Back to Journals » Journal of Inflammation Research » Volume 18
Mechanistic Insights into Qin Bi Yin for Psoriasis Treatment Using Network Pharmacology, Experimental Validation, and Molecular Docking
Authors Zhang S, Du Y, Zhang H, Duan X, Li L
Received 31 May 2025
Accepted for publication 23 September 2025
Published 1 October 2025 Volume 2025:18 Pages 13535—13554
DOI https://doi.org/10.2147/JIR.S544019
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Anish R. Maskey
Shouxu Zhang,1,2 Yue Du,1,2 Haomin Zhang,1,2 Xingwu Duan,1 Lingling Li1
1Department of Dermatology, Dongzhimen Hospital, Beijing University of Chinese Medicine, Beijing, 100700, People’s Republic of China; 2Graduate School, Beijing University of Chinese Medicine, Beijing, 100029, People’s Republic of China
Correspondence: Lingling Li, Department of Dermatology, Dongzhimen Hospital, Beijing University of Chinese Medicine, Beijing, 100700, People’s Republic of China, Email [email protected]
Objective: In this study, we investigated the potential mechanism of action of Qing Bi Yin (QBY) in psoriasis treatment via regulation of the sphingosine-1-phosphate (S1P)/sphingosine-1-phosphate receptor 1 (S1PR1) pathway and Th17 cell differentiation.
Methods: Network pharmacology was used to identify potential targets and elucidate the potential mechanisms of QBY in psoriasis. The predicted mechanisms were validated with in vitro Th17 cell differentiation assays using isolated mouse splenic CD4+ T cells. Molecular docking was performed to evaluate the binding affinities between active compounds and key target proteins.
Results: We identified 262 overlapping QBY psoriasis target genes. Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis showed significant associations with the sphingolipid signaling pathway, Th17 cell differentiation, and IL-17 signaling pathway. Network algorithms were used to identify 10 key active compounds and five hub genes. QBY treatment suppresses S1P synthesis and S1pr1 expression in Th17-polarized cells. QBY inhibited Th17 cell proliferation and differentiation, reduced inflammatory cytokine secretion by Th17 cells via the S1P/S1PR1 pathway, and modulated STAT3 and SMAD2 phosphorylation. Molecular docking showed strong binding affinities between active compounds (glabridin, luteolin, licoflavone A, and isobutyrylshikonin) from QBY and key targets (STAT3, SMAD2, SPHK1, and RORγt) in Th17 cells.
Conclusion: QBY ameliorates psoriatic inflammation by regulating Th17 cell differentiation via the S1P/S1PR1 signaling pathway. These findings underscore the clinical translational potential of QBY and its active constituent glabridin.
Keywords: psoriasis, network pharmacology, S1P, S1PR1, Th17 differentiation
Introduction
Psoriasis is a chronic autoimmune-mediated inflammatory skin disease affecting approximately 2–3% of the global population and substantially affects patients’ quality of life.1,2 Interleukin-23 (IL-23)/Th17 axis is a central mediator in psoriasis pathogenesis.2 IL-23 triggers Th17 cell differentiation and IL-17 release. These inflammatory mediators cause keratinocyte hyperproliferation, neutrophil recruitment, and enhanced angiogenesis, resulting in persistent inflammation and the characteristic psoriatic lesions. Although biological therapies targeting IL-17, IL-23, and related pathways have shown clinical efficacy, substantial challenges remain, including long-term safety concerns, high treatment costs, and emerging drug resistance in certain patient populations.3 These limitations highlight the importance of developing novel therapeutic targets and strategies for psoriasis treatment.
Recent advances in sphingolipid pathway research have highlighted their crucial role in the pathogenesis of psoriasis. This signaling system mediates diverse biological processes, including immune cell trafficking, angiogenesis regulation,4 skin barrier maintenance, and keratinocyte proliferation control,5 all of which are critically involved in psoriasis pathogenesis. As a pivotal lipid mediator, sphingosine-1-phosphate (S1P) modulates diverse physiological and pathological processes through sphingosine-1-phosphate receptors 1–5 (S1PR1–S1PR5, collectively referred to as S1PRs).6 The S1P/S1PR1 pathway plays an important role in Th17 cell differentiation and function. Aberrant activation of this signaling axis enhances Th17 cell polarization and amplifies inflammatory factor secretion, creating a cascade effect that intensifies inflammatory responses and accelerates lesion development in psoriasis.7 Therefore, the S1P/S1PR1 signaling pathway is a crucial mediator of psoriasis pathogenesis and offers novel therapeutic targets.8
Traditional Chinese medicine provides a unique therapeutic approach for psoriasis management through its multi-component, multi-target strategy.9 Qing Bi Yin (QBY) is derived from Xiaoyin Jiedu granules (XYJD), an empirical psoriasis formula developed by Prof. Jin Qifeng. Our previous clinical studies demonstrated that XYJD ameliorates psoriatic symptoms and modulates Th17-related immune responses. We observed that XYJD could reduce peripheral blood levels of retinoic acid-related orphan receptor gamma t (ROR-γt), IL-17, interleukin-22 (IL-22), and interleukin-23 (IL-23),10 suggesting its role in regulating Th17 cell-related immune responses. The addition of Lonicerae japonicae flos and Forsythiae fructus in the QBY formula makes it particularly effective in clinical practice for early-stage psoriasis, characterized mainly by erythematous papules.11 Although biological agents are widely used to treat plaque psoriasis, clinical experience has shown that QBY offers a safe and effective alternative for managing guttate psoriasis. However, the characterization of QBY’s bioactive compounds, molecular targets, and underlying mechanisms of action of QBY requires further investigation.
In this study, we used ultra-high-performance liquid chromatography coupled with Q Exactive Orbitrap mass spectrometry (UHPLC-QE-MS) to characterize QBY’s chemical constituents of QBY. We also used network pharmacology approaches to investigate potential therapeutic targets and molecular mechanisms of psoriasis treatment. The predicted mechanisms were validated experimentally using mouse CD4+ T cells. We first validated the QBY-mediated inhibition of S1P synthesis and S1PR1 mRNA expression using enzyme-linked immunosorbent assay (ELISA) and Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR). We confirmed QBY’s suppressive effects of QBY on Th17 cell proliferation and differentiation via the S1P/S1PR1 pathway. Molecular docking was used to evaluate the interactions between the key active components and hub targets. The findings provide molecular evidence for understanding QBY’s therapeutic effects of QBY in psoriasis treatment. A flowchart of the study design is shown in Figure 1.
 |
Figure 1 The schematic diagram of the current research. |
Materials and Methods
Reagents and Instruments
The following instruments were used: ultra-high performance liquid chromatography (Thermo Fisher Scientific, Vanquish), high resolution mass spectrometry (Thermo Fisher Scientific, Q Exactive HF-X), microplate reader (Thermo Scientific, Multiskan MK3), shaker (Qilinbeier, TS-1000), refrigerated centrifuge (Sigma, 3K15), ABI 7500 Real Time PCR System (Applied Biosystems, 7500), flow cytometer (BD-FACSVerse, C6), Chemiluminescence imager (ChemiScope 5300 Pro).
The following reagents were used: S1P-receptor-agonist-1 (TargetMOI, T4031), APC anti-mouse CD4 antibody (Biolegend, B318293), IL-17A Monoclonal Antibody, PE (eBioscience, 12-7177-81), Recombinant human IL-6 (Solarbio, P00022), Recombinant Human TGF-beta 1/TGFB1 (Solarbio, P00121), Recombinant human IL1B (Solarbio, P02454), Recombinant Mouse IL-23 (Solarbio, P00230), Anti-IFNG Antibody (Solarbio, K009380P), Anti-IL4 Antibody (Solarbio, K114858P), Trizol (Thermo Fisher, 15596018), RevertAid First Strand cDNA Synthesis Kit (Thermo, K1622), DyNAmo Flash SYBR Green qPCR Kit (Thermo Fisher, F-415XL), Cell Counting Kit-8 (MedChemExpress, HY-K0301), Mouse ELISA kits (Buya-TEK, BYK4004, BYK1596, BYK0827), BCA Protein Assay Kit (BL521A). Primary antibodies were purchased from Affinity Biosciences, including STAT3 (AF6294), P-STAT3 (AF3293), SMAD2 (AF6449), P-SMAD2 (AF3449), GAPDH (AF7021).
Animal
Female BALB/c mice (8 weeks old, 18±2 g) were obtained from Beijing Vital River Laboratory Animal Technology Co., Ltd. (Certificate No. SCXK [Beijing] 2016-0006). The animals were housed in specific-pathogen-free (SPF) facilities at the Key Laboratory of Dongzhimen Hospital, Beijing University of Chinese Medicine, with ad libitum access to water and standard laboratory chow. Following a one-week acclimatization period, experimental procedures were initiated. All animal care and experimental protocols were conducted in accordance with the Guide for the Care and Use of Laboratory Animals and approved by the Institutional Animal Care and Use Committee of Dongzhimen Hospital, Beijing University of Chinese Medicine (Approval No. 2021DZMEC-056-02).
