Back to Journals » International Journal of Nanomedicine » Volume 20
Macrophage Membrane-Coated Nanocarriers in Myocardial Infarction: A Paradigm Shift in Targeted Cardiac Therapy
Authors Wang L, Wang K, Zheng H
Received 13 June 2025
Accepted for publication 1 October 2025
Published 7 November 2025 Volume 2025:20 Pages 13499—13525
DOI https://doi.org/10.2147/IJN.S546817
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Jie Huang
Liping Wang,1 Kaili Wang,2 Hongjian Zheng2
1Department of Cardiology, Chun’an First People’s Hospital, Hangzhou, Zhejiang Province, 311700, People’s Republic of China; 2Nursing Department, Chun’an First People’s Hospital, Hangzhou, Zhejiang Province, 311700, People’s Republic of China
Correspondence: Hongjian Zheng, Nursing Department, Chun’an First People’s Hospital, No. 1869 Huanhubei Road, Hangzhou, Zhejiang Province, 311700, People’s Republic of China, Email [email protected]
Abstract: Myocardial infarction remains a major contributor to global morbidity and mortality, with current therapeutic strategies often falling short in addressing post-infarction inflammation, fibrotic remodeling, and suboptimal drug localization. Macrophages are key modulators of the cardiac immune microenvironment, play a dual role in tissue damage and repair by polarizing into distinct inflammatory and reparative phenotypes. Recent advancements in biomimetic nanotechnology have facilitated the development of macrophage membrane-coated nanocarrier engineered systems that replicate the functional surface characteristics of native macrophages. These nanocarriers offer enhanced therapeutic precision by enabling immune evasion, targeted delivery to infarcted myocardium, and sustained release of bioactive agents. Their prolonged systemic circulation further augments therapeutic efficacy. Current clinical strategies remain insufficient in preventing long-term complications such as adverse cardiac remodeling and the development of heart failure, highlighting a critical need for targeted and effective post-MI therapies. This review comprehensively evaluates the biological role of macrophages in the context of myocardial infarction and highlights current innovations in the fabrication and functional optimization of macrophage membrane-coated nanocarriers. We further discuss their mechanisms of action, therapeutic benefits demonstrated in preclinical models, and their prospective integration into regenerative treatment paradigms. Critical challenges, including regulatory approval, manufacturing scalability, and clinical translation, are also discussed. These nanocarriers represent a promising frontier in myocardial infarction therapy, offering targeted, biocompatible, and immune-responsive drug delivery platforms.
Keywords: macrophage membrane-coated nanocarriers, myocardial infarction therapy, biomimetic drug delivery systems, cardiac tissue regeneration, immune modulation in heart repair, targeted nanomedicine, translational nanotechnology
Introduction
Acute myocardial infarction (AMI) is characterized by irreversible ischemic necrosis of cardiomyocytes, primarily due to the abrupt cessation of coronary blood flow following thrombotic occlusion of an atherosclerotic artery.1 This process is initiated by plaque rupture or erosion, leading to platelet aggregation and intraluminal thrombus formation.2 Resultant oxygen deprivation triggers anaerobic metabolism, ATP depletion, and ultimately, myocyte death. The extent of myocardial injury depends on the duration and severity of ischemia, with transmural infarction representing the most severe form.3 A substantial proportion of myocardial infarction (MI) cases present without typical anginal symptoms, a clinical phenomenon termed “silent” myocardial ischemia.4 Myocardial infarction arises from the interplay of modifiable (eg, hypertension, diabetes, smoking) and non-modifiable (eg, age, chronic kidney disease) risk factors that drive endothelial dysfunction, atherosclerotic plaque progression, and inflammation-induced thrombosis, ultimately leading to coronary occlusion.5,6
Myocardial infarction is diagnosed through electrocardiographic changes and elevated cardiac biomarkers, primarily troponin and CK-MB. ECG differentiates STEMI from NSTEMI, guiding acute management strategies such as antiplatelet therapy, vasodilators, and supplemental oxygen.7 STEMI requires prompt reperfusion via primary PCI or, if unavailable, thrombolysis. PCI provides mechanical revascularization; thrombolysis enables pharmacologic clot dissolution. Treatment choice depends on ischemia duration, clinical status, and facility access, with both aiming to restore perfusion and minimize infarct size.8
PCI is indicated in high-risk patients with significant coronary artery disease to restore blood flow and prevent infarction.9 In diabetic patients with multivessel disease, CABG is preferred over PCI for more complete and durable revascularization.10,11 Even though these techniques dramatically lower the patient death rate,12 problems such hemorrhage, ischemia reperfusion damage, and coronary restenosis13,14 might happen at any time. New therapies focusing on angiogenesis, inflammation, and fibrosis after myocardial infarction aim to preserve cardiac function and reduce progression to heart failure.15,16 Ischemic hypoxia triggers cardiomyocyte apoptosis within hours to days post-MI, initiating inflammatory cell infiltration and cytokine release. This response drives granulation tissue formation via extracellular matrix deposition and activates reparative as well as maladaptive remodeling.17
To provide historical context, early pioneering work by Hu et al. (2011) demonstrated erythrocyte membrane-coated polymeric nanoparticles with extended circulation times, laying the foundation for the field of biomimetic nanocarriers.18 Recent advances in nanomedicine have introduced non-biomimetic platforms, including liposomes, polymeric nanoparticles, and inorganic carriers for myocardial infarction therapy, primarily aimed at enhancing pharmacokinetics and site-specific drug accumulation. Despite these benefits, such systems often encounter rapid clearance, limited myocardial targeting, and off-target immune activation. In contrast, macrophage membrane-coated nanocarriers (MM-NCs) exhibit superior biological interface compatibility by leveraging surface proteins that facilitate immune evasion, inflammation-responsive homing, and prolonged systemic retention.19,20 Moreover, understanding the temporal progression of myocardial infarction is crucial to optimizing therapeutic interventions. Clinically, myocardial infarction progresses through distinct temporal phases: an acute phase (hours to days) characterized by ischemia-reperfusion injury and inflammation, and a chronic phase (weeks to months) dominated by fibrotic remodeling and ventricular dilation. MM-NCs offer phase-specific therapeutic integration acutely, as modulators of post-reperfusion inflammation, and chronically, to attenuate maladaptive remodeling and promote cardiac repair. These attributes position MM-NCs as promising candidates to complement existing MI management protocols across its clinical timeline.21,22
Pathological Characteristics of Myocardial Ischemia and Infarction
The pathological definition of myocardial infarction centers on cardiomyocyte death caused by prolonged ischemia. Figure 1 depicts a schematic representation of the roles of cardiomyocytes, fibroblasts, immune cells, and endothelial subtypes in the progression and remodeling of the failing heart. Myocardial ischemia triggers ultrastructural changes within 10–15 minutes, including sarcolemmal disruption and mitochondrial swelling, visible via electron microscopy. While animal models show biochemical evidence of cell death within minutes, human myocardial necrosis develops over hours, progressing from subendocardial to transmural regions. This timeline can be altered by ischemic preconditioning, reduced oxygen demand, or collateral circulation. Modern diagnostics detect these changes through sensitive biomarkers and advanced imaging, identifying even minimal injury.23
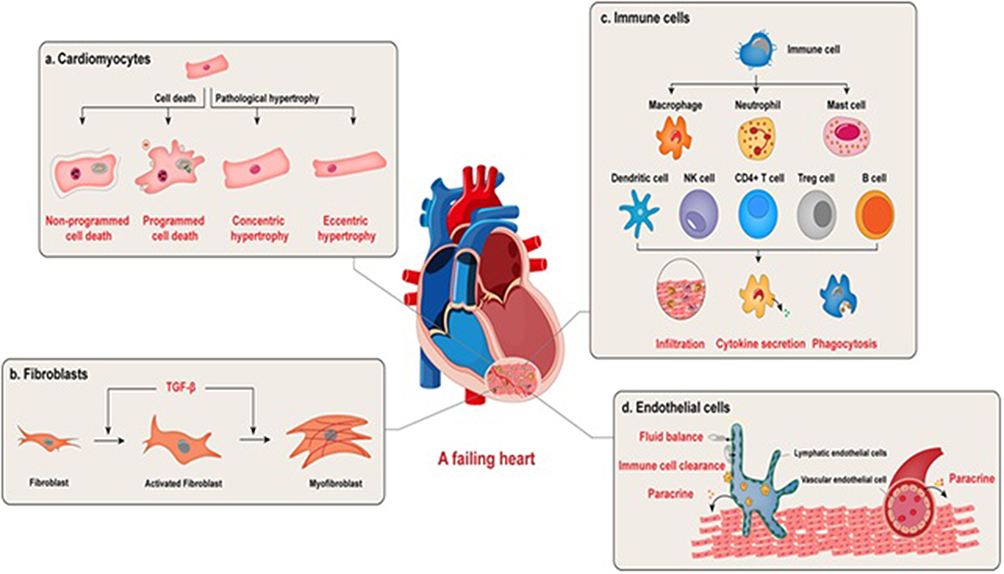 |
Figure 1 Cellular contributors to the pathophysiology of heart failure: Cardiomyocytes respond to pathological stimuli through maladaptive hypertrophy or undergo cell death via necrotic or apoptotic mechanisms, contributing to contractile dysfunction and loss of viable myocardium. Fibroblasts, upon activation, differentiate into myofibroblasts and drive cardiac fibrosis, a hallmark of structural remodeling. The immune compartment, including infiltrating leukocytes, modulates myocardial injury and repair by secreting inflammatory cytokines, clearing cellular debris, and influencing the process of regeneration. Endothelial cells also play essential roles: vascular endothelial cells modulate cardiac function through paracrine signaling to adjacent cells, whereas lymphatic endothelial cells facilitate cardiac repair post-infarction by maintaining interstitial fluid homeostasis, promoting immune cell clearance, and secreting regenerative mediators. Adapted from He X, Du T, Long T, Liao X, Dong Y, Huang ZP. Signaling cascades in the failing heart and emerging therapeutic strategies. Signal Transduct Target Ther. 2022 Apr 23;7(1):134. © The Author(s) 2022.24 under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/). |
Introduction to Macrophage Membrane-Coated Nanocarriers as an Innovative Solution
Traditional drug delivery systems face significant limitations, including systemic toxicity, nonspecific distribution, rapid clearance, poor bioavailability, and delayed therapeutic onset. To address these challenges in myocardial infarction treatment, targeted drug delivery to the ischemic myocardium has emerged as a critical therapeutic strategy. Recent advances highlight biomimetic nanoparticle platforms as a promising approach, offering enhanced precision, prolonged circulation, and improved therapeutic efficacy for cardiac repair.25 Biomimetic nanoparticles inherit special biofunctions from the original cells, including long circulation times, active targeting capabilities, and high biocompatibility, while retaining the beneficial physical characteristics of conventional nanoparticles. This is achieved by fusing bioactive cell components onto the surfaces of synthetic nanoparticles.18,26 Figure 2 depicts cell membrane-coated nanoparticles combine customizable synthetic cores with biologically derived membrane coatings, enabling tailored functionalities for targeted biomedical applications. Red blood cell membrane-coated nanoparticles, for instance, have a half-life of up to 40 hours and can improve blood circulation. 10 Porous silicon nanoparticles coated with leukocyte membranes show effective accumulation at tumor locations and extended circulation durations.27 Nanoparticles enclosed in neutrophil membranes can alleviate inflammatory arthritis.28 Furthermore, injured blood arteries can be selectively adhered to by platelet membrane-coated nanoparticles.29
 |
Figure 2 Cell membrane-coated nanoparticles combine customizable synthetic cores with biologically functionalized coatings, where core material selection depends on application needs while membrane wrappings (eg, from RBCs, WBCs, or stem cells) confer source cell-specific properties. This modular design enables tailored biomedical functionality, from targeted drug delivery to immune modulation. Adapted from Krishnan N, Fang RH, Zhang L. Cell membrane-coated nanoparticles for the treatment of cancer. Clin Transl Med. 2023 Jun;13(6):e1285. © 2023 The Authors. Clinical and Translational Medicine published by John Wiley & Sons Australia, Ltd on behalf of Shanghai Institute of Clinical Bioinformatics,30 under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/). |
Macrophages, as an immune cells, are the main mediators of inflammation and tissue repair.31 Figure 3 represents schematic overview of macrophage crosstalk with neutrophils, eosinophils, basophils, and T cells regulating inflammatory resolution and reparative polarization post-MI. In different phases of myocardial infraction, macrophages have been involved.32,33 The infarcted myocardium exhibits rapid recruitment of inflammatory monocytes within 30 minutes post-MI, which subsequently differentiate into macrophages and localize predominantly within the ischemic core and margin zones.34 These macrophages have very vital role in promoting cardiac repair and for removing necrotic cellular debris.35,36 Adhesion molecules, such as PSGL-1 and LAF-1 facilitates the transfer of monocyte-derived macrophages.37 Macrophage membranes (MM) are potential candidates to target injured myocardium because these intrinsic surface proteins bind to vascular cell adhesion molecules-1 (ICAM-1, VCAM-1), which are substantially increased in damaged endothelial cells during MI27. Additionally, by helping polymer nanoparticles avoid immune clearance, this MM coating technique may improve their biocompatibility.
