Back to Journals » International Journal of Nanomedicine » Volume 21
Mechanism-Guided Nanoengineered Therapeutic Peptides for Bone Healing
Authors Zou SQ ![]() , Yu JC, Li Y, Chai LZ, Ye YJ
, Yu JC, Li Y, Chai LZ, Ye YJ
Received 30 March 2026
Accepted for publication 10 June 2026
Published 29 June 2026 Volume 2026:21 613192
DOI https://doi.org/10.2147/IJN.S613192
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Dong Wang
Shan-Qi Zou,1,* Jing-Chao Yu,2,* Yang Li,3 Ling-Zhi Chai,4 Yong-Jing Ye5
1Foot and Ankle Group of Orthopedics, The Second People’s Hospital of Quzhou, Quzhou, 324000, People’s Republic of China; 2Quzhou Hospital of Traditional Chinese Medicine, Quzhou, 324000, People’s Republic of China; 3Orthopedics, The Second People’s Hospital of Quzhou, Quzhou, 324000, People’s Republic of China; 4The Second People’s Hospital of Quzhou, Quzhou, 324000, People’s Republic of China; 5Anesthesiology Department, The Second People’s Hospital of Quzhou, Quzhou, 324000, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Ling-Zhi Chai, The Second People’s Hospital of Quzhou, Quzhou, 324000, People’s Republic of China, Email [email protected] Yong-Jing Ye, Anesthesiology Department, The Second People’s Hospital of Quzhou, Quzhou, 324000, People’s Republic of China, Email [email protected]
Abstract: Bone healing is frequently compromised not by the absence of a single osteogenic factor, but by the breakdown of coordinated biological processes, including inflammatory resolution, vascular invasion, endogenous cell recruitment, matrix mineralization, and remodeling. Therapeutic peptides are attractive for bone repair because they are chemically defined, modular, and readily engineered to encode diverse functions such as immunomodulation, angiogenic activation, osteogenic signaling, antimicrobial activity, and mineral binding. However, in their free or conventionally delivered forms, peptides often suffer from rapid degradation, diffusion, burst release, poor local retention, and inadequate spatiotemporal presentation, limiting their efficacy in complex defects. Nanoengineering provides the missing level of control by converting sequence-defined peptides into retained, clustered, responsive, and matrix-integrated nanoscale signals that can be interpreted by cells within the evolving defect niche. Unlike previous reviews that are typically organized by material platform (eg, hydrogels, scaffolds) or by peptide category (eg, biomimetic, antimicrobial), this Review adopts a mechanism-guided framework that is structured around the sequential biological bottlenecks of bone healing—from inflammatory resolution and vascular invasion to mineralization and remodeling—and critically examines how nanoengineering transforms peptide sequence information into spatiotemporally effective signals at each stage. It first discusses how sequence features encode regenerative potential and how nanoscale presentation determines whether that potential becomes biologically effective. The field is then organized according to the major biological bottlenecks that peptide nanoengineering can overcome, including immuno-osteogenesis, angiogenic–osteogenic coupling, mineralization guidance, and endogenous cell recruitment. Problem-oriented applications in compromised healing are further examined, with emphasis on infected bone defects and osteoporotic bone repair, and it is argued that biomaterialization acts as a mechanism-amplifying extension of peptide nanoengineering by adding four critical functions: retention, geometry, responsiveness, and defect-specific context matching. Finally, emerging opportunities in AI-assisted peptide design, stage-matched delivery, mineralization-guiding nanoarchitectures, and translational validation in load-bearing and nonunion-relevant models are highlighted. Together, this Review positions nanoengineered therapeutic peptides as a promising preclinical platform that may advance toward more targeted and mechanism-informed strategies for bone regeneration. The infographic illustrates four main sections: 1) Peptide sequence design includes charge and amphiphilicity, self-assembly propensity, receptor-binding motifs and mineral-binding/phosphoserine motifs. 2) Nanoengineering and biomaterialization involve self-assembled nanofibers, supramolecular hydrogels, mesoporous nanocarriers, scaffold coatings and responsive biomaterial matrices. Key features include retention, geometry and responsiveness to stimuli like enzymes, light and ultrasound. 3) Mechanism-guided regenerative functions cover immuno-osteogenesis with M1-like and M2-like cells, angiogenic-osteogenic coupling, mineralization guidance and endogenous cell recruitment. 4) Defect-specific bone healing addresses infected defects, osteoporotic defects and implant interfaces, highlighting reduced bacteria, improved vascularization and osseointegration. Key highlights include sequence-encoded precision, multifunctional integration and stimulus-responsiveness.Infographic on peptide design, nanoengineering, regenerative functions and bone healing processes.
Keywords: bone healing, therapeutic peptides, nanoengineering, biomaterialization, osseointegration
Introduction
Bone possesses a remarkable capacity for regeneration after injury; however, this reparative program becomes strikingly vulnerable when the local healing niche is destabilized by excessive or unresolved inflammation, vascular insufficiency, impaired progenitor-cell function, infection, age- or osteoporosis-associated microenvironmental decline, or the combined biological and mechanical burden of large segmental defects and nonunion.1,2 In such settings, healing failure rarely reflects the absence of a single osteogenic factor. Rather, it arises from the progressive breakdown of coordinated regenerative events spanning immune regulation, angiogenic invasion, endogenous cell recruitment, matrix deposition, mineralization, and remodeling.3 This multiscale complexity helps explain why static graft substitutes and one-dimensional osteoinductive strategies often underperform in clinically difficult defects, and why effective regenerative interventions must be designed to operate across interconnected inflammatory, vascular, osteogenic, and remodeling axes.3,4
Therapeutic peptides are particularly attractive in this context because they offer a chemically defined and highly programmable molecular toolkit for addressing multiple regenerative bottlenecks.5 Unlike full-length proteins, peptides can be synthesized with high precision, readily modified, and modularly combined to encode specific functions such as cell adhesion, osteogenic signaling, angiogenic activation, antimicrobial activity, immunomodulation, protease responsiveness, or mineral binding.4,6–8 Sequence-level parameters—including charge, amphiphilicity, secondary-structure propensity, phosphorylated or acidic motifs, substrate-binding segments, and multifunctional fusion design—allow peptide bioactivity to be tailored with unusual precision.6,9 As a result, peptides are not simply reduced versions of growth factors; they can be engineered as mechanism-specific regulators of the bone-healing microenvironment.10
Accordingly, a growing body of work has explored peptide-based strategies for bone repair from multiple perspectives.11 Previous reviews have discussed the determinants of osteogenic performance in peptide-modified bone-repair materials, the design and multifunctionality of peptide-functionalized hydrogels, biomimetic peptide categories for skeletal regeneration, extracellular-matrix-mimetic peptide systems, and peptide-conjugated biodegradable polyester scaffolds.12–14 (Table 1) In parallel, recent literature has also expanded toward problem-oriented and translationally framed discussions, including antimicrobial-peptide-enabled strategies for infected bone defects (eg, AMP-integrated scaffold systems and printing/AI-enabled design directions) and broader nanomedicine-driven design logic for complex bone defect repair. Other reviews have focused on peptide biofunctionalization of implants and hydrogels, or more broadly on biomimetic peptides in bone tissue engineering.15–18 Collectively, these studies have established peptides as versatile bioactive building blocks for skeletal repair. However, the literature is still organized predominantly by peptide class, nominal biofunction, or scaffold platform.19 While such categorizations are informative, they tend to treat each peptide or material as an independent variable and do not directly reveal why a given peptide strategy succeeds or fails at a specific stage of the healing cascade. In particular, this organizational logic makes it difficult to identify which biological bottleneck—whether unresolved inflammation, insufficient vascularization, disordered mineralization, or poor endogenous cell participation—is being addressed, and whether the nanoscale presentation format is mechanistically matched to that bottleneck. Reorganizing the field around these sequential biological checkpoints, rather than around peptide identity or carrier type, can offer several distinct advantages: it clarifies the mechanistic rationale for choosing a particular peptide–nanoengineering combination, exposes gaps where specific healing stages remain inadequately targeted, and provides a more direct bridge between material design decisions and the pathological logic of defect-specific repair failure.20
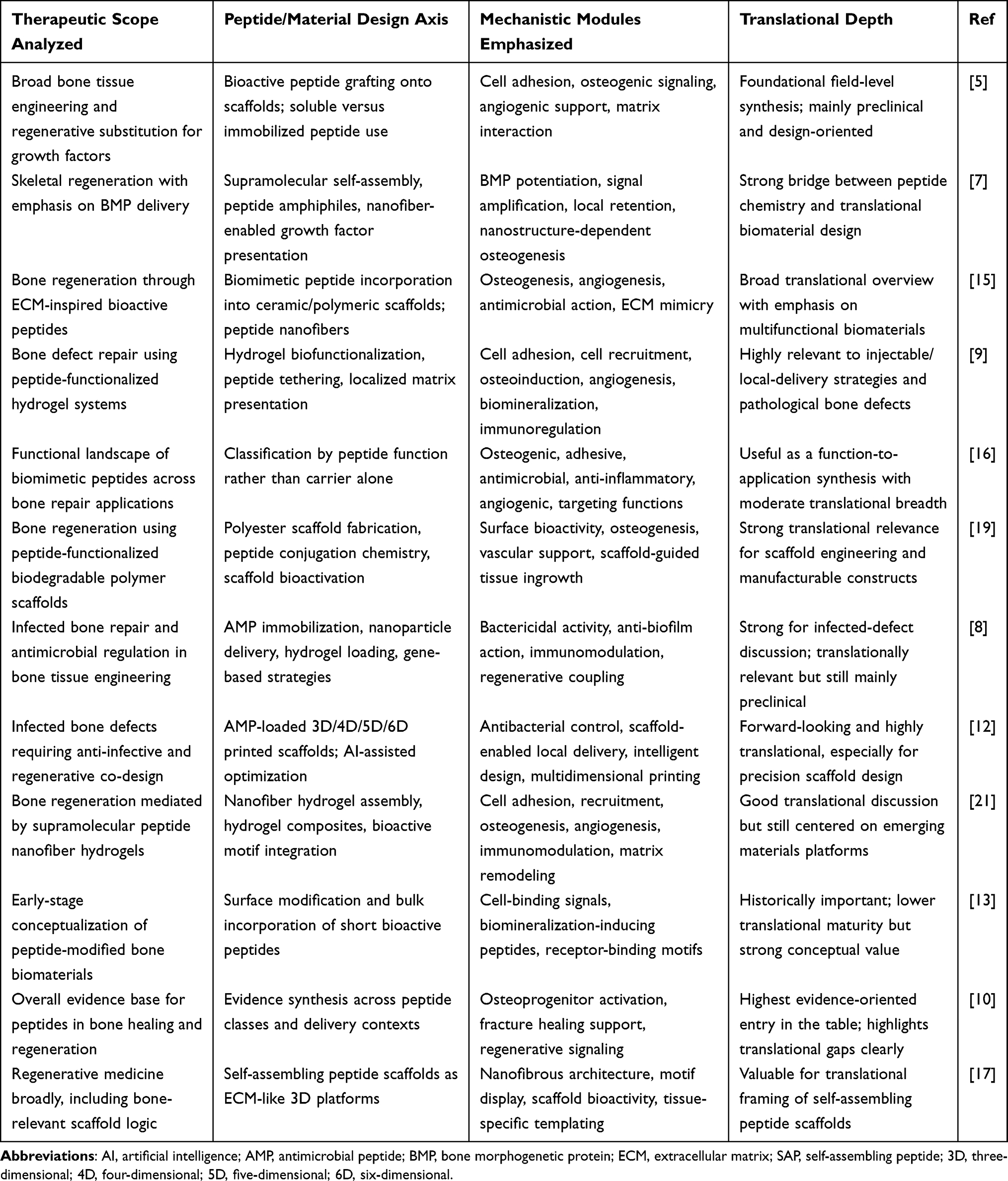 |
Table 1 Review Literature on Peptide-Enabled Bone Repair Deconstructed by Therapeutic Scope, Engineering Axis, Mechanistic Modules, and Translational Depth |
The main limitation of peptide therapeutics in bone repair is usually not lack of biological relevance, but lack of effective presentation.22 In their free or conventionally delivered form, peptides often suffer from rapid diffusion, enzymatic degradation, burst release, poor site retention, weak receptor clustering, and inadequate synchronization with the evolving phases of healing.16,19,23 These limitations are especially problematic in complex defects, where regenerative success depends on sustained local signaling, matrix-associated cue presentation, microenvironment-responsive release, and precise temporal coordination among immune, vascular, osteogenic, and remodeling processes.24,25 These delivery-related shortcomings are particularly consequential in complex defects, where the gap between a peptide’s intrinsic bioactivity and its actual therapeutic impact widens considerably.26
Nanoengineering provides that missing level of control.27 Once peptide motifs are self-assembled into nanofibers, organized into supramolecular hydrogels, clustered on scaffold surfaces, confined within mesoporous carriers, immobilized at implant interfaces, or integrated into responsive biomaterial systems, they no longer function merely as diffusible ligands.28 Instead, they become nanoscale instructive signals whose activity is shaped by multivalent presentation, local retention, interfacial density, proteolytic protection, stimulus responsiveness, and matrix-like organization. Such nanoengineered presentation can amplify peptide function in several ways: by increasing local concentration at the defect site, promoting receptor engagement and clustering, stabilizing peptide conformation, coupling signal exposure to inflammation- or enzyme-dependent events, and transforming soluble bioactivity into a structured regenerative microenvironment.29 Thus, nanoengineering introduces a qualitative shift: peptide cues are no longer passively available but become spatially encoded, temporally gated, and mechanistically coupled to the evolving biology of the defect.
On this basis, this Review focuses specifically on nanoengineered therapeutic peptides for bone healing, with emphasis on the mechanistic value of combining peptide design with nanoscale presentation. Rather than treating peptides and nanotechnology as separate topics, we examine how their integration generates stage-matched and defect-relevant regenerative functions that neither component can achieve as effectively alone. We first outline a conceptual framework linking peptide sequence features to nanoscale presentation and biological outcome. We then discuss how nanoengineered peptides act across major mechanistic bottlenecks in bone repair, including immunomodulation, angiogenic–osteogenic coupling, matrix mineralization, and endogenous cell recruitment. Finally, we consider how these principles can be translated toward problem-oriented applications in compromised healing, including infected defects, osteoporotic bone repair, and other clinically challenging regenerative contexts.
This is a narrative review. The relevant literature was identified through searches of PubMed, Web of Science, and Scopus using keyword combinations related to peptides, nanoengineering, and bone regeneration, primarily covering publications from 2010 to 2025. Studies were selected based on mechanistic relevance rather than formal systematic inclusion/exclusion criteria, given the high heterogeneity of material platforms, peptide classes, and biological endpoints across the field.
Conceptual Framework: Nanoengineering Converts Peptide Sequence into Bone-Healing Function
Therapeutic peptides for bone repair should not be viewed as isolated soluble ligands.30 They are better understood as sequence-defined bioactive instructions whose regenerative function depends on nanoscale presentation.31 In bone healing, the same peptide motif can produce markedly different outcomes depending on whether it is freely diffusible, self-assembled into nanofibers, immobilized on a scaffold, clustered at an implant interface, or revealed within a degradable and cell-responsive matrix.32 This distinction is mechanistically important because the major barriers to bone repair—immune dysregulation, poor vascular invasion, defective mineralization, and insufficient endogenous-cell participation—are all spatially and temporally organized processes.33,34 Sequence therefore encodes what a peptide can potentially do, whereas nanoengineering determines whether that potential is retained, amplified, and made biologically legible within the evolving defect niche.16
Sequence Features Encode Regenerative Potential
At the molecular level, peptide function is shaped by a small set of high-value sequence variables.35 Charge and amphiphilicity regulate membrane interaction, antimicrobial activity, adsorption to charged surfaces, and the propensity for supramolecular assembly.36,37 These features are particularly relevant in contaminated or implant-associated defects, where anti-infective activity must be balanced against host compatibility and interfacial retention.38 Modular fusion further expands peptide utility by integrating multiple functions—such as antimicrobial, angiogenic, osteogenic, and substrate-binding motifs—into a single construct, as illustrated by multifunctional titanium-binding peptide systems for infected or poorly integrating interfaces.39
A second major determinant is the relationship between secondary structure and receptor recognition. β-sheet-forming self-assembling peptides can generate fibrillar networks that resemble aspects of extracellular matrix (ECM) topology while improving local retention and supporting mineral-related activity. The self-assembling peptide P11-4 is a representative example: when physically adsorbed onto electrospun polycaprolactone nanofibers to form a hybrid fibrous scaffold—with a fiber diameter of approximately 500 nm—it preserved nanoscale fibrillar architecture, promoted mineral precipitation in simulated body fluid, enhanced cell-mediated mineral deposition, and improved bone regeneration in a rat calvarial critical-size defect model compared with peptide-free controls.40 A complementary in vivo demonstration was provided in a rat cranial defect study, in which P11-4 delivered as a self-assembling hydrogel promoted bone regeneration, with histological evidence of osseous island formation containing blood vessels.41 Together, these studies indicate that the fibrillar organization of P11-4 is integral to its mineralization-competent and regenerative performance, and that delivery format—whether as an adsorbed coating on electrospun fibers or as an injectable self-assembling gel—directly determines the functional state in which this β-sheet propensity is expressed.