Preparation of QBY
QBY was prepared using Lonicerae Japonicae Flos 15g, Forsythiae Fructus 15g, Arctii Fructus 15g, Rehmanniae Radix 30g, Moutan Cortex 12g, Paeoniae Radix Rubra 12g, Scutellariae Radix 10g, Imperatae Rhizoma 30g, Houttuyniae Herba 20g, Arnebiae Radix 12g, Isatidis Radix 10g, Glycyrrhizae Radix et Rhizoma 10g. All the herbal ingredients were from the Traditional Chinese Medicine Pharmacy of Dongzhimen Hospital, Beijing, China. All the herbs were soaked with 10 times distilled water for 1 hour, decocted twice. The combined extracts were concentrated via rotary evaporation, pre-frozen at −80°C for 12 hours, and lyophilized under vacuum conditions. The lyophilized powder was stored at −20°C for later use.
UHPLC-QE-MS Analysis of QBY
Chromatographic separation was performed using a Vanquish UHPLC system coupled with an ACQUITY UPLC® HSS T3 column (2.1 mm × 100 mm, 1.8 μm particle size). The binary mobile phase system consisted of (A) 0.1% formic acid in water and (B) acetonitrile, delivered at a flow rate of 0.3 mL·min−1. The column temperature was maintained at 40°C. The gradient elution program was optimized over a 17-minute run: initial conditions of 10% B were maintained for 2 minutes, followed by a linear increase to 95% B from 2 to 11 minutes, held at 95% B for 3 minutes (11–14 min), then returned to initial conditions at 14.1 minutes and re-equilibrated until 17 minutes. The sample injection volume was 5 μL.
Mass spectrometric detection was performed on a Q Exactive HF-X high-resolution mass spectrometer equipped with a heated electrospray ionization (HESI) source, operating in both positive and negative ion modes. The optimized HESI source parameters were: spray voltage +3.8/-3.2 kV, probe heater temperature 350°C, capillary temperature 320°C, sheath gas flow rate 30 arbitrary units, and auxiliary gas flow rate 10 arbitrary units. Data acquisition was conducted in Full MS/dd-MS2 mode, with Full MS scans ranging from m/z 100–1500 at a resolution of 120,000. For dd-MS2, the resolution was 30,000, with an AGC target of 1E6, maximum IT of 50 ms, and collision energy of 30 eV.
Network Pharmacological Analysis
Active Components and Targets of QBY
Sixty-nine compounds were identified in QBY by UHPLC-QE-MS analysis. The relevant molecular structures were searched in PubChem database (https://pubchem.ncbi.nlm.nih.gov/), corresponding drug targets were identified through Swiss Target Prediction (http://swisstargetprediction.ch/), PubChem, HERB (http://herb.ac.cn/) databases. We also supplemented some experimentally validated compound targets derived from peer-reviewed literature accessed through Google Scholar (https://scholar.google.com/).
Psoriasis Disease Targets
Psoriasis-associated targets were collected from four comprehensive databases: Online Mendelian Inheritance in Man (OMIM, https://www.omim.org/), GeneCards (https://www.genecards.org/), Therapeutic Target Database (TTD, http://db.idrblab.net/ttd), and DisGeNET (http://www.disgenet.org/). The search was conducted using “psoriasis” as the keyword. For GeneCards database, targets with relevance scores ≥1 were selected. The targets from all databases were integrated and duplicates were removed. The overlapped targets between QBY compounds and psoriasis-related genes were identified using the Venny 2.1.0 online tool (https://bioinfogp.cnb.csic.es/tools/venny/).
Functional Enrichment Analysis
Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis was performed on the obtained overlapped targets using the clusterProfiler package in R. Results were visualized by https://www.bioinformatics.com.cn, an online platform for data analysis and visualization.
Network Construction and Analysis
To investigate QBY’s effects on immune regulation and signal transduction, we selected five significant pathways from the top 25 enriched KEGG pathways (https://www.genome.jp/kegg/pathway.html), which are categorized under immune system and signal transduction in the KEGG secondary classification system. The targets from these selected pathways were submitted to the STRING database (https://cn.string-db.org/) to construct a protein-protein interaction (PPI) network. The top 5 hub genes were identified using the MCC algorithm implemented in the cytoHubba plug-in of Cytoscape3.9.1.
A compound-target-pathway network was constructed by integrating the active compounds from QBY, their corresponding targets, and the selected pathways. The topological features were analyzed using the Analyze Network tool in Cytoscape3.9.1, and the top 10 compounds with the highest degree values were identified as key active components of QBY in the treatment of psoriasis.
Verification Experiments in vitro
Isolation of CD4+T Cells and Th17 Polarization
CD4+ T cells were isolated from mouse spleen single-cell suspensions using the MojoSort™ Mouse CD4 Naïve T Cell Isolation Kit, achieving >94.1% purity (Figure S1). The isolated cells were seeded at 2 × 106 cells/well in 12-well plates pre-coated with anti-CD3 (5 μg/mL) and anti-CD28 (2 μg/mL) antibodies. Th17 cell differentiation was induced using a cytokine cocktail containing TGF-β (5 ng/mL), IL-6 (20 ng/mL), IL-1β (10 ng/mL), IL-23 (15 ng/mL), anti-IL-4 (10 μg/mL), and anti-IFN-γ (10 μg/mL) for 3 days.12 Given our previous results, cells were treated with S1P (0.5 μM) for 24 hours and QBY (100 ng/mL) for 48 hours.11 To examine S1P/S1PR1 pathway involvement, we used the S1PR1 antagonist W146.
The experimental design comprised nine groups: Group A: Th17 induction medium only, Group B: Th17 induction medium + vehicle, Group C: Th17 induction medium + QBY freeze-dried powder solution (100 ng/mL), Group D: S1P (0.5 μM) + Th17 induction medium, Group E: S1P + Th17 induction medium + vehicle, Group F: S1P + Th17 induction medium + QBY freeze-dried powder solution, Group G: W146 + S1P + Th17 induction medium, Group H: W146 + S1P + Th17 induction medium + vehicle, Group I: W146 + S1P + Th17 induction medium + QBY freeze-dried powder solution.
RT-qPCR
Total RNA was extracted from Th17 cells using Trizol reagent. 1μg of total RNA was reverse transcribed to cDNA using a first-strand cDNA synthesis kit. PCR amplification was performed using SYBR Green qPCR Master Mix. Glyceraldehyde-3-phosphate dehydrogenase (Gapdh) was used as an internal reference gene. Differential expression of target genes was analyzed using the 2−ΔΔCT method. Primer sequences are listed in Table 1.
 |
Table 1 Primer Sequences |
Cell Counting Kit-8 (CCK-8) Assay
The viability of Th17 cells was assessed using Cell Counting Kit-8. After each group of cells was cultured to the corresponding time point, 20 μL of CCK-8 was added to each well and incubated at 37°C in a 5% CO2 incubator protected from light for 4 h. The OD value of the same point was measured at 450 nm by an enzyme marker, and cell viability was analyzed by the measured OD value.
Flow Cytometry
Cells were washed twice with PBS and incubated with antibodies at 4°C for 30 minutes. Cell surface staining was performed using CD4-APC antibody. Following fixation and permeabilization with Fixation/Permeabilization solution, cells were subjected to intracellular staining with IL-17-PE antibody. The stained cells were analyzed using flow cytometry, and data analysis was performed using FlowJo software.
ELISA
Cell supernatants were taken and the levels of S1P, IL-22, IL-23 and IL-17 in the culture medium were determined using an ELISA kit according to the manufacturer’s instructions.
Western Blotting Assay
RIPA lysate containing PMSF was added to the cell precipitate and thoroughly lysed. After centrifugation, the supernatant was collected. Protein concentration in the supernatant was determined using a BCA protein assay kit. Equal amounts of protein from each experimental group were subjected to electrophoresis at 80 V for 30 min followed by 120 V for 60 min. The proteins were then transferred to a PVDF membrane, which was rinsed in TBST and blocked in 5% skimmed milk for 2 h. Primary antibodies were added at the following dilutions: GAPDH (1:3000), STAT3 (1:1000), P-STAT3 (1:1000), SMAD2 (1:1000), and P-SMAD2 (1:1000). The membrane was incubated overnight at 4°C, washed, and then incubated with secondary antibody (1:5000) for 1 hour, followed by additional washing steps. ECL luminescent solution was applied to the membrane, and images were captured using a chemiluminescence imaging system. Band intensities were quantified using Image J software. The relative protein expression levels were calculated by normalizing the target protein band intensities to those of GAPDH.
Molecular Docking Analysis
Molecular docking was performed using the CB-Dock2 platform (https://cadd.labshare.cn/cb-dock2/php/index.php), which is an AutoDock Vina-based docking tool that automatically identifies and analyzes ligand-receptor binding sites with enhanced accuracy and simplified workflow. The 3D structures of the protein targets were obtained from the Protein Data Bank (PDB) database (https://www.rcsb.org/), while the 3D structures of molecular ligands were retrieved from the PubChem database (https://pubchem.ncbi.nlm.nih.gov/). The 3D structures of both proteins and ligands were uploaded to the CB-Dock2 website (https://cadd.labshare.cn/cb-dock2/php/example.php) for automated blind docking.13,14 Binding affinities (kcal/mol) were calculated using CB-Dock2, and the conformations with the lowest binding energies were selected for visualization. The detailed ligand-receptor interactions were analyzed and visualized using BIOVIA Discovery Studio Visualizer (version 2021).