 |
Figure 3 Immunoregulatory interactions between macrophages and immune cells in the infarcted myocardium. Neutrophil-derived extracellular DNA promotes macrophage polarization through activation of the toll-like receptor 9 (TLR9) pathway. Additionally, neutrophil secretion of neutrophil gelatinase-associated lipocalin (NGAL) enhances macrophage efferocytosis capacity by upregulating the Mer tyrosine kinase (MertK) receptor. In turn, macrophages limit excessive neutrophil infiltration by releasing matrix metalloproteinase-12 (MMP-12). Crosstalk with adaptive immune cells is also critical: regulatory T cells (Tregs) secrete interleukin-35 (IL-35), which triggers glycoprotein 130 (GP130)-dependent signaling via interleukin-12 receptor beta 2 (IL-12Rβ2) in macrophages, resulting in phosphorylation of signal transducer and activator of transcription 1 and 4 (STAT1 and STAT4), and subsequent upregulation of C-X3-C motif chemokine receptor 1 (CX3CR1) and transforming growth factor-beta 1 (TGF-β1). Furthermore, macrophage-derived interleukin-5 (IL-5) promotes eosinophil accumulation, which drives macrophage phenotypic switching through the interleukin-4/signal transducer and activator of transcription 6 (IL-4/STAT6) axis. Basophils also contribute to alternative macrophage activation by secreting interleukin-4 (IL-4) and interleukin-13 (IL-13). Collectively, these interactions orchestrate the balance between pro-inflammatory and reparative macrophage states during myocardial infarction healing. Adapted from Jian Y, Zhou X, Shan W et al. Crosstalk between macrophages and cardiac cells after myocardial infarction. Cell Commun Signal. 2023 May 11;21(1):109. © The Author(s) 2023,38 under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/). |
Certain receptors, such as CXCR4, CXCR2, interleukin (IL)-1R, and tumor necrosis factor (TNF)-R, are found in the membrane of macrophages. Consequently, nanoparticles coated with macrophage membranes exhibit biomimetic properties similar to those of macrophages found in nature.39,40 Macrophage membrane-coated VEGF sustained-release PLGA nanoparticles can migrate more specifically towards the inflammatory site when CXCR2 and CXCR4 receptors are present,39,40 and.41 This allows the particles to accumulate in the infarcted myocardium and guarantees that VEGF is continuously released in the infarcted area, which promotes neovascularization. The enhanced biosafety profile of biomimetic membrane-coated nanoparticles is one of their most convincing benefits over alternative VEGF delivery techniques to the infarcted myocardium.42 Conventional physical methods needed the modification of paramagnetic nanoparticles (Fe3O4) in order to concentrate them in the infarcted myocardium via magnetic targeting.43 There is a chance of further cardiac injury because this technique has been linked to myocardial iron overload.44 In contrast to immunoliposome-based delivery systems, biomimetic membrane-coated PLGA nanoparticles offer superior sustained-release capabilities while maintaining targeted delivery properties.45
According to reports, when macrophages are exposed to endotoxins like LPS, they begin secreting many cytokines, including interleukins and TNFα, within 4 to 5 hours.46 Based on findings it is suggested that the duration of lipopolysaccharide (LPS) induction was systematically varied to optimize membrane-bound tumor necrosis factor-alpha (TNFα) expression on macrophage surfaces. These modified membranes demonstrated time-dependent antiproliferative effects against HeLa, MCF-7, and MDA-MB-231 cancer cell lines, confirming preserved bioactivity. Subsequent mechanical extrusion of the LPS-activated macrophage membranes produced functional nanovesicles for targeted therapeutic applications.47
Role of Macrophages in Inflammation and Repair After MI
Infarct macrophages orchestrate debris clearance via phagocytosis (necrotic material/matrix fragments) and efferocytosis (apoptotic cells). This dual function resolves inflammation while promoting repair, with efferocytosis additionally suppressing pro-inflammatory cascade. In vivo studies demonstrate infarct macrophages actively phagocytose dying cardiomyocytes and neutrophils, confirming their essential role in debris clearance. Impaired phagocytic capacity disrupts inflammation resolution and tissue repair, directly linking macrophage function to cardiac recovery post-infarction.48–50
Infarct macrophages adopt reparative M2 phenotypes through efferocytosis-induced anti-inflammatory signaling, lymphocyte/neutrophil crosstalk via contact or soluble factors, and neutrophil-derived NGAL that enhances efferocytosis. This multilevel regulation ensures timely inflammation resolution and tissue repair.51 The recruitment of anti-inflammatory T cell subsets particularly regulatory T cells (Tregs), further promotes macrophage polarization toward a reparative phenotype through multiple mechanisms: paracrine IL-10 signaling and direct cell-contact-mediated interactions. These coordinated immunomodulatory actions enhance the transition to an M2-like macrophage state critical for cardiac repair.52 Cytokine activation in the infarct microenvironment triggers infarct macrophages to generate endogenous anti-inflammatory signals that block proinflammatory pathways. The activation of IL-1 receptor-associated kinase (IRAK)-M by infarct macrophages has been demonstrated to prevent inflammatory activity induced by TLRs and IL-1, protecting the infarcted heart from damaging remodeling.53 Transforming growth factor (TGF)-bs, IL-4, IL-10, and IL-13 are among the anti-inflammatory cytokines that are induced by the infarct and collectively prevent proinflammatory macrophage activation.54 Both increased de novo production55 and activation of latent reserves56 in the infarcted heart activate TGF-b signaling cascades, which may be crucial in mediating the anti-inflammatory effects of macrophages.57–59 It was discovered that TGF-b activates Smad3 signaling but not Smad2 signaling, which results in anti-inflammatory effects on infarct macrophages.60,61 The modulation of transcription factors belonging to the IRF family may be one way whereby secreted mediators affect the phenotypic and functional character of infarct macrophages.62
Monocyte/Macrophage Response After MI
During the acute phase of myocardial ischemia, neutrophils rapidly infiltrate the infarcted myocardium through a well-orchestrated adhesion cascade, achieving maximal accumulation within 24 hours post-occlusion. This recruitment is mechanistically governed by selectin-mediated rolling (L- and P-selectin), firm adhesion via intercellular adhesion molecule-1 (ICAM-1), and chemotactic gradient formation by interleukin-8 (CXCL8) and CXCL1, which collectively guide neutrophils from the vasculature into the ischemic tissue.63 The post-infarct cellular infiltrate is predominantly composed of monocytes and their macrophage derivatives. Extensive studies in both rodent and large animal models have characterized the temporal dynamics, persistence, and functional significance of these phagocytic populations in cardiac repair and remodeling processes.64,65 Studies demonstrate that monocytes and macrophages play pivotal roles in orchestrating infarct healing and regulating cellular recruitment during the initial 14-day post-MI period. Monocyte infiltration into the infarct zone depends critically on the MCP-1/CCR2 chemokine axis, as evidenced by impaired recruitment in germline CCR2/MCP-1 knockout models. Additionally, upregulated expression of adhesion molecules - particularly vascular cell adhesion molecule 1 (VCAM-1) on activated endothelium facilitates leukocyte extravasation into ischemic myocardium.64,66 VCAM-1-mediated monocyte recruitment via VLA-4 binding paradoxically worsens ischemia-reperfusion injury, as activated neutrophils and macrophages release cytotoxic ROS and proteases that injure viable cardiomyocytes. Preclinical evidence confirms that modulating this inflammatory response reduces infarct expansion (measured by infarct size/area-at-risk ratio), highlighting the therapeutic potential of targeted anti-inflammatory approaches in reperfusion injury.67,68
The clinical translatability of these approaches remains unestablished, with an unclear role for reparative monocytes in injury modulation. Non-reperfused MI models show dichotomous results monocyte/macrophage infiltration associates with both improved healing (via tissue remodeling) and worsened dysfunction (through inflammatory exacerbation).64,69–72 Effective infarct healing demands balanced monocyte/macrophage activity, merging inflammation and repair, particularly in unreperfused MI, where scar formation requires phagocyte-driven clearance. Excessive suppression or amplification of this response impairs remodeling, mirroring macrophages’ dual roles in skin wounds. Thus, therapeutic strategies must preserve this equilibrium to avoid disrupting the phased transition from debris removal to fibrosis:73–75 Proteases like matrix metalloproteinases,76–78 urokinase-type plasminogen activator, and cathepsins are released by the cells, which break down the preexisting collagen network and promote cell movement;76 phagocytose apoptotic and necrotic myocytes and neutrophils and other debris;79 release inflammatory mediators like inducible nitrous oxide synthase, reactive oxygen species, interferon, TNF-, IL-1, IL-6, and macrophage inflammatory protein 1;80 promote angiogenesis through the secretion of vascular endothelial growth factor and fibroblast growth factor;81 transport reparative enzymes and prosurvival factors like transglutaminases; stimulate collagen synthesis and deposition by myofibroblasts through the release of transforming growth factor and fibroblast growth factor.63,82
Nanofibers serve as versatile scaffolds in regenerative medicine, particularly for skin, bone, vascular, and nerve tissues, due to their highly porous and interconnected architecture. This unique structure facilitates efficient nutrient exchange, waste removal, and cellular communication while mimicking the natural extracellular matrix to guide tissue regeneration. Their adaptable composition and tunable physical properties allow customization for diverse therapeutic applications.83 Unlike solid Macrophage biology and its role in MI.
M1 (Pro-Inflammatory) vs M2 (Anti-Inflammatory) Macrophage
In order to promote an inflammatory response in the wounded tissues, macrophages or monocytes first polarize into a pro-inflammatory (M1) phenotype and then change into an anti-inflammatory (M2) phenotype for tissue regeneration.84,85 The uncontrolled inflammation caused by the M1 phenotype results in a severe foreign body reaction, where macrophages aggregate to form massive cells called foreign body granuloma with fibrous encapsulation.86,87 Biomaterials can guide post-infarct healing by modulating macrophage polarization through engineered physical (stiffness, topography) and biochemical (cytokine release, ECM mimicry) properties. These precisely designed matrices promote regenerative M2 phenotypes while suppressing damaging inflammation, shifting from passive scaffolds to active immunomodulatory platforms that orchestrate cardiac repair.88,89 Biomaterial design critically influences macrophage polarization through specific physical cues: implants with sharp edges, high stiffness, moderate roughness, and small-scale topography (nanofibers/micropores) promote pro-inflammatory M1 responses, while smooth surfaces, low stiffness, high roughness, and macro-scale architecture (large fibers/pores) favor anti-inflammatory M2 phenotypes. These structural parameters enable targeted immunomodulation to control foreign body reactions and tissue repair processes.90 While early approaches focused on immunosuppression, current evidence highlights that a controlled, transient inflammatory response is essential for effective myocardial regeneration. Given the critical role of M1-to-M2 transitions in post-MI healing, targeted delivery systems capable of modulating this shift while minimizing systemic toxicity are urgently needed this is where macrophage membrane-coated nanocarriers offer significant promise. However, recent evidence highlights that controlled early inflammation is indispensable for effective tissue regeneration, shifting the paradigm toward immunomodulatory biomaterial design rather than immunosuppression polycaprolactone (PCL) implants, nanofibrous PCL scaffolds demonstrate superior biocompatibility, as evidenced by host cell infiltration in murine subcutaneous models. This structural advantage promotes a more favorable tissue response through improved biomaterial-tissue integration.91 Electrospun nanofibers enable bioactive molecule delivery but their hydrophobicity limits cell adhesion, risking fibrosis. Surface modifications can enhance biocompatibility while preserving its function.92,93 Peptides, intercellular adhesion molecule-1, epidermal growth factors, antigen-mimetic GM3-lactone molecules, granulocyte colony-stimulating factors, and proteins (EphA5-Fc and ephrin A5-Fc) have all been functionalized into the nanofibers to improve cellular interactions.94–98 Additionally, immunoregulatory substances like mechano-growth factor (MGF) and interleukin-4 (IL-4) can be added to the exterior of electrospun nanofibers to encourage macrophages to transition to the M2 phenotype.99,100 To prevent protein denaturation during physical and chemical processes, nanofibrous scaffolds have been coated with synthetic lipid bilayers.101 According to a recent study, macrophages are promoted to the M2 phenotype by PCL scaffolds functionalized with mesenchymal stromal exosomes.102 PCL/nano-hydroxyapatite-incorporated chitin-derived hydrogels loaded with mesenchymal stem cells induce macrophages to adopt an M2 phenotype, which improves bone regeneration.103
The Role of M1 and M2 Macrophages in MI
Macrophage polarization critically regulates wound healing and tissue regeneration across physiological contexts.104 Despite incomplete understanding of polarization pathways, macrophage modulation represents a promising therapeutic strategy for diseases requiring tissue regeneration.105–107 MI represents a prime candidate for macrophage polarization therapy, as temporal control of M1-to-M2 transition could simultaneously resolve inflammation and promote myocardial regeneration.108 Cardiac macrophages exhibit transcriptional and functional heterogeneity beyond M1/M2 dichotomies, with infarcted hearts containing distinct subsets (eg, CCR2+ vs TIMD4+ macrophages) that differentially contribute to inflammation, clearance, and fibrosis.109
According to mouse models, cardiac resident macrophages make up 6–8% of non-cardiomyocytes and come from two different lineages: fetal monocytes in the postnatal phase and erythromyeloid progenitors in the yolk sac during early embryogenesis.33,110 The expression of CCR2 distinguishes these two groups of macrophages from one another. Therefore, macrophages generated from the yolk sac are CCR2-, while macrophages derived from monocytes are CCR2+.110 CCR2-macrophages are mainly found in the heart’s myocardium and are renewed by self-renewal, whereas CCR2+ macrophages are mainly concentrated in the endocardium and are restored by the recruitment of circulating monocytes.111,112 Based on the expression of MHC-II and Ly6C, CCR2-macrophages are further divided into three subsets: MHC-IIhigh, MHC-IIlow, and Ly6C+.113 Overall, resident cardiac macrophages can be divided into four subsets: CCR2-MHC-IIhigh, CCR2-MHC-IIlow, CCR2-Ly6C+, and CCR2+ macrophages, based on the surface markers described. However, a study conducted recently by Dick and associates has offered a rather different explanation of the local cardiac macrophage morphologies. The scientists used genetic fate mapping, long-term parabiosis studies, and single-cell RNA sequencing in mice to identify four subsets of macrophages in an adult heart: TIMD4+LYVE1+MHC-IIlowCCR2– macrophages, TIMD4−LYVE1–MHC-IIhighCCR2 macrophages, and two CCR2+MHC-IIhigh subsets. Blood monocytes are the only ones that can restore CCR2+MHC-IIhigh subsets, and circulating monocytes may partially replace TIMD4−LYVE1–MHC-IIhighCCR2− macrophages. It was shown that TIMD4+LYVE1+MHC-IIlowCCR2– macrophages are maintained by local proliferation distinct from circulating monocytes.36
Since animals with lower levels of macrophages exhibit poorer wound recovery, macrophages are most likely the primary source of growth factors needed for scar formation.114–120 By phagocytosing necrotic cells and releasing angiogenic and growth factors, macrophages in the post-MI LV primarily aid in wound repair. Macrophages travel in response to signals from the injured myocardium, including those from resident cells (myocytes) and acute inflammatory cells (neutrophils). The local production of adhesion molecules, chemo attractants, and other proteins attracts macrophages to the injury site and the surrounding border zone.121 In turn, activated macrophages generate a variety of cytokines, chemokines, and proteases, including MMPs.122,123
As master regulators of post-MI inflammation, macrophages dictate clinical outcomes through phased activities: early debris clearance (preventing secondary necrosis) and late matrix remodeling (preventing wall thinning). Therapeutic strategies now target their polarization dynamics rather than global suppression.124 Reperfusion accelerates and amplifies the inflammatory response in ischemic tissue, yet paradoxically enhances healing outcomes. Early clinical trials demonstrated that anti-inflammatory therapy with methylprednisolone increased mortality due to left ventricular rupture, underscoring the essential role of inflammation in post-ischemic repair.125 Figure 4 represents key features and roles of M1 versus M2 macrophage polarization during myocardial infarction-related inflammation and tissue repair.