By contrast, collagen-mimetic sequences such as glycine-phenylalanine-hydroxyproline-glycine-glutamate-arginine (GFOGER) show how peptide design can reproduce receptor-specific signaling rather than generic adhesiveness.23,42 When covalently immobilized within α2β1 integrin-specific PEG hydrogels and implanted in rat femoral segmental defects, GFOGER promoted integrin-mediated mesenchymal stem cell adhesion, survival, and osteogenic differentiation, and supported bone formation in vivo in a BMP-2 co-delivery setting.29,43,44 In a further extension to a critical-sized defect model, GFOGER-coated biomaterial scaffolds enhanced bone repair through α2β1-dependent engagement, confirming that the efficacy of this motif depends not on chemical presence alone but on its presentation state and ligand density at the material interface.23 In this context, sequence is not simply a biochemical code; it determines how regenerative information can be structurally organized and interpreted by cells.
For bone healing in particular, mineral-interactive motifs form a critical bridge between organic sequence design and inorganic tissue formation. Phosphoserine-containing peptide amphiphiles established that peptide nanofiber surfaces can directly nucleate hydroxyapatite and enhance regeneration in vivo: when self-assembled into nanofiber gels and implanted into a 5-mm rat femoral critical-size defect, these phosphorylated nanofiber matrices produced significantly greater bone formation than non-phosphorylated controls and achieved regeneration comparable to that of a clinically used allogenic bone matrix.6,21 A complementary mineralization-directing strategy was demonstrated by a DPP-mimetic peptide nanofiber system, in which ALP-mediated dephosphorylation triggered self-assembly and calcium ions further promoted nanofiber network formation in vitro, generating a mineralized ECM-like material capable of facilitating apatite-like deposition—a result that established the principle that mineralization guidance can be programmed at the sequence level and activated through nanoscale self-assembly.45 Similar principles apply to acidic and mineral-binding motifs more broadly: in a rat cranial defect model, a biomimetic self-assembling acidic peptide delivered as a hydrogel promoted bone regeneration, supporting the view that such motifs can define where and how mineral deposition is initiated within the developing repair matrix.41 Higher-order variables such as chirality and multifunctional fusion further extend this design space by tuning supramolecular packing, degradation kinetics, and signal persistence.46 Collectively, these studies show that sequence defines a peptide’s regenerative potential—its capacity for receptor engagement, mineral interaction, interfacial binding, self-assembly, or multifunctionality. Yet sequence alone does not determine therapeutic performance.39,47,48
Coupling Modes That Amplify Function
If sequence defines biological potential, nanoengineering determines how that potential is experienced by cells and tissues. This is the level at which a peptide becomes more than a bioactive molecule and starts to function as a regenerative material signal. Self-assembly is the most direct example. Peptide amphiphiles can organize into nanofibrous architectures that improve local retention, concentrate bioactive motifs, and transform otherwise inert substrates into instructive interfaces, as shown in porous titanium systems.9 Co-assembly extends this principle by integrating endogenous biological components into the material itself; peptide amphiphile–blood gels, for example, embed peptide function within a living, growth-factor-rich provisional matrix that supports bone repair.16,49
Nanoengineering is equally important for spatially confined ligand presentation. Surface immobilization and covalent tethering allow peptide density, orientation, and accessibility to be controlled at cell-relevant length scales. This is especially important for receptor-specific ligands such as GFOGER, whose efficacy depends not simply on chemical presence but on nanoscale display.23,50 In such systems, ligand geometry becomes part of the biological message: a peptide that is poorly exposed or weakly clustered may be chemically intact yet biologically ineffective.29,51
A third function of nanoengineering is temporal control. Bone healing is stage-dependent, and peptide cues are rarely most effective when delivered continuously in free form. Responsive matrices and degradable linkers can instead couple peptide exposure to cell invasion, inflammatory protease activity, or tissue remodeling. Foundational poly(ethylene glycol) (PEG)-based synthetic extracellular matrices demonstrated how protease-sensitive peptide linkers can support cell-ingrowth-dependent bone regeneration.52 More recent systems have extended this logic to inflammatory or osteoclast-associated enzyme responsiveness, allowing peptide activity to be revealed only when the defect microenvironment is prepared to use it.53 These considerations—where peptide cues are displayed, when they become available, and in what structural form cells encounter them—form the basis of the design logic discussed in the following section.54,55
A Design Logic for Mechanism-Guided Bone Repair
This framework suggests a simple but useful design rule: regenerative outcome is determined by the interaction between sequence-encoded potential, nanoengineered presentation, and defect-stage biology. Sequence specifies what a peptide is capable of doing; nanoengineering determines whether that capability is protected, clustered, localized, or temporally revealed; and the local defect environment determines whether the resulting signal is meaningful at that phase of repair. This is why free peptides and nanoengineered peptides cannot be treated as functionally equivalent formulations of the same cue.
Accordingly, bone-healing peptides should not be classified only by motif type or scaffold platform. A more informative framework is to organize them by the biological bottlenecks they are designed to overcome and by the nanoscale presentation strategies that make them effective in that setting.27 This perspective underpins the following sections, in which nanoengineered peptide systems are discussed according to their mechanistic roles in immunomodulation, angiogenic–osteogenic coupling, matrix mineralization, and endogenous-cell recruitment30 (Table 2).
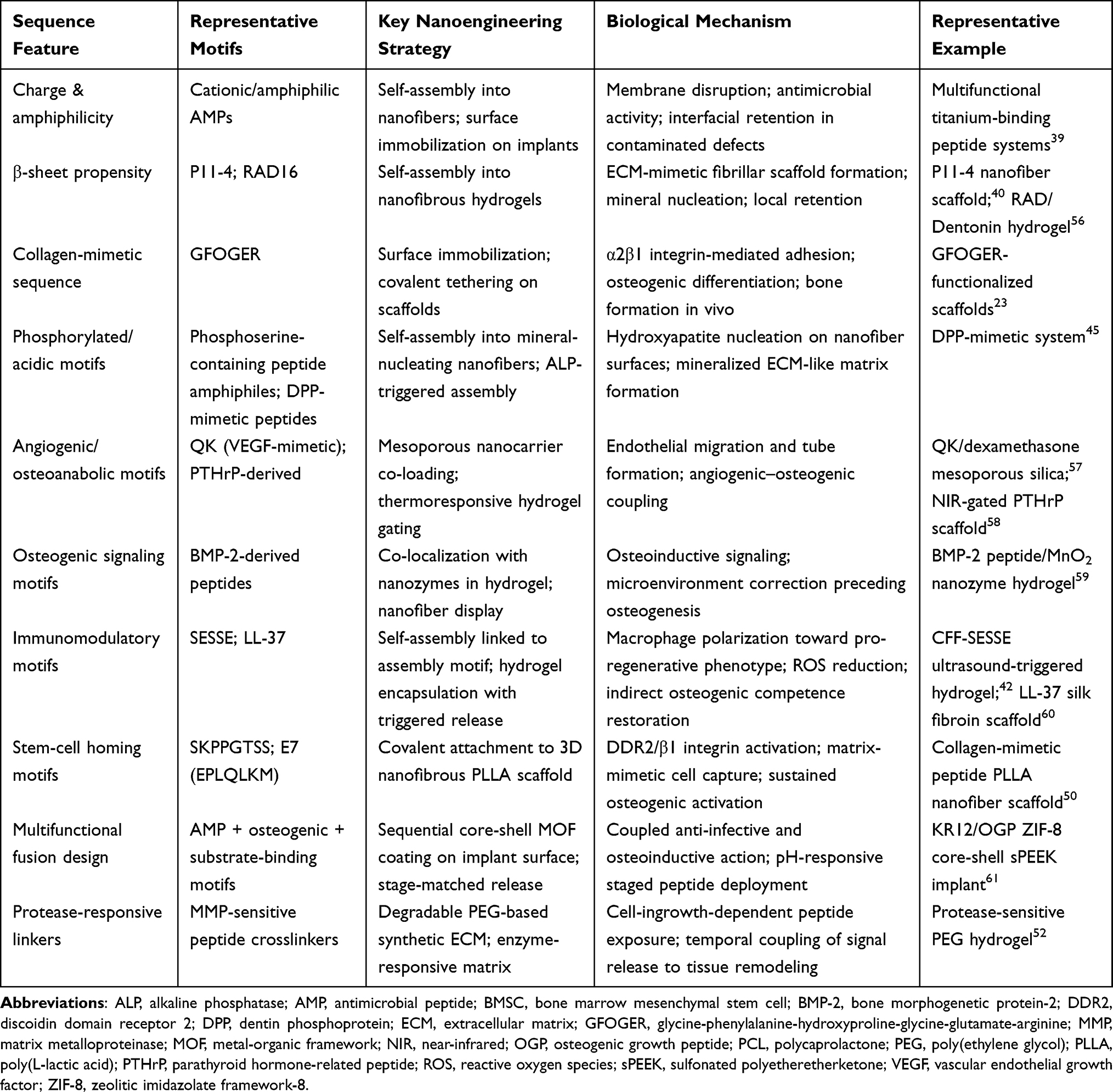 |
Table 2 Summary of Peptide Sequence Features, Nanoengineering Strategies, Biological Mechanisms, and Representative Examples Within the Conceptual Framework for Bone Healing |
Mechanism-Guided Regenerative Functions
Bone-healing nanoengineered peptides are most informative when discussed according to the biological bottlenecks they overcome rather than the platforms into which they are incorporated.39 In this context, nanoengineered peptides should be understood as sequence-programmed instructive modules whose effects emerge only after they are spatially clustered, temporally revealed or dynamically released within the defect niche.
Mechanism I – Immuno-Osteogenesis
Immuno-osteogenesis is the first gatekeeper of successful bone healing.62 Immediately after injury or implantation, the regenerative trajectory is strongly conditioned not simply by the presence of inflammation, but by its quality, magnitude and duration.63 A transient early pro-inflammatory response is physiologically necessary for debris clearance, antimicrobial defence and the recruitment of reparative cells. However, macrophage responses in bone repair do not follow a simple binary switch between discrete pro-inflammatory and anti-inflammatory states; rather, they exist along a continuum of activation phenotypes shaped by evolving local signals including damage-associated molecular patterns, cytokines, matrix cues, and microbial stimuli.64 When the early pro-inflammatory activation program—characterized by sustained ROS production, elevated interleukin-1β (IL-1β)/tumor necrosis factor-α (TNF-α) secretion, and persistent nuclear factor kappa B (NF-κB) signaling—fails to resolve in a timely manner, it suppresses the osteogenic differentiation of stromal cells, impair angiogenic progression, and favour fibrotic encapsulation rather than osseous integration.65 In this setting, the key therapeutic bottleneck is therefore not simply insufficient osteogenic signalling, but the failure to transition from an injury-responsive inflammatory program to a pro-resolving, matrix-remodeling-permissive immune state.66
Peptides are attractive in this setting because they can be designed to act at precisely this early bottleneck: they can modulate macrophage behaviour, reduce oxidative and inflammatory stress, and indirectly restore the osteogenic competence of the local niche.67,68 Yet, in their free or conventionally delivered form, immunomodulatory peptides often fail to do so effectively in vivo.69 Their major limitations are rapid enzymatic degradation, poor local retention, burst-like loss from the defect, and insufficient spatiotemporal control over when and where immune cells encounter the signal. These shortcomings are particularly consequential in immuno-osteogenesis, where therapeutic efficacy depends on acting within a narrow early inflammatory window and maintaining sufficient local exposure to reprogram macrophage fate before maladaptive inflammation becomes self-sustaining.24 Nanoengineering addresses these barriers by converting peptide motifs into retained, clustered, and stage-controllable nanoscale signals that can persist within the defect, interact more effectively with immune cells, and be released or exposed in a temporally relevant manner.
A representative example that directly addresses what nanoengineering contributes—rather than simply what a peptide can do—is the ultrasound-triggered biomimetic ultrashort peptide nanofiber hydrogel reported by Zhang et a.42 The central nanoengineering question this study addresses is whether converting a rapidly degradable immunomodulatory peptide (SESSE) into a supramolecular nanostructure changes its biological outcome, and if so, through which physical mechanism. By linking SESSE to a self-assembly motif to generate CFF-SESSE—which forms crosslinked nanofiber networks through hydrophobic interactions and aromatic stacking—the authors transformed a short soluble peptide into a nanostructure with measurably improved stability and local persistence. This step is not trivial: without self-assembly, SESSE would be expected to diffuse rapidly and undergo enzymatic degradation before achieving sustained macrophage engagement at the defect site. Incorporation into a calcium alginate hydrogel further added a second layer of nanoengineering control—stimulus-responsive release—by gating nanofiber deployment to ultrasound activation rather than allowing passive burst exposure.70 Critically, the study provides evidence that this physical presentation state, rather than peptide sequence alone, determined macrophage accessibility: the released nanofibers preferentially interacted with macrophages and shifted their activation toward a pro-resolving state through the signal transducer and activator of transcription 6 (STAT6)/peroxisome proliferator-activated receptor gamma (PPAR-γ)/suppressor of cytokine signaling 3 (SOCS3) axis, increasing macrophage-derived bone morphogenetic protein-2 (BMP-2) and insulin-like growth factor-I (IGF-I) and thereby indirectly restoring osteogenic competence. What this study therefore proves about nanoengineering is that local retention and temporally controlled release are not merely pharmacokinetic conveniences—they are prerequisites for converting an immunomodulatory peptide sequence into a defect-relevant biological signal (Figure 1A).