Statistical Analysis
All data are expressed as mean ± standard deviation (x ± s). Data were analyzed and plotted using GraphPad Prism 9.0 (San Diego, CA, USA). Differences between groups were assessed by one-way ANOVA, and multiple comparisons were performed using the Tukey-Kramer test, with P<0.05 considered statistically significant.
Results
Analysis of the Chemical Composition of QBY
The chemical composition of QBY was analyzed using UHPLC-QE-MS to obtain the retention time and mass spectrometry data for each compound (Figure 2A and B), and 69 fractions were identified with high confidence based on scores, including quercetin, baicalein, glycyrrhizic acid, glabridin, forsythin, and licoflavone A (Table S1).
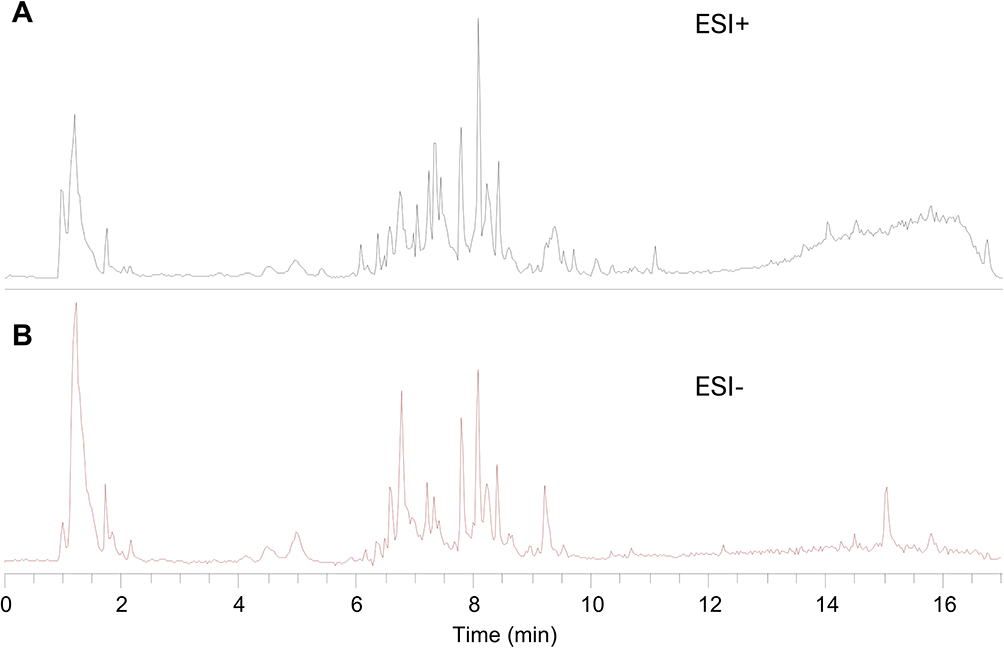 |
Figure 2 Chemical composition analysis of QBY. (A) The total ions current (TIC) in positive ion mode; (B) The TIC in negative ion mode. |
Potential Target Genes for Psoriasis Treatment by QBY
Target prediction analysis using these databases yielded 754 potential targets for QBY chemical compounds (Table S2). A comprehensive collection of 1869 non-duplicated psoriasis-related target genes was obtained from GeneCards, OMIM, TTD, and DisGeNET databases using “psoriasis” as the search keyword. Intersection analysis between the QBY targets and psoriasis-associated genes identified 262 overlapping targets, representing potential therapeutic targets for psoriasis treatment (Figure 3A).
 |
Figure 3 Functional enrichment analysis of intersecting target genes. (A) Venn diagram of QBY targets for psoriasis; (B) GO functional enrichment analysis of the targets of QBY in the treatment of psoriasis (top10). (C) KEGG pathway enrichment analysis of the target of QBY in the treatment of psoriasis (top 25). Different colors indicate different KEGG Classification. The red box highlights the pathway of interest. |
GO and KEGG Pathway Enrichment Analysis
GO and KEGG pathway enrichment analyses were performed on 262 overlapping targets using a significance threshold of P< 0.05. A total of 182 KEGG pathways and 3440 GO functional entries were obtained, including 124 CC, 282 MF, and 3034 BP terms. The top ten enriched terms in each GO category are shown as bar graphs (Figure 3B). Biological process enrichment predominantly involved immune and inflammatory responses, including responses to lipopolysaccharides, bacterial molecules, and regulation of the inflammatory response. Cellular component analysis showed enrichment of membrane rafts, membrane microdomains, and membrane region binding. Molecular function terms were significantly associated with G protein-coupled peptide receptor activity and related signaling mechanisms. KEGG pathway analysis further elucidated the functional landscape of overlapping genes (Table S3). Several signaling cascades critical for environmental information processing and organismal systems demonstrated significant enrichment, with particular emphasis on the sphingolipid, Th17 cell differentiation, and IL-17 signaling pathways (Figure 3C). These findings suggest that QBY exerts its anti-psoriasis effects by modulating multiple interconnected pathways involved in immune regulation and inflammatory responses.
Component–Target–Pathway Network Construction
Focusing on immune system processes and S1P-S1PR1-mediated signal transduction, we prioritized the top five pathways within these two secondary KEGG classification categories from the enriched pathway pool. The selected pathways included Th17 cell differentiation, PI3K-Akt signaling pathway, C-type lectin receptor signaling pathway, sphingolipid signaling pathway, and IL-17 signaling pathway. We identified 85 target genes associated with these pathways (Figure 4A). Further analyses showed that 60 compounds targeted these genes. To visualize the complex interactions, we constructed a compound–target–pathway network using the Cytoscape software (version 3.9.1) (Figure 4B). Through degree centrality-based network topology analysis, we identified the top ten bioactive compounds ranked by degree values for molecular docking, with baicalein being the most prominent candidate.
 |
Figure 4 Network analysis of enriched pathways, target genes, and bioactive compounds. (A) Sankey diagram of the distribution of 85 target genes across five significant KEGG pathways. (B) Compound-target-pathway network. Purple circular nodes indicate hub targets, green triangular nodes indicate bioactive compounds, and pink nodes indicate pathways. |
PPI Network Interaction Analysis
The obtained 85 intersection targets were constructed on the STRING platform using a PPI network. The data were imported into Cytoscape 3.9.1 software for visual analysis (Figure 5A). The MCC algorithm using Cytoscape software yielded the top five hub genes, namely, STAT3, Jun proto-oncogene (JUN), B-cell lymphoma 2 (BCL2), interleukin 6 (IL-6), and tumor necrosis factor (TNF) (Figure 5B).
 |
Figure 5 Network Pharmacology diagram of 85 target genes. (A) Protein-protein interaction network diagram (B) Hub genes of 85 target genes. |
QBY Regulates S1P/S1PR1 Axis Signaling in Th17 Polarized Cells
To investigate the effect of QBY on the S1P/S1PR1 pathway, we treated Th17 polarized cells with QBY for 48 h and detected S1P secretion using ELISA. Untreated and vehicle-treated groups were designated as negative groups. We found that QBY significantly reduced S1P content in an in vitro Th17 cell model (Figure 6A). S1P is produced by sphingosine kinases 1 and 2 (SPHK1 and SPHK2, respectively), and S1P lyase (encoded by the Sgpl1 gene) irreversibly breaks down S1P. At the mRNA level, QBY significantly reduced the expression of Sphk1 and Spnk2 in Th17 cells compared to that in the untreated group and elevated the expression of Sgpl1. Meanwhile, the addition of solvent alone showed no significant change (Figure 6B–D). This suggested the inhibitory effect on S1P expression. To further investigate how QBY regulates signaling downstream of S1P, we blocked S1P/S1PR1 pathway signaling using the S1PR1 receptor antagonist W146. This significantly the mRNA expression levels of Il17a and Rorc (encoding the master transcription factor RORγt) in S1P-treated Th17-polarized cells (Figure S2). The addition of 5 mM W146 significantly inhibited the expression of both genes. Subsequent experiments were conducted at this concentration. As shown in Figure 6E, compared to untreated cells, QBY reduced the expression of S1pr1 in Th17 polarized cells. However, the difference was not significant. Compared with the S1P-treated cell group, QBY and W146 significantly reduced the expression of S1pr1. Compared to W146 alone, the combination with QBY further reduced the level of S1pr1 expression, and there was no significant difference compared to the blank group of Th17 cells.