 |
Figure 4 Functional roles and phenotypic transitions of macrophages in myocardial injury and repair. Following cardiomyocyte damage, peripheral monocytes are recruited to the injured myocardium, where they differentiate into pro-inflammatory M1 macrophages. These macrophages activate key signaling pathways, including the C-C motif chemokine ligand 2 (CCL2) and nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) pathways, resulting in elevated secretion of tumor necrosis factor-alpha (TNF-α), interleukin-1 beta (IL-1β), and interleukin-6 (IL-6). This pro-inflammatory response exacerbates cardiomyocyte necrosis and hinders tissue repair. Over time, M1 macrophages undergo phenotypic switching to reparative M2 macrophages, a transition mediated in part by the cannabinoid receptor type 2 (CB2) signaling axis, which promotes resolution of inflammation and supports tissue remodeling. M2 macrophages suppress pro-inflammatory cytokines, facilitate tissue repair by promoting angiogenesis and fibroblast activation, and share functional characteristics with resident cardiac macrophages that support myocardial regeneration. Adapted from Chen G, Jiang H, Yao Y, Tao Z, Chen W, Huang F, Chen X. Macrophage, a potential targeted therapeutic immune cell for cardiomyopathy. Front Cell Dev Biol. 2022 Sep 30;10:908790. Copyright © 2022 Chen, Jiang, Yao, Tao, Chen, Huang and Chen.126 under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/). |
M-CSF administration post-MI enhances macrophage recruitment, improving functional recovery and accelerating infarct repair. Conversely, in cryoinjury models, clodronate liposome-mediated macrophage depletion impairs wound healing, underscoring their essential role in cardiac repair.71,127,128 Post-MI healing requires inflammatory chemotaxis: apoptotic neutrophils induce endothelial/monocytic adhesion molecule expression (ICAM-1, β2 integrins), enabling macrophage infiltration. These cells coordinate essential repair processes, underscoring inflammation’s dual role in injury and recovery.129 Activated macrophages secrete over 150 distinct proteins cytokines, chemokines, and growth factors, underscoring their central role in immune regulation and tissue remodeling.130 Among the proteins released are IL-1 α and β, IL-6, TNFα, and the macrophage inflammatory proteins 1α, 1β, 2α, and 2β 90.131 The release of growth factors and cytokines at the site of injury controls the fibroblast proliferation, angiogenesis, and inflammatory component.125
Enhancing Drug Delivery and Immune Evasion, Macrophage Membranes Coated with Nanocarriers
Macrophage membrane-coated nanomaterials inherit the biological functions of natural membranes, enhancing biocompatibility, immune evasion, and targeted delivery to inflammatory and malignant tissues. This biomimetic strategy has shown therapeutic potential across diverse pathologies, including cancer, infectious diseases, cardiovascular disorders, neurological conditions, and immune dysregulation. The preserved surface receptors such as integrins (eg, α4β1), MAC-1 (CD11b/CD18), and CSF1R enable nanoparticle homing to disease sites (eg, tumors, infected tissues) through innate recognition pathways.125
Atherosclerosis (AS) is a chronic inflammatory vasculopathy characterized by lipid-rich plaque accumulation within the arterial intima. This process is initiated when endothelial cells, stimulated by oxidized low-density lipoprotein (Ox-LDL), overexpress adhesion molecules (eg, VCAM-1, ICAM-1) and generate excessive reactive oxygen species (ROS), thereby recruiting immune cells (monocytes, T cells) that perpetuate vascular inflammation and plaque progression.132 Local inflammation contributes to arterial lumen narrowing and plaque destabilization, leading to stroke and other fatal cardiovascular events.133 Integrin α4 and β1, two crucial leukocyte receptors on the macrophage membrane, draw macrophages to atherosclerotic plaques by recognizing VCAM-1 on the surface of vascular endothelial cells.134,135 Fe3O4 is a typical magnetic resonance imaging (MRI) agent. It was discovered that Fe3O4 nanoparticles coated with a RAW264.7 membrane (Fe3O4@M) were successful in focusing on the first phases of AS.136 Fe3O4@Ms was more effective at imaging than Fe3O4@PEG and was safer for research items in vivo or in vitro when compared to the traditional MR agent. The molecular mechanism analysis demonstrated that macrophages’ upregulation of integrins α4 and β1 specifically identified VCAM-1 on endothelial cells. In order to target atherosclerotic lesions, Wang et al created a macrophage membrane coating on the surface of rapamycin-loaded PLGA (MM/RAPNPs).137
Fabrication of Macrophage Cell Membrane-Coated Systems
There are a few sequential procedures needed to fabricate MCM-coated NPs. This is accomplished by applying a nanovesicle formed from MCM to the previously constructed NP inner core. In order to create MCM-coated NPs, the preparation steps include 1) removing the outer membrane and isolating membrane vesicles, also known as “ghost cells”, 2) choosing and creating the NP core, and 3) fusing the MCM nanovesicles with the NP core via coextrusion, sonication, or electrostatic interaction.138,139 The source cells have been emptied and their intracellular contents extracted using a variety of delicate methods that cause cell lysis and disturb the cell structure.
These methods include the use of a Dounce homogenizer, sonication, extrusion, freeze-thaw cycles, and hypotonic lysis buffer. To preserve the cell membranes’ biofunctionality for fruitful biological interactions, these methods preserve the proteins that make up the surface membrane.138 To fabricate macrophage membrane-coated nanoparticles (MCM-NPs), the plasma membranes of isolated macrophages must be extracted. Unlike red blood cells, nucleated white blood cells require more complex protocols due to their intracellular components. Membrane isolation typically involves cell disruption methods such as hypotonic lysis and extrusion, followed by purification through differential centrifugation.140
Isolated membranes are physically extruded through polycarbonate membranes with defined pore sizes and then sonicated to form nanoscale vesicles that preserve the native proteolipid profile of MCMs. These vesicles are subsequently fused with nanoparticle (NP) cores pre-loaded with diagnostic or therapeutic payloads. A variety of core materials have been employed, including lipid-based NPs, gold NPs, Fe₃O₄ NPs, mesoporous silica NPs, upconversion NPs (UCNPs), and other inorganic platforms. Organic polymeric cores such as PLGA, albumin, and chitosan NPs have also been widely reported.141 In situ packaging of nanoparticles (NPs) via live-cell treatment enables the production of high-quality, membrane-wrapped NPs, preserving membrane integrity and surface protein functionality. For example, PLGA NPs loaded with rapamycin—an mTOR inhibitor—were encapsulated using macrophage-derived membranes to actively target atherosclerotic lesions. This dual-targeting strategy leverages macrophage surface α4β1 integrins and chemokine receptors, which bind VCAM-1 and inflammatory cytokines on activated endothelium, respectively. The resultant biomimetic NPs demonstrated effective in vivo accumulation in atherosclerotic plaques and reduced phagocytosis in vitro. Additionally, combining simvastatin with apolipoprotein A-I peptide (AP) enhanced foam cell cholesterol efflux and attenuated inflammatory cytokine expression, significantly reducing lesion formation. These findings underscore the therapeutic potential of macrophage membrane-coated nanoplatforms for targeted atherosclerosis diagnosis and treatment.142
Acute ischemic stroke, primarily caused by thrombotic occlusion of cerebral vessels, remains a leading cause of mortality globally. Therapeutic strategies aim to preserve neurons within the ischemic penumbra. A biomimetic nanosystem, Ma@(MnO2 + FTY), was developed by coating fingolimod-loaded manganese dioxide (MnO2) nanospheres with macrophage membranes to target inflamed brain regions. Leveraging macrophage-mediated inflammation-homing, the system exhibited enhanced biocompatibility, prolonged circulation, and selective accumulation in ischemic lesions. MnO2 scavenged excess ROS by converting H2O2 into O2, mitigating oxidative stress, while fingolimod promoted M1-to-M2 microglial polarization, shifting the proinflammatory microenvironment toward a neuroprotective state. This dual mechanism led to significant neuroprotection and lesion-targeting in vivo, positioning Ma@(MnO2 + FTY) as a promising synergistic therapy for acute ischemic stroke.143
Types of Core Nanocarriers
Lipid nanocarriers’ pharmacokinetics and biodistribution are dictated by their composition, size, surface charge, and steric stabilization. These factors influence plasma protein interactions, MPS clearance, and organ targeting. PEGylation reduces opsonization but does not prevent hepatic elimination. Administration routes alter systemic exposure: i.v. delivery leads to rapid organ distribution, while s.c. or i.m. routes may favor slower absorption and lymphatic transport. Additionally, lipid–drug affinity and membrane fluidity affect drug retention and release dynamics.144
The non-specific adsorption of charged nanoparticles onto phospholipid bilayers results in steric and electrostatic repulsions. These adsorbed nanoparticles work density-dependently to prevent liposomes from combining to form larger vesicles.145 Stabilizers promote lipid surface remodeling at nanoparticle adsorption sites, reducing interfacial tension and enhancing liposomal stability.146 This stabilization strategy, utilizing nanoparticles such as polystyrene, gold, and silica, facilitates the development of advanced liposome formulations with enhanced drug delivery performance.145 Stabilizing liposomes with nanoparticles allows precise modulation of surface charge and charge density, enabling stimuli-responsive attachment and detachment. This facilitates controlled liposome fusion and intelligent, on-demand cargo release. For instance, at neutral pH, carboxyl-modified gold nanoparticles adsorbed on liposomal surfaces inhibit membrane fusion. Lipid–polymer hybrid nanoparticles further enable systematic analysis of surface functionalities; studies have employed them to assess immunocompatibility of terminal groups such as methoxyl, carboxyl, and amine moieties.147
Polymeric nanoparticles offer tunable physicochemical and drug release properties through control of size, surface charge, hydrophobicity, and stimuli-responsive behavior. Surface functionalization with polymer end groups or conjugation with targeting ligands such as antibodies, peptides, or small molecules—enables tissue- or receptor-specific delivery. Their high surface-area-to-volume ratio enhances epithelial interaction and facilitates both receptor-mediated and nonspecific uptake. Polymeric NPs can encapsulate diverse therapeutics, including small molecules, biologics, and nucleic acids (eg, siRNA, DNA), and are often composed of biodegradable, FDA-approved materials suitable for clinical application.148
Lipid-Core Polymeric Micelles for Intracellular Drug Delivery
Endocytosis-mediated lysosomal degradation limits therapeutic micelle efficacy. Enhancing intracellular delivery can be achieved by targeting ligands (eg, mAb 2C5) and controlling micelle surface charge. Positively charged nanoparticles show increased cellular uptake via endocytosis, the primary mechanism for cationic lipid–DNA complexes. Lipofectin® (DOTMA/DOPE), a cationic lipid formulation, enhances intracellular drug and DNA delivery by promoting endosomal membrane disruption and endosomal escape. PEG-PE micelles carry a net negative charge, reducing cellular internalization. Incorporating cationic lipids into PEG-PE micelles neutralizes this charge, enhancing uptake and facilitating endosomal escape. Paclitaxel-loaded PEG-PE/LL (Lipofectin® lipids) micelles exhibited improved cytoplasmic distribution and higher in vitro cytotoxicity in BT-20 breast cancer and A2780 ovarian cancer cells, compared to PEG-PE micelles without cationic lipids or free paclitaxel. MTT assay results showed IC50 values for paclitaxel-loaded PEG-PE/LL micelles at 1.2 µM (A2780) and 6.4 µM (BT-20), significantly lower than free drug (22.5 µM and 24.3 µM, respectively) and non-cationic PEG-PE micelles (5.8 µM and 9.5 µM). This highlights the role of surface charge modulation and endosomal escape in enhancing micellar drug delivery and anticancer efficacy.149
Isolation and Coating of Macrophage Membranes Around the Nanocarriers
Macrophage-based delivery systems are emerging as targeted strategies for transporting therapeutics to tumor microenvironments and sites of inflammation, potentially enhancing efficacy while minimizing off-target effects. These approaches include: (1) drug delivery using live macrophages; (2) utilization of macrophage-derived extracellular vesicles (EVs) and EV-mimetic nanoparticles; (3) delivery via proteolipid nanovesicles isolated from macrophages; and (4) macrophage cell membrane (MCM)-coated nanoparticles.150,151 Macrophages’ intrinsic ability to infiltrate hypoxic tumor regions enables targeted delivery of tirapazamine-loaded PLGA nanoparticles to hypoxic zones in 4T1 breast cancer models, enhancing intratumoral drug accumulation and tumor growth inhibition in vivo. Various nanomaterials have been successfully encapsulated within live macrophages for biomedical applications. For example, macrophages loaded with MFe₃O₄-Cy5.5 photothermal nanoprobes demonstrate effective blood-brain barrier penetration and enable targeted photothermal ablation of gliomas.142
After manufacture, the membrane and inner core NPs must be joined to form a membrane-coated core and cell membrane biomimetic NPs. There are several methods for creating cell-membrane coated NPs. In situ packaging (by natural endocytosis and exocytosis), coextrusion (by mechanical force), extrusion/sonication and stir (by ultrasonic energy/endocytosis and exocytosis), extrusion/electroporation and extrusion/microfluidic electroporation (by electric pulse), and extrusion/sonication and freeze-thaw/sonication (by ultrasonic energy)152 are some of the suggested methods that have gained widespread recognition. Co-extrusion, sometimes referred to as co-injection, is the technique of using an extruder and an auxiliary system to continuously fill one material with another in order to create a single product out of two separate materials. However, some products are made using the so-called “true co-extrusion” process, which combines two extruders to extrude two products into one at the same time. The generated nanoparticles are mechanically extruded onto membranes in therapeutic settings. For several minutes before bath sonication, the “NP-membrane” mixture is repeatedly extruded through porous membranes of different diameters. For NPs to accomplish the vesicle-particle fusion, they must cross through a lipid bilayer. Following many extrusions, the precipitates are gathered and employed as final products, while the excess vesicles are centrifuged and disposed of.153