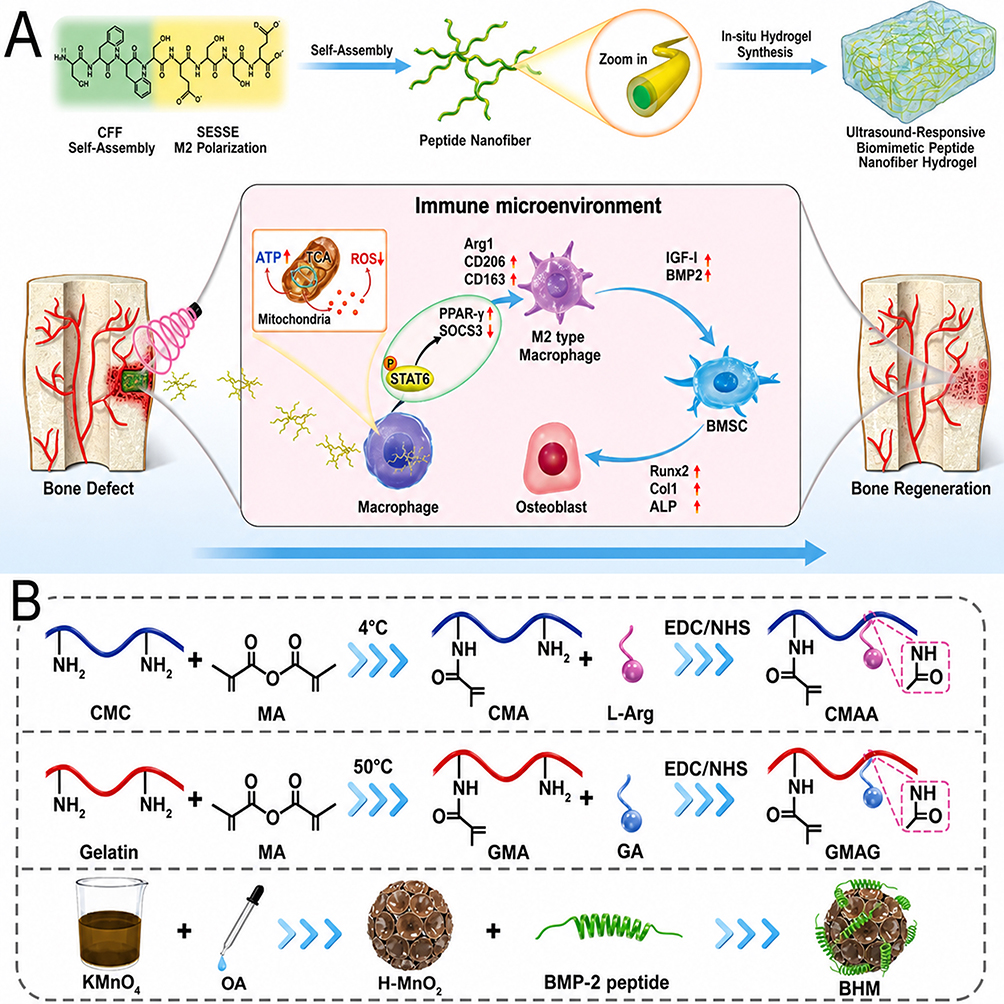 |
Figure 1 Nanoengineered immunomodulatory peptides for immuno-osteogenesis. (A) Schematic illustration of an ultrasound-triggered ultrashort peptide nanofiber hydrogel for immunomodulation-enhanced bone regeneration. The ultrashort peptide was modularly designed to self-assemble into nanofibers and further incorporated into a calcium alginate hydrogel for defect filling and stimulus-responsive release. Upon ultrasound stimulation, the released nanofibers reduced macrophage oxidative stress, promoted M2 polarization through the STAT6/PPAR-γ/SOCS3 axis, and enhanced macrophage paracrine secretion of BMP-2 and IGF-I, thereby facilitating the osteogenic differentiation of bone marrow mesenchymal stem cells (BMSCs). Adapted with permission from Ref.42 (B) Schematic of a multifunctional hybrid hydrogel (BHM@CG) integrating a BMP-2-derived peptide-coupled nanozyme within an L-Arg-modified CMA/GA-grafted GMA hydrogel matrix. This nanoengineered system combines ROS scavenging, inflammatory microenvironment regulation, and osteogenic signaling to promote bone repair. Adapted with permission from Ref.59 Abbreviations: BMSCs, bone marrow mesenchymal stem cells; STAT6, signal transducer and activator of transcription 6; PPAR-γ, peroxisome proliferator-activated receptor gamma; SOCS3, suppressor of cytokine signaling 3; BMP-2, bone morphogenetic protein-2; IGF-I, insulin-like growth factor-I; ROS, reactive oxygen species; L-Arg, L-arginine; CMA, catechol-modified alginate; GA, gallic acid; GMA, glycidyl methacrylate. |
A second example provides complementary but distinct evidence about what nanoengineering contributes in the context of ROS-dominated, inflammation-impaired defects.59 The core nanoengineering insight here is not about release timing or retention per se, but about defect-specific responsiveness and spatial co-localization: when a BMP-2-derived peptide is delivered alone into a niche dominated by excessive ROS and unresolved inflammation, its osteoinductive potential is largely negated by the hostile microenvironment before receptor-level engagement can occur.71 This study addresses that problem not by modifying the peptide sequence, but by co-localizing it with a honeycomb manganese dioxide (MnO2) nanozyme within a photoenhanced hybrid hydrogel, thereby creating a nanoscale division-of-labour architecture. Crucially, the nanozyme component does not simply serve as a co-therapeutic additive; it functions as a prerequisite gate that must first correct the microenvironment—depleting ROS, suppressing inflammatory amplification, and restoring stromal cell viability—before the peptide can operate within a niche that is biologically receptive to osteogenic signaling. This sequential logic could not be achieved by mixing the same components in free solution, where spatial co-localization and staged action would be lost. What this study therefore proves about nanoengineering is that defect-specific context matching—the ability to physically organize multiple therapeutic functions so that microenvironment correction precedes and enables downstream peptide signaling—is a property that emerges from nanoarchitecture rather than from sequence design alone. Together, these two studies establish that nanoengineering contributes at least two mechanistically distinct and non-redundant functions to immuno-osteogenic peptide therapy: controlled local retention and temporal deployment, and spatially organized, defect-responsive cascade regulation59 (Figure 1B).
Despite these advances, several limitations remain. First, many studies still rely heavily on macrophage polarization markers to infer immune reprogramming, whereas the causal linkage between peptide-enabled immunomodulation and downstream improvements in angiogenesis, matrix deposition, and mechanically relevant bone repair is not always rigorously established.64 Second, the therapeutic window is narrow: excessive or prolonged suppression of early inflammation may also compromise host defence and impair initial recruitment events.24 Finally, the field still lacks a clear understanding of which nanoengineering parameters—such as nanofiber architecture, ligand density, release kinetics, or degradation behaviour—most strongly determine immune-stage reprogramming in vivo.72 Future studies should therefore define these structure–function relationships more rigorously and test whether immunomodulatory peptide nanoarchitectures remain effective under more clinically demanding conditions.
Mechanism II – Angiogenic-Osteogenic Coupling
Bone regeneration is often described as a sequence in which angiogenesis precedes osteogenesis, yet accumulating evidence suggests that this view is too linear.33 In compromised defects, vascularization is not simply a parallel outcome that accompanies new bone formation; rather, it is a rate-limiting prerequisite that determines whether osteogenesis can proceed efficiently at all.73 When vascular invasion is delayed or inadequate, the defect remains poorly perfused and hypoxic, progenitor recruitment is weakened, endothelial–stromal crosstalk is diminished, and subsequent matrix deposition becomes disorganized or incomplete.74 The core pathological bottleneck is therefore not merely a shortage of osteogenic cues, but the failure to establish a vascularly permissive microenvironment capable of supporting cell ingress, nutrient delivery, and coordinated bone formation.
Peptides are well suited to this bottleneck because they can activate endothelial migration, tube formation, stromal–endothelial communication, and downstream osteogenic signaling. Angiogenic peptides such as vascular endothelial growth factor (VEGF)-mimetic motifs or QK-like sequences, as well as osteoanabolic peptides with vascular effects, offer a chemically defined way to promote vascular priming without relying exclusively on full-length growth factors. However, when delivered in free or conventional form, these peptide cues often show rapid diffusion, short local residence, weak gradient formation, and poor temporal coordination with subsequent osteogenesis. As a result, even biologically relevant angiogenic peptides may fail to generate sustained vascular support within the defect. Nanoengineering overcomes these limitations by organizing peptides into nanostructures that retain them locally, cluster them at cell-interactive interfaces, co-localize them with osteogenic signals, or release them according to the evolving biology of repair. In this mechanism, the role of nanoengineering is not simply to prolong peptide exposure, but to transform angiogenic stimulation into a spatiotemporally structured program that can support later osteogenesis.75
This principle is illustrated by the peptide amphiphile nanofiber scaffold reported by Bakshi et al (2021), in which self-assembled peptide nanostructures were used to control VEGF presentation and generate a chemotactic microenvironment that enhanced stem-cell recruitment and angiogenesis before facilitating BMP-2-mediated bone regeneration76 (Figure 2A–D). The key advantage of nanoengineering in this case was that the angiogenic signal was not released as a transient soluble cue, but displayed within a nanofibrous assembly capable of sustaining local vascular priming. A related strategy was described by Sun et al (2019), who used biodegradable mesoporous silica nanocarriers to spatially couple the angiogenic QK peptide with dexamethasone, thereby integrating endothelial activation and osteogenic stimulation within a single nanoscale platform57 (Figure 2E–G). Although the engineering formats differed, both studies support the same design logic: peptide-driven vascular priming becomes more effective when angiogenic signals are stabilized, localized, and coordinated with subsequent osteogenic events.
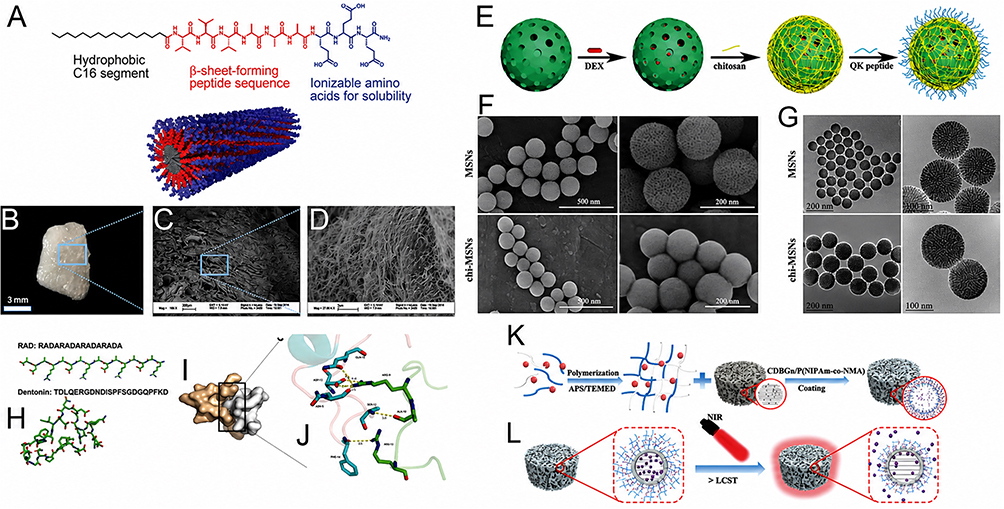 |
Figure 2 Nanoengineered peptide systems for angiogenic–osteogenic coupling. (A) Chemical structure of the peptide amphiphile. (B) Molecular representation of peptide co-assembly. (C and D) Microscopic and scanning electron microscopy (SEM) images of peptide amphiphile nanofiber-incorporated collagen sponges. (A–D) adapted with permission from Ref.76 (E) Schematic illustration of the preparation of QK/DEX@chi-MSNs. (F and G) SEM and transmission electron microscopy (TEM) images of MSNs and chi-MSNs at different magnifications. (E–G) adapted with permission from Ref.57 (H) Initial modeled structures of RAD and Dentonin peptides. (I) Composite model of the RAD/Dentonin peptide system. (J) Hydrogen-bonding network of the RAD/Dentonin peptide complex, with hydrogen bonds indicated by dashed lines. (H-J) adapted with permission from Ref.58 (K) Schematic diagram of the synthesis and mechanism of CBP/MBGS/PTHrP-2 bioscaffolds for the coordinated regulation of angiogenesis and osteogenesis. (L) Fluorescence image of CDBGn aqueous suspension under 365 nm UV irradiation, showing bright blue emission centered at 485 nm upon excitation at 380 nm. (K and J) adapted with permission from Ref.56 Abbreviations: SEM, scanning electron microscopy; TEM, transmission electron microscopy; QK, VEGF-mimetic peptide QK; DEX, dexamethasone; chi-MSNs, chitosan-modified mesoporous silica nanoparticles; MSNs, mesoporous silica nanoparticles; RAD, arginine–alanine–aspartic acid-based self-assembling peptide; CBP, collagen-binding peptide; MBGS, mesoporous bioactive glass scaffold; PTHrP-2, parathyroid hormone-related peptide-2; CDBGn, carbon dot-functionalized bioactive glass nanoparticles; UV, ultraviolet; VEGF, vascular endothelial growth factor. |
An even more explicit spatiotemporal example was provided by Liu et al (2023), who developed a near-infrared (NIR)-activable hydrogel-coated mesoporous bioactive glass scaffold for dual-mode delivery of the parathyroid hormone (PTH)-related peptide (PTHrP).58 In this system, PTHrP-2 was first loaded into the hierarchical mesopores of mesoporous bioactive glass and then gated by a thermoresponsive photothermal hydrogel, converting a bioactive peptide into a nanostructurally regulated release system. Under on/off NIR irradiation, the hydrogel underwent reversible phase transition, enabling both pulsatile on-demand release and prolonged slow release of the peptide. This design improved human umbilical vein endothelial cells (HUVEC) proliferation, migration, and tube formation in vitro, promoted cluster of differentiation 31 (CD31)/a α-smooth muscle actin (α-SMA)-positive vessel formation in vivo, and enhanced repair of critical-sized femoral defects. Importantly, the advantage of nanoengineering here was not merely prolonged retention, but the imposition of a stage-adaptable delivery profile more compatible with the biological dynamics of PTH-family signaling58 (Figure 2H–J).
A further step beyond nanocarrier-mediated delivery was reported by Liu et al (2024), who nanostructured the peptide system itself rather than using an external carrier.56 In their RAD/Dentonin hydrogel, the functional peptide Dentonin was incorporated into a self-assembling RAD matrix that formed highly ordered β-sheet supramolecular nanofiber networks through non-covalent interactions. This design is particularly relevant because both the structural matrix and the therapeutic function were peptide-derived. Mechanistically, the RAD/Dentonin scaffold supported BMSCs adhesion, proliferation, and three-dimensional migration, promoted osteogenic differentiation through activation of Wnt/β-catenin signaling, and simultaneously enhanced BMSCs-mediated endothelial activation, including HUVEC migration, tube formation, and angiogenic gene expression. Unlike systems that simply co-deliver separate angiogenic and osteogenic additives, this self-assembled nanofibrous peptide scaffold generated coupling from within the peptide microenvironment itself, allowing vascular and osteogenic programs to reinforce each other through matrix-guided cell crosstalk56 (Figure 2K and L).
Taken together, these studies show that peptide nanoengineering can support angiogenic–osteogenic coupling through several convergent routes: self-assembled nanofibers can sustain local vascular priming, mesoporous nanocarriers can co-localize angiogenic and osteogenic cues, gated nanoreservoirs can impose stage-matched release dynamics, and peptide-only supramolecular matrices can generate vascularized osteogenic niches intrinsically. The defining feature of successful angiogenic–osteogenic coupling in these systems is therefore temporal coordination: nanoengineering does not simply sustain vascular signals, but organizes their delivery relative to the osteogenic program so that the two processes reinforce rather than compete with each other.
Despite strong proof-of-concept data, several limitations remain. First, many studies still evaluate success primarily through endothelial migration, tube formation, or vessel density markers, whereas the more clinically relevant question is whether peptide nanoarchitectures generate functional vascular networks with adequate perfusion, maturity, and coupling to bone formation.73 Second, the optimal temporal relationship between angiogenic stimulation and osteogenesis is still incompletely defined; premature, excessive, or prolonged vascular signaling may not translate into better bone repair.77 Finally, the field still lacks a quantitative understanding of which nanoengineering parameters—such as ligand density, release kinetics, matrix porosity, and co-presentation with osteogenic cues—most strongly determine productive angiogenic–osteogenic coupling in vivo.78 Future studies should therefore move beyond “more vessels plus more bone” as a descriptive endpoint and instead determine how nanoengineered peptide systems can generate stable, perfused, and osteogenesis-supportive vascular niches under clinically demanding conditions.
Mechanism III – Mineralization Guidance and Matrix-Instructive Bone Formation
Mineralization guidance and matrix-instructive bone formation represent a third major mechanistic bottleneck in bone repair.79 It is important to distinguish at the outset between mineral deposition and functional bone regeneration: the former refers to the nucleation and growth of calcium phosphate mineral phases within the repair matrix, whereas the latter requires that this mineral be hierarchically organized, mechanically integrated with the collagen scaffold, remodelable by osteoclasts, and progressively replaced by load-bearing lamellar bone.80 Calcium phosphate deposition that is ectopic, disordered, or poorly coupled to collagen fibril organization does not necessarily translate into mechanically competent or biologically functional bone tissue.80 With this distinction in mind, a critical bottleneck in many difficult defects is not only the absence of osteogenic stimulation, but the inability to establish an extracellular environment that is competent to guide where, when, and in what structural form mineral deposition occurs.81,82 Nanoengineered peptides are particularly relevant to this bottleneck because they can potentially recreate the physicochemical logic of native mineralizing proteins—not simply to promote calcium phosphate deposition, but to template its spatial organization and coupling to the organic matrix in a manner that supports progression toward functional bone architecture.