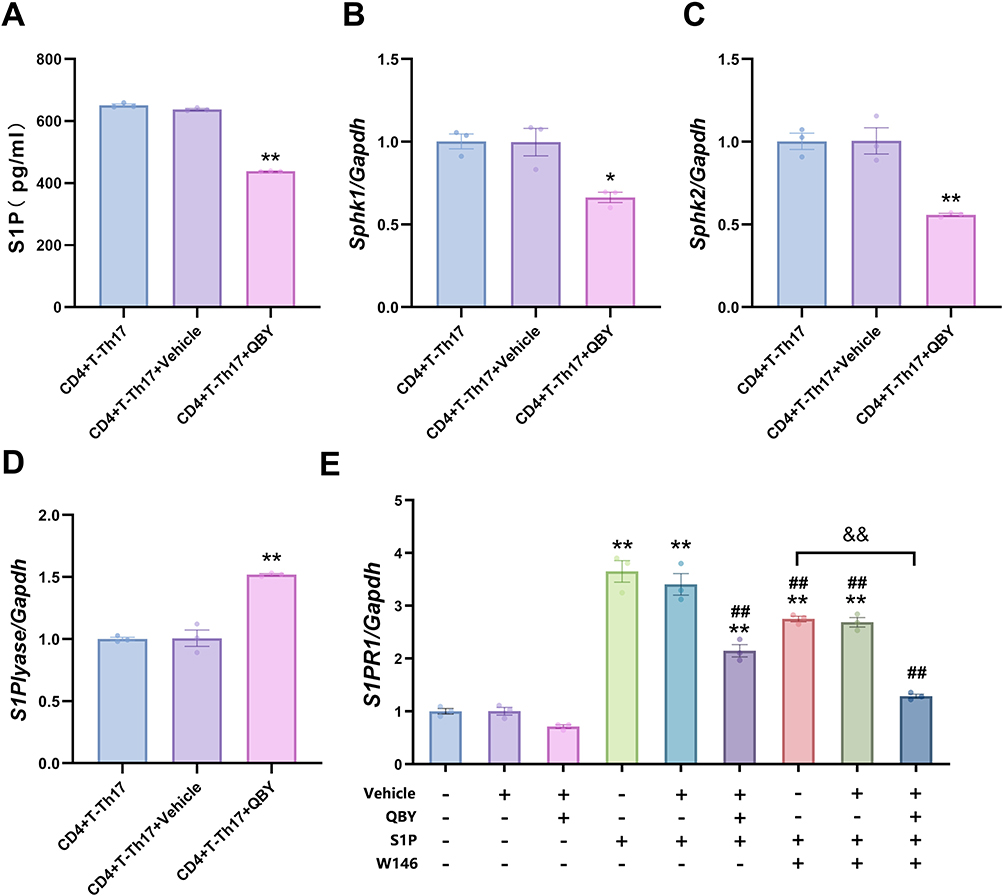 |
Figure 6 Effect of QBY on the S1P/S1PR1 pathway. (A) The concentration of S1P was measured by ELISA. (B–E) The mRNA expression levels of SphK1, SphK2, Sgpl1 and S1pr1 were detected by RT-qPCR. Data are expressed as mean ± SD (n=3). *P<0.05 and **P <0.01 versus Th17-CD4+T cell group; ##P <0.01 versus Th17-CD4+T cells +S1P group; &&P<0.01 versus Th17-CD4+T cells +S1P+W146 group. |
QBY Affects Th17 Proliferation and Differentiation Through the S1P/S1PR1 Axis
To examine the influence of QBY on Th17 cell proliferation and differentiation in vitro, we used CCK8 assays and flow cytometry to assess proliferation activity and differentiation rates. Treatment with S1P significantly increased Th17 polarization compared to that in untreated controls (Figure 7A and B). W146 and QBY effectively inhibited S1P-induced Th17 polarization. Cells treated with a combination of W146 and QBY showed a greater reduction in Th17 polarization than cells treated with W146 alone. The CCK8 assay results demonstrated similar trends in cell proliferation across all treatment groups (Figure 7C). These findings indicate that QBY could suppress the differentiation and proliferation of CD4+ Th17 cells, potentially through downregulation of the S1P/S1PR1 signaling pathway.
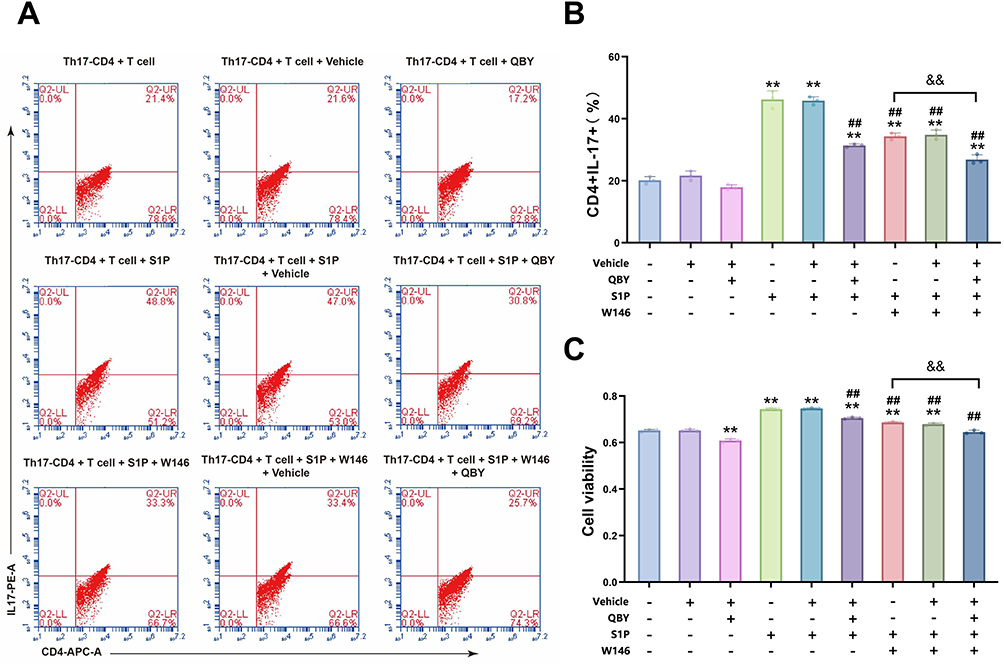 |
Figure 7 Effect of QBY on the proliferation and differentiation of Th17 cells in vitro. (A and B) Flow cytometric analysis of CD4+ Th17 cells. (C) Th17 Cell viability assessment using CCK-8 assay. Data are expressed as mean ± SD (n=3). **P<0.01 versus Th17-CD4+T cell group; ##P<0.01 versus Th17-CD4+T cells +S1P group; &&P<0.01 versus Th17-CD4+T cells +S1P+W146 group. |
QBY Affects the Secretion of Inflammatory Factors in Th17 Cells Through the S1P/S1PR1 Axis
Th17 cells produce key inflammatory mediators, including IL-17, IL-22, and IL-23, which play crucial roles in the pathogenesis of psoriasis. To assess the effect of QBY on the expression and secretion of inflammatory factors in Th17 cells, we conducted RT-qPCR and ELISA analyses for IL-17, IL-22, and IL-23 (Figure 8). RT-qPCR results showed that S1P treatment significantly upregulated the expression of Il-17 mRNA in Th17 cells (P<0.01; Figure 8A). This enhanced expression was significantly attenuated by W146 treatment (P<0.01). In cells treated with both S1P and W146, QBY treatment further reduced Il-17 mRNA levels (P<0.01).
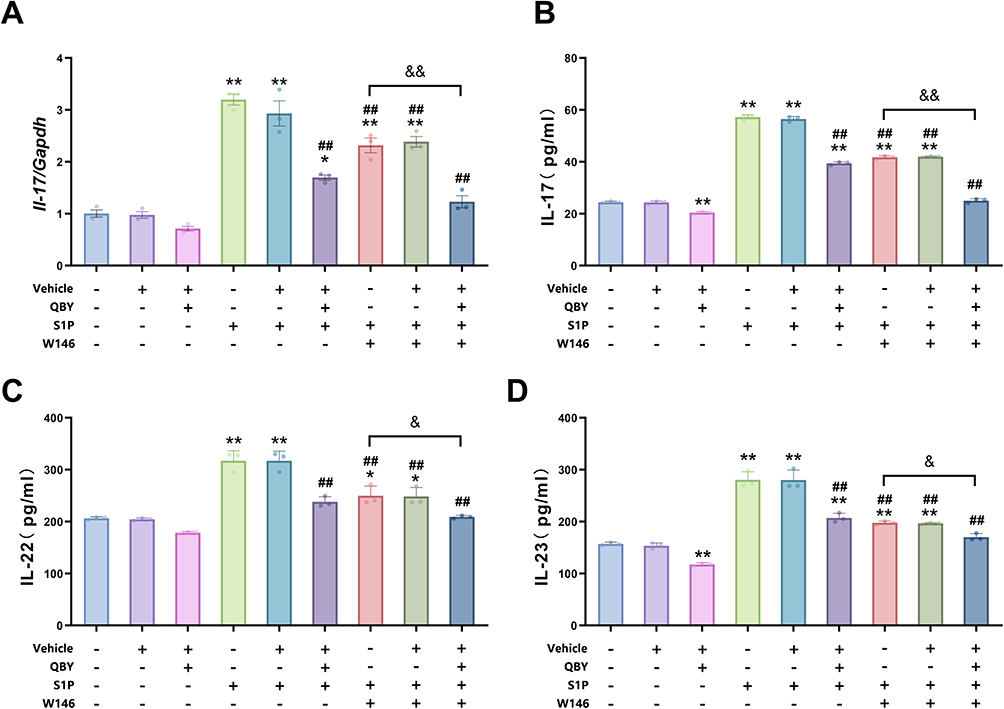 |
Figure 8 Regulation of the Expression and Secretion of the Inflammatory Factors IL-17, IL-22 and IL-23 by QBY in Th17 Cells. (A) Effects of S1P, W146 and QBY treatments on Il-17 mRNA expression in Th17 cells. (B–D) Effects of S1P, W146 and QBY treatments on the secretion level of IL-17, IL-22, IL-23 in Th17 cells. Data are expressed as mean ± SD (n=3). *P<0.05 and **P <0.01 versus Th17-CD4+T cell group; ##P <0.01 versus Th17-CD4+T cells +S1P group; &P <0.05 and &&P<0.01 versus Th17-CD4+T cells +S1P+W146 group.c. |
ELISA showed that S1P stimulation significantly increased the secretion of IL-17, IL-22, and IL-23 (all P<0.01, Figure 8B–D) from Th17 cells. Meanwhile, W146 treatment significantly inhibited this effect (P<0.01). In the presence of S1P and W146, QBY reduced the secretion of three inflammatory factors (P<0.01 for IL-17 and P<0.05 for both IL-23 and IL-22). QBY treatment resulted in a slight but statistically significant inhibition of IL-17 and IL-23 secretion from Th17 cells under basal conditions (P<0.01). This suggested that QBY may have an inflammation-suppressive effect independent of S1P. The results indicated that S1P promoted the expression and secretion of inflammatory factors in Th17 cells. This effect was inhibited by W146, and further enhanced inhibition was obtained by the combined application of QBY.