Co-extrusion methodology has been used to coat NPs with RBCs (eg, on PLGA, AuNP, Mesoporous silica, Cu2-xSe, upconversion NP, and nano pharmaceuticals), platelets (on magnetic NPs), cancer cells (on PLGA, gold NPs, and PGL), stem cells (on gelatin NPs), and immune cells (on liposome and PLGA). Recently, erythrocyte membrane-disguised anti-cancer drug delivery systems were created by co-extrusion coating red blood cell membrane-derived vesicles (RDVs) with poly (acrylic acid)-cysteine hydrochloride-D-α-tocopherol succinate (PAAssVES) nanoparticles (NPs). Polycarbonate membranes were used in this application to create homogenous, consistent, and co-extruded mixtures of RDVs and sorafenib (SFN)-PAAssVES NPs. In a different application, MDA-MB-231 breast cancer cell-derived CCM vesicles were used to coat and co-extrusion TOPSi 22 NPs. These NPs were subsequently ingested in polymeric particles of AcDEX or SpAcDEX.154 Synthetic liposomes are frequently prepared by coextrusion, in which the mechanical force exerted breaks down the membrane’s integrity and permits reconstruction around the NP core. Creating multi-layer and multifunctional structures, cutting down on the number of steps, and enabling tailored performance through the use of specific membranes are some advantages of co-extrusion technology. Through extrusion/sonication, NPs have been coated with RBCs (on PLGA and metal-organic framework), stem cells, platelets and platelet/RBCs (on PLGA), cancer cells (on liposome and mesoporous silica NPs), and immune cells (on PLGA). Ultrasonic energy induces the assembly of core nanoparticles and cell membranes into core–shell nanostructures, offering several advantages. This method preserves nanoparticle properties akin to physical extrusion while minimizing material loss. It also enables fusion of multiple cell membranes, integrating diverse functionalities. By combining membrane characteristics such as stability and charge asymmetry with nanoparticle structural integrity, this approach achieves right-side-out membrane orientation and favorable energetic configuration. Additional membrane-coating techniques include freeze-thaw/sonication (eg, platelet coating on PLGA), extrusion/sonication with stirring (platelet coating on nanogel and dextran), extrusion/electroporation (platelet coating on Au nanorods), and in situ packaging (RBC coating on perfluorocarbon-PLGA and cancer cell coating on PLGA).152
Surface Functionalization to Improve Targeting and Drug Release
The structure of the NP surface plays a critical role in determining the effectiveness of NPs both in vitro and in vivo because it affects how they interact with blood proteins, the immune system, and tumor tissue. Figure 5 is an Illustration of macrophage membrane-coated nanoparticles engineered for immune evasion and targeted delivery in inflammatory conditions such as myocardial infarction. Hydrophilic polymers, particularly polyethylene glycol (PEG, ~2–20 kDa), are the most commonly used surface coatings for nanoparticles (NPs). PEG forms a steric barrier that reduces plasma protein adsorption (opsonization) and delays immune clearance, thus prolonging NP circulation and enhancing tumor accumulation. Besides PEG, other hydrophilic polymers such as dextran, chitosan, hyaluronic acid (HA), polyvinyl pyrrolidone (PVP), polyvinyl alcohol (PVA), polyamino acids, and poloxamers have been employed as stealth coatings. An alternative stealth strategy involves coating NPs with biomimetic materials that mimic “self” components recognized by the immune system, thereby evading clearance. Examples include porphyrin-like lipoproteins (PLP), membrane proteins such as CD47, and intact red blood cell membranes. However, the “PEG dilemma” highlights a trade-off: while PEG improves NP circulation time, its presence can hinder NP interaction with target cells, reducing cellular uptake and therapeutic efficacy.155 To overcome the PEG dilemma, nanoparticles (NPs) are functionalized with cell- or receptor-specific ligands, enhancing targeted cellular interaction and uptake by tumor tissues. Ligand decoration also facilitates drug delivery to metastatic or poorly vascularized tumors where the enhanced permeability and retention (EPR) effect is insufficient. These ligands recognize specific markers on cancer cell surfaces or intracellular organelles, promoting receptor-mediated endocytosis. Enhanced cellular internalization improves therapeutic efficacy, particularly when drug targets are intracellular or when cancer cells exhibit drug efflux. Lipid-based nanocarriers have attracted significant interest due to their favorable properties, including low toxicity, improved bioavailability, biocompatibility, controlled drug release, high loading capacity, gastrointestinal stability, and scalability. Their excipients critically influence carrier characteristics and are vital for optimizing drug therapeutic outcomes. These systems effectively deliver hydrophilic, lipophilic, and amphiphilic drugs via various administration routes.156
 |
Figure 5 Conceptual overview of engineered cell membrane-camouflaged nanomaterials. The illustration highlights the design principle of coating synthetic nanoparticle cores with natural cell membranes, such as those derived from macrophages, to enhance biocompatibility, prolong systemic circulation, and enable targeted delivery through membrane-bound recognition ligands. Adapted from Guan X, Xing S, Liu Y. Engineered Cell Membrane-Camouflaged Nanomaterials for Biomedical Applications. Nanomaterials (Basel). 2024 Feb 23;14(5):413. © 2024 by the authors.157 Licensee MDPI, Basel, Switzerland under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/). |
Modifying nanocarriers with cell membranes is an effective strategy due to their long circulation, biosafety, biocompatibility, and biodegradability. The adhesive properties of cell membranes enhance antibiotic localization and site-specific delivery. Moreover, membrane coatings mimic host–pathogen interactions, improving bacterial targeting and binding.158 Developing efficient nanocarriers requires overcoming complex biological barriers to maximize therapeutic efficacy. White blood cell-derived nanoparticles (WBC-NPs) offer unique advantages, including reduced phagocytic clearance via self-recognition, enhanced trans-endothelial transport, and targeted delivery to diseased sites through inherent cellular surface ligands. These properties have enabled WBC-NPs to be explored for drug delivery in bacterial infections, cancer, and inflammatory diseases, particularly by targeting inflammatory microenvironments.159
Mechanisms of Action in Myocardial Infarction Therapy
The targeting efficiency of macrophage membrane-coated DiD nanoparticles (MM/DiDNPs) to injured myocardium was assessed using a myocardial infarction (MI) mouse model. Mice received intravenous injections of DiDNPs or MM/DiDNPs one day post-MI. Fluorescence imaging at 6 hours post-injection showed that MM/DiDNPs accumulated in the infarcted myocardium with an average intensity 1.5-fold higher than DiDNPs alone. At 12 hours, fluorescence predominantly localized to the liver, spleen, lungs, and kidneys, indicating clearance via the reticuloendothelial system, consistent with previous reports highlighting macrophage-mediated nanoparticle phagocytosis in the liver and spleen.160,161 Macrophage membrane-coated nanoparticles (MM/DiDNPs) reduce off-target accumulation and associated toxicity by lowering nonspecific uptake in key organs. Fluorescence signals in the liver and spleen were significantly decreased for MM/DiDNPs compared to DiDNPs. Therapeutic efficacy of MM/RESNPs was evaluated in a myocardial infarction (MI) mouse model via tail vein injection of saline, RES, RESNPs, or MM/RESNPs. After 28 days, echocardiography revealed that MM/RESNPs significantly improved cardiac function, with left ventricular ejection fraction (EF%) and fractional shortening (FS%) increased by approximately 1.46- and 1.56-fold over the RES group (p < 0.05, p < 0.001). MM/RESNPs also showed superior improvements in EF%, and reductions in left ventricular internal diameter in systole (LVIDs) and end-systolic volume (ESV), indicating enhanced contractile function. Histological analyses using Masson’s trichrome and TTC staining confirmed reduced fibrosis and infarct size, underscoring the therapeutic advantage of macrophage membrane coating in cardiac repair.37
Prolonged Circulation: Immune Evasion via Macrophage-Derived Proteins (Eg, CD47)
Macrophage membranes express receptors such as CXCR4, CXCR2, IL-1R, and TNF-R, conferring biomimetic targeting capabilities to macrophage membrane-coated nanoparticles. VEGF-loaded PLGA nanoparticles coated with macrophage membranes exhibit enhanced homing to inflammatory sites via CXCR2 and CXCR4 interactions, enabling targeted accumulation and sustained VEGF release within infarcted myocardium to promote neovascularization. This biomimetic coating improves the biosafety profile compared to other VEGF delivery methods. MI involves complex inflammatory responses, with M1 macrophages dominating early post-infarct phases and exacerbating cardiomyocyte injury through pro-inflammatory cytokine release. Promoting polarization from M1 to reparative M2 macrophages during early MI stages is critical for improved cardiac repair and prognosis.42 Recent studies indicate that M1 macrophages primarily promote angiogenesis initiation, while M2 macrophages support vessel maturation and stabilization. Prolonged M1 macrophage activity leads to the degradation of existing vasculature, highlighting the importance of timely M1-to-M2 polarization following myocardial ischemia/reperfusion (MI/R) injury. The macrophage phenotypic switch is regulated by diverse signals, but further research is needed to determine the optimal timing for therapeutic interventions targeting this transition to maximize treatment efficacy.162
Targeted Drug Delivery: Homing to Inflamed MI Sites
Characteristics of Drug Delivery Systems For targeted delivery to areas of vascular injury and inflammation, a variety of DDS classes have been employed, including liposomes, polymeric nanoparticles, solid lipid nanoparticles, magnetic nanoparticles, antibody- and polymer-drug conjugates, and dendrimers.163
Comparisons with Other Biomimetic Nanocarriers (Platelet-Coated, Neutrophil-Coated, Etc)
Platelets-Derived Nanostructures
Table 1 demonstrates comparison of Macrophage Membrane-Coated Nanocarriers with Other Biomimetic Nanocarriers. Platelet membranes are promising for drug delivery due to their roles in hemostasis, angiogenesis, inflammation, and wound healing, combined with high payload and storage capacity. Key membrane ligands, such as CD47 and CD55/59, enable immune evasion by inhibiting phagocytosis and complement activation, respectively. CD47 interacts with signal-regulatory protein α (SIRPα) on immune cells, reducing nanoparticle clearance and prolonging circulation time. Jin et al developed PLGA nanoparticles coated with platelet membranes, loaded with curcumin and resveratrol, for acute lung injury treatment. In vitro studies demonstrated enhanced biocompatibility, reduced hemolytic activity compared to free drugs, and anti-inflammatory effects evidenced by macrophage polarization from proinflammatory M1 to anti-inflammatory M2 phenotypes.164
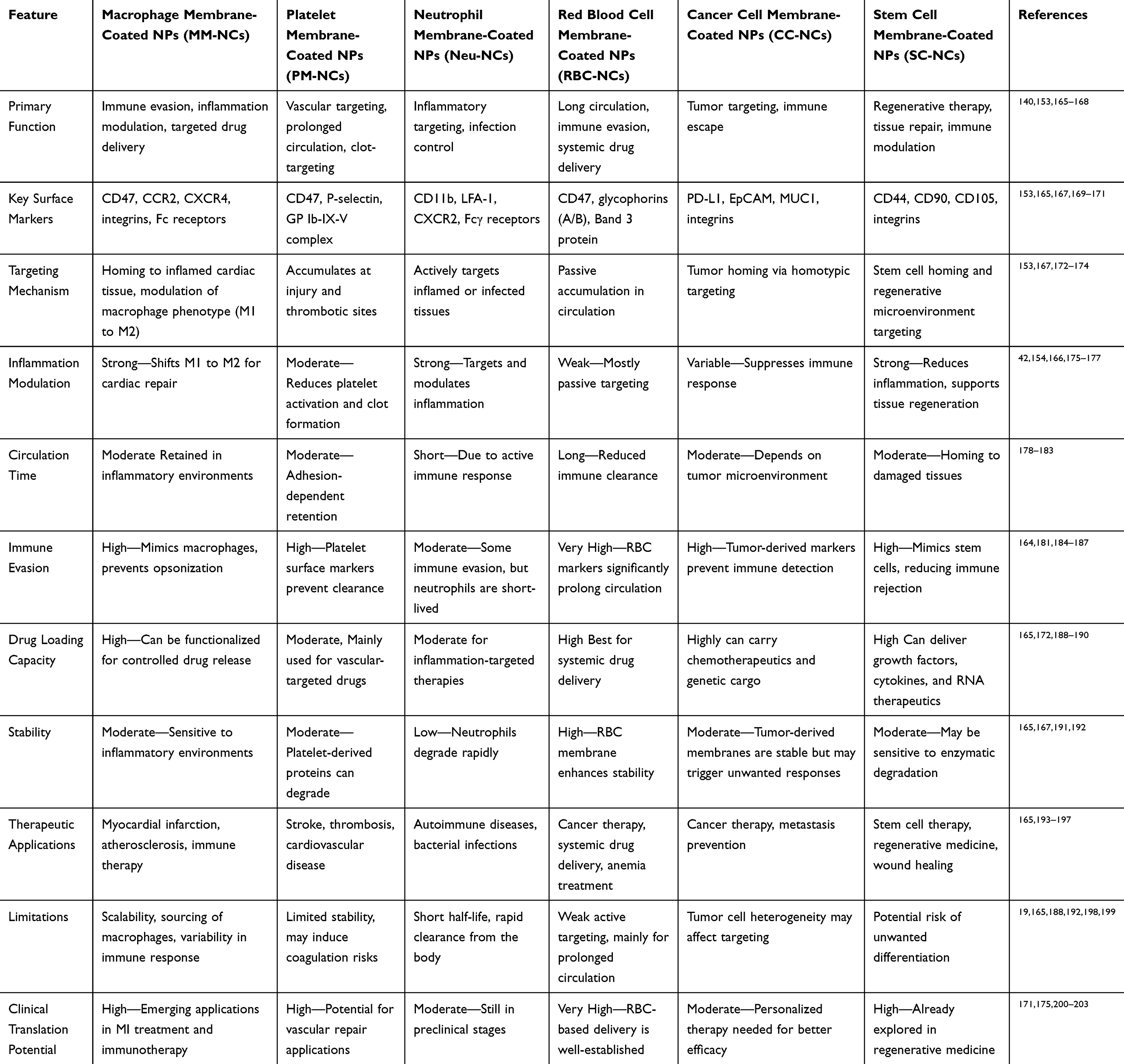 |
Table 1 Comparison of Macrophage Membrane-Coated Nanocarriers with Other Biomimetic Nanocarriers |
In a different approach, He et al also used PLGA NPs loaded with FK506 (tacrolimus) and coated with platelet membrane to treat rheumatoid arthritis. Using an in vitro model of the synovial membrane composed of collagen IV and synoviocytes, it was observed that the NPs under research may adhere to intercellular collagen IV due to the affinity between collagen and the glycoprotein VI present in the platelet membrane. According to fluorescence signals acquired in vivo using the NIRF imaging apparatus, NPs coated with the platelet membrane selectively aggregate in the inflamed joint, lowering inflammation. Blood circulation time was increased, and biodistribution studies show a preferential buildup in inflammatory regions, even though the presence of NPs in the liver and spleen—the natural pathways for platelet clearance was confirmed. Additionally, using NPs coated with platelet membrane decreased the breakdown of bone tissue.204
Leukocytes-Derived Nanostructures
Leukocytes are favored drug carriers due to their intrinsic ability to target inflammatory sites and extend drug half-life. Their prolonged circulation, capacity to traverse biological barriers, recognition of pathological tissues, and transendothelial migration make them ideal for drug delivery. Leukocytes possess diverse surface proteins, specific targeting ligands, and membrane receptors that facilitate precise targeting.205 Although physicochemical properties such as size, composition, surface ligand functionalization, and homogeneity can be controlled, replicating the complexity and integrity of leukocyte membranes remains challenging. Leukocyte membranes comprise diverse proteins and antigens essential for preserving cellular functions, including signal transduction and intercellular communication.206 Drug attachment to leukocyte membranes for cell surface functionalization is mainly achieved through three methods: covalent binding, which ensures a strong and durable linkage; receptor-mediated adhesion, a reliable and repeatable approach that can activate signaling pathways—for example, B cells utilize CD44 receptors to bind hyaluronic acid (HA)-based nanoparticles; and selectin-mediated adhesion, which enables rapid drug conjugation by targeting selectin ligands, often involving functional groups like thiol (cysteine-SH) or amino (lysine-NH2) on membrane proteins.207
Neutrophils-Derived Nanocarriers
The following characteristics of neutrophils make them appealing for the development of nanotherapeutics: (1) although neutrophils have a brief lifespan in the bloodstream, [6,117] their numbers can be raised by tens of hundreds of hours during a brief inflammatory response, which would significantly boost delivery, (2) Since neutrophils make about 50–70% of circulating human leukocytes, targeting them may improve therapeutic efficacy and have translational implications; (3) The cytokine/chemokine gradient and different membrane proteins control their migration, and this chemotaxis behavior is a very interesting characteristic to be implemented in nanosystems.208
Limitations and Challenges (Stability, Scalability, Regulatory Approval)
Recent landmark preclinical investigations significantly reinforce both the therapeutic potential and translational challenges of MM-NCs in myocardial infarction. Specifically, Xue et al (2021) demonstrated that macrophage membrane-coated nanoparticles delivering miR-199a-3p attenuate infarct size, promote cardiac repair, and modulate inflammatory cytokine profiles in murine MI models.163 Likewise, Li et al (2024) developed resveratrol-loaded PLGA nanoparticles coated with macrophage membranes (MM/RESNPs), which showed enhanced myocardial targeting, reduced infarction area, improved cardiac function, and favorable biosafety upon long-term administration.37 In addition, a study by Liao et al (2025) introduced ceria-based nanoparticles encapsulating Nrf2 (CeO2/Nrf2 nanocomposites) that accumulated in infarcted myocardium via macrophage uptake, reduced reactive oxygen species, and attenuated adverse remodeling.209 These studies provide compelling evidence for the efficacy of MM-NC strategies but also underscore the need for addressing issues such as long-term biocompatibility, membrane stability, and scalable synthesis as discussed.