Peptides are particularly attractive for this bottleneck because they can be engineered not only to stimulate cells, but also to mimic the mineral-directing functions of native ECM proteins. Sequence motifs such as phosphoserine, acidic residues, calcium-binding domains, or dentin phosphoprotein-inspired segments can provide ion-binding capacity, nucleation potential, and matrix-mimetic recognition. However, in free or conventionally delivered form, these peptide functions are often poorly realized. Soluble mineral-binding peptides may diffuse away from the target site, lose spatial organization, and fail to maintain the local density and orientation of functional groups required for effective mineral templating.83 This is especially important because mineral nucleation is inherently an interfacial nanoscale process: sequence chemistry alone is insufficient unless it is translated into a stable architecture that can present ion-binding motifs in an ordered and persistent manner. Nanoengineering overcomes this limitation by converting mineral-interactive peptides into nanostructured templates—such as self-assembled nanofibers, electrospun hybrid matrices, or mineralized interfaces—that can guide apatite formation, localize osteoinductive signals, and organize matrix maturation in a spatially instructive way.84
A strong mechanistic foundation for this concept is provided by Gulseren et al (2019), who developed a dentin phosphoprotein (DPP)-mimetic peptide nanofiber system to recapitulate the mineral-directing role of phosphorylated non-collagenous matrix proteins.45 In this study, peptide molecules self-assembled through ALP-mediated dephosphorylation, while calcium ions further promoted nanofiber network formation, ultimately generating a mineralized extracellular-matrix-like material capable of facilitating apatite-like deposition. The significance of this work lies in showing that mineralization guidance can be programmed at the sequence level and activated through nanoscale self-assembly. An in vivo extension of the same principle had been demonstrated earlier by Mata et al (2010), who engineered self-assembling peptide amphiphile nanofiber gels containing phosphoserine residues that nucleated hydroxyapatite on nanofiber surfaces.6 When implanted into a 5-mm rat femoral critical-size defect, these phosphorylated nanofiber matrices produced significantly greater bone formation than non-phosphorylated controls and achieved regeneration comparable to that of a clinically used allogenic bone matrix. Together, these studies establish that mineral nucleation can be actively initiated by nanoengineered peptide architectures that recreate the physicochemical logic of native mineralizing proteins. However, it should be noted that neither study provides direct evidence that the resulting mineral phase is hierarchically organized or mechanically equivalent to native lamellar bone. The in vitro DPP-mimetic system demonstrated apatite-like deposition competence,45 and the in vivo phosphoserine nanofiber study showed greater bone area by histology compared with controls,6 but the mechanical properties, collagen–mineral coupling, and long-term remodeling behavior of the regenerated tissue were not characterized. These remain important open questions for evaluating whether peptide-guided nucleation translates into structurally and functionally competent bone.
This sequence-to-nanostructure-to-mineralization paradigm was further advanced by de Souza Araújo et al (2022), who translated the self-assembling mineralization peptide P11-4 into a hybrid fibrous scaffold for guided mineralized tissue regeneration.40 In this case, P11-4 was physically adsorbed onto electrospun polycaprolactone nanofibers, generating a peptide-functionalized scaffold with an approximately 500-nm fiber diameter that preserved nanoscale fibrillar architecture while adding mineralization functionality. Mechanistically, the electrospun matrix was not simply a passive carrier; it served as an ECM-like support that stabilized peptide presentation and prolonged its mineralization-competent state. This hybrid design promoted mineral precipitation in simulated body fluid, supported apatite-like formation and crystallization, enhanced cell-mediated mineral deposition, and upregulated mineralization-related gene expression compared with pristine scaffolds. In a rat calvarial critical-size defect model, it also improved bone regeneration, with histology showing osseous islands containing blood vessels in the higher-concentration peptide group. Here again, nanoengineering was essential because it transformed an intrinsically mineralization-competent sequence into a structurally stable and biologically instructive nanofibrous matrix rather than a diffusible peptide additive. Nevertheless, the evidence for improved bone regeneration in this study rested primarily on mineral precipitation in simulated body fluid, mineralization-related gene expression, and histological observation of osseous islands.40 Whether the regenerated tissue possessed appropriate mechanical integrity, collagen–mineral co-organization, or remodeling capacity was not directly assessed, underscoring the need to move beyond mineral deposition metrics when evaluating the functional outcomes of peptide-guided mineralization strategies.
Overall, the convergent design logic is clear: mineral-guiding peptide motifs become therapeutically effective only when nanoengineering preserves them in a local, structured, and interfacially active state. However, participation in the construction of the mineralizing niche is a necessary but not sufficient condition for functional bone regeneration. The ultimate goal is not to maximize calcium phosphate deposition, but to guide its spatial organization, collagen coupling, and progressive maturation into hierarchically structured, mechanically load-bearing, and remodelable bone tissue. Current nanoengineered peptide systems have demonstrated compelling control over mineral nucleation and early matrix organization, but whether this translates into biomechanically competent bone remains underexplored. Future studies should therefore incorporate mechanical testing, polarized light microscopy or small-angle X-ray scattering for collagen–mineral organization, and long-term remodeling assessments alongside conventional histological and radiographic endpoints.
Despite these advances, several limitations remain. First, and most fundamentally, the field has not yet established a clear mechanistic link between peptide-guided mineral nucleation and the emergence of hierarchically organized, mechanically competent bone. Most studies evaluate success through mineral deposition assays, alkaline phosphatase activity, osteocalcin/osteopontin expression, or micro-CT bone volume measurements—endpoints that capture the quantity of mineralized tissue but not its structural quality, collagen–mineral co-organization, stiffness, toughness, or susceptibility to remodeling.80,85 This distinction matters clinically: disorganized or ectopic calcium phosphate may occupy a defect radiographically while providing little load-bearing function, and may even impair normal remodeling by resisting osteoclast resorption.86 Future studies should therefore adopt a tiered outcome framework that distinguishes mineral deposition, mineral organization, mechanical competence, and remodeling capacity as separate endpoints requiring separate validation. Third, the optimal nanoarchitectural parameters for effective mineral guidance—including nanofiber diameter, charge density, phosphate spacing, degradation rate, and dynamic assembly behaviour—remain insufficiently defined.86 Future work should therefore move beyond “more mineral” as an endpoint and determine whether peptide nanoarchitectures can reproducibly generate hierarchically organized, remodelable, and mechanically functional bone matrix under clinically demanding conditions.
Mechanism IV – Endogenous Cell Participation: From Surface Capture to Host-Driven Regeneration
Cell recruitment and endogenous regeneration represent a distinct mechanistic bottleneck in bone repair.87 Rather than relying primarily on exogenous cell transplantation, this strategy depends on the ability to attract host stem/progenitor cells to the defect, retain them within a permissive matrix, and subsequently direct their osteogenic participation through localized biochemical and biophysical cues. In many compromised defects, however, this sequence fails at an early stage: too few reparative cells enter the lesion, recruited cells are not retained long enough to participate meaningfully, or the local matrix does not provide the receptor-level and microenvironmental signals required for survival and lineage progression. The pathological problem is therefore not simply insufficient osteoinduction, but the inability to convert host regenerative potential into a spatially organized and biologically productive endogenous repair response.88
Peptides are particularly well suited to this bottleneck because they can function as stem-cell-homing motifs, affinity ligands, or extracellular-matrix-mimetic cues that selectively recruit, capture, and activate endogenous reparative cells. Short motifs such as SKPPGTSS or E7 can increase the likelihood of host BMSCs accumulation, whereas receptor-targeting or collagen-mimetic peptides can provide a more instructive interface for sustained engagement and downstream differentiation. Yet, in free or conventionally delivered form, these peptides often face substantial limitations: soluble homing signals are rapidly diluted, chemotactic gradients dissipate quickly, local retention is weak, and monovalent peptide presentation may be insufficient to support stable cell capture or receptor activation. Nanoengineering addresses these shortcomings by displaying such motifs at high local density on hydrogels, nanofibers, or implant surfaces, thereby converting simple affinity sequences into localized, multivalent, and matrix-integrated recruitment signals. In this context, the value of nanoengineering lies not only in improving peptide persistence, but in coupling recruitment with retention and niche conditioning so that incoming cells can subsequently contribute to tissue reconstruction.56
A representative example is the biomimetic hydrogel reported by Wei et al (2024) for osteoporotic bone defects, in which stem-cell homing was explicitly integrated with nanostructure-enabled microenvironment regulation89 (Figure 3A). In this system, the hydrogel acted as a reservoir for S-nitrosoglutathione and Ca2⁺, activated the nitric oxide (NO)/cyclic guanosine monophosphate (cGMP) pathway, and simultaneously recruited BMSCs through conjugation of the stem-cell-homing peptide SKPPGTSS. The presence of mesoporous silica nanoparticles (MSNs) and an MSNs@CM intermediate within the SA-MSNs@CM-Stiff platform further established the nanoscale basis of the construct. Mechanistically, recruitment was positioned as the initiating event of repair: the peptide increased the probability that host BMSCs would enter the defect niche, while the nanoengineered hydrogel environment then supported osteogenic and angiogenic coupling as well as local immunoregulation. In a rat distal femoral defect model, the system increased both new bone and new vessel formation, illustrating that peptide-guided homing becomes more effective when integrated into a nanostructured microenvironment rather than delivered as a transient soluble signal.
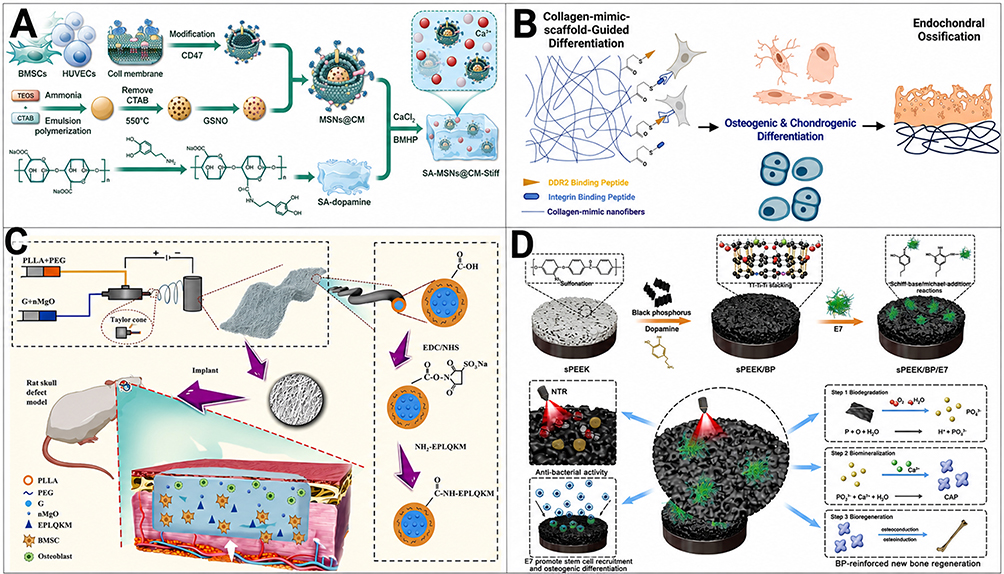 |
Figure 3 Nanoengineered peptide platforms for endogenous cell recruitment and regeneration. (A) Schematic illustration of the preparation of membrane-biomimetic MSNs@CM. Mesoporous silica nanoparticles (MSNs) were first synthesized and loaded with GSNO, followed by coating with cell membrane (CM) derived from ultracentrifuged BMSCs and HUVECs to obtain MSNs@CM. (A) adapted with permission from Ref.89 (B) Schematic of a PLLA-based nanofibrous scaffold enabling controllable peptide conjugation while preserving the nanofiber-forming properties of PLLA. Peptide-decorated nanofibrous scaffolds were evaluated in a critical-sized bone defect model to assess the individual and synergistic regenerative effects of GFOGER and GVMGFO peptides. (B) adapted with permission from Ref.50 (C) Fabrication of coaxial fiber membranes functionalized with nano-MgO (nMgO) and the BMSC-affinitive peptide E7 via coaxial electrospinning and subsequent EDC/NHS-mediated grafting, followed by implantation into a rat calvarial defect model. (C) adapted with permission from Ref.90 (D) Schematic illustration of the multifunctional sPEEK/BP/E7 implant surface, fabricated by integrating black phosphorus (BP) nanosheets and the E7 peptide onto sulfonated PEEK to endow the implant with osteogenic and antibacterial properties. (D) adapted with permission from Ref.91 Abbreviations: MSNs, mesoporous silica nanoparticles; CM, cell membrane; GSNO, S-nitrosoglutathione; BMSCs, bone marrow mesenchymal stem cells; HUVECs, human umbilical vein endothelial cells; PLLA, poly(L-lactic acid); GFOGER, Gly-Phe-Hyp-Gly-Glu-Arg; GVMGFO, Gly-Val-Met-Gly-Phe-Hyp; nMgO, nano-magnesium oxide; EDC, 1-ethyl-3-(3-dimethylaminopropyl)carbodiimide; NHS, N-hydroxysuccinimide; sPEEK, sulfonated polyetheretherketone; BP, black phosphorus; PEEK, polyetheretherketone. |
A complementary but conceptually deeper strategy was presented by Zhou et al (2025), who showed that endogenous regeneration depends not only on chemotaxis, but also on selective capture and receptor-level activation of host reparative cells50 (Figure 3B). Their scaffold consisted of poly(L-lactic acid) (PLLA)-based nanofibers with covalently attached synthetic helical peptides designed to mimic native collagen motifs and activate discoidin domain receptor 2 (DDR2) and β1 integrins, two major collagen-responsive receptors in bone cells. Here, the decisive factor was the nanoscale presentation state of the ligand: because the peptides were displayed on an interconnected three-dimensional nanofibrous scaffold rather than released in soluble form, the system could mimic extracellular-matrix signaling at both molecular and nanometre scales. This peptide-decorated scaffold produced 7.8-fold greater mineralized bone regeneration than peptide-free controls after 8 weeks in a critical-sized defect model, supporting the view that nanoengineered peptide presentation can liberate endogenous regenerative potential not merely by attracting cells, but by engaging them through a matrix-like interface that sustains downstream osteogenic activation.
A more explicitly recruitment-oriented design was described by Long et al (2025), who fabricated core–shell coaxial fibre membranes using PLLA/PEG as shell materials and nanomagnesium oxide combined with gelatin as core materials, followed by grafting of the BMSCs-affinitive peptide E7 (EPLQLKM) onto the fibre surface90 (Figure 3C). This system is especially informative because recruitment of endogenous BMSCs was treated as the central therapeutic objective rather than as a secondary benefit. The E7-functionalized membranes improved hydrophilicity and mechanical properties, maintained pH stability in vitro, and significantly enhanced BMSCs proliferation, migration, and osteogenic differentiation. Importantly, the in vivo analysis included early recruitment-related evaluation using cluster of differentiation 29 (CD29) and cluster of differentiation 90 (CD90) staining at days 3 and 7 after implantation, directly linking the nanoengineered peptide interface to host-cell accumulation within the defect. In rat cranial defects, this translated into improved new bone formation. Compared with soluble peptide administration, this study highlights why nanoengineering matters: the coaxial nanofibrous architecture provided both a recruitment substrate and a retention structure, allowing the affinity peptide to function in a sustained and spatially localized manner.
An implant-centered version of endogenous regeneration was recently reported by Wang et al (2025), who constructed a photoresponsive sPEEK/BP/E7 surface through self-assembly of black phosphorus (BP) nanosheets, polydopamine, and the short peptide E7 on sPEEK91 (Figure 3D). This multifunctional nanoscale interface improved cytocompatibility, adhesion, proliferation, alkaline phosphatase (ALP) activity, matrix mineralization, osteogenesis-related gene expression, antibacterial performance, and in vivo osseointegration in a rat femoral infection model under NIR irradiation. Although direct in vivo evidence for stem-cell homing at the implant interface remains limited, the incorporation of E7 into this nanoscale surface suggests that endogenous regeneration may also be initiated at implants when peptide presentation is combined with infection control and interfacial osteogenic support. This possibility is mechanistically important because it broadens the concept of recruitment-competent peptide nanoengineering beyond defect-filling scaffolds to include osseointegrative implant surfaces.
Taken together, these studies show that peptide nanoengineering can promote endogenous regeneration through several convergent routes: homing-peptide-functionalized hydrogels can recruit host cells while conditioning the local niche; nanofiber-displayed receptor ligands can capture and activate reparative cells through matrix-mimetic signaling; and multifunctional implant interfaces may extend recruitment-competent regeneration to the peri-implant environment. The common principle is that peptide value does not lie in sequence alone, but in the nanoscale mode of presentation that allows recruitment, retention, and cell-instructive signaling to occur in the same localized microenvironment.
Despite promising advances, several limitations remain. First, many studies use the term “recruitment” broadly, even when the available evidence more directly supports early adhesion, local accumulation, or enhanced retention rather than true chemotactic homing and long-term functional participation.92 Without lineage tracing, fate mapping, or equivalent strategies, it remains difficult to determine the origin, persistence, and actual reparative contribution of the recruited cells.93 Second, the design rules governing productive endogenous regeneration are still incompletely defined: ligand density, spatial distribution, matrix degradability, and the balance between firm capture and subsequent migration/differentiation likely all matter, but their relative importance in vivo remains unclear.94 Finally, evidence for recruitment-competent implant interfaces remains less mature than that for defect-filling scaffolds. Future studies should therefore define endogenous regeneration more rigorously and determine whether nanoengineered peptide systems can reproducibly recruit, retain, and activate reparative host cells under clinically demanding conditions.