QBY Affected the Expression of STAT3 and SMAD2 by Regulating S1P/S1PR1 Axis
To elucidate the mechanisms by which QBY modulates Th17 differentiation, we investigated its effects on the TGF-β/SMAD and JAK/STAT signaling pathways. These play crucial roles in regulating Th17 differentiation (Figure 9A). SMAD2 and STAT3 play opposing roles in modulating STAT3-induced transcription relevant to Th17 differentiation.15 Western blot analysis showed that QBY significantly increased phosphorylated SMAD2 (p-SMAD2) levels and the p-SMAD2/SMAD2 ratio (P<0.01, Figure 9B and C). Meanwhile, total SMAD2 remained unchanged (Figure 9D). Conversely, S1P reduced p-SMAD2 levels, an effect that was attenuated by QBY co-treatment (P<0.01 vs S1P alone). Similarly, the S1P1 antagonist, W146, counteracted the S1P-mediated reduction in p-SMAD2, with W146 and QBY showing synergistic effects (P<0.01 vs W146 alone). QBY also significantly downregulated phosphorylated STAT3 (p-STAT3) expression and the p-STAT3/STAT3 ratio (P<0.01, Figure 9E and F), countering S1P-induced STAT3 phosphorylation. Meanwhile, total STAT3 remained unchanged (Figure 9G). The combination of QBY and W146 exhibited a more pronounced inhibitory effect on STAT3 phosphorylation than W146 alone (P<0.01). These findings suggest that QBY modulates Th17 cell differentiation by upregulating SMAD2 phosphorylation and downregulating STAT3 phosphorylation via the S1P/S1PR1 axis, with synergistic effects observed in combination with W146.
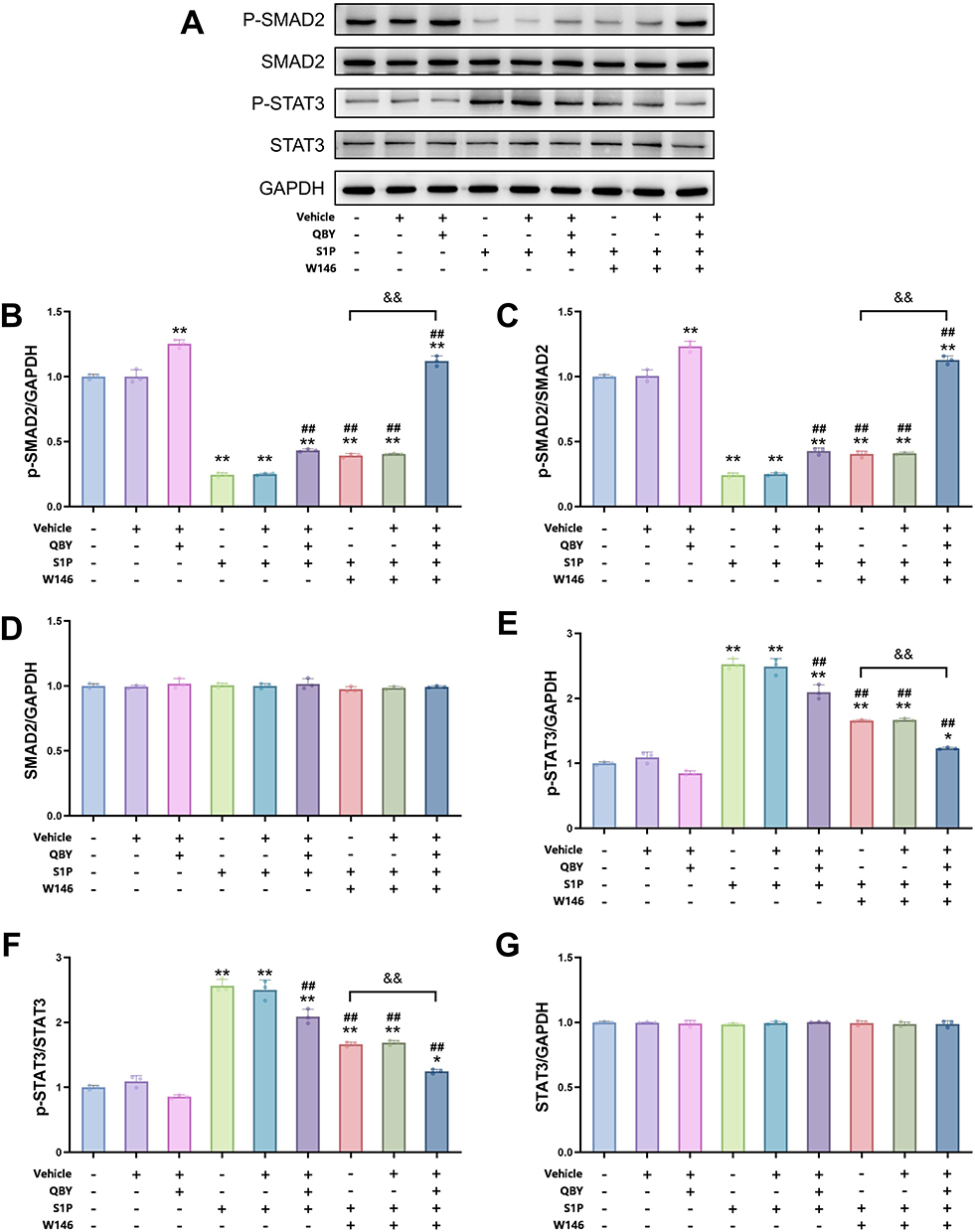 |
Figure 9 Effect of QBY on STAT3 and SMAD2 phosphorylation via the S1P/S1PR1 pathway. (A) Western blot images of p-SMAD2, SMAD2, p-STAT3, STAT3, and GAPDH expression in response to vehicle, QBY, S1P, and W146 treatments. (B–G) Densitometric analysis of the corresponding band intensity ratios: (B) p-SMAD2/GAPDH, (C) p-SMAD2/SMAD2, (D) SMAD2/GAPDH, (E) p-STAT3/GAPDH, (F) p-STAT3/STAT3, and (G) STAT3/GAPDH. Data are expressed as mean ± SD (n=3). *P<0.05 and **P<0.01 versus Th17-CD4+T cell group; ##P<0.01 versus Th17-CD4+T cells +S1P group; &&P<0.01 versus Th17-CD4+T cells +S1P+W146 group. |
Molecular Docking Evaluation of the Binding of Hub Targets
Combining the results of the screening of network pharmacological hub targets and in vitro experiments, we investigated the affinity between STAT3, JUN, BCL2, IL-6, TNF, SMAD2, S1PR1, SPHK1, RORγt, IL-17A, and the main active ingredients of QBY. Molecular docking analysis was performed to investigate the binding affinity between the 10 compounds and 10 target proteins using the CB-Dock2 platform. Based on the receptor-ligand docking hypothesis, docking energy is inversely proportional to binding affinity. As shown in Figure 10A, the heat map of docking scores showed that glabridin exhibited the strongest binding affinity with multiple targets, particularly with S1PR1 (−11 kcal/mol) and SPHK1 (−9.8 kcal/mol). This suggests that glabridin may be a potent modulator of S1P/S1PR1 signaling. Licoflavone A demonstrated considerable binding energy with TNF (−12.1 kcal/mol), while isobutyrylshikonin showed strong affinity with RORγt (−13 kcal/mol). Both of these are crucial regulators in Th17-mediated inflammatory responses.