Cell membrane isolation requires a minimum of 100 million cells that maintain their phenotypic, purity, and quality across passages. A proven and standardized cell culture process specific to each type of cell is necessary for large-scale production. For the ultralarge scale-up of stem cells, T cells, and dendritic cells, the existing, well-established biomanufacturing platforms that employ 3D bioreactors (such as the stirred-tanked bioreactor, WAVE bioreactor, etc.) can be beneficial in this regard.210–213 Conventional multi-step, type-specific lab-scale methods for isolating cell membranes often result in sample loss, receptor degradation, and contamination from nuclear, mitochondrial, and cytoplasmic components—particularly in nucleated cells. To achieve high-yield, high-purity membrane isolation, a streamlined, minimally manual protocol is essential. Precise control over membrane layer deposition and uniform coating is critical for assembling cell membrane-coated (CMC) mimics, although the physiological impact of membrane layer variability remains unclear. Automated techniques, such as microfluidic electroporation, offer a promising alternative by enhancing coating efficiency and producing uniformly sized RBC-coated CMC mimics.214
Enhancing the stability and storage conditions of cell membrane-coated (CMC) mimics is essential to extend their shelf life. Lyophilized membranes can be stored and later reconstituted; however, long-term studies on their functional integrity remain limited. To reduce batch-to-batch variability, quantitative characterization is necessary, assessing parameters such as membrane coverage, transmembrane protein orientation, and functional protein retention. Ensuring membranes are free of microbial and pyrogenic contaminants is critical, necessitating standardized quality control and sterile, GMP-compliant fabrication processes. Selective removal of denatured or immunogenic proteins while preserving functional surface proteins can optimize CMC efficacy, especially for targeting and immune evasion. Although several membrane modification strategies exist, few maintain correct orientation and protein functionality. Noncovalent modifications preserve bioactivity but suffer from weak binding affinity, highlighting the need for a balance between structural integrity and functional preservation.215 While covalent bonding offers strong membrane-template attachment, it risks altering native membrane functionality and disrupting protein profiles. Current evaluation methods primarily rely on changes in membrane potential, highlighting the gap between qualitative observations and quantitative assessments. Additionally, detecting small-molecule conjugation and evaluating membrane integrity remain technically challenging, limiting precise characterization of membrane modifications.201
Future Prospects and Clinical Implications
Nanoparticles (NPs) have revolutionized drug delivery by addressing many limitations of conventional therapeutics. However, their clinical translation remains constrained by biological barriers that limit targeted delivery and promote immune clearance. While synthetic NPs can be engineered with specific physicochemical properties, their recognition as foreign entities by the immune system hampers in vivo efficacy. To overcome this, biomimetic NPs have emerged as a promising strategy, leveraging cell membrane coatings to mimic endogenous interactions. These coatings confer immune evasion, prolonged circulation, and enhanced targeting by transferring surface functionalities from source cells to NPs. Hybrid membrane systems further expand functionality by combining attributes from multiple cell types. Despite these advancements, clinical application is challenged by non-standardized, small-scale production techniques that limit reproducibility and scalability. Among available sources, red blood cells (RBCs) are particularly advantageous due to their abundance, transfusion compatibility, and immunological inertness, contributing to improved biocompatibility of biomimetic nanosystems.164
Although membrane extraction techniques are relatively simple, they require further optimization to minimize contamination from intracellular components. Red blood cells (RBCs) are advantageous in this context due to their nucleus-free structure and ease of membrane isolation through basic methods like lysis and centrifugation, which also improve reproducibility in CMCNP synthesis. Ensuring complete and uniform membrane coating of synthetic NPs demands robust and standardized validation protocols.
While preclinical studies demonstrate promising therapeutic potential of macrophage membrane-coated nanocarriers (MM-NCs), most models use young, healthy rodents that do not fully represent the clinical scenario of myocardial infarction, which primarily affects aged patients with comorbidities such as diabetes and hypertension. The diverse macrophage subsets involved in cardiac injury and repair may behave differently under these conditions, potentially impacting therapeutic efficacy. Therefore, future research should focus on validating MM-NCs in aged and comorbid animal models to better reflect human pathology and improve clinical translation prospects. Therefore, future investigations should prioritize validation of MM-NCs in aged or comorbid animal models to enhance the translational relevance of preclinical findings. For successful clinical translation, stringent safety and quality control regulations must be established. The use of autologous or patient-derived membranes may enable personalized nanotherapy approaches by minimizing immunogenicity and enhancing targeting specificity. In parallel, early assessments of toxicity and long-term biocompatibility are essential to ensure safety for chronic administration. Emerging advances in genetic engineering, membrane functionalization, and the development of novel biomaterials offer promising opportunities to improve targeting precision, therapeutic performance, and overall clinical viability of biomimetic nanodrug delivery systems.216
Future Prospects, Clinical Implications, and the Nursing Role in Biomimetic Nanoparticle Therapy
The use of biomimetic nanocarriers (BNCs) in contemporary medicine has enormous promise for boosting therapeutic efficacy, decreasing immunological rejection, and increasing drug delivery. But in order to successfully integrate these cutting-edge systems into clinical practice, it is crucial to address not just the technical and regulatory issues but also the part that nurses and other healthcare professionals play in administering them, educating patients, and keeping an eye on safety.
Advanced Nursing Responsibilities in the Clinical Integration of Biomimetic Nanoparticle Therapies
The clinical application of biomimetic nanoparticle (BNP) therapies, particularly in complex conditions such as cancer, cardiovascular diseases, and inflammatory disorders, necessitates an evolved role for nurses in ensuring safety, efficacy, and patient-centered care. As these therapies utilize cell membrane-coated nanoparticles to enhance biocompatibility and targeted delivery, nursing professionals are pivotal in bridging laboratory innovation with bedside implementation. A primary nursing responsibility involves the proper management and administration of BNP-based therapeutics, which often require specialized handling protocols, such as aseptic preparation techniques, advanced infusion systems, and temperature-controlled storage. Nurses must be adept at recognizing and responding to potential adverse events, including immune-mediated hypersensitivity reactions, unexpected biodistribution of nanoparticles, or delayed systemic responses. Real-time monitoring and clinical vigilance are essential to mitigate these risks and ensure therapeutic benefit. In parallel, nurses serve a critical role in patient education and engagement. Given the novel nature of biomimetic nanocarriers, nurses must provide comprehensive information regarding the treatment’s mechanism of action, anticipated outcomes, and potential side effects. Patients should be informed about differences in pharmacokinetics and biodistribution compared to conventional therapies, as well as the importance of strict adherence to treatment regimens and follow-up schedules. Empowering patients through education not only improves compliance but also facilitates early recognition of complications. Moreover, nurses function as integral members of the multidisciplinary care team, fostering coordination among clinicians, pharmacists, biomedical engineers, and research scientists. Their clinical insights and hands-on experience can contribute significantly to protocol development, adverse event reporting, and refinement of treatment workflows. Involvement in clinical research and post-market surveillance also positions nurses to influence the safe translation of BNP technologies into routine practice.
Ethical and safety considerations further underscore the importance of the nursing role. Since many biomimetic nanocarriers utilize cell-derived membranes—including autologous or donor-derived materials—nurses are uniquely positioned to support informed consent processes by ensuring that patients understand the origin, nature, and implications of such therapies. Additionally, maintaining rigorous infection control and biocompatibility protocols is vital to prevent contamination and maintain therapeutic integrity. To support these expanded responsibilities, nursing education must evolve accordingly. Curricular reforms should incorporate core concepts in nanomedicine, targeted drug delivery systems, and biomimetic technologies. Simulation-based training and interprofessional workshops can equip nurses with practical skills for administering and troubleshooting BNP therapies. Continuing education opportunities, particularly those emphasizing collaborative learning with pharmacists, physicians, and nanotechnology researchers, are essential for sustaining clinical competency in this rapidly advancing field.
Conclusion
The limitations of conventional myocardial infarction therapies may be addressed through utilizing macrophage membrane-coated nanocarriers. These biomimetic nanocarriers leverage the immune-modulating properties and innate homing capabilities of macrophage membranes, thereby enhancing circulation longevity, reducing systemic toxicity, and improving targeted delivery to the ischemic myocardium. Preclinical studies have demonstrated that macrophage membrane-coated nanocarriers can modulate macrophage polarization, shifting the balance from pro-inflammatory M1 phenotypes to anti-inflammatory M2 phenotypes, resulting in attenuated inflammation, reduced fibrosis, and promotion of cardiac tissue regeneration. By overcoming poor drug localization and off-target inflammation, MM-NCs may reduce the incidence of post-MI heart failure, addressing a critical unmet clinical need. Despite these promising developments, several challenges remain, including the need for standardized and scalable manufacturing protocols, regulatory considerations, and ensuring long-term biocompatibility. Future research should prioritize the development of GMP-compliant membrane isolation techniques and initiate pilot clinical trials, particularly in post-PCI patients, to facilitate clinical translation. Furthermore, integrating macrophage membrane-coated nanocarriers with complementary therapeutic strategies, such as gene therapy, stem cell therapy, and personalized medicine approaches, could maximize their clinical efficacy. The critical role of interdisciplinary collaboration in advancing myocardial infarction treatment is underscored by the essential contributions of healthcare professionals, particularly nurses, in patient education, safe administration, and monitoring of nanomedicine-based therapies. As macrophage membrane-coated nanocarriers progress toward clinical translation, they hold significant potential to improve patient outcomes and mitigate the global burden of myocardial infarction.
Data Sharing Statement
Not Applicable. This is a review article, and all relevant information is provided in the article.
Ethical Approval and Consent to Participate
Not Applicable. This is a review paper and does not involve direct research on humans or animals.
Consent for Publication
“Not applicable” as this manuscript does not contain data from any person.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work. All the authors listed meet the criteria for authorship as per the ICMJE guidelines, read the final manuscript and agree to publish this work.
Funding
There is no funding to report.
Disclosure
The Authors declare that they have no competing interests financial or non-financial or any other interests that might be perceived to influence the results and/or discussion reported in this paper.
References
1. Młynarska E, Czarnik W, Fularski P, et al. From atherosclerotic plaque to myocardial infarction—The leading cause of coronary artery occlusion. Int J Mol Sci. 2024;25(13):7295. doi:10.3390/ijms25137295
2. Heusch G. Myocardial ischemia/reperfusion: translational pathophysiology of ischemic heart disease. Med. 2024;5(1):10–31. doi:10.1016/j.medj.2023.12.007
3. Lu L, Liu M, Sun R, Zheng Y, Zhang P. Myocardial infarction: symptoms and treatments. Cell Biochem Biophys. 2015;72:865–867. doi:10.1007/s12013-015-0553-4
4. Valensi P, Lorgis L, Cottin Y. Prevalence, incidence, predictive factors and prognosis of silent myocardial infarction: a review of the literature. Arch Cardiovasc Dis. 2011;104(3):178–188. doi:10.1016/j.acvd.2010.11.013
5. Devlin RJ, Henry JA. Clinical review: major consequences of illicit drug consumption. Critical Care. 2008;12:1–7. doi:10.1186/cc6166
6. Graham I, Atar D, Borch-Johnsen K, et al. European guidelines on cardiovascular disease prevention in clinical practice: executive summary: Fourth Joint Task Force of the European Society of Cardiology and Other Societies on Cardiovascular Disease Prevention in Clinical Practice (Constituted by representatives of nine societies and by invited experts). Eur Heart J. 2007;28(19):2375–2414. doi:10.1093/eurheartj/ehm316
7. Erhardt L, Herlitz J, Bossaert L, et al. Task Force on the management of chest pain. Italian Heart J Supplement. 2004;5(4):298–323.
8. Roe MT, Messenger JC, Weintraub WS, et al. Treatments, trends, and outcomes of acute myocardial infarction and percutaneous coronary intervention. J Am Coll Cardiol. 2010;56(4):254–263. doi:10.1016/j.jacc.2010.05.008
9. O’Connor RE, Brady W, Brooks SC, et al. Part 10: acute coronary syndromes: 2010 American Heart Association guidelines for cardiopulmonary resuscitation and emergency cardiovascular care. Circulation. 2010;122(18_suppl_3):S787–S817. doi:10.1161/CIRCULATIONAHA.110.971028
10. Authors/Task Force M, Hamm CW, Bassand J-P, et al. ESC Guidelines for the management of acute coronary syndromes in patients presenting without persistent ST-segment elevation: the Task Force for the management of acute coronary syndromes (ACS) in patients presenting without persistent ST-segment elevation of the European Society of Cardiology (ESC). Eur Heart J. 2011;32(23):2999–3054. doi:10.1093/eurheartj/ehr236
11. Mekonnen W, Gebremariam A. Causes of maternal death in Ethiopia between 1990 and 2016: systematic review with meta-analysis. Ethiop J Health Dev. 2018;32(4).
12. Sabatine MS, Bergmark BA, Murphy SA, et al. Percutaneous coronary intervention with drug-eluting stents versus coronary artery bypass grafting in left main coronary artery disease: an individual patient data meta-analysis. Lancet. 2021;398(10318):2247–2257. doi:10.1016/S0140-6736(21)02334-5
13. Doenst T, Haverich A, Serruys P, et al. PCI and CABG for treating stable coronary artery disease: JACC review topic of the week. J Am Coll Cardiol. 2019;73(8):964–976. doi:10.1016/j.jacc.2018.11.053
14. McCarthy CP, Vaduganathan M, McCarthy KJ, Januzzi JL, Bhatt DL, McEvoy JW. Left ventricular thrombus after acute myocardial infarction: screening, prevention, and treatment. JAMA Cardiol. 2018;3(7):642–649. doi:10.1001/jamacardio.2018.1086
15. Fraccarollo D, Galuppo P, Bauersachs J. Novel therapeutic approaches to post-infarction remodelling. Cardiovasc Res. 2012;94(2):293–303. doi:10.1093/cvr/cvs109
16. Wu X, Reboll MR, Korf-Klingebiel M, Wollert KC. Angiogenesis after acute myocardial infarction. Cardiovasc Res. 2021;117(5):1257–1273. doi:10.1093/cvr/cvaa287
17. Viola M, de Jager SCA, Sluijter JPG. Targeting inflammation after myocardial infarction: a therapeutic opportunity for extracellular vesicles? Int J Mol Sci. 2021;22(15):7831.
18. C-MJ H, Zhang L, Aryal S, Cheung C, Fang RH, Zhang L. Erythrocyte membrane-camouflaged polymeric nanoparticles as a biomimetic delivery platform. Proc Natl Acad Sci. 2011;108(27):10980–10985. doi:10.1073/pnas.1106634108
19. Khatoon N, Zhang Z, Zhou C, Chu M. Macrophage membrane coated nanoparticles: a biomimetic approach for enhanced and targeted delivery. Biomater Sci. 2022;10(5):1193–1208. doi:10.1039/D1BM01664D
20. Hilgendorf I, Frantz S, Frangogiannis NG. Repair of the infarcted heart: cellular effectors, molecular mechanisms and therapeutic opportunities. Circ Res. 2024;134(12):1718–1751. doi:10.1161/CIRCRESAHA.124.323658
21. Welt FG, Batchelor W, Spears JR, et al. Reperfusion injury in patients with acute myocardial infarction: JACC scientific statement. J Am Coll Cardiol. 2024;83(22):2196–2213. doi:10.1016/j.jacc.2024.02.056
22. Zhang W, Mehta A, Tong Z, Esser L, Voelcker NH. Development of polymeric nanoparticles for blood–brain barrier transfer—strategies and challenges. Adv Sci. 2021;8(10):2003937. doi:10.1002/advs.202003937
23. Smit M, Coetzee AR, Lochner A. The pathophysiology of myocardial ischemia and perioperative myocardial infarction. J Cardiothorac Vasc Anesthesia. 2020;34(9):2501–2512. doi:10.1053/j.jvca.2019.10.005
24. He X, Du T, Long T, Liao X, Dong Y, Huang Z-P. Signaling cascades in the failing heart and emerging therapeutic strategies. Signal Transduct Target Ther. 2022;7(1):134. doi:10.1038/s41392-022-00972-6
25. Tang W, Yang Y, Yang L, Tang M, Chen Y, Li C. Macrophage membrane-mediated targeted drug delivery for treatment of spinal cord injury regardless of the macrophage polarization states. Asian J Pharm Sci. 2021;16(4):459–470. doi:10.1016/j.ajps.2021.03.005
26. Chen T, Wang Y, Zhu L, et al. Hybrid membrane camouflaged chemodrug-gene nanoparticles for enhanced combination therapy of ovarian cancer. ACS Appl Mater Interfaces. 2023;15(50):58067–58078. doi:10.1021/acsami.3c10586
27. Parodi A, Quattrocchi N, Van De Ven AL, et al. Synthetic nanoparticles functionalized with biomimetic leukocyte membranes possess cell-like functions. Nature Nanotechnol. 2013;8(1):61–68. doi:10.1038/nnano.2012.212
28. Zhang Q, Dehaini D, Zhang Y, et al. Neutrophil membrane-coated nanoparticles inhibit synovial inflammation and alleviate joint damage in inflammatory arthritis. Nature Nanotechnol. 2018;13(12):1182–1190. doi:10.1038/s41565-018-0254-4
29. Huang J-Q, Zhang Q, Wei F. Multi-functional separator/interlayer system for high-stable lithium-sulfur batteries: progress and prospects. Energy Storage Mater. 2015;1:127–145.