Problem-Oriented Applications in Compromised Bone Healing
Mechanism-guided peptide engineering becomes most clinically relevant when it is aligned with the pathological logic of specific bone-healing failures.95 In compromised defects, the same osteogenic sequence may be insufficient, or even ineffective, if the dominant barrier is persistent infection, osteoporotic niche dysfunction, failed vascular invasion or mechanically unstable nonunion.96 Defect-specific design therefore requires more than selecting a bioactive peptide motif; it requires matching sequence features, nanoengineered presentation and release logic to the dominant pathological bottleneck of each clinical scenario.97 From this perspective, infected defects, osteoporotic fractures and large segmental defects should not be viewed as interchangeable indications for “bone regeneration”, but as distinct regenerative problems that demand different peptide-centred design priorities.
Infected Bone Defects
Infected bone defects represent one of the most challenging scenarios in skeletal repair because treatment failure is driven not only by impaired osteogenesis, but also by persistent bacterial colonization, biofilm formation, inflammatory amplification, and a hostile microenvironment that suppresses vascular and matrix regeneration.98 In this setting, therapeutic peptides are attractive because they can provide antimicrobial, immunomodulatory, or osteogenic functions within a molecularly programmable format.99 However, free peptides are often limited by rapid diffusion, enzymatic degradation, and poor site-specific retention.100 Nanoengineering therefore becomes particularly important in infected defects, as it enables peptides to be integrated into responsive nanostructures that confine antibacterial action to the pathological niche, coordinate infection control with inflammatory correction, and preserve subsequent regenerative potential.55
A representative example of this strategy was reported by Lin et al (2025), who developed hyaluronic acid-based hydrogel microspheres for Staphylococcus aureus-infected skull defects by combining antimicrobial peptides (AMP)-conjugated oxidized hyaluronic acid (AOHA) with ZnO nanoparticles in a multi-responsive system101 (Figure 4A and B). The value of this design lies not simply in loading an AMP into a scaffold, but in making peptide action infection-responsive: AMP release was enhanced under acidic conditions and in the presence of hyaluronidase, both of which are relevant to infected tissue environments. Mechanistically, this enabled the microspheres to reduce bacterial growth and biofilm burden while also supporting BMSCs osteogenic differentiation, angiogenesis-related responses, and anti-inflammatory remodeling in vivo. From a problem-oriented perspective, this study is important because it shows that nanoengineered AMP can be deployed as microenvironment-triggered therapeutics, rather than being constitutively released in a nonselective manner.101
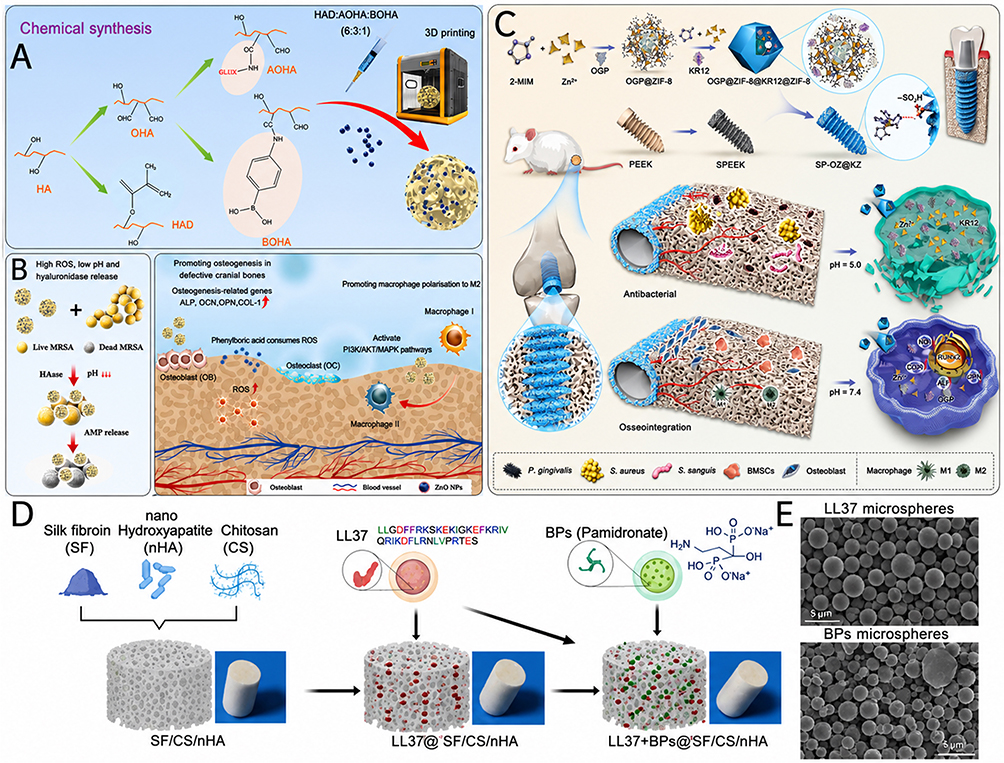 |
Figure 4 Nanoengineered peptide systems for infected bone defects. (A) Schematic illustration of the synthesis procedure of ABOHA@ZnO hydrogel microspheres. (B) Incorporation of ZnO nanoparticles into the hydrogel microspheres to enhance antibacterial activity and promote bone defect repair through broad-spectrum bactericidal effects and activation of multiple signaling pathways. (A and B) adapted with permission from Ref.101 (C) Schematic overview of the pH-responsive implant interface based on sulfonated PEEK and bilayer core–shell ZIF-8 nanostructures for staged delivery of KR12 and osteogenic growth peptide (OGP). Abbreviations: ZIF-8, zeolitic imidazolate framework-8; 2-MIM, 2-methylimidazole; PEEK, polyetheretherketone; sPEEK, sulfonated PEEK; SP-OZ@KZ, sPEEK-OGP@ZIF-8@KR12@ZIF-8; iNOS, inducible nitric oxide synthase; RUNX2, runt-related transcription factor 2; ALP, alkaline phosphatase; OPN, osteopontin; BMSCs, bone marrow mesenchymal stem cells. (C) adapted with permission from Ref.61 (D) Schematic of the fabrication of a silk fibroin (SF)/chitosan (CS)/nano-hydroxyapatite (nHA) composite scaffold modified with LL-37 peptide- and bisphosphonate-functionalized microspheres. (E) Scanning electron microscopy (SEM) images and size distribution histograms of LL-37 and bisphosphonate microspheres. Scale bar: 200 μm. (D and E) adapted with permission from Ref.60 Abbreviations: ABOHA@ZnO, antimicrobial peptide-conjugated oxidized hyaluronic acid hydrogel microspheres containing zinc oxide nanoparticles; ZnO, zinc oxide; PEEK, polyetheretherketone; sPEEK, sulfonated polyetheretherketone; ZIF-8, zeolitic imidazolate framework-8; KR12, KR12 antimicrobial peptide; OGP, osteogenic growth peptide; SF, silk fibroin; CS, chitosan; nHA, nano-hydroxyapatite; LL-37, human cathelicidin antimicrobial peptide; SEM, scanning electron microscopy. |
A more interface-oriented version of the same principle was described by Ma et al (2025) in a pH-responsive sulfonated polyetheretherketone (sPEEK) implant modified with a bilayer core-shell zeolitic imidazolate framework-8 (ZIF-8) mmetal-organic framework (MOF)61 (Figure 4C). In this system, the outer shell carried the AMP KR12, whereas the inner core contained osteogenic growth peptide (OGP), thereby establishing a staged peptide-delivery architecture at the implant surface. This design is especially relevant to infected bone defects because it addresses a common but underappreciated problem: regeneration failure can arise not only within defect-filling scaffolds, but also at the peri-implant interface. Under acidic inflammatory conditions, the core-shell ZIF-8 coating degraded more rapidly and released KR12 and Zn2⁺ to suppress bacterial adhesion and proliferation, while simultaneously promoting macrophage transition toward an anti-inflammatory, pro-resolving; OGP release then supported subsequent peri-implant osseointegration. In rat distal femur models with and without infection, the implant inhibited bacterial growth, reduced inflammation, and improved peri-implant bone formation, with the benefits being particularly evident under infected, lower-pH conditions. Thus, this work illustrates how nanoengineered peptides can be organized into a sequential anti-infective-to-regenerative programme at a clinically relevant implant interface.61
Whereas the first two systems focused on infection-responsive antibacterial control and interface regulation, Ye et al (2024) highlighted another key pathological feature of infected bone defects: infection-associated bone loss and excessive resorption60 (Figure 4D and E). Their 3D scaffold integrated the human cathelicidin peptide LL-37, pamidronate, and nano-hydroxyapatite within a silk fibroin/chitosan-based porous architecture. The significance of this design is that it moved beyond the conventional assumption that bacterial killing alone is sufficient for repair. Instead, the scaffold was engineered to couple broad-spectrum antimicrobial activity from LL-37 with antiresorptive support from pamidronate and structural osteoconductivity from nano-hydroxyapatite. In a murine infectious bone defect model, this co-delivery system reduced inflammation and bacterial load while promoting bone regeneration. Conceptually, the study suggests that nanoengineered peptides for infected defects may need to function not only as anti-infective cues, but also as part of a broader therapeutic programme that counteracts the secondary skeletal damage caused by infection.60
Taken together, these studies indicate that infected bone defects should not be approached as a simple osteogenic deficiency. Instead, they require nanoengineered peptide systems capable of solving a sequence of coupled problems: eliminating bacteria, disrupting biofilm-favoring conditions, correcting inflammatory and acidic microenvironments, protecting against infection-driven bone loss, and only then supporting vascularized bone regeneration or osseointegration.12,55 In this framework, nanoengineered peptides do not merely add antimicrobial activity to a bone material; they reorganize the infected defect niche into one that is again permissive for repair.
Osteoporotic Bone Defects/Osteoporotic Fracture Healing
Osteoporotic bone repair is not simply a weakened version of normal healing. Rather, it is characterized by impaired osteogenesis, insufficient angiogenesis, excessive osteoclast activity, and a microenvironment that is less supportive of endogenous cell function.102,103 In osteoporotic fractures, these deficits are further compounded by delayed cell recruitment and disrupted neurovascular signaling. Under such conditions, therapeutic peptides such as PTH-related sequences or calcitonin gene-related peptide (CGRP) are highly attractive because they are biologically instructive and mechanistically specific;104 however, their free administration is often limited by burst release, rapid diffusion, and poor local retention.105 Nanoengineering is therefore particularly valuable in this setting because it converts these peptides from transient soluble signals into locally anchored, microenvironment-responsive, and sometimes stage-programmed regenerative cues.106
A representative example is the injectable antioxidant supramolecular peptide nanofiber hydrogel developed by Feng et al (2025). In this study, the therapeutic peptide itself was nanoengineered into the material architecture: the PTH-related peptide P1R16 formed a supramolecular nanofiber hydrogel and was further combined with an oxidized alginate/gelatin (OAG) matrix to generate the P1R16-OAG composite hydrogel107 (Figure 5A). This design is important because it addresses one of the central limitations of local PTH delivery in osteoporotic defects—initial burst release—by converting a diffusible anabolic peptide cue into a locally retained nanofibrous matrix. Mechanistically, the composite hydrogel reversed oxidative stress-induced cellular damage, restored the viability, proliferation, migration, and senescence state of both BMSCs and endothelial cells, and shifted BMSCs gene expression toward osteogenesis while suppressing adipogenic drift. In vivo, it showed good biocompatibility and promoted repair of osteoporotic bone defects, indicating that in osteoporosis, peptide nanoengineering can function not only as a delivery strategy but also as a way to rebuild a redox-resilient regenerative niche.
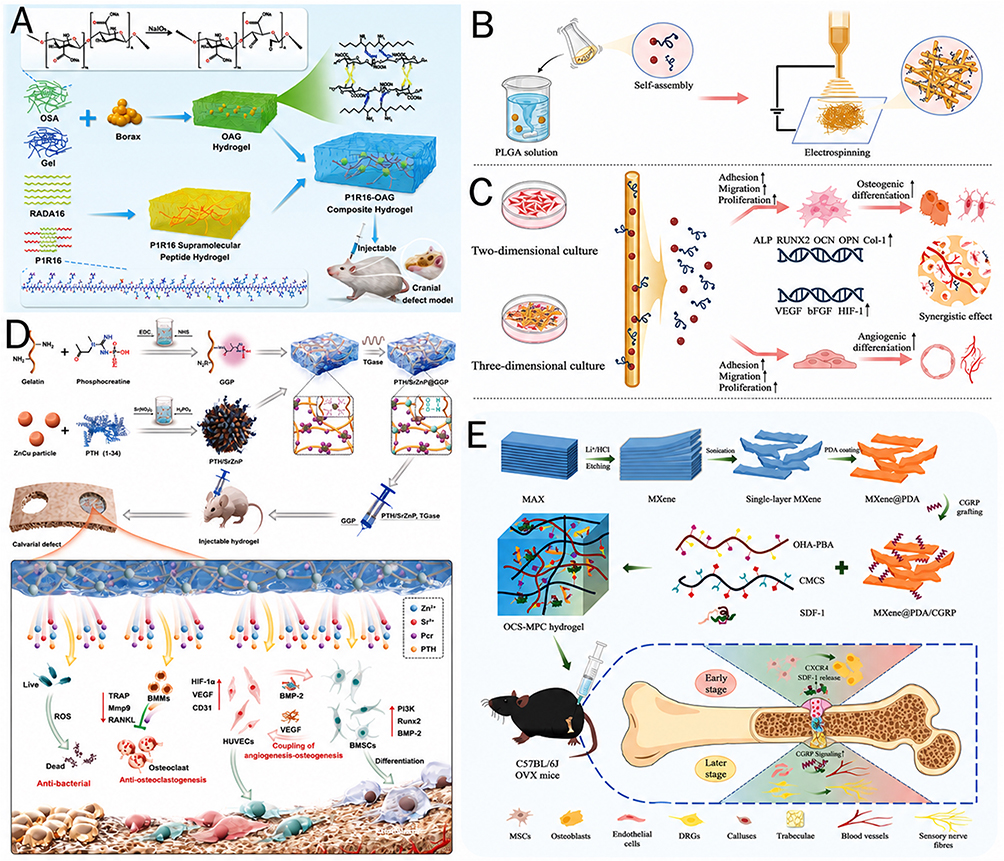 |
Figure 5 Nanoengineered peptide strategies for osteoporotic bone repair and fracture healing. (A) Schematic illustration of the synthesis and multifunctional properties of the P1R16–OAG composite hydrogel. (A) adapted with permission from Ref.107 (B) Schematic illustration of the fabrication of PLGA-PLL-based electrospun membranes loaded with PTHrP1 for bone tissue regeneration. (C) In vitro validation of PLGA-PLL-based membranes in promoting osteogenic differentiation and angiogenic differentiation using 2D culture and 3D co-culture systems. (B and C) adapted with permission from Ref.108 (D) Schematic illustration of the synthesis and mechanism of action of the PTH/SrZnP@GGP hydrogel for osteoporotic bone regeneration. This injectable hydrogel integrates therapeutic peptide signaling with ion-releasing nanostructures to promote osteogenesis, angiogenesis, anti-osteoclastogenesis, antibacterial activity, and bone–vascular coupling. (D) adapted with permission from Ref.109 (E) Schematic illustration of the OHA-PBA/CMCS/SDF-1/MXene@PDA/CGRP (OCS-MPC) dual-crosslinked hydrogel designed for osteoporotic fracture healing. The system enables burst release of SDF-1 to promote early cell recruitment and sustained release of CGRP to support neurovascular reconstruction, callus maturation, and bone repair. (E) adapted with permission from Ref.110 Abbreviations: OAG, oxidized alginate/gelatin; PLGA, poly(lactic-co-glycolic acid); PLL, poly(L-lysine); PTHrP1, parathyroid hormone-related peptide 1; PTH, parathyroid hormone; SrZnP, strontium-zinc phosphate; GGP, phosphocreatine-grafted gelatin platform; OHA, oxidized hyaluronic acid; PBA, phenylboronic acid; CMCS, carboxymethyl chitosan; SDF-1, stromal cell-derived factor-1; MXene, two-dimensional transition metal carbide/nitride nanosheet; PDA, polydopamine; CGRP, calcitonin gene-related peptide; OCS-MPC, OHA-PBA/CMCS/SDF-1/MXene@PDA/CGRP hydrogel. |
A related but structurally distinct strategy was reported by Chen et al (2024), who used electrospun poly(lactic-co-glycolic acid) (PLGA)- poly(L-lysine) (PLL) nanofibrous membranes to achieve local delivery and sustained release of a novel PTHrP1 peptide108 (Figure 5B and C). Here, the key advance was not merely loading a PTH-related peptide into a scaffold, but presenting it within an electrospun nanofibrous matrix that could stabilize peptide residence, support cell adhesion, and mimic aspects of extracellular fibrillar structure. The resulting membrane promoted osteogenic differentiation and angiogenesis in vitro, recruited endogenous stem cells in vivo, and enhanced their ectopic osteogenic differentiation in an osteoporotic environment. It also improved the local osteoporotic microenvironment and facilitated defect repair. From a problem-oriented perspective, this study underscores an important point: for PTHrP-like peptides in osteoporotic bone defects, therapeutic efficacy depends less on intrinsic peptide activity alone than on whether the peptide can be persistently and structurally presented by a nanofibrous matrix.