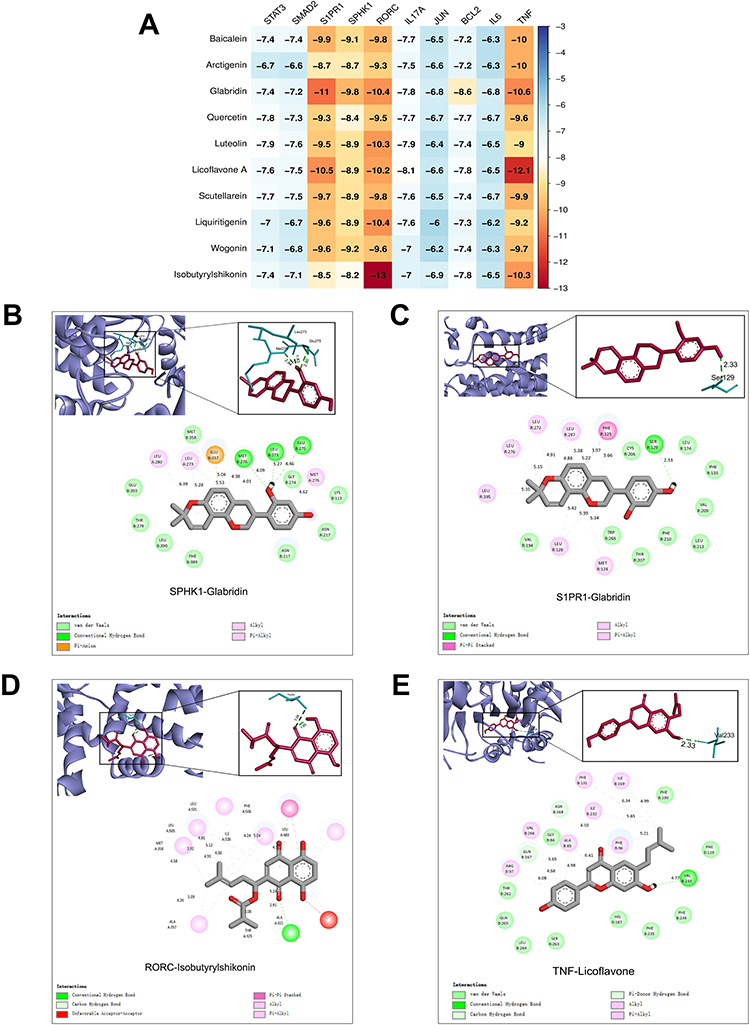 |
Figure 10 Molecular docking evaluation of the binding affinity of the main targets and components. (A) The docking score of the main targets and components. (B) 2D, 3D interaction diagrams of SPHK1 and glabridin molecular docking. (C) 2D, 3D interaction diagrams of SPPR1 and glabridin molecular docking. (D) 2D, 3D interaction diagrams of RORγt and isobutyrylshikonin molecular docking. (E) 2D, 3D interaction diagrams of TNF and licoflavone A molecular docking. |
To further elucidate the binding mechanisms, detailed molecular interactions were analyzed for the complexes with the highest binding affinities. The 3D and 2D interaction diagrams (Figure 10B) showed that glabridin formed stable interactions within SPHK1’s binding pocket, primarily via van der Waals forces with multiple residues. Conventional hydrogen bonds were observed, which contributed to the complex stability. Similarly, when docked with S1PR1 (Figure 10C), glabridin established a network of interactions, including pi-alkyl interactions and hydrogen bonds, with key binding site residues.
The interaction between isobutyrylshikonin and RORγt exhibited the strongest binding affinity (−13 kcal/mol) among all tested combinations (Figure 10A). The 3D structural analysis (Figure 10D) revealed that isobutyrylshikonin fits well within RORγt’s binding pocket, forming critical hydrogen bonds with key residues. The 2D interaction diagram shows various types of non-covalent interactions, including hydrogen bonds and hydrophobic interactions, which stabilize the ligand-receptor complex. Similarly, the TNF-Licoflavone A complex exhibited significant binding, with Licoflavone A establishing multiple stabilizing contacts within the active site of TNF (Figure 10E). These molecular docking results suggested that isobutyrylshikonin could potentially be a RORγt inhibitor, licoflavone A could function as a TNF modulator, and glabridin may act as a dual regulator of the S1P/S1PR1 signaling pathway. These findings provide a structural basis for further experimental validation of the therapeutic potential.
Discussion
Psoriasis is a common chronic inflammatory disease characterized by epidermal hyperplasia. Th17 cells are major effector cells in psoriasis.2 The positive feedback loop formed by Th17 cells and keratinocytes around the IL-23/Th17 axis augments the inflammatory response in psoriasis through self-amplification.16 S1P is a metabolite of sphingosine. S1P and its receptor S1PRs play roles in various skin diseases, among which the S1P/S1PR1 pathway is closely related to psoriasis pathogenesis.5 Some researchers believe that S1P the S1PR1 axis has the same effect as IL-23.17
QBY was optimized from Xiaoyin Jiedu granules. Xiaoyin Jiedu granules have been clinically demonstrated to significantly reduce Th17 cell numbers and IL-23 levels in the peripheral blood of patients with psoriasis, while inhibiting the mRNA expression of Il-17, Rorc, Il-22, and Il-23 related to Th17 cells and improving psoriatic skin lesions in mice.10,18 The optimized QBY was confirmed to inhibit KC proliferation in vitro.11 In this study, we investigated QBY’s effect of QBY on Th17 cell differentiation and the inflammatory response through the S1P/S1PR1 axis using network pharmacology, in vitro experiments, and molecular docking approaches.
Network pharmacology analysis showed 212 overlapping targets between QBY and psoriasis. GO enrichment results suggested that the active compounds in QBY may inhibit psoriasis through various biological processes, including regulation of the inflammatory response and G protein-coupled peptide receptor activity. This suggests that S1PRs, a class of G protein-coupled receptors, are involved in the interaction between QBY and psoriasis. KEGG pathway analysis further strengthened these findings, showing QBY’s potential of QBY to modulate multiple key pathways, including Th17 cell differentiation, PI3K-Akt, IL-17, and sphingolipid signaling pathways. The identification of these interconnected pathways demonstrates that QBY achieves its therapeutic effects by modulating multiple disease-relevant mechanisms simultaneously rather than targeting a single pathway.
Through component–target–network analysis, we identified key components targeting signal transduction and immune system-related pathways, including baicalein, glabridin, luteoli, as well as five hub genes: STAT3, JUN, BCL2, IL-6, and TNF. STAT3 is crucial for Th17 cell differentiation. IL-6 triggers STAT3 phosphorylation and enhances Th17 cells differentiation.19 TNF, as a common proinflammatory factor, cooperates with IL-6 to promote STAT3 phosphorylation20 and works with TGF-β to induce RORγt expression in naive CD4+ T cells independent of IL-6 and STAT3.21 In IL-23-dependent pathogenic Th17 cells, JUNB plays a crucial role in RORγt expression by enhancing the DNA binding of BATF, IRF4, and STAT3 to the RORC locus.22 These reports suggest that these hub genes participate in the progression of Th17 inflammation.
It was reported23 that baicalein inhibits the S1P-STAT3 signaling pathway by reducing the expression of SPHK1, S1PR1, and decreasing phosphorylation of STAT3. Quercetin inhibits the SPHK1/S1P signaling pathway.24 Shikonin, the parent compound of isobutyrylshikonin, reduced STAT3 dimer expression.25 Shikonin exhibits promising anti-psoriatic effects when delivered through keratinocyte membrane-mediated nanodelivery systems. This suggests that optimized delivery methods could enhance the clinical efficacy of traditional herbal formulations.26 These findings suggest that QBY may regulate both the upstream and downstream components of S1P and Th17 inflammatory processes through multiple bioactive compounds with established therapeutic potential.
S1P is a signal transduction lipid that controls immune cell migration, vascular development, and multiple inflammation processes.27,28 Given that psoriasis is characterized by local immune cell infiltration and angiogenesis, the significantly elevated S1P levels in the circulation and skin of patients with psoriasis compared to healthy individuals.29,30 S1P synthesis occurs via two sphingosine kinases, SPHK1 and SPHK2, collectively referred to as SPHKs. It is then degraded by S1P lyase, S1P phosphatase 1, or S1P phosphatase 2.4 Among them, S1P lyase irreversibly degrades S1P and is essential for attenuating S1P signaling.31
SPHKs promote human Th17 cell differentiation and IL-17 secretion.32 Our results showed that QBY downregulated SPHKs expression in Th17 cells while upregulating S1P lyase and decreasing S1P content. This suggests that QBY may improve psoriasis via the S1P-related pathway. Extracellular S1P binds to the membrane S1PRs for signal transduction. S1P enhances Th17 cell development via S1PR1-mediated STAT3 activation.33 The S1P/S1PR1 axis promotes Th17 differentiation both in vivo and in vitro.7,17 S1PR1 mediates Th17 cell migration from the thymus to the skin.
Our previous study18 found that Xiaoyin Jiedu granules modulate S1PR1 expression and suppress Th17 cells in mice. QBY reduces S1pr1 and Il-17 mRNA levels in murine Th17-polarized cells in vitro. The CCK8 and flow cytometry results showed that QBY inhibited Th17 cell proliferation and CD4 + T cell polarization to Th17 cells through the S1P/S1PR1 axis.