30. Krishnan N, Fang RH, Zhang L. Cell membrane‐coated nanoparticles for the treatment of cancer. Clin Transl Med. 2023;13(6):e1285. doi:10.1002/ctm2.1285
31. Hamidzadeh K, Christensen SM, Dalby E, Chandrasekaran P, Mosser DM. Macrophages and the recovery from acute and chronic inflammation. Ann Rev Physiol. 2017;79(1):567–592. doi:10.1146/annurev-physiol-022516-034348
32. Li T, Yan Z, Fan Y, et al. Cardiac repair after myocardial infarction: a two-sided role of inflammation-mediated. Front Cardiovasc Med. 2023;9:1077290. doi:10.3389/fcvm.2022.1077290
33. Peet C, Ivetic A, Bromage DI, Shah AM. Cardiac monocytes and macrophages after myocardial infarction. Cardiovasc Res. 2020;116(6):1101–1112. doi:10.1093/cvr/cvz336
34. van der Laan AM, Ter Horst EN, Delewi R, et al. Monocyte subset accumulation in the human heart following acute myocardial infarction and the role of the spleen as monocyte reservoir. Eur Heart J. 2014;35(6):376–385. doi:10.1093/eurheartj/eht331
35. Honold L, Nahrendorf M. Resident and monocyte-derived macrophages in cardiovascular disease. Circ Res. 2018;122(1):113–127. doi:10.1161/CIRCRESAHA.117.311071
36. Dick SA, Macklin JA, Nejat S, et al. Self-renewing resident cardiac macrophages limit adverse remodeling following myocardial infarction. Nat Immunol. 2019;20(1):29–39. doi:10.1038/s41590-018-0272-2
37. Li T, Feng J, Ding W, Zhang Z. Targeted treatment of myocardial infarction by macrophage membrane coated with resveratrol nanoparticles. ACS omega. 2024;9(47):47145–47155. doi:10.1021/acsomega.4c07573
38. Jian Y, Zhou X, Shan W, et al. Crosstalk between macrophages and cardiac cells after myocardial infarction. Cell Commun Signaling. 2023;21(1):109. doi:10.1186/s12964-023-01105-4
39. Liu T, Sun F, Cui J, et al. Morroniside enhances angiogenesis and improves cardiac function following acute myocardial infarction in rats. Eur J Pharmacol. 2020;872:172954. doi:10.1016/j.ejphar.2020.172954
40. Rieck S, Heun Y, Heidsieck A, et al. Local anti-angiogenic therapy by magnet-assisted downregulation of SHP2 phosphatase. J Control Release. 2019;305:155–164. doi:10.1016/j.jconrel.2019.05.031
41. Brozovich FV, Nicholson CJ, Degen CV, Gao YZ, Aggarwal M, Morgan KG. Mechanisms of vascular smooth muscle contraction and the basis for pharmacologic treatment of smooth muscle disorders. Pharmacol Rev. 2016;68(2):476–532. doi:10.1124/pr.115.010652
42. Zhu K, Wang K, Yao Y, et al. N-acetyl-L-cysteine-modified macrophage membrane-coated VEGF sustained-release nanoparticles for treatment of myocardial infarction: a biomimetic nano-buffer for neutralization of detrimental factors and promotion of mature angiogenesis. Chem Eng J. 2024;489:151438. doi:10.1016/j.cej.2024.151438
43. Chen S, Han B, Zhao Y, et al. Injectable EC-BMSC hydrogel with prolonged VEGF action for enhanced angiogenesis. Polym Test. 2023;125:108109. doi:10.1016/j.polymertesting.2023.108109
44. Kinoshita M, Uchida T, Sato A, et al. Characterization of two F4/80-positive Kupffer cell subsets by their function and phenotype in mice. J Hepatol. 2010;53(5):903–910. doi:10.1016/j.jhep.2010.04.037
45. González A, Schelbert EB, Díez J, Butler J. Myocardial interstitial fibrosis in heart failure: biological and translational perspectives. J Am Coll Cardiol. 2018;71(15):1696–1706. doi:10.1016/j.jacc.2018.02.021
46. Rossol M, Heine H, Meusch U, et al. LPS-induced cytokine production in human monocytes and macrophages. Critical Rev Immunol. 2011;31(5):379–446. doi:10.1615/critrevimmunol.v31.i5.20
47. Bhattacharyya S, Ghosh SS. Transmembrane TNFα-expressed macrophage membrane-coated chitosan nanoparticles as cancer therapeutics. ACS omega. 2020;5(3):1572–1580. doi:10.1021/acsomega.9b03531
48. Wen RX, Shen H, Huang SX, et al. P2Y6 receptor inhibition aggravates ischemic brain injury by reducing microglial phagocytosis. CNS Neurosci Ther. 2020;26(4):416–429. doi:10.1111/cns.13296
49. Wan E, Yeap XY, Dehn S, et al. Enhanced efferocytosis of apoptotic cardiomyocytes through MER tyrosine kinase links acute inflammation resolution to cardiac repair after infarction. Circ Res. 2013;113(8):10–1161. doi:10.1161/CIRCRESAHA.113.301198
50. Zhang G, Li Q, Tao W, et al. Sigma-1 receptor-regulated efferocytosis by infiltrating circulating macrophages/microglial cells protects against neuronal impairments and promotes functional recovery in cerebral ischemic stroke. Theranostics. 2023;13(2):543. doi:10.7150/thno.77088
51. Horckmans M, Ring L, Duchene J, et al. Neutrophils orchestrate post-myocardial infarction healing by polarizing macrophages towards a reparative phenotype. Eur Heart J. 2017;38(3):187–197. doi:10.1093/eurheartj/ehw002
52. Weirather J, Hofmann UDW, Beyersdorf N, et al. Foxp3+ CD4+ T cells improve healing after myocardial infarction by modulating monocyte/macrophage differentiation. Circ Res. 2014;115(1):55–67. doi:10.1161/CIRCRESAHA.115.303895
53. Chen W, Saxena A, Li N, et al. Endogenous IRAK-M attenuates postinfarction remodeling through effects on macrophages and fibroblasts. Arteriosclerosis Thrombosis Vasc Biol. 2012;32(11):2598–2608. doi:10.1161/ATVBAHA.112.300310
54. Song J, Frieler RA, Whitesall SE, et al. Myeloid interleukin-4 receptor α is essential in postmyocardial infarction healing by regulating inflammation and fibrotic remodeling. Am J Physiol Heart Circ Physiol. 2021;320(1):H323–H337. doi:10.1152/ajpheart.00251.2020
55. Dewald O, Ren G, Duerr GD, et al. Of mice and dogs: species-specific differences in the inflammatory response following myocardial infarction. Am J Pathol. 2004;164(2):665–677. doi:10.1016/S0002-9440(10)63154-9
56. Frangogiannis NG, Ren G, Dewald O, et al. Critical role of endogenous thrombospondin-1 in preventing expansion of healing myocardial infarcts. Circulation. 2005;111(22):2935–2942. doi:10.1161/CIRCULATIONAHA.104.510354
57. Gong D, Shi W, S-j Y, Chen H, Groffen J, Heisterkamp N. TGFβ signaling plays a critical role in promoting alternative macrophage activation. BMC Immunol. 2012;13:1–10. doi:10.1186/1471-2172-13-31
58. Frangogiannis NG. Transforming growth factor-β in myocardial disease. Nat Rev Cardiol. 2022;19(7):435–455. doi:10.1038/s41569-021-00646-w
59. Li J, Li R, Tuleta I, et al. The role of endogenous Smad7 in regulating macrophage phenotype following myocardial infarction. THE FASEB Journal. 2022;36(7):e22400. doi:10.1096/fj.202101956RR
60. Chen B, Huang S, Su Y, et al. Macrophage Smad3 protects the infarcted heart, stimulating phagocytosis and regulating inflammation. Circ Res. 2019;125(1):55–70. doi:10.1161/CIRCRESAHA.119.315069
61. Chen B, Li R, Hernandez SC, et al. Differential effects of Smad2 and Smad3 in regulation of macrophage phenotype and function in the infarcted myocardium. J Mol Cell Cardiol. 2022;171:1–15. doi:10.1016/j.yjmcc.2022.06.009
62. Kubota A, Frangogiannis NG. Macrophages in myocardial infarction. Am J Physiol Cell Physiol. 2022;323(4):C1304–C1324. doi:10.1152/ajpcell.00230.2022
63. Nahrendorf M, Pittet MJ, Swirski FK. Monocytes: protagonists of infarct inflammation and repair after myocardial infarction. Circulation. 2010;121(22):2437–2445. doi:10.1161/CIRCULATIONAHA.109.916346
64. Nahrendorf M, Swirski FK, Aikawa E, et al. The healing myocardium sequentially mobilizes two monocyte subsets with divergent and complementary functions. J Exp Med. 2007;204(12):3037–3047. doi:10.1084/jem.20070885
65. Geissmann F, Jung S, Littman DR. Blood monocytes consist of two principal subsets with distinct migratory properties. Immunity. 2003;19(1):71–82. doi:10.1016/S1074-7613(03)00174-2
66. Nahrendorf M, Keliher E, Panizzi P, et al. 18F-4V for PET–CT imaging of VCAM-1 expression in atherosclerosis. JACC Cardiovasc Imaging. 2009;2(10):1213–1222. doi:10.1016/j.jcmg.2009.04.016
67. Yellon DM, Hausenloy DJ. Myocardial reperfusion injury. N Engl J Med. 2007;357(11):1121–1135. doi:10.1056/NEJMra071667
68. Prasad A, Stone GW, Holmes DR, Gersh B. Reperfusion injury, microvascular dysfunction, and cardioprotection: the “dark side” of reperfusion. Circulation. 2009;120(21):2105–2112. doi:10.1161/CIRCULATIONAHA.108.814640
69. Leor J, Rozen L, Zuloff-Shani A, et al. Ex vivo activated human macrophages improve healing, remodeling, and function of the infarcted heart. Circulation. 2006;114(1_supplement):I–94. doi:10.1161/CIRCULATIONAHA.105.000331
70. Morimoto H, Takahashi M, Izawa A, et al. Cardiac overexpression of monocyte chemoattractant protein-1 in transgenic mice prevents cardiac dysfunction and remodeling after myocardial infarction. Circ Res. 2006;99(8):891–899. doi:10.1161/01.RES.0000246113.82111.2d
71. van Amerongen MJ, Harmsen MC, van Rooijen N, Petersen AH, van Luyn MJA. Macrophage depletion impairs wound healing and increases left ventricular remodeling after myocardial injury in mice. Am J Pathol. 2007;170(3):818–829. doi:10.2353/ajpath.2007.060547
72. Swirski FK, Nahrendorf M, Etzrodt M, et al. Identification of splenic reservoir monocytes and their deployment to inflammatory sites. Science. 2009;325(5940):612–616. doi:10.1126/science.1175202
73. Cleutjens JPM, Blankesteijn WM, Daemen MJAP, Smits JFM. The infarcted myocardium: simply dead tissue, or a lively target for therapeutic interventions. Cardiovasc Res. 1999;44(2):232–241. doi:10.1016/S0008-6363(99)00212-6
74. Ertl G, Frantz S. Healing after myocardial infarction. Cardiovasc Res. 2005;66(1):22–32. doi:10.1016/j.cardiores.2005.01.011
75. Frantz S, Bauersachs J, Ertl G. Post-infarct remodelling: contribution of wound healing and inflammation. Cardiovasc Res. 2009;81(3):474–481. doi:10.1093/cvr/cvn292
76. Pfeffer MA, Braunwald E, Moyé LA, et al. Effect of captopril on mortality and morbidity in patients with left ventricular dysfunction after myocardial infarction: results of the survival and ventricular enlargement trial. N Engl J Med. 1992;327(10):669–677. doi:10.1056/NEJM199209033271001
77. Pfeffer MA, Braunwald E. Ventricular remodeling after myocardial infarction. Experimental observations and clinical implications. Circulation. 1990;81(4):1161–1172. doi:10.1161/01.cir.81.4.1161
78. Auffray C, Sieweke MH, Geissmann F. Blood monocytes: development, heterogeneity, and relationship with dendritic cells. Ann Rev Immunol. 2009;27(1):669–692. doi:10.1146/annurev.immunol.021908.132557
79. on Population H B, Public Health P. Committee on a National Surveillance System for C, Select Chronic D. A Nationwide Framework for Surveillance of Cardiovascular and Chronic Lung Diseases. National Academies Press; 2011.
80. Anversa P, Kajstura J, Leri A, Bolli R. Life and death of cardiac stem cells: a paradigm shift in cardiac biology. Circulation. 2006;113(11):1451–1463. doi:10.1161/CIRCULATIONAHA.105.595181
81. Chien KR, Domian IJ, Parker KK. Cardiogenesis and the complex biology of regenerative cardiovascular medicine. Science. 2008;322(5907):1494–1497. doi:10.1126/science.1163267
82. Tabas I, Bornfeldt KE. Macrophage phenotype and function in different stages of atherosclerosis. Circ Res. 2016;118(4):653–667. doi:10.1161/CIRCRESAHA.115.306256
83. Libby P, Pasterkamp G, Crea F, Jang I-K. Reassessing the mechanisms of acute coronary syndromes: the “vulnerable plaque” and superficial erosion. Circ Res. 2019;124(1):150–160. doi:10.1161/CIRCRESAHA.118.311098
84. Hulsmans M, Clauss S, Xiao L, et al. Macrophages facilitate electrical conduction in the heart. Cell. 2017;169(3):510–522. doi:10.1016/j.cell.2017.03.050
85. Cahill TJ, Sun X, Ravaud C, et al. Tissue-resident macrophages regulate lymphatic vessel growth and patterning in the developing heart. Development. 2021;148(3):dev194563. doi:10.1242/dev.194563
86. Leid J, Carrelha J, Boukarabila H, Epelman S, Jacobsen SEW, Lavine KJ. Primitive embryonic macrophages are required for coronary development and maturation. Circ Res. 2016;118(10):1498–1511. doi:10.1161/CIRCRESAHA.115.308270
87. Galis ZS, Sukhova GK, Kranzhöfer R, Clark S, Libby P. Macrophage foam cells from experimental atheroma constitutively produce matrix-degrading proteinases. Proc Natl Acad Sci. 1995;92(2):402–406. doi:10.1073/pnas.92.2.402
88. Moore KJ, Tabas I. Macrophages in the pathogenesis of atherosclerosis. Cell. 2011;145(3):341–355. doi:10.1016/j.cell.2011.04.005
89. Bäck M, Yurdagul JA, Tabas I, Öörni K, Kovanen PT. Inflammation and its resolution in atherosclerosis: mediators and therapeutic opportunities. Nat Rev Cardiol. 2019;16(7):389–406. doi:10.1038/s41569-019-0169-2
90. Barrett TJ. Macrophages in atherosclerosis regression. Arteriosclerosis Thrombosis Vasc Biol. 2020;40(1):20–33. doi:10.1161/ATVBAHA.119.312802
91. Frangogiannis NG. The immune system and the remodeling infarcted heart: cell biological insights and therapeutic opportunities. J Cardiovasc Pharmacol. 2014;63(3):185–195. doi:10.1097/FJC.0000000000000003
92. Jennings RB, Murry CE, Steenbergen JC, Reimer KA. Development of cell injury in sustained acute ischemia. Circulation. 1990;82(3 Suppl):II2–12.