If the previous two studies primarily highlight stabilization and local retention of PTH-family peptides, the system developed by Qin et al (2025) illustrates why osteoporotic bone defects often require multifunctional nanoengineered peptide platforms rather than single-axis osteoanabolic stimulation109 (Figure 5D). Their injectable phosphocreatine-grafted gelatin hydrogel incorporated hierarchically structured Zn-Cu particles functionalized with a teriparatide/strontium-zinc phosphate hybrid coating, thereby integrating a therapeutic peptide with ion-releasing nanostructures inside one injectable matrix. This architecture yielded improved mechanical properties and sustained release of Zn2⁺, Sr2⁺, and PTH. Functionally, the hydrogel enhanced BMSCs proliferation, osteogenic differentiation, and mineralization, promoted HUVEC migration and tube formation, and inhibited osteoclastogenesis. Importantly, the authors further identified a dual paracrine mechanism underlying osteogenesis-angiogenesis coupling, involving BMSCs-derived hypoxia-inducible factor 1-alpha (HIF-1α)/VEGF signaling and HUVEC-derived phosphoinositide 3-kinase (PI3K)-protein kinase B (Akt)/BMP-2 signaling. In ovariectomized rat calvarial defects, the system accelerated both neovascularization and bone regeneration. This study is therefore particularly useful for your section because it shows that osteoporotic repair failure is not a single “lack of bone formation” problem, but a coupled deficit of osteogenesis, angiogenesis, and bone remodeling that can be more effectively addressed by peptide-centered nanocomposite systems.
For osteoporotic fracture healing, a more explicitly spatiotemporal strategy was demonstrated by Wang et al (2025), who developed a smart composite hydrogel (OCS-MPC) by embedding CGRP-functionalized polydopamine-coated MXene nanosheets into an oxidized hyaluronic acid (OHA)-PBA/carboxymethyl chitosan (CMCS) hydrogel loaded with stromal cell-derived factor-1 (SDF-1)110 (Figure 5E). This design is highly relevant because it aligns peptide nanoengineering with the staged biology of fracture repair. Rather than treating osteoporotic fracture healing as a uniform process, the system was designed to release SDF-1 and CGRP in a controlled manner so as to support different regenerative tasks at different times. In the osteoporotic femoral fracture model, the hydrogel enhanced early MSC recruitment through the SDF-1/C-X-C chemokine receptor type 4 (CXCR4) axis, thereby improving early callus formation, and subsequently promoted late-stage callus mineralization and remodeling through enhanced CGRP signaling. The study also showed increased expression of tubulin beta-3 class III (TUBB3), CGRP, and CD31, supporting regeneration of the neurovascular network. This work is especially valuable for your review because it extends the concept of nanoengineered peptides in osteoporosis from defect filling to true fracture healing, where the therapeutic goal is not only to stimulate bone formation, but to restore the temporal sequence of recruitment, neurovascular reconstitution, callus maturation, and remodeling.
Taken together, these studies indicate that nanoengineered peptides for osteoporotic bone repair are most effective when they are designed to match the specific pathological bottlenecks imposed by osteoporosis. In osteoporotic bone defects, PTH-related peptides benefited from conversion into supramolecular nanofiber hydrogels or electrospun nanofibrous membranes, which improved local retention and broadened function from direct osteoanabolism to microenvironmental correction and endogenous-cell support. In more complex osteoporotic defects, peptide-functionalized nanocomposite hydrogels enabled simultaneous regulation of osteogenesis, angiogenesis, and osteoclast activity. In osteoporotic fracture healing, peptide nanoengineering further evolved toward stage-programmed delivery, allowing early recruitment and later remodeling signals to be integrated within one platform. In osteoporotic repair, nanoengineering therefore serves a dual purpose: it compensates for the pharmacokinetic vulnerabilities of free peptides, and simultaneously converts peptide bioactivity into a format that is compatible with the compromised cellular, vascular, and metabolic state of the osteoporotic niche.
Biomaterialization as a Mechanism-Amplifying Extension of Peptide Nanoengineering
Biomaterialization should be understood not as a secondary packaging step for therapeutic peptides, but as a functional extension of peptide nanoengineering.27 Once peptides are nanoengineered into supramolecular assemblies, nanofiber-displayed ligands, nanoparticle-coupled signals, or nanoscale interfacial motifs, their biological activity is no longer determined by sequence alone. Instead, their therapeutic performance becomes increasingly dependent on how these nanoengineered states are stabilized, spatially organized, and deployed within the defect environment.28 In this sense, biomaterialization amplifies mechanism not simply by improving peptide delivery, but by converting nanoengineered peptides into persistent, structured, and context-responsive regenerative signals.111 What is ultimately enhanced is not only peptide residence time, but also the way peptide cues are presented to cells, integrated with matrices, and synchronized with the pathological logic of bone healing.112
Retention: Sustaining Peptide Activity Within the Defect
The first level at which biomaterialization amplifies nanoengineered peptide function is signal stabilization.113 Therapeutic peptides are intrinsically vulnerable to enzymatic degradation, rapid diffusion, and premature dilution after local administration, which often prevents them from maintaining a biologically meaningful concentration within bone defects.44 Nanoengineering partially addresses this problem by transforming peptides into nanofibers, nanocomplexes, or nanocarrier-associated signals; biomaterialization then further secures these nanoengineered states within injectable matrices, nanofibrous scaffolds, or defect-filling hydrogels.14,84 As a result, peptide activity is no longer transiently exposed to the defect site, but retained as a localized and persistent signal reservoir.44 Mechanistically, this shift is crucial because many osteogenic, angiogenic, or immunoregulatory peptide cues require sustained local presence rather than brief molecular exposure.114 Thus, biomaterialization improves nanoengineered peptides not merely by prolonging retention in a pharmacokinetic sense, but by converting unstable peptide signaling into a persistent regenerative niche.89
Geometry: Fixing Peptide Signals in a Cell-Readable Matrix Context
A second and more profound effect of biomaterialization is that it changes the ontological role of nanoengineered peptides in the regenerative system. Before biomaterialization, even nanoengineered peptides may still behave primarily as mobile bioactive entities. After biomaterialization, however, they can become part of the structural and instructive architecture of the matrix itself.28 This transition is particularly important in bone healing, where matrix organization is not passive background, but an active determinant of cell behavior, mineral deposition, and tissue integration.81 By immobilizing or structurally embedding nanoengineered peptides within nanofibrous, mineralized, or hybrid matrix systems, biomaterialization transforms them from soluble cues into matrix-bound instructions. In this state, peptide function is no longer limited to receptor activation after release; it extends to guiding where signals accumulate, how minerals nucleate, how cells attach, and how tissue ingrowth is spatially organized.6 Therefore, nanoengineering makes peptides structurally definable, whereas biomaterialization makes those structures biologically instructive.
Multivalent Presentation: Changing the Geometry of Bioactivity
The third mechanism amplification arises from nanoscale presentation.115 Free peptides are usually encountered by cells as dispersed and short-lived molecules, which limits their ability to achieve multivalent engagement, receptor clustering, and stable interfacial recognition.116 Nanoengineering overcomes this by organizing peptides into dense and spatially controlled nanoscale displays, while biomaterialization fixes these displays within matrices or interfaces that cells can physically interrogate.50,117 This combination creates a mode of signaling that is qualitatively different from soluble stimulation.118 Rather than acting as isolated ligands, biomaterialized nanoengineered peptides can be perceived as clustered, multivalent, and topologically organized cues that favor stronger receptor binding, more efficient downstream signaling, and more selective recruitment of target cell populations such as BMSCs, endothelial cells, or macrophages.89 In this context, nanoengineering does not simply increase the amount of peptide at the interface; it changes the geometry of bioactivity. Biomaterialization then preserves this geometry long enough for cells to interpret it as a stable regenerative instruction. The improvement in peptide function therefore lies not only in dose retention, but in the emergence of a nanoscale signaling logic that soluble peptides cannot readily achieve.
Context Matching: Adapting Peptide Function to Defect-Specific Pathology
A peptide may encode osteogenic, angiogenic, antimicrobial, or cell-homing potential, but whether that potential is realized depends on the pathological environment in which it is deployed.110 Biomaterialization provides the format through which nanoengineered peptides can be matched to the dominant bottlenecks of specific defects. In infected bone defects, this may involve integrating antimicrobial or immunomodulatory peptides into responsive matrices that suppress bacteria while restoring a repair-permissive niche.53 In osteoporotic defects, it may require embedding osteoanabolic or recruitment-promoting peptides into injectable or nanofibrous systems that compensate for oxidative stress, poor cell responsiveness, and impaired vascular coupling.24 In fracture healing, it may involve temporally staged materials that align early recruitment with later callus maturation and remodeling.54,107 At implant interfaces, it may require surface-bound peptide nanoarchitectures that promote selective host-cell engagement and osseointegration.119 Biomaterialization thus adds what sequence cannot provide on its own: pathology-adapted deployment.
Taken together, biomaterialization contributes four functions that are not encoded by peptide sequence itself—retention, geometry, responsiveness, and context matching. These are not peripheral formulation issues. They are the factors that determine whether a nanoengineered peptide remains present long enough, is displayed in a cell-interpretable form, acts at the right stage of repair, and is deployed in a format appropriate for the defect being treated. Biomaterialization should therefore be understood as a mechanism-amplifying extension of peptide nanoengineering rather than a downstream packaging step.
Outlook and Conclusion
Outlook
A first major opportunity lies in artificial intelligence (AI)-assisted peptide sequence design. Recent advances in generative and structure-based design are rapidly changing what is possible in peptide engineering. In Nature, RoseTTAFold diffusion (RFdiffusion)-enabled approaches generated high-affinity binders for bioactive helical peptides, demonstrating that flexible peptide recognition can now be designed with minimal starting information.120 Deep learning was used to design self-assembling AMP de novo, showing that sequence, assembly behaviour and function can be co-optimized computationally rather than discovered only by trial and error.121 Importantly, this trajectory is now beginning to reach bone repair directly: a 2026 study reported an AI-selected multifunctional oligopeptide-coated biphasic calcium phosphate (BCP) scaffold that combined osteogenic and angiogenic activities with hydroxyapatite-binding capacity and promoted bone defect repair in vivo.122 Together with recent bone-regeneration reviews advocating database-driven and AI-enabled biomaterial development, these studies suggest that AI-assisted approaches have the potential to accelerate peptide discovery and optimization, though the integration of computational design with rigorous in vivo mechanistic validation and defect-specific testing remains at an early stage and will require substantial further development before clinical translation becomes feasible.
A second priority is defect-stage-matched peptide release, coupled with a transition from binary pathway targeting toward immuno-angiogenic-osteogenic tri-coupling. Sequentially regulated systems are already demonstrating why this matters. A recent study showed that an inflammation-responsive peptide coating can first induce M2 macrophage polarization and then expose angiogenic and osteogenic motifs in a physiologically aligned sequence.55 Likewise, stem-cell-homing hydrogels for osteoporotic defects have shown that recruitment, angiogenesis, osteogenesis and even immunoregulation can be coordinated within a single dynamic matrix, while sequential SDF-1/CGRP delivery systems further illustrate the value of matching signal order to fracture-healing stages.110 Future peptide platforms should therefore move beyond the simple question of whether they promote bone formation, and instead ask whether they can orchestrate immune correction, vascular support and osteogenesis in the right order, at the right location and with the right persistence. This need for context sensitivity is reinforced by recent research showing that calvarial healing differs fundamentally from femoral fracture repair, including a striking uncoupling of angiogenesis from osteogenesis in flat bone regeneration.26
A third direction is the development of mineralization-guiding nanostructured peptides. Much of the current literature still evaluates osteogenic materials mainly through lineage markers, yet functional bone repair also depends on where, when and how mineral is deposited within the evolving matrix. Recent peptide systems are beginning to move decisively in this direction. Sequential protein-instructed peptide co-assembly has been used to generate adaptive biphasic scaffolds in which peptide assembly, ECM association and calcium phosphate deposition evolve together.123 Earlier phosphoserine-containing peptide amphiphile nanofibers had already demonstrated that biomimetic mineralization can be encoded directly in peptide sequence and translated into in vivo regeneration.6 Looking forward, one of the most promising frontiers will be to integrate mineral-templating motifs, dynamic enzymatic responsiveness and mechanically relevant supramolecular organization within the same peptide nanoarchitecture, so that nanostructured peptides can guide not only osteogenic differentiation but also the emergence of bone-like matrix hierarchy.124
Finally, the field must move from calvarial proof-of-concept to load-bearing and nonunion-relevant validation. Rodent calvarial defects have been invaluable for mechanistic discovery, but they remain non-load-bearing and do not reproduce the biomechanical demands, tissue ingress distances or failure modes of long-bone nonunion and segmental reconstruction.125 A recent translational analysis of rodent calvarial models explicitly highlighted the absence of physiological mechanical loading as a major limitation, and recent intravital studies further showed that calvarial repair differs biologically from long-bone healing.126 Encouragingly, the field is beginning to move toward more relevant validation: Nature Communications recently reported long-term reconstruction of ovine segmental mandibulectomy defects using a patient-matched, mechanobiologically optimized scaffold-guided system, illustrating the type of large-animal, mechanically demanding evidence that future peptide-based constructs will ultimately need to generate.127 For nanoengineered therapeutic peptides, the immediate priority is not to claim translational readiness, but to generate the mechanically relevant, large-animal, and clinically contextualized preclinical evidence that would justify progression toward first-in-human studies—evidence that, for the vast majority of systems reviewed here, remains to be produced.
In this sense, the future of the field is unlikely to be defined by a single “best peptide”, but by whether mechanism-guided peptide systems can be computationally discoverable, biomaterially programmable, and scalable for clinically relevant use.
Conclusion
This Review highlights that peptide-based bone regeneration is increasingly moving from empirical motif grafting toward a mechanism-guided paradigm in which peptide sequence, nanoscale presentation, defect-stage biology, and translational constraints are treated as an integrated design problem. A central message is that peptides should not be viewed merely as miniaturized growth-factor substitutes: when nanoengineered and biomaterialized, they can function as programmable molecular instructions that shape immune resolution, vascular invasion, endogenous cell participation, matrix organization, mineralization, and remodeling in a context-dependent manner.
Despite clear progress, the current evidence base remains largely preclinical and model-dependent, with many studies performed in simplified or non-load-bearing defects that do not reproduce clinically challenging scenarios such as mechanically demanding segmental defects, nonunion, established infection, or osteoporotic microenvironments. Direct, quantitative comparisons between free peptides and nanoengineered peptide presentations are still uncommon, which limits the ability to isolate the incremental benefit of nanoscale presentation. In addition, commonly reported endpoints (eg, marker expression, mineral deposition, vessel density, or radiographic bone volume) do not always establish mechanically competent, hierarchically organized, and remodelable bone. Translation-facing issues—including scalable manufacturing, batch consistency, sterilization stability, long-term biodegradation, immunological safety, and regulatory pathways for multifunctional nanoarchitectures—also remain insufficiently addressed.
Future progress will likely depend on developing predictive structure–presentation–function relationships and stage-matched, defect-adaptive delivery strategies that coordinate immune, vascular, osteogenic, and remodeling cues in the right sequence and at the right location. Promising directions include data-driven and AI-assisted peptide design, mineralization-guiding nanoarchitectures that prioritize matrix organization and mechanical competence rather than mineral quantity alone, and more rigorous validation in clinically relevant load-bearing and nonunion-prone models with standardized functional outcome reporting. Overall, nanoengineered therapeutic peptides represent a highly programmable platform, but their clinical impact will ultimately be determined by reproducible, safe, and surgically practical performance under clinically realistic conditions.