IL-23/Th17 cells are crucial in the pathogenesis of psoriasis. IL-23 binding to its receptor activates the JAK/STAT pathway, particularly STAT3, which stimulates Th17 cell proliferation and activation. Activated Th17 cells secrete cytokines, such as IL-17 and IL-22, which act on keratinocytes to cause excessive proliferation and epidermal thickening. Simultaneously, stimulated keratinocytes produce a variety of inflammatory factors and chemokines, including IL-23, which recruit more immune cells to the skin. The recruitment of immune cells produces more IL-23 and IL-17, which perpetuate and amplify the inflammatory cycle and explain the chronic nature of psoriasis.34,35 QBY reduced the levels of the inflammatory cytokines IL-17, IL-23, and IL-22 by inhibiting the S1P/S1PR1 pathway. It inhibited the secretion of inflammatory factors beyond the blocking effect of W146. This suggests that the multitarget approach of TCM provides advantages over single-target blocking in inflammation inhibition.
Th17 differentiation is regulated by multiple pathways, including JAK/STAT and TGF-β/SMAD. In the JAK/STAT pathway, STAT3 acts as a major positive regulator and translocates to the nucleus after activation to regulate RORγt expression in Th17 cells.36 Lee37 suggested that there may be a positive feedback loop between S1PR1 and STAT3, with STAT3 serving as an S1PR1 gene transcription factor while directly promoting its expression. Conversely, increased S1PR1 expression activates STAT3 and upregulates IL-6 gene expression. In the TGF-β/SMAD pathway, SMAD2 positively regulates Th17 differentiation through interaction with RORγt, while SMAD3 acts as a negative regulator. Th17 differentiation is reduced in Smad2-deficient mice but remains intact in Smad3-deficient mice.38
QBY promoted SMAD2 phosphorylation and inhibited STAT3 phosphorylation through the S1P/S1PR1 axis, achieving multitarget regulation. Molecular docking analysis provided structural insights into the interactions between the QBY components and their molecular targets. Glabridin showed a strong binding affinity to S1P/S1PR1 pathway targets, including S1PR1 and SPHK2. Meanwhile, isobutyrylshikonin, licoflavone A, and luteolin demonstrated preferential binding to Th17 inflammatory pathway genes.
Glabridin was reported to inhibit TNF-α-induced sphingosine kinase activity39 and downregulate IL-17A, IL-22, and IL-23 expression in HaCaT cells following TNF-α stimulation.40 Glabridin appears to be QBY’s main active component in regulating Th17 differentiation through the S1P/S1PR1 pathway. Although molecular docking simulations provide initial insights, future studies should experimentally validate glabridin binding to SPHK1 and S1PR1 to elucidate their precise molecular interactions and binding mechanisms.
Dysregulation of the sphingolipid pathway has been mentioned as a potential link between psoriasis and comorbidities such as cardiovascular diseases and metabolic disorders.41 Altered sphingolipid metabolism can affect multiple organ systems and may explain the systemic nature of psoriasis. Future research should explore the effects and mechanisms of TCM on psoriasis comorbidities, using sphingolipid metabolism as an entry point. QBY’s ability of QBY to improve psoriasis symptoms by targeting multiple pathways reflects the holistic theoretical basis of traditional Chinese medicine. Compared to traditional single-target therapies, QBY may offer more comprehensive disease control and potentially reduce the likelihood of therapeutic resistance.
This study has several limitations. Network pharmacology provides valuable mechanistic insights and computational predictions with inherent limitations. This method primarily considers molecular solubility and permeability, while excluding the actual content of active components in herbs, potentially excluding important bioactive compounds that may show enhanced bioavailability through supramolecular and intermolecular interactions during decoction preparation. Although well-controlled, the in vitro experimental system may not fully recapitulate the complex tissue environment and systemic factors present in human psoriasis. Molecular docking studies provide structural binding predictions but do not confirm the actual biological interactions or functional consequences.
Future research should focus on validating these key findings in vivo, using appropriate animal models for psoriasis. Investigating individual component contributions versus whole-formula effects could optimize therapeutic formulations. Patient applicability studies are needed to identify optimal treatment populations and determine QBY positioning within the current treatment algorithms. To facilitate the clinical translation of QBY, future efforts should focus on the development of a more convenient and stable granule formulation. Establishing a standardization system is crucial. This includes standardized extraction processes and fingerprint chromatography for quality consistency. These measures will lay the groundwork for the development of a marketable, standardized traditional Chinese medicine preparation. Large-scale clinical trials are warranted to rigorously evaluate the long-term efficacy and safety of providing higher-level evidence-based medical support for its clinical application.
Conclusion
This study demonstrated that QBY treats psoriasis through multipathway regulation, particularly via the S1P/S1PR1 axis. QBY modulates S1P synthesis and catabolism, inhibits Th17 cell differentiation, and reduces secretion of inflammatory cytokines. Glabridin was identified as the key active component targeting S1PR1 and SPHK1 based on molecular docking and its documented anti-inflammatory properties. Although network pharmacology provides mechanistic predictions, experimental validation has confirmed key aspects of the S1P/S1PR1 pathway modulation. However, distinguishing between in silico predictions and experimentally validated mechanisms requires further study. These findings provide mechanistic evidence for QBY’s anti-psoriatic effects of QBY. However, in vivo and clinical validation are essential to establish its translational relevance. Future studies should incorporate biomarker analyses to establish the direct correlations between pathway modulation and therapeutic outcomes in patients with psoriasis.
Abbreviations
QBY, Qing Bi Yin; STAT3, signal transducer and activator of transcription 3; SMAD2, SMAD family member 2; IL-23, Interleukin-23; IL-22, Interleukin-22; IL-17, Interleukin-17; S1P, Sphingosine-1-phosphate; S1PR1–S1PR5, sphingosine-1-phosphate receptors 1 to 5; XYJD, Xiaoyin Jiedu granules; ROR-γt, Retinoic acid-related orphan receptor gamma t; UHPLC-QE-MS, ultra-high-performance liquid chromatography coupled with Q Exactive Orbitrap mass spectrometry; HESI, heated electrospray ionization; GO, Gene Ontology; KEGG, Kyoto Encyclopedia of Genes and Genomes; PPI, protein-protein interaction; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; CCK-8, cell counting Kit-8; ELISA, enzyme-linked immunosorbent assay; JUN, jun proto-oncogene; BCL2, B-cell lymphoma 2; IL-6, interleukin 6; TNF, tumor necrosis factor; SPHK1, sphingosine kinases 1; SPHK2, sphingosine kinases 2; Sgpl1, S1P lyase gene; p-STAT3, phosphorylated STAT3; p-SMAD2, phosphorylated SMAD2.
Ethics Approval
This study utilized data from publicly accessible databases, fully anonymized. According to Article 32 of the Measures for Ethical Review of Life Science and Medical Research Involving Human Subjects (issued February 18, 2023, China), research that (1) uses legally obtained public data or data generated by observing public behaviors without interference, or (2) uses anonymized information data, and does not involve sensitive personal information or commercial interests, is exempt from ethical review. As this study meets the criteria outlined in items (1) and (2) of Article 32, it is exempt from ethical approval (available at https://www.gov.cn/zhengce/zhengceku/2023-02/28/content_5743658.htm).
Acknowledgments
The authors are grateful to the support and assistance in terms of instruments and facilities provided by Dongzhimen Hospital, Beijing University of Traditional Chinese Medicine, Beijing, PR China. We would like to thank Editage (www.editage.cn) for English language editing.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was supported by the National Natural Science Foundation of China (No. 81874393) and The Fifth Batch of National Excellent Talents Training Program for Traditional Chinese Medicine Clinicians.
Disclosure
The authors declare that they have no known competing interests in this work.
References
1. Greb JE, Goldminz AM, Elder JT, et al. Psoriasis. Nat Rev Dis Primers. 2016;2(1):1–17. doi:10.1038/nrdp.2016.82
2. Griffiths CEM, Armstrong AW, Gudjonsson JE, Barker JNWN. Psoriasis. Lancet. 2021;397(10281):1301–1315. doi:10.1016/S0140-6736(20)32549-6
3. Al-Janabi A, Yiu ZZN. Biologics in psoriasis: updated perspectives on long-term safety and risk management. Psoriasis. 2022;12:1. doi:10.2147/PTT.S328575
4. Baeyens AAL, Schwab SR. Finding a way out: S1P signaling and immune cell migration. Ann Rev Immunol. 2020;38:759–784. doi:10.1146/annurev-immunol-081519-083952
5. Masuda-Kuroki K, Alimohammadi S, Di Nardo A. The role of sphingolipids and Sphingosine-1-phosphate—Sphingosine-1-phosphate-receptor signaling in psoriasis. Cells. 2023;12(19):2352. doi:10.3390/cells12192352
6. Kunkel GT, Maceyka M, Milstien S, Spiegel S. Targeting the sphingosine-1-phosphate axis in cancer, inflammation and beyond. Nat Rev Drug Discov. 2013;12(9):688–702. doi:10.1038/nrd4099
7. Shin SH, Cho KA, Hahn S, et al. Inhibiting sphingosine kinase 2 derived-sphingosine-1-phosphate ameliorates psoriasis-like skin disease via blocking Th17 differentiation of naïve CD4 T lymphocytes in mice. Acta Derm Venereol. 2019;99(6):594–601. doi:10.2340/00015555-3160
8. Liu L, Wang J, Li HJ, et al. Sphingosine-1-phosphate and its signal modulators alleviate psoriasis-like dermatitis: preclinical and clinical evidence and possible mechanisms. Front Immunol. 2021;12:759276. doi:10.3389/fimmu.2021.759276
9. Luo Y, Chen J, Kuai L, et al. Chinese herbal medicine for psoriasis: evidence from 11 high-quality randomized controlled trials. Front Pharmacol. 2021;11:599433. doi:10.3389/fphar.2020.599433
10. Chen X, Zhang R, Duan X, Xue M, Qu T, Li L. Effectiveness of xiaoyin jiedu granules in the treatment of psoriasis vulgaris in patients with blood-heat symptom patterns in terms of traditional Chinese medicine. J Tradit Chin Med. 2020;40(5):863–869. doi:10.19852/j.cnki.jtcm.2020.05.017
11. Zhang H, Wang Z, Guo Y, Li L, Hang H. Effects of Qingbi decoction on S1P/S1PR5 signaling pathway in HaCaT inflammatory model. Glob Tradit Chin Med. 2023;16(12):2434–2443.