93. Reimer KA, Lowe JE, Rasmussen MM, Jennings RB. The wavefront phenomenon of ischemic cell death. 1. Myocardial infarct size vs duration of coronary occlusion in dogs. Circulation. 1977;56(5):786–794. doi:10.1161/01.CIR.56.5.786
94. Andrassy M, Volz HC, Igwe JC, et al. High-mobility group box-1 in ischemia-reperfusion injury of the heart. Circulation. 2008;117(25):3216–3226. doi:10.1161/CIRCULATIONAHA.108.769331
95. Rohde D, Schön C, Boerries M, et al. S100A1 is released from ischemic cardiomyocytes and signals myocardial damage via Toll‐like receptor 4. EMBO Mol Med. 2014;6(6):778–794. doi:10.15252/emmm.201303498
96. Tian J, Guo X, Liu X-M, et al. Extracellular HSP60 induces inflammation through activating and up-regulating TLRs in cardiomyocytes. Cardiovasc Res. 2013;98(3):391–401. doi:10.1093/cvr/cvt047
97. Kono H, Rock KL. How dying cells alert the immune system to danger. Nat Rev Immunol. 2008;8(4):279–289. doi:10.1038/nri2215
98. Jaén RI, Val-Blasco A, Prieto P, et al. Innate immune receptors, key actors in cardiovascular diseases. Basic Transl Sci. 2020;5(7):735–749. doi:10.1016/j.jacbts.2020.03.015
99. Arslan F, De Kleijn DP, Pasterkamp G. Innate immune signaling in cardiac ischemia. Nat Rev Cardiol. 2011;8(5):292–300. doi:10.1038/nrcardio.2011.38
100. Frangogiannis NG, Lindsey ML, Michael LH, et al. Resident cardiac mast cells degranulate and release preformed TNF-α, initiating the cytokine cascade in experimental canine myocardial ischemia/reperfusion. Circulation. 1998;98(7):699–710. doi:10.1161/01.CIR.98.7.699
101. Singh MV, Swaminathan PD, Luczak ED, Kutschke W, Weiss RM, Anderson ME. MyD88 mediated inflammatory signaling leads to CaMKII oxidation, cardiac hypertrophy and death after myocardial infarction. J Mol Cell Cardiol. 2012;52(5):1135–1144. doi:10.1016/j.yjmcc.2012.01.021
102. Favre J, Musette P, Douin-Echinard V, et al. Toll-like receptors 2-deficient mice are protected against postischemic coronary endothelial dysfunction. Arteriosclerosis Thrombosis Vasc Biol. 2007;27(5):1064–1071. doi:10.1161/ATVBAHA.107.140723
103. Bajpai G, Bredemeyer A, Li W, et al. Tissue resident CCR2− and CCR2+ cardiac macrophages differentially orchestrate monocyte recruitment and fate specification following myocardial injury. Circ Res. 2019;124(2):263–278. doi:10.1161/CIRCRESAHA.118.314028
104. Kloc M, Ghobrial RM, Wosik J, Lewicka A, Lewicki S, Kubiak JZ. Macrophage functions in wound healing. J Tissue Eng Regener Med. 2019;13(1):99–109. doi:10.1002/term.2772
105. Spiller KL, Koh TJ. Macrophage-based therapeutic strategies in regenerative medicine. Adv Drug Delivery Rev. 2017;122:74–83. doi:10.1016/j.addr.2017.05.010
106. Chan MWY, Viswanathan S. Recent progress on developing exogenous monocyte/macrophage-based therapies for inflammatory and degenerative diseases. Cytotherapy. 2019;21(4):393–415. doi:10.1016/j.jcyt.2019.02.002
107. Barman PK, Koh TJ. Macrophage dysregulation and impaired skin wound healing in diabetes. Front Cell Develop Biol. 2020;8:528. doi:10.3389/fcell.2020.00528
108. Giannarelli C, Fernandez DM. Manipulating macrophage polarization to fix the broken heart: challenges and hopes. Am Coll Cardiol Foundation. 2018;72:905–907. doi:10.1016/j.jacc.2018.06.019
109. Wang Z, Koenig AL, Lavine KJ, Apte RS. Macrophage plasticity and function in the eye and heart. Trends Immunol. 2019;40(9):825–841. doi:10.1016/j.it.2019.07.002
110. O’Rourke SA, Dunne A, Monaghan MG. The role of macrophages in the infarcted myocardium: orchestrators of ECM remodeling. Front Cardiovasc Med. 2019;6:101. doi:10.3389/fcvm.2019.00101
111. Nicolás-ávila JA, Hidalgo A, Ballesteros I. Specialized functions of resident macrophages in brain and heart. J Leukocyte Biol. 2018;104(4):743–756. doi:10.1002/JLB.6MR0118-041R
112. Poller WC, Nahrendorf M, Swirski FK. Hematopoiesis and cardiovascular disease. Circ Res. 2020;126(8):1061–1085. doi:10.1161/CIRCRESAHA.120.315895
113. Swirski FK, Nahrendorf M. Cardioimmunology: the immune system in cardiac homeostasis and disease. Nat Rev Immunol. 2018;18(12):733–744. doi:10.1038/s41577-018-0065-8
114. Singer AJ, Clark RAF. Cutaneous wound healing. N Engl J Med. 1999;341(10):738–746. doi:10.1056/NEJM199909023411006
115. Danenberg HD, Fishbein I, Gao J, et al. Macrophage depletion by clodronate-containing liposomes reduces neointimal formation after balloon injury in rats and rabbits. Circulation. 2002;106(5):599–605. doi:10.1161/01.CIR.0000023532.98469.48
116. Espinosa-Heidmann DG, Suner IJ, Hernandez EP, Monroy D, Csaky KG, Cousins SW. Macrophage depletion diminishes lesion size and severity in experimental choroidal neovascularization. Invest Ophthalmol Visual Sci. 2003;44(8):3586–3592. doi:10.1167/iovs.03-0038
117. Sakurai E, Anand A, Ambati BK, van Rooijen N, Ambati J. Macrophage depletion inhibits experimental choroidal neovascularization. Invest Ophthalmol Visual Sci. 2003;44(8):3578–3585. doi:10.1167/iovs.03-0097
118. Takayama J, Koyamada N, Abe T, et al. Macrophage depletion prevents accelerated rejection and results in long-term survival in hamster to rat cardiac xenotransplantation. In: Transplantation Proceedings. Vol. 2000:1016.
119. Liu T, van Rooijen N, Tracey DJ. Depletion of macrophages reduces axonal degeneration and hyperalgesia following nerve injury. Pain. 2000;86(1–2):25–32. doi:10.1016/S0304-3959(99)00306-1
120. Duffield JS, Forbes SJ, Constandinou CM, et al. Selective depletion of macrophages reveals distinct, opposing roles during liver injury and repair. J Clin Invest. 2005;115(1):56–65. doi:10.1172/JCI200522675
121. Li Q, Park PW, Wilson CL, Parks WC. Matrilysin shedding of syndecan-1 regulates chemokine mobilization and transepithelial efflux of neutrophils in acute lung injury. Cell. 2002;111(5):635–646. doi:10.1016/S0092-8674(02)01079-6
122. Suzuki T, S-i H, Toyoda N, et al. Comprehensive gene expression profile of LPS-stimulated human monocytes by SAGE. Blood J Am Soc Hematol. 2000;96(7):2584–2591.
123. Aikawa M, Rabkin E, Sugiyama S, et al. An HMG-CoA reductase inhibitor, cerivastatin, suppresses growth of macrophages expressing matrix metalloproteinases and tissue factor in vivo and in vitro. Circulation. 2001;103(2):276–283. doi:10.1161/01.CIR.103.2.276
124. Kakio T, Matsumori A, Ono K, Ito H, Matsushima K, Sasayama S. Roles and relationship of macrophages and monocyte chemotactic and activating factor/monocyte chemoattractant protein-1 in the ischemic and reperfused rat heart. Lab Invest. 2000;80(7):1127–1136. doi:10.1038/labinvest.3780119
125. Frangogiannis NG, Smith CW, Entman ML. The inflammatory response in myocardial infarction. Cardiovasc Res. 2002;53(1):31–47. doi:10.1016/S0008-6363(01)00434-5
126. Chen G, Jiang H, Yao Y, et al. Macrophage, a potential targeted therapeutic immune cell for cardiomyopathy. Front Cell Develop Biol. 2022;10:908790. doi:10.3389/fcell.2022.908790
127. Okazaki T, Ebihara S, Asada M, et al. Macrophage colony-stimulating factor improves cardiac function after ischemic injury by inducing vascular endothelial growth factor production and survival of cardiomyocytes. Am J Pathol. 2007;171(4):1093–1103. doi:10.2353/ajpath.2007.061191
128. Yano T, Miura T, Whittaker P, et al. Macrophage colony-stimulating factor treatment after myocardial infarction attenuates left ventricular dysfunction by accelerating infarct repair. J Am Coll Cardiol. 2006;47(3):626–634. doi:10.1016/j.jacc.2005.09.037
129. Frangogiannis NG. The mechanistic basis of infarct healing. Antioxid Redox Signaling. 2006;8(11–12):1907–1939.
130. Lambert JM, Lopez EF, Lindsey ML. Macrophage roles following myocardial infarction. Int J Cardiol. 2008;130(2):147–158. doi:10.1016/j.ijcard.2008.04.059
131. Kobusiak-Prokopowicz M, Orzeszko J, Mazur G, Mysiak A, Orda A, Mazurek W. Kinetics of chemokines in acute myocardial infarction. Polish Heart J. 2005;62(4):309–314.
132. Cho D-I, Kim MR, H-y J, et al. Mesenchymal stem cells reciprocally regulate the M1/M2 balance in mouse bone marrow-derived macrophages. Exp Mol Med. 2014;46(1):e70–e70. doi:10.1038/emm.2013.135
133. Im Cho D, H-j K, Jeon JH, et al. Antiinflammatory activity of ANGPTL4 facilitates macrophage polarization to induce cardiac repair. JCI Insight. 2019;4(16):e125437. doi:10.1172/jci.insight.125437
134. Zhang B, Zhao N, Zhang J, Liu YU, Zhu D, Kong YE. Mesenchymal stem cells rejuvenate cardiac muscle through regulating macrophage polarization. Aging. 2019;11(12):3900. doi:10.18632/aging.102009
135. Zhao J, Li X, Hu J, et al. Mesenchymal stromal cell-derived exosomes attenuate myocardial ischaemia-reperfusion injury through miR-182-regulated macrophage polarization. Cardiovasc Res. 2019;115(7):1205–1216. doi:10.1093/cvr/cvz040
136. Xu R, Zhang F, Chai R, et al. Exosomes derived from pro‐inflammatory bone marrow‐derived mesenchymal stem cells reduce inflammation and myocardial injury via mediating macrophage polarization. J Cell Mol Med. 2019;23(11):7617–7631. doi:10.1111/jcmm.14635
137. Hu H, Wu J, Cao C, Ma L. Exosomes derived from regulatory T cells ameliorate acute myocardial infarction by promoting macrophage M2 polarization. IUBMB Life. 2020;72(11):2409–2419. doi:10.1002/iub.2364
138. Liu G, Zhao X, Zhang Y, et al. Engineering biomimetic platesomes for pH‐responsive drug delivery and enhanced antitumor activity. Adv Mater. 2019;31(32):1900795. doi:10.1002/adma.201900795
139. Epelman S, Lavine KJ, Randolph GJ. Origin and functions of tissue macrophages. Immunity. 2014;41(1):21–35. doi:10.1016/j.immuni.2014.06.013
140. Wu Y, Wan S, Yang S, et al. Macrophage cell membrane-based nanoparticles: a new promising biomimetic platform for targeted delivery and treatment. J Nanobiotechnol. 2022;20(1):542. doi:10.1186/s12951-022-01746-6
141. Ison MG, Hirsch HH. Community-acquired respiratory viruses in transplant patients: diversity, impact, unmet clinical needs. Clin Microbiol Rev. 2019;32(4):10–1128.
142. Lopes J, Lopes D, Pereira‐Silva M, et al. Macrophage cell membrane‐cloaked nanoplatforms for biomedical applications. Small Methods. 2022;6(8):2200289.
143. Li C, Zhao Z, Luo Y, et al. Macrophage‐disguised manganese dioxide nanoparticles for neuroprotection by reducing oxidative stress and modulating inflammatory microenvironment in acute ischemic stroke. Adv Sci. 2021;8(20):2101526. doi:10.1002/advs.202101526
144. Kraft JC, Freeling JP, Wang Z, Ho RJY. Emerging research and clinical development trends of liposome and lipid nanoparticle drug delivery systems. J Pharmaceut Sci. 2014;103(1):29–52. doi:10.1002/jps.23773
145. Michel R, Plostica T, Abezgauz L, Danino D, Gradzielski M. Control of the stability and structure of liposomes by means of nanoparticles. Soft Matter. 2013;9(16):4167–4177.
146. Wang B, Zhang L, Bae SC, Granick S. Nanoparticle-induced surface reconstruction of phospholipid membranes. Proc Natl Acad Sci. 2008;105(47):18171–18175. doi:10.1073/pnas.0807296105
147. Gao W, Hu C-MJ, Fang RH, Zhang L. Liposome-like nanostructures for drug delivery. J Mat Chem B. 2013;1(48):6569–6585.
148. Pridgen EM, Alexis F, Farokhzad OC. Polymeric nanoparticle drug delivery technologies for oral delivery applications. Expert Opin Drug Delivery. 2015;12(9):1459–1473.
149. Torchilin VP. Lipid-core micelles for targeted drug delivery. Current Drug Delivery. 2005;2(4):319–327. doi:10.2174/156720105774370221
150. Jahromi LP, Shahbazi MA, Maleki A, Azadi A, Santos HA. Chemically engineered immune cell‐derived microrobots and biomimetic nanoparticles: emerging biodiagnostic and therapeutic tools. Adv Sci. 2021;8(8):2002499. doi:10.1002/advs.202002499
151. Xia Y, Rao L, Yao H, Wang Z, Ning P, Chen X. Engineering macrophages for cancer immunotherapy and drug delivery. Adv Mater. 2020;32(40):2002054. doi:10.1002/adma.202002054
152. Choi B, Park W, Park S-B, Rhim W-K, Han DK. Recent trends in cell membrane-cloaked nanoparticles for therapeutic applications. Methods. 2020;177:2–14. doi:10.1016/j.ymeth.2019.12.004
153. Xia Q, Zhang Y, Li Z, Hou X, Feng N. Red blood cell membrane-camouflaged nanoparticles: a novel drug delivery system for antitumor application. Acta Pharmaceutica Sinica B. 2019;9(4):675–689. doi:10.1016/j.apsb.2019.01.011
154. Oroojalian F, Beygi M, Baradaran B, Mokhtarzadeh A, Shahbazi MA. Immune cell Membrane‐Coated biomimetic nanoparticles for targeted cancer therapy. Small. 2021;17(12):2006484.