Abbreviations
ALP, Alkaline phosphatase; M2, M2-like macrophage; AMP, Antimicrobial peptide; AOHA, Antimicrobial peptide-conjugated oxidized hyaluronic acid; AI, Artificial intelligence; BCP, Biphasic calcium phosphate; BP, Black phosphorus; BMSCs, Bone marrow mesenchymal stem cells; BMP-2, Bone morphogenetic protein-2; CGRP, Calcitonin gene-related peptide; CMCS, Carboxymethyl chitosan; M1, M1-like macrophage; CD31, Cluster of differentiation 31; CD90, Cluster of differentiation 90; cGMP, Cyclic guanosine monophosphate; CXCR4, C-X-C chemokine receptor type 4; DDR2, Discoidin domain receptor 2; DPP, Dentin phosphoprotein; ECM, Extracellular matrix; HUVEC, Human umbilical vein endothelial cells; HIF-1α, Hypoxia-inducible factor 1-alpha; IGF-I, Insulin-like growth factor-I; MnO2, Manganese dioxide; MSNs, Mesoporous silica nanoparticles; MOF, Metal-organic framework; NIR, Near-infrared; NO, Nitric oxide; OAG, Oxidized alginate/gelatin; OHA, Oxidized hyaluronic acid; OGP, Osteogenic growth peptide; PTH, Parathyroid hormone; PTHrP, Parathyroid hormone-related peptide; PPAR-γ, Peroxisome proliferator-activated receptor gamma; PI3K, Phosphoinositide 3-kinase; PEG, Poly(ethylene glycol); PEEK, Polyetheretherketone; PLGA, Poly(lactic-co-glycolic acid); PLLA, Poly(L-lactic acid); PLL, Poly(L-lysine); Akt, Protein kinase B; ROS, Reactive oxygen species; RFdiffusion, RoseTTAFold diffusion; STAT6, Signal transducer and activator of transcription 6; SDF-1, Stromal cell-derived factor-1; sPEEK, Sulfonated polyetheretherketone; SOCS3, Suppressor of cytokine signaling 3; TUBB3, Tubulin beta-3 class III; VEGF, Vascular endothelial growth factor; ZIF-8, Zeolitic imidazolate framework-8; α-SMA, Alpha-smooth muscle actin.
Disclosure
Shan-Qi Zou and Jing-Chao Yu are first authors. Dr Yong-Jing Ye provided all support for the present manuscript, during the conduct of the study. The authors report no conflicts of interest in this work.
References
1. Wang W, Yeung KWK. Bone grafts and biomaterials substitutes for bone defect repair: a review. Bioact Mater. 2017;2(4):224–28. doi:10.1016/j.bioactmat.2017.05.007
2. Schlundt C, El Khassawna T, Serra A, et al. Macrophages in bone fracture healing: their essential role in endochondral ossification. Bone. 2018;106:78–89. doi:10.1016/j.bone.2015.10.019
3. Xu G, Chen J, Xiao Z, et al. Bioaugmented osteosynthesis: precise monitoring and intervention of the bone healing microenvironment. Bone Res. 2025;13(1):95. doi:10.1038/s41413-025-00466-5
4. Ho-Shui-Ling A, Bolander J, Rustom LE, et al. Bone regeneration strategies: engineered scaffolds, bioactive molecules and stem cells current stage and future perspectives. Biomaterials. 2018;180:143–162. doi:10.1016/j.biomaterials.2018.07.017
5. Visser R, Rico-Llanos GA, Pulkkinen H, et al. Peptides for bone tissue engineering. J Control Release. 2016;244:122–135. doi:10.1016/j.jconrel.2016.10.024
6. Mata A, Geng Y, Henrikson KJ, et al. Bone regeneration mediated by biomimetic mineralization of a nanofiber matrix. Biomaterials. 2010;31(23):6004–6012. doi:10.1016/j.biomaterials.2010.04.013
7. Chen CH, Hsu EL, Stupp SI. Supramolecular self-assembling peptides to deliver bone morphogenetic proteins for skeletal regeneration. Bone. 2020;141:115565. doi:10.1016/j.bone.2020.115565
8. Hao Z, Chen R, Chai C, et al. Antimicrobial peptides for bone tissue engineering: diversity, effects and applications. Front Bioeng Biotechnol. 2022;10:1030162. doi:10.3389/fbioe.2022.1030162
9. Li G, Luo Y, Hu Z, et al. Recent advances in peptide-functionalized hydrogels for bone tissue engineering. ACS Biomater Sci Eng. 2025;11(4):1970–1989. doi:10.1021/acsbiomaterials.4c02198
10. Pountos I, Panteli M, Lampropoulos A, et al. The role of peptides in bone healing and regeneration: a systematic review. BMC Med. 2016;14(1):103. doi:10.1186/s12916-016-0646-y
11. Liao J, Wu S, Li K, et al. Peptide-modified bone repair materials: factors influencing osteogenic activity. J Biomed Mater Res A. 2019;107(7):1491–1512. doi:10.1002/jbm.a.36663
12. Li M, Zhao P, Wang J, et al. Functional antimicrobial peptide-loaded 3D scaffolds for infected bone defect treatment with AI and multidimensional printing. Mater Horizons. 2025;12(1):20–36. doi:10.1039/D4MH01124D
13. Lee JY, Choi YS, Lee SJ, et al. Bioactive peptide-modified biomaterials for bone regeneration. Curr Pharm Des. 2011;17(25):2663–2676. doi:10.2174/138161211797416011
14. Niu Z, Fan Y, Yin L, et al. Advancing nanomedicine: for bone defect repair and regeneration. Int J Nanomed. 2025;20:15043–15062. doi:10.2147/IJN.S545353
15. Azadi S, Yazdanpanah MA, Afshari A, et al. Bioinspired synthetic peptide-based biomaterials regenerate bone through biomimicking of extracellular matrix. J Tissue Eng. 2024;15:20417314241303818. doi:10.1177/20417314241303818
16. Wang S, Liu Y, Zhou Y, et al. Biomimetic peptides in bone tissue engineering: from function to application. Colloids Surf B Biointerfaces. 2025;256(Pt 1):115017. doi:10.1016/j.colsurfb.2025.115017
17. Ravichandran R, Griffith M, Phopase J. Applications of self-assembling peptide scaffolds in regenerative medicine: the way to the clinic. J Mater Chem B. 2014;2(48):8466–8478. doi:10.1039/C4TB01095G
18. Wang C, Liu Y, Fan Y, et al. The use of bioactive peptides to modify materials for bone tissue repair. Regen Biomater. 2017;4(3):191–206. doi:10.1093/rb/rbx011
19. Zhou T, Ma PX. Peptide-conjugated biodegradable polyester scaffolds for bone regeneration. Biomaterials. 2026;329:123919. doi:10.1016/j.biomaterials.2025.123919
20. Hao Z, Li H, Wang Y, et al. Supramolecular peptide nanofiber hydrogels for bone tissue engineering: from multihierarchical fabrications to comprehensive applications. Adv Sci. 2022;9(11):2103820. doi:10.1002/advs.202103820
21. Wan L, Yao X, Pan J, et al. Crafting the future of bone regeneration: the promise of supramolecular peptide nanofiber hydrogels. Front Bioeng Biotechnol. 2025;13. doi:10.3389/fbioe.2025.1514318
22. Wu J, Zheng A, Liu Y, et al. Enhanced bone regeneration of the silk fibroin electrospun scaffolds through the modification of the graphene oxide functionalized by BMP-2 peptide. Int J Nanomed. 2019;14:733–751. doi:10.2147/IJN.S187664
23. Wojtowicz AM, Shekaran A, Oest ME, et al. Coating of biomaterial scaffolds with the collagen-mimetic peptide GFOGER for bone defect repair. Biomaterials. 2010;31(9):2574–2582. doi:10.1016/j.biomaterials.2009.12.008
24. Duda GN, Geissler S, Checa S, et al. The decisive early phase of bone regeneration. Nat Rev Rheumatol. 2023;19(2):78–95. doi:10.1038/s41584-022-00887-0
25. Wen C, Xu X, Zhang Y, et al. Bone targeting nanoparticles for the treatment of osteoporosis. Int J Nanomed. 2024;19:1363–1383. doi:10.2147/IJN.S444347
26. Bixel MG, Sivaraj KK, Timmen M, et al. Angiogenesis is uncoupled from osteogenesis during calvarial bone regeneration. Nat Commun. 2024;15(1):4575. doi:10.1038/s41467-024-48579-5
27. Webber MJ, Appel EA, Meijer EW, et al. Supramolecular biomaterials. Nature Mater. 2016;15(1):13–26. doi:10.1038/nmat4474
28. Levin A, Hakala TA, Schnaider L, et al. Biomimetic peptide self-assembly for functional materials. Nat Rev Chemist. 2020;4(11):615–634. doi:10.1038/s41570-020-0215-y
29. Clark AY, Martin KE, García JR, et al. Integrin-specific hydrogels modulate transplanted human bone marrow-derived mesenchymal stem cell survival, engraftment, and reparative activities. Nat Commun. 2020;11(1):114. doi:10.1038/s41467-019-14000-9
30. Dalby MJ, Gadegaard N, Oreffo ROC. Harnessing nanotopography and integrin–matrix interactions to influence stem cell fate. Nature Mater. 2014;13(6):558–569. doi:10.1038/nmat3980
31. Li W, Zheng Y, Zhao X, et al. Osteoinductive effects of free and immobilized bone forming peptide-1 on human adipose-derived stem cells. PLoS One. 2016;11(3):e0150294. doi:10.1371/journal.pone.0150294
32. Huang J, Gräter SV, Corbellini F, et al. Impact of order and disorder in RGD nanopatterns on cell adhesion. Nano Lett. 2009;9(3):1111–1116. doi:10.1021/nl803548b
33. Ramasamy SK, Kusumbe AP, Wang L, et al. Endothelial Notch activity promotes angiogenesis and osteogenesis in bone. Nature. 2014;507(7492):376–380. doi:10.1038/nature13146
34. Gultian KA, Gandhi R, Decesari K, et al. Injectable hydrogel with immobilized BMP-2 mimetic peptide for local bone regeneration. Front Biomater Sci. 2022. doi:10.3389/fbiom.2022.948493
35. Frederix PWJM, Scott GG, Abul-Haija YM, et al. Exploring the sequence space for (tri-)peptide self-assembly to design and discover new hydrogels. Nat Chemist. 2015;7(1):30–37. doi:10.1038/nchem.2122
36. Fjell CD, Hiss JA, Hancock REW, et al. Designing antimicrobial peptides: form follows function. Nat Rev Drug Discov. 2012;11(1):37–51. doi:10.1038/nrd3591
37. Randall JR, Vieira LC, Wilke CO, et al. Deep mutational scanning and machine learning for the analysis of antimicrobial-peptide features driving membrane selectivity. Nat Biomed Eng. 2024;8(7):842–853. doi:10.1038/s41551-024-01243-1
38. Boix-Lemonche G, Guillem-Marti J, D’este F, et al. Covalent grafting of titanium with a cathelicidin peptide produces an osteoblast compatible surface with antistaphylococcal activity. Colloids Surf B Biointerfaces. 2020;185:110586. doi:10.1016/j.colsurfb.2019.110586
39. Chen J, Hu G, Li T, et al. Fusion peptide engineered “statically-versatile” titanium implant simultaneously enhancing anti-infection, vascularization and osseointegration. Biomaterials. 2021;264:120446. doi:10.1016/j.biomaterials.2020.120446
40. De Souza Araújo IJ, Ferreira JA, Daghrery A, et al. Self-assembling peptide-laden electrospun scaffolds for guided mineralized tissue regeneration. Dent Mater. 2022;38(11):1749–1762. doi:10.1016/j.dental.2022.09.011
41. Saha S, Yang XB, Wijayathunga N, et al. A biomimetic self-assembling peptide promotes bone regeneration in vivo: a rat cranial defect study. Bone. 2019;127:602–611. doi:10.1016/j.bone.2019.06.020
42. Zhang F, Lv M, Wang S, et al. Ultrasound-triggered biomimetic ultrashort peptide nanofiber hydrogels promote bone regeneration by modulating macrophage and the osteogenic immune microenvironment. Bioact Mater. 2024;31:231–246. doi:10.1016/j.bioactmat.2023.08.008
43. Sargeant TD, Guler MO, Oppenheimer SM, et al. Hybrid bone implants: self-assembly of peptide amphiphile nanofibers within porous titanium. Biomaterials. 2008;29(2):161–171. doi:10.1016/j.biomaterials.2007.09.012
44. Shekaran A, García JR, Clark AY, et al. Bone regeneration using an alpha 2 beta 1 integrin-specific hydrogel as a BMP-2 delivery vehicle. Biomaterials. 2014;35(21):5453–5461. doi:10.1016/j.biomaterials.2014.03.055
45. Gulseren G, Tansik G, Garifullin R, et al. Dentin phosphoprotein mimetic peptide nanofibers promote biomineralization. Macromol Biosci. 2019;19(1):e1800080. doi:10.1002/mabi.201800080
46. Yu H, Song J, Zhang X, et al. Hydroxyapatite-tethered peptide hydrogel promotes osteogenesis. Gels. 2022;8(12):804. doi:10.3390/gels8120804
47. Liu J, Zhao B, Shen X, et al. Enhanced vascularization and osseointegration under osteoporotic conditions through functional peptide coating on implant surfaces. Mater Today Bio. 2024;27:101150. doi:10.1016/j.mtbio.2024.101150
48. Chen W, Sheng S, Tan K, et al. Injectable hydrogels for bone regeneration with tunable degradability via peptide chirality modification. Mater Horizons. 2024;11(18):4367–4377. doi:10.1039/D4MH00398E
49. Padilla-Lopategui S, Ligorio C, Bu W, et al. Biocooperative regenerative materials by harnessing blood-clotting and peptide self-assembly. Adv Mater. 2024;36(52):e2407156. doi:10.1002/adma.202407156
50. Zhou T, C Cavalcante R, Ge C, Franceschi RT, Ma PX. Synthetic helical peptides on nanofibers to activate cell-surface receptors and synergistically enhance critical-sized bone defect regeneration. Bioact Mater. 2025;43:98–113. doi:10.1016/j.bioactmat.2024.08.017
51. Wei Y, Pan H, Yang J, et al. Aligned cryogel fibers incorporated 3D printed scaffold effectively facilitates bone regeneration by enhancing cell recruitment and function. Sci Adv. 2024;10(6):eadk6722. doi:10.1126/sciadv.adk6722
52. Lutolf MP, Weber FE, Schmoekel HG, et al. Repair of bone defects using synthetic mimetics of collagenous extracellular matrices. Nat Biotechnol. 2003;21(5):513–518. doi:10.1038/nbt818
53. Zhang M, Yu T, Li J, et al. Matrix metalloproteinase-responsive hydrogel with on-demand release of phosphatidylserine promotes bone regeneration through immunomodulation. Adv Sci. 2024;11(20):e2306924. doi:10.1002/advs.202306924
54. Fischer NG, Chen X, Astleford-Hopper K, et al. Antimicrobial and enzyme-responsive multi-peptide surfaces for bone-anchored devices. Mater Sci Eng C Mater Biol Appl. 2021;125:112108. doi:10.1016/j.msec.2021.112108
55. Zhou W, Liu Y, Nie X, et al. Peptide-based inflammation-responsive implant coating sequentially regulates bone regeneration to enhance interfacial osseointegration. Nat Commun. 2025;16(1):3283. doi:10.1038/s41467-025-58444-8
56. Liu Y, Li L, He M, et al. Self-assembled peptide hydrogel loaded with functional peptide Dentonin accelerates vascularized bone tissue regeneration in critical-size bone defects. Regen Biomater. 2024;11:rbae106. doi:10.1093/rb/rbae106
57. Sun P, Zhang Q, Nie W, et al. Biodegradable mesoporous silica nanocarrier bearing angiogenic QK peptide and dexamethasone for accelerating angiogenesis in bone regeneration. ACS Biomater Sci Eng. 2019;5(12):6766–6778. doi:10.1021/acsbiomaterials.9b01521
58. Liu S, Han Z, Hao JN, et al. Engineering of a NIR-activable hydrogel-coated mesoporous bioactive glass scaffold with dual-mode parathyroid hormone derivative release property for angiogenesis and bone regeneration. Bioact Mater. 2023;26:1–13. doi:10.1016/j.bioactmat.2023.02.008
59. Chen J, Zhao Y, Ruan R, et al. Bone morphogenetic protein-2–derived peptide-conjugated nanozyme-integrated photoenhanced hybrid hydrogel for cascade-regulated bone regeneration. ACS Nano. 2025;19(15):14707–14726. doi:10.1021/acsnano.4c13690
60. Ye P, Yang Y, Qu Y, et al. LL-37 and bisphosphonate co-delivery 3D-scaffold with antimicrobial and antiresorptive activities for bone regeneration. Int J Biol Macromol. 2024;277(Pt 1):134091. doi:10.1016/j.ijbiomac.2024.134091