12. Zhou L, Lopes JE, Chong MMW, et al. TGF-β-induced Foxp3 inhibits TH17 cell differentiation by antagonizing RORγt function. Nature. 2008;453(7192):236–240. doi:10.1038/nature06878
13. Liu Y, Yang X, Gan J, Chen S, Xiao ZX, Cao Y. CB-Dock2: improved protein-ligand blind docking by integrating cavity detection, docking and homologous template fitting. Nucleic Acids Res. 2022;50(W1):W159–W164. doi:10.1093/nar/gkac394
14. Pan J, Lu Y, Wang S, et al. Synergistic neuroprotective effects of two natural medicinal plants against CORT-induced nerve cell injury by correcting neurotransmitter deficits and inflammation imbalance. Phytomedicine. 2023;121:155102. doi:10.1016/j.phymed.2023.155102
15. Yoon JH, Sudo K, Kuroda M, et al. Phosphorylation status determines the opposing functions of Smad2/Smad3 as STAT3 cofactors in TH17 differentiation. Nat Commun. 2015;6(1):7600. doi:10.1038/ncomms8600
16. Deng Y, Chang C, Lu Q. The inflammatory response in psoriasis: a comprehensive review. Clin Rev Allerg Immunol. 2016;50(3):377–389. doi:10.1007/s12016-016-8535-x
17. Liao JJ, Huang MC, Goetzl EJ. Cutting edge: alternative signaling of Th17 cell development by sphingosine 1-phosphate. J Immunol. 2007;178(9):5425–5428. doi:10.4049/jimmunol.178.9.5425
18. Wang Z, Zhang G, Zhang H, Li L. Xiaoyin Jiedu Granules may alleviate psoriasis-like skin diseases in mice by regulating sphingosine 1-phosphate receptor expression and reducing Th17 cells. Heliyon. 2023;9(8):e19109. doi:10.1016/j.heliyon.2023.e19109
19. Xue C, Yao Q, Gu X, et al. Evolving cognition of the JAK-STAT signaling pathway: autoimmune disorders and cancer. Signal Transduct Target Ther. 2023;8(1):1–24. doi:10.1038/s41392-023-01468-7
20. Alam MS, Otsuka S, Wong N, et al. TNF plays a crucial role in inflammation by signaling via T cell TNFR2. Proc Natl Acad Sci USA. 2021;118(50):e2109972118. doi:10.1073/pnas.2109972118
21. Zheng Y, Sun L, Jiang T, Zhang D, He D, Nie H. TNFα promotes Th17 cell differentiation through IL-6 and IL-1β produced by monocytes in rheumatoid arthritis. J Immunol Res. 2014;2014(1):385352. doi:10.1155/2014/385352
22. Hasan Z, Koizumi SI, Sasaki D, et al. JunB is essential for IL-23-dependent pathogenicity of Th17 cells. Nat Commun. 2017;8(1):15628. doi:10.1038/ncomms15628
23. Yao J, Liu T, Chen RJ, Liang J, Li J, Wang CG. Sphingosine-1-phosphate signal transducer and activator of transcription 3 signaling pathway contributes to baicalein-mediated inhibition of dextran sulfate sodium-induced experimental colitis in mice. Chin Med J. 2020;133(03):292–300. doi:10.1097/CM9.0000000000000627
24. Zhang X, Cai Y, Zhang W, Chen X. Quercetin ameliorates pulmonary fibrosis by inhibiting SphK1/S1P signaling. Biochem Cell Biol. 2018;96(6):742–751. doi:10.1139/bcb-2017-0302
25. Kaur K, Sharma R, Singh A, et al. Pharmacological and analytical aspects of alkannin/shikonin and their derivatives: an update from 2008 to 2022. Chin Herb Med. 2022;14(4):511. doi:10.1016/j.chmed.2022.08.001
26. Jing Q, Ruan H, Li J, et al. Keratinocyte membrane-mediated nanodelivery system with dissolving microneedles for targeted therapy of skin diseases. Biomaterials. 2021;278:121142. doi:10.1016/j.biomaterials.2021.121142
27. Obinata H, Hla T. Sphingosine 1-phosphate and inflammation. Int Immunol. 2019;31(9):617–625. doi:10.1093/intimm/dxz037
28. Tsai HC, Han MH. Sphingosine-1-Phosphate (S1P) and S1P signaling pathway: therapeutic targets in autoimmunity and inflammation. Drugs. 2016;76(11):1067–1079. doi:10.1007/s40265-016-0603-2
29. Checa A, Xu N, Sar DG, Haeggström JZ, Ståhle M, Wheelock CE. Circulating levels of sphingosine-1-phosphate are elevated in severe, but not mild psoriasis and are unresponsive to anti-TNF-α treatment. Sci Rep. 2015;5(1):12017. doi:10.1038/srep12017
30. Matwiejuk M, Mysliwiec H, Lukaszuk B, et al. The interplay between bioactive sphingolipids in the psoriatic skin and the severity of the disease. Int J Mol Sci. 2023;24(14):11336. doi:10.3390/ijms241411336
31. Kumar A, Zamora-Pineda J, Degagné E, Saba JD. S1P lyase regulation of thymic egress and oncogenic inflammatory signaling. Mediators Inflamm. 2017;2017:e7685142. doi:10.1155/2017/7685142
32. Barra G, Lepore A, Gagliardi M, et al. Sphingosine kinases promote IL-17 expression in human T lymphocytes. Sci Rep. 2018;8(1):13233. doi:10.1038/s41598-018-31666-1
33. Garris CS, Blaho VA, Hla T, Han MH. Sphingosine-1-phosphate receptor 1 signalling in T cells: trafficking and beyond. Immunology. 2014;142(3):347. doi:10.1111/imm.12272
34. Bugaut H, Aractingi S. Major role of the IL17/23 axis in psoriasis supports the development of new targeted therapies. Front Immunol. 2021;12:621956. doi:10.3389/fimmu.2021.621956
35. Fitch E, Harper E, Skorcheva I, Kurtz SE, Blauvelt A. Pathophysiology of psoriasis: recent advances on IL-23 and Th17 cytokines. Curr Rheumatol Rep. 2007;9(6):461–467.
36. Martín P, Gómez M, Lamana A, et al. CD69 association with Jak3/Stat5 proteins regulates Th17 cell differentiation. Mol Cell Biol. 2010;30(20):4877–4889. doi:10.1128/MCB.00456-10
37. Lee H, Deng J, Kujawski M, et al. STAT3-induced S1PR1 expression is crucial for persistent STAT3 activation in tumors. Nat Med. 2010;16(12):1421–1428. doi:10.1038/nm.2250
38. Zhang S. The role of transforming growth factor β in T helper 17 differentiation. Immunology. 2018;155(1):24–35. doi:10.1111/imm.12938
39. Kang JS, Yoon YD, Han MH, et al. Glabridin suppresses intercellular adhesion molecule-1 expression in tumor necrosis factor-α-stimulated human umbilical vein endothelial cells by blocking sphingosine kinase pathway: implications of akt, extracellular signal-regulated kinase, and nuclear factor-κB/rel signaling pathways. Mol Pharmacol. 2006;69(3):941–949. doi:10.1124/mol.105.017442
40. Li P, Li Y, Jiang H, et al. Glabridin, an isoflavan from licorice root, ameliorates imiquimod-induced psoriasis-like inflammation of BALB/c mice. Int Immunopharmacol. 2018;59:243–251. doi:10.1016/j.intimp.2018.04.018
41. Matwiejuk M, Mysliwiec H, Chabowski A, Flisiak I. The role of sphingolipids in the pathogenesis of psoriasis. Metabolites. 2022;12(12):1171. doi:10.3390/metabo12121171
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Potential Targets and Mechanisms of Saikosaponin D in Psoriasis: A Bioinformatic and Experimental Study on Oxidative Stress
Ding J, Sun A, Hu H, Li J, Lu S, Song J, He J, Song X, Qian S, Tian Z
Journal of Inflammation Research 2025, 18:16487-16507
Published Date: 26 November 2025
Network Pharmacology and Animal Experimental Validation on the Therapeutic Mechanisms of Prunus cerasifera Extract in Psoriasis
Wang H, Ding Y, Wang Z, Wang Y, Liu Y, Ma Y, Wu W
Clinical, Cosmetic and Investigational Dermatology 2026, 19:608837
Published Date: 27 June 2026