitrin attenuate the inflammatory response and oxidative stress in LPS-Induced RAW264.7 Cells: in vitro assessment and a theoretical model. Biomed Res Int. 2019;2019:1–8. doi:10.1155/2019/7039802
79. Mayadas TN, Cullere X, Lowell CA. The multifaceted functions of neutrophils. Annu Rev Pathol Mech Dis. 2014;9(1):181–218. doi:10.1146/annurev-pathol-020712-164023
80. Rosales C. Neutrophil: a cell with many roles in inflammation or several cell types? Front Physiol. 2018;9:113. doi:10.3389/fphys.2018.00113
81. Lawrence T, Willoughby DA, Gilroy DW. Anti-inflammatory lipid mediators and insights into the resolution of inflammation. Nat Rev Immunol. 2002;2(10):787–795. doi:10.1038/nri915
82. Li WS, Lin SC, Chu CH, et al. The gastroprotective effect of naringenin against ethanol-induced gastric ulcers in mice through inhibiting oxidative and inflammatory responses. Int J Mol Sci. 2021;22(21):11985. doi:10.3390/ijms222111985
83. Jones RJ, Smith SME, Gao YT, et al. The function of the small insertion in the hinge subdomain in the control of constitutive mammalian nitric-oxide synthases. J Biol Chem. 2004;279(35):36876–36883. doi:10.1074/jbc.M402808200
84. Osawa Y, Lowe ER, Everett AC, Dunbar AY, Billecke SS. Proteolytic degradation of nitric oxide synthase: effect of inhibitors and role of hsp90-based chaperones. J Pharmacol Exp Ther. 2003;304(2):493–497. doi:10.1124/jpet.102.035055
85. Sadek J, Hall DT, Colalillo B, et al. Pharmacological or genetic inhibition of iNOS prevents cachexia‐mediated muscle wasting and its associated metabolism defects. EMBO Mol Med. 2021;13(7). doi:10.15252/emmm.202013591
86. Zeng LH, Fatima M, Syed SK, et al. Anti-inflammatory and anti-oxidant properties of Ipomoea nil (Linn.) Roth significantly alleviates cigarette smoke (CS)-induced acute lung injury via possibly inhibiting the NF-κB pathway. Biomed Pharmacother. 2022;155:113267. doi:10.1016/j.biopha.2022.113267
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Medicinal Plants That Modulate Nitric Oxide Synthase Activity: Implications in Inflammation and Oxidative Stress
Prayoga DK, Aulifa DL, Budiman A, Jiranusornkul S, Levita J
Drug Design, Development and Therapy 2026, 20:582323
Published Date: 13 March 2026