61. Ma S, Yao S, Li Y, et al. A pH-responsive polyetheretherketone implant modified with a core-shell metal-organic framework to promote antibacterial and osseointegration abilities. Biomater Res. 2025;29:0188. doi:10.34133/bmr.0188
62. Hachemi Y, Perrin S, Ethel M, et al. Multimodal analyses of immune cells during bone repair identify macrophages as a therapeutic target in musculoskeletal trauma. Bone Res. 2024;12(1):56. doi:10.1038/s41413-024-00347-3
63. Raggatt LJ, Wullschleger ME, Alexander KA, et al. Fracture healing via periosteal callus formation requires macrophages for both initiation and progression of early endochondral ossification. Am J Pathol. 2014;184(12):3192–3204. doi:10.1016/j.ajpath.2014.08.017
64. Murray PJ, Allen JE, Biswas SK, et al. Macrophage activation and polarization: nomenclature and experimental guidelines. Immunity. 2014;41(1):14–20. doi:10.1016/j.immuni.2014.06.008
65. Vi L, Baht GS, Whetstone H, et al. Macrophages promote osteoblastic differentiation in-vivo: implications in fracture repair and bone homeostasis. J Bone Miner Res. 2015;30(6):1090–1102. doi:10.1002/jbmr.2422
66. Wasnik S, Rundle CH, Baylink DJ, et al. 1,25-Dihydroxyvitamin D suppresses M1 macrophages and promotes M2 differentiation at bone injury sites. JCI Insight. 2018;3(17). doi:10.1172/jci.insight.98773
67. Yu X, Quan J, Long W, et al. LL-37 inhibits LPS-induced inflammation and stimulates the osteogenic differentiation of BMSCs via P2X7 receptor and MAPK signaling pathway. Exp Cell Res. 2018;372(2):178–187. doi:10.1016/j.yexcr.2018.09.024
68. Kong Q, Gao S, Li P, et al. Calcitonin gene-related peptide-modulated macrophage phenotypic alteration regulates angiogenesis in early bone healing. Int Immunopharmacol. 2024;130:111766. doi:10.1016/j.intimp.2024.111766
69. Xiao W, Jiang W, Chen Z, et al. Advance in peptide-based drug development: delivery platforms, therapeutics and vaccines. Signal Transduct Targeted Ther. 2025;10(1):74. doi:10.1038/s41392-024-02107-5
70. Yin X, Yan L, Jun HD, et al. Calcium alginate template-mineral substituted hydroxyapatite hydrogel coated titanium implant for tibia bone regeneration. Int J Pharm. 2020;582:119303. doi:10.1016/j.ijpharm.2020.119303
71. Jeon EY, Um SH, Park J, et al. Precisely localized bone regeneration mediated by marine-derived microdroplets with superior BMP-2 binding affinity. Small. 2022;18(24):e2200416. doi:10.1002/smll.202200416
72. Loxley GA, Coser C, Ghaemmaghami AM, et al. Long-term interleukin-4 release from 3D printable affinity hydrogels promotes M2-like macrophage polarisation in vitro. Biomater. Sci. 2025;13(9):2489–2502. doi:10.1039/D4BM01623H
73. Ramasamy SK, Kusumbe AP, Schiller M, et al. Blood flow controls bone vascular function and osteogenesis. Nat Commun. 2016;7(1):13601. doi:10.1038/ncomms13601
74. Hu K, Olsen BR. Osteoblast-derived VEGF regulates osteoblast differentiation and bone formation during bone repair. J Clin Invest. 2016;126(2):509–526. doi:10.1172/JCI82585
75. Burger MG, Grosso A, Briquez PS, et al. Robust coupling of angiogenesis and osteogenesis by VEGF-decorated matrices for bone regeneration. Acta Biomater. 2022;149:111–125. doi:10.1016/j.actbio.2022.07.014
76. Bakshi R, Hokugo A, Khalil D, et al. A chemotactic functional scaffold with VEGF-releasing peptide amphiphiles facilitates bone regeneration by BMP-2 in a large-scale rodent cranial defect model. Plast Reconstr Surg. 2021;147(2):386–397. doi:10.1097/PRS.0000000000007551
77. Grosso A, Lunger A, Burger MG, et al. VEGF dose controls the coupling of angiogenesis and osteogenesis in engineered bone. NPJ Regen Med. 2023;8(1):15. doi:10.1038/s41536-023-00288-1
78. Wang WY, Kent RN, Huang SA, et al. Direct comparison of angiogenesis in natural and synthetic biomaterials reveals that matrix porosity regulates endothelial cell invasion speed and sprout diameter. Acta Biomater. 2021;135:260–273. doi:10.1016/j.actbio.2021.08.038
79. Thrivikraman G, Athirasala A, Gordon R, et al. Rapid fabrication of vascularized and innervated cell-laden bone models with biomimetic intrafibrillar collagen mineralization. Nat Commun. 2019;10(1):3520. doi:10.1038/s41467-019-11455-8
80. Nair AK, Gautieri A, Chang SW, et al. Molecular mechanics of mineralized collagen fibrils in bone. Nat Commun. 2013;4(1):1724. doi:10.1038/ncomms2720
81. Petersen A, Princ A, Korus G, et al. A biomaterial with a channel-like pore architecture induces endochondral healing of bone defects. Nat Commun. 2018;9(1):4430. doi:10.1038/s41467-018-06504-7
82. Wang S, Yang Y, Wang R, et al. Mineralization of calcium phosphate controlled by biomimetic self-assembled peptide monolayers via surface electrostatic potentials. Bioact Mater. 2020;5(2):387–397. doi:10.1016/j.bioactmat.2020.03.003
83. Choi YS, Lee JY, Suh JS, et al. The mineralization inducing peptide derived from dentin sialophosphoprotein for bone regeneration. J Biomed Mater Res A. 2013;101(2):590–598. doi:10.1002/jbm.a.34352
84. Lee SS, Huang BJ, Kaltz R, et al. Bone regeneration with low dose BMP-2 amplified by biomimetic supramolecular nanofibers within collagen scaffolds. Biomaterials. 2013;34(2):452–459. doi:10.1016/j.biomaterials.2012.10.005
85. Nudelman F, Pieterse K, George A, et al. The role of collagen in bone apatite formation in the presence of hydroxyapatite nucleation inhibitors. Nature Mater. 2010;9(12):1004–1009. doi:10.1038/nmat2875
86. Newcomb CJ, Bitton R, Velichko YS, et al. The role of nanoscale architecture in supramolecular templating of biomimetic hydroxyapatite mineralization. Small. 2012;8(14):2195–202,4. doi:10.1002/smll.201102150
87. Bai J, Ge G, Wang Q, et al. Engineering Stem Cell Recruitment and Osteoinduction via Bioadhesive Molecular Mimics to Improve Osteoporotic Bone-Implant Integration. Research. 2022;2022:9823784. doi:10.34133/2022/9823784
88. Kim SH, Hur W, Kim JE, et al. Self-assembling peptide nanofibers coupled with neuropeptide substance P for bone tissue engineering. Tissue Eng Part A. 2015;21(7–8):1237–1246. doi:10.1089/ten.tea.2014.0472
89. Wei FL, Zhai Y, Wang TF, et al. Stem cell-homing biomimetic hydrogel promotes the repair of osteoporotic bone defects through osteogenic and angiogenic coupling. Sci Adv. 2024;10(44):eadq6700. doi:10.1126/sciadv.adq6700
90. Long S, Wang W, Chen Y, et al. E7peptide and magnesium oxide-functionalized coaxial fibre membranes enhance the recruitment of bone marrow mesenchymal stem cells and promote bone regeneration. BMC Biotechnol. 2025;25(1):80. doi:10.1186/s12896-025-01017-w
91. Wang X, Zhang S, Zhu A, et al. Black phosphorus and E7-Functionalized sulfonated polyetheretherketone with effective osteogenicity and antibacterial activity. Engineering. 2025;46:147–161. doi:10.1016/j.eng.2024.07.019
92. He Y, Mu C, Shen X, et al. Peptide LL-37 coating on micro-structured titanium implants to facilitate bone formation in vivo via mesenchymal stem cell recruitment. Acta Biomater. 2018;80:412–424. doi:10.1016/j.actbio.2018.09.036
93. Duchamp De Lageneste O, Julien A, Abou-Khalil R, et al. Periosteum contains skeletal stem cells with high bone regenerative potential controlled by Periostin. Nat Commun. 2018;9(1):773. doi:10.1038/s41467-018-03124-z
94. Trappmann B, Baker BM, Polacheck WJ, et al. Matrix degradability controls multicellularity of 3D cell migration. Nat Commun. 2017;8(1):371. doi:10.1038/s41467-017-00418-6
95. Wang C, Ying J, Nie X, et al. Targeting angiogenesis for fracture nonunion treatment in inflammatory disease. Bone Res. 2021;9(1):29. doi:10.1038/s41413-021-00150-4
96. He W, Wang S, He Z, et al. Injectable osteogenic hydrogel based on antibiotic recognition for infected radial bone defect regeneration. Mater Des. 2024;241:112904. doi:10.1016/j.matdes.2024.112904
97. Hestehave Pedersen R, Rasmussen M, Overgaard S, et al. Effects of P-15 Peptide Coated Hydroxyapatite on Tibial Defect Repair In Vivo in Normal and Osteoporotic Rats. Biomed Res Int. 2015;2015:253858. doi:10.1155/2015/253858
98. Chen H, Wei L, Yu Q, et al. Injectable hydrogels for osteomyelitis treatment induce metabolic reprogramming for protection against reinfection. Nat Commun. 2026;17(1):1613. doi:10.1038/s41467-026-68318-2
99. Choe H, Narayanan AS, Gandhi DA, et al. Immunomodulatory peptide IDR-1018 decreases implant infection and preserves osseointegration. Clin Orthop Relat Res. 2015;473(9):2898–2907. doi:10.1007/s11999-015-4301-2
100. Xu H, Feng J, Dai N, et al. Self-assembling peptide hydrogel scaffold integrating stem cell-derived exosomes for infected bone defects. J Biomater Sci Polym Ed. 2024;35(10):1511–1522. doi:10.1080/09205063.2024.2336316
101. Lin X, Deng S, Fu T, et al. Hyaluronic acid-based hydrogel microspheres with multi-responsive properties for antibacterial therapy and bone regeneration in Staphylococcus aureus-infected skull defects. Mater Today Bio. 2025;32:101676. doi:10.1016/j.mtbio.2025.101676
102. Su S, Xie J, Gao J, et al. A bone adhesive enhances osteoporotic fracture repair by regulating bone homeostasis. NPG Asia Mater. 2024;16(1):21. doi:10.1038/s41427-024-00539-7
103. Ambrosi TH, Marecic O, Mcardle A, et al. Aged skeletal stem cells generate an inflammatory degenerative niche. Nature. 2021;597(7875):256–262. doi:10.1038/s41586-021-03795-7
104. Zhang Y, Xu J, Ruan YC, et al. Implant-derived magnesium induces local neuronal production of CGRP to improve bone-fracture healing in rats. Nat Med. 2016;22(10):1160–1169. doi:10.1038/nm.4162
105. Wang Q, Chen Y, Ding H, et al. Optogenetic activation of mechanical nociceptions to enhance implant osseointegration. Nat Commun. 2025;16(1):3093. doi:10.1038/s41467-025-58336-x
106. Wang Y, Hu Y, Lan S, et al. A recombinant parathyroid hormone-related peptide locally applied in osteoporotic bone defect. Adv Sci. 2023;10(22):e2300516. doi:10.1002/advs.202300516
107. Feng Q, Li H, Wang Y, et al. An injectable antioxidant parathyroid hormone related supramolecular peptide nanofiber hydrogel for repairing osteoporotic bone defects. ACS Appl Mater Interfaces. 2025;17(27):38890–38911. doi:10.1021/acsami.5c06674
108. Chen R, Wang J, Chen L, et al. Electrospunned nanofiber membranes of PTH-related peptide loaded biopolymers for osteoporotic bone defect repair. Mater Des. 2024;244:113179. doi:10.1016/j.matdes.2024.113179
109. Qin H, Guan Z, Wang Y, et al. An injectable phosphocreatine-grafted hydrogel incorporating hierarchically structured teriparatide/SrZnP-functionalized Zn-Cu particles for osteogenesis-angiogenesis coupling and osteoporotic bone regeneration. Mater Today Bio. 2025;35:102272. doi:10.1016/j.mtbio.2025.102272
110. Wang Y, Pan Z, Wang Q, et al. Sequential SDF-1/CGRP-releasing smart composite hydrogel promotes osteoporotic fracture healing by targeting sensory nerve-regulated bone remodeling. Mater Today Bio. 2025;32:101750. doi:10.1016/j.mtbio.2025.101750
111. Darnell M, Mooney DJ. Leveraging advances in biology to design biomaterials. Nature Mater. 2017;16(12):1178–1185. doi:10.1038/nmat4991
112. Koons GL, Diba M, Mikos AG. Materials design for bone-tissue engineering. Nature Rev Mater. 2020;5(8):584–603.
113. Gaharwar AK, Singh I, Khademhosseini A. Engineered biomaterials for in situ tissue regeneration. Nature Rev Mater. 2020;5(9):686–705. doi:10.1038/s41578-020-0209-x
114. Wojda SJ, Marozas IA, Anseth KS, et al. Impact of release kinetics on efficacy of locally delivered parathyroid hormone for bone regeneration applications. Tissue Eng Part A. 2021;27(3–4):246–255. doi:10.1089/ten.tea.2020.0119
115. Hu X, Roy SR, Jin C, et al. Control cell migration by engineering integrin ligand assembly. Nat Commun. 2022;13(1):5002. doi:10.1038/s41467-022-32686-2
116. Englund EA, Wang D, Fujigaki H, et al. Programmable multivalent display of receptor ligands using peptide nucleic acid nanoscaffolds. Nat Commun. 2012;3(1):614. doi:10.1038/ncomms1629
117. Wang T, Bai J, Lu M, et al. Engineering immunomodulatory and osteoinductive implant surfaces via mussel adhesion-mediated ion coordination and molecular clicking. Nat Commun. 2022;13(1):160. doi:10.1038/s41467-021-27816-1
118. Ge C, Li Y, Wu F, et al. Synthetic peptides activating discoidin domain receptor 2 and collagen-binding integrins cooperate to stimulate osteoblast differentiation of skeletal progenitor cells. Acta Biomater. 2023;166:109–118. doi:10.1016/j.actbio.2023.05.039
119. Sun Y, Yang X, Liu Y, et al. Adaptive cationic peptide bundles hydrogel for the repair of infected mandibular defects. Biomaterials. 2026;325:123613. doi:10.1016/j.biomaterials.2025.123613
120. Vázquez Torres S, Leung PJY, Venkatesh P, et al. De novo design of high-affinity binders of bioactive helical peptides. Nature. 2024;626(7998):435–442. doi:10.1038/s41586-023-06953-1
121. Liu H, Song Z, Zhang Y, et al. De novo design of self-assembling peptides with antimicrobial activity guided by deep learning. Nat Mater. 2025;24(8):1295–1306. doi:10.1038/s41563-025-02164-3
122. Ma S, Liu X, Xiao B, et al. An artificial intelligence-selected multifunctional oligopeptide coated Biphasic Calcium Phosphate (BCP) scaffold promotes bone defect repair. Adv Healthc Mater. 2026;15:e05458. doi:10.1002/adhm.202505458
123. Chen Y, Zhang Q, Yang S, et al. Formulate adaptive biphasic scaffold via sequential protein-instructed peptide co-assembly. Adv Sci. 2024;11(29):e2401478. doi:10.1002/advs.202401478
124. Dai K, Geng Z, Zhang W, et al. Biomaterial design for regenerating aged bone: materiobiological advances and paradigmatic shifts. Natl Sci Rev. 2024;11(5):nwae076. doi:10.1093/nsr/nwae076
125. Banu SA, Manjusha KM, Mamachan M, et al. From flat bones to functional healing: redefining rodent calvarial defect models for translational bone regeneration research. Ann Med Surg. 2025;87(10):6382–6385. doi:10.1097/MS9.0000000000003528
126. Song P, Zhou D, Wang F, et al. Programmable biomaterials for bone regeneration. Mater Today Bio. 2024;29:101296. doi:10.1016/j.mtbio.2024.101296
127. Clark JR, Al Maruf DSA, Tomaskovic-Crook E, et al. Mechanobiologically-optimized non-resorbable artificial bone for patient-matched scaffold-guided bone regeneration. Nat Commun. 2025;16(1):9422. doi:10.1038/s41467-025-64466-z
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.