155. Hatakeyama H, Akita H, Harashima H. A multifunctional envelope type nano device (MEND) for gene delivery to tumours based on the EPR effect: a strategy for overcoming the PEG dilemma. Adv Drug Delivery Rev. 2011;63(3):152–160. doi:10.1016/j.addr.2010.09.001
156. Abd Ellah NH, Abouelmagd SA. Surface functionalization of polymeric nanoparticles for tumor drug delivery: approaches and challenges. Expert Opin Drug Delivery. 2017;14(2):201–214. doi:10.1080/17425247.2016.1213238
157. Guan X, Xing S, Liu Y. Engineered cell membrane-camouflaged nanomaterials for biomedical applications. Nanomaterials. 2024;14(5):413. doi:10.3390/nano14050413
158. Priya S, Desai VM, Singhvi G. Surface modification of lipid-based nanocarriers: a potential approach to enhance targeted drug delivery. ACS omega. 2022;8(1):74–86. doi:10.1021/acsomega.2c05976
159. Osman N, Devnarain N, Omolo CA, Fasiku V, Jaglal Y, Govender T. Surface modification of nano‐drug delivery systems for enhancing antibiotic delivery and activity. Wiley Interdiscip Rev Nanomed Nanobiotechnol. 2022;14(1):e1758. doi:10.1002/wnan.1758
160. Anchordoquy TJ, Barenholz Y, Boraschi D, et al. Mechanisms and Barriers in Cancer Nanomedicine: Addressing Challenges, Looking for Solutions. ACS Publications; 2017.
161. Pelaz B, Alexiou C, Alvarez-Puebla RA, et al. Diverse applications of nanomedicine. ACS Nano. 2017;11(3):2313–2381. doi:10.1021/acsnano.6b06040
162. Li S, Li F, Wang Y, et al. Multiple delivery strategies of nanocarriers for myocardial ischemia-reperfusion injury: current strategies and future prospective. Drug Delivery. 2024;31(1):2298514. doi:10.1080/10717544.2023.2298514
163. Xue Y, Zeng G, Cheng J, Hu J, Zhang M, Li Y. Engineered macrophage membrane‐enveloped nanomedicine for ameliorating myocardial infarction in a mouse model. Bioeng Transl Med. 2021;6(2):e10197.
164. Marinho A, Nunes C, Reis S. On the design of cell membrane-coated nanoparticles to treat inflammatory conditions. Nanoscale Horiz. 2024;10:38–55. doi:10.1039/d4nh00457d
165. Wang S, Duan Y, Zhang Q, et al. Drug targeting via platelet membrane–coated nanoparticles. Small Struct. 2020;1(1):2000018. doi:10.1002/sstr.202000018
166. Zhang Q, Hu C, Feng J, et al. Anti-inflammatory mechanisms of neutrophil membrane-coated nanoparticles without drug loading. J Control Release. 2024;369:12–24. doi:10.1016/j.jconrel.2024.03.030
167. Harris JC, Scully MA, Day ES. Cancer cell membrane-coated nanoparticles for cancer management. Cancers. 2019;11(12):1836. doi:10.3390/cancers11121836
168. Wang M, Xin Y, Cao H, et al. Recent advances in mesenchymal stem cell membrane-coated nanoparticles for enhanced drug delivery. Biomater Sci. 2021;9(4):1088–1103. doi:10.1039/D0BM01164A
169. Ou Z, Zhong H, Zhang L, et al. Macrophage membrane-coated nanoparticles alleviate hepatic ischemia-reperfusion injury caused by orthotopic liver transplantation by neutralizing endotoxin. Int J Nanomed. 2020;15:4125–4138. doi:10.2147/IJN.S253125
170. Cao L, Du M, Cai M, et al. Neutrophil membrane-coated nanoparticles for targeted delivery of toll-like receptor 4 siRNA ameliorate LPS-induced acute lung injury. Int J Pharm. 2025;668:124960.
171. Joshi S, Allabun S, Ojo S, et al. Enhanced drug delivery system using mesenchymal stem cells and membrane-coated nanoparticles. Molecules. 2023;28(5):2130. doi:10.3390/molecules28052130
172. Wang H, Zang J, Zhao Z, Zhang Q, Chen S. The advances of neutrophil-derived effective drug delivery systems: a key review of managing tumors and inflammation. Int J Nanomed. 2021;Volume 16:7663–7681. doi:10.2147/IJN.S328705
173. Wu L, Xie W, Zan H-M, et al. Platelet membrane-coated nanoparticles for targeted drug delivery and local chemo-photothermal therapy of orthotopic hepatocellular carcinoma. J Mat Chem B. 2020;8(21):4648–4659. doi:10.1039/D0TB00735H
174. Wang J, Gu X, Ouyang Y, et al. Engineering of neutrophil membrane camouflaging nanoparticles realizes targeted drug delivery for amplified antitumor therapy. Int J Nanomed. 2021;16:1175–1187. doi:10.2147/IJN.S288636
175. Han H, Bártolo R, Li J, Shahbazi M-A, Santos HA. Biomimetic platelet membrane-coated nanoparticles for targeted therapy. Eur J Pharm Biopharm. 2022;172:1–15. doi:10.1016/j.ejpb.2022.01.004
176. Liu H, Su -Y-Y, Jiang X-C, Gao J-Q. Cell membrane-coated nanoparticles: a novel multifunctional biomimetic drug delivery system. Drug Delivery Transl Res. 2023;13(3):716–737. doi:10.1007/s13346-022-01252-0
177. Gao W, Xiao Y. Advances in cell membrane-encapsulated biomaterials for tissue repair and regeneration. Appl Mater Today. 2022;26:101389. doi:10.1016/j.apmt.2022.101389
178. Qu Y, Chu B, Li J, Deng H, Niu T, Qian Z. Macrophage‐biomimetic nanoplatform‐based therapy for inflammation‐associated diseases. Small Methods. 2024;8(7):2301178. doi:10.1002/smtd.202301178
179. Wang A, Wang X, Li D, et al. A superior method for antitumor therapy and application: dual-ligand nanomedicines. J Mat Chem B. 2025;13:6574–6596. doi:10.1039/d5tb00044k
180. Rao Z, Tang Y, Zhu J, et al. Enhanced FGF21 delivery via neutrophil-membrane-coated nanoparticles improves therapeutic efficacy for myocardial ischemia–reperfusion injury. Nanomaterials. 2025;15(5):346. doi:10.3390/nano15050346
181. Desai N, Tambe V, Pofali P, Vora LK. Cell membrane‐coated nanoparticles: a new frontier in immunomodulation. Adv NanoBiomed Res. 2024;4(8):2400012. doi:10.1002/anbr.202400012
182. Jiang T, Zhan Y, Ding J, et al. Biomimetic cell membrane‐coated nanoparticles for cancer theranostics. ChemMedChem. 2024;19(22):e202400410. doi:10.1002/cmdc.202400410
183. Zewail MB, Yang G, Fan Y, Hui Y, Zhao CX, Liu Y. Cell membrane‐coated lipid nanoparticles for drug delivery. Aggregate. 2025;6:e70054. doi:10.1002/agt2.70054
184. Boraei SBA, Javadpour S, Nejad FK, Otaghvari FH, Zare Y, Rhee KY. Recent advances on the application of nanobiomimetic structures as drug delivery systems. J Drug Delivery Sci Technol. 2024;106009.
185. Bagheri E, Sameiyan E, Hassibian S, et al. Modification of cell membrane-coated platforms for targeted drug delivery. In: Cell Membrane Surface-Engineered Nanoparticles: Biomimetic Nanomaterials for Biomedical Applications. ACS Publications; 2024:57–88.
186. Zhang D, Wang G, Ma N, et al. Biomedical applications of cell membrane‐based biomimetic nano‐delivery system. Adv Ther. 2024;7(2):2300304.
187. Huang D, Wang X, Wang W, Li J, Zhang X, Xia B. Cell-membrane engineering strategies for clinic-guided design of nanomedicine. Biomater Sci. 2024.
188. Khan M, Ullah R, Wang G, Chu M. Macrophage membrane-functionalized nanotherapeutics for tumor targeted therapy. Theranostics. 2025;15(10):4823. doi:10.7150/thno.108875
189. Villa CH, Anselmo AC, Mitragotri S, Muzykantov V. Red blood cells: supercarriers for drugs, biologicals, and nanoparticles and inspiration for advanced delivery systems. Adv Drug Delivery Rev. 2016;106:88–103. doi:10.1016/j.addr.2016.02.007
190. Yaman S, Chintapula U, Rodriguez E, Ramachandramoorthy H, Nguyen KT. Cell-mediated and cell membrane-coated nanoparticles for drug delivery and cancer therapy. Cancer Drug Resistance. 2020;3(4):879. doi:10.20517/cdr.2020.55
191. Li J, Wei Y, Zhang C, et al. Cell-membrane-coated nanoparticles for targeted drug delivery to the brain for the treatment of neurological diseases. Pharmaceutics. 2023;15(2):621. doi:10.3390/pharmaceutics15020621
192. Zhang W, Huang X. Stem cell membrane-camouflaged targeted delivery system in tumor. Mater Today Bio. 2022;16:100377. doi:10.1016/j.mtbio.2022.100377
193. Zhu C, Ma J, Ji Z, Shen J, Wang Q. Recent advances of cell membrane coated nanoparticles in treating cardiovascular disorders. Molecules. 2021;26(11):3428. doi:10.3390/molecules26113428
194. Zhao Q, Jiang D, Sun X, et al. Biomimetic nanotherapy: core–shell structured nanocomplexes based on the neutrophil membrane for targeted therapy of lymphoma. J Nanobiotechnol. 2021;19:1–19. doi:10.1186/s12951-021-00922-4
195. Liang S, Wang M, Wang J, Chen G. Red‐blood‐cell‐membrane‐coated metal‐drug nanoparticles for enhanced chemotherapy. ChemBioChem. 2021;22(22):3184–3189. doi:10.1002/cbic.202100313
196. Sun H, Su J, Meng Q, et al. Cancer cell membrane‐coated gold nanocages with hyperthermia‐triggered drug release and homotypic target inhibit growth and metastasis of breast cancer. Adv Funct Mater. 2017;27(3):1604300. doi:10.1002/adfm.201604300
197. Zhang W, Huang X. Stem cell-based drug delivery strategy for skin regeneration and wound healing: potential clinical applications. Inflamm Regen. 2023;43(1):33. doi:10.1186/s41232-023-00287-1
198. Rezaei N, Zadory M, Babity S, Marleau S, Brambilla D. Therapeutic applications of nanoparticles targeting neutrophil and extracellular traps. J Control Release. 2023;358:636–653. doi:10.1016/j.jconrel.2023.05.018
199. Zou S, Wang B, Wang C, Wang Q, Zhang L. Cell membrane-coated nanoparticles: research advances. Nanomedicine. 2020;15(6):625–641. doi:10.2217/nnm-2019-0388
200. Wang C, Li C, Zhang R, Huang L. Macrophage membrane-coated nanoparticles for the treatment of infectious diseases. Biomed Mater. 2024;19(4):042003. doi:10.1088/1748-605X/ad4aaa
201. Chugh V, Vijaya Krishna K, Pandit A. Cell membrane-coated mimics: a methodological approach for fabrication, characterization for therapeutic applications, and challenges for clinical translation. ACS Nano. 2021;15(11):17080–17123. doi:10.1021/acsnano.1c03800
202. Malhotra S, Dumoga S, Singh N. Red blood cells membrane‐derived nanoparticles: applications and key challenges in their clinical translation. Wiley Interdiscip Rev. 2022;14(3):e1776. doi:10.1002/wnan.1776
203. He Y, Zhang S, She Y, et al. Innovative utilization of cell membrane‐coated nanoparticles in precision cancer therapy. In Exploration. Wiley Online Library; 2024:20230164.
204. He Y, Li R, Liang J, et al. Drug targeting through platelet membrane-coated nanoparticles for the treatment of rheumatoid arthritis. Nano Res. 2018;11:6086–6101. doi:10.1007/s12274-018-2126-5
205. Narain A, Asawa S, Chhabria V, Patil-Sen Y. Cell membrane coated nanoparticles: next-generation therapeutics. Nanomedicine. 2017;12(21):2677–2692. doi:10.2217/nnm-2017-0225
206. Ag Seleci D, Maurer V, Barlas FB, et al. Transferrin-decorated niosomes with integrated InP/ZnS quantum dots and magnetic iron oxide nanoparticles: dual targeting and imaging of glioma. Int J Mol Sci. 2021;22(9):4556. doi:10.3390/ijms22094556
207. Sun Y, Su J, Liu G, et al. Advances of blood cell-based drug delivery systems. Eur J Pharm Sci. 2017;96:115–128. doi:10.1016/j.ejps.2016.07.021
208. Xuan M, Shao J, Li J. Cell membrane-covered nanoparticles as biomaterials. Natl Sci Rev. 2019;6(3):551–561. doi:10.1093/nsr/nwz037
209. Liao W, Lin J, Wang W, et al. Assembly of ceria-Nrf2 nanoparticles as macrophage-targeting ROS scavengers protects against myocardial infarction. Front Pharmacol. 2025;15:1503757. doi:10.3389/fphar.2024.1503757
210. Wu S-Y, Fu T, Jiang Y-Z, Shao Z-M. Natural killer cells in cancer biology and therapy. Mol Cancer. 2020;19:1–26. doi:10.1186/s12943-019-1085-0
211. Chester C, Fritsch K, Kohrt HE. Natural killer cell immunomodulation: targeting activating, inhibitory, and co-stimulatory receptor signaling for cancer immunotherapy. Front Immunol. 2015;6:601. doi:10.3389/fimmu.2015.00601
212. Lorenzo-Herrero S, López-Soto A, Sordo-Bahamonde C, Gonzalez-Rodriguez AP, Vitale M, Gonzalez S. NK cell-based immunotherapy in cancer metastasis. Cancers. 2018;11(1):29. doi:10.3390/cancers11010029
213. Vivier E, Ugolini S, Blaise D, Chabannon C, Brossay L. Targeting natural killer cells and natural killer T cells in cancer. Nat Rev Immunol. 2012;12(4):239–252. doi:10.1038/nri3174
214. Rao L, Cai B, Bu -L-L, et al. Microfluidic electroporation-facilitated synthesis of erythrocyte membrane-coated magnetic nanoparticles for enhanced imaging-guided cancer therapy. Acs Nano. 2017;11(4):3496–3505. doi:10.1021/acsnano.7b00133
215. Bjij I, Khan S, Ramharak P, Cherqaoui D, Soliman MES. Distinguishing the optimal binding mechanism of an E3 ubiquitin ligase: covalent versus noncovalent inhibition. J Cell Biochem. 2019;120(8):12859–12869. doi:10.1002/jcb.28556
216. Gu Y, Du L, Wu Y, et al. Biomembrane-modified biomimetic nanodrug delivery systems: frontier platforms for cardiovascular disease treatment. Biomolecules. 2024;14(8):960. doi:10.3390/biom14080960
